 Xiao-Kang Lu
Xiao-Kang Lu Tao Deng
Tao Deng Luca Pandolfi
Luca Pandolfi
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 16 February 2023
Sec. Paleontology
Volume 11 - 2023 | https://doi.org/10.3389/fevo.2023.1005126
This study presents the first phylogenetic analysis focused on the subfamily Aceratheriinae to date, with 392 characters (361 parsimony-informative characters) coded from 50 taxa at the species level. We added 80 newly defined and 33 revised characteristics to an existing matrix, covering features of the skull, teeth, and postcranial bones. Based on the results of ordered and unordered analyses, combined with a diagnosis in accordance with traditional morphological taxonomy, we revised the diagnosis of Aceratheriinae and reconstructed the phylogeny of Aceratheriinae. The tribe Teleoceratini, as well as the tribe Aceratheriini, was reclassified within Aceratheriinae; however, the traditionally established contents of each tribe were changed somewhat. Aceratheriinae underwent evolutionary adaptation several times during the early stages of its evolution, and several genera are herein reconstructed as early-diverging taxa, such as Floridaceras, Chilotheridium, and Plesiaceratherium. Turkanatherium and Protaceratherium are excluded from Aceratheriinae in this study. We suggest another two subclades of Aceratheriinae, containing Hoploaceratherium and Aprotodon, respectively. Aceratheriini and Teleoceratini are redefined as two highly specialized groups of Aceratheriinae.
Aceratheres are an extinct group of rhinocerotids widespread throughout Eurasia, North America, and Africa during the Neogene and characterized by the absence of nasal and/or frontal horns. The first hornless genus Aceratherium, from the Late Miocene, was recorded by Kaup (1832) in Eppelsheim, Germany. Based on this record, Dollo (1885) established the subfamily Aceratheriinae as a member of the family Rhinocerotidae, which has another two subfamilies, Rhinocerotinae and Elasmotheriinae (Prothero et al., 1989; Heissig, 1999; Antoine, 2002). Osborn (1900) recognized the subfamily Aceratheriinae and included some Oligocene and Miocene European species within it, giving the following diagnosis: “dolichocephalic with long, narrow nasals; smooth or with rudimentary horns at sides of the tips; frontals finally developing horns; large cutting teeth; relatively persistent tetradactyl manus; long-limbed” (Osborn, 1900, p. 240). Osborn (1900) included the following species within Aceratheriinae: “Aceratherium” filholi, “A.” lemanense, “A.” platyodon, “A.” blanfordi, “A.” tetradactylum, A. incisivum. Heissig (1973) included within Aceratheriinae the tribe Teleoceratini Hay, 1902, which contains the brachypotheres Teleoceras Hatcher, 1894, Brachypotherium Roger, 1904, Aprotodon Forster-Cooper, 1915, Diaceratherium Dietrich, 1931, and the tribe Aceratherini Dollo, 1885, which includes the genera Aceratherium Kaup, 1832, Plesiaceratherium Young, 1935, Chilotherium Ringström, 1924, and Aphelops Cope, 1873. In 1989, Heissig included within Aceratherinii the genera Mesaceratherium Heissig, 1969, Alicornops Ginsburg & Guérin, 1979, Aceratherium Kaup, 1832, Plesiaceratherium Young, 1935, Hoploaceratherium Ginsburg & Heissig, 1989, Aphelops Cope, 1873, Peraceras Cope, 1880, Chilotheridium Hooijer, 1971, Turkanatherium Deraniyagala, 1951, Chilotherium Ringström, 1924, Subchilotherium Heissig, 1972, and Acerorhinus Kretzoi, 1942.
In the many years since being established, over 90 species have been referred to Aceratheriinae. However, according to Prothero (2005), this taxon has been used as a taxonomic wastebasket for all hornless rhinoceroses. Not every hornless rhinoceros can be classified as Aceratheriinae, which is advanced and even includes rhinoceroses with a small nasal horn. Other features that characterize Aceratheriinae include a deep nasal notch, reduced upper incisor I1, considerably large lower incisor i2, brachycephalic skull, tetradactyl manus, and shortened and massive metapodials (Heissig, 1989; Cerdeño, 1995).
The content of Aceratheriinae has been in dispute for a long time. Pavlow (1892) united Aphelops, Teleoceras, and Brachypotherium, but assigned Aceratherium to the subfamily Rhinocerotinae. Scott and Osborn (1898) recognized the close relationship between Peraceras and Aphelops. Heissig (1973, 1999) was the first to include the tribe Teleoceratini in Aceratheriinae. Prothero et al. (1986) and Prothero et al. (1989) suggested that only the members of Aceratheriini could be recognized as Aceratheriinae; thus, the tribe Teleoceratini should be a member of Rhinocerotinae. Using 72 characters coded from 43 rhinocerotid taxa, Cerdeño (1995) found that Teleoceratini was more closely related to Aceratheriini than the other groups within Rhinocerotidae.
In order to explore the phylogeny of the elasmotheres, Antoine (2002) expanded the matrix to 282 characters. Deng (2008) used this matrix to discuss the taxonomic identity of new elasmotheres. However, this combination of characters could not resolve the phylogeny of the aceratheres (Antoine et al., 2003, 2010), despite the addition of several features by Lu (2013).
Aceratheriinae (Aceratheriini or Aceratheriina) represents a monophyletic group within the subfamily Rhinocerotinae according to Antoine (2002), Antoine et al. (2003, 2010), Becker et al. (2013). The aceratheres were classified as the subfamily Aceratheriinae by Prothero (2005) and also by Heissig (2012), who considered this group the sister to the Rhinocerotinae and Elasmotheriinae clade. The latest phylogenetic analysis of Rhinocerotidae found the traditional contents of Aceratheriinae, Rhinocerotinae, and Elasmotheriinae to form polyphyletic groups with confusing relationships (Lu et al., 2016).
This study aims to produce a new inclusive phylogenetic analysis of Aceratheriinae, for both taxa and characters, partly including previously defined characters. We also aim to reappraise the phylogenetic definition of Aceratheriinae and discuss the taxonomic positions of several genera. In order to provide a comprehensive suggestion, the following discussion is not only based on the results of the present cladistic analysis but also on the traditional ways of morphological comparison.
The characters in the present matrix were mainly from the works by Antoine (2002) and Lu (2013). After an extensive examination of the morphology of the members of Rhinocerotidae, we added 80 newly defined characters, and revised 33 previously used characters, covering features of the skull, teeth, and postcranial bones. These were directly from the unpublished doctoral dissertation of one author (Lu, 2014). In total, 392 characters were used in this analysis, 93 characters of the skull, 168 of the teeth, and 131 of the postcranium (Supplementary materials 1, 2).
In this matrix, some characters were potentially continuous, such as the widening of the mandibular symphysis. As many authors have pointed out that such characters should be ordered (Slowinski, 1993; Wiens, 2001; Grand et al., 2013; Rineau et al., 2015, 2018; briefly reviewed in Marjanović and Laurin, 2019), we designed 98 characters as ordered: 6, 9, 14–16, 18, 35, 43–46, 50, 52–53, 58–61, 63, 69, 77, 79–80, 82, 84, 89, 91, 95–98, 108, 114–117, 119–120, 124–125, 127, 129, 133, 148, 153, 155–156, 161–162, 166–169, 171, 173–176, 184–187, 191, 196, 198, 211, 213–214, 216, 220–221, 223–227, 233–234, 253, 258, 260, 279, 293, 308, 312, 314, 318–319, 323, 331–332, 348, 357, 368, 370, 372–373, and 384. However, we also performed another analysis, treating all characters as unordered. Finally, we will present and discuss the results generated from these different assumptions.
Because there is no consensus on the phylogeny of Aceratheriinae, the present analysis incorporated all the genera once referred to as Aceratheriini and Teleoceratini, with 25 taxa, and other genera of rhinocerotoids, with 24 taxa, which provided a larger basis for reconstructing a reliable topology. The outgroup is Tapirus terrestris. All the taxa in this analysis were at the species level, represented mainly by the type species of each genus. Teleoceras is known for its graviportal limbs, but the relevant materials are at present not accessible for the type species Teleoceras major, so we further included the most characterized species, Teleoceras fossiger. The type species of Aprotodon is only known from a crushed skull and tooth fragments; thus, we chose the Chinese species Aprotodon lanzhouensis, the phylogenetically informative parts of whose body are well known, particularly the lower incisor i2. For the same reasons, we included two taxa of Hispanotherium. The character scoring relied on direct observation or data from the literature (Supplementary material 3).
