 Wanrong Wei1
Wanrong Wei1 Yan Zhang2,3Zhongmin Tang4Shenghui An4Qiaoyan Zhen1Mingsen Qin1Jundong He5,6*
Yan Zhang2,3Zhongmin Tang4Shenghui An4Qiaoyan Zhen1Mingsen Qin1Jundong He5,6* Maria K. Oosthuizen7,8*
Maria K. Oosthuizen7,8*- 1Key Laboratory of Southwest China Wildlife Resources Conservation, College of Life Sciences, China West Normal University, Nanchong, China
- 2National Demonstration Center for Experimental Grassland Science Education, Lanzhou University, Lanzhou, China
- 3College of Pastoral Agricultural Science and Technology, Lanzhou University, Lanzhou, China
- 4Gannan Grassland Workstation in Gansu Province, Hezuo, China
- 5Institute of Ecology, China West Normal University, Nanchong, China
- 6Administration Bureau of Jiuzhaigou National Nature Reserve, Jiuzhaigou, China
- 7Department of Zoology and Entomology, University of Pretoria, Hatfield, South Africa
- 8Mammal Research Institute, University of Pretoria, Hatfield, South Africa
Vegetation is a crucial component of any ecosystem and to preserve the health and stability of grassland ecosystems, species diversity is important. The primary form of grassland use globally is livestock grazing, hence many studies focus on how plant diversity is affected by the grazing intensity, differential use of grazing time and livestock species. Nevertheless, the impact of the grazing time on plant diversity remains largely unexplored. We performed a field survey on the winter pastures in alpine meadows of the Qinghai-Tibetan Plateau (QTP) to examine the effects of grazing time on the vegetation traits. Livestock species, grazing stocking rates and the initiation time of the grazing were similar, but termination times of the grazing differed. The grazing termination time has a significant effect on most of the vegetation traits in the winter pastures. The vegetation height, above-ground biomass, and the Graminoids biomass was negatively related to the grazing termination time in the winter pastures. In contrast, vegetation cover and plant diversity initially increased and subsequently decreased again as the grazing termination time was extended. An extension of the grazing time did not have any effect on the biomass of forbs. Our study is the first to investigate the effects of grazing during the regrowth period on vegetation traits and imply that the plant diversity is mediated by the grazing termination time during the regrowth period in winter pastures. These findings could be used to improve the guidelines for livestock grazing management and policies of summer and winter pasture grazing of family pastures on the QTP from the perspective of plant diversity protection.
Introduction
Grassland is the largest terrestrial ecosystem on earth and is also classified as one of the most vulnerable systems to environmental stress (Vans et al., 2015). Biological diversity, and in particular plant diversity, is important for the maintenance of the health, function, structure, and stability of grassland ecosystems (Xiong et al., 2016; Hopping et al., 2018; Bennett et al., 2020). Globally, grassland is primarily used as grazing for livestock (Bai et al., 2007; Vermeire et al., 2008; Herrero and Thornton, 2013). Hence, a lot of research has been conducted on the effects of grazing on plant diversity in grasslands in terms of differential use in livestock species, intensity, time, and space (Bai et al., 2007; Vermeire et al., 2008; Herrero and Thornton, 2013; Liu et al., 2019). However, the impact of the grazing time on plant diversity, and more importantly, when it is terminated, has been overlooked. Understanding how plant diversity responds to grazing with different termination times is crucial for the protection of biodiversity and ecosystem function. It also provides valuable insight regarding the role of livestock grazing in grassland degradation (Vermeire et al., 2008; Zhu et al., 2019).
Alpine meadow represents one of the largest grassland ecosystems in the Qinghai-Tibetan Plateau (QTP) (Harris, 2010; Wei et al., 2020a,b). Similar to other grassland ecosystems, alpine meadow ecosystems provide numerous essential ecological functions and services, including pastoral production, water resource regulation, climate control, biodiversity conservation, carbon storage, tourism, and recreation (Strömberg et al., 2013; Wei et al., 2014; Wang et al., 2019). However, during the past few decades, alpine meadows have been degraded to varying degrees (Harris, 2010; Wang et al., 2015), which directly threaten the ecological security of QTP as well as down-stream regions (Dong and Sherman, 2015). Alpine meadow degradation can impair the ecosystem function and structure, thereby reducing the quality of grasslands (Gao et al., 2006, 2019; Wu et al., 2014). Declining biodiversity is one of the most direct manifestations of degrading grasslands (Harris, 2010; Hopping et al., 2018). Several factors can be responsible for alpine meadow degradation, for example rapidly increasing grazing pressure, rodent damage, and climate change (Li et al., 2013; Wei et al., 2019, 2020c). In addition, the mismanagement of livestock grazing is an important factor which can causes alpine meadow degradation (Harris, 2010).
At present, the grassland grazing in the pastoral sector is rotated on a seasonal basis in the QTP in accordance with the household responsibility system (Yan et al., 2005; Wei et al., 2020b). Nomadic herders across the QTP have developed different types of grazing systems under the guidance of governmental policies (Brantingham and Xing, 2006). The most common method of grazing management in the QTP grasslands is nomadic, where livestock moves seasonally between fixed summer and winter pastures (Wei et al., 2020a,b). Under normal circumstances, winter pastures are only grazed during winter (WG, from early October to late April) and is allowed to recover during the summer growing season, whereas the summer pastures are grazed during the growing season (SG, from the end of April to the beginning of October). However, the beginning and end times of grazing on the winter and summer pastures are not uniform since the movement of livestock is determined by the individual herders and depends on when sufficient plant growth has taken place in the summer pastures to sustain the livestock (according to our survey data). The livestock transition from winter pastures to summer pastures overlaps with the regrowth period when plants turn green again after the winter (Yan et al., 2005). Grazing during this time therefore has a significant impact on the plant community composition and structure, as foraging and trampling by livestock will affect the relationship between different plant species and soil properties (Nie and Zollinger, 2012; Selemani et al., 2013). However, few studies focus on the grazing termination time, and our understanding of the relationship between plant community traits and the grazing termination time remains vague (Selemani et al., 2013), especially when the transition between grazing pastures overlaps with the regrowth period of plants.
We investigated the plant community traits of winter pastures in Maqu Country (northeastern QTP), where pastures had a similar grazing onset time, but the termination time differed. We proposed three possible hypotheses with the extension of grazing termination time in winter pastures: (a) extended grazing will decrease vegetation height and coverage since tall grasses with numerous stems and leaves are more susceptible to grazing during the regrowth period; (b) the above-ground biomass and the Graminoids biomass will decrease due to selective feeding by livestock such that grasses and sedges are more likely to be eaten by livestock during the regrowth period; and (c) the suitable grazing during the regrowth period is conducive to a higher plant diversity (plant richness and the Shannon-Wiener diversity) as grazing decreases the vegetation height, which increase light reaching low-growing plants and promoting their growth. We aim to explore how to effectively increase plant diversity in winter pastures under the current grazing model.
