 Amir Ali Khoddamzadeh
Amir Ali Khoddamzadeh Jason Flores
Jason Flores M. Patrick Griffith
M. Patrick Griffith Bárbara Nogueira Souza Costa
Bárbara Nogueira Souza Costa
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 12 May 2023
Sec. Biogeography and Macroecology
Volume 11 - 2023 | https://doi.org/10.3389/fevo.2023.1127679
This article is part of the Research Topic Environmental Threats to the State of Florida—Climate Change and Beyond, volume II View all 10 articles
Climate change will alter natural areas on a global scale within the next century, especially in low-lying coastal areas where sea-level rise is predicted to severely degrade or destroy many ecosystems. As sea-level rise continues, it is expected that salinity due to saltwater intrusion will impact soil health and agricultural production, this is of even greater importance in areas such as South Florida where the surface and groundwater resources are hydrologically connected due to the shallow and highly permeable limestone soils. The chlorophyll concentrations in leaf tissue were chosen as the primary health indicator to assess whether this is a valuable factor to consider for plant health risk assessment and whether optical sensor technology such as the SPAD and NDVI, are valuable tools when understanding the impact seawater encroachment has on plant nitrogen uptake. Therefore, the aim of this study was to evaluate the effects of salt concentrations on plant growth and health of four palm species. The treatments were from pure reverse osmosis water, Fresh water, brackish water, seawater and hypersaline water (0, 0.5, 5, 10, 15, 25, and 50 ppt). Control treatment that received reverse osmosis filtrated water with a salinity concentration of 0 ppt had the highest health-indicator averages compared to treatment 50 ppt. As the salinity increased, all health indicators gradually declined or remained rather constant and declined as they approached the greater salinity concentrations. The growth parameters and chlorophyll content were reduced over time, particularly at higher salt concentrations for all palm species studied. The chlorophyll content by SPAD and the normalized difference vegetation index (NDVI) can be used as health indicators of the palm species studied, as it was observed in this study a decline in the chlorophyll content as there was an increase in salt concentrations. It is important to emphasize that leaf chlorosis and subsequent decline of palm species were observed in the treatment with higher salt concentration, thus highlighting the importance of chlorophyll content as an indicator of plant health. The species Roystonea regia was very susceptible to salinity, and the palm Sabal palmetto is more susceptible to salinity than the Thrinax radiata and Pseudophoenix sargentii palms.
Climate change will alter natural areas on a global scale within the next century, especially in low-lying coastal areas where sea-level rise is predicted to severely degrade or destroy many ecosystems (Braun de Torrez et al., 2021). Climate change and variability contribute to increased warmer conditions, increased frequency of heavy rain that accounts for an increasing proportion of total rainfall, extreme weather characterized by spatially variable cycles of drought and wetness, increased frequency of tropical storms/hurricanes, increased frequency of storm surges, and accelerated rate of sea-level rise (SLR). As SLR continues, it is expected that salinity due to saltwater intrusion (SWI) will impact soil health and agricultural production, this is of even greater importance in areas such as South Florida where the surface and groundwater resources are hydrologically connected due to the shallow and highly permeable limestone soils (Bayabil et al., 2021). South Florida is home to a diversity of palms (Arecaceae) with 9 native species and many more represented in cultivation for commercial production (Broschat and Black, 2021).
Native palms provide structural integrity and resources for the ecosystems they inhabit, and non-native palms are not only emblematic of the tropics, but generators of novel self-containing ecosystems (Fehr et al., 2020). Montgomery Botanical Center is a nonprofit botanical garden and research institution located in the city of Coral Gables, Florida, just south of Miami. As a coastal landsite hosting an outdoors living collection of nationally accredited palms and cycads that is recognized as vulnerable to sea level-rise and “King Tide” events, we chose to design an ex situ experiment based on observations and inferences from this site (Griffith et al., 2017). The coastal location of the botanic garden results in frequent inundation from encroaching oceanwater coming from coastal mangrove forest in the Southeastern section of the property. Previous studies have investigated the effects of saltwater on palms, focusing primarily on the commercially and economically valuable Phoenix dactylifera (Yaish and Kumar, 2015). Specific to South Florida, the Cabbage palm (Sabal palmetto) has been the subject of many salinity trials (Perry and Williams, 1996).
The relative “greenness” of a leaf can serve as a proxy for chlorophyll concentration and leaf nitrogen levels. This is the very correlation that remote sensing and vegetation indices technologies exploit (Inman et al., 2005). Nitrogen is known to affect plant health indicators such as leaf size, weight, overall plant size, and transpiration rates (Basyouni and Dunn, 2013). The soil plant analysis development (SPAD) sensor is a diagnostic tool used to estimate the nitrogen status of crop foliage in an efficient and non-destructive manner (Sim et al., 2015). Normalized difference vegetation index (NDVI) has been proven to be a useful tool in gaining insights into photosynthetic efficiency, productivity, and yield (Inman et al., 2005).
Thereby, chlorophyll concentrations in leaf tissue were chosen as the primary health indicator to assess whether this is a valuable factor to consider for plant health risk assessment and whether optical sensor technology such as the SPAD and NDVI, are valuable tools when understanding the impact seawater encroachment has on plant nitrogen uptake. SPAD and NDVI sensors offer an insight into the uptake of macronutrients and if any known environmental changes have affected the plant’s ability to produce favorable or desirable amounts of chlorophyll. In the context of this experiment optical sensors were utilized to determine any changes in Nitrogen-plant utilization in the presence of saltwater. Thus, assessing the correlation between leaf tissue chlorophyll concentration and saltwater intrusion presents an opportunity for new technology adoption for concerns regarding plants and salinity exposure. Therefore, the aim of this study was to evaluate the effects of salt concentrations on plant growth and plant health of four palm species.
Four-year-old palm saplings were used for the species Pseudophoenix sargentii, Roystonea regia, and Thrinax radiata collected from Montgomery Botanic Center. Saplings were transplanted from 25.4 cm nursery pots to 35.56 cm nursery pots using a potting soil blend that consists of 25% Canadian peat, 40% pine bark, 25% perlite, and 10% coarse sand. Three-year-old palms were used for the species S. palmetto “Lisa.” This group of palms had not outgrown their original nursery pots of 15.25 cm, and therefore were not transplanted. A low-rate dosage of 14-4-14 Nutricote slow-release fertilizer was administered to all palms a month prior to the experiment. This was done to rule out the possibility of nutrient deficiencies and to achieve uniformity across each plant. A total of 140 palms were involved in this study, 35 of each species. P. sargentii and S. palmetto “Lisa” were donated by Montgomery Botanical Center and the remaining species were purchased at a local South Florida plant nursery.
The plants were grown in the Montgomery Botanical Center. The site for the ex situ experiment was exposed to plenty of sunlight with Southern exposure. Along the edges of the site some neighboring trees cast little shade during early parts of the day, but plants were shuffled in random blocks to address this. With no shade cloth these palms were exposed to all possible rainfall which was accounted for as an analogous for natural freshwater recharge as would occur in the area. A ratio of 45:7 was recorded for rainfall days. Throughout the trial a total of 38.88 cm (15.31 inch) of rainwater was documented. This was accounted for as natural freshwater buffering also occurs in natural settings. Data was recorded during the months of January through June, which had average temperatures of 26.6°C or 79.88°F, which is favorable conditions for these palm species. Palms were arranged in no particular order in the study site and were shuffled in random order twice throughout the study in order to rule out environmental differences such as sunlight exposure, rainwater, and proximity to other elements.