The parsimony analyses were performed using TNT version 1.5 (Goloboff et al., 2008; Goloboff and Catalano, 2016), with 1,000 replicates, tree bisection-reconnection (TBR) branch swapping, and 100 trees saved per replicate. All characters were equally weighted. The Bremer support values were calculated in TNT by running a traditional search on the most parsimonious tree, with 1,000 replications and TBR branch swapping.
We present the results generated by the ordered and unordered analyses, based on which we discuss the diagnosis and definition of Aceratheriinae in the following section. By ordering or not ordering certain characters in our dataset, we received two results with little difference. With all characters unordered, the maximum parsimony analysis resulted in one tree with 1,825 steps (CI = 0.28, RI = 0.51; Figure 1). When 98 characters were treated as ordered, with 361 parsimony-informative characters, three most parsimonious trees were generated, and the consensus tree had 2,018 steps (CI = 0.25, RI = 0.54; Figure 2). The Bremer support values were low for the clades of interest under both assumptions. This is likely due to the high proportion of missing data and homoplastic characters, which could increase the instability in the suboptimal trees, thereby causing the clades to collapse.

Figure 1. Consensus tree of the three most parsimonious trees with 98 characters ordered. Numbers above nodes are counts of synapomorphies and below nodes are Bremer support values. Colored areas and labeled thick lines on the right side indicate phylogeny frameworks.
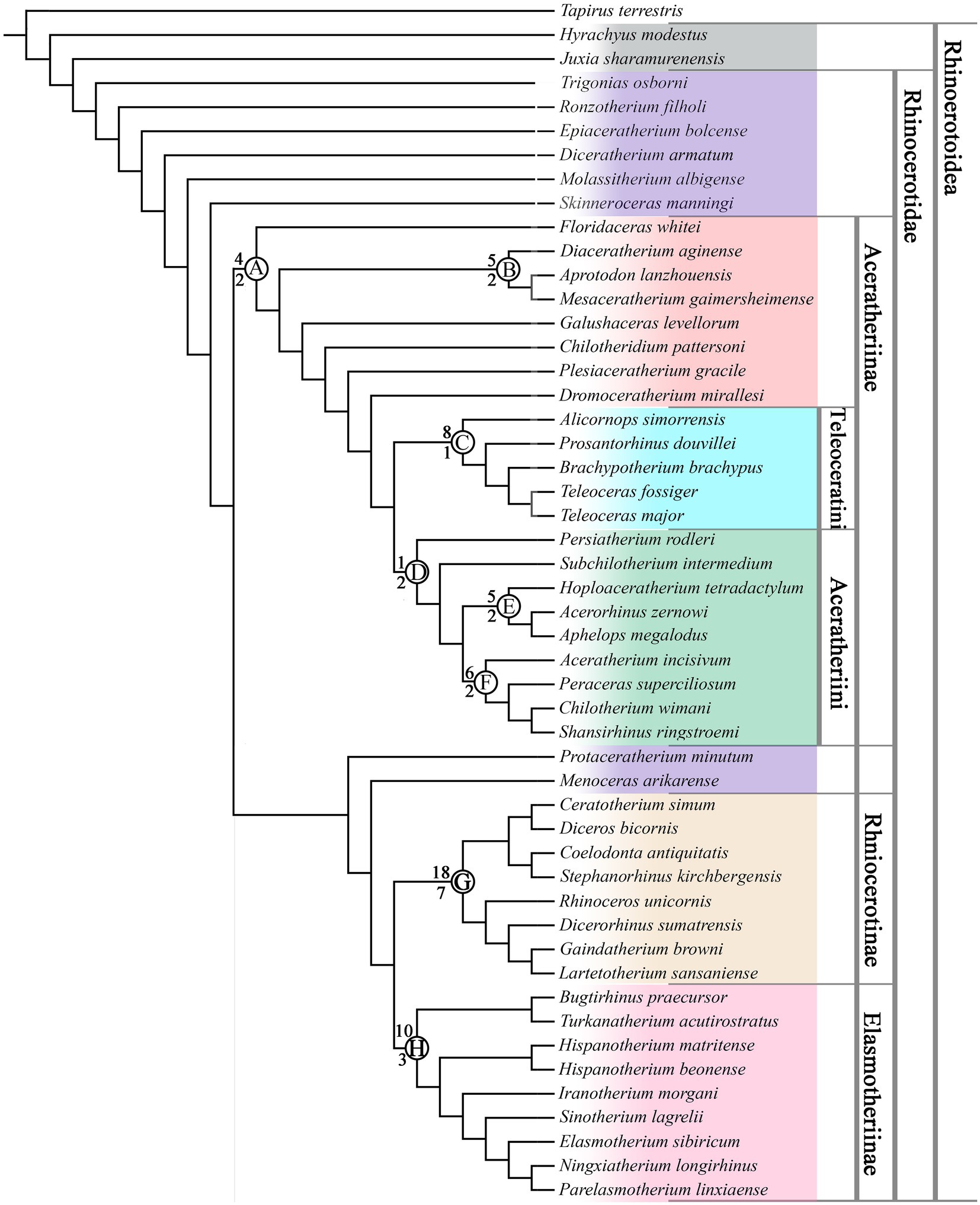
Figure 2. Single most parsimonious tree with all characters unordered. Numbers above nodes are counts of synapomorphies and below nodes are Bremer support values. Colored areas and labeled thick lines on the right side indicate phylogeny frameworks.
Both reconstructed trees were roughly consistent with the widely recognized contents of Rhinocerotinae (Figure 1, node 8, Char. 61–0, 150–1, 170–1, 670–1, 1070–1,1230–1, 1643–0, 1741–0, 19612–0, and 2133–2; Figure 2, node G, Char. 60–2, 150–1, 170–1, 610–2, 670–1, 1240–1, 1643–0, 1741–0, 1962–0, 2400–1, 2420–1, 2650–1, 3011–0, 3081–2, 3301–2, 3311–2, 3800–1, and 3881–0) and Elasmotheriinae (Figure 1, node 9, Char. 141–0, 380–1, 390–1, 420–1, 1331–0, 1482–1, and 1532–1; Figure 2, node H, Char. 380–1, 390–1, 991–2, 1422–3, 1490–1, 1532–1, 1582–3, 1750–3, 2020–2, and 3660–1; Cerdeño, 1995; Antoine, 2002; Antoine et al., 2010).
Nearly all taxa that were previously referred to as the tribe Aceratheriini or the tribe Teleoceratini were clustered as a large clade (Figure 1, node 1, Char. 310–1, 890–1, 1070–1, 1080–1, 1752–3, 1760–1, and 2010–1; Figure 2, node A, Char. 1070–1, 1090–1, 1180–1, and 1560–1). It was unexpected that Protaceratherium was excluded from the clade of Aceratheriinae. Furthermore, Turkanatherium was found to be a member of Elasmotheriinae, similar to the results by Geraads and Zouhri (2021).
However, the results of the present analysis did not fully support the previously suggested content of Aceratheriini in detail, and likewise for Teleoceratini (Heissig, 1989; Prothero and Schoch, 1989; Cerdeño, 1995). With 98 characters ordered, Plesiaceratherium, Chilotheridium, and Galushaceras were united as one group, supported by four synapomorphies (Figure 1, node 3, Char. 80–1, 161–2, 680–1, and 771–0). Dromoceratherium and Persiatherium were an independent pair of sister groups (Figure 1, node 4, 1490–1, 1541–2, and 1680–2).
On the other hand, when all characters were unordered, the content of Aceratheriinae was slightly different from above. Floridaceras, Galushaceras, Chilotheridium, Plesiaceratherium, and Dromoceratherium were placed as early diverged single-genus clades in Aceratheriinae. There was a clade clustering Diaceratherium, Mesaceratherium, and Aprotodon, consistent with their mosaic characters: less advanced upper premolars, greatly specialized incisors (I1 and i2), and symphysis (Deng, 2013; Figure 2, node B, Char. 321–0, 590–1, 981–2, 1561–3, and 2170–1). In the alternative result, only Mesaceratherium and Aprotodon were clustered, supported by similarities such as the incisors (Figure 1, node 2, Char. 801–2, 820–1, 1081–2, and 2230–1).