Materials and methods
Study site
Our study site is situated at the Hequ racecourse, in Maqu County, eastern QTP in the Gansu Province County, northwestern China, at an average elevation of 3,430°m (Figure 1). The Hequ Racecourse covers an area of 39 866 hectares, has five natural villages and a total of 527 households. This area has a typical alpine continental climate. The mean annual temperature is –2.3°C, and the average annual rainfall is 643.9 mm, that falls predominantly in the summer period (June to September) (Wei et al., 2022). The soil type is an alpine meadow soil that with high organic matter content, thick humus stratum, good soil aggregate structure and sufficient moisture (Wei et al., 2022). The vegetation in this area is dominated by Poaceae, Cyperaceae, Ranunculaceae, and Compositae, such as Kobresia pygmaea, Elymus nutans griseb, Cremanthodium lineare, Anemone rivularis, Leontopium leontopodioides, and Anemone rivularis. Plants started to turn green at the end of April, and yellow at the end of September (Liu et al., 2019). The growing season is approximately 148 days, and the 2020 growing season information originates from the grassland monitoring department. All winter pastures are flat in this region. Our study site is limited to Hequ Racecourse and we believe that the general climatic conditions are the same.
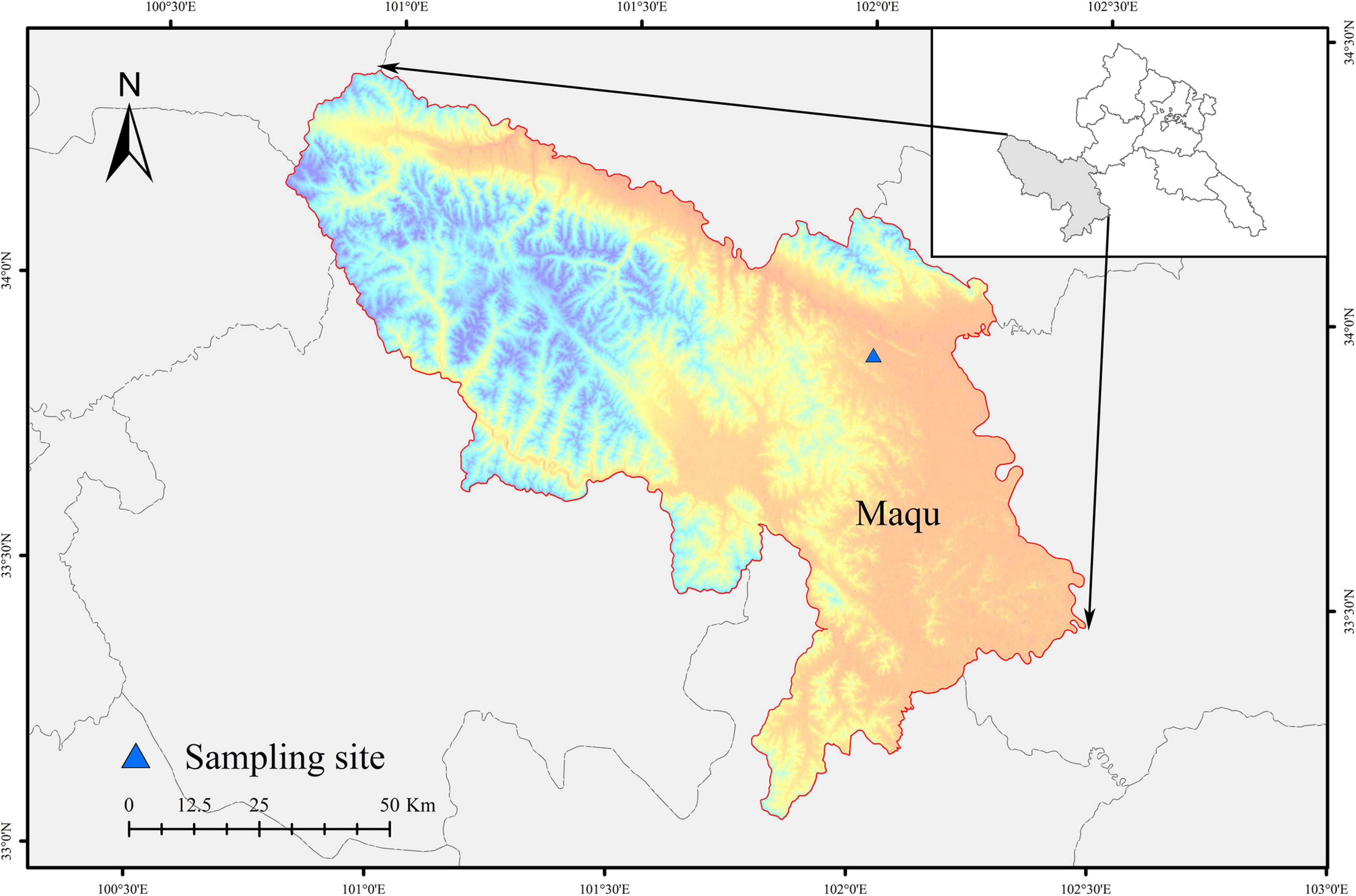
Figure 1. Locations of the study site on the Qinghai-Tibetan plateau.
Experimental design
This study was carried out between April and August of 2020. We conducted a detailed survey of the winter pastures of 214 herdsmen in five natural villages at Hequ racecourse from April to June 2020. We selected a total of 17 winter pastures that meet our requirements in terms of the grazing livestock species, grazing intensity, starting date of grazing (November 22 of 2019), but with a different end date of the grazing (from April to June). We selected the yak (Bos grunniens) as the grazing livestock species, and yak older than 4 years weighs more than 250 kg (Fan et al., 2019; Liu et al., 2019). The winter pastures for all herders are flat and lower in elevation than summer pastures. The grazing intensity was limited between 3.5 and 4.8 head of yak ha–1, which was calculated based on the pasture area and the number of yaks older than 4 years. In this region, this is a moderate grazing intensity as calculated by the grazing intensity classification standard (Xiao et al., 2019). In the five natural villages on the Hequ racecourse, 2, 3, 6, 4, and 2 family pastures met our requirements per village, respectively. Information regarding the livestock species, grazing intensity and the starting and ending dates of grazing from 2019 to 2020 was provided by herders.
Plant sampling
The vegetation survey was conducted in early August 2020. Vegetation was randomly sampled from ten quadrats (50 × 50°cm) in each winter pasture of a selected herder. The interval between quadrats was greater than 100 meters and the quadrats were representative of the vegetation composition and community of this winter pasture. We measured the vegetation height and cover, species richness, species abundance and above-ground biomass. The height of the vegetation was measured by selecting 20 random plants and averaging their heights. Vegetation cover was assessed with the point frame method. A 50°cm long frame with pins at 5 cm intervals was placed over the vegetation and all the plants that touched a pin were identified (Guo et al., 2012a). Species richness was evaluated by recording the number of the species in each of the 0.25°m2 quadrats (Guo et al., 2012b). The abundance of a species was determined by the number of plants of that species in a quadrat. Finally, the plants in each quadrat were harvested, sorted into grass, sedges and forb, brought back to the lab where it was dried at 80°C for 24 h and then weighed. The total above-ground biomass was calculated as the collective biomass of the plants in a quadrat.
Calculation method
The Shannon-Wiener’s diversity index was calculated with the following formula:
H = –Pilog10Pi
where Pi is the proportion of plants of the i-th species.
Pielou eveness index (J) was determined as:
J = H/lnS
With H as the Shannon-Wiener’s diversity index, and S as the species richness.
The plant biomass was divided into three functional groups namely Grass, Sedge, and Forbs (Guo et al., 2012a). Grass and Sedge are the preferred forage of livestock, therefore we calculated the Graminoid biomass as the combination of grasses and sedges biomass.