Treatments were formulated using commercially available aquarium salts in order to closely mimic seawater and retain elements naturally found in the water encroaching coastal and vulnerable landsites. Salinity concentrations were measured using a commercially available refractometer that measures concentrations in ppt. Using reverse osmosis water as the base for the solution, this ensured no unwanted factors impacting results coming from tap water treatments and residues. Treatments of saltwater were administered uniformly on the same date with a frequency of once every 7 days including with rainfall. Root zones were saturated with solution until leachate was observed at the bottom of the pots. This was administered over a period of 6 months beginning in January 2021 and ending in June 2021. Pure reverse osmosis water was used for the control. So, the treatments were 0 ppt which served as the control, 0.5, 5, 10, 15, 25, and 50 ppt. Fresh water represented by the concentration 0.5 ppt. Brackish water was represented by concentrations 5, 10, and 15 ppt. Seawater was represented by the concentration 25 ppt. Hypersaline water was represented by 50 ppt. The waters used for the treatments were collected from the lakes around the Montgomery Center, salt was not added to the waters used, but the amount of salt in each of them was measured. Plant health parameters consisting of plant height (cm), leaf count (unit), SPAD sensor readings, and Normalized Difference Vegetation Index—NDVI sensor readings were collected uniformly once per month. The Konica Minolta Chlorophyll meter SPAD-502 and the Trimble Greenseeker handheld crop sensor were used to record chlorophyll readings for all treatment and palm species. Three readings were taken from each plant per sensor and averaged out.
The experiment was established in a completely randomized design with seven treatments replicated 5 times with single pot replications (one plant in each pot), totaling 20 plants per species. Data were subjected to analysis of variance (ANOVA) and the means compared by Tukey’s test (p ≤ 0.05) using the SISVAR statistical program (Ferreira, 2011).
In this study two factors were studied, salt concentrations and evaluation periods represented by days after treatment (DAT). For some plant growth and health parameters there was no interaction between these factors and therefore they were evaluated separately, for all palm species used in this study.
The treatment 0 ppt of salt increased leaf numbers (4.33) compared to 0.5 and 5 ppt (3.87, and 3.80), respectively in of P. sargentii. The salt concentrations (25 and 50 ppt) provided an increased number of leaves (4.27 and 4.23) of P. sargentii plants, respectively then 5 ppt treatment (3.80). Greater plant height of P. sargentii was observed in the treatment control (0 ppt; 152.44), compared to (5, 15, 25, and 50 ppt) treatments (119.65, 138.47, 130.18, 122.19, and 106.31), respectively. The 0.5 ppt treatment provided an increase plant height (141.94) of P. sargentii compared to (5, 25, and 50 ppt) treatments (119.65, 122.19, and 106.31), respectively, and the 10 ppt provided treatment an increase plant height (138.47) than (5 ppt—119.65 and 50 ppt—106.31) treatments. However, the (0, 0.5, and 5 ppt) treatments provided an increase for NDVI values (0.81, 0.81, and 0.80), correspondingly than (25 ppt—0.75 and 50 ppt—0.72) treatments, and the 10 and 15 ppt treatments provided an increase for NDVI values (0.79 and 0.78), subsequently compared to 50 ppt treatment (0.72; Table 1).
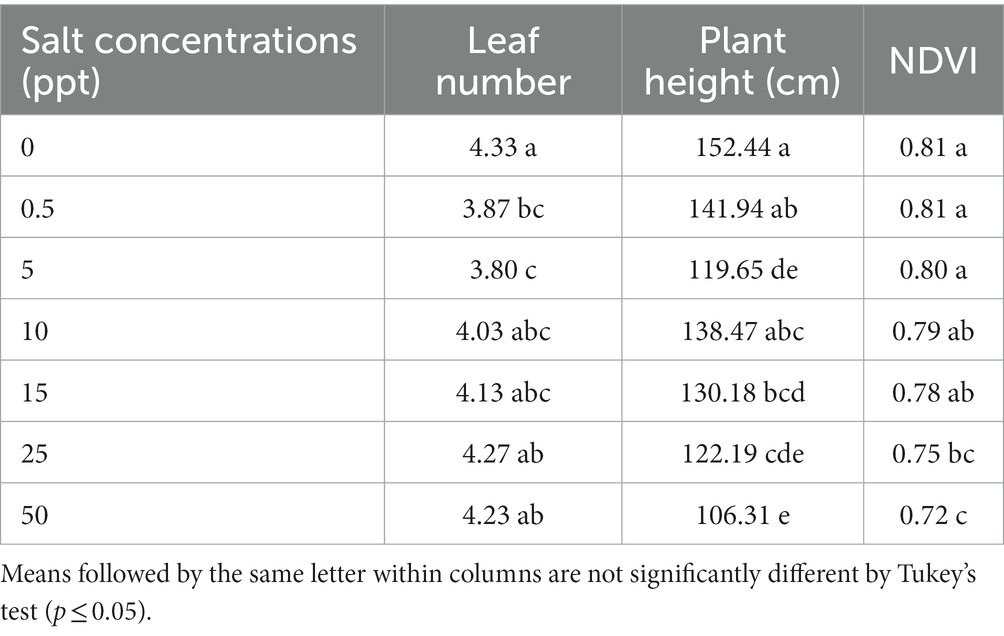
Table 1. Leaf number, plant height, and normalized difference vegetation index values (NDVI) of Pseudophoenix sargentii plants grown in saline conditions.
Greater leaf number of P. sargentii was observed at 150 DAT (4.37) compared to 60 DAT (3.94) and 90 DAT (3.91). For plant height an increase was observed in this characteristic at 90 DAT (137.01), 120 DAT (137.01), and 150 DAT (139.14) than 0, 30, and 60 DAT (118.91, 122.29, and 126.64), correspondingly. Greater NDVI values of P. sargentii was observed at 30, 60, 90, 120 and 150 DAT (0.79, 0.79, 0.79, 0.80 and 0.78), respectively than 0 DAT (0.73; Table 2).
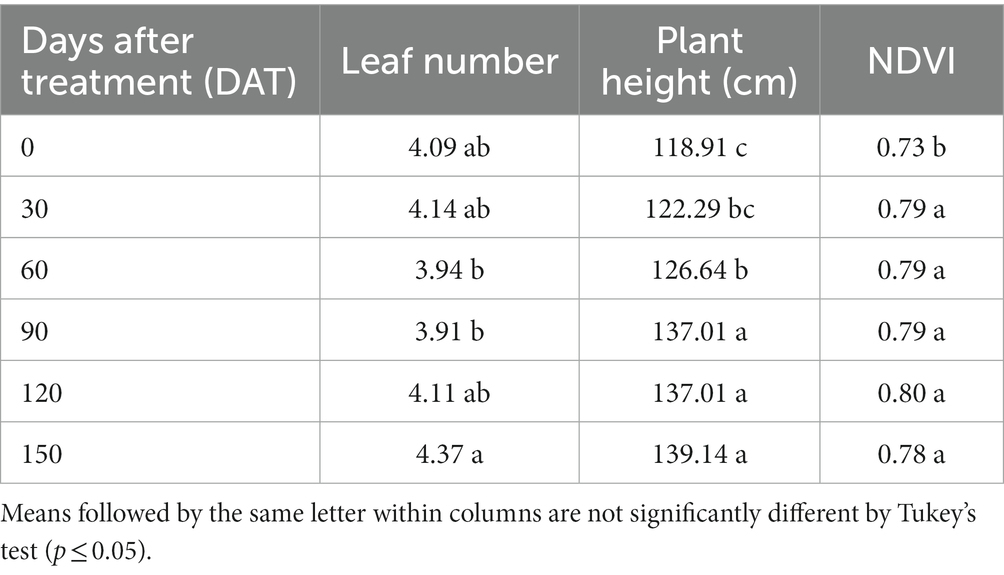
Table 2. Leaf number, plant height and normalized difference vegetation index values (NDVI) of Pseudophoenix sargentii plants at 0, 30, 60, 90, 120 and 150 days after treatment.
There was significantly interaction (p ≤ 0.05) between salt concentrations and evaluation periods represented by days after treatment (DAT) for SPAD values of P. sargentii plants. The 0 ppt treatment provided increase in SPAD values at 90 DAT (90.24; Table 3).
