 Madeleine Scott
Madeleine Scott W. Douglas Robinson
W. Douglas Robinson
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 31 January 2023
Sec. Behavioral and Evolutionary Ecology
Volume 11 - 2023 | https://doi.org/10.3389/fevo.2023.1092323
Mobbing is a widespread but risky behavior where prey actually approach and may even attack their predators. Small songbirds gather in multi-species flocks to direct mobbing at birds of prey such as the Northern Pygmy-Owl (Glaucidium gnoma), a small diurnal species of owl in mountainous western North America. We hypothesized that intensity of mobbing behavior would vary according to risk to songbirds and that risk varied seasonally and across elevations. In particular, we expected mobbing intensity to change seasonally as a function of two primary sources of risk: seasonal dietary changes of pygmy-owls and distribution of pygmy-owls as a function of elevation. Pygmy-owls are known to shift their diet from mostly mammals in early spring to birds in early summer, so we hypothesized risk of predation would vary in parallel. Likewise, risk was expected to vary as the distribution of pygmy-owls was thought to vary seasonally across elevations. To elicit mobbing in a controlled manner, we conducted owl call playback experiments in the Oregon Coast Range mountains, United States. Overall, we found mobbing to be rare, observing it in 8.1% of 663 experiments. The frequency of mobbing did not appear to vary across elevations. Instead, mobbing varied mostly by season. The frequency of mobbing was highest during fall at lower elevations. Mobbing was rarest (1.3% of trials) during winter. Mobbing occurred during seasons when pygmy-owls tend to consume more birds, consistent with the hypothesis that seasonal variation in risk influences mobbing. In our study, mobbing happened when risk was higher. However, the increased risk may be offset by the influence of mobbing group size. Mobbing was more likely to occur at sites where the number of songbirds, especially chickadees (Poecile sp.) and nuthatches (Sitta sp.), was larger. Yet, even when songbird numbers were high, mobbing was rare during winter, suggesting an interaction between risk and energetic costs of mobbing. Additional experiments manipulating food resource availability may be able to reveal the strength and influence of the interaction on mobbing decisions.
Life is dangerous for small birds. Many fall victim to predators, leading to the evolution of behavioral strategies that may reduce risks of predation (Lima, 1993). Although most strategies involve hiding or fleeing from predators, mobbing is a paradoxical behavioral strategy where prey actually approach predators. Mobbing is when “birds of one or more species assemble around a stationary or moving predator (potentially dangerous animal), change locations frequently, perform (mostly) stereotyped wing and/or tail movements and emit loud calls” (Curio, 1978). Mobbing makes prey more conspicuous but may reduce risk of predation by alerting conspecifics to the presence of danger (Curio et al., 1978; Hurd, 1996), as well as relaying information about the predator’s location (McPherson and Brown, 1981). Predator harassment may also prevent or reduce future surprise attacks (Flasskamp, 1994; Templeton and Greene, 2007). Since mobbing usually happens in large groups, the risk of being singled out by a predator is reduced (Solheim, 1984).
Songbirds mob Northern Pygmy-Owls (Glaucidium gnoma; Solheim, 1984), a small, diurnal owl species of western North America. Yet, the behavioral dynamics of mobbing assemblages, including which species initiate and participate in mobbing events, and the factors that influence occurrence of mobbing have not been described well. A significant part of the Northern Pygmy-Owl’s diet is composed of small bird species, including species that partake in mobbing behavior (Altmann, 1956; McPherson and Brown, 1981; Holt and Leroux, 1996). In studies of the congeneric Eurasian Pygmy-Owl (Glaucidium passerinum), songbird species comprising a larger portion of the owl’s diet had a higher propensity for mobbing than those that were never or rarely consumed (Dutour et al., 2017). Given that prey species can distinguish when they are the target of predation (Hamerstrom, 1957), and which predator species are most likely to attack them (Motta-Junior and Santos-Filho, 2012; Carlson et al., 2017), songbird mobbing of Northern Pygmy-Owls is inherently dangerous (Sordahl, 1990).
Prey assess changes in predation risk, which are expected to vary temporally and spatially (Lima and Bednekoff, 1999). Predation risk to songbirds from Northern Pygmy-Owls may vary seasonally. The dominant dietary items of Northern Pygmy-Owls switch seasonally between birds and small mammals in the Oregon Coast Range (Deshler, 2020). The proportion of small birds relative to small mammals in Northern Pygmy-Owl diets almost doubles from spring to summer, making birds the primary food source in summer presumably because of the increasing availability of fledged offspring. This seasonal variation in diet suggests that mobbing behavior may also vary seasonally in its occurrence or intensity if songbirds are sensitive to such changes in predation risk. Mobbing of Eurasian Pygmy-Owls was also correlated with dietary preference, occurring more frequently in fall by those species frequently preyed upon (Dutour et al., 2019). Timing of the Northern Pygmy-Owl diet shift aligns with the owl’s breeding season, which begins with the selection of nesting territories in March and April, followed by incubation in April and May, and feeding of nestlings in the summer (Sater et al., 2006). We hypothesized that songbirds could be aware of this seasonal change in risk from Northern Pygmy-Owls and alter their propensity for mobbing. We predicted that mobbing would be most likely to occur when the risk is higher, so mobbing behavior should vary seasonally in accordance with owl dietary shifts.
The seasonal variation in risk also has a spatial component. In mountainous areas, bird abundance and, potentially, occurrences of Northern Pygmy-Owls may vary across elevations (Boyle, 2017). If awareness in songbirds of predation risk correlates with the rate at which pygmy-owls are encountered across elevations and seasons, then we expect parallel responses in mobbing behavior. Altogether, we hypothesized that mobbing of Northern Pygmy-Owls by songbirds would be highest in seasons when owl diets included more birds, when numbers of songbirds available to join mobbing assemblages were highest, and at elevations where owls were seasonally present.