Data analysis
All data was normally distributed. We used a linear regression to determine whether different grazing termination times significantly altered the vegetation traits (vegetation height, vegetation cover, species richness, above-ground biomass, Shannon-Wiener’s diversity, and Pielou evenness), the biomass of three functional groups, and Graminoids biomass. The grazing termination time was used as the independent variable; and the vegetation traits, the biomass of the three functional groups and Graminoid biomass as dependent variables. Simple linear and polynomial regression models were compared with a likelihood ratio test to select the final regression models. Regression models were employed to determine the relationship between the grazing termination time and the vegetation traits, biomass of the three functional groups, and the Graminoid biomass, and the fit was assessed using the coefficient of determination (R2). The significance level was maintained at P < 0.05. We used the statistical program SPSS19.0 (SPSS, Chicago, IL, United States) to perform statistical analyses while the graphs were produced in Sigmaplot 12.5.
Results
Vegetation height and coverage
Both the vegetation height and cover was strongly influenced by the termination time of the grazing in winter pastures (Figures 2A,B). The vegetation height showed a significant negative linear relationship with the time grazing ended (Figure 2A). However, vegetation cover showed a significant unimodal curvilinearity as the end of the grazing time increased (Figure 2B).
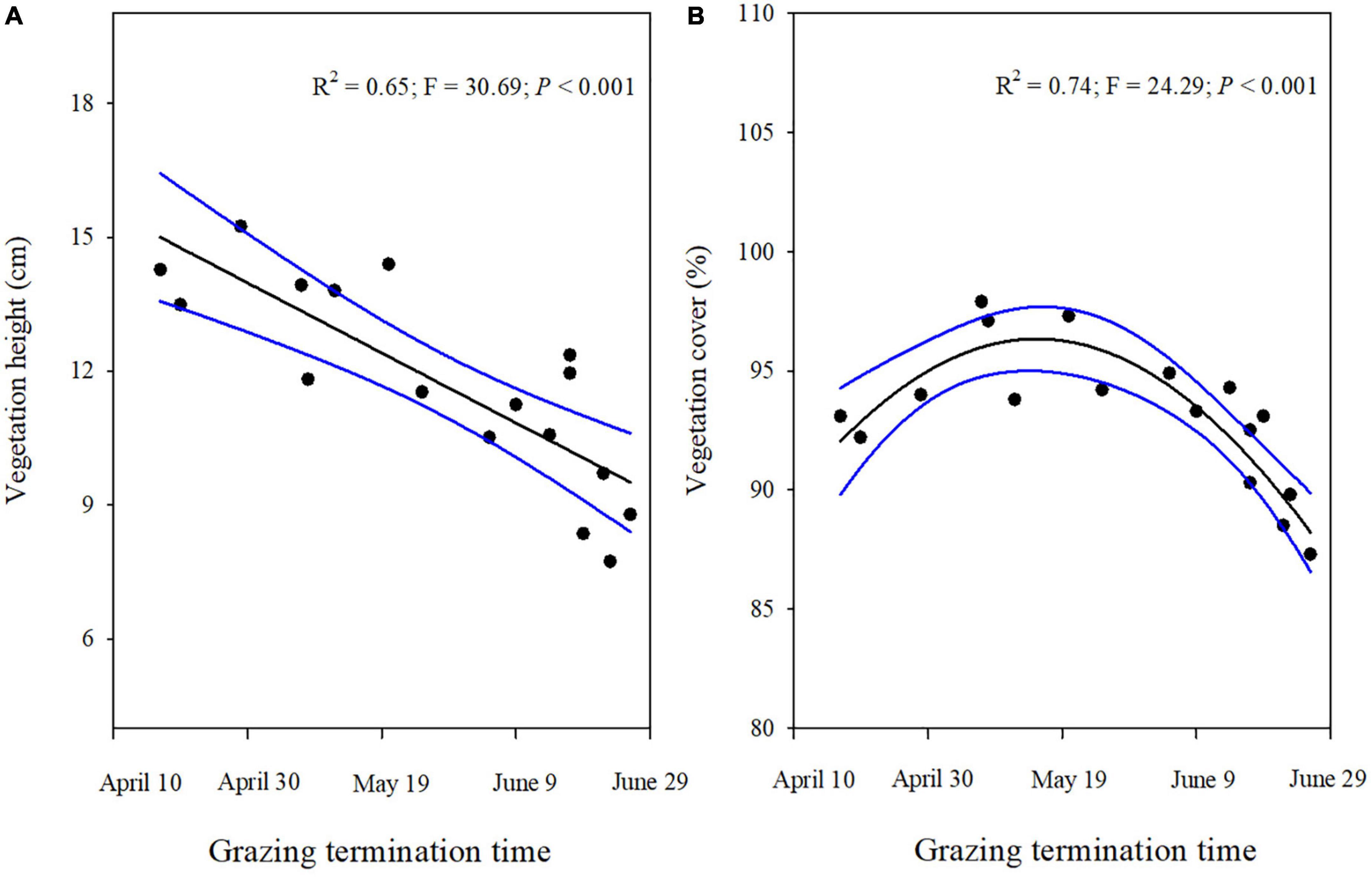
Figure 2. Vegetation height (A) and Vegetation cover (B) for different grazing termination times on winter pastures in alpine meadows based on linear regressions. The black lines indicate significant regression relationships and blue lines indicate the smoothed regressions with its 95% confidence intervals.
Biomass of grasses, sedges and forbs, and graminoid biomass and the total biomass
Grass and sedge biomass showed a significant decrease as the termination time of grazing increased (Figures 3A,B), but the forbs biomass did not change when grazing time was extended in winter pastures (Figure 3C). Similar to changes in the grass and sedge biomass, the graminoids biomass also decreased linearly the longer the livestock was allowed to graze in winter pastures (Figure 4A). In addition, the above-ground biomass showed a significant negative linear relationship with the time that grazing ended (Figure 4B).