Two subclades were slightly different in the two trees (Figures 1, 2). Five genera previously referred to as Aceratheriini, namely Aceratherium, Peraceras, Chilotherium, Shansirhinus, and Subchilotherium, were clustered as a clade among the most advanced group of Aceratheriinae (Figure 1, node 7, Char. 30–1, 250–1, 960–1, 1100–1, 1190–1, 2590–1, and 3610–1). However, with all characters unordered, Subchilotherium was placed as a single-genus clade and the first four genera were united (Figure 2, node F, Char. 560–1, 610–1, 691–2, 1610–3, 3410–1, and 3471–0). Under this assumption, three genera, Brachypotherium, Prosantorhinus, and Teleoceras, were retained within Teleoceratini. Alicornops, which has long been referred to as Aceratheriini (Prothero et al., 1986; Cerdeño, 1995), was placed as a sister group to the clade merging these three genera of Teleoceratini (Figure 2, node C, Char. 830–1, 1552–1, 2131–2, 3151–0, 3480–1, 3571–2, 3611–0, and 3911–0). On the other hand, with 98 characters ordered, these three genera were also united (Figure 1, node 6, 51–3, 80–1, 431–2, 680–1, 991–0, 1170–1, 1240–1, 1671–0, 2060–1, 2580–1, 2791–2, 2930–2, 2960–1, 3000–1, 3011–0, 3290–1, 3301–2, 3312–3, 3340–1, 3571–2, 3640–1, 3690–1, 3890–1, and 3920–1), and Alicornops was present as a sister group of this clade.
There was one stable subclade with three genera in the results of both analyses, Hoploaceratherium, Aphelops, and Acerorhinus (Figure 1, node 5, Char. 30–1, 250–1, 960–1, 1220–1, 1840–1, 2132–3, and 2940–1; Figure 2, node E, Char. 341–0, 1220–1, 1272–1, 2991–0, and 3611–0). With all characters unordered, this subclade was united with the subclade of Aceratherium, forming a pair of sister groups. Two single-genus clades, namely Persiaceratherium and Subchilotherium, showed a close relationship with them (Figure 2, node D, Char. 1611–0).
Our phylogenetic analyses provide characters supporting the Aceratheriinae node; those would be used as diagnostic features, but there are only four or seven characters, far from enough to provide a distinct outline of aceratheres. On the other hand, some features that have long been considered as diagnostic of Aceratheriini or Teleoceratini were not found as synapomorphies, such as features concerning the tusk i2, the brachycephalic skull, and the graviportal limbs. These features demonstrate a series of gradually changing tendencies and should be considered when reamending the diagnosis.
The clear starting point when studying the characteristics of hornless rhinoceroses is the horn. A small horn and the enlarged upper incisors I1 were two crucial characteristics that Prothero et al. (1986) used to include Teleoceratini in Rhinocerotinae rather than Aceratheriinae. However, among members once classified as Aceratheriini, at least four genera developed a small nasal horn, namely Chilotheridium, Hoploaceratherium, Peraceras, and Shansirhinus. In Teleoceratini, the presence of a nasal horn has interspecific variation in Diaceratherium and Brachypotherium (Répelin, 1917; Heissig, 1999, 2017; Figure 3). Nasal horns developed in different lineages of rhinocerotids several times.
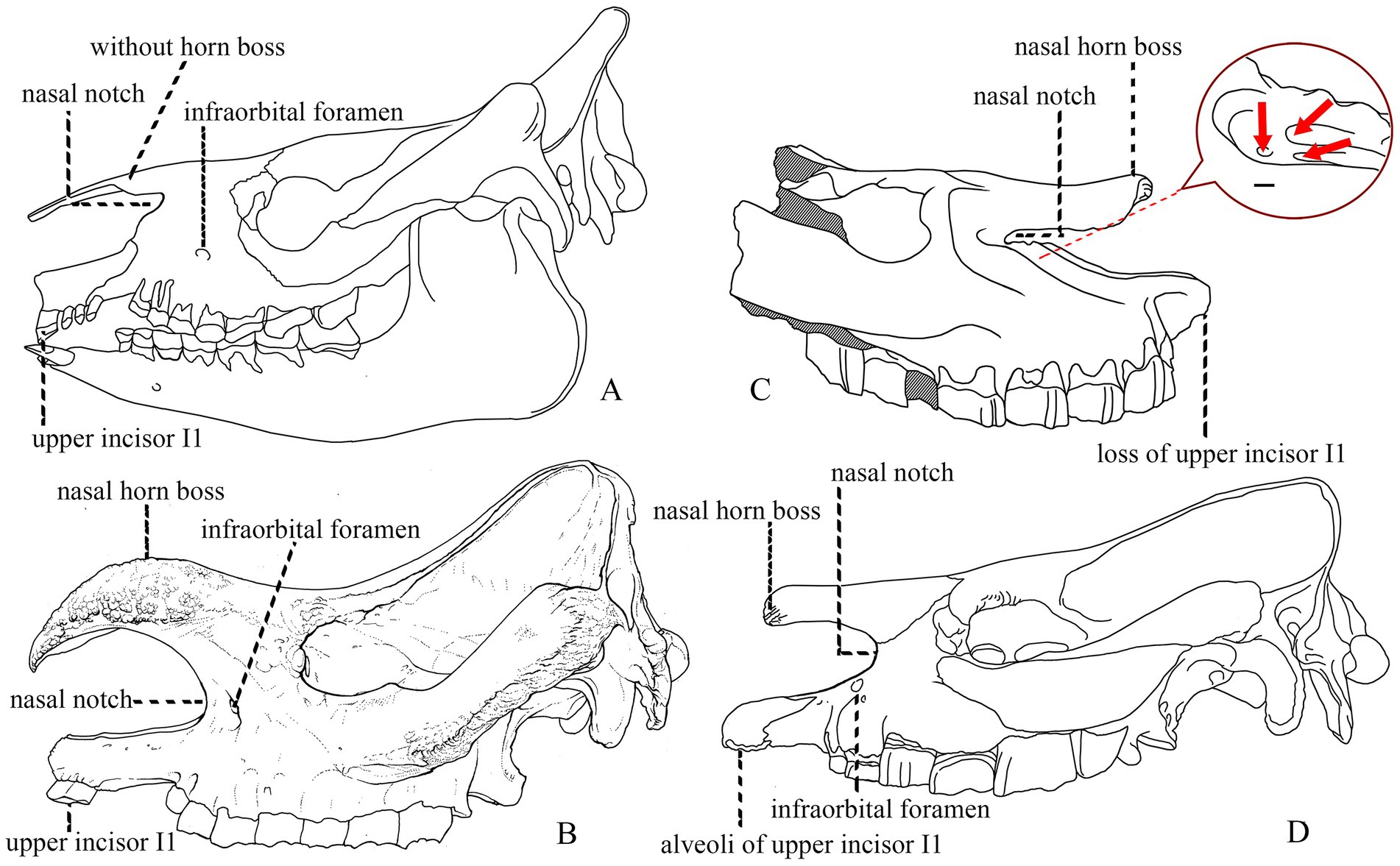
Figure 3. Skulls of rhinocerotids. (A) Trigonias osborni, AMNH 9847, Scott (1941). (B) Rhinoceros unicornis, IVPP OV 1383, new specimen. (C) Shansirhinus ringstroemi, Qiu and Yan (1982), the red circle marks three infraorbital foramina. (D) Teleoceras fossiger, Osborn (1900). Not to scale.
Another notable structure lies in the rostral end of the skull and involves two aspects, namely the retracted nasal notch and the specialized incisors, both of which are closely related to feeding behavior (Prothero et al., 1986; Heissig, 1989, 1999; Cerdeño, 1995). The enlarged lower incisor (i2) is a typical feature differentiating rhinocerotids from other rhinocerotoids and was maximized in the evolution of Aceratheriini and Teleoceratini, exemplified by Aceratherium, Chilotherium, and Teleoceras (Ringström, 1924; Deng, 2001; Prothero, 2005; Figure 3). However, adaptations of the upper incisor I1 diverged in the two tribes; most taxa retained the upper incisor I1 (Figure 2, node B, Char. 98), while in the advanced members, I1 was lost (Figure 1, node 5 and 7, Char. 96), and the enlarged lower incisor i2 subsequently lost the occlusal surface on the internal edge and developed a labially upturning occlusal surface (Figure 1, node 1, Char. 107; Figure 2, node A, Char. 107, 109), such as in Chilotherium (Deng, 2001; Lu, 2013; Figure 4).
