 Koyo Sato
Koyo Sato Takao Sato
Takao Sato Mugino O. Kubo
Mugino O. Kubo
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 10 October 2022
Sec. Paleoecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.957038
This article is part of the Research Topic Reconstructing Paleodiets: Challenges and Advances View all 15 articles
Sika deer (Cervus nippon) is the most abundant ruminant in the Japanese archipelago and has been the primary hunting target, including during the prehistoric ages. Abundant skeletal remains of sika deer have been excavated from archeological sites of the Jomon periods (ca. 15,000–2,400 BP). We reconstructed the feeding habits of sika deer from the Torihama Shell Midden site in Fukui Prefecture, western Japan. The Torihama site is one of the most well-preserved archeological sites of the Jomon period, and we investigated materials from the layers of the Early Jomon period (ca. 6,000 BP). In this study, we obtained three-dimensional tooth surface texture from the lower molars of excavated deer and measured microwear texture using international surface roughness parameters (ISO 25178-2) to infer their habitat use. Next, we estimated the percentage of grasses in the diet by using the reference dataset for extant sika deer with known diets. The results show that the Torihama deer overlapped with mixed feeding and grazing sika deer populations. Moreover, the proportion of grasses in the diet was estimated to be 50.7% on average but showed a wide range among the Torihama deer. This result implied that Torihama deer were mixed feeders of dicot leaves and grasses and had a flexible diet adapted to the vegetation of its habitat. Our results support the paleoenvironmental estimation that the Torihama Shell Midden site contained mixed vegetation of evergreen, deciduous, and coniferous trees around the Mikata Five Lakes in the Early Jomon period. These findings provide insights into the highly plastic diets of the extant sika deer in the Japanese archipelago.
Sika deer (Cervus nippon), a medium-sized species of deer that inhabit a wide area of East Asia, from Amurland in the north to Vietnam in the south, are distributed in the Japanese archipelago from Hokkaido to the Kerama Islands, where there are five subspecies (Ohtaishi, 1986; Miura, 2008). Sika deer are thought to have migrated from the Asian continent to the Japanese archipelago during MIS12 (approximately 430,000 years ago) (Kawamura, 2009). Although the acidic soil in Japan is not conducive to the preservation organic remains, animal skeletal remains have been found, along with artifacts in prehistoric shell middens, caves, and low wetland sites. Skeletal remains of sika deer and wild boar (Sus scrofa) have often been excavated from those archeological sites, and these animals are thought to be among the major animals hunted during the Jomon period (ca. 15,000–2,400 BP) (Nishimoto, 1991).
From Torihama shell midden, abundant skeletal remains of sika deer have been excavated (Figure 1). This site was occupied by Paleolithic people from the Incipient to Early Jomon period. Because of the environmental condition of wetlands, this site has yielded various artifacts, for example, Jomon pottery and stone and bone tools; botanical artifacts, namely, fibers and wooden tools, represented by a well-preserved comb coated with red lacquer (Torihama Shell Midden Study Group, 1987); and many plant and animal remains. Therefore, Torihama Shell Midden has been one of the most intensively studied archeological sites in Japan. This research has been conducted by an interdisciplinary research team of archeologists and natural science researchers motivated by this slogan, “the reconstruction of the Jomon lifestyle focusing on their subsistence” (Kojima, 2015). Analyses of abundant sika deer and wild boar remains have been conducted based on various objectives and methodologies, for example, species identification and the quantitative analysis of each skeletal element (Morikawa, 1963; Nishida, 1979; Inami, 1983; Shigehara et al., 1991; Uchiyama, 2000; Anezaki et al., 2005; Sato, 2021a), the estimation of hunting seasons (Nishida, 1980; Ohtaishi, 1980; Uchiyama, 2005; Sato, 2021b), the examination of cut marks (Hongo, 1991), and the manufacturing of bone tools (Yamakawa, 1992a,b).
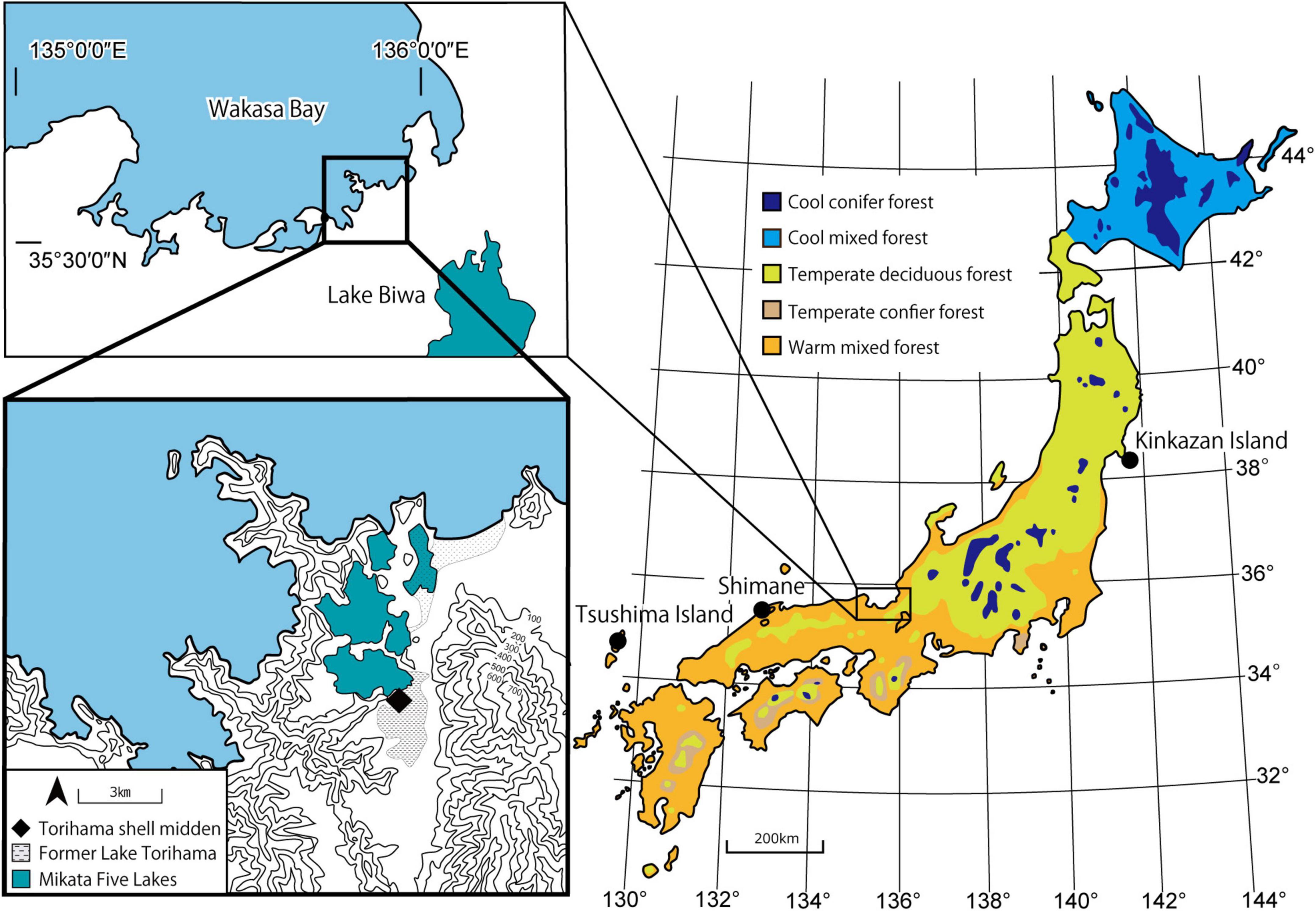
Figure 1. Location of Torihama Shell Midden (♦); location of comparative specimen collection (•). We based the estimated shoreline of the Jomon period and the location of former Lake Torihama on Morikawa (2002) and the present vegetation classification on Yoshioka (1973).
Studies have also been conducted to reconstruct the paleoenvironment and the use of plant resources by analyzing pollen (Yasuda, 1979), seed (Kasahara, 1983, 1984), phytolith (Sasaki, 1983), natural wood (Noshiro and Suzuki, 1990), and wood tools (Noshiro et al., 1996; Kudo et al., 2016a) in the sediments of the site. In addition, pollen analysis of lakes and lowland sediments has provided insights into long-term climatic changes around the site (Yasuda, 1982; Takahara and Takeoka, 1992; Nakagawa et al., 2002, 2005; Kitagawa et al., 2018).
The vegetation around the site in the Early Jomon period was estimated to be evergreen broadleaved forests, consisting of evergreen oak (Quercus subgen Cyclobalanopsis), chinquapin (Catanopisis sieboldii), and camellia (Camellia japonica Linn.) on the top of the hill; Japanese cedar (Cryptomeria japonica) on the hillsides; and deciduous broadleaved forests consisting of willow (Salix) and alder (Alnus sect. Gymnothyrsus) and Japanese ash (Fraxinus) in the waterside low areas (Noshiro and Suzuki, 1990). Subsequently, Yoshikawa et al. (2016) analyzed excavated pollen and seeds; their results suggest that the flora around the site was mixed vegetation: evergreen trees, broadleaved trees, and coniferous trees.