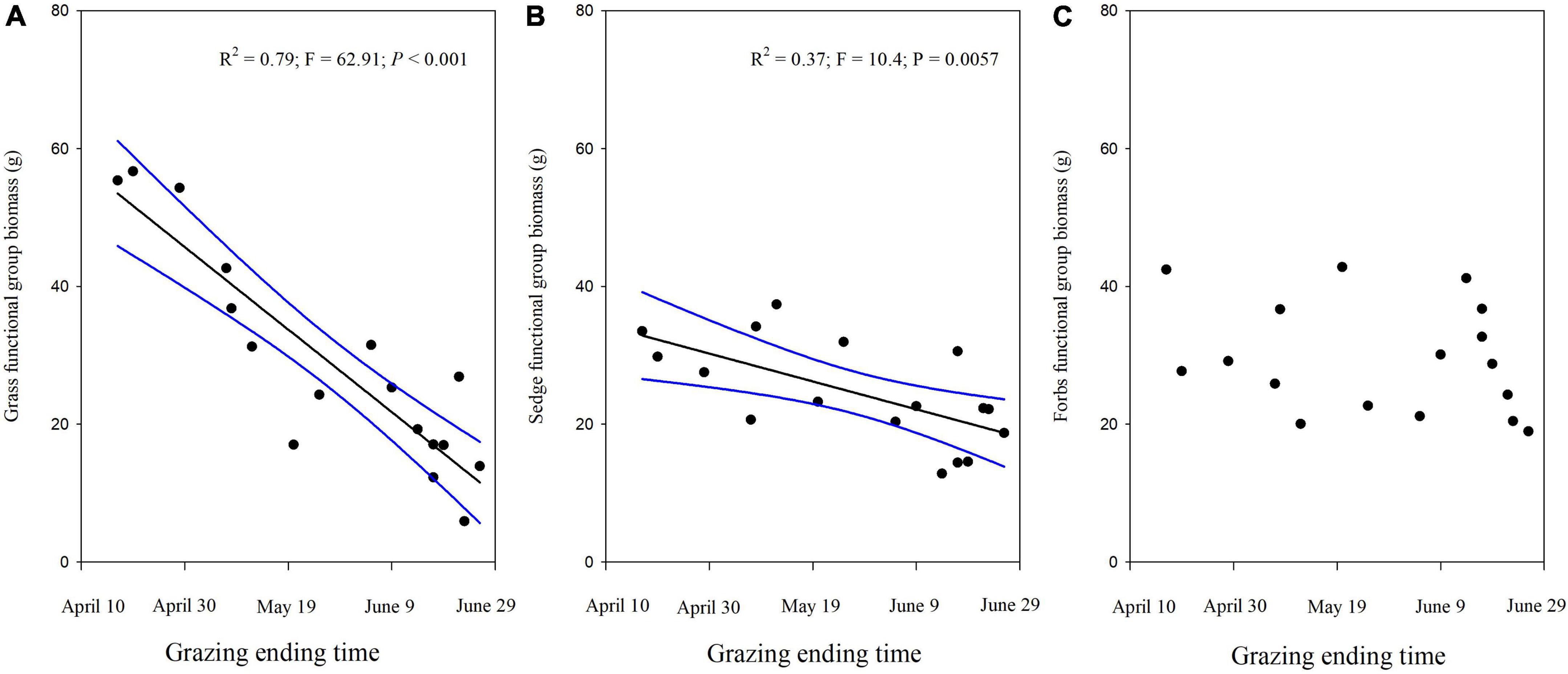
Figure 3. Biomass of the three functional groups (A) Grass biomass. (B) Sedge biomass. (C) Forbs biomass for different grazing termination times on winter pastures in alpine meadows based on a linear regression. The black lines indicate significant regression relationships and blue lines indicate the smoothed regressions with its 95% confidence intervals.
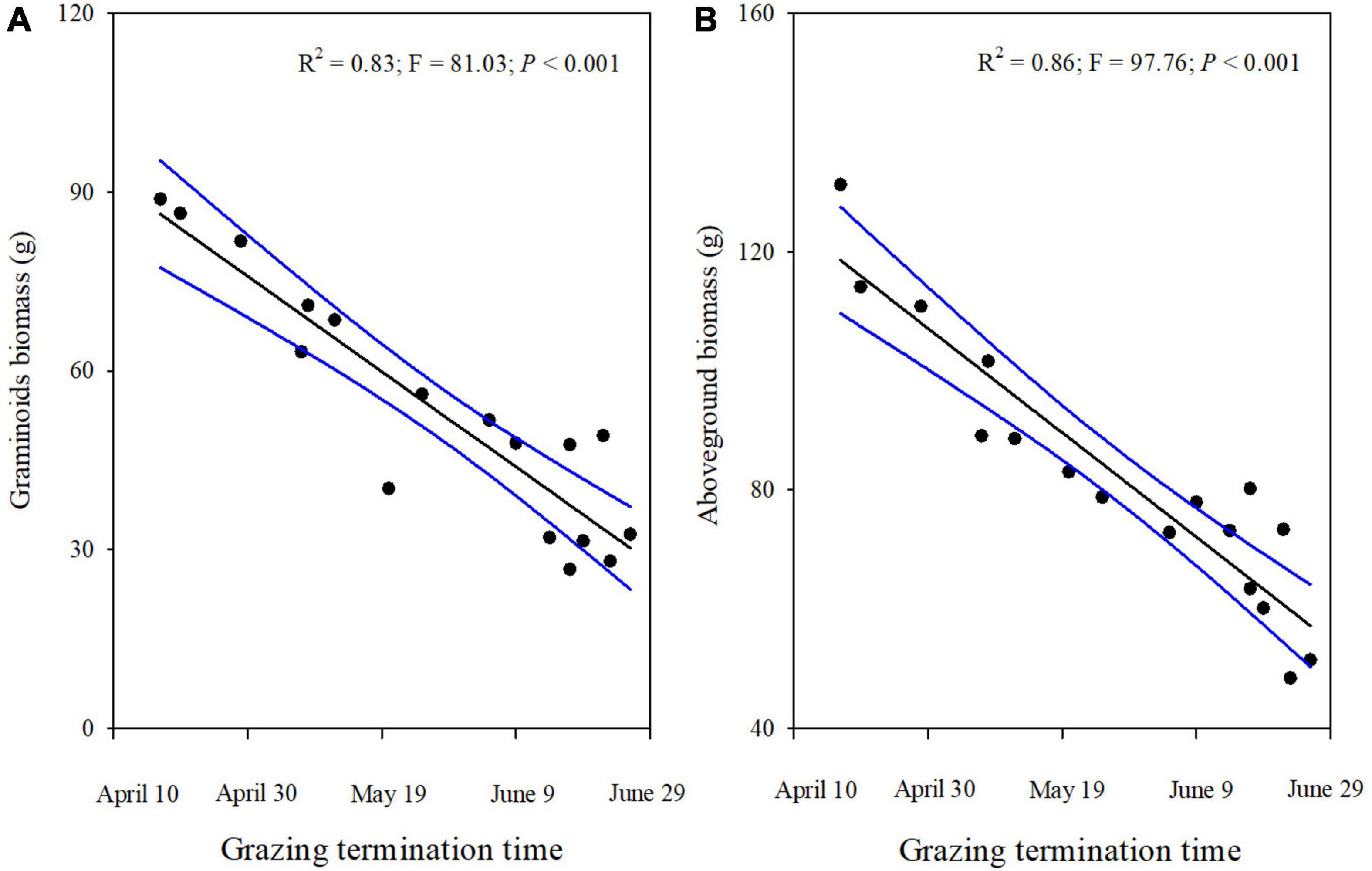
Figure 4. The Graminoid biomass (A) and the aboveground biomass (B) for different grazing termination times on winter pastures in alpine meadows, according to a linear regression. The black lines indicate significant regression relationships and blue lines indicate the smoothed regressions with its 95% confidence intervals.
Plant diversity (species richness and Shannon-Wiener’s diversity) and Pielou evenness
The plant diversity (species richness and Shannon-Wiener’s diversity) was strongly influenced by the end of the grazing time in winter pastures (Figures 5A,B). However, the Pielou evenness was not significantly correlated with grazing termination time (Figure 5C). Species richness and the Shannon-Wiener index showed a significant unimodal curvilinearity as the end of the grazing time increased (Figures 5A,B). As the grazing time was extended, species richness and Shannon diversity initially increased and subsequently decreased, with peaks on May 23 and May 29, respectively.