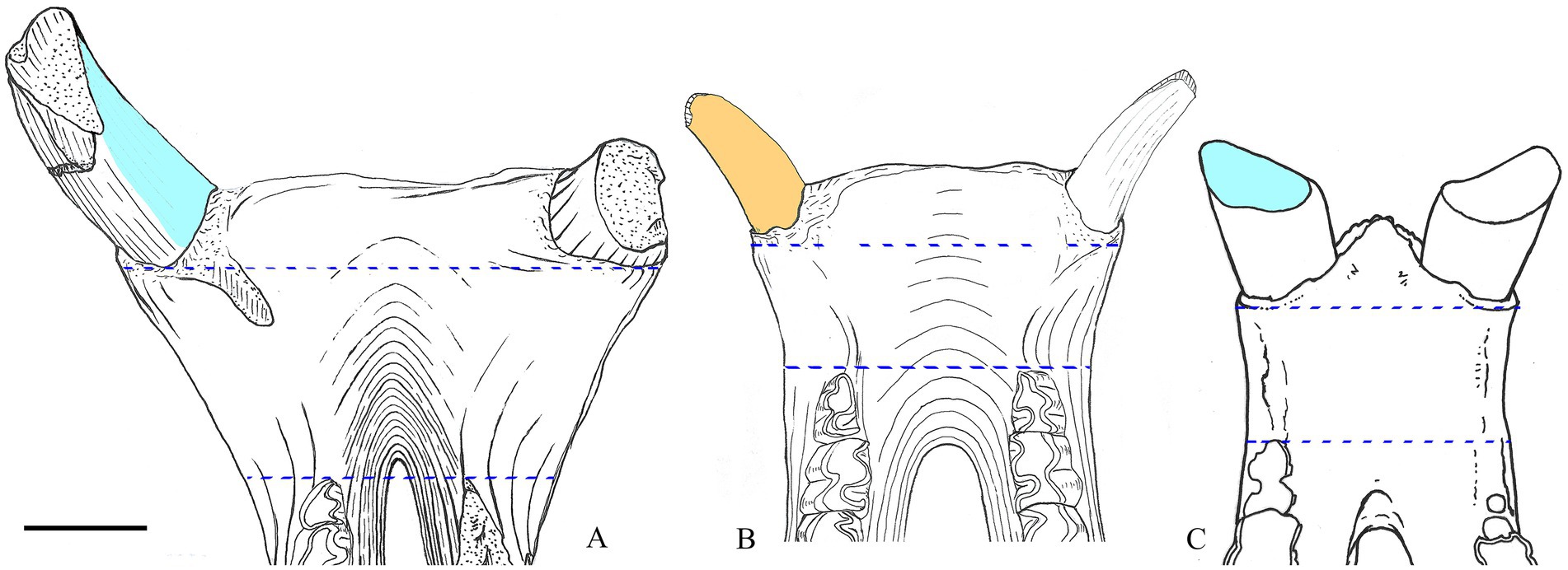
Figure 4. Lower incisors i2 of rhinocerotids. (A) Aprotodon lanzhouensis, IVPP V 11060, based on a specimen by Qiu and Xie (1997). (B) Chilotherium wimani, IVPP V 12505, based on a specimen by Deng (2001). (C) Rhinoceros unicornis, IVPP OV 1383, new specimen. Blue dash lines mark the width of symphysis, sky blue areas mark the occlusal surface for upper incisor I1, and the orange areas mark the surface wear with the upper lip when I1 was lost. Scale bar=50mm.
Another feature in the rostral area is the retracted nasal notch, even to the level of M1, which evolved gradually and occurred in nearly all rhinocerotids (Prothero et al., 1986; Heissig, 1989; Cerdeño, 1995; Antoine, 2002; Prothero, 2005). Its consistent appearance in the members of Aceratheriini and Teleoceratini could be evaluated by its position relative to the infraorbital foramen: the position of the infraorbital foramen moved from the posterior to the nasal notch to below it. In the primitive rhinocerotids from the Eocene Hyrachyus and Trigonias to the extant Rhinoceros, the nasal notch is shallow, and the infraorbital foramen is behind the notch and above the level of P2–P4 (Gregory and Cook, 1928; Prothero, 2005). However, in the Oligocene Aprotodon, the nasal notch was moderately retracted, at P3, and the infraorbital foramen moved to below its posterior end. By the Late Miocene, this foramen had increased to three openings in Shansirhinus and Teleoceras but remained below the ventral edge of the nasal notch (Figure 3; Qiu and Yan, 1982; Qiu and Xie, 1997; Deng, 2005; Prothero, 2005; Lihoreau et al., 2009).
The broadening and shortening tendencies of the skull occured exclusively in Aceratheriini and Teleoceratini and have important systematic value (Heissig, 1989; Cerdeño, 1995). However, both are gradually changing characteristics, with a long history of evolution; their maximum degree only occurred in very advanced taxa. Based on the maximum length/maximum width ratio, several genera had a brachycephalic skull, namely Peraceras, Teleoceras, Brachypotherium, and Prosantorhinus (Prothero, 2005; Heissig, 2017). In this analysis, the skull outline was found to be one of the synapomorphies uniting Teleoceratini (Figure 1, node 6, Char. 43). The graviportal limbs were exemplified in several aspects, and we have added new features to discern, such as the length ratio between the dorsal and lateral sides of the magnum (Char. 312), the length/width ratio of the third metacarpal (Char. 330, 331), and the outline of the unciform (Char. 317; Figure 5). The brachycephalic skull, accompanied by graviportal limb, occured in Peraceras, Teleoceras, and Prosantorhinus (Figure 1, node 6, Char. 330, 331, 357; Prothero, 2005; Heissig, 2017). Both characters are related to a short and massive body and are notable aspects of the appearance of Aceratheriini and Teleoceratini.
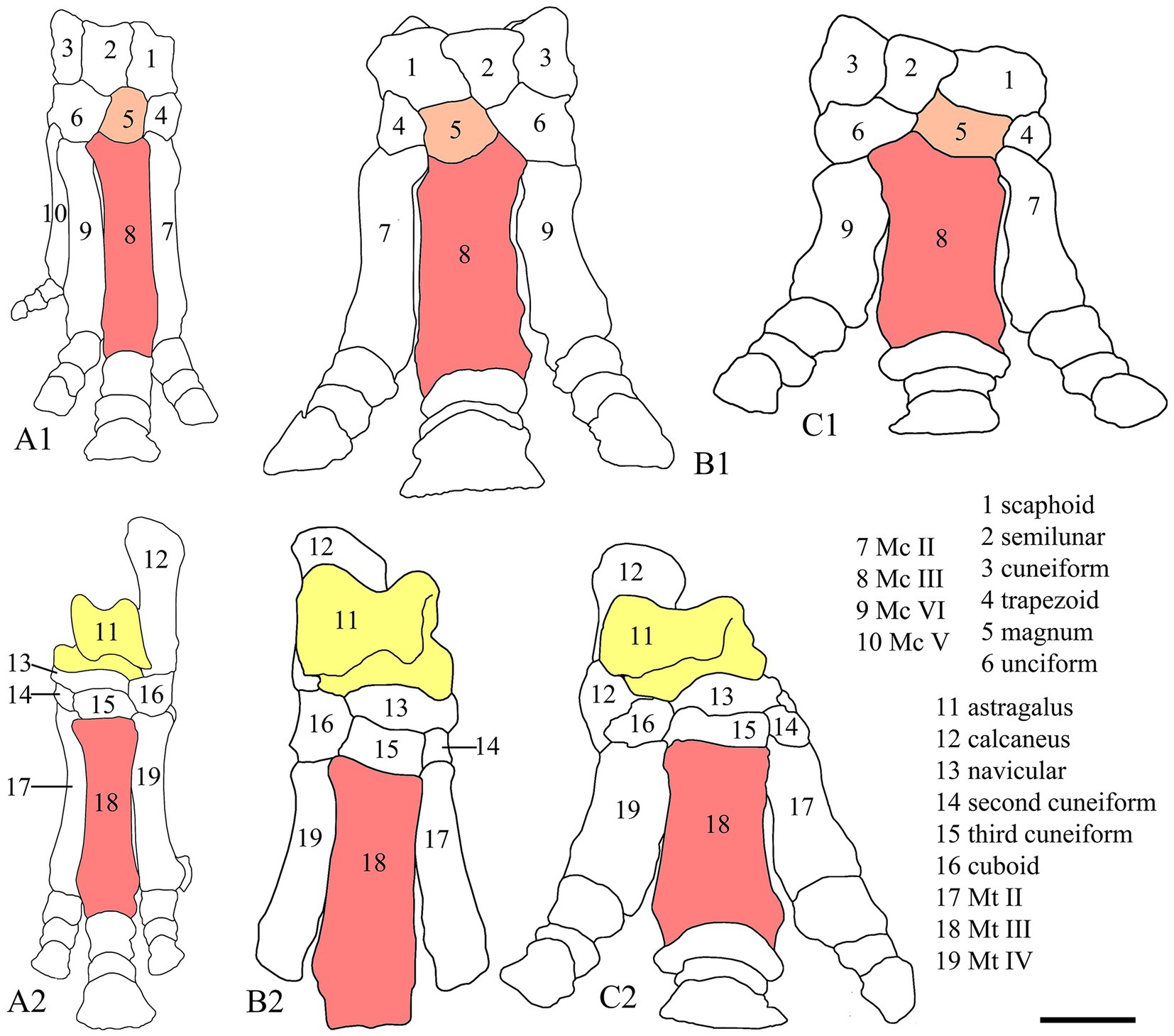
Figure 5. Drawings of distal limb bones of rhinocerotids. 1, fore-limb; 2, hind-limb. (A) Trigonias osborni, AMNH 9847, Scott (1941). (B) Aphelops megalodus, A1/A2 AMNH 9745/5293, based on a specimen by Matthew (1918). (C) Teleoceras fossiger, AMNH 2650, based on a specimen by Matthew (1918). The scale of the latter two are from measurements by Prothero (2005). Color areas mark magnum, Mc III, astragalus, and Mt III on different rhinoceroses. Scale bar = 50 mm.