Therefore, clarifying the diets of sika deer in such a vegetation environment would help uncover the habitat use of the deer in the ecosystem and humans’ hunting activities. To estimate the diets of the excavated deer, we applied dental microwear texture analysis (DMTA) for the first time in the Jomon archeological sites. The DMTA measured the surface roughness of microscopic tooth enamel surfaces in three dimensions, obtained by optical profilometers such as confocal microscopes. The application of DMTA has been expanded to various types of vertebrates for dietary estimation (DeSantis, 2016) and has been most intensively applied to herbivores (e.g., Merceron et al., 2010; Berlioz et al., 2017), including extant, extinct, and excavated animals (Kubo and Fujita, 2021).
Because tooth surfaces retain dietary signals for a shorter period (days to weeks) than stable carbon and nitrogen isotopes of soft tissues and bones (Matsubayashi and Tayasu, 2019; Winkler et al., 2020), seasonal differences in a population can be detected, providing insights into hunting seasons and sites occupied by humans (Rivals et al., 2009, 2015). These results reveal human activity but are based on observations at low magnification; thus, they are not applicable to the method used in this study to quantitatively observe the three-dimensional (3D) shape of microwear. Kubo et al. (2017) and Kubo and Fujita (2021) have provided the reference dataset of extant sika deer sampled from various environments with quantitative dietary information; thus, the dental microwear texture (DMT) of the excavated deer can be compared with theirs.
In this study, we estimate the foraging behavior of the hunted sika deer from the Torihama Shell Midden site (hereafter “Torihama deer”) by using DMTA to infer the habitat use of the deer that lived during the Early Jomon period (ca. 6,000 BP).
We used mandibles of the Torihama deer (n = 56) housed in the Wakasa History Museum and analyzed lower second molars (M2), which were not separated from the mandibles (Figure 2 and Supplementary Data 1). According to the correspondence between age in months and eruption of the molars (Niimi, 1997), the mandibles were 29 months old or older. Therefore, we observed the second molars, which had begun to wear down and had relatively few defects. They might have been derived from the same individual because the hemi-mandibles had been excavated in isolated conditions from the archeological site. However, in order to obtain the maximum number of samples, we included the left and right sides in the analysis. They were excavated during the ninth survey in 1984 (Torihama Shell Midden Study Group, 1985) and obtained from sediment layers yielding potteries of the Early Jomon period (Kitashirakawa Kaso II types). The layers, organic soils or shell deposits consisting primarily of freshwater mollusks, were dated to ca. 5,900–5,655 cal BP by radiocarbon dating (Kudo et al., 2016b). The deer remains showed evidence of human use, namely, scars from removal and dismantling and traces of extracting bone marrow and brain (Inami, 1983; Hongo, 1991; Yamakawa, 1992a; Uchiyama, 2000; Sato, 2021a). Additionally, many stone arrowheads, thought to have been used for hunting, were excavated (Torihama Shell Midden Study Group, 1987). These suggested that the bones probably originated from hunted individuals and were disposed of in the lowlands after being used as resources for food and tools. We expected those materials to include dead individuals from all seasons. The Torihama Shell Midden site yielded the evidence of hunting activity throughout the year, though it seemed to be intensively used from fall to winter based on the death-season of sika deer remains which were estimated from observations of antlers, the cementum layer, and mandibular tooth eruption (Nishida, 1980; Ohtaishi, 1980; Sato, 2021b).
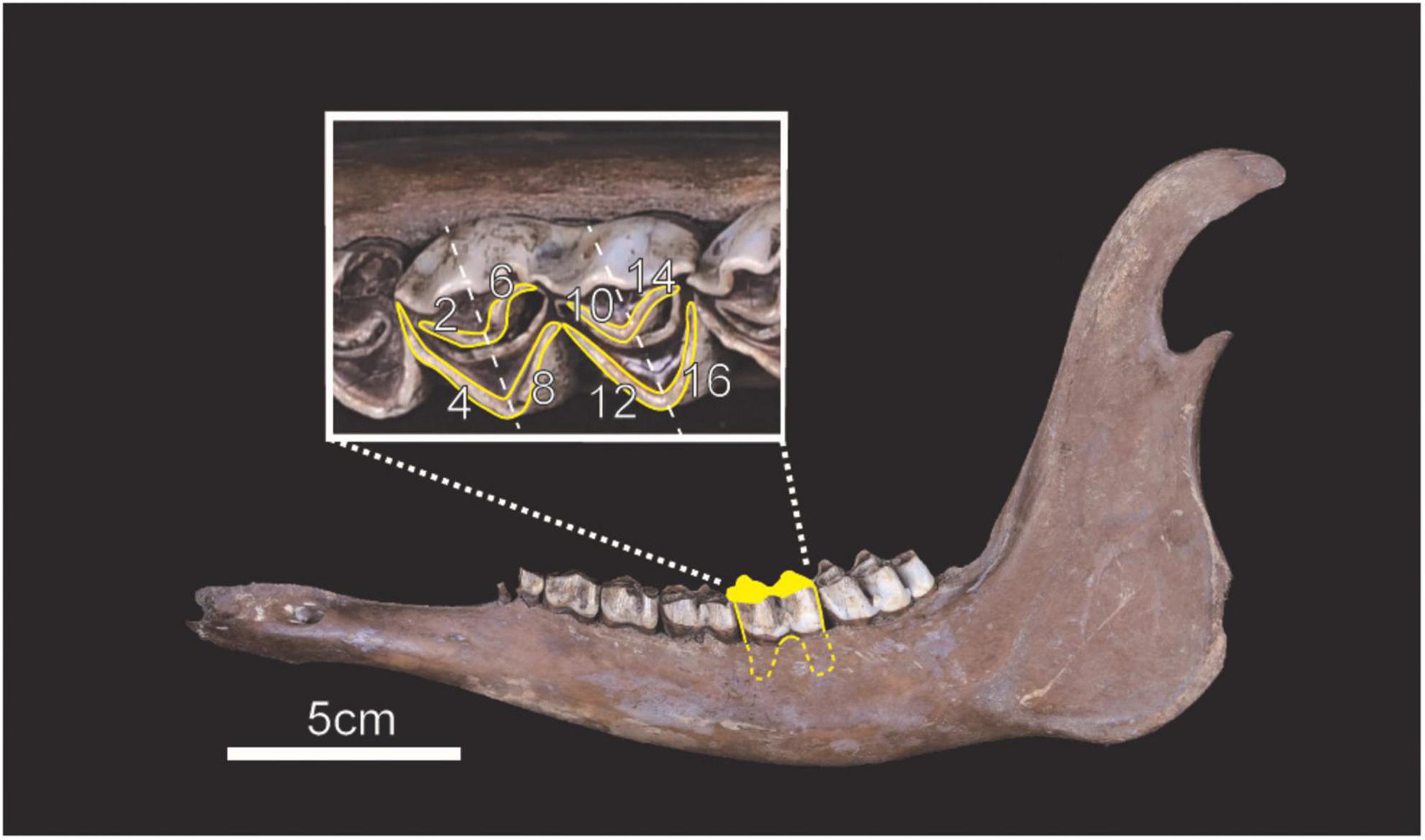
Figure 2. Example of analyzed materials and position of scanned enamel facets. We investigated the occlusal surface of lower second molars enclosed in yellow (i.e., facet No. 2, 4, 6, 8, 10, 12, 14, and 16).
We compared the data from the archeological remains with the published DMT data of extant sika deer. Among the 15 sika deer populations in Japan that Kubo and Fujita (2021) analyzed, we selected three populations (Figure 1 and Table 1): typical grazing (Kinkazan Island), browsing (Tsushima Island), and mixed-feeding (Shimane) populations. The Shimane population was selected because of its habitat’s geological proximity and environmental similarity to the Torihama Shell Midden location. We selected the Kinkazan and Tsushima populations because of their contrasting north–south diets in the Japanese archipelago, and many samples have been analyzed. Sika deer on Kinkazan Island inhabit a small island near the Oshika Peninsula in Miyagi Prefecture, facing the Pacific Ocean. The main vegetation is deciduous broadleaved forest with some open grasslands. Kinkazan deer feed mainly on grasses such as silver grass (Miscanthus sinensis) and lawn grass (Zoysia japonica) (Takatsuki, 1980). The Tsushima deer inhabited an evergreen broadleaved forest. Stomach content analysis revealed that they fed mainly on woody plants, followed by herbaceous plants, seeds, and fruits; graminoids were a minor component of their diet (Suda, 1997). Deer from the Shimane population inhabited the Misen Mountains of Shimane Prefecture, in the San-in region, at the same latitude as the Torihama site. Evergreen broadleaved forests are the primary vegetation in the area. Stomach content analysis of Shimane deer showed that they consumed both graminoids and leaves of broadleaved trees and forbs year-round, although there was seasonal variation (Shimane Prefectural Government, 2002). The dietary differences among the three populations were represented in the percentage of grasses in the diets: 65.8% in Kinkazan (grazers), 38.1% in Shimane (mixed feeders), and 3.4% in Tsushima (browsers) (Kubo and Yamada, 2014; Kubo and Fujita, 2021).
