 Yuan Wang
Yuan Wang Long Ren†
Long Ren† Dong-po Xu
Dong-po Xu- Key Laboratory of Freshwater Fisheries and Germplasm Resources Utilization, Ministry of Agriculture and Rural Affairs, Freshwater Fisheries Research Center, Chinese Academy of Fishery Sciences, Wuxi, China
This study found significant differences between the standard length distributions of humpback and redfin culter and between Mongolian culter and topmouth culter. A stable isotope analysis (δ13C and δ15N) was used to investigate the interspecies differences between the feeding habits and trophic niches of four carnivorous species of Cultrinae fish. The results showed that the δ13C and δ15N values were significantly correlated with the standard length and species. However, the δ13C values of humpback and redfin culter were not significantly different, suggesting that these two fish species had similar food sources. The δ15N values of Mongolian culter and topmouth culter were the highest, suggesting that they occupied a higher trophic level and that animal prey was more important in their diets. Moreover, variations in δ13C and δ15N indicated considerable niche overlap and interspecific competition among the four species. We also evaluated trophic niches, diversity, redundancy, and evenness utilizing isotopic niche metrics, and we estimated asymmetrical niche overlaps. The analysis revealed that four carnivorous species of Cultrinae displayed similar trophic niche sizes and trophic diversity. The trophic traits of topmouth culter clearly defined them as a trophic generalist in terms of the inter-individual variability in their isotopic niches. A significant finding was that the average niche overlap between them was as high as 64.02%; topmouth culter had the greatest trophic overlap with redfin culter (95.52%) and humpback (90.38%), followed by Mongolian culter onto redfin culter (85.32%), indicating that topmouth culter and Mongolian culter benefit the most from the food supply in the habitat, or they are more competitive in the presence of limited resources.
Introduction
“Niche” refers to the relationships between a species and all other species in a specific ecosystem (Elton, 1927). A niche includes the spatial and temporal positions occupied by a particular species, the status of environmental adaptation, and the utilization of resources by the species (Zhang and Xie, 1997; Li et al., 2006). Multidimensional hypervolume ecology not only emphasizes the habitat meaning of the niche but also includes the resource utilization by organisms and the relationship between them and the environment (Hutchinson, 1957). An occupied niche space implies resource utilization; moreover, understanding the factors that lead to changes in trophic niches is important in assessing food web structures (Yao et al., 2016), resource utilization (Ortega-Cisneros et al., 2017), and trophic interactions (Wallace et al., 2009). Different predators may occupy similar trophic niches, and the trophic overlaps indicate similar diets in freshwater ecosystems (Chen et al., 2011; Wang et al., 2019). Carbon and nitrogen stable isotopes have been proven to effectively quantify niches and niche overlaps of species (Post, 2002; Bearhop et al., 2004). The method allows for the estimation of the overlap among species' isotopic niches to suggest that feeding competition can occur if resources are to be limited and if niche partitioning occurs (Balzani et al., 2020). In conclusion, it is feasible and scientific to use stable isotopes to study isotopic niche overlap.
The topmouth culter (Culter alburnus), Mongolian culter (Culter mongolicus), humpback (Culter dabryi), and redfin culter (Cultrichthys erythropterus) are related carnivorous species in the genus Cultrinae (Cypriniformes and Cyprinidae) (Hu et al., 2011). The topmouth culter and Mongolian culter occupy higher trophic levels than the other two species because of their piscivorous feeding habits and larger sizes, although all four culter fish species are common predators in the aquatic food web. In addition, humpback and redfin culter are widely known as connecting links between forage living and top predators in some lake and reservoir ecosystems (Ye, 2006; Li et al., 2013). They are important commercial freshwater fishes and are widely distributed in China, inhabiting the middle and upper levels of large water bodies rich in aquatic plants (Chen, 1998; Zhang X. G. et al., 2008; Zhang X. L. et al., 2008). Mongolian culter and topmouth culter have the highest economic values because of their large size, rapid growth, and the desirable flavor of their meat. Redfin culter and humpback are smaller, more aggressive fish that are also economically valuable (Feng et al., 2007). Previous research has shown that culters, important top predators in lakes, primarily feed on lake anchovy (Coilia nasus taihuensis), followed by other small fish and shrimp. Hence, because they have obvious inhibitory effects on small fish and shrimp, they maintain the ecosystem's stability (Zhou et al., 2011).
In recent years, overfishing and the destruction of the spawning grounds of large fish such as Mongolian culter and topmouth culter have caused changes in the populations and distributions of humpback and redfin culter. Wild populations of four species are declining so quickly that have resulted in the domination of their communities by lake anchovy and other small fish (Liu et al., 2005, 2007; Xiong et al., 2022), and the overlap of their isotopic niches has become an extremely important issue (Wang et al., 2007). Ontogenetic niche shifts could change trophic relationships among species and would play diverse roles in food webs (Woodward and Hildrew, 2002). It has been established that culters experience significant increases in body size during their development, during which they utilize different resources, which results in different feeding habits and ontogenetic niche shifts (Zhou et al., 2011). Domestic and overseas scholars have conducted much research on the domestication and cultivation of culters (Lin et al., 2013; Wang F. et al., 2015), with the goals of protecting germplasm resources, increasing body sizes and economic benefits, and strengthening the ecological regulation of lakes. Restraining the biomass of small fish with culters could decrease the predation pressure on zooplankton. This would mitigate the competition for filter-feeding fish, such as silver carp and bighead carp, reinforcing the effects of control on algae growth. It would be extremely beneficial for populations of the culters to rebound (Liu et al., 2007; Hu et al., 2011). The improved ecological balance of waters and the prevention of fish resources being dominated by smaller individuals could be achieved by artificial propagation and release and by strengthening the management of lake eutrophication.