Natural mobbing events are uncommonly observed, and pygmy-owls are usually ambush predators, attacking prey from a hidden location, so we stimulated mobbing behavior by playing recordings of Northern Pygmy-Owls. The use of playbacks to elicit mobbing responses was successfully done with Eurasian Pygmy-Owls and Ferruginous Pygmy-Owls (G. brasilianum), without the use of an owl decoy (Sandoval and Wilson, 2012; Dutour et al., 2016). We conducted playback trials with Northern Pygmy-Owl calls to: (1) characterize the species assemblage participating in mobbing events; (2) measure the influence of numbers of songbirds on the occurrence of mobbing events; and (3) quantify seasonal and altitudinal differences in occurrence of mobbing.
We conducted experiments in western Oregon, United States, near Corvallis and in the nearby Coast Range Mountains spanning elevations from 80 to 1,220 m. To replicate sites with the highest elevations, we included all three of the highest peaks (Marys Peak, Grass Mountain, and Prairie Mountain) available in our study area (all above 1,000 m). Across the elevational gradient, we chose study sites based on stratified sampling. We first selected three elevation zones: low (0–299 m), mid (300–999 m), and high (above 1,000 m). We then used the Public Land Survey System, which grids the state into one-square-mile sections, and randomly selected 10 one-square-mile sections in each elevation zone. After that, we inspected each section for accessibility (publicly accessible road or trail had to be present). If access was not possible, we chose the next nearest section that did have access and was within the same elevation zone. We supplemented our sample sizes with additional convenience sampling outside of the original sections in surrounding areas to ensure inclusion of the entire elevational gradient across the study area. Elevation was measured (+/− 5 m accuracy) at each location with a handheld GPS unit. We sampled a variety of wooded habitat types, but the focus was on mixed coniferous-deciduous forests, the known habitat preference of Northern Pygmy-Owls (Sater et al., 2006).
We played calls of Northern Pygmy-Owls to prompt songbirds to respond to standardized stimuli presented across the study area. Northern Pygmy-Owl advertising calls vary little among individuals, aside from minor pitch differences in males vs. females (Holt and Petersen, 2020). Owls are widespread across our study area and routinely call for long bouts, sometimes calling for 30–60 min. They are known to call at all seasons (Noble, 1990). We used two recordings we made in the study area and two recordings of owl calls from Oregon we downloaded from xento-canto.org. We ensured that recordings had no alarm calls of songbirds in the background. Calls were chosen for playback at each site in a non-systematic fashion. Since songbirds in the forest understory and subcanopy have been observed mobbing Northern Pygmy-Owls, we positioned a portable speaker (Bluetooth Boom 3) mounted on a 3.3-m pole in vegetation. After noting the date, start time, latitude, longitude, and elevation, we began each playback trial by taking an initial count of the number of birds of all species seen and heard in the area (within 50 m) for 3 min. We then played the owl calls for 1 min. In our preliminary evaluations, the owl calls were audible to humans to at least 100 m. After the first 1-min playback was completed, we again counted the birds of each species in the area for 3 min. We also noted when vocalizations increased or intensified, if birds moved toward the playback speaker and, if they did, if mobbing behavior occurred. We considered approach to the speaker within 5 m combined with the stereotypical wing-flicking and increased intensity of vocalizations as mobbing (Curio, 1978; Curio et al., 1978). We scored each species’ behavior. We then conducted an additional 1-min playback, followed by a subsequent 3-min silent period where we noted the same variables and scored each species’ behavior. Our preliminary data indicated that songbirds normally responded quickly to the 1-min playback but, on occasion, responded only to the second 1-min playback. If we detected evidence of mobbing after the whole 11-min trial period was completed, we further characterized the mobbing patterns by playing an additional 3 min of the owl calls. When no mobbing behaviors were detected during the 11-min trial, we did not conduct the subsequent 3 min of additional playback.
It was not possible to score behavior of each individual bird because no birds were individually marked. If a Northern Pygmy-Owl appeared or was heard during a playback trial, we noted its presence, and whether the songbirds directed their movements toward the speaker or the owl. We conducted our trials year-round to characterize seasonal changes in behavior. For analyses and presentation of results, we divided the year into four seasons (spring: March–May, summer: June–August, fall: September–November, and winter: December–February). Most sites (n = 351) were visited just once during the study. Others were visited twice (n = 117), three (n = 65), or four times (n = 41). Some sites were visited up to three times per season and as many as 12 times during the 2-year study but such numbers of multiple visits were rare (n = 88 playback experiments total with 5–12 visits at a site). Prior to data analyses, we ensured that habituation to playbacks was not an issue, finding that number visits and mobbing response were unrelated statistically (r2 = 0.02).
To analyze predictors of occurrence of mobbing, we used logistic regression to quantify odds of mobbing occurring as a function of season and elevation in one model and number of songbirds present in another model. Because mobbing was uncommon (n = 54 of 663 trials), the number of variables our models could support was limited. Mobbing was considered a binary response variable, and season and elevation were included as categorical variables. Elevation was categorized as low (0–299 m above sea level), mid (300–999 m), or high (1,000 m and above). An interaction of season and elevation was excluded because of model overfitting problems. We excluded winter from the model because mobbing was extremely rare (1.3% of trials) and model convergence failed when winter data were included. We created an additional model examining mobbing as a response with number of birds and season (again excluding winter) as fixed-effect variables. To determine the number of birds present, we restricted the species set to those known to participate in mobbing (Altmann, 1956, personal observation; Table 1). We calculated the number of each species as the maximum count within any of the three 3-min observation periods within each trial. Because owl calls stimulate the conspicuousness and therefore the availability of the species of interest, we did not adjust our counts for potential detectability issues. We evaluated the potential influence on mobbing behavior of real Northern Pygmy-Owls stimulated by our playbacks to vocalize. When we compared trials where real owls were and were not detected we found minimal differences so ignored the presence or absence of owls for subsequent analyses. All analyses were performed with R (R Core Team, 2020).