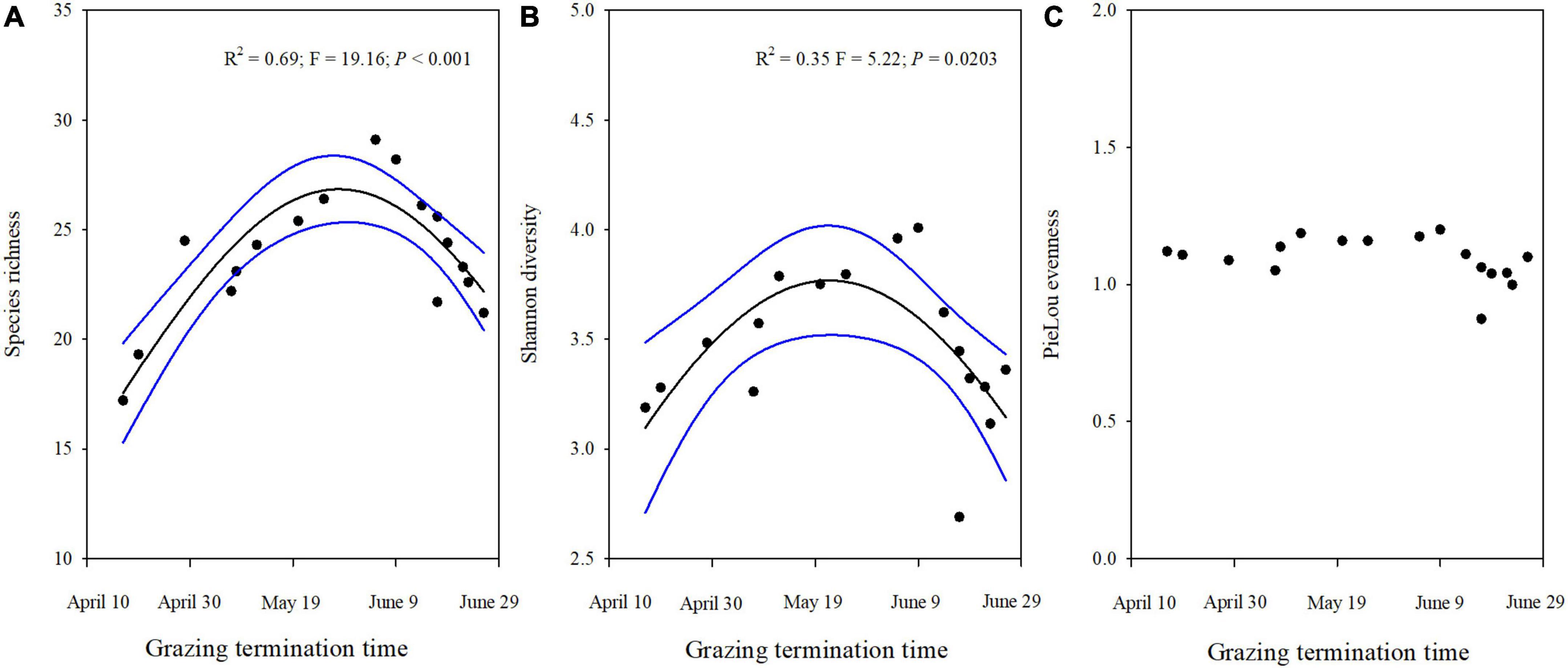
Figure 5. Plant diversity (A) Species richness; (B) Shannon diversity, and PieLous evenness (C) for different grazing termination times on winter pastures in alpine meadows based on linear regressions. The black lines indicate significant regression relationships and blue lines indicate the smoothed regressions with its 95% confidence intervals.
Discussion
The influence of grazing termination time on vegetation composition, structure, and biomass
Grasslands represent some of the largest ecosystems worldwide and its primary land use is grazing. Grazing can modify the composition and productivity of grasslands, and thus have to be carefully managed to ensure that plant diversity is maintained (Vermeire et al., 2008). In this study, we provide the first comprehensive evaluation of the relative consequences of the grazing termination time on community traits in winter pastures. Our results indicate that the decrease in plant diversity may be caused by winter grazing being terminated either too early or too late during the plant regrowth period.
We show distinct patterns in the vegetation traits corresponding to the grazing termination time in winter pastures. This supports the hypothesis that the grazing termination time during plant regrowth period has a powerful effect on vegetation traits and that the extension of the grazing time results in lower height, lower above-ground biomass, lower Grass and Sedge biomass and much-reduced Graminoids biomass. Our results fill a paucity in knowledge regarding the effects of the grazing termination time on vegetation traits.
As we expected, vegetation height decreased with the extension of the grazing time in winter pastures. This is caused by the decreased height of tall dominant species such as grasses and sedges as a result of livestock foraging and trampling (Wang et al., 2019). However, the vegetation cover first showed an increase, that subsequently decreased again as the grazing time extended into the regrowth period, which is inconsistent with our first hypothesis. The appropriate extension of grazing time in the regrowth period is beneficial for the increase of the vegetation coverage in winter pastures and may be the result of several factors. Firstly, it limits the growth of tall plants such as grasses and sedges because they are the preferred by livestock, this can relieve the competitive pressure on low-growing plants (Yang et al., 2019; Filazzola et al., 2020) and increase the growth potential of the shorter plants in the community (Wei et al., 2019). Secondly, the foraging and trampling by livestock often decreases the upward growth of tall plants while encouraging the production of auxiliary shoots (Wang Q. et al., 2018). Thirdly, opportunistic species, in particular broad-leaved plants, are encouraged to germinate (Yang et al., 2019; Yao et al., 2019). The increased plant height when grazing ends too early may promote a self-thinning effect that decreases the density of the plants (Yao et al., 2019), thereby resulting in a decrease in vegetation coverage. The vegetation coverage decreased when the grazing time exceeds a certain range as a result of the decrease in the proportion of grasses with lush stems and leaves since livestock grazing and trampling can cause failure to germinate in some plants (Kattenborn et al., 2017; Wang Q. et al., 2018).
This study illustrates that grazing termination time affects the above-ground biomass and the Graminoids biomass significantly, and this finding supports the second hypothesis. It is clear from our results that above-ground biomass and the Graminoids biomass decreased as a result of the decline in grasses and sedges (Figures 3A,B, 4A,B), but not the forbs biomass (Figure 3C). The regrowth period is the primary period when the forages grow and its nutrients supply mainly depends on the root reserve (Buttolph and Coppock, 2004), especially for grasses and sedges which provides a competitive advantage. Continuous grazing will inevitably quickly deplete the root nutrient accumulation, and it will also lead to insufficient new leaves, and the photosynthetic index of the new leaves will not be able to meet the growth needs of the plant itself during this critical growth period (Buttolph and Coppock, 2004; Nie and Zollinger, 2012). The decrease in the grass and sedge biomass is thought to be due to the selective ingestion by the Tibetan yak (Wang et al., 2019). Interestingly, the forbs biomass did not change significantly with the extension of the grazing time in winter pasture. Theoretically, the forbs biomass should increase significantly as the biomass of dominant plants such as grasses and sedges decrease (Yang et al., 2019), but this is not the case. This may be because the forbs are low growing plants with limited competitiveness and occupies little space compared to grasses and sedges (Wang Q. et al., 2018), and resulting in little change in its biomass.