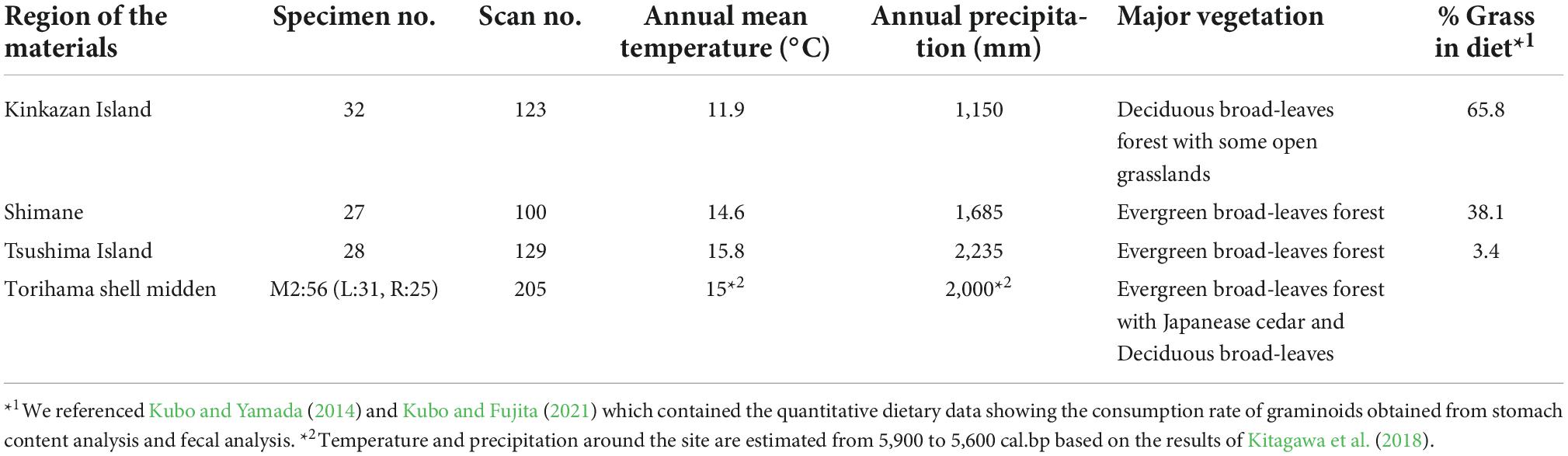
Table 1. Habitat environment and food habits related to the materials in this study.
We used the methodology of Kubo et al. (2017) and made molds of the M2. To remove dirt adhered during the sedimentation, we cleaned the molar occlusal surfaces with cotton swabs soaked with acetone. Next, the surface was molded using high-resolution A-silicone dental impression material (Dr. Silicon regular type, BSA Sakurai, Japan). We selected the occlusal enamel band as our target for scanning (Figure 2) because we expected it to be less affected than other parts by factors other than diet (Schulz et al., 2010).
We scanned the dental impressions by using a confocal laser microscope (VK-9700, Keyence, Osaka, Japan) equipped with a 100× long-distance lens (N.A. = 0.95). Three-dimensional data was obtained with a lateral (x, y) sampling interval of 0.138 μm and a vertical resolution (z) of 0.001 μm. Occlusal enamel facets were scanned on areas of 140 × 105 μm, and grayscale images of the tooth surfaces were acquired for graphical comparison. The 3D surface data were processed by applying the methods in Yamada et al. (2018), Aiba et al. (2019), and Kubo and Fujita (2021), using MountainsMap Imaging Topography (7.4.8872, Digital Surf, Besançon, France). The methods provide an objective evaluation of quantitative data and scratch depth by measuring microwear three-dimensionally. First, we scanned and trimmed the 135 × 100 μm area of the microwear impressions, avoiding dust and large scratches. Because the scanned images were mirror images of real tooth surfaces, the coordinates were mirrored in the x- and z-axes. Second, through the least square plane by subtraction, the surfaces were leveled to remove the inclination of the molds. Next, we applied a robust Gaussian filter with a cutoff value of 0.8 μm to remove measurement noise (S-filter as defined in ISO25178) and the form removal function (polynomial of increasing power = 2) to remove the large-scale curvature of the enamel bands (F-operation in ISO25178). We used the automated outlier removal function of MountainsMap, which removes features with a slope > 80°, and a threshold to remove the upper and lower 0.1% of the data. Scans with a non-measured point above 0.5% of total data points were discarded from the following analyses. The non-measured points were filled by using the smoothing function of MountainsMap. Finally, the values of 31 parameters of ISO 25178-2 were calculated, of which the names and definitions are shown in Table 2. These 31 ISO parameters were used in Kubo and Fujita (2021) to investigate the relationship between DMTA parameters and the extant sika deer diet. Because we used the same analytical template of the same software (MountainsMap) that Kubo and Fujita (2021) used, their published parameters are directly comparable to ours.
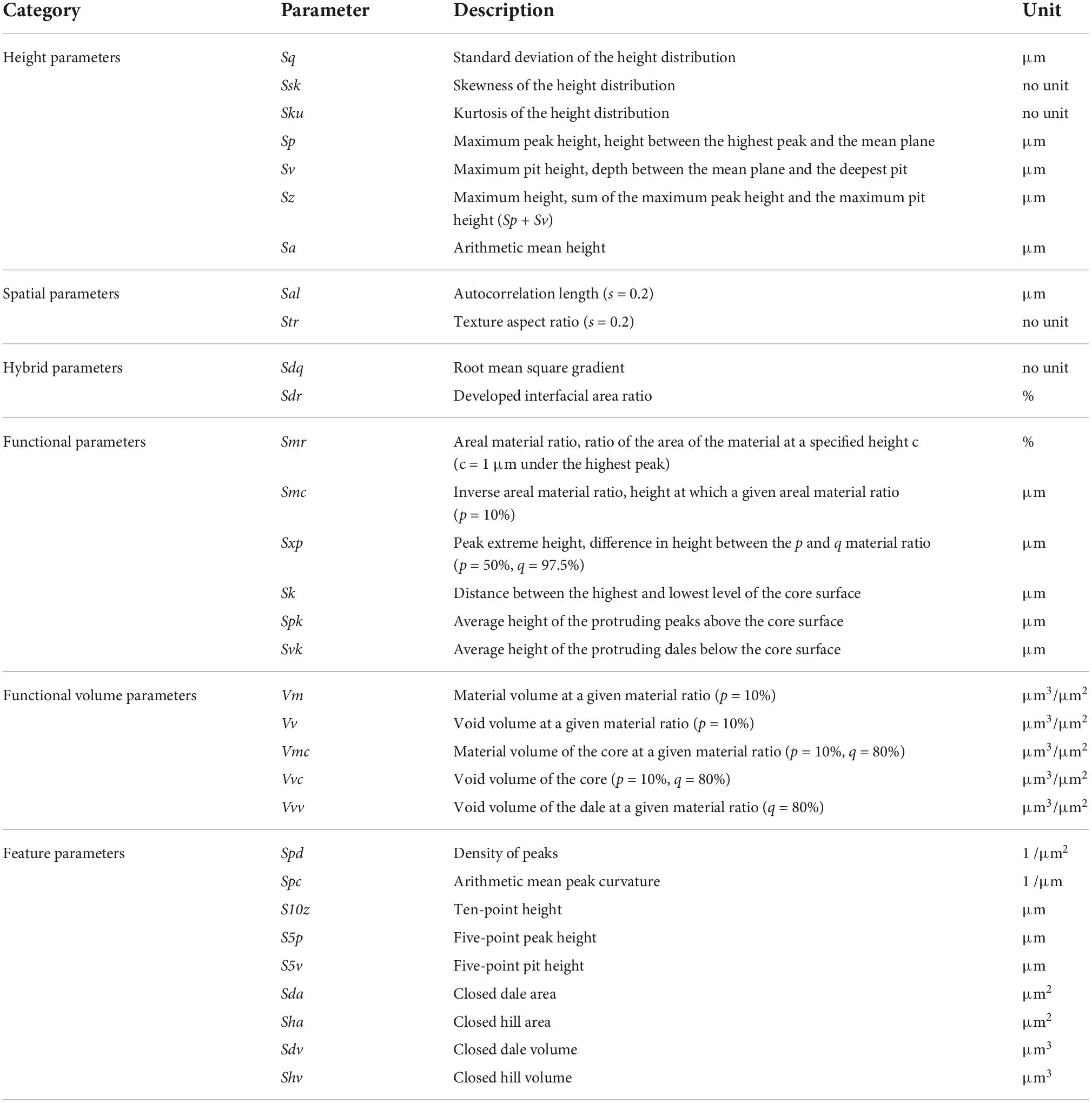
Table 2. Names and definitions of 31 DMT parameters.
To clarify the feeding habits of the Torihama deer, we compared their DMT parameters with those of the three populations of sika deer (Kinkazan Island, Shimane, and Tsushima Island). We conducted principal component analysis (PCA) followed by a varimax rotation to interpret the 31 parameters and summarize them into a few principal components (PCs). We also conducted the Mann–Whitney U-test to determine differences in the parameters and the PC scores between Torihama and the three populations. The statistical analyses were conducted using SPSS statistics ver.28 (IBM, Chicago, USA).