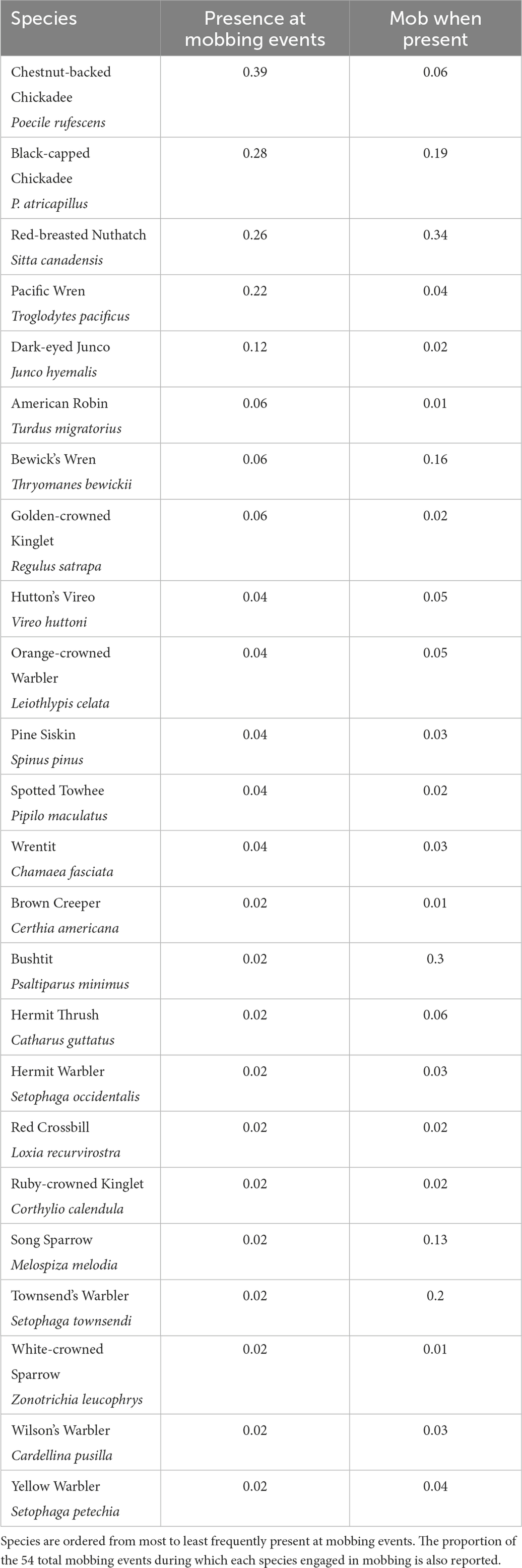
Table 1. All species observed to participate in mobbing events (N = 54) in response to playbacks (N = 663) of calls of Northern Pygmy-Owls.
Mobbing in response to Northern Pygmy-Owl call playbacks was relatively rare. We observed birds approaching to within 5 m of the playback speaker while engaging in typical mobbing behaviors in 8.1% (N = 54) of 663 trials. In seven trials, bird behavior did not indicate mobbing according to our definition, but an increase in vocalization rates and movement toward the speaker caused us to attempt the additional 3-min playback period to determine if mobbing might subsequently occur; it did not.
The species composition of mobbing assemblages was diverse, including at least 24 species of songbird (Table 1). All were small songbird species of woodland and woodland edge habitats. The largest species participating in mobbing was American Robin (Turdus migratorius), which averages 75 g. Larger songbirds that commonly mob large owl species did not mob Northern Pygmy-Owls (e.g., Aphelocoma and Cyanocitta jays). The most commonly observed participants in mobbing events were small (<30 g), including the two chickadee species (Poecile atricapillus and P. rufescens) and Red-breasted Nuthatch (Sitta canadensis). Pacific Wrens (Troglodytes pacificus) and Dark-eyed Juncos (Junco hyemalis) were the two additional species regularly detected at mobbing events. We detected the presence of those five species in 12–39% of all mobbing events. The remaining species were uncommonly present (<6% of events) during mobbing.
Occurrence of mobbing peaked in late summer and in fall when the highest proportion of trials eliciting mobbing occurred during October (23%; Supplementary Table 1). It was rare in spring 2020 and winter 2021. We observed mobbing most often at mid and lower elevations (Table 2) during the summer and fall when mobbing occurred in 10–31% of trials. Mobbing was most frequently observed (31% of trials) at low elevation during fall. The lowest frequency of mobbing behavior was detected during winter (two of 148 trials, 1.3%) and also during spring, especially at middle elevations (2%).