In the 1950s and 1960s, the water quality was good because of a high degree of abundant macrophytes (Chen et al., 2009). However, subsequently, due to increasing eutrophication and the resulting lack of macrophytes, Lihu Lake became the most highly polluted in Taihu Lake (Nian et al., 2006). The monitoring of Lihu Lake revealed that the water quality changed due to severe eutrophication as reported in 2001 (Gu and Lu, 2004; Bai et al., 2010). Since 2003, several environmental protection and ecological restoration projects have been implemented by the central government and local authorities. Technological measures, such as water purification fisheries, were conducted in Lihu Lake by releasing suspension-feeding animals between 2007 and 2010 (Chen et al., 2006; Huang et al., 2012; Xu et al., 2012). As a result of the program, the fish community structure in Lihu Lake has been adjusted to a certain extent, and the trophic level of the ecosystem has increased from 3.63 to 3.95 (Di, 2014). Furthermore, large-scale stock enhancement has been implemented in recent years by the Taihu Lake Fishery Management Committee Office. However, the phenomenon that lake anchovy has been the dominant species in Taihu Lake, while the four carnivorous Cultrinae fish resources are still extremely limited (Liu et al., 2005; Zhang et al., 2021). The main causes of this phenomenon were human overfishing, the destruction of spawning grounds (Mao et al., 2011), and considerable niche overlap. Now, following the comprehensive Yangtze River Ten-Year Fishing Ban in October 2020, a comprehensive fishing ban was also implemented in Taihu Lake. Regardless of whether the structure of the fish community in Taihu Lake and Lihu Lake, particularly the biomass of large culters, has improved since the ban's implementation, there is an urgent need for future research on the resources and trophic niche of the culters. This study focused on the effects of individual size on δ13C and δ15N values. Besides, the proportion of niche overlap was analyzed, showing considerable niche overlap among the four species examined in this study. The study aimed to provide new knowledge and sufficient data to support fishery management and biological resource conservation.
Materials and methods
Fish sampling sites
Lihu Lake is an extensive semi-enclosed bay toward to the northern Taihu Lake, located at 120°13′-120°27′E and 31°28′-31°35′N. It covers an area of 8.6 km2 and has a perennial water level of 3.07 m and an average water depth of 1.60 m. In this study, we examine the overlap and competition among four fish species from the perspective of feeding ecology. During the fishing period in October 2013, samples from 46 topmouth culter, 35 Mongolian culter, 129 humpback, and 49 redfin culter were collected in Lihu Lake at three sampling sites, labeled S1, S2, and S3, based on previous research by our team (Figure 1).
Figure 1. Sampling sites in Lihu Lake.
Collection and pre-treatment of fish
The samples of redfin culter, topmouth culter, Mongolian culter, and humpback were collected in October 2013 by multi-mesh gillnets (standard length: 125 m; width: 1.5 m; mesh size: 1.2, 2, 4, 6, 8, 10, and 14 cm, average distribution in the entire net). Then the fish were preserved in ice, the standard length and body weight of each sample were measured at approximately 0.01 mm and 0.1 g, respectively, and the white dorsal muscle tissues were dissected in the laboratory (Pinnegar and Polunin, 1999). Samples of different ranges of standard lengths of the four species were selected for statistical comparison. The following groups were compared: 40–59, 60–79, 80–99, 100–119, 120–139, 140–159, 160–179, 180–199, 200–219, 220–239, and >280 mm. After 48 h of drying at 60°C, the muscle samples were ground into powder and stored in centrifuge tubes before measurement.
The biometrics including the standard length-weight relationship of 16 topmouth culter, 13 Mongolian culter, 101 humpback, and 30 redfin culter specimens were statistically analyzed. A one-way analysis of variance (ANOVA) was performed on 16 topmouth culter, 10 Mongolian culter, 14 humpback, and 8 redfin culter specimens. To determine the isotopic niche metrics and overlaps, 13 topmouth culter, 11 Mongolian culter, 14 humpback, and 11 redfin culter specimens were examined.
Stable isotope analysis
Carbon and nitrogen stable isotope ratios were determined using Vario EL cube-Iso Prime 100 (Elementar, GER) by isotope-ratio mass spectrometry and elemental analysis. The results were expressed in conventional delta (δ) notation as parts per thousand (‰) defined as follows:
where X is 13C or 15N, R is 13C/12C or 15N/14N, Rstandard is the isotopic ratio in certified reference material, and Rsample is the isotopic ratio measured (Peterson and Fry, 1987). The isotopic ratio of C was expressed in relation to its abundance in Vienna Pee Dee Belemnite (VPDB); the isotopic ratio of N was expressed in relation to its abundance in standard atmospheric nitrogen, provided as a certified reference. We inserted one standard sample for every 10 sample measurements, and we randomly selected one or two samples for reanalysis. The standard errors of repeated measurements of δ13C and δ15N values were less than 0.2‰ and 0.3‰, respectively.
Statistical analysis
The data were analyzed using IBM SPSS Statistics. The relationships between the values of δ13C and δ15N of muscle and standard length were analyzed by partial correlation (Wang X. et al., 2015). Kruskal-Wallis test analysis was used to analyze whether there were significant differences in standard length distribution among the four fish species, which gives us a way to evaluate ordinal data in more depth and draw strict statistical interpretations from the results, including comparison of means (Gao and Chen, 2011). To identify whether there were any statistically significant differences between the standard length and δ13C and δ15N values in the four species, a multivariate analysis of variance (MANOVA) was performed, with δ13C and δ15N values as variables and standard length as covariates. Post-hoc multiple comparison tests using the least significant difference (LSD; α = 0.05) were also performed to identify the significance of standard length and δ13C and δ15N values between the four species.
To assess the overlap between the isotopic niches, complementary approaches were used. The direct overlap of the maximum likelihood fitted standard ellipses (SEAc) was calculated using the R package SIBER (Jackson et al., 2011). To measure the total extent of spacing and trophic redundancy within a δ13C and δ15N biplot for fish communities or species, all data were calculated in the R package SIBER. These metrics were adapted from the community-wide metrics proposed by Layman (Layman et al., 2007, 2012; Jackson et al., 2012): carbon range (CR), nitrogen range (NR), mean distance to centroid (CD), total area (TA), standard ellipse corrected area (SEAc), mean nearest neighbor distance (MNND), and its standard deviation (SDNND). CR is the difference between individuals with the most enriched and the most depleted δ13C and is a measure of basal resource diversity. NR is the difference between species with the most enriched and most depleted δ15N values and is a measure of trophic length within a population. CD and TA are indices of species niche width. SEAc indicates the species' core isotopic niches, calculated from the variance and covariance of bivariate isotopic data by evaluating the trophic niche area of a species. MNND is the mean of the Euclidean distances to each specie's nearest neighbor in a biplot space and thus is a measure of the overall packing density of the species. SDNND is a measure of the evenness of species packing in a biplot space. Low SDNND values indicated an even distribution of trophic niches.
To measure the trophic niche size and to test whether trophic niche overlap was not equivalently weighted among species, we used the nicheROVER R package developed by Swanson et al. (2015). In this method, a Monte Carlo resampling routine (n = 10,000 draws) is applied to randomly draw from the sampled population and calculate a mean and 95% (or user-defined) credible interval of niche overlap probabilities. Swanson et al. (2015) defined the niche overlap of species A onto species B as the fraction of the intersection area between niche A and niche B over the total niche area of B and vice versa. We used a 95% probability for niche size and overlap.