In addition to the graviportal tendency of the limbs, Aceratheriini and Teleoceratini demonstrate another diagnostic character regarding the postcranial bones: the primitive members maintained a functional fifth metacarpal (Heissig, 1989, 1999; Prothero, 2005). The functional fifth metacarpal was inherited from the primitive ancestor of the Rhinocerotidae, and has been reduced to a small sesamoid-like bone in the advanced members of Aceratheriini and Teleoceratini (Figure 1, node 6, Char. 334; Figure 4; Ringström, 1924; Deng, 2002).
Traditionally, cheek teeth are an important diagnosis of Aceratheriini or Teleoceratini: deep constriction of both lingual cusps of the upper cheek teeth (Figure 1, node 1, Char. 175; Figure 2, node A, Char. 156) with a maximum degree that is just slightly weaker than that of elasmotheres; and the continuous lingual cingulum of the upper premolars, which was inherited from a primitive ancestor (Heissig, 1989, 1999; Figure 5). The former could complicate the occlusal pattern of the cheek teeth, and the latter would reinforce the lingual enamel wall of significantly worn teeth. Both features are quite useful for determining the taxonomic identity of fossil material of aceratheres.
The present two analyses found that Aceratheriini and Teleoceratini were closely related and united as Aceratheriinae, consistent with the results of Heissig (1989, 1999) and Cerdeño (1995). Here, we used the node-based definition of Aceratheriinae as the last common ancestor of Aceratherium, Teleoceras, and Aprotodon and all of its descendants. On the other hand, Elasmotheriinae and Rhinocerotinae showed a closer relationship relative to Aceratheriinae. This result is partly consistent with Cerdeño’s work (1995), in which Aceratheriinae formed a clade, and Elasmotheriinae and Rhinocerotinae were united but each did not form their monophyly, respectively. Antoine and his colleagues established the subfamily Elasmotheriinae but placed aceratheres and rhinoceroses together as a paraphyletic group (Antoine, 2002; Antoine et al., 2003, 2010). In this study, Elasmotheriinae and Rhinocerotinae were united as sister groups, supported by many characters (Supplementary material 4). These synapomorphies revealed that, compared to aceratheres, elasmotheres and rhinoceroses were more advanced in their skull, particularly the large nasal and frontal horns, and in their cheek teeth, such as the hypsodont crown of elasmotheres and the much higher crown, widened crest, and strong ectoloph (mesostyle and metacone rib) of rhinoceroses, but less advanced in their limb bones, preserving the tetradactyl manus and slender or strong limb.
Among the 25 taxa previously referred to as Aceratheriinae, Turkanatherium was excluded from this group and recovered as Elasmotheriinae. Deraniyagala (1951) described and established Turkanatherium based on materials from the middle Miocene of Moruorot, Kenya, while, Arambourg (1959) and Hooijer (1966) referred to this taxon as Aceratherium without further discussion. However, Geraads (2010) re-identified it as Turkanatherium based on the differences between Turkanatherium acutirostratum and Aceratherium incisivum, and attributed it to the subtribe Aceratheriina. The position of T. acutirostratum was recently reported within Elasmotheriinae by Geraads et al. (2016), who, however, suggested that their results should be interpreted cautiously. Furthermore, four features of this genus differed from Aceratheriinae; the nasal notch was shallow at the level of P2, and the infraorbital foramen was behind the nasal notch; the upper premolars had a metacone rib on the ectoloph and had lost the lingual cingulum. Turkanatherium maintained a notably primitive structure of the upper premolars; the metaloph had yet to be fully formed. Evidently, Turkanatherium was not an acerathere, so we tentatively followed the result of the analysis and assigned it as a basal elasmothere.
Protaceratherium was represented only by the type species P. minutum, found in the early Miocene of Western Europe, and had long been considered a member of Aceratheriinae (Cerdeño, 1995; Heissig, 1999). This genus had a pair of enlarged lower incisors i2, long and narrow nasal bones, and primitive cheek teeth Roman, 1912. Antoine (2002) and Becker et al. (2013) have suggested that it was close to the group of Aceratheriinae and Rhinocerotinae. In this analysis, its position was within Rhinocerotinae.
Teleoceratini: This tribe was established based on the genus Teleoceras from the Late Miocene in North America, previously with five widely recognized genera, Aprotodon, Diaceratherium, Brachypotherium, Teleoceras, and Prosantorhinus (Heissig, 1989, 1999; Prothero and Schoch, 1989; Prothero, 2005). However, the present analysis, which sampled all genera of Aceratheriini and Teleoceratini, questioned previous taxonomic opinions and reconstructed a different clade that included Brachypotherium, Teleoceras, and Prosantorhinus, but excluded Aprotodon and Diaceratherium; both the latter, together with Mesaceratherium, were recovered as an early-diverging clade of Aceratheriinae.
Aprotodon from the late Oligocene in Asia has long been recognized as an early member of Teleoceratini: the skull has a slender outline, the parietal crests fuse and form the sagittal crest, and the upper premolar has yet to be fully molarized (Forster-Cooper, 1915; Borissiak, 1954). In Cerdeño’s (1995) analysis, it was recovered as the most basal Rhinocerotinae. The description of the new materials of Aprotodon from China, Qiu and Xie (1997) likewise suggested it should be an early-diverging lineage of Rhinocerotidae. Mesaceratherium from the late Oligocene of Eurasia has long been considered the earliest genus of Aceratheriini, with less molarized premolars (Heissig, 1969). Antoine et al. (2010) placed it as a sensu lato acerathere, and in the newest paper this genus and some other Paleogene rhinoceroses were placed as an early clade of Rhinocerotinae (Tissier et al., 2020). Given its enlarged upper and lower incisors, like Aprotodon, we suggested Mesaceratherium should be an acerathere, consistent with the present analyses.
Diaceratherium from the late Oligocene to the early Miocene was widely distributed around the old world (Heissig, 1999; Becker et al., 2009; Lu et al., 2021). It was a large-sized rhinoceros, with a small nasal horn, dolichocephalic skull, large upper and lower incisors, and long and robust distal limb bones (Lu et al., 2021). This genus has long been considered an early ancestor of some advanced teleoceratini genera. Sizov et al. (2022) tried to classify all species of Diaceratherium except the type species to Brachydiceratherium, indicating a complicated relationship of this genus, and put D. tomerdingense in the basal position of Teleoceratini. Together with the discussion of the relationship between Diaceratherium and Prosantorhinus by Heissig (2017), the taxonomic and phylogenetic relationships are debatable and complex. The present analyses suggested that Diaceratherium was a single-genus clade of Aceratheriinae, with the position unstable. Consequently, meanwhile, the diagnosis of Teleoceratini no longer primarily depended on features of the nasal horn and the upper incisors; the skull outline and metapodial proportions were more significant, as discussed in the above section.
Alicornops has long been referred to as Aceratheriini (Prothero et al., 1986; Cerdeño, 1995). This genus from the middle Miocene was a moderately specialized genus, with a medium-sized skull and moderately specialized cheek teeth (Cerdeño and Sánchez, 2000; Deng, 2004). Heissig (1999, 2012) once considered Alicornops a subgenus of Aceratherium. This referral is confusing because the latter had lost the upper incisors I1 and its limb bones were slender. In addition to the synapomorphies at node C, Alicornops showed a robust distal limb bone, similar to that of Teleoceratini. The lateral and medial sides of the diaphysis of the third metacarpal were irregular, so the width of the diaphysis was similar from the proximal to distal portions—there was no narrowest position. Alicornops was distinct from Aceratherium and Chilotherium, because, in the latter two, the narrowest position of the diaphysis was at the proximal extremity (Hünermann, 1989; Cerdeño and Sánchez, 2000; Deng, 2002). The outlines of the astragalus in Alicornops were more flattened than Chilotherium, which was the most advanced member of Aceratheriini (Ringström, 1924; Deng, 2002). We tentatively consider Alicornops a single-genus clade with an unstable phylogenetic position.