Kubo and Fujita (2021) presented an equation, as follows, to estimate the % grass in diets from an ISO 25178-2 parameter, Sk, which showed the highest correlation with the % grass in the diet in their analysis:
where logit (% grass) is transformed into the % grass by the equation
We used those two equations to estimate the % grass in the diet of 56 Torihama deer.
Representative microwear images and 3D models of the tooth surfaces are shown in Figure 3. Of the two-dimensional (2D) images, we excluded those in which the tooth surface was clearly damaged or the non-measured point was above 0.5% of the total data points. As a result, of the 224 scans (N = 224), we used data from 205 scans (Supplementary Data 1). Although some materials showed heavily worn surfaces, microwear features were well-preserved in the Torihama deer specimens, and the diagenetic alteration was considered negligible. Small pits were commonly observed among the Torihama deer samples. Some samples had flat surfaces with a visible enamel prism structure (Figure 3A: FTS84-086), and others showed abundant scratches (Figure 3B: FTS084-096 and Figure 3C: FTS84-054). Coarse scratches were observed on the surface of some individuals (Figure 3C).

Figure 3. Representative microwear 2D images and 3D models of tooth surfaces of Torihama deer [specimen number (A) FTS084-086, (B) FTS084-096, (C) FTS084-054]. Enamel prism structures can be observed in (A). (B,C) Show show scratches. We also plotted the principal component scores of these specimens in Figure 4. White bars in the 2D micrographs are 10 μm.
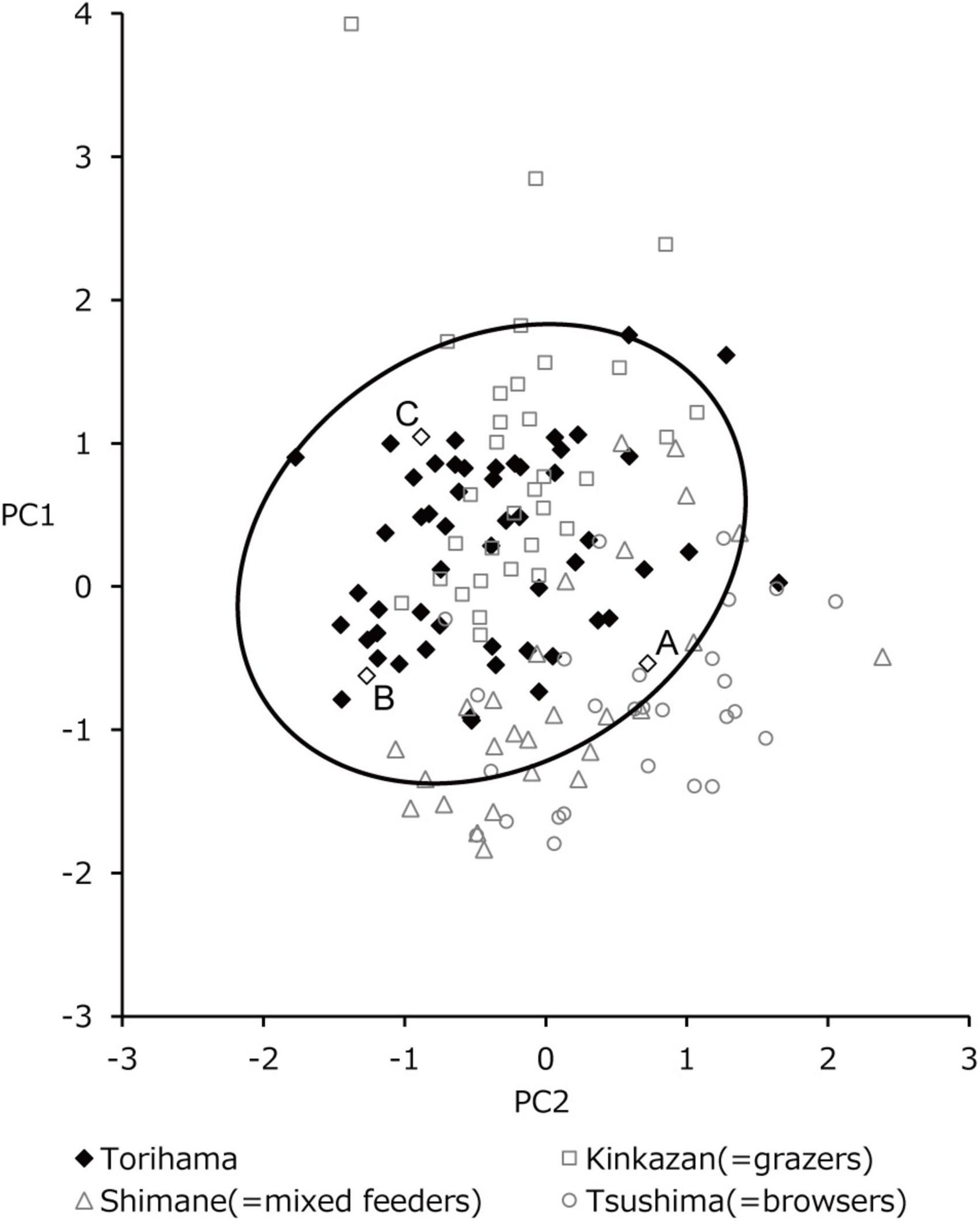
Figure 4. Scatter plot of the first (PC1) and second (PC2) principal component scores calculated by PC analysis using 31 parameters of ISO 25178-2. PC1 is interpreted as overall surface roughness because the height and volume parameters had positive loads. PC2 presents the fineness of the microwear features because the parameters related to surface segmentation had positive loads: larger PC2 values indicate that surfaces are segmented into larger hills and dales. The 95% confidence range of Torihama deer is indicated by the bold line. Kinkazan, Shimane, and Tsushima deer are typical grazers, mixed feeders, and browsers, respectively.
The PCA of the 31 DMT parameters showed that the first, second, and third components explained approximately 52.6, 15.9, and 12.8% of the total variance, respectively. Table 3 indicates the factor loadings of the 31 parameters. The first component (PC1) can be interpreted as overall surface roughness because 22 of the 31 parameters had factor loadings higher than the critical threshold value of 0.4, and 20 parameters were larger than 0.8. The height (e.g., Sq, Sz, Sa) and volume (e.g., Vm, Vv, Vmc, Vvc, Vvv) parameters had positive loads; thus, the larger the PC1 value, the larger the surface relief. On the other hand, parameters related to surface segmentation (Sda, Sha, Sdv, Shv, and Spd) contributed significantly to PC2. The factor loadings of Sda, Sha, Sdv, and Shv, which indicate the areas and volumes of segmented hills and dales on the surface, were positive values, and that of Spd, which indicates the density of the peaks, was negative. Therefore, PC2 can be interpreted as the fineness of the microwear features, with the smaller values indicating a surface characterized by abundant fine features.
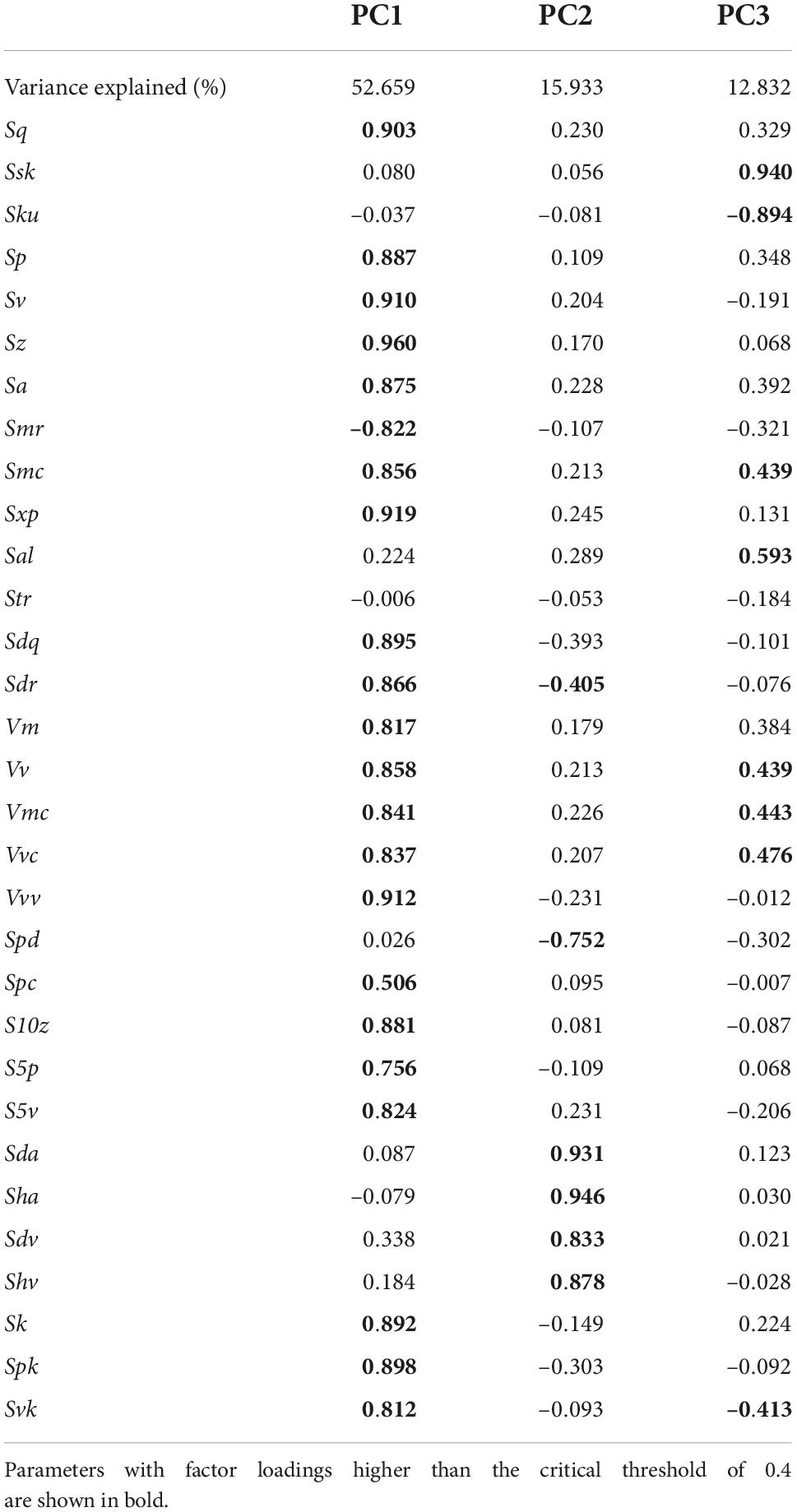
Table 3. Factor loadings obtained from PCA conducted using 31DMT parameters.