Results
Relationship between standard length and weight
In this study, all fish samples were measured as shown in the table. Standard length and body–weight growth relationships were fitted to power functions (Figure 2) represented by W = aLb: humpback W = 3 × 10−5 L2.8555 (R2 = 0.9859, P < 0.01), redfin culter W = 1 × 10−5 L3.047 (R2 = 0.9666, P < 0.01), Mongolian culter W = 3 × 10−5 L2.8627 (R2 = 0.9911, P < 0.01), and topmouth culter W = 2 × 10−5 L2.8664 (R2 = 0.9941, P < 0.01). The mean standard length of humpback (122.07 ± 36.62 mm) was similar to that of the redfin culter (136.81 ± 16.32 mm); however, both were much lower than that of the Mongolian culter (185.87 ± 72.46 mm) and topmouth culter (165.20 ± 61.22 mm).
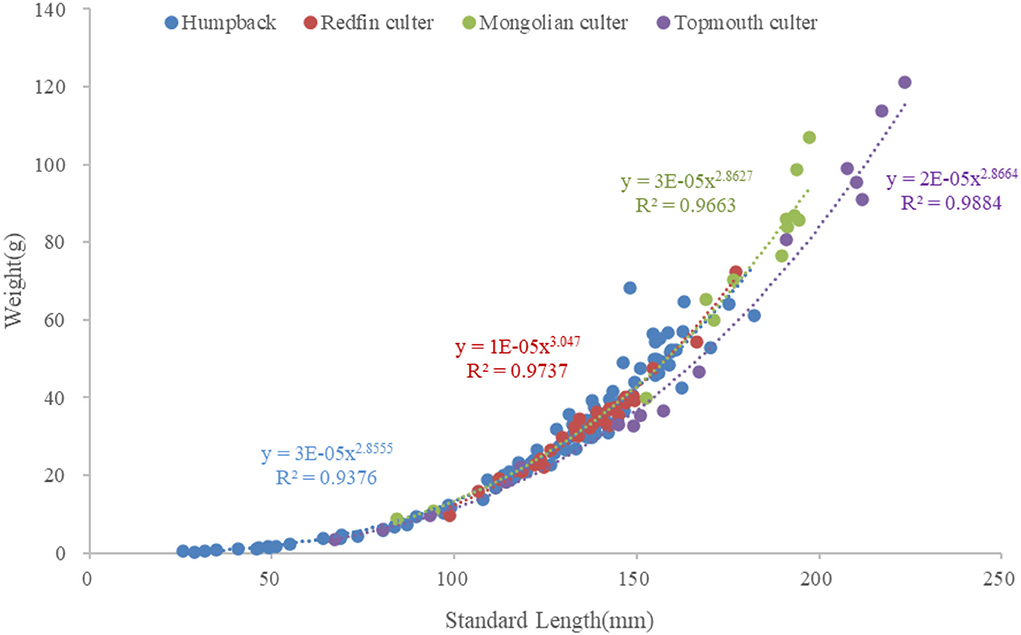
Figure 2. Relationships between the weights and standard lengths of humpback, redfin culter, Mongolian culter, and topmouth culter.
The results of the Kruskal-Wallis test showed that there was a significant difference in the overall distribution of standard length among the four species (χ2 = 22.99, P < 0.05). There were significant differences between humpback to Mongolian culter and topmouth culter (χ2 = 17.15, P < 0.05; χ2 = 6.61, P < 0.05), and there was also a significant difference between redfin culter to Mongolian culter (χ2 = 11.64, P < 0.05), but no significant differences between humpback and topmouth culter (χ2 = 2.39, P > 0.05; χ2 = 3.24, P > 0.05), and there was also no significant difference between Mongolian culter and topmouth culter (χ2 = 0.38, P > 0.05) (Table 1).
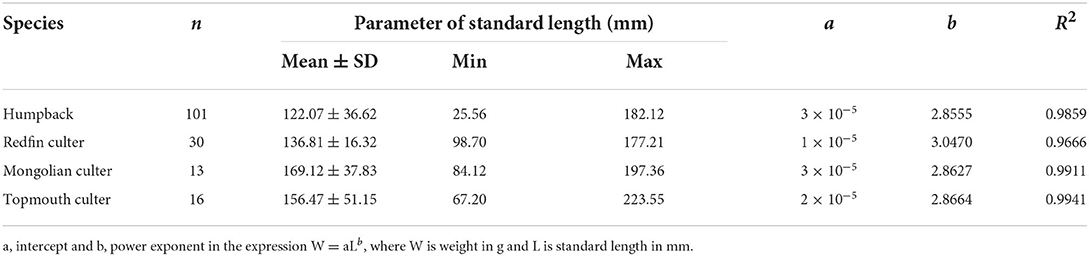
Table 1. Standard length–weight relationships for four fish species in Lihu Lake.
δ13C and δ15N values and interspecies relationships
The results of the partial correlation analysis demonstrated a significant negative correlation in the δ13C value of humpback with standard length (P < 0.01) and a significant positive correlation of the δ15N value of humpback with standard length (P < 0.01). The δ13C and δ15N values of redfin culter, Mongolian culter, and topmouth culter all showed significant positive correlations with standard length (P < 0.05).
The mean δ13C values for humpback, redfin culter, Mongolian culter, and topmouth culter were (−25.76 ± 0.65‰), (−25.93 ± 0.61‰), (−25.94 ± 0.69‰), and (−26.59 ± 0.90‰), respectively. The value of humpback was the highest, and that of the topmouth culter was the lowest within the same standard length group. The results of multivariate analysis indicated no significant variations in the distribution of δ13C values (P > 0.05).
The mean δ15N values for humpback, redfin culter, Mongolian culter, and topmouth culter were (12.08 ± 0.74‰), (12.80 ± 0.68‰), (13.60 ± 1.56‰), and (12.63 ± 1.21‰), respectively. The value of Mongolian culter was the highest and that of humpback was the lowest. The results of the multivariate analysis indicated significant variations in the distribution of δ15N values (P < 0.05). The results showed that the δ15N value of the redfin culter did not significantly differ from those of the others (P > 0.05) while that of the Mongolian culter was significantly different from that of the topmouth culter (P < 0.05) and humpback (P < 0.01).
Isotopic niche metrics and overlaps
The results showed that the isotopic niches of the four species were confirmed by the considerable overlap of SEAc areas (2.95).