Plant diversity was assessed by the species richness and the Shannon-Wiener diversity index. Our results indicated that the species richness and the Shannon-Wiener diversity index had a parabolic shape, showing that beyond a certain point, the species diversity steadily decreases as the grazing termination time is extended in winter pastures, in agreement with our third hypothesis. The pattern of the plant diversity may result from increased plant competition in the grassland with shorter grazing times. Early in the growing season, plants are all short and do not have to compete for light and nutrients. However, in time, taller plants may limit these resources for shorter-statured plants such as forbs and outcompete them (Olff and Ritchie, 1998; Borer et al., 2014). The substantial increase in tall grass and sedge biomass in the grasslands with an earlier end time of winter grazing, supports the increased competition hypothesis (Grime, 1979). The lower plant diversity in the grasslands with an extended grazing time is likely the result of intense feeding and continued livestock trampling that prevent some low-growing plants from germinating, since the overall vegetation height is much lower compared to grasslands where grazing terminated earlier. Furthermore, the relative abundances of the different plant functional groups may also influence the pattern of the plant diversity. The abundance of grasses and sedges decreased with the extension of grazing time in winter pastures. Grasses and sedges are reported to be preferred by yaks (Cai et al., 2015), especially during the regrowth period. The above results suggest that plant diversity was strongly affected by the grazing termination time in winter pastures, implying that the appropriate extension of grazing time in winter pastures is conducive to maximize the protection of plant diversity under specific livestock species and grazing intensity. However, if grazing is terminated too early or too late, the plant diversity is not conserved.
Under the current grazing system, the above-ground biomass and graminoid biomass cannot be used to objectively evaluate whether the grassland is degraded in winter pastures (Zhang et al., 2018; Wei et al., 2019). The above-ground biomass in winter pastures is lower but does not necessarily imply a reduction in productivity (Wei et al., 2020a), it only seems lower because the biomass consumed by livestock during the regrowth period is not taken into account. The same is true for the graminoid biomass. The plant diversity is related to the grazing termination time during the plant regrowth period in winter pastures. Grazing after plants turn green will increase plant diversity (plant richness and Shannon diversity) to varying degrees compared to grazing terminated before plants turn green in winter pastures (Figures 5A,B). This can explain why studies have shown that grassland degradation mainly occurs in summer pastures and four-season pastures from the perspective of plant diversity, while it is rare in winter pastures (Tang and Gavin, 2015; Zhao et al., 2016; Wang et al., 2019; Sainnemekh et al., 2022).
Although our study clearly demonstrates that suitable grazing time during the regrowth period can increase plant diversity in winter pastures, our study has some limitations that should be acknowledged. First, different climatic years may lead to variations in plant diversity (Alatalo et al., 2016; Wang et al., 2021). We surveyed for a single year and did not account for the effect of interannual climatic factors. Second, as with interannual climate change, we did not take into account the effects of microclimate which can affects community traits in alpine regions, such as plant diversity (Stark et al., 2017). Third, soil heterogeneity can also change species diversity (Stark et al., 2017; Dong et al., 2019), grazing affects the physical and chemical properties of soil which can indirectly affect the vegetation community during the plant growth season and may potentially also influence our results. Previous studies found that grassland plant diversity is primarily controlled by light, that is mainly affected by grazing (Hautier et al., 2009; Borer et al., 2014), regardless of soil nutrient and livestock utilization (Borer et al., 2014). However, the effects of climate, microclimate and soil heterogeneity on plant diversity cannot be ignored (Borer et al., 2014; Dong et al., 2019), so further studies would be required to show the contribution of climate, microclimate, and soil heterogeneity on plant diversity under grazing disturbance. A follow-up study could focus on plant diversity under global climate change.
Management implications
In the past, a large volume of research has been devoted to grassland vegetation composition and structure in response to grazing (Herrero and Thornton, 2013; Zhang et al., 2018; Dai et al., 2019; Liu et al., 2019). Nevertheless, the consequences of the grazing termination time in grasslands have not been carefully considered, especially in relation to the plant regrowth period. Overall, we found that the grazing termination time is closely related to the vegetation composition and structure in winter pastures. Our findings show that the grazing termination time has a negative impact on vegetation height, aboveground biomass, grass, sedge, and graminoid biomass, but that forbs biomass is not affected. However, it increases plant diversity when the grazing termination is timed appropriately. Our results are significant for conservation efforts that are aimed at maintaining plant diversity–the appropriate extension of the grazing time into the plant regrowth period is advantageous for the increase of plant diversity in winter pastures.
We recommend that the transition from winter (the winter pastures with elevations between 3,300 and 3,600°m) to summer pastures should be carried out in late May under the family ranch model. Ideally, rotational grazing pastures should be grazed and rested on a regular basis, either on set schedule or intermittently as needed (Li et al., 2019). This implies that the current grazing rotation system used on the QTP is not optimal as it affects the biodiversity of alpine meadows negatively and can have long-term implications for conservation. Many studies have shown that the prohibition of grazing in the growing season aids the recovery of degraded grasslands (Xiong et al., 2016; Guo et al., 2018; Wang Y. et al., 2018). However, our study shows that a complete prohibition of grazing in the growing season is not optimal for the maintenance of plant diversity (Figures 5A,B). Therefore, in addition to minding the grazing intensity, we urge pastoralists and policymakers to be more attentive to the impact of grazing termination times under the rotational grazing system and to implement integrative social-ecological management plans to maintain the sustainability of the Tibetan rangelands.
Data availability statement
The original contributions presented in this study are included in the article/supplementary material, further inquiries can be directed to the corresponding authors.
Author contributions
WW, JH, and MO designed the study, analyzed the data, and revised the manuscript. WW, YZ, JH, ZT, SA, QZ, and MQ conducted the fieldwork. WW wrote the first draft of the manuscript. All authors read, commented, and approved the final manuscript.
Funding
This study was supported by the Fundamental Research Funds of China West Normal University (18Q046 and 18Q051), the Gansu Provincial Science and Technology Program (1054nkcp159), and the Natural Science Foundation of Sichuan (No. 2022NSFSC1708).
Acknowledgments
We thank Xin Chen and Yang Jia for assistance with fieldwork.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alatalo, J. M., Jägerbrand, A. K., and Molau, U. (2016). Impacts of different climate change regimes and extreme climatic events on an alpine meadow community. Sci. Rep. 6:21720. doi: 10.1038/srep21720
Bai, Y. F., Wu, J. G., Pan, Q. M., Huang, J. H., Wang, Q. B., Li, F. S., et al. (2007). Positive linear relationship between productivity and diversity: evidence from the Eurasian Steppe. J. Appl. Ecol. 44, 1023–1034. doi: 10.1111/j.1365-2664.2007.01351.x
Bennett, J. A., Koch, A. M., Forsythe, J., Johnson, N. C., Tilman, D., and Klironomos, J. (2020). Resistance of soil biota and plant growth to disturbance increases with plant diversity. Ecol. Lett. 23, 119–128. doi: 10.1111/ele.13408
Borer, E. T., Seabloom, E. W., Gruner, D. S., Harpole, W. S., Hillebrand, H., Lind, E. M., et al. (2014). Herbivores and nutrients control grassland plant diversity via light limitation. Nature 508:517. doi: 10.1038/nature13144
Brantingham, P. J., and Xing, G. (2006). Peopling of the northern Tibetan Plateau. World Archaeol. 38, 387–414. doi: 10.1080/00438240600813301
Buttolph, L. P., and Coppock, D. L. (2004). Influence of deferred grazing on vegetation dynamics and livestock productivity in an Andean pastoral system. J. Appl. Ecol. 41, 664–674. doi: 10.1111/j.0021-8901.2004.00921.x
Cai, R. J., Wei, W. R., and Zhang, W. G. (2015). The primary investigation of food niche differentiation between livestock and grassland rodents. Pratacul. Sci. 32, 802–808.