We found significant differences between the Torihama deer and the three populations of extant deer (P < 0.05; Table 4). The comparison with the Kinkazan deer showed statistically significant differences in 23 parameters, except for some hybrid, functional, and feature parameters. Statistically significant differences between the Torihama deer and the Shimane deer were found in 21 parameters, excluding the spatial, functional volume, and feature parameters. The largest number of statistical differences (28 parameters) was found in the comparison between the Torihama deer and the Tsushima deer, indicating that the difference between the Torihama deer and the Tsushima deer is larger than that between the deer in Kinkazan and Shimane.
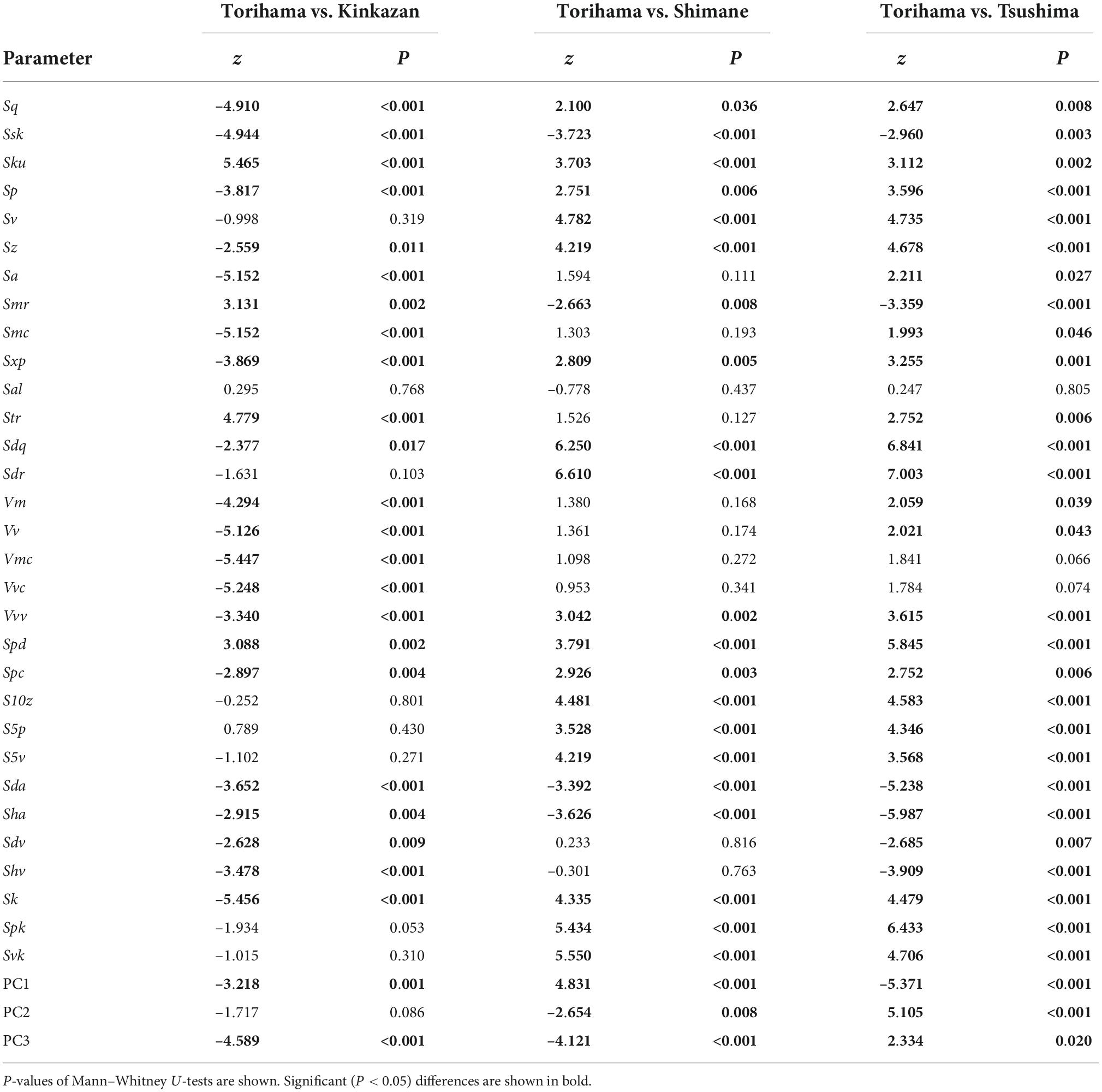
Table 4. Comparison of DMT parameters and principal component scores between Torihama deer and extant deer.
The results of the statistical comparison of the PC scores between the Torihama and extant deer populations are also shown in Table 4. A scatter plot of PC1 and PC2 scores are shown in Figure 4 for the populations of Torihama, Shimane, Kinkazan, and Tsushima. Deer on Kinkazan Island, typical grazers, were characterized by higher PC1 scores (i.e., large surface roughness), whereas those deer on Tsushima Island, typical browsers, were located at the lower bottom. The Shimane population, mixed feeders, was located between them. Thus, we found a separation between the grazing, mixed feeding, and browsing populations of the extant deer, although the Shimane and Tsushima populations overlapped. The Torihama deer overlapped with the distribution of the grazing and mixed-feeding sika deer but not with the browsing population.
We applied the estimation equation of Kubo and Fujita (2021) to estimate the % grass in the diet of 56 Torihama deer (Figure 5). The Torihama deer were estimated to have consumed a diet of, on average, 50.7% grass, which supported a general mixed-feeding habit of the Torihama deer. Browsing and grazing individuals occurred with a similar frequency, and there was a wide range in % grass within the Torihama deer population.