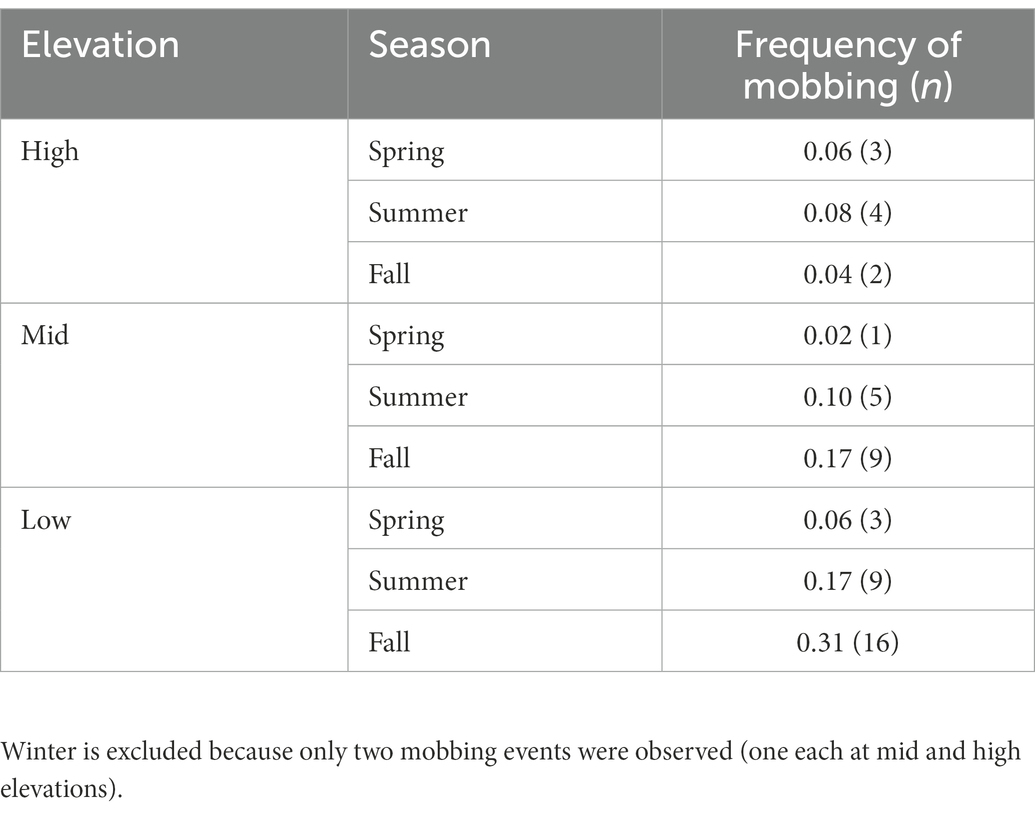
Table 2. Proportion of experiments (n = 663) where mobbing was observed (number of mob events; n = 54 total) across the three elevation zones (high ≥ 1,000 m; mid = 300–999 m; and low < 300 m) and three seasons.
The mean number of individual birds present during a mobbing event was 12.8 (±4.8 SD) compared to the mean number across all trials of 8.1 (±6.6). In general, number of songbirds was higher in spring and summer at high elevations and peaked at low elevations in winter but numbers of birds were highly variable (Table 3).
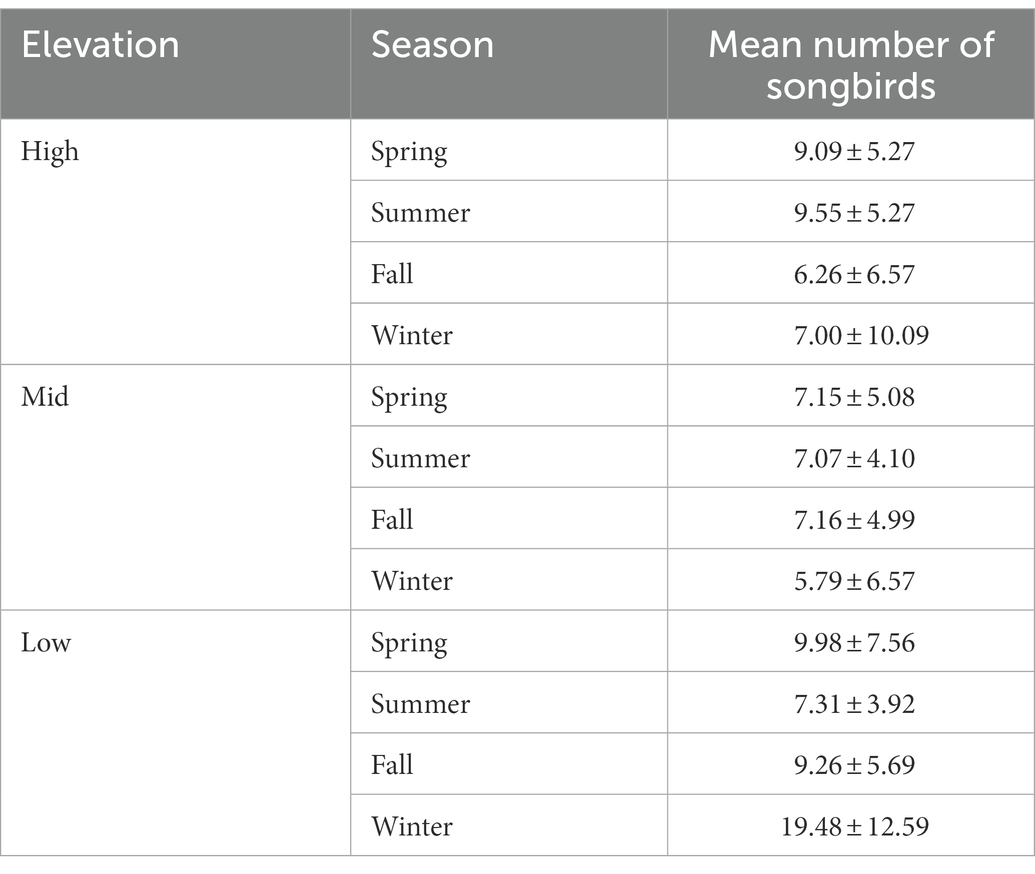
Table 3. Mean (±SD) number of songbirds across elevational zones and season.