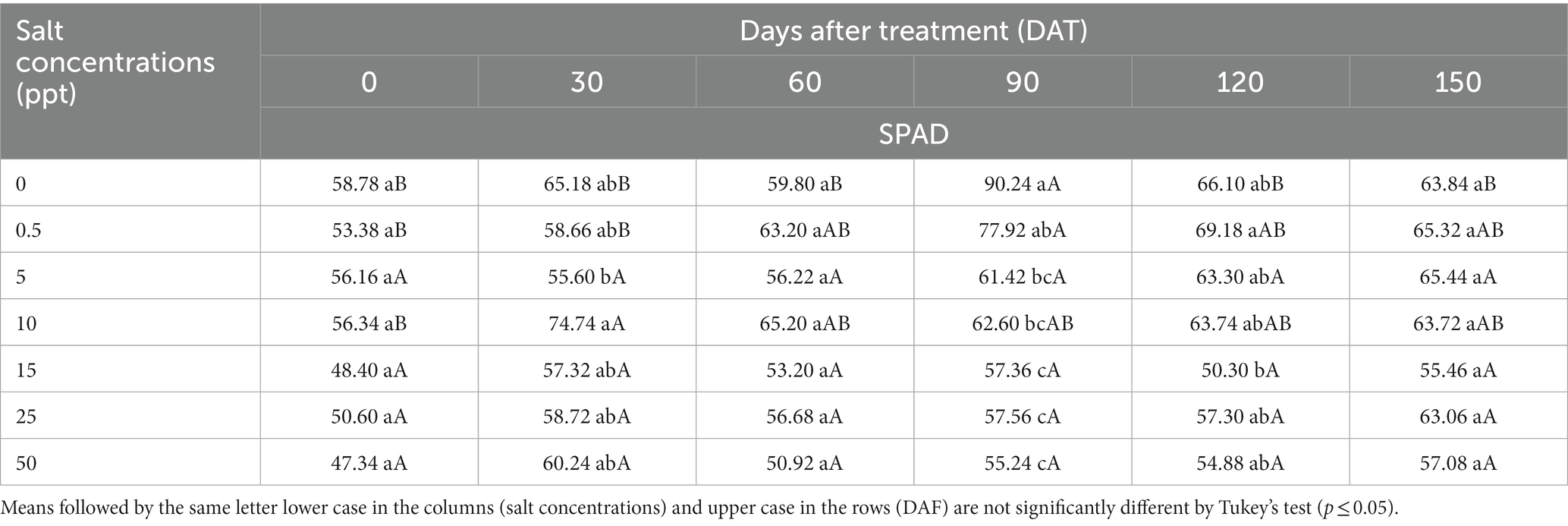
Table 3. SPAD values of Pseudophoenix sargentii plants grown in saline conditions at 0, 30, 60, 90,120, and 150 days after treatment.
Control (0 ppt) treatment provided an increase in plant height (133.68) of R. regia plants compared to (0.5, 10, and 25 ppt) treatments (118.88, 120.55, and 114.69), respectively (Table 4).
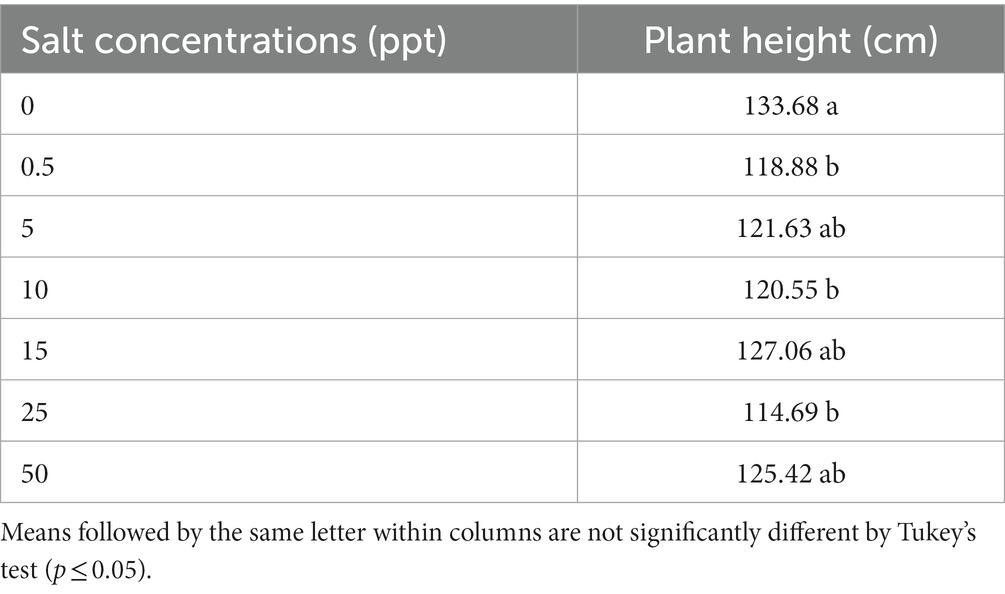
Table 4. Plant height of Roystonea regia grown in saline conditions.
No significant differences (p ≤ 0.05) were observed for plant height of R. regia at 0, 30 and 60 DAT (Table 5). It is important to note that R. regia is a salinity susceptible palm specie because the plants exposed to 25 and 50 ppt treatments died, so we do not have data for the 90, 120, and 150 DAT for this species. Therefore, to homogenize we evaluated all treatments only at 0, 30, and 60 DAT.
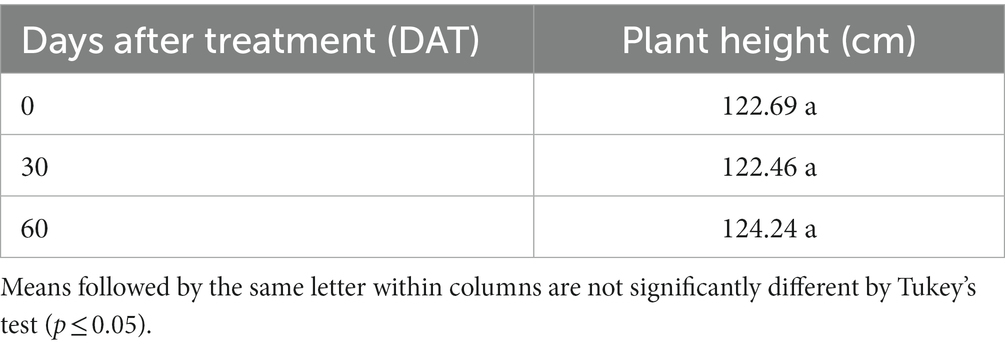
Table 5. Plant height of Roystonea regia at 0, 30, and 60 days after treatment.
There was significantly interaction (p ≤ 0.05) between salt concentrations and evaluation periods represented by days after treatment (DAT) for leaves number, SPAD and NDVI values of R. regia. The 50 ppt treatment provided a decreased leaves number (1.40), SPAD (29.96) and NDVI values (0.31) at 60 DAT compared to other treatments (Table 6).