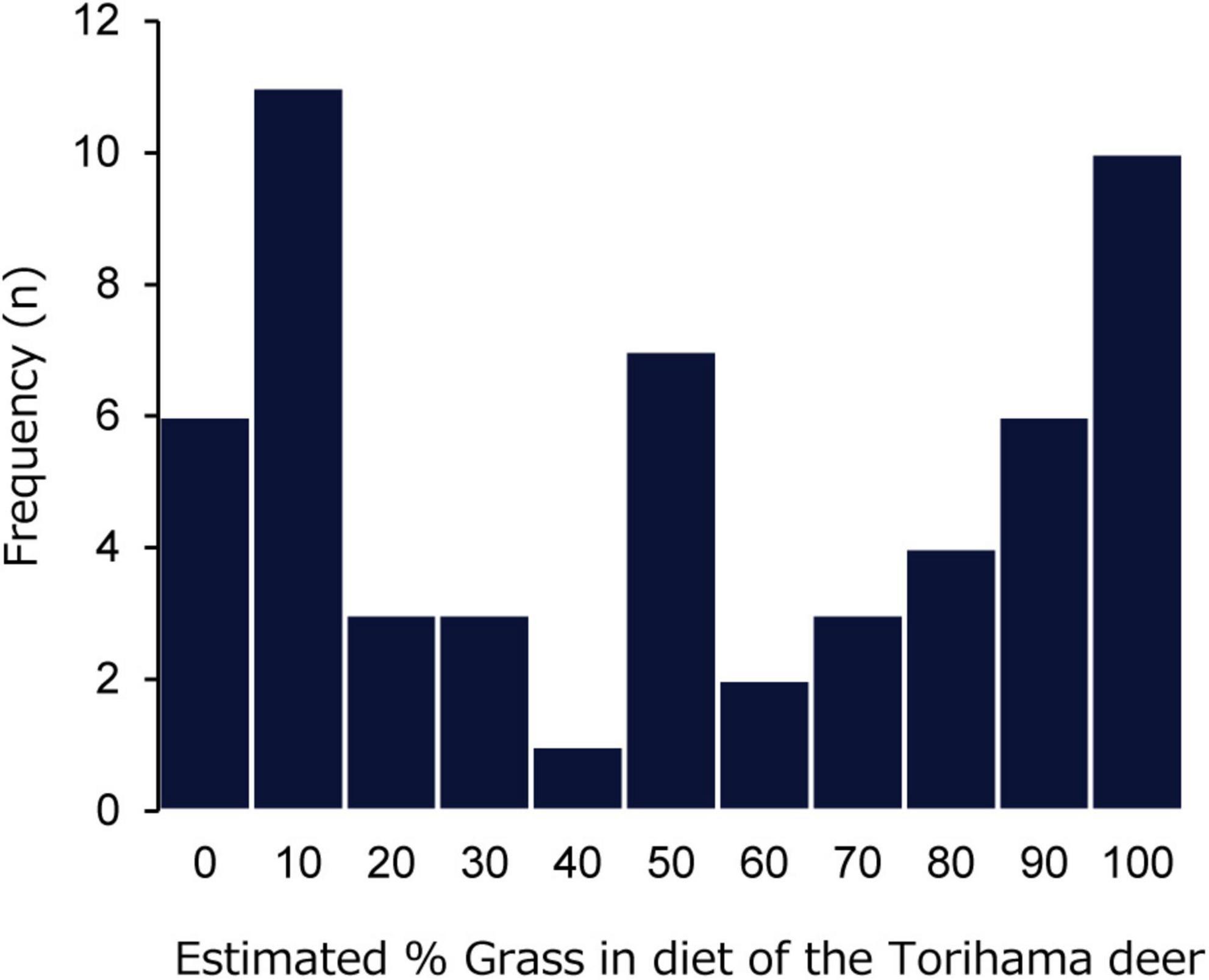
Figure 5. Histogram of estimated % grass in the diet of the Torihama deer, using the estimation equation of Kubo and Fujita (2021).
In this study, we used DMTA to estimate the diets of Torihama deer inhabited approximately 6,000 years ago. Deer mandibles excavated from the site were in satisfactory preservation conditions; thus, observing the antemortem microwear was possible. The results of the comparison with the extant sika deer showed that the Torihama deer overlapped with the populations that had mixed-feeding and grazing-feeding habits and showed a wide range in % grass in the diet.
The results of PCA on 31 parameters of surface roughness showed that the Torihama deer overlapped with the grazing and mixed-feeding deer. However, they did not overlap with the browsing Tsushima deer. The Tsushima deer did not have well-developed microwear. A reason for this observation is that their diets were less abrasive than those of the Torihama deer. For example, the former ate dicot leaves, which polished tooth surfaces rather than abrading them. The absence of overlap between the Tsushima and the Torihama deer indicates that the diet of the Torihama deer was not primarily dicots. The Torihama deer overlapped with the Kinkazan deer, a typical grazer type that developed microwear by feeding on grass, and with the Shimane deer, a mixed feeder type, which demonstrates the possibility that the Torihama deer were feeding on both grass plants and the leaves and fruits of dicotyledonous plants.
Furthermore, some Torihama deer developed abundant scratches on their tooth surfaces, which were finer than those of grazing deer on Kinkazan Island (Figure 3B). These individuals did not overlap with the Kinkazan deer in the scatter plot of PC1 and PC2 (Figure 4). These results may indicate that some Torihama deer foraged on grass species, which differ from those foraged by the Kinkazan deer. The Kinkazan deer feed primarily on short-statured lawn grass (Z. japonica) (Takatsuki, 1980), which may force them to ingest the soil secondarily when feeding on lawn grass at the ground level (Rivals et al., 2014; Rivals and Takatsuki, 2015). Microwear can also be affected by sand and dust attached to foods (Maas, 1994; Gügel et al., 2001; Lucas et al., 2013). However, experimental feeding has clarified that the effects of external abrasives are mediated by the rumination process to some extent (Ackermans et al., 2020; Schulz et al., 2020). The comparison of 15 populations of sika deer demonstrated that the Kinkazan deer had rougher surfaces than other grazing deer (Kubo and Fujita, 2021). Therefore, the Torihama deer might not have fed on short grasses contaminated by soil. Around the Torihama Shell Midden site grew graminoid (Poaceae and Cyperaceae) plants, indicated by the analyses of pollen and macrobotanical remains (Kasahara, 1983, 1984; Kitagawa et al., 2018). The analysis of phytoliths has also identified grasses such as the reed Phragmites and Bambusoideae species (Sasaki, 1983). Therefore, the Torihama deer could also have fed on these grasses when they were available.
The proportion of grass in the diets of the Torihama deer was estimated to be 50.7% on average, with a wide range of 0 to 100% among these individuals (Figure 5). This estimate indicates that Torihama deer are mixed feeders and may reflect the dietary flexibility of sika deer during the Jomon period. It is also assumed that the Torihama deer would change diets according to seasonal availability. The Torihama deer in this study were over 2 years old, representing the diets of weaned individuals. Furthermore, the average age in the Torihama population was estimated to be 6.96 years (Koike and Ohtaishi, 1985); thus, the tendencies of sexually mature individuals can be observed. We could not estimate the mortality seasons for each material used in this study. However, the literature has suggested that the people who lived at the Torihama Shell Midden site may have hunted sika deer year-round, mainly during fall and winter (Nishida, 1980; Ohtaishi, 1980; Sato, 2021b).
The result of stomach content analysis of the Shimane populations, which we used as the reference in this study, showed a seasonal change in diets of deer with a high consumption rate of grasses in summer, seeds and nuts in fall, and tree leaves in winter (Shimane Prefectural Government, 2002). Such seasonal change in diets was also identified for sika deer populations in Hyogo, the closest area to the archeological site; the consumption of seeds and nuts was high in fall; and that of evergreen leaves was high in winter (Yokoyama, 2009). Therefore, although we did not estimate the mortality seasons, the individuals that died in each season were analyzed by the DMTA. Based on the seasonal changes in the diets of the extant sika deer with geographical proximity and the background information that the Torihama deer may have been hunted year-round, the DMT of the Torihama deer may reflect their seasonal dietary changes. As mentioned in the introduction, the flora around the site was mixed vegetation of evergreen trees, broadleaved trees, and coniferous trees, which allowed the Torihama deer to change their forage plants seasonally.
Additionally, the extant sika deer have dietary flexibility and can inhabit a wide range of habitat environments in Japan (Takatsuki, 2009). Takatsuki (2006) suggested that the ecological plasticity of the sika deer is associated with the extinction of large herbivores in the Japanese archipelago around the Pleistocene–Holocene boundary, through the process that vacant ecological niches after those extinctions were occupied by a survived species, sika deer. The argument of Takatsuki (2006) is vague regarding the cause–effect relationship, namely, “sika deer could survive the extinction because they had the ability to adapt to changing environments” or “since there were vacant ecological niches after the extinction, the sika deer became ecologically diverse according to the respective habitats.” To clarify this issue, information on the ecological variability of Pleistocene and Holocene sika deer, as well as other extinct ruminants, is necessary. This study is the first step toward this clarification. Our study indicates that sika deer in the Early Jomon period (approximately 6,000 years ago) had a flexible feeding habit of adapting to the vegetation of their habitats. Further investigation of excavated sika deer from older archeological sites than the site we examined would provide insights into when and how they attained high ecological plasticity.
We investigated the feeding habits of the sika deer in the Early Jomon period by analyzing the DMT of abundant skeletal remains excavated from an archeological site. The Torihama deer were mixed feeders, with a wide range in the estimated percentage of foraged grasses. These results are consistent with the estimate of paleo-vegetation around the Torihama Shell Midden site. Similar to the extant deer populations inhabiting the evergreen broadleaved forest in Honshu, the Torihama deer might have changed their foraging plants seasonally, depending on the growth of plants around their habitat. Their flexible diets are important for understanding how they survived the extinction of large mammals at the end of the Pleistocene and are widely present around the Japanese archipelago today. Most of the Japanese extant samples were collected during the opening of hunting seasons, and in the unhunted population of Kinkazan deer, they were individuals that died of natural causes. Therefore, because the comparative data are mainly from autumn to early spring, further research on seasonal variation in microwear should be conducted by obtaining samples that can be compared among seasons.
For further research, DMTA results will be able to further elucidate the paleoecology of deer by including studies of the mortality season and range of behavior in individuals.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material. The original scan data of all individuals in common 3D surface format (“.sur” format) and 3D surface analytical files including raw scans and procedures of analyses in DMTA standard format (“.mnt”) were uploaded to an online repository (Zenodo): https://doi.org/10.5281/zenodo.7058460.
Ethical review and approval was not required for the animal study because it is focused on archeological materials.