In addition, the results of the SIBER package analysis indicated that the topmouth culter had the highest CR, TA, SEAc, and CD values, followed by the Mongolian culter, but the NR, MNND, and SDNND values were higher in the Mongolian culter. Furthermore, the NR of the Mongolian culter (6.08‰) and topmouth culter (5.03‰) was greater than the magnitude of the average isotope fraction (3.4‰) per trophic level. In addition, the humpback displayed the lowest CR, TA, MNND, and SDNND values, and the redfin culter also possessed the lowest NR, SEAc, and CD values. CR, NR, TA, CD, MNND, SDNND, and SEAc showed that the topmouth culter and Mongolian culter were much higher than the humpback and redfin culter. Therefore, the topmouth culter, Mongolian culter, humpback, and redfin culter shared some parts of similar niche space sizes (Figure 3, Table 2), and the niche space of the former two species was wider than the others.
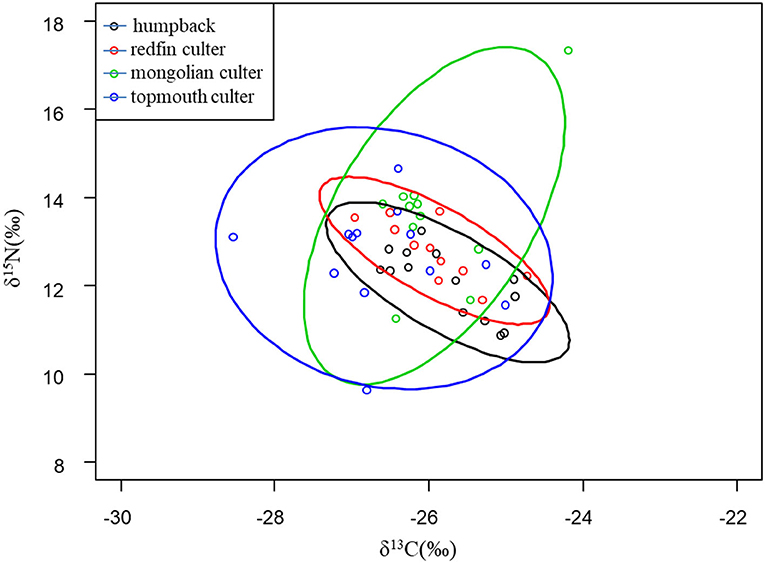
Figure 3. Isotopic biplot of four fish species in Lihu Lake.
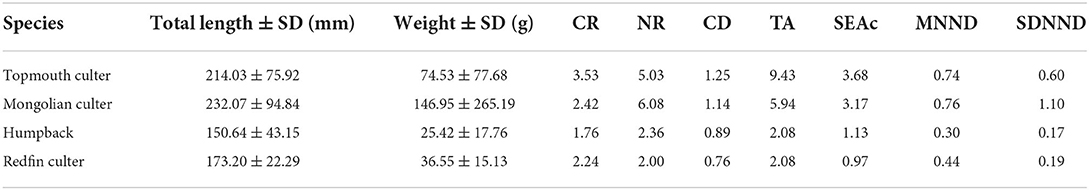
Table 2. Layman's metrics and corrected standard ellipse area of topmouth culter, Mongolian culter, humpback, and redfin culter in Lihu Lake.
The results also showed various degrees of trophic overlap among the four species, with an overall average of 64.02% (nicheROVER). High trophic overlaps (>80%) were shown for topmouth culter onto redfin culter (95.52%) and topmouth culter onto humpback (90.38%) and Mongolian culter onto redfin culter (85.22%). The lowest trophic overlap was found between humpback and Mongolian culter (30.95%), while higher overlaps were found between redfin culter and Mongolian culter (33.70%), between redfin culter and topmouth culter (35.72%), and between humpback and topmouth culter (42.10%). The average trophic overlap between topmouth culter and other species was as high as 85.93%, followed by Mongolian culter (74.75%) onto the other three fish, which was greater than the trophic overlap between humpback (50.36%) and redfin culter (45.04%) (Figure 4).

Figure 4. Posterior distribution of the probabilistic niche overlap metric (%) and niche region size (α) = 0.95, indicating the probability of one species (row) overlapping onto the niche of another species (column) among topmouth culter, Mongolian culter, humpback, and redfin culter in Lihu Lake.
Discussion
Body dimension relationships among the four species
In humpback and topmouth culter, the power exponent was b < 3, indicating that the two populations experienced positive allometric growth (i.e., the growth rate of standard length was higher than weight). The redfin culter and Mongolian culter showed negative allometric growth (Wang et al., 2020).
The growth rates of the four fish species have been shown to vary among water bodies, depending on geographical location, physical and chemical factors, food resources, and fishing intensity (Feng et al., 2007). Other factors that affect the standard length–weight relationship include habitat, seasons, gonad maturation state, and sex (Lv et al., 2018). In this study, we found significant differences in these relationships among the four fish species living in the same habitat.
Relationships between species and δ13C and δ15N values
Many species have been found to display similar patterns of increasing δ13C and δ15N values with size, presumably due to the ability of larger fish to capture larger and more diverse prey at a higher trophic level (Graham et al., 2007; Tsai et al., 2014). As shown in Figure 5, the values of δ13C and δ15N in the four species increased with standard length (P < 0.05), implying that their food source changed as they grew (Li et al., 2011; Wang F. et al., 2015). These observations are consistent with the results of a previous dietary analysis, which revealed that their food sources changed as they developed (Chen et al., 1994).