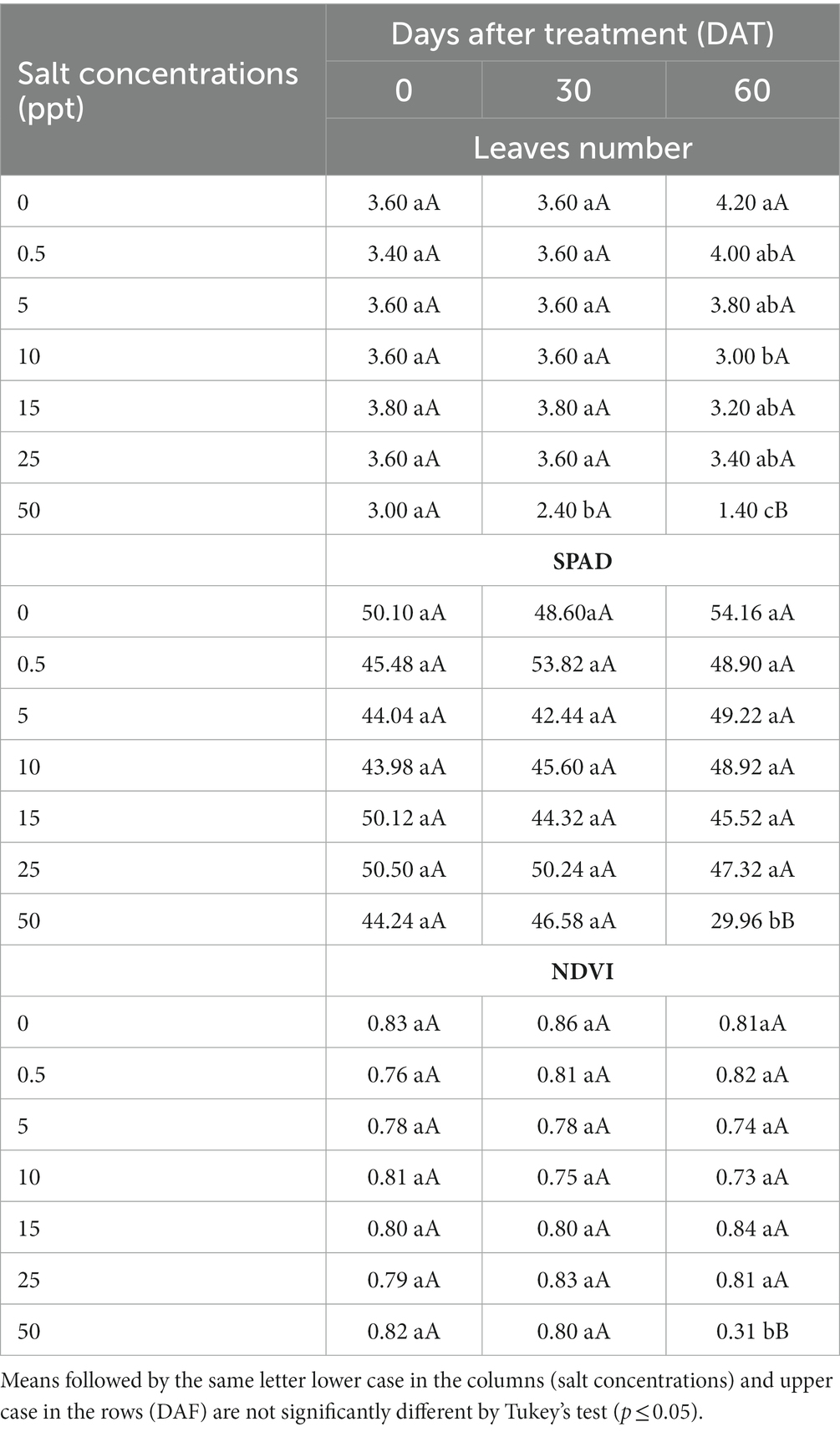
Table 6. Leaves number, SPAD values, and normalized difference vegetation index values (NDVI) of Roystonea regia plants grown in saline conditions at 0, 30, and 60 days after treatment.
The (10 ppt—52.03 and 25 ppt—50.43) treatments provided an increase SPAD values of leaves of T. radiata plants compared to 50 ppt treatment (43.60). However, the (0, 0.5, 5, 10, 15, and 25 ppt) treatments provided an increase for NDVI values (0.86, 0.86, 0.85, 0.86, 0.85, and 0.84), correspondingly than 50 ppt treatment (0.78; Table 7).
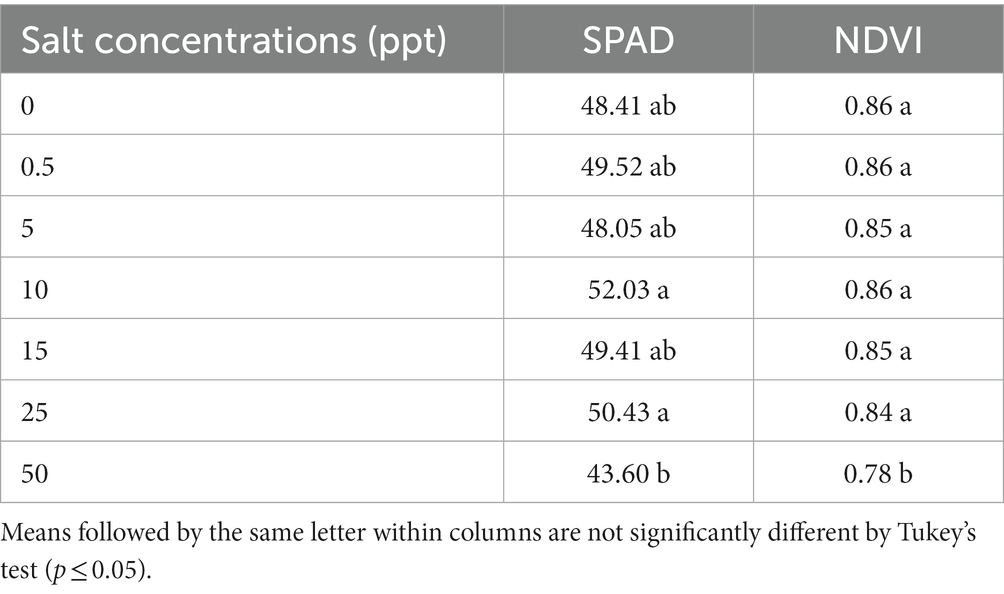
Table 7. SPAD values, and normalized difference vegetation index values (NDVI) of Thrinax radiata plants grown in saline conditions.
Greater SPAD values of T. radiata was observed at 90 DAT (51.22), and 120 DAT (50.87) compared to 0 DAT (45.45). For NDVI values an increase was observed in this characteristic at 30 DAT (0.88) than 0, 90, 120 and 150 DAT (0.83, 0.83, 0.84, and 0.81), respectively (Table 8).
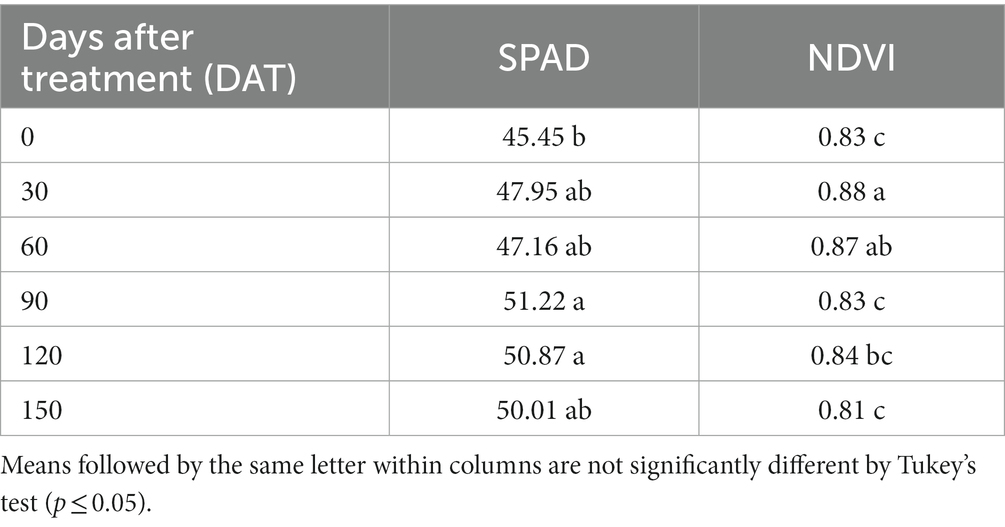
Table 8. SPAD values, and normalized difference vegetation index values (NDVI) of Thrinax radiata plants at 0, 30, 60, 90,120, and 150 days after treatment.