Aceratheriini: The results of the present analyses do not support the previously established contents of Aceratheriini and Aceratheriinae (Heissig, 1989; Prothero and Schoch, 1989; Cerdeño, 1995). Turkanatherium and Mesaceratherium have been mentioned above. Three subclades were consistently reconstructed in both trees, but two were unstable (Figures 1, 2).
In the clade of Hoploaceratherium, Acerorhinus, and Aphelops (Figure 1, node 5; Figure 2, node E), the relationship between the former two has been noted in previous studies because of their longer nasal bones (Heissig, 1989). Additionally, among the synapomorphies supporting this clade, the most significant was a uniquely derived feature, the expansion of the lingual cusps of the upper cheek teeth (Char. 122). The constriction of the lingual cusps was an efficient way to complicate the occlusal pattern of the cheek teeth and was present in nearly all lineages of rhinocerotids, excluding the members of Rhinocerotini that expanded the lingual cusps while losing the constriction. The combination of both characters in Acerorhinus was first noted by Deng (2000): the expanded and rounded lingual cusps in the upper cheek teeth and the shallow constriction in the upper molars; however, the antecrochet remains present, although with a narrow outline (Figure 6). This synapomorphy, found only in these three genera, powerfully demonstrates a closer evolutionary relationship.
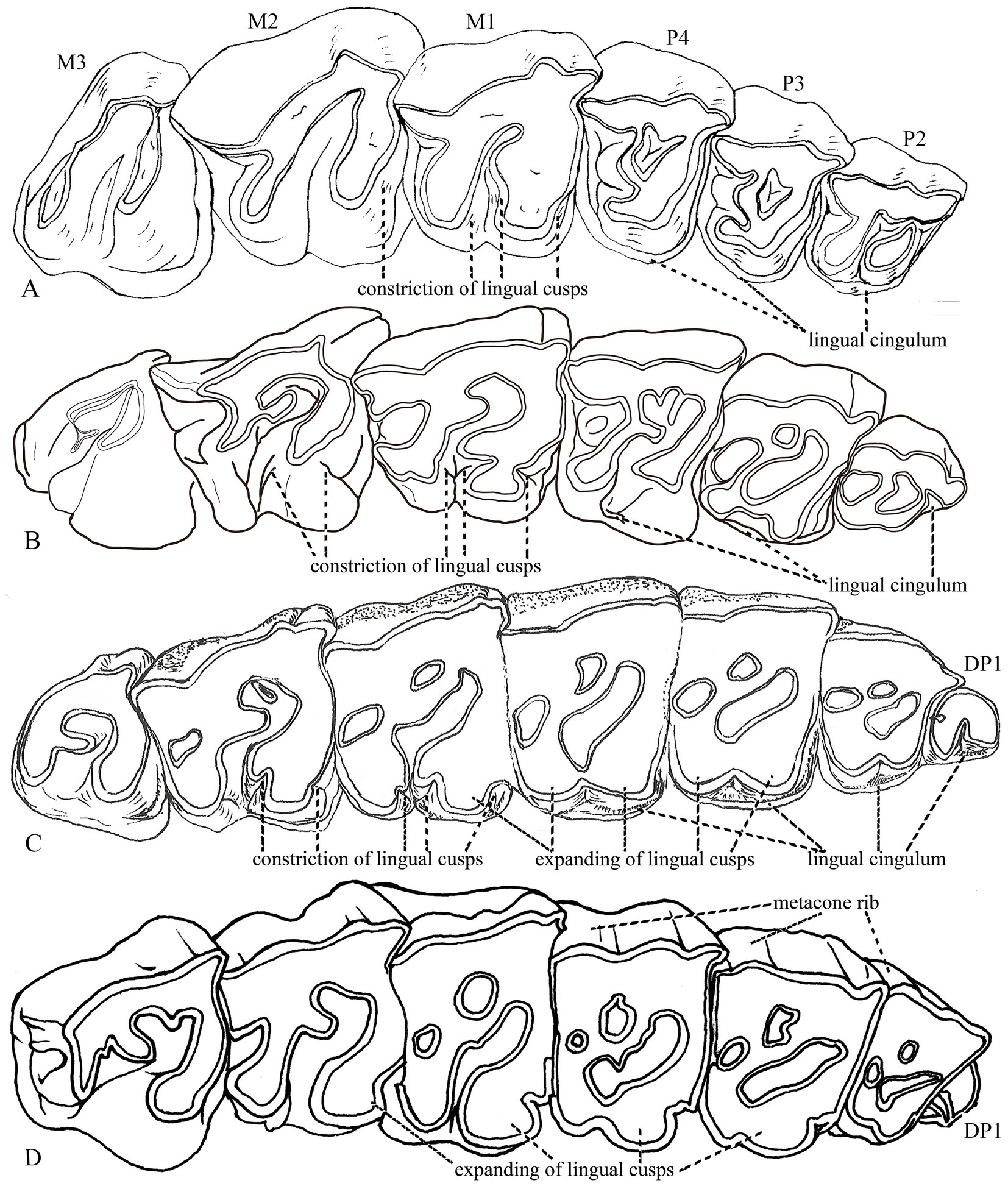
Figure 6. Upper cheek teeth of rhinocerotids. (A) Trigonias osborni, DMNH 414, based on a specimen by Gregory and Cook (1928). (B) Teleoceras fossiger, Osborn (1904), Figure 21. (C) Acerorhinus hezhengensis, HMV 0639, new specimen. (D) Rhinoceros unicornis, IVPP OV 1383, new specimen. Not to scale.
Another subclade of Aceratheriinae clustered five Late Miocene genera: Subchilotherium, Aceratherium, Peraceras, Chilotherium, and Shansirhinus (Figure 1, node 7). The most obvious synapomorphy of these five genera was related to the specialization of the rostral end of the skull: the premaxillae were slender, the upper incisor I1 were fully reduced, and the occlusal face of i2 directed outward. Furthermore, when all characters were unordered, and consequently Subchilotherium was excluded from this clade, the most obvious feature shared by these four genera related to the skull outline: the occipital crest was wide (Char. 56), and the parietal crest was not fused (Char. 61).
Persiatherium from Maraghen, Iran, was exemplified by a crushed skull with the cheek tooth row. For this rhinoceros from the Late Miocene, the most outstanding feature was the presence of the lingual cingulum on the upper molars. Reduction of the cingulum was a common tendency widely occurring among contents of rhinocerotids since the Oligocene (Heissig, 1989). Dromoceratherium had limited materials, consisting of fragments of upper and lower teeth, and limbs (Deraniyagala, 1951). As for now, no exclusive similarity was found between these two genera, and we did not expect them to form a clade.
Chilotheridium from the early Miocene in Africa was also an early-diverging genus, which is consistent with its mosaic morphology: dolichocephalic skull, slender limb bones, complicated cheek teeth, and a horned nasal bone at the subterminal position (Hooijer, 1971; Geraads, 2010). Plesiaceratherium was a well-known acerathere from the early Miocene of China, and its differences from Chilotheridium are many, such as the nasal horn (Yan and Heissig, 1986; Lu et al., 2016). It was also true for Galushaceras. This subclade and the subclade of Persiatherium and Dromoceratherium were not consistently present in the two analyses using different assumptions; their relationships were unstable and reconstructed as single-genus clades (Figures 1, 2).
When discussing the phylogeny of Aceratheriinae, Heissig (1973, 1989), Prothero et al. (1986), and Cerdeño (1995) did not distinguish its content from that of the tribe Aceratheriini. It is not reasonable and possible to perfectly refer each taxon of Aceratheriinae to either Aceratheriini or Teleoceratini. Here, some taxa were tentatively recovered as a set of single-genus clades. With 98 characters ordered, the subclade of Hoploaceratherium was placed outside Aceratheriini, indicating the possibility of a new tribe, as well as a subclade of Aprotodon. Tribes Aceratheriini and Teleoceratini were restricted to include taxa that were more specialized compared to a set of basal clades.
1. According to the results of the phylogenetic analyses of the newly established matrix, Aceratheriinae is defined as the last common ancestor of Aceratherium, Teleoceras, and Aprotodon and all of its descendants (node-based).