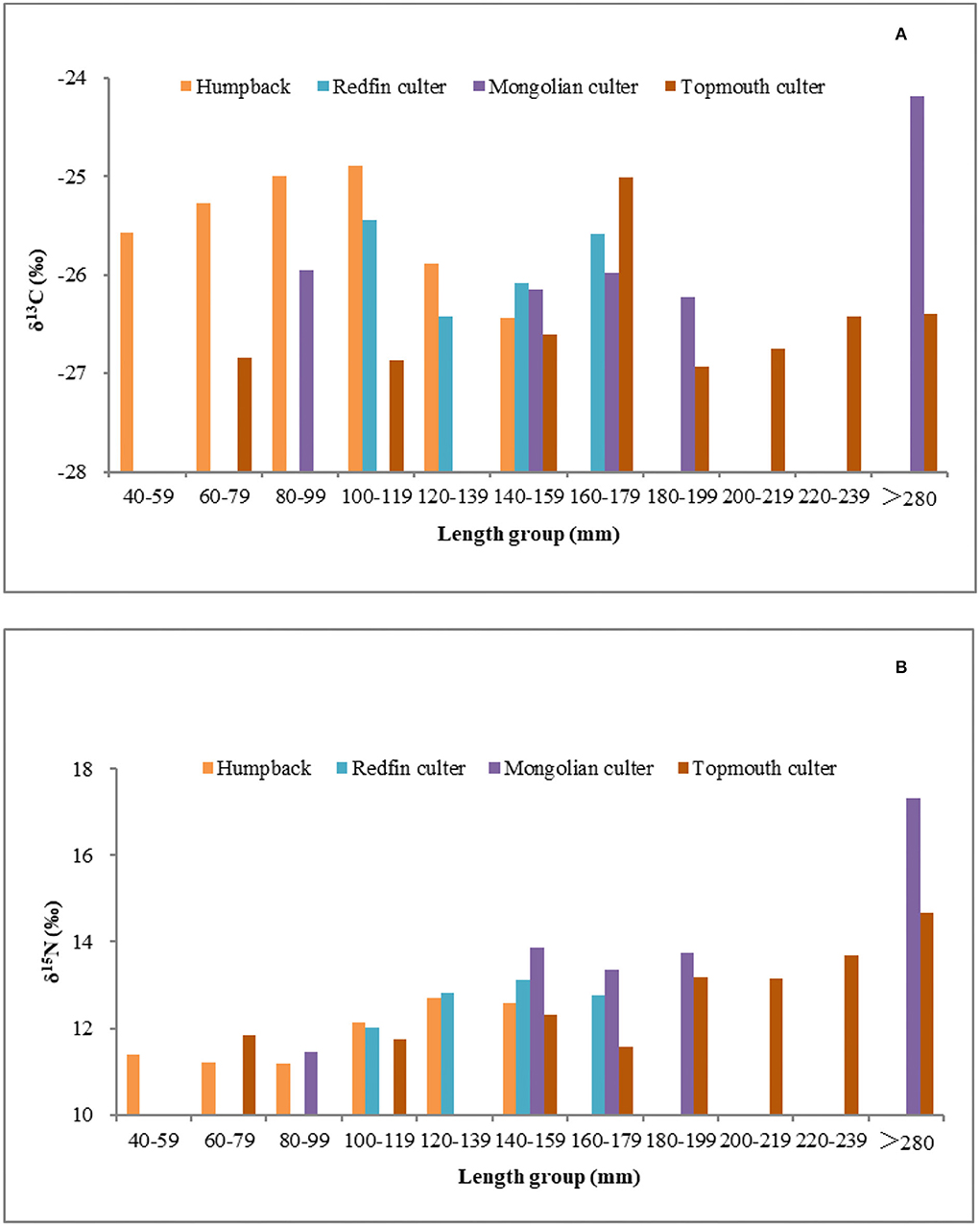
Figure 5. Relationships between δ13C (A) and δ15N (B) values and standard lengths of humpback, redfin culter, Mongolian culter, and topmouth culter.
In this study, significantly larger bodies and higher values of the δ13C were found in the topmouth culter than in the other three species. These results indicated that the carbon sources of the topmouth culter were more diverse and similar to previous studies; although they were in the same habitat, the humpback, redfin culter, and Mongolian culter had a smaller size, perhaps because of various individual sizes or different feeding habits, such as the topmouth culter could make better use of zooplankton, small prey fish, and shrimp, while the redfin culter can make better use of phytoplankton (Li, 2014).
Within the same standard length range, the δ15N values of the Mongolian culter and topmouth culter were generally higher than those of the humpback and redfin culter (Figure 5), thus reflecting the former species' higher trophic level. The Mongolian culter and topmouth culter are more carnivorous than the other two carnivorous Cultrinae species, and animal prey accounts for a large proportion of their food resources (Wang et al., 2009).
Trophic niche of the four carnivorous Cultrinae fish species and overlaps
Individuals of the four fish species could be defined as more or less “generalist” by comparing either the inter-individual variability of their trophic niches or their intra-individual variability or both. In this study, we evaluated the inter-individual components of the realized trophic niches of different species by analyzing their isotopic niches and metrics. Several generalist species have low within-individual and high inter-individual diet variations, which indicates that the species includes specialists that utilize different resources. The overlap and breadth of generalist species' niches, such as those of topmouth culter and Mongolian culter, are much greater than those of the others. Moreover, the generalist species typically have a wider spectrum of trophic resources and feed at multiple trophic levels. They also have lower trophic redundancy, higher trophic evenness, higher trophic diversity, and larger trophic niches (Shea and Chesson, 2002; Ribeiro et al., 2007; Hayden et al., 2014; Cicala et al., 2020), indicating that these species take full advantage of the food supply in the habitat and could be more competitive in the presence of limited resources. In addition, previous studies have shown that adults of the Mongolian culter are carnivorous, mainly feeding on sharpbelly, while topmouth culters mainly feed on various fishes, such as fishes in Rhodeus, Culter, and so on (Xing and Yang, 1992; Ni and Wu, 2006; Zhang, 2008). Our study also showed that although both Mongolian culter and topmouth culter could be bigger and regarded as “generalist,” the CR of Mongolian culter was lower than that of topmouth culter, indicating that the food sources of the former were narrower, and its viability would be more sensitive to environmental turbulence, which might be necessary to pay attention to by managers. Moreover, the researchers proposed that artificial release or formulating reasonable fishing ban periods, or prohibiting harmful fishing gear fishing laws were effective measures to proliferate large and medium-sized commercial fish, adjust community composition, and stabilize interspecies relationships in Taihu Lake (Yin and Miu, 1991). So as for the redfin culter and topmouth culter, the competitive ability was weaker than the other two carnivorous Cultrinae fish species. We can take appropriate proliferation measures, such as setting a fishing ban period, fishing ban, or taking necessary artificial proliferation measures to maintain the stability of the Lihu Lake ecosystem.
Small topmouth culter and Mongolian culter are omnivorous, feeding on small zooplankton and small fish (e.g., Rhodeinae and Pseudorasbora parva) (Yang et al., 2002; Liu et al., 2007; Liu, 2008). However, large individuals are carnivorous, preferring high-energy lake anchovy and other small fish, such as redfin culter. Adult culters prefer open upper–middle water layers, whereas juveniles prefer slow-moving shoal water (Tan et al., 2009). Thus, their different food sources may reflect their different habitats. Food intake increases and the capacity for predation strengthens according to their body growth (Yu et al., 2012). Because of the dual effects of inter- and intraspecific competition, the culters tend to feed on high-energy food organisms that meet their growth requirements. While optimal foraging theories predict trophic niche broadening as a consequence of reduced food availability, where consumers relying on insufficient preferred food items are forced to add less profitable resources to their diet, hence widening their trophic niche. Other biological factors also influence the niche of a species, including food density and diversity, population size, and predator-prey relationships. In addition, seasonal changes in aquatic environmental factors also influence their distribution and food resources, thus affecting fish biomass in the trophic niche (Bearhop et al., 2004; Sha et al., 2015).