There was significantly interaction (p ≤ 0.05) between salt concentrations and evaluation periods represented by days after treatment (DAT) for leaves number and plant height of T. radiata plants. The 50 ppt treatment provided a decreased leaves number at 150 DAT (4.40) compared to other treatments. However, it is important to note that the plant height increase was observed in the 0 ppt treatment at 90, 120 and 150 DAT (107.70, 115.00, and 119.40), subsequently (Table 9).
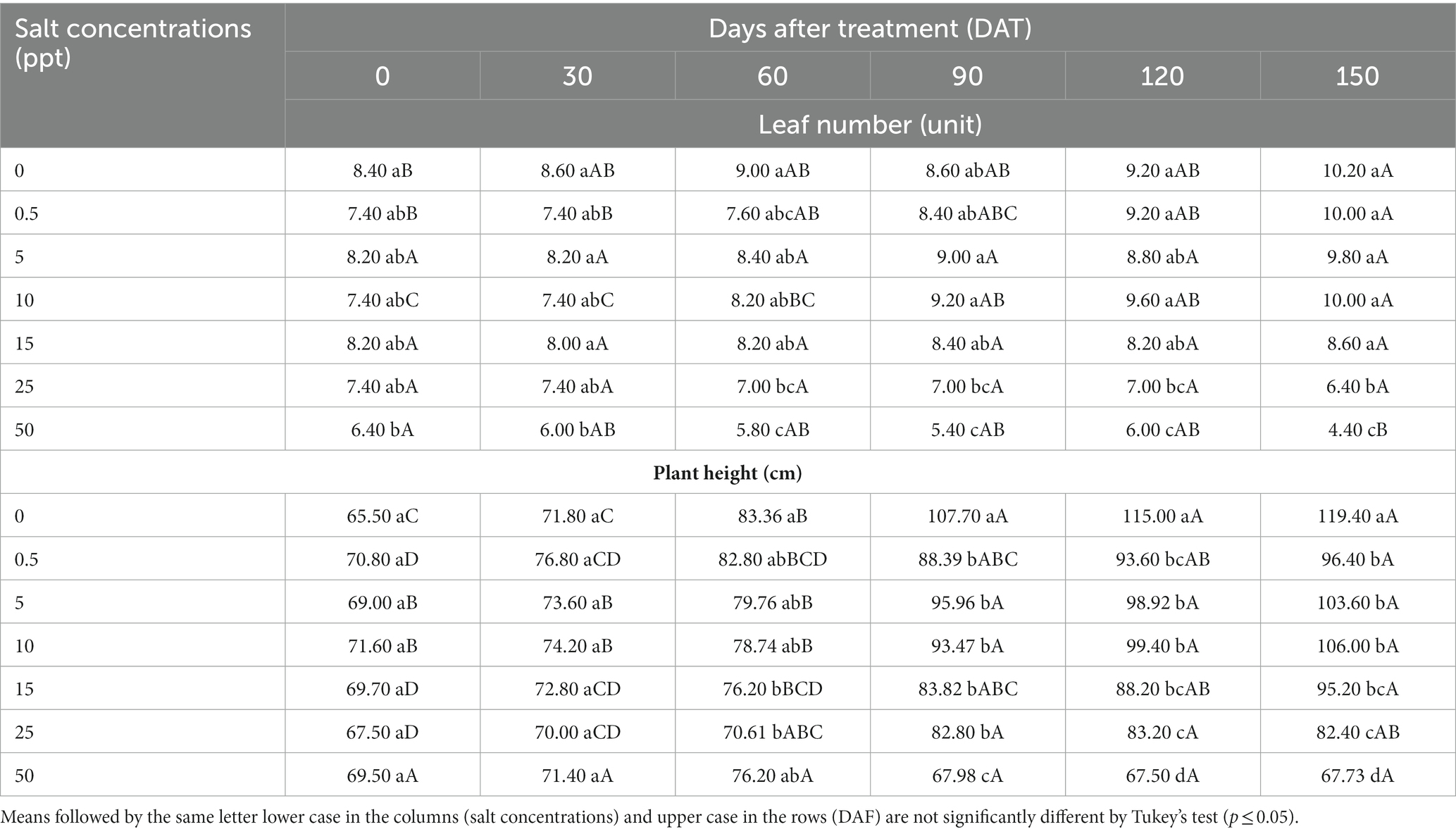
Table 9. Leaf number, and Plant height of Thrinax radiata plants grown in saline conditions at 0, 30, 60, 90,120, and 150 days after treatment.
The 0.5 ppt treatment provided an increase plant height (31.64) of S. palmetto plants compared to (0, 5, 10, 15, 25, and 50 ppt) treatments (27.40, 27.19, 26.95, 21.99, 21.55, and 20.44), appropriately. However, the (0 ppt—27.40, 5 ppt—27.19, and 10 ppt—26.95) treatments provided an increase plant height of S. palmetto plants than (15 ppt—21.99, 25 ppt—21.55, and 50 ppt—20.44; Table 10).
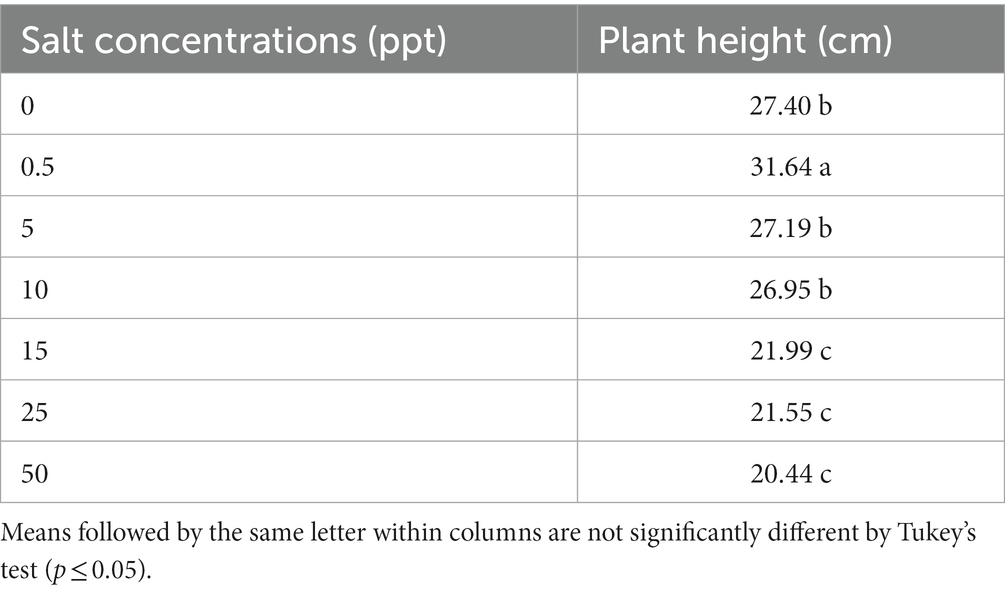
Table 10. Plant height of Sabal palmetto grown in saline conditions.
No significant differences (p ≤ 0.05) were observed for plant height of S. palmetto at 0, 30, 60, 90, 120, and 150 DAT (Table 11).