2. The diagnosis of Aceratheriinae is revised as follows. It was of medium-large size, tended to evolve a brachycephalic skull, and the presence of a nasal horn varied. It had a deep nasal notch, to the level of P3–M1, with an infraorbital foramen below the nasal notch. It exhibited an enlarged or lost upper incisor I1, the lower incisor i2 was tusk-like, even extremely enlarged. It had moderately specialized cheek teeth: the labial cingula of the upper and lower cheek teeth were reduced with residuals, but the lingual cingulum of the upper premolars was always unreduced; the lingual cusps of the upper cheek teeth could be constricted, or sometimes also expanded. It had a functional fifth metacarpal and the limbs had a tendency toward graviportality, their distal parts were always massive and flattened.
3. Aceratheriini and Teleoceratini are sister-groups. Teleoceratini includes Prosantorhinus, Brachypotherium, and Teleoceras. Aceratheriini includes Subchilotherium, Aceratherium, Peraceras, Chilotherium, and Shansirhinus. On the other hand, Floridaceras, Plesiaceratherium, Chilotheridium, Galushaceras, Dromoceratherium, Persiaceratherium, Diaceratherium, and Alicornops are tentatively considered single-genus clades.
4. In addition, we suggest another two subclades of Aceratheriinae. One consists of two genera from the late Oligocene to the early Miocene, Aprotodon and Mesaceratherium. Another unites three genera from the middle to Late Miocene, Hoploaceratherium, Acerorhinus, and Aphelops.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
Ethical review and approval was not required for the animal study because there has no animal experiment.
This research was supported by the National Natural Science Foundation of China (42172001); Strategic Priority Cultivating Research Program, Chinese Academy of Sciences (XDA20070203 and XDB26000000), and State Key Laboratory of Palaeobiology and Stratigraphy (Nanjing Institute of Geology and Palaeontology, CAS) (No. 203113).
We thank Chen Shanqin and He Wen from the Hezheng Paleozoological Museum, Linxia, China; Zheng Xiaoting, Yi Shiyin, Yi Dawei from the Tianyu Museum, Linyi, China; Wang Shiqi, Hou Sukuan, Shi Qinqin, Sun Boyang, Sun Danhui, Li Shijie, Chen Jin, and Si Hongwei from the Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China; Ji Xueping from Kunming Institute of Zoology, Chinese Academy of Sciences, Beijing, China; Gao Feng from the Yunnan Institute of Cultural Relics and Archaeology & Research Center for Southeast Asian Archeology, Kunming, China. Special thanks go to the editors and reviewers for their constructive suggestions for the improvement of the manuscript. We are grateful to Chen Yu and Xu Yong for preparing the drawings.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1005126/full#supplementary-material
Antoine, P.-O. (2002). Phylogénie et évolution des Elasmotheriina (Mammalia, Rhinocerotidae). Mém. Mus. Nat. Hist. Nat. 188, 1–359.
Antoine, P.-O., Downing, K. F., Crochet, J.-Y., Duranthon, F., Flynn, L. J., Marivaux, L., et al. (2010). A revision of Aceratherium blanfordi Lydekker, 1884 (Mammalia: Rhinocerotidae) from the early Miocene of Pakistan: postcranials as a key. Zool. J. Linnean Soc. 160, 139–194. doi: 10.1111/j.1096-3642.2009.00597.x
Antoine, P.-O., Duranthon, F., and Welcomme, J.-L. (2003). Alicornops (Mammalia, Rhinocerotidae) dans le Miocène superiéur des collines Bugti (Balouchistan, Pakistan): implications phylogénétiques. Geodiversitas 25, 575–603.
Becker, D., Bürgin, T., Oberli, U., and Scherler, L. (2009). Diaceratherium lemanense (Rhinocerotidae) from Eschenbach (eastern Switzerland): systematics, palaeoecology, palaeobiogeography. N. Jb. Geol. Paläont. Abh. 254, 5–39.
Becker, D., Antoine, P.-O., and Maridet, O. (2013). A new genus of Rhinocerotidae (Mammalia, Perissodactyla) from the Oligocene of Europe. J. Sys. Palaeont. 11, 947–972. doi: 10.1080/14772019.2012.699007
Borissiak, A. (1954). The oldest Aceratherium from Kazakhstan. Trans. Paleontol. Inst. Acad. Sci. USSR 47, 5–23.
Cerdeño, E. (1995). Cladistic analysis of the family Rhinocerotidae (Perissodactyla). Am. Mus. Novit. 3143, 1–25.
Cerdeño, E., and Sánchez, B. (2000). Intraspecific variation and evolutionary trends of Alicornops simorrense (Rhinocerotidae) in Spain. Zool. Scri. 29, 275–305. doi: 10.1046/j.1463-6409.2000.00047.x
Deng, T. (2000). A new species of Acerorhinus (Perissodactyla, Rhinocerotidae) from the Miocene in Fugu, Shaanxi, China. Vert. PalAsiat. 38, 203–217.
Deng, T. (2001). New materials of Chilotherium wimani (Perissodactyla, Rhinocerotidae) from the late Miocene of Fugu, Shaanxi. Vert. PalAsiat. 39, 129–138.
Deng, T. (2002). Limb bones of Chilotherium wimani (Perissodactyla, Rhinocerotidae) from the Late Miocene fo the Linxia Basin in Gansu, China. Vert. PalAsiat. 40, 305–316.
Deng, T. (2004). A new species of the rhinoceros Alicornops from the Middle Miocene of the Linxia Basin, Gansu, China. Palaeontology 47, 1427–1439. doi: 10.1111/j.0031-0239.2004.00420.x
Deng, T. (2005). New cranial material of Shansirhinus (Rhinocerotidae, Perissodactyla) from the Lower Pliocene of the Linxia Basin in Gansu, China. Geobios 38, 301–313. doi: 10.1016/j.geobios.2003.12.003
Deng, T. (2008). A new elasmothere (Perissodactyla, Rhinocerotidae) from the late Miocene of the Linxia Basin in Gansu, China. Geobios 41, 719–728. doi: 10.1016/j.geobios.2008.01.006
Deng, T. (2013). Incisor fossils of Aprotodon (Perissodactyla, Rhinocerotidae) from the early Miocene Shangzhuang formation of the Linxia Basin in Gansu, China. Vert. PalAsiat. 51, 131–140.
Deraniyagala, P. E. P. (1951). A hornless rhinoceros from the Mio-Pliocene deposits of East Africa. Spolia Zeylanica 26, 133–135.
Forster-Cooper, C. (1915). New genera and species of mammals from the Miocene deposits of Baluchistan. Ann. Mag. Nat. Hist. 16, 404–410. doi: 10.1080/00222931508693732
Geraads, D. (2010). “Rhinocerotidae” in Cenozoic Mammals of Africa. eds. L. Werdelin and W. J. Sanders (Berkeley: University of California Press), 669–683.
Geraads, D., Lehmann, T., Peppe, D. J., and McNulty, K. P. (2016). New Rhinocerotidae from the Kisingiri localities (lower Miocene of western Kenya). J. Vert. Paleontol. 36:e1103247. doi: 10.1080/02724634.2016.1103247
Geraads, D., and Zouhri, S. (2021). A new late Miocene elasmotheriine rhinoceros from Morocco. Acta Palaeontol. Polon. 66, 753–765. doi: 10.4202/app.00904.2021
Goloboff, P. A., and Catalano, S. A. (2016). TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32, 221–238. doi: 10.1111/cla.12160
Goloboff, P. A., Farris, J. S., and Nixon, K. C. (2008). TNT, a free program for phylogenetic analysis. Cladistics 24, 774–786. doi: 10.1111/j.1096-0031.2008.00217.x
Grand, A., Corvez, A., Duque Velez, L. M., and Laurin, M. (2013). Phylogenetic inference using discrete characters: performance of ordered and unordered parsimony and of three-item statements. Biol. J. Linn. Soc. 110, 914–930. doi: 10.1111/bij.12159
Gregory, W. K., and Cook, H. J. (1928). New materials for the study of evolution: a series of primitive fossil rhinoceros skulls (Trigonias) from the lower Oligocene of Colorado. Proc. Colorado. Mus. Nat. Hist. 7, 1–32.
Heissig, K. (1969). Die Rhinocerotidae (Mammalia) aus der oberoligozänen Spaltenfüllung von Gaimersheim bei Ingolstadt in Bayern und ihre phylogenetische Stellung. Abh. Bayer. Akad. Wiss., Math. Naturwiss. Kl., N. F. 138, 1–133.
Heissig, K. (1972). Paläontologische und geologische Untersuchungen im Tertiär von Pakistan. 5. Rhinocerotidae (Mamm.) aus den unteren und mittleren Siwalik-Schichten. Abh. Bayer. Akad. Wiss., Math. Naturwiss. Kl., N. F. 152, 1–112.
Heissig, K. (1973). Die Unterfamilien und Tribus der recenten und fossilen Rhinocerotidae (Mammalia). Säugetierkundl. Mitt. 21, 25–30.