KS: conceptualization, investigation, formal analysis, data curation, visualization, and writing—original draft preparation. TS: conceptualization, resources, supervision, and writing—review and editing. MOK: resources, supervision, and writing—review and editing. All authors contributed to the article and approved the submitted version.
This study was partially funded by JSPS KAKENHI (Grant No. 16H03106) to TS and the Keio University Doctorate Student Grant-in-Aid Program from Ushioda Memorial Fund to KS.
We thank the following institutions and people for their support. The Wakasa History Museum permitted us to use the collections. Mayumi Ajimoto (Wakasa History Museum) and Hideaki Kojima (Wakasa Town Section of History and Culture) provided us with suggestions and support. In addition, we received valuable comments from professors of the Department of Archaeology and Ethnology, Keio University, and the Mammal Society of Japan. We would like to thank Enago (www.enago.jp) for the English language review. Finally, we appreciate the two reviewers for their constructive comments on the earlier version of this manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.957038/full#supplementary-material
Ackermans, N. L., Winkler, D. E., Martin, L. F., Kaiser, T. M., Clauss, M., and Hatt, J.-M. (2020). Dust and grit matter: Abrasives of different size lead to opposing dental microwear textures in experimentally fed sheep (Ovis aries). J. Exp. Biol. 223:jeb220442. doi: 10.1242/jeb.220442
Aiba, K., Miura, S., and Kubo, M. O. (2019). Dental microwear texture analysis in two ruminants, Japanese serow (Capricornis crispus) and sika deer (Cervus nippon), from central Japan. Mamm. Study 44, 183–192. doi: 10.3106/ms2018-0081
Anezaki, T., Nishimoto, T., and Niimi, M. (2005). Faunal analysis of the Torihama shell midden, materials from the area 1 of the 1985 excavation (In Japanese). Torihama Shell Midden Pap. 4-5, 19–30.
Berlioz, E., Azorit, C., Blondel, C., Ruiz, M. S. T., and Merceron, G. (2017). Deer in an arid habitat: Dental microwear textures track feeding adaptability. Hystrix 28, 222–230. doi: 10.4404/hystrix-28.2-12048
DeSantis, L. (2016). Dental microwear textures: Reconstructing diets of fossil mammals. Surf. Topogr. 4:e023002. doi: 10.1088/2051-672X/4/2/023002
Gügel, I. L., Grupe, G., and Kunzelmann, K. H. (2001). Simulation of dental microwear: Characteristic traces by opal phytoliths give clues to ancient human dietary behavior. Am. J. Phys. Anthropol. 114, 124–138. doi: 10.1002/1096-8644(200102)114:2<124::AID-AJPA1012<3.0.CO;2-S
Hongo, H. (1991). Analysis of cut marks on mammal remains from section 85 of the Torihama shell midden (In Japanese). Bull. Natl. Mus. Jpn. Hist. 29, 149–195.
Inami, M. (1983). “Remains of sika deer and wild boar from the Torihama shell midden,” in The summary report of excavation in 1981 and 1982 and research results of Torihama shell midden – research 3 on archaeological site located in lowland area from the early Jomon period (In Japanese), ed. Torihama Shell Midden Study Group (Wakasa Town: Board of Fukui Prefecture and Wakasa Museum of History and Folklore), 65–81.
Kasahara, Y. (1983). “The detection and identification of plant seeds from Torihama shell midden (the 6th excavation),” in The summary report of excavation in 1981 and 1982 and research results of Torihama shell midden – research 3 on archaeological site located in lowland area from the early Jomon period (In Japanese), ed. Torihama Shell Midden Study Group (Wakasa Town: Board of Fukui Prefecture and Wakasa Museum of History and Folklore), 47–64.
Kasahara, Y. (1984). “The detection and identification of plant seeds from the Torihama shell midden (7th excavation),” in The summary report of excavation in 1983 and research results of Torihama shell midden – research 4 on archaeological site located in lowland area from the early Jomon period (In Japanese), ed. Torihama Shell Midden Study Group (Wakasa Town: Board of Fukui Prefecture and Wakasa Museum of History and Folklore), 49–90.
Kawamura, Y. (2009). “Fossil record of sika deer in Japan,” in Sika deer, eds D. R. McCullough, S. Takatsuki, and K. Kaji (Tokyo: Springer), 11–25. doi: 10.1007/978-4-431-09429-6_2
Kitagawa, J., Kojima, H., Yoshida, T., and Yasuda, Y. (2018). Adaptations of the early Jomon people in their settlement relocation to climate change around Lake Mikata, Central Japan. Archaeol. Res. Asia 16, 66–77. doi: 10.1016/j.ara.2018.03.002
Koike, H., and Ohtaishi, N. (1985). Prehistoric hunting pressure estimated by the age composition of excavated sika deer (Cervus nippon) using the annual layer of tooth cement. Archaeol. Sci. 12, 443–456. doi: 10.1016/0305-4403(85)90004-4
Kojima, H. (2015). The consideration of the excavation and study history of Torihama shell midden and its contribution to the zooarchaeological study (In Japanese with English abstract). Zooarchaeology 32, 11–24.
Kubo, M. O., and Fujita, M. (2021). Diets of Pleistocene insular dwarf deer revealed by dental microwear texture analysis. Paleogeogr. Palaeoclimatol. Palaeoecol. 562:e110098. doi: 10.1016/j.palaeo.2020.110098
Kubo, M. O., and Yamada, E. (2014). The inter-relationship between dietary and environmental properties and tooth wear: Comparisons of mesowear, molar wear rate, and hypsodonty index of extant sika deer populations. PLoS One 9:e90745. doi: 10.1371/journal.pone.0090745
Kubo, M. O., Yamada, E., Kubo, T., and Kohno, N. (2017). Dental microwear texture analysis of extant sika deer with considerations on inter-microscope variability and surface preparation protocols. Biosurf. Biotribol. 3, 155–165. doi: 10.1016/j.bsbt.2017.11.006
Kudo, Y., Suzuki, M., Noshiro, S., Ajimoto, M., and Amitani, K. (2016a). Radiocarbon dating of Castanea crenata wood from the incipient and initial Jomon periods excavated from the Torihama shell midden site, Fukui Prefecture, Japan (In Japanese with English abstract). Jpn. Assoc. Hist. Bot. 24, 59–68. doi: 10.34596/hisbot.24.2_59
Kudo, Y., Amitani, K., Yoshikawa, J., Sasaki, Y., Ajimoto, M., and Noshiro, S. (2016b). Radiocarbon dating of plant macrofossils excavated from the Torihama shell midden site, Fukui Prefecture, Japan—a chronological reconsideration of sediment layers and types of pottery from the incipient to early Jomon periods (In Japanese with English abstract). Jpn. Assoc. Hist. Bot. 24, 43–58. doi: 10.34596/hisbot.24.2_43
Lucas, P. W., Omar, R., Al-Fadhalah, K., Almusallam, A. S., Henry, A. G., Michael, S., et al. (2013). Mechanisms and causes of wear in tooth enamel: Implications for hominin diets. J. R. Soc. Interface 10:e20120923. doi: 10.1098/rsif.2012.0923
Maas, M. C. (1994). A scanning electron microscopic study of in vitro abrasion of mammalian tooth enamel under compressive loads. Arch. Oral Biol. 39, 1–11. doi: 10.1016/0003-9969(94)90028-0
Matsubayashi, J., and Tayasu, I. (2019). Collagen turnover and isotopic records in cortical bone. J. Archaeol. Sci. 106, 37–44. doi: 10.1016/j.jas.2019.03.010
Merceron, G., Escarguel, G., Angibault, J.-M., and Verheyden-Tixier, H. (2010). Can dental microwear textures record inter-individual dietary variations? PLoS One 5:e9542. doi: 10.1371/journal.pone.0009542
Miura, S. (2008). “Sika deer,” in A guide to the mammals of Japan, ed. Japan Wildlife Research Center (Tokyo: Tokai university press), 110–111.
Morikawa, M. (1963). Some problems concerning the Torihama shell midden site in Fukui Prefecture (In Japanese). Mater. Cult. 1, 19–34.
Nakagawa, T., Kitagawa, H., Yasuda, Y., Tarasov, P. E., Gotanda, K., and Sawai, Y. (2005). Pollen/event stratigraphy of the varved sediment of Lake Suigetsu, Central Japan from 15,701 to 10,217 SG vyr BP (Suigetsu varve years before present): Description, interpretation, and correlation with other regions. Quat. Sci. Rev. 24, 1691–1701. doi: 10.1016/j.quascirev.2004.06.022
Nakagawa, T., Tarasov, P. E., Nishida, K., Gotanda, K., and Yasuda, Y. (2002). Quantitative pollen-based climate reconstruction in Central Japan: Application to surface and Late Quaternary spectra. Quat. Sci. Rev. 21, 2099–2113. doi: 10.1016/S0277-3791(02)00014-8
Niimi, M. (1997). The preliminary report of the determination of deer hunting seasons by the teeth eruption and wearing (In Japanese with English abstract). Zooarchaeology 9, 21–32.