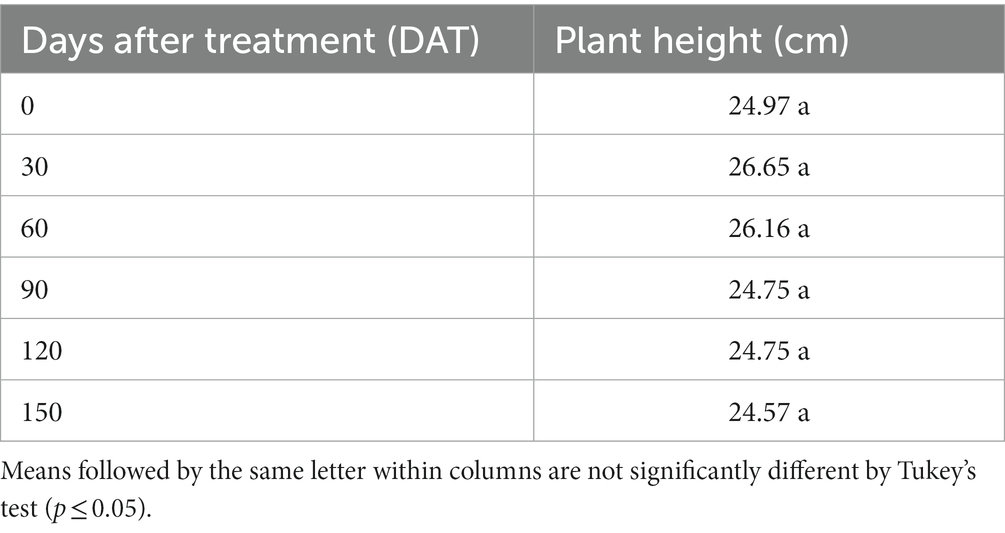
Table 11. Plant height of Sabal palmetto at 0, 30, 60, 90,120, and 150 days after treatment.
There was significantly interaction (p ≤ 0.05) between salt concentrations and evaluation periods represented by days after treatment (DAT) for leaves number, SPAD and NDVI values of S. palmetto plants. The 50 ppt treatment provided a decreased leaves number at 90 DAT (1.80), and 120 DAT (1.80), for SPAD values at 120 DAT (22.82), and 150 DAT (19.22), and for NDVI at 90 DAT (0.32), 120 DAT (0.32), and 150 DAT (0.26) compared to other treatments (Table 12).
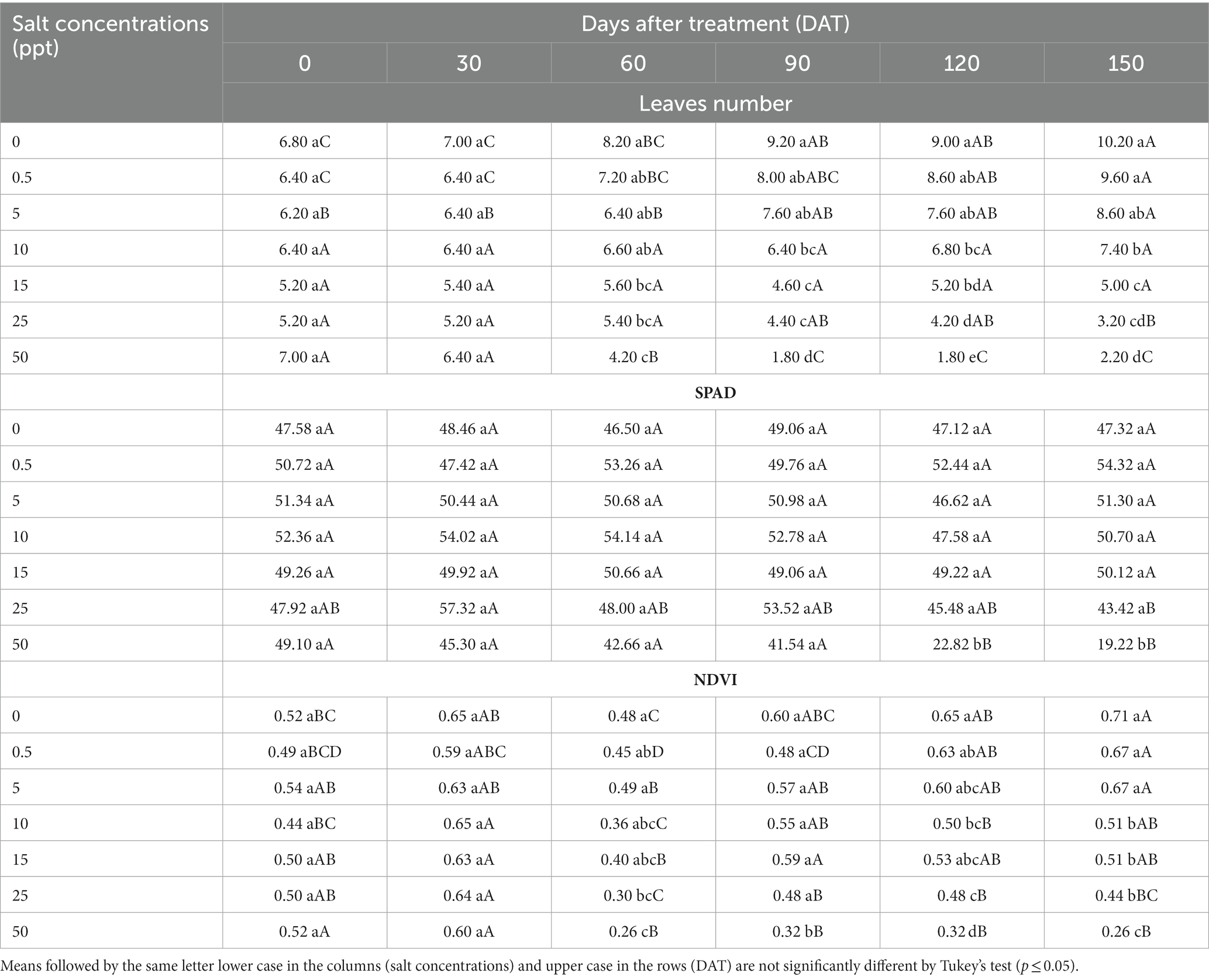
Table 12. Leaves number, SPAD values, and normalized difference vegetation index values (NDVI) of Sabal palmetto plants grown in saline conditions at 0, 30, 60, 90,120, and 150 days after treatment.
In order to verify which palm species was more resistant to saline conditions in this study, the interaction was performed between the factors salt concentrations and palm species, so that there was a significant interaction (p ≤ 0.05) between the factors studied salt concentrations and palm species (Table 13).
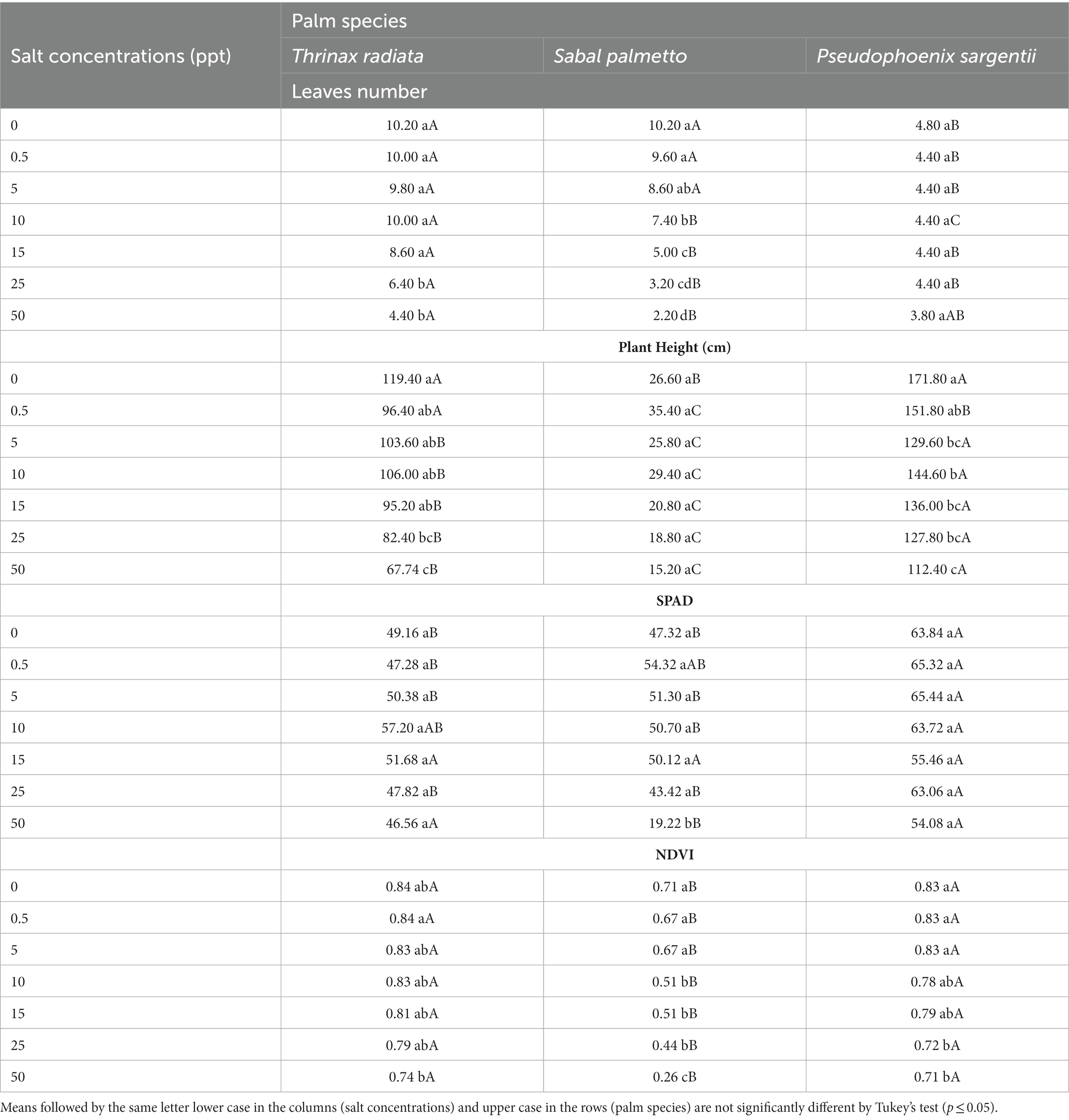
Table 13. Leaf number, plant height, SPAD values, and normalized difference vegetation index values (NDVI) of palm species grown in saline conditions at 150 days after treatment.