Dai, L., Guo, X., Ke, X., Zhang, F., Li, Y., Peng, C., et al. (2019). Moderate grazing promotes the root biomass in Kobresia meadow on the northern Qinghai-Tibet Plateau. Ecol. Evol. 9, 9395–9406. doi: 10.1002/ece3.5494
Dong, S. K., Sha, W., Su, X. K., Zhang, Y., Shuai, L. I., Gao, X., et al. (2019). The impacts of geographic, soil and climatic factors on plant diversity, biomass and their relationships of the alpine dry ecosystems: cases from the Aerjin Mountain Nature Reserve, China. Ecol. Eng. 127, 170–177. doi: 10.1016/j.ecoleng.2018.10.027
Dong, S. K., and Sherman, R. (2015). Enhancing the resilience of coupled human and natural systems of alpine rangelands on the Qinghai-Tibetan Plateau. Rangeland J. 37, i–iii. doi: 10.1071/RJ14117
Fan, Q., Wanapat, M., and Hou, F. (2019). Mineral nutritional status of yaks (Bos Grunniens) grazing on the Qinghai-Tibetan plateau. Animals 9:468. doi: 10.3390/ani9070468
Filazzola, A., Brown, C., Dettlaff, M. A., Batbaatar, A., Grenke, J., Bao, T., et al. (2020). The effects of livestock grazing on biodiversity are multi-trophic: a meta-analysis. Ecol. Lett. 23, 1298–1309. doi: 10.1111/ele.13527
Gao, Q., Li, Y., and Wan, Y. (2006). Grassland degradation in Northern Tibet based on remote sensing data. J. Geogr. Sci. 16, 165–173. doi: 10.1007/s11442-006-0204-201
Gao, X. X., Dong, S. K., Xu, Y. D., Wu, S. N., Wu, X. H., Zhang, X., et al. (2019). Resilience of revegetated grassland for restoring severely degraded alpine meadows is driven by plant and soil quality along recovery time: a case study from the Three-river Headwater Area of Qinghai-Tibetan Plateau. Agr. Ecosyst. Environ. 279, 169–177. doi: 10.1016/j.agee.2019.01.010
Grime, J. P. (1979). Primary strategies in plants. Trans. Bot. Soc. 43, 151–160. doi: 10.1080/03746607908685348
Guo, N., Wang, A., Degen, A. A., Deng, B., Shang, Z., Ding, L., et al. (2018). Grazing exclusion increases soil CO2 emission during the growing season in alpine meadows on the Tibetan Plateau. Atmos. Environ. 174, 92–98. doi: 10.1016/j.atmosenv.2017.11.053
Guo, Z. G., Li, X. F., Liu, X. Y., and Zhou, X. (2012a). Response of alpine meadow communities to burrow density changes of plateau pika (Ochotona curzoniae) in the Qinghai-Tibet Plateau. Acta Ecolog. Sin. 32, 44–49. doi: 10.1016/j.chnaes.2011.12.002
Guo, Z. G., Zhou, X. R., and Yuan, H. (2012b). Effect of available burrow densities of plateau pika (Ochotona curzoniae) on soil physicochemical property of the bare land and vegetation land in the Qinghai-Tibetan Plateau. Acta Ecol. Sin. 32, 104–110. doi: 10.1016/j.chnaes.2012.02.002
Harris, R. B. (2010). Rangeland degradation on the Qinghai-Tibetan Plateau: a review of the evidence of its magnitude and causes. J. Arid Environ. 74, 1–12. doi: 10.1016/j.jaridenv.2009.06.014
Hautier, Y., Niklaus, P. A., and Hector, A. (2009). Competition for light causes plant biodiversity loss after eutrophication. Science 324, 636–638. doi: 10.1126/science.1169640
Herrero, M., and Thornton, P. K. (2013). Livestock and global change: emerging issues for sustainable food systems. Proc. Natl. Acad. Sci. U S A. 110, 20878–20881. doi: 10.1073/pnas.1321844111
Hopping, K. A., Knapp, A. K., Dorji, T., and Klein, J. A. (2018). Warming and land use change concurrently erode ecosystem services in Tibet. Global Change Biol. 24, 5534–5548. doi: 10.1111/gcb.14417
Kattenborn, T., Fassnacht, F. E., Pierce, S., Lopatin, J., Grime, J. P., and Schmidtlein, S. (2017). Linking plant strategies and plant traits derived by radiative transfer modelling. J. Veg. Sci. 28, 717–727. doi: 10.1111/jvs.12525
Li, X. L., Gao, J., Brierley, G., Qiao, Y. M., Zhang, J., and Yang, Y. W. (2013). Rangeland degradation on the Qinghai-Tibet plateau: implications for rehabilitation. Land Degrad. Dev. 24, 72–80. doi: 10.1002/ldr.1108
Li, Y., Dong, S., Gao, Q., Zhang, Y., Liu, S., Swift, D., et al. (2019). Grazing promotes plant functional diversity in alpine meadows on the Qinghai-Tibetan Plateau. Rangeland J. 41, 73–81. doi: 10.1071/RJ18091
Liu, P., Ding, L., Zhou, Y., Jing, X., and Degen, A. A. (2019). Behavioural characteristics of yaks grazing summer and winter pastures on the Qinghai-Tibetan Plateau. Appl. Anim. Behav. Sci. 218:104826. doi: 10.1016/j.applanim.2019.06.007
Nie, Z. N., and Zollinger, R. P. (2012). Impact of deferred grazing and fertilizer on plant population density, ground cover and soil moisture of native pastures in steep hill country of southern Australia. Grass Forage Sci. 67, 231–242. doi: 10.1111/j.1365-2494.2011.00838.x
Olff, H., and Ritchie, M. E. (1998). Effects of herbivores on grassland plant diversity. Trends Ecol. Evol. 13, 261–265. doi: 10.1016/S0169-5347(98)01364-0
Sainnemekh, S., Barrio, I. C., Densambuu, B., Bestelmeyer, B., and Aradóttir, ÁL. (2022). Rangeland degradation in Mongolia: a systematic review of the evidence. J. Arid Environ. 196:104654. doi: 10.1016/j.jaridenv.2021.104654
Selemani, I. S., Eik, L. O., Holand, Ø, Ådnøy, T., Mtengeti, E., and Mushi, D. (2013). The effects of a deferred grazing system on rangeland vegetation in a north-western, semi-arid region of Tanzania. Afr. J. Range For. Sci. 30, 141–148. doi: 10.2989/10220119.2013.827739
Stark, J., Lehman, R., Crawford, L., Enquist, B. J., and Blonder, B. (2017). Does environmental heterogeneity drive functional trait variation? a test in montane and alpine meadows. Oikos 126, 1650–1659. doi: 10.1111/oik.04311
Strömberg, C. A. E., Dunn, R. E., Madden, R. H., Kohn, M. J., and Carlini, A. A. (2013). Decoupling the spread of grasslands from the evolution of grazer-type herbivores in South America. Nat. Commun. 4:1478. doi: 10.1038/ncomms2508
Tang, R., and Gavin, M. C. (2015). Degradation and re-emergence of the commons: the impacts of government policies on traditional resource management institutions in China. Environ. Sci. Policy 52, 89–98. doi: 10.1016/j.envsci.2015.04.023