Nishida, M. (1979). “Animal remains,” in Torihama shell midden – research 1 on archaeological site located in lowland area from the early Jomon period (In Japanese), ed. Torihama Shell Midden Study Group (Wakasa Town: Board of Fukui Prefecture), 164–166.
Nishida, M. (1980). Food resources and subsistence activities in the Jomon period on the organic remains of the Torihama shell midden (In Japanese). Anthropol. Q. 11, 3–41.
Nishimoto, T. (1991). Hunted deer and wild boar in the Jomon period (In Japanese). Kodai 91, 114–132.
Noshiro, S., and Suzuki, M. (1990). A palaeobotanical study of natural woods from the Torihama shell midden and reconstruction of the forest vegetation (In Japanese with English abstract). Bull. Jpn. Sea Res. Inst. 22, 63–152.
Noshiro, S., Suzuki, M., and Amitani, K. (1996). Species of wooden products from the Torihama shell midden (In Japanese). Torihama Shell Midden Pap. 1, 23–79.
Ohtaishi, N. (1980). Estimation of sex, age, and season of death using mandibles of Cervus nippon excavated from an archaeological site (In Japanese). Archaeol. Nat. Sci. 13, 51–74.
Ohtaishi, N. (1986). Preliminary memorandum of classification, distribution and geographic variation on sika deer (In Japanese). Honyurui Kagaku 53, 13–17.
Rivals, F., Schulz, E., and Kaiser, T. M. (2009). A new application of dental wear analyses: Estimation of duration of hominid occupations in archaeological localities. J. Hum. Evol. 56, 329–339. doi: 10.1016/j.jhevol.2008.11.005
Rivals, F., Takatsuki, S., Albert, R. M., and Macià, L. (2014). Bamboo feeding and tooth wear of three sika deer (Cervus nippon) populations from northern Japan. J. Mammal. 95, 1043–1053. doi: 10.1644/14-MAMM-A-097
Rivals, F., and Takatsuki, S. (2015). Within-island local variations in tooth wear of sika deer (Cervus nippon centralis) in northern Japan. Mammal. Biol. 80, 333–339. doi: 10.1016/j.mambio.2015.02.001
Rivals, F., Prignano, L., Semprebon, G. M., and Lozano, S. (2015). A tool for determining duration of mortality events in archaeological assemblanges using extant ungulate microwear. Sci. Rep. 5:e17330. doi: 10.1038/srep17330
Sasaki, S. (1983). “Plant opal analysis of soil from Torihama shell midden,” in The summary report of excavation in 1981 and 1982 and research results of Torihama shell midden – research 3 on archaeological site located in lowland area from the early Jomon period (In Japanese), ed. Torihama Shell Midden Study Group (Wakasa Town: Board of Fukui Prefecture and Wakasa Museum of History and Folklore), 37–41.
Sato, K. (2021a). The resource utilization of sika deer from Torihama shell midden (In Japanese). Annu. Bull. Shiga Prefect. Assoc. Cult. Herit. 34, 1–12.
Sato, K. (2021b). Death-season of sika deer remains from the Torihama shell midden: Nondestructive analyses on mandibular molariform tooth development (In Japanese with English abstract). Zooarchaeology 38, 11–22.
Schulz, E., Calandra, I., and Kaiser, T. M. (2010). Applying tribology to teeth of hoofed mammals. Scanning 32, 162–182. doi: 10.1002/sca.20181
Schulz, E., Winkler, D. E., Clauss, M., Carlsson, J., Ackermans, N. L., Martin, L. F., et al. (2020). Everything matters: Molar microwear texture in goats (Capra aegagrus hircus) fed diets of different abrasiveness. Palaeogeogr. Palaeoclimatol. Palaeoecol. 552:109783. doi: 10.1016/j.palaeo.2020.109783
Shigehara, N., Hongo, H., and Amitani, K. (1991). Mammalian remains from the Torihama shell midden excavation in 1985 (In Japanese). Bull. Natl. Mus. Jpn. Hist. 29, 329–341.
Shimane Prefectural Government (2002). A survey on the sika deer (Cervus nippon) in Misen mountainous region on the Shimane Peninsula, Japan (In Japanese). Shimane: Shimane Prefectural Government.
Suda, K. (1997). Rumen contents and food selectivity of sika deer (Cervus nippon) on Tsushima Islands (In Japanese with English abstract). Wildl. Conserv. Jpn. 2, 125–134.
Takahara, H., and Takeoka, M. (1992). Vegetation history since the last glacial period in the Mikata lowland, the sea of Japan area, western Japan. Ecol. Res. 7, 371–386. doi: 10.1007/BF02347104
Takatsuki, S. (1980). Food habits of sika deer on Kinkazan Island. Sci. Rep. Tohoku Univ. Ser. IV Biol. 38, 7–31.
Takatsuki, S. (2009). “Geographical variations in food habits of sika deer: The Northern Grazer vs. the Southern Browser,” in Sika deer, eds D. R. McCullough, S. Takatsuki, and K. Kaji (Tokyo: Springer), 231–237. doi: 10.1007/978-4-431-09429-6_17
Torihama Shell Midden Study Group (1985). Summary report of excavation in 1984 and research results of Torihama shell midden – research 5 on archaeological site located in lowland area from the early Jomon period (In Japanese). Wakasa Town: Board of Fukui Prefecture and Wakasa Museum of History and Folklore.
Torihama Shell Midden Study Group (1987). Torihama shell midden – summary of research from 1980-1985 (In Japanese). Wakasa Town: Board of Fukui Prefecture and Wakasa Museum of History and Folklore.
Uchiyama, J. (2000). The problem of deer and wild boar in Torihama shell midden: The function of the site revealed by the remains of sika deer and wild boar excavated in 1984 (In Japanese). Torihama Shell Midden Pap. 2, 1–22.
Uchiyama, J. (2005). Hunting seasonality of sika deer (Cervus nippon) and wild boar (Sus scrofa) at Torihama shell midden (Early Jomon): A zooarchaeological analysis of the faunal remains of the excavation in 1984 (In Japanese). Torihama Shell Midden Pap. 4-5, 79–113.
Winkler, D. E., Schulz-Kornas, E., Kaiser, T. M., Codron, D., Leichliter, J., Hummel, J., et al. (2020). The turnover of dental microwear texture: Testing the “last supper” effect in small mammals in a controlled feeding experiment. Palaeogeogr. Palaeoclimatol. Palaeoecol. 557:e1099930. doi: 10.1016/j.palaeo.2020.109930
Yamada, E., Kubo, O. M., Kubo, T., and Kohno, N. (2018). Three-dimensional tooth surface texture analysis on stall-fed and wild boars (Sus scrofa). PLoS One 13:e0204719. doi: 10.1371/journal.pone.0204719
Yamakawa, F. (1992a). Method of making bone piercing tools in the Jomon: Analysis of animal remains excavated from Torihama shell midden (In Japanese). J. Archaeol. Soc. Nippon 78, 61–106.
Yamakawa, F. (1992b). Bone piercing tools from Torihama shell midden (In Japanese). Bull. Wakasa Mus. Hist. Folk. 4, 1–42.
Yasuda, Y. (1979). “Pollen analysis,” in Torihama shell midden – research 1 on archaeological site located in lowland area from the early Jomon period (In Japanese), ed. Torihama Shell Midden Study Group (Wakasa Town: Board of Fukui Prefecture), 176–196.
Yasuda, Y. (1982). Pollen analytical study of the sediment from the Lake Mikata in Fukui Prefecture, Central Japan –especially on the fluctuation of precipitation since the Last Glacial age on the side of Sea of Japan (In Japanese with English abstract). Q. Res. 21, 255–271. doi: 10.4116/jaqua.21.255
Yokoyama, M. (2009). “Biology of sika deer in Hyogo: Characteristics of reproduction, food habits, growth, and condition,” in Sika deer, eds D. R. McCullough, S. Takatsuki, and K. Kaji (Tokyo: Springer), 193–205. doi: 10.1007/978-4-431-09429-6_14
Yoshikawa, M., Yoshikawa, J., Noshiro, S., Kudo, Y., Sasaki, Y., Suzuki, M., et al. (2016). Vegetation history and use of plant resources from the incipient to early Jomon periods at the Torihama shell midden site, Fukui Prefecture, central Japan (In Japanese with English abstract). Jpn. Assoc. Hist. Bot. 24, 69–82. doi: 10.34596/hisbot.24.2_69
Keywords: Archaeology, Jomon period, Torihama Shell Midden site, Sika deer, Microwear, ISO 25178
Citation: Sato K, Sato T and Kubo MO (2022) Reconstructing diets of hunted sika deer from Torihama Shell Midden site (ca. 6,000 years ago) by dental microwear texture analysis. Front. Ecol. Evol. 10:957038. doi: 10.3389/fevo.2022.957038
Received: 30 May 2022; Accepted: 16 September 2022;
Published: 10 October 2022.
Edited by:
Ferran Estebaranz-Sánchez, Milá y Fontanals Institution for Research in Humanities (CSIC), SpainReviewed by:
Florent Rivals, Catalan Institution for Research and Advanced Studies (ICREA), SpainCopyright © 2022 Sato, Sato and Kubo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Koyo Sato, a295by5zYXRvQGtlaW8uanA=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.