Conclusion
This study presents the trophic niche characteristics of four carnivorous Cultrinae fish species in Lihu Lake. The results support the preview hypothesis that the topmouth culter has a wide spectrum of trophic resources and feeds at multiple trophic levels compared with the Mongolian culter, humpback, and redfin culter. Based on comparisons of four similar species, we know that the competitive ability of redfin culter and humpback is weaker than the topmouth culter and Mongolian culter, and the Mongolian culter is more sensitive to environmental turbulence than the topmouth culter. All these findings show the influence of trophic interactions of culter and Cultrichthys in lake ecosystems and provide information that assists in resource protection with respect to nutritional ecology. Future research should be conducted to determine the mechanism of the differences in the nutrition acquisition of the four species examined in this study.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was reviewed and approved by Regulations for the Administration of Affairs Concerning Experimental Animals and the State Council of the People's Republic of China.
Author contributions
YW and LR led the manuscript writing and analyzed the data. D-pX and D-aF contributed to the manuscript writing. YW designed the data analysis. LR determined and validated the species' determination. D-pX and YW designed the experiment and were the scientific supervisors the experiment. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by the National Key Research and Development Program of China (NO. 2020YFD0900500), Central Public-Interest Scientific Institution Basal Research Fund, CAFS (NO. 2020XT13 and NO. 2020TD61).
Acknowledgments
We thank Xiao-ling Wang for collecting the samples. Furthermore, we thank the Third Institute of Oceanography, State Oceanic Administration for measurement of stable isotopes.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Bai, X., Chen, K. N., Huang, W., and Chen, X. M. (2010). Current status and variation tendency of water quality in Lihu Lake, Jiangsu Province. Water Resour. Protect. 26, 6–10.
Balzani, P., Gozlan, R. E., and Haubrock, P. J. (2020). Overlapping niches between two co-occurring invasive fish: the topmouth gudgeon Pseudorasbora parva and the common bleak Alburnus alburnus. J. Fish Biol. 97, 1385–1392. doi: 10.1111/jfb.14499
Bearhop, S., Adams, C. E., Waldron, S., Fuller, R. A., and Macleod, H. (2004). Determining trophic niche width: a novel approach using stable isotope analysis. J. Anim. Ecol. 73, 1007–1012. doi: 10.1111/j.0021-8790.2004.00861.x
Chen, G., Wu, Z., Gu, B., Liu, D., Li, X., and Wang, Y. (2011). Isotopic niche overlap of two planktivorous fish in southern China. Limnology 12, 151–155. doi: 10.1007/s10201-010-0332-2
Chen, J. Z., Meng, S. L., You, Y., Hu, G. D., Qu, J. H., Wu, W., et al. (2009). Characteristic of phytoplankton community in Lihu Lake, Taihu Lake. Ecol. Environ. Sci. 18, 1358–1367. doi: 10.3969/j.issn.1674-5906.2009.04.028
Chen, K. N., Bao, X. M., Shi, L. X., Chen, W. M., Lan, C. J., Xu, H., et al. (2006). Ecological restoration engineering in Lake Wuli, Lake Taihu: a large enclosure experiment. J. Lake Sci. 18, 139–149.
Chen, Y. R., Yang, J. X., Zhou, W., Cui, G. H., Kuang, F. R., Wang, Y., et al. (1994). The biology of Cultrichthys erythropterus in Dianchi Lake with evaluation on its influence on Neosalanx taihuensis. Zool. Res. 15, 88–95.
Cicala, D., Polgar, G., Mor, J. R., Piscia, R., Brignone, S., Zaupa, S., et al. (2020). Trophic niches, trophic positions, and niche overlaps between non-native and native fish species in a subalpine lake. Water 12:3475. doi: 10.3390/w12123475
Di, Y. (2014). Technology Research of Waterpurification Fisheries in Lihu Lake (D). Nanjing Agricultural University.
Feng, G. P., Ye, S. W., Li, Z. J., and Xie, C. X. (2007). Age and growth of Cluter alburnus and Cultrichthys erythropterus in Niushan Lake. Reser. Fish. 37, 39–42. doi: 10.3969/j.issn.1000-6907.2007.01.008
Gao, Y. W., and Chen, D. Y. (2011). Study on the distribution of groundwater contamination based on Kruskal-Wallis Test method: taking Changchun area an example. Progr. Geophys. 26, 2249–2254. doi: 10.3969/j.issn.1004-2903.2011.06.045
Graham, B. S., Grubbs, D., Holland, K., and Popp, B. N. (2007). A rapid ontogenetic shift in the diet of juvenile yellowfin tuna from Hawaii. Mar. Biol. 150, 647–658. doi: 10.1007/s00227-006-0360-y
Gu, G., and Lu, G. F. (2004). On the integrated control of water environment of Lihu Lake, Taihu Lake. J. Lake Sci. 16, 56–60. doi: 10.18307/2004.0108
Hayden, B., Massa-gallucci, A., Harrod, C., O'grady, M., Caffrey, J., and Kelly-quinn, M. (2014). Trophic flexibility by roach Rutilus rutilus in novel habitats facilitates rapid growth and invasion success. J. Fish Biol. 84, 1099–1116. doi: 10.1111/jfb.12351
Hu, H. Y., Di, Y., Zhao, Y. F., Song, Q. H., and Bing, X. W. (2011). Comparative study on the morphological characteristics of four species of Culter and Culterichthys in Lihu Lake. J. Yunn. Agric. Univ. 26, 488–494.