Numbers tended to be lowest at mid-elevation sites during winter. The predicted probability of mobbing was related to the number of songbirds we detected and season. The smallest numbers predicted for mobbing to occur were during fall and the largest were during spring. Rates predicted for summer overlapped those of both spring and fall. Overall, for every increase of an additional songbird, the odds of mobbing increased 1.19 times (CI 1.13–1.26; Figure 1). Mobbing was more likely to occur when numbers of chickadees and nuthatches, in particular, were larger (Figure 2).
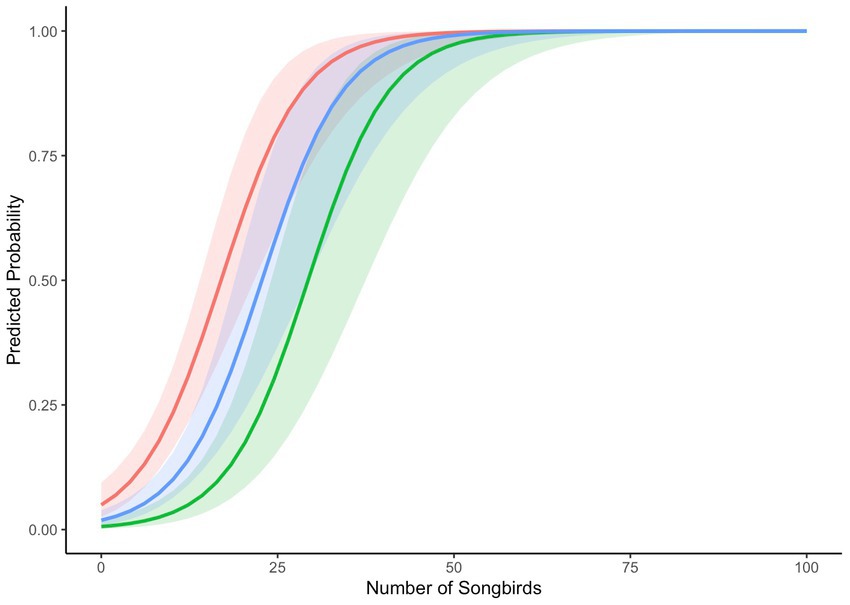
Figure 1. Predicted probabilities of mobbing given the total number of participating individual birds across seasons (fall=red, summer=blue, spring=green). Winter was not included in the model. CIs (95%) are shown around each seasonal prediction.
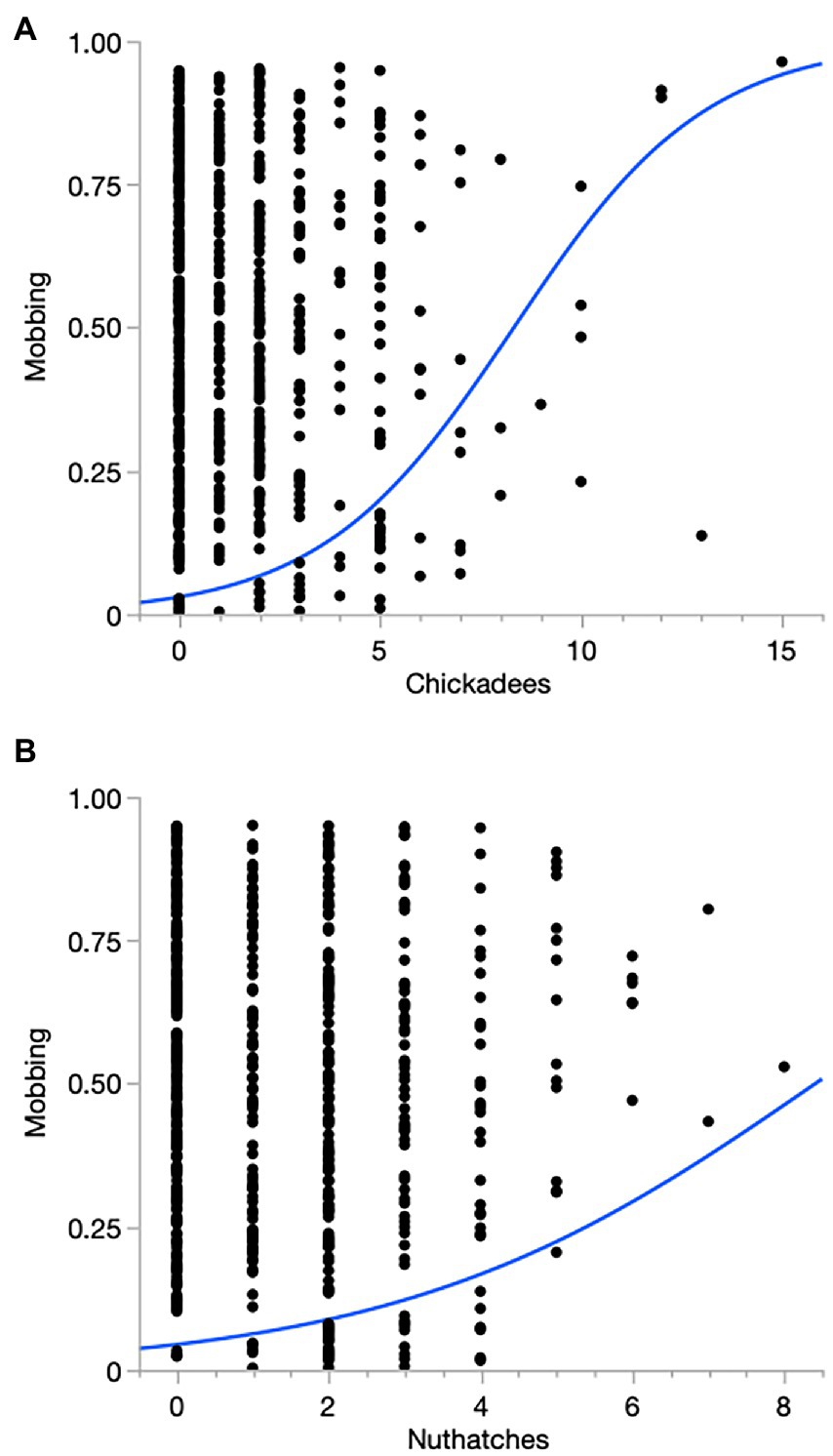
Figure 2. Relationships between numbers of (A) chickadees (Poecile atricapillus and P. rufescens) and (B) Red-breasted Nuthatches (Sitta canadensis). Logistic fits: Chickadees (R2 = 0.158, Chi-squared = 59.3, df = 1, p < 0.0001); Nuthatches (R2 = 0.05, Chi-squared = 17.6, df = 1, p < 0.0001).