In general, with the increase in the salt concentration, there was a decrease in the values to growth parameters and chlorophyll content for all species. The 50 ppt treatment for the palm S. palmetto provided a decreased leaves number (2.20), plant height (15.20), SPAD (19.22) and NDVI (0.26) at 150 DAT compared to the other palm species (Table 13). Therefore, the palm S. palmetto is more susceptible to high salt concentrations than the T. radiata and P. sargentii palms.
In response to increasing exposure to high concentrations of salinity in simulated seawater, palm health indicators follow a declining trend. Lower concentrations of salinity also lead health indicators to decline, but with less magnitude. Over the six-month period that data was collected from the ex situ trial, it became visible that the hypersaline treatments (50 ppt) were negatively affecting the palms at a much faster rate. Chlorosis and leaf tissue burn became visible in just 3 months, with some palms completely declining the following month. By the end of the trial a total of 8 palms out of the 140 reached complete decline. Five R. regia, one T. radiata, and two S. palmetto “Lisa” all belonging to treatment that received the hypersaline solution with a salt concentration of 50 ppt. Control treatment that received reverse osmosis filtrated water with a salinity concentration of 0 ppt had the highest health-indicator averages compared to treatment 50 ppt. As the salinity increased, all health indicators gradually declined or remained rather constant and declined as they approached the greater salinity concentrations. At the end of the study, the effects of increased salinity concentration became very apparent simply by observing the palms, this can be appreciated in Figure 1 as it shows S. palmetto “Lisa” grow in saline conditions at 150 days after treatment, each treatment in ascending order from left to right (Figure 1). Pure reverse osmosis water was used for the control. So, the treatments were 0 ppt which served as the control. Fresh water represented by the concentration 0.5 ppt. Brackish water was represented by concentrations 5, 10, and 15 ppt. Seawater was represented by the concentration 25 ppt. Hypersaline water was represented by 50 ppt.

Figure 1. S. palmetto ‘Lisa’ grown in saline conditions at 150 days after treatment, each treatment in ascending order from left to right.
This study examines the ecophysiological effects of palms native to South Florida facing pressures from climate change such as sea level rise. A major consequence affecting plants with sea level rise is saltwater intrusion. Increased exposure to saltwater over time for established palms appears to have less of an effect on their longevity when compared to seedlings or saplings. Plant health decline can be measured and observed in as little as 6 months with the salinity gradient present in this study. This was documented across the four palm species found in this trial, suggesting the possibility of many palm seedlings and saplings not overcoming a similar salinity gradient in vulnerable landsites.
Ecophysiological data is necessary to interpret and model proper responses to seawater encroachment. The data gathered and visible evidence suggest that chlorophyll tissue concentration and plant health declines as exposure to high concentrations of salinity prolongs. It is expected that as chronic exposure to brackish, oceanwater, and hypersaline concentration of salinity continues, plant health will decline appreciably. The data also suggests that extended exposure to lower concentrations of salinity also negatively impacts plant health, but over an extended period. Within the six-month time frame of this trial, chlorosis and leaf burn became apparent and was documented. In comparison to the control which received irrigation with complete absence of salinity, all plants treated with any amount of salinity scored lower in health indicators. This occurred quickly withing a 6-month period. If this translates to habitats, then it could suggest that saplings and seedlings would have a very tough time establishing themselves into the vegetative state.
Other authors also observed negative effects of salinity on the growth of some plants, corroborating the results found in this study, such as Nascimento et al. (2019) observed saline stress reduces the levels of the photosynthetic pigments and affects adversely the growth, causing reductions in plant height, stem diameter, leaf area and number of leaves of the cotton cultivars. Oliveira et al. (2018) observed that increasing salinity in water (CEa) levels significantly affected the initial growth and gas exchange of cotton cultivars, both at 15 and 30 DAT. Ait-El-Mokhtar et al. (2022) concluded that salt stress induced significant adverse effects on growth, physiological and biochemical attributes of date palm plants. Yu et al. (2021) observed that the plant growth and vigorousness of Hibiscus rosa-sinensis and Mandevilla splendens grown in nursery crops in south Florida were significantly limited when the salinity concentrations increased. The same authors highlight the need to monitor the salinity levels and prepare groundwater salinity mitigation plans under the projected increases in sea level and the resulting increases in the frequency and severity of saltwater intrusion in south Florida.
The plants, when cultivated under soil with salinity or irrigated with water saline, have their growth compromised due to the osmotic effect, which reduces the water absorption by the plant and/or as a function of the specific effect of the ions that cause functional disorders and damages, especially in leaves, thus affecting plant metabolism (Nobre et al., 2013). Thus, salinity can be considered as one of the abiotic stresses that most limits plant growth (Pedrotti et al., 2015). Biochemical and physiological processes are triggered in plants submitted to saline stress, which consequently interfere with their growth, stomatal behavior, and photosynthetic capacity (Nunkaewa et al., 2014), due to the osmotic effects of the salts and the specific characteristics of Na+ and Cl− (Graciano et al., 2011). Qiu et al. (2014) reported that saline stress, specifically, promotes the inhibition of the chlorophyllase enzyme, which results in a reduction in the levels of chlorophyll and its precursor, 5-aminolevulinate.
Despite continuous weekly exposure to irrigation containing more salinity than is ideal, most palms across genera and species continued to show signs of growth and development. All plants increased in average height, leaf count, and chlorophyll, when compared to baseline data (0 DAT). However, there was a decline in health markers the closer the experiment came to the 6-month mark. This can only lead to the conclusion that prolonging exposure would lead to eventual decline of most if not all plants, given sufficient time and continued salt exposure, mainly those exposed to the highest concentrations.
Furthermore, it is important to point out that the specie R. regia was very susceptible to salinity, as the plants exposed to the treatments of 25 and 50 ppt died at 90 days after treatment, and the palm S. palmetto is more susceptible to salinity than the T. radiata and P. sargentii palms.
This data is important in driving forward informed decision-making and proper technology adoption for conservation horticulture, horticultural production, landscaping, and botanic gardens in South Florida. As anthropogenic activities continue to alter natural systems, the consequences of sea-level rise and saltwater intrusion should be anticipated for and responded to in an informed and data-driven approach.