Huang, X. F., Bing, X. W., and Chen, J. Z. (2012). Energy flow and trophic structure of the Lihu Lake ecosystem based on an ecopath model. J. Fish. Sci. China 19, 471–481. doi: 10.3724/SP.J.1118.2012.00471
Hutchinson, G. E. (1957). Concluding remarks. Cold Spring Harb. Symp. Quant. Biol. 22, 415–427. doi: 10.1101/SQB.1957.022.01.039
Jackson, A. L., Inger, R., Parnell, A. C., and Bearhop, S. (2011). Comparing isotopic niche widths among and within communities: SIBER - stable isotope Bayesian ellipses in R. J. Anim. Ecol. 80, 595–602. doi: 10.1111/j.1365-2656.2011.01806.x
Jackson, M. C., Donohue, I., Jackson, A. L., Britton, J. R., Harper, D. M., and Grey, J. (2012). Population-level metrics of trophic structure based on stable isotopes and their application to invasion ecology. PLoS ONE 7:e31757. doi: 10.1371/journal.pone.0031757
Layman, C. A., Araujo, M. S., Boucek, R., Hammerschlag-Peyer, C. M., Harrison, E., Jud, Z. R., et al. (2012). Applying stable isotopes to examine food-web structure: an overview of analytical tools. Biol. Rev. 87, 545–562. doi: 10.1111/j.1469-185X.2011.00208.x
Layman, C. A., Arrington, D. A., Montana, C. G., and Post, D. M. (2007). Can stable isotope ratios provide for community-wide measures of trophic structure? Ecology 88, 42–48. doi: 10.1890/0012-9658(2007)88(42:CSIRPF)2.0.CO;2
Li, B., Wang, Z. J., Jin, L., Ye, F. F., Chen, D. Q., and Zhang, Y. G. (2011). Study on diet shift of Culter mongolicus with stable isotope technology. J. Fish. China 35, 1419–1425. doi: 10.3724/SP.J.1231
Li, B., Wang, Z. J., Yang, J. P., Yue, X. J., Qi, Z. M., and Zhang, Y. G. (2013). The dynamic and seasonal variation of the fish food webs in the mainstream of Three Gorges Reservoir. J. Fish. China 37, 1015–1022. doi: 10.3724/SP.J.1231.2013.38428
Li, D. Z., Shi, Q., Zang, R. G., Wang, X. P., Sheng, L. J., Zhu, Z. L., et al. (2006). Models for niche breadth and niche overlap of species or populations. Sci. Silv. Sin. 95–103.
Li, H. C. (2014). Diet and Morphological Differences of Feeding Organs Among Five Culters in Changhu Lake. Yangtze University.
Lin, M. L., Li, Z. J., Xia, Y. G., and Wang, Q. D. (2013). Preliminary study on feeding domestication and fingerling cultivation of Culter mongolicus. Acta Hydrobiol. Sin. 37, 974–977. doi: 10.7541/2013.127
Liu, E. S. (2008). A study on diet composition of dominant fishes in Taihu Lake. J. Fish. China 32, 395–401. doi: 10.3724/SP.J.00001
Liu, E. S., Liu, Z. W., Chen, W. M., and Chen, K. N. (2005). Changes in the yield and composition of the fish catches and their relation to the environmental factors in Taihu Lake. J. Lake Sci. 17, 251–255. doi: 10.3321/j.issn:1003-5427.2005.03.010
Liu, E. S., Wu, L. K., Cao, P., Ding, S. Q., and Yang, Q. C. (2007). Quantity variation and ecological effect analysis of Culter in Taihu Lake. Reserv. Fish. 27, 70–73. doi: 10.3969/j.issn.1003-1278.2007.03.031
Lv, D. W., Zhou, Y. F., Ge, Y., Wang, C. H., and You, Y. (2018). Age structure and growth characteristics of Culter alburnus from Dianshan Lake. Acta Hydrobiol. Sin. 42, 762–769. doi: 10.7541/2018.093
Mao, X. W., Gu, X. H., Zeng, Q. F., Zhou, L. H., Wang, X. L., Wu, L. K., et al. (2011). Community structure and diversity of fish in Taihu Lake. Chin. J. Ecol. 30, 2836–2842. doi: 10.13292/j.1000-4890.2011.0435
Nian, Y. G., Nie, Z. D., and Chen, J. (2006). The ecological restoration theory and practice for Lihu lake. China Water Resour. 17, 37–39. doi: 10.3969/j.issn.1000-1123.2006.17.013
Ortega-Cisneros, K., Lecea, A. M., Smit, A. J., and Schoeman, D. S. (2017). Resource utilization and trophic niche width in sandy beach macrobenthos from an oligotrophic coast, Estuar. Coast. Shelf Sci. 184, 115–125. doi: 10.1016/j.ecss.2016.11.011
Peterson, B. J., and Fry, B. (1987). Stable isotopes in ecosystem studies. Annu. Rev. Ecol. Syst. 18, 293–320. doi: 10.1146/annurev.es.18.110187.001453
Pinnegar, J. K., and Polunin, N. V. C. (1999). Differential fractionation of δ13c and δ15n among fish tissues: implications for the study of trophic interactions. Funct. Ecol. 13, 225–231. doi: 10.1046/j.1365-2435.1999.00301.x
Post, D. M. (2002). Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology 83, 703–718. doi: 10.1890/0012-9658(2002)083(0703:USITET)2.0.CO;2
Ribeiro, F., Orjuela, R. L., Magalhaes, M. F., and Collares-Pereira, M. J. (2007). Variability in feeding ecology of a South American cichlid: a reason for successful invasion in mediterranean-type rivers? Ecol. Freshw. Fish 16, 559–569. doi: 10.1111/j.1600-0633.2007.00252.x
Sha, Y. C., Zhang, P. Y., Zhang, H., Su, G. H., and Xu, J. (2015). Impacts of habitat environment on trophic niches of a local population: a case study of yellow catfish. Acta Ecol. Sin. 35, 1321–1328. doi: 10.5846/stxb201305121024
Shea, K., and Chesson, P. (2002). Community ecology theory as a framework for biological invasions. Trends Ecol. Evol. 17, 170–176. doi: 10.1016/S0169-5347(02)02495-3
Swanson, H. K., Lysy, M., Power, M., Stasko, A. D., Johnson, J. D., and Reist, J. D. (2015). A new probabilistic method for quantifying n-dimensional ecological niches and niche overlap. Ecology 96, 318–324. doi: 10.1890/14-0235.1
Tan, L., Xiong, B. X., Wang, J. S., Cheng, Z. X., and Wang, D. W. (2009). Ecological function of Culter in natural water and countermeasures of resources proliferation. Hubei Agric. Sci. 48, 233–236. doi: 10.3969/j.issn.0439-8114.2009.01.067