The relationship between season and elevation showed that the highest probability of a mobbing event occurring was during fall at low elevations (Figure 3). The odds of mobbing at lower elevations in fall averaged 7.65 (CI 1.70–34.42, p = 0.0044) times more likely than in spring, and 2.78 (CI 0.90–8.63, p = 0.0864) times more likely than in summer. At our mid-elevation range, the odds of mobbing in fall were 9.56 (CI 1.15–79.49, p = 0.0335) times more likely than in spring, and 3.63 (CI 0.96–13.78, p = 0.0603) times more likely than in summer. At higher elevations, we did not see significant differences in the likelihood of mobbing between seasons. Across all elevations, there was not a significant difference in the odds of mobbing between spring and summer.
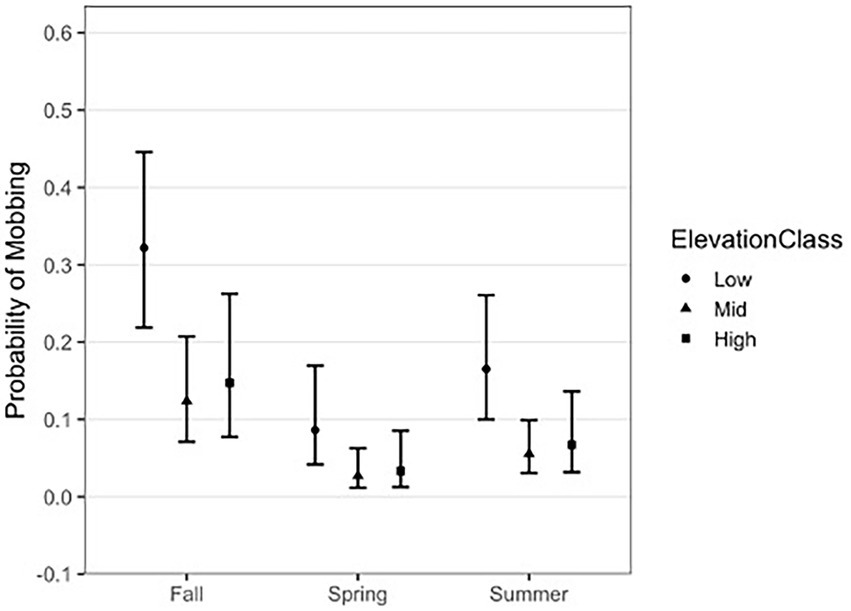
Figure 3. Mean probabilities (95% CIs) of mobbing behavior across three elevational zones for three seasons in the Oregon Coast Range. Winter is excluded because mobbing was rarely detected.
Mobbing of Northern Pygmy-Owl calls by songbirds was most likely to occur during fall at lower elevations. Two dozen species were detected at mobbing events. The most frequent participants were two species of chickadee, Red-breasted Nuthatch, Pacific Wren, and Dark-eyed Junco. Presence and activities of those five species appeared to strongly influence the formation of mobbing events, a similar outcome to controlled experiments in Germany revealing that a few core species (also mostly tits and nuthatches) responded to heterospecific mobbing alarm calls while other mobbing species were much less responsive (Randler and Vollmer, 2013). All five of the core species in our study are common and their numbers influenced the formation of mobs. Numbers during fall, when many young of the year are typically present on the landscape, were generally higher than numbers at other seasons but we saw substantial variability in numbers at all seasons. Greater numbers of birds are expected to dilute the risk to each individual during mobbing (Solheim, 1984), offering a potential explanation for its prevalence during fall in our study area. The occurrence of mobbing may also be influenced by energetic considerations. Avian behavioral decisions, such as foraging, breeding, or searching for predators, have been explained by the costs and benefits to overall fitness (Lima, 1993; Roth and Lima, 2007). The peak number of birds we observed during our trials occurred at low elevations during winter, when we observed no mobbing. This suggests that, despite possibilities for dilution of risk to individuals in large winter assemblages, mobbing may not occur because its occurrence may be a function of both number of participants and environmental conditions that permit expenditure of energy on mobbing. Mobbing is energetically costly (Sordahl, 1990), and individuals need the resources to be able to support this expenditure. We assume resources are less prevalent in winter. A potential experimental manipulation to evaluate this assumption could involve food supplementation. Four species in our set of five that mob most often also regularly attend bird feeders. If energetic costs reduce mobbing because of food shortages in winter, mobbing might increase if food were made more readily available. Although we know of no such experimental results yet, experiments have verified that very small weight gains in small songbirds influence their behavioral decisions in situations where predators are present (Lima, 1988).