Heissig, K. (1989). “Rhinocerotidae” in The Evolution of Perissodactyls. eds. D. R. Prothero and R. M. Schoch (New York: Oxford University Press), 399–417.
Heissig, K. (1999). “Family Rhinocerotidae” in The Miocene Land Mammals of Europe. eds. G. E. Rössner and K. Heissig (München: Verlag Dr. Friedrich Pfeil), 175–188.
Heissig, K. (2012). “Les Rhinocerotidae (Perissodactyla) de Sansan” in Mammifères de Sansan. eds. S. Peigné and S. Sen (Paris: Muséum national d’Histoire naturelle, Publications scientifiques du Muséum), 317–485.
Heissig, K. (2017). Revision of the European species of Prosantorhinus Heissig, 1974 (Mammalia, Perissodactyla, Rhinocerotidae). Foss. Impr. 73, 236–274. doi: 10.2478/if-2017-0014
Hooijer, D. A. (1966). Fossil mammals of Africa 21: Miocene rhinoceroses of East Africa. Bull. Brit. Mus. Nat. Hist. Geol. Ser. 13, 117–190.
Hooijer, D. A. (1971). A new Rhinoceros from the late Miocene of Loperot, Turkana district, Kenya. Bull. Mus. Comp. Zool. 142, 339–392.
Hünermann, K. A. (1989). Die Nashornskelette (Aceratherium incisivum Kaup 1832) aus dem Jungtertiär vom Höwenegg im Hegau (Südwestdeutschland). Andrias 6, 1–116.
Kaup, J. (1832). Über Rhinoceros incisivus Cuv., und eine neue Art Rhinoceros schleiermacheri. Isis von Oken 8, 898–904.
Lihoreau, F., Ducrocq, S., Antoine, P.-O., Monique, V.-L., Rafaÿ, S., Garcia, G., et al. (2009). First complete skulls of Elomeryx crispus (Gervais, 1849) and of Protaceratherium albigense (Roman, 1912) from a new Oligocene locality near Moissac (SW France). J. Vert. Paleontol. 29, 242–253. doi: 10.1671/039.029.0114
Lu, X. K. (2013). A juvenile skull of Acerorhinus yuanmouensis (Mammalia: Rhinocerotidae) from the later Miocene hominoid fauna of the Yuanmou Basin (Yunnan, China). Geobios 46, 539–548. doi: 10.1016/j.geobios.2013.10.001
Lu, X. K. (2014). A survey of Acerorhinus yuanmouensis (Perissodactyla, Rhinocerotidae) from the Late Miocene of the Yuanmou Basin (Yunnan, China) and the Phylogeny of Aceratheriinae. Doctoral Dissertation. Beijing, China: University of Chinese Academy of Science.
Lu, X. K., Zheng, X. T., Corwin, S., and Tan, J. (2016). A skull of Plesiaceratherium gracile (Rhinocerotidae, Perissodactyla) from a new lower Miocene locality in Shandong Province, China, and the phylogenetic position of Plesiaceratherium. J. Vert. Paleontol. 36:e1095201. doi: 10.1080/02724634.2016.1095201
Lu, X., Cerdeño, E., Zheng, X., Wang, S., and Deng, T. (2021). The first Asian skeleton of Diaceratherium from the early Miocene Shanwang Basin (Shandong, China), and implications for its migration route. J. Asian Earth Sci. X:6. doi: 10.1016/j.jaesx.2021.100074
Marjanović, D., and Laurin, M. (2019). Phylogeny of Paleozoic limbed vertebrates reassessed through revision and expansion of the largest published relevant data matrix. PeerJ 6:e5565. doi: 10.7717/peerj.5565
Matthew, W. D. (1918). Contributions to the Snake Creek Fauna, with notes upon the Pleistocene of Western Nebraska American museum expedition of 1916. Bull. Am. Mus. Nat. Hist. 38, 183–229.
Osborn, H. F. (1900). Phylogeny of the rhinoceroses of Europe. Bull. Am. Mus. Nat. Hist. 13, 229–267.
Osborn, H. F. (1904). New Miocene rhinoceroses with revision of known species. Bull. Am. Mus. Nat. Hist. 20, 307–326.
Pavlow, M. (1892). Études sur l’Histoire Paléontologiques des Ongulés, VI. Les Rhinocéridae de la Russie et le développement des Rhinocéridae en général. Bull. Soc. Imp. Naturalistes. Mos. 6, 147–231.
Prothero, D. R., Guérin, C., and Manning, E. (1989). The history of the Rhinocerotoidea in The Evolution of Perissodactyls. eds. D. R. Prothero and R. M. Schoch (New York: Oxford University Press), 321–340.
Prothero, D. R. (2005). The Evolution of North American Rhinoceroses. Cambridge: Cambridge University Press.
Prothero, D. R., Manning, E. M., and Hanson, C. B. (1986). The phylogeny of the Rhinocerotoidea (Mammalia, Perissodactyla). Zool. J. Linnean Soc. 87, 341–366. doi: 10.1111/j.1096-3642.1986.tb01340.x
Prothero, D. R., and Schoch, R. M. (1989). “Classification of the Perissodactyla” in The Evolution of Perissodactyls. eds. D. R. Prothero and R. M. Schoch (New York: Oxford University Press), 530–537.
Qiu, Z. X., and Xie, J. Y. (1997). A new species of Aprotodon (Perissodactyla, Rhinocerotidae) from Lanzhou Basin, Gansu, China. Vert. PalAsiat. 35, 250–267.
Qiu, Z. X., and Yan, D. (1982). A horned Chilotherium skull from Yushe, Shansi. Vert. PalAsiat. 20, 122–132.
Répelin, J. (1917). Études paléontologiques dans le sud-ouest de la France (Mammifères)–Les Rhinocérotidés de l’Aquitanien supérieur de l’Agenais (Laugnac). Ann. Mus. Hist. Nat. Marseille 16, 1–45.
Rineau, V., Grand, A., Zaragüeta, R., and Laurin, M. (2015). Experimental systematics: sensitivity of cladistic methods to polarization and character ordering schemes. Contr. Zool. 84, 129–148. doi: 10.1163/18759866-08402003
Rineau, V., Zaragüetai Bagils, R., and Laurin, M. (2018). Impact of errors on cladistic inference: simulation-based comparison between parsimony and three-taxon analysis. Contr. Zool. 87, 25–40. doi: 10.1163/18759866-08701003
Roman, M. F. (1912). Les rhinocérotidés de l’Oligocène d’Europe. Arch. Mus. Sci. Nat. Lyon 11, 1–92.
Scott, W. B. (1941). The mammalian Fauna of the White River Oligocene, Part V, Perissodactyla. Trans. Am. Phil. Soc. 28, 747–980. doi: 10.2307/1005518
Scott, W. B., and Osborn, H. F. (1898). On the skull of the Eocene rhinoceros, Orthocynodon, and the relations of this genus to other members of the group. Contrib. Mus. Geol. Archeol. Princet. Coll. 3, 1–22.
Sizov, A., Klementiev, A., and Antoine, P.-O. (2022). An early Miocene skeleton of Brachydiceratherium Lavocat, 1951 (Mammalia, Perissodactyla) from the Baikal area, Russia, and a revised phylogeny of Eurasian teleoceratines. bioRxiv. doi: 10.1101/2022.07.06.498987
Slowinski, J. B. (1993). “Unordered” versus “ordered” characters. Syst. Biol. 42, 155–165. doi: 10.1093/sysbio/42.2.155
Tissier, J., Antoine, P.-O., and Becker, D. (2020). New material of Epiaceratherium and a new species of Mesaceratherium clear up the phylogeny of early Rhinocerotidae (Perissodactyla). R. Soc. Open Sci. 7:200633. doi: 10.1098/rsos.200633
Wiens, J. J. (2001). Character analysis in morphological phylogenetics: problems and solutions. Syst. Biol. 50, 689–699. doi: 10.1080/106351501753328811
Keywords: Rhinocerotidae, Aceratheriinae, Aceratheriini, Teleoceratini, morphology, phylogeny
Citation: Lu X, Deng T and Pandolfi L (2023) Reconstructing the phylogeny of the hornless rhinoceros Aceratheriinae. Front. Ecol. Evol. 11:1005126. doi: 10.3389/fevo.2023.1005126
Edited by:
Chenyang Cai, Nanjing Institute of Geology and Paleontology (CAS), ChinaReviewed by:
Damien Becker, JURASSICA Museum, SwitzerlandCopyright © 2023 Lu, Deng and Pandolfi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Deng Tao, ✉ ZGVuZ3Rhb0BpdnBwLmFjLmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.