Tsai, C. N., Chiang, W. C., Sun, C. L., Shao, K. T., Chen, S. Y., and Yeh, S. Z. (2014). Trophic size-structure of sailfish Istiophorus platypterus in eastern Taiwan estimated by stable isotope analysis. J. Fish Biol. 84, 354–371. doi: 10.1111/jfb.12290
Wallace, B. P., Avens, L., Braun-McNeill, J., and McClellan, C. M. (2009). The diet composition of immature loggerheads: insights on trophic niche, growth rates, and fisheries interactions. J. Exp. Mar. Biol. Ecol. 373, 50–57. doi: 10.1016/j.jembe.2009.03.006
Wang, F., Zhang, Y. K., and Chen, D. (2015). Artificial breeding and seedling cultivation of Culter alburnus in Xingkai Lake. Sci. Fish Farm. 2, 6–7. doi: 10.14184/j.cnki.issn1004-843x.2015.02.003
Wang, P. Z., Song, R., Zhang, Y. Z., Qian, J., Gong, K., Ye, S. W., et al. (2020). Resource composition, length-weight relationship and condition factor of fishes. Biotic Resour. 42, 181–187. doi: 10.14188/j.ajsh.2020.02.004
Wang, W., Chen, L. Q., Yang, P., Hou, L., He, C. B., Gu, Z. M., et al. (2007). Assessing genetic diversity of populations of topmouth culter (Culter alburnus) in China using AFLP markers. Biochem. Syst. Ecol. 35, 662–669. doi: 10.1016/j.bse.2007.04.008
Wang, X. L., Xu, D., Liu, K., Duan, J., and Shi, W. (2015). The change of δ13C and δ15N values in Cultrichthys erythropterus from three water areas in Taihu Lake. J. Lake Sci. 27, 925–931. doi: 10.18307/2015.0520
Wang, Y. Y., Yu, X. B., Zhang, L., and Xu, J. (2009). Food web structure of Poyang Lake during the dry season by stable carbon and nitrogen isotopes analysis. Acta Ecol. Sin. 29, 1181–1188. doi: 10.3321/j.issn:1000-0933.2009.03.014
Wang, Y. Y., Zhang, H., Chen, Y. W., Zhang, L., and Lei, G. C. (2019). Trophic niche width and overlap of three benthic living fish species in Poyang Lake: a stable isotope approach. Wetlands 39(Suppl 1), 17–23. doi: 10.1007/s13157-018-0995-8
Woodward, G., and Hildrew, A. G. (2002). Body-size determinants of niche overlap and intraguild predation within a complex food web. J. Anim. Ecol. 71, 1063–1074. doi: 10.1046/j.1365-2656.2002.00669.x
Xing, D. Y., and Yang, G. R. (1992). Feeding habits of six ferocious fish species in Danjiangkou Reservoir. Reserv. Fish. 3, 10–12.
Xiong, M. H., Ren, L., and Xu, D. P. (2022). Community structure and diversity of fish in Taihu Lake from 2016 to 2020 and its relationship with water environment. J. Shanghai Ocean Univ. 1–13. Available online at: http://kns.cnki.net/kcms/detail/31.2024.S.20220311.1902.006.html
Xu, W. D., Mao, X. W., Wu, D. H., and Lin, Q. B. (2012). Analysis and evaluation of water ecological restoration effect of Lihu Lake, Taihu Lake. Water Resour. Dev. Res. 12, 60–63. doi: 10.13928/j.cnki.wrdr.2012.08.005
Yang, R. B., Xie, C. X., and Yang, X. F. (2002). Study of food composition from 6 kinds of ferocious fish in Liangzi Lake. Reserv. Fish. 22, 1–3.
Yao, X., Huang, G., Xie, P., and Xu, J. (2016). Trophic niche differences between coexisting omnivores silver carp and bighead carp in a pelagic food web. Ecol. Res. 31, 831–839. doi: 10.1007/s11284-016-1393-4
Ye, J. L. (2006). Studies on Community Composition and Feeding Ecologyof Fish in the Littoral Zone of Mayliang Bay, Taihu Lake. Huazhong Agricultural University.
Yin, M. C., and Miu, X. Z. (1991). Ecological characteristic of common fishes and propagation measures in Taihu Lake. J. Lake Sci. 3, 25–34. doi: 10.18307/1991.0104
Yu, J. L., He, H., Li, K. Y., Chen, F. Z., and Liu, Z. W. (2012). Seasonal variations in the diets of Coilia ectenes taihuensis Yen et Lin in Gonghu Bay of Taihu Lake. J. Lake Sci. 24, 765–770. doi: 10.18307/2012.0518
Zhang, G. M., and Xie, S. C. (1997). Evolution and prospects of niche concept. Chin. J. Ecol. 16, 47–52. Available online at: http://ir.xtbg.org.cn/handle/353005/2879
Zhang, X., Shen, W., and Zhou, G. D. (2021). Status of the fish community and canonical correspondence analysis of environmental factors in Lake 'Taihu from 2018 to 2020. J. Ecol. Rural Environ. 37, 674–680. doi: 10.19741/j.issn.1673-4831.2020.0447
Zhang, X. G. (2008). Comparative Studies on Morphology and Growth of Four Culters in Lake Poyang. Huazhong Agricultural University.
Zhang, X. G., Cao, L., Liu, J. Y., Bai, L. H., and Yu, C. J. (2008). Growth pattern of Redfin Culter, Cultrichthys erythropterus Basilewsky 1855, in Poyang Lake. J. Nanchang Univ. 32, 70–74. doi: 10.3969/j.issn.1006-0464.2008.01.017
Zhang, X. L., Yang, P. L., Xu, L. M., and Wang, X. Y. (2008). The research to characteristics and growth of the age for Culter alburnus and Culter mongolicus. Inland Aquat. Prod. 33, 7–10.
Keywords: trophic niche, Cultrinae fish, interspecies relationship, Lihu Lake, stable isotope
Citation: Wang Y, Ren L, Xu D-p and Fang D-a (2022) Exploring the trophic niche characteristics of four carnivorous Cultrinae fish species in Lihu Lake, Taihu Basin, China. Front. Ecol. Evol. 10:954231. doi: 10.3389/fevo.2022.954231
Received: 27 May 2022; Accepted: 17 August 2022;
Published: 08 September 2022.
Edited by:
Tian Zhao, Key Laboratory of Mountain Ecological Rehabilitation and Biological Resource Utilization, Chengdu Institute of Biology (CAS), ChinaReviewed by:
Yanyi Zeng, Pearl River Fisheries Research Institute (CAFS), ChinaZachary Feiner, Wisconsin Department of Natural Resources, United States
Copyright © 2022 Wang, Ren, Xu and Fang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dong-po Xu, eHVkcEBmZnJjLmNu
†These authors have contributed equally to this work