Results from our playback trials with Northern Pygmy-Owls and from similar studies with Eurasian Pygmy-Owls (Dutour et al., 2019) and Eastern Screech-Owls (Megascops asio; Shedd, 1983) showed seasonal changes in mobbing behavior. Both studies revealed a higher prevalence of mobbing during fall. The seasonal pattern of mobbing may be driven by seasonal changes in bird numbers on the landscape, which is probably largely a result of more juveniles being present after the summer breeding season (Shedd, 1982) and as information about predator identity is transmitted from adults to fledged offspring before the young disperse from their home territory (Curio et al., 1978). In addition, songbirds may perceive a shift in predation risk as Northern Pygmy-Owls shift dietary preference from small mammals to small birds during summer (Deshler, 2020). If this seasonal pattern of dietary shifts is inconsistent from year to year or differs between geographic locations, alternative assessments of risk by songbirds may occur. Prey are known to alter the intensity of mobbing behavior in relation to their perceived risk (Sandoval and Wilson, 2012; Dutour et al., 2016, 2017). We found that mobbing occurred during seasons when the risk to songbirds was higher, but it was more likely to occur within that timeframe if the number of birds present was greater and therefore risk to an individual could be diluted within the larger mobbing group.
An important caveat is that we did not see consistent patterns between all seasons of different years. In 2021, mobbing occurred more frequently and began earlier in the spring than in 2020 when we primarily detected mobbing only in late summer and fall. Possible explanations for this annual difference could involve changes in bird numbers, although we saw minor variation in those numbers during our trials. Alternatively, variable amounts of food resources could influence songbird behaviors that are energetically costly. Our data included wide confidence intervals around the odds ratios suggesting large variability and the potential for minimal biologically relevant differences in some cases. However, the odds of mobbing occurring in fall, especially at lower elevations, appears to be a robust result. Additional trials are needed to verify seasonal differences outside of fall, but similar seasonal patterns were shown in recent studies of other Glaucidium owls (Dutour et al., 2016, 2019).
Across most seasons, we observed a higher occurrence of mobbing at lower elevations. We expected to see an alignment of mobbing with hypothesized seasonal altitudinal movements of pygmy-owls where mobbing was expected to increase at higher elevations during the warmer months and at lower elevations during the cooler months. However, we did not detect seasonal shifts in the altitudinal distribution of pygmy-owls (Scott, 2021). Similarly, our playback trials did not indicate an increase in the occurrence of mobbing at higher elevations during the warmer months. Instead, the frequency of mobbing remained relatively concordant with the frequency of owl detections. We conclude that minimal differences in the frequency of mobbing occur across elevations within our study area.
The composition of species participating in mobbing events in our study area contained two dozen species but was dominated by five species. Relatives of those same species also dominate mobbing events directed at pygmy-owls in Europe (Altmann, 1956; Dutour et al., 2016). The groups primarily mobbing pygmy-owls are small songbirds, such as parids, nuthatches, thrushes, warblers, wrens, and finches. Within our study, those species that were most frequently observed to mob Northern Pygmy-Owls comprise a high percentage of their avian diet, including Pacific Wrens, warblers, sparrows, chickadees, and Red-breasted Nuthatches (Deshler, 2020). The small size of mob participants, relative to the size of pygmy-owls, and the consistency of the types of birds mobbing in North America and Europe, aligns with the hypothesis that species at risk of being prey are the most likely to expend energy on mobbing. Larger species that commonly mob larger owls or hawks, such as thrushes and jays, rarely mob pygmy-owls and have rarely been observed as prey of pygmy-owls.
Although additional experimentation is still required to exclude competing hypotheses such as cultural transmission to related offspring, our playback experiments revealed patterns of behavior indicating that (1) birds regularly involved in mobbing are also likely to be prey of Northern Pygmy-Owls; (2) birds were more likely to mob when larger numbers of songbirds are present to participate in such mobs; and (3) numbers of birds alone were not the exclusive predictor of the occurrence of mobbing because mobbing was rare to absent during winter at low elevations when we found the largest number of potential mobbers within our study area. Additional experiments evaluating strength of mobbing responses when calls are augmented with owl decoys or real owls are also needed. Although previous studies (Sandoval and Wilson, 2012; Dutour et al., 2016, 2019) found similarly strong responses to call playbacks alone, the connection of visual stimuli with the perception of risk in songbirds may produce different behavioral responses.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethical review and approval was not required for the animal study because our study was observational. We used brief (2 min) playbacks of owl calls to attempt to elicit mobbing behavior from songbirds. According to Guidelines to the Use of Wild Birds in Research maintained by the American Ornithological Society, animal care permits are not required for playback experiments such as ours.