Vans, D. M., Villar, N., Littlewood, N. A., Pakeman, R. J., Evans, S. A., Dennis, P., et al. (2015). The cascading impacts of livestock grazing in upland ecosystems: a 10- year experiment. Ecosphere 6, 1–15. doi: 10.1890/ES14-00316.1
Vermeire, L. T., Heitschmidt, R. K., and Haferkamp, M. R. (2008). Vegetation response to seven grazing treatments in the Northern Great Plains. Agr. Ecosyst. Environ. 125, 111–119. doi: 10.1016/j.agee.2007.12.003
Wang, C., Gao, Q., and Yu, M. (2019). Quantifying trends of land change in Qinghai-Tibet Plateau during 2001-2015. Remote Sens-basel 11:2435. doi: 10.3390/rs11202435
Wang, J., Yu, C., and Fu, G. (2021). Warming reconstructs the elevation distributions of aboveground net primary production, plant species and phylogenetic diversity in alpine grasslands. Ecol. Indic. 133:108355. doi: 10.1016/j.ecolind.2021.108355
Wang, L., Delgado-baquerizo, M., Wang, D. L., Isbell, F., Liu, J., Feng, C., et al. (2015). A comparison of biodiversity-ecosystem function relationships in alpine grasslands across a degradation gradient on the Qinghai-Tibetan Plateau. Rangeland J. 37, 45–55. doi: 10.1071/RJ14081
Wang, Q., Yu, C., Pang, X. P., Jin, S. H., Zhang, J., and Guo, Z. G. (2018). The disturbance and disturbance intensity of small and semi-fossorial herbivores alter the below-ground bud density of graminoids in alpine meadows. Ecol. Eng. 113, 35–42. doi: 10.1016/j.ecoleng.2018.01.003
Wang, Y., Sun, Y., Wang, Z., Chang, S., and Hou, F. (2018). Grazing management options for restoration of alpine grasslands on the Qinghai-Tibet Plateau. Ecosphere 9:e02515. doi: 10.1002/ecs2.2515
Wei, D., Ri, X., Liu, Y. W., Wang, Y. H., and Wang, Y. S. (2014). Three-year study of CO2 efflux and CH4/N2O fluxes at an alpine steppe site on the central Tibetan Plateau and their responses to simulated N deposition. Geoderma 232, 88–96. doi: 10.1016/j.geoderma.2014.05.002
Wei, W., Knops, J. M., and Zhang, W. (2019). The impact of plateau pikas (Ochotona curzoniae) on alpine grassland vegetation and soil is not uniform within the home range of pika families. Plant Ecol. Divers. 12, 417–426. doi: 10.1080/17550874.2019.1628113
Wei, W. R., He, J. D., and Zheng, Q. Y. (2020a). Plateau pikas (Ochotona curzoniae) at low densities have no destructive effect on winter pasture in alpine meadows. Rangeland J. 42, 55–61. doi: 10.1071/RJ19042
Wei, W. R., He, J. D., Zheng, Q. Y., He, Y. L., and Oosthuizen, M. K. (2020b). A comparison of density estimation methods in plateau pika populations in an alpine meadow ecosystem. J. Mammal. 101, 1091–1096. doi: 10.1093/jmammal/gyaa049
Wei, W. R., Zhen, Q. Y., Tang, Z. M., and Oosthuizen, M. K. (2020c). Risk assessment in the plateau pika (Ochotona curzoniae): intensity of behavioral response differs with predator species. BMC Ecol. 20:41. doi: 10.1186/s12898-020-00309-3
Wei, W. R., Zhen, Q. Y., Deng, J., Yue, H. L., Qin, M. S., and Oosthuizen, M. K. (2022). Grazing during the grassland periop promotes plant species richness in alpine grassland in winter pasture. Front. Plant Sci. 13:973662. doi: 10.3389/fpls.2022.973662
Wu, J., Shen, Z., Shi, P., Zhou, Y., and Zhang, X. (2014). Effects of grazing exclusion on plant functional group diversity alpine grasslands along a precipitation gradient on the Northern Tibetan Plateau. Arct. Ant. Alp. Res. 46, 419–429. doi: 10.1657/1938-4246-46.2.419
Xiao, X., Ge, R. C. D., and Hou, F. J. (2019). Effects of grazing and topography on α diversity and physical properties of the soil of alpine meadow communities in the Qinghai-Tibet Plateau. Pratacul. Sci. 36, 3041–3051. doi: 10.11829/j.issn.1001-0629.2019-0392
Xiong, D. P., Shi, P. L., Zhang, X. Z., and Zou, C. B. (2016). Effects of grazing exclusion on carbon sequestration and plant diversity in grasslands of China-a meta-analysis. Ecol. Eng. 94, 647–655. doi: 10.1016/j.ecoleng.2016.06.124
Yan, Z. L., Wu, N., Dorji, Y., and Ru, J. (2005). A review of rangeland privatization and its implications in the Tibetan Plateau, China. Nomadic Peoples 9, 31–51. doi: 10.3167/082279405781826155
Yang, C. T., Zhang, Y., Hou, F. J., Millner, J. P., Wang, Z. F., and Chang, S. H. (2019). Grazing activity increases decomposition of yak dung and litter in an alpine meadow on the Qinghai-Tibet plateau. Plant Soil 444, 239–250. doi: 10.1007/s11104-019-04272-x
Yao, X., Wu, J., Gong, X., Lang, X., Wang, C., Song, S., et al. (2019). Effects of long term fencing on biomass, coverage, density, biodiversity and nutritional values of vegetation community in an alpine meadow of the Qinghai-Tibet Plateau. Ecol. Eng. 130, 80–93. doi: 10.1016/j.ecoleng.2019.01.016
Zhang, C., Dong, Q., Chu, H., Shi, J., Li, S., Wang, Y., et al. (2018). Grassland community composition response to grazing intensity under different grazing regimes. Rangeland Ecol. Manag. 71, 196–204. doi: 10.1016/j.rama.2017.09.007
Zhao, Y., Feng, D., Yu, L., Wang, X., Chen, Y., Bai, Y., et al. (2016). Detailed dynamic land cover mapping of Chile: accuracy improvement by integrating multi-temporal data. Remote Sens. Environ. 183, 170–185. doi: 10.1016/j.rse.2016.05.016
Keywords: plant diversity, alpine meadow, winter pasture, vegetation traits, grazing
Citation: Wei W, Zhang Y, Tang Z, An S, Zhen Q, Qin M, He J and Oosthuizen MK (2022) Suitable grazing during the regrowth period promotes plant diversity in winter pastures in the Qinghai-Tibetan plateau. Front. Ecol. Evol. 10:991967. doi: 10.3389/fevo.2022.991967
Received: 12 July 2022; Accepted: 26 September 2022;
Published: 18 October 2022.
Edited by:
María Rosa Mosquera-Losada, University of Santiago de Compostela, SpainReviewed by:
Xia Zhichao, Hangzhou Normal University, ChinaHuan Li, Lanzhou University, China
Wangya Han, Nanjing University of Information Science and Technology, China
Copyright © 2022 Wei, Zhang, Tang, An, Zhen, Qin, He and Oosthuizen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jundong He, anVuZGhlQDE2My5jb20=; Maria K. Oosthuizen, bW9vc3RodWl6ZW5Aem9vbG9neS51cC5hYy56YQ==