The growth parameters and chlorophyll content are reduced over time, particularly at higher salt concentrations for all palm species studied. The chlorophyll content by SPAD and the normalized difference vegetation index (NDVI) can be used as health indicators of the palm species studied, as it was observed in this study a decline in the chlorophyll content as there was an increase in salt concentrations. It is important to emphasize that leaf chlorosis and subsequent decline of palm species were observed in the treatment with higher salt concentration, thus highlighting the importance of chlorophyll content as an indicator of plant health. The specie R. regia was very susceptible to salinity, and the palm S. palmetto is more susceptible to salinity than the T. radiata and P. sargentii palms.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
AK and MG: conceptualization, supervision, and project administration. JF: methodology. BS: software and formal analysis. JF: investigation. BS and JF: writing—original draft preparation. AK, MG, and BS: writing—review and editing. AK: funding acquisition. All authors contributed to the article and approved the submitted version.
Funding for this project was provided by the Hispanic Serving Institutions Higher Education Grants Program 2016-38422-25549 from the USDA National Institute of Food and Agriculture.
We would like to thank Montgomery Botanical Center (MBC) for providing palm seedlings, materials, and space. In addition to MBC’s greenhouse manager, Vickie Murphy and Xavier Gratacos, Superintendent for facilitating resources and support throughout the experiment.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Ait-El-Mokhtar, M., Fakhech, A., Ben-Laouane, R., Anli, M., Boutasknit, A., Ait-Rahou, Y., et al. (2022). Compost as an eco-friendly alternative to mitigate salt-induced effects on growth, nutritional, physiological and biochemical responses of date palm. Int. J. Recycl. Org. Waste Agric. 11, 85–100. doi: 10.30486/IJROWA.2021.1927528.1233
Basyouni, R., and Dunn, B. (2013). Use of reflectance sensors to monitor plant nitrogen status in horticultural plants. Oklahoma Cooperative Extension ServiceHLA-6719, 1–4. Available at: https://extension.okstate.edu/fact-sheets/print-publications/hla/use-of-optical-sensors-to-monitor-plant-nitrogen-status-in-horticultural-plants-hla-6719.pdf
Bayabil, H. K., Li, Y., Tong, Z., and Gao, B. (2021). Potential management practices of saltwater intrusion impacts on soil health and water quality: a review. J. Water Clim. Change. 12, 1327–1343. doi: 10.2166/wcc.2020.013
Braun de Torrez, E. C., Frock, C. F., Boone, W. W., Sovie, A. R., and McCleery, R. A. (2021). Seasick: why value ecosystems severely threatened by sea-level rise? Estuar. Coasts 44, 899–910. doi: 10.1007/s12237-020-00850-w
Broschat, T. K., and Black, R. J. (2021). Ornamental palms for South Florida. Enh21/EP009:Ornamental Palms for South Florida. University of Florida. Available at: https://edis.ifas.ufl.edu/publication/EP009
Fehr, V., Buitenwerf, R., and Svenning, J. (2020). Non-native palms (Arecaceae) as generators of novel ecosystems: a global assessment. Divers. Distrib. 26, 1523–1538. doi: 10.1111/ddi.13150
Ferreira, D. F. (2011). Sisvar: a computer statistical analysis system. Cienc. Agrotec. 35, 1039–1042. doi: 10.1590/S1413-70542011000600001
Graciano, E. S. A., Rejane, J. M. C., Nogueira, R. J. M. C., Lima, D. R. M., Pacheco, C. M., and Santos, S. R. C. (2011). Crescimento e capacidade fotossintética da cultivar de amendoim BR 1 sob condições de salinidade. Rev. Bras. Eng. Agric. Ambient. 15, 794–800. doi: 10.1590/S1415-43662011000800005
Griffith, M. P., Barber, G., Lima, J. T., Barros, M., Calonje, C., Noblick, L. R., et al. (2017). Plant collection “half life:” can botanic gardens weather the climate? Curator. 60, 395–410. doi: 10.1111/cura.12229
Inman, D., Khosla, R., and Mayfield, T. (2005). On the go active remote sensing for efficient crop nitrogen management. Sens. Rev. 25, 209–214. doi: 10.1108/02602280510606499
Nascimento, E. C. S., do Nascimento, R., da Silva, A. A. R., de Castro Bezerra, C. V., Batista, M. C., de Sá Almeida Veloso, L. L., et al. (2019). Growth and photosynthetic pigments of cotton cultivars irrigated with saline water. Agric. Sci. 10, 81–91. doi: 10.4236/as.2019.101007
Nobre, R. G., Lima, G. S., Gheyi, H. R., Lourenço, G. S., and Soares, L. A. A. (2013). Emergência, Crescimento e produção da mamoneira sob estresse salino e adubação nitrogenada. Rev. Cienc. Agron. 44, 76–85. doi: 10.1590/S1806-66902013000100010
Nunkaewa, T., Kantachote, D., Kanzaki, H., Nitoda, T., and Ritchie, R. J. (2014). Effects of 5-aminolevulinic acid (ALA)-containing supernatants from selected Rhodopseudomonas palustris on rice growth under NaCl stress, with mediating effects on chlorophyll, photosynthetic electron transport and antioxidative enzymes. Electron. J. Biotechnol. 17, 19–26. doi: 10.1016/j.ejbt.2013.12.004
Oliveira, H., do Nascimento, R., Silva, S., Cardoso, J. A. F., Guimarães, R. F. B., and Nascimento, E. C. S. (2018). Initial growth and gas exchanges of plants of colored cotton submitted to saline stress. Agric. Sci. 9, 1652–1663. doi: 10.4236/as.2018.912115
Pedrotti, A., Chagas, R. M., Ramos, V. C., Prata, A. P. M., Lucas, A. A. T., and Santos, P. B. (2015). Causas e consequências do processo de salinização dos solos. Rev. Eletr. Gest. Educ. Tecnol. Amb. 19, 1308–1324. doi: 10.5902/2236117016544
Perry, L., and Williams, K. (1996). Effects of salinity and flooding on seedlings of cabbage palm (Sabal palmetto). Oecologia 105, 428–434. doi: 10.1007/BF00330004
Qiu, Z. B., Guo, J., Jhu, A. J., Zhang, L., and Zhang, M. M. (2014). Exogenous Jasmonic acid can enhance tolerance of wheat seedlings to salt stress. Ecotoxicol. Environ. Saf. 104, 202–208. doi: 10.1016/j.ecoenv.2014.03.014
Sim, C. C., Zaharah, A. R., Tan, M. S., and Goh, K. J. (2015). Rapid determination of leaf chlorophyll concentration, photosynthetic activity and NK concentration of Elaiesguineensis via correlated SPAD-502 chlorophyll index. Asian J. Agric. Res. 9, 132–138. doi: 10.3923/ajar.2015.132.138
Yaish, M. W., and Kumar, P. P. (2015). Salt tolerance research in date palm tree (Phoenix dactylifera L.), past, present, and future perspectives. Front. Plant Sci. 6:348. doi: 10.3389/fpls.2015.00348
Keywords: Arecaceae, palms, saline stress, chlorophyll content, NDVI
Citation: Khoddamzadeh AA, Flores J, Griffith MP and Souza Costa BN (2023) Saltwater intrusion ecophysiological effects on Pseudophoenix sargentii, Roystonea regia, Sabal palmetto “Lisa,” and Thrinax radiata in South Florida. Front. Ecol. Evol. 11:1127679. doi: 10.3389/fevo.2023.1127679
Edited by:
Frank S. Gilliam, University of West Florida, United StatesReviewed by:
Hussein J. Shareef, University of Basrah, IraqCopyright © 2023 Khoddamzadeh, Flores, Griffith and Souza Costa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Amir Ali Khoddamzadeh, YWtob2RkYW1AZml1LmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.