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
We thank N. Clements, J. Hedges, for their assistance with observations. M. Landys, J. Armstrong provided helpful guidance during the study and comments on earlier versions of the manuscript. This work was part of a graduate thesis (Scott, 2021).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1092323/full#supplementary-material
Altmann, S. A. (1956). Avian mobbing behavior and predator recognition. Condor 58, 241–253. doi: 10.2307/1364703
Boyle, W. A. (2017). Altitudinal bird migration in North America. Auk 134, 443–465. doi: 10.1642/AUK-16-228.1
Carlson, N. V., Healy, S. D., and Templeton, C. N. (2017). Hoo are you? Tits do not respond to novel predators as threats. Anim. Behav. 128, 79–84. doi: 10.1016/j.anbehav.2017.04.006
Curio, E. (1978). The adaptive significance of avian mobbing. Z. Tierpsychol. 48, 175–183. doi: 10.1111/j.1439-0310.1978.tb00254.x
Curio, E., Ernst, U., and Vieth, W. (1978). Cultural transmission of enemy recognition: one function of mobbing. Science 202, 899–901. doi: 10.1126/science.202.4370.899
Deshler, J. F. (2020). Northern pygmy-owl (Glaucidium gnoma) nesting ecology in northwestern Oregon. Wilson J. Ornithol. 132, 352–365. doi: 10.1676/1559-4491-132.2.352
Dutour, M., Cordonnier, M., Léna, J.-P., and Lengagne, T. (2019). Seasonal variation in mobbing behaviour of passerine birds. J. Ornithol. 160, 509–514. doi: 10.1007/s10336-019-01630-5
Dutour, M., Lena, J.-P., and Lengagne, T. (2016). Mobbing behaviour varies according to predator dangerousness and occurrence. Anim. Behav. 119, 119–124. doi: 10.1016/j.anbehav.2016.06.024
Dutour, M., Lena, J.-P., and Lengagne, T. (2017). Mobbing behaviour in a passerine community increases with prevalence in predator diet. Ibis 159, 324–330. doi: 10.1111/ibi.12461
Flasskamp, A. (1994). The adaptive significance of avian mobbing V. an experimental test of the ‘move on’ hypothesis. Ethology 96, 322–333. doi: 10.1111/j.1439-0310.1994.tb01020.x
Hamerstrom, F. (1957). The influence of a Hawk’s appetite on mobbing. Condor 59, 192–194. doi: 10.2307/1364723
Holt, D. W., and Leroux, L. A. (1996). Diets of northern pygmy-owls and northern saw-whet owls in west-Central Montana. Wilson Bull. 108, 123–128.
Holt, D. W., and Petersen, J. L. (2020). “Northern pygmy-owl (Glaucidium gnoma)” in Birds of the World. eds. A. F. Poole and F. B. Gill (Ithaca, New York: Cornell Lab of Ornithology)
Hurd, C. R. (1996). Interspecific attraction to the mobbing calls of black-capped chickadees (Parus atricapillus). Behav. Ecol. Sociobiol. 38, 287–292. doi: 10.1007/s002650050244
Lima, S. L. (1988). Initiation and termination of daily feeding in dark-eyed juncos: influences of predation risk and energy reserves. Oikos 53:3. doi: 10.2307/3565656
Lima, S. L. (1993). Ecological and evolutionary perspectives on escape from predatory attack: a survey of north American birds. Wilson Bull. 105, 1–47.
Lima, S. L., and Bednekoff, P. A. (1999). Temporal variation in danger drives antipredator behavior: the predation risk allocation hypothesis. Am. Nat. 153, 649–659. doi: 10.1086/303202
McPherson, R., and Brown, R. (1981). Mobbing responses of some passerines to the calls and location of the screech owl. Raptor Res. 15, 23–30.
Motta-Junior, J. C., and Santos-Filho, P. D. S. (2012). Mobbing on the striped owl (Asio clamator) and barn owl (Tyto alba) by birds in Southeast Brazil: do owl diets influence mobbing? Ornitol. Neotrop. 23, 159–168.
Noble, P. L. (1990). Distribution and density of owls at Monte Bello open space preserve, Santa Clara County, California. Western Birds 21, 11–16.
R Core Team (2020). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Randler, C., and Vollmer, C. (2013). Asymmetries in commitment in an avian communication network. Naturwissenschaften 100, 199–203. doi: 10.1007/s00114-013-1009-6
Roth, T. C., and Lima, S. L. (2007). The predatory behavior of wintering accipiter hawks: temporal patterns in activity of predators and prey. Oecologia 152, 169–178. doi: 10.1007/s00442-006-0638-2
Sandoval, L., and Wilson, D. R. (2012). Local predation pressure predicts the strength of mobbing responses in tropical birds. Curr. Zool. 58, 781–790. doi: 10.1093/czoolo/58.5.781
Sater, D., Forsman, E., Ramsey, F., and Glenn, E. (2006). Distribution and habitat associations of northern pygmy-owls in Oregon. J. Raptor Res. 40, 89–97. doi: 10.3356/0892-1016(2006)40[89:DAHAON]2.0.CO;2
Scott, M. (2021). Songbird mobbing of northern pygmy-owls: a function of risk across landscapes and seasons. Available at: https://ir.library.oregonstate.edu/concern/graduate_thesis_or_dissertations/js956p80v
Shedd, D. H. (1982). Seasonal variation and function of mobbing and related antipredator bheaviors of the American Robin (Turdus migratorius). Auk 99, 342–346.
Shedd, D. H. (1983). Seasonal variation in mobbing intensity in the black-capped chickadee. Wilson Bull. 95, 343–348.
Solheim, R. (1984). Caching behaviour, prey choice and surplus killing by pygmy owls Glaucidium passerinum during winter, a functional response of a generalist predator. Ann. Zool. Fenn. 21, 301–308.
Sordahl, T. (1990). The risks of avian mobbing and distraction behavior: an anecdotal review. Wilson Bull. 102, 349–352.
Keywords: anti-predator behavior, Glaucidium gnoma, Poecile atricapillus, Poecile rufescens, risky behavior, seasonal behavioral shifts, Sitta canadensis
Citation: Scott M and Robinson WD (2023) Mobbing behavior of songbirds in response to calls of an ambush-predator, the Northern Pygmy-Owl. Front. Ecol. Evol. 11:1092323. doi: 10.3389/fevo.2023.1092323
Edited by:
Anindita Bhadra, Indian Institute of Science Education and Research Kolkata, IndiaReviewed by:
Roi Dor, Open University of Israel, IsraelCopyright © 2023 Scott and Robinson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Madeleine Scott, ✉ bWFkZGllMTMzMUBnbWFpbC5jb20=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.