 Chong Yu
Chong Yu Hao Zhao2
Hao Zhao2 Chuenyan Ng
Chuenyan Ng- 1School of Sociology and Anthropology, Sun Yat-sen University, Guangzhou, China
- 2School of Archaeology and Museology, Peking University, Beijing, China
- 3Shaanxi Provincial Institute of Cultural Relics and Archaeology, Xi’an, China
The introduction and the later dispersal of domestic cattle in China had played an enormous part in food production, handicraft manufacture, and ritual activities. However, we lack the details of pastoral economies, herding strategies, and utilization patterns of cattle in ancient China. This study, for the first time, uses biometrical assessments to investigate post-domesticated cattle husbandry and how it changed over time. Biometric data from eight sites dating from the Late Neolithic to the Late Bronze Age were evaluated using kernel analysis and mixture analysis. Results indicated that the general body size of cattle first decrease and then increase. By combining the estimated distribution of male and female individuals, we can see that the sex ratios affect size variation during Bronze Age times. Although no breed improvement has been detected so far, the herding and slaughtering strategies over time are different.
Introduction
First domesticated in the Near East during the late 9th millennium BC and soon spread into the neighboring regions (Peters et al., 1999; Helmer et al., 2005; Arbuckle and Makarewicz, 2009; Hongo et al., 2009), cattle (Bos taurus), the domesticated form of the extinct aurochs (Bos primigenius) has been an important animal to many ancient societies since prehistoric times as it provides not only meat and milk for subsistence but also hide, blood, dung, traction, and even bones that contribute to the organization of human beliefs, cultural attitudes, and social complexity (Sherratt, 1981; Evershed et al., 2008; Campbell et al., 2011).
Changes in the animal body size were proven to be a powerful indicator in explaining both hunting and husbandry activities in ancient times, such as selective hunting (Zhang et al., 2022), early domestication (Grigson, 1989; Zeder and Hesse, 2000), herding strategies (Manning et al., 2015), and husbandry improvements (Davis and Beckett, 1999; Albarella et al., 2008; Davis, 2008; Wright, 2021). Previous biometrical analysis suggested that aurochs were widely hunted in China during the Early Neolithic and the Middle Neolithic times (−2300 BC), based on their relatively large body size compared with the standard animal and later samples. The small-sized domesticated cattle make its first appearance in the middle Yellow River Valley not earlier than 2300 BC (Yu, 2020). Bimolecular investigations confirmed that the domestication of cattle is a single event (Bollongino et al., 2006, 2008, 2012; Edwards et al., 2007), and domestic cattle in China were descendants of their Near East ancestors (Cai et al., 2014). However, the time and by what routes domestic cattle were introduced into China are still unclear (Yu, 2020). In addition, no previous study has investigated the details of cattle utilization after its introduction. This study, for the first time, reveals the patterns of change and diversity in cattle body size that are linked with husbandry practices and slaughtering strategies across northern China from the Late Neolithic to the Late Bronze Age, which helps to tease apart questions about livestock raising development and socio-economic organization in the past.
Archeological background and site description
In this study, biometrical data of domesticated cattle from eight archeological sites spanning the Late Neolithic through the Late Bronze Age periods (2300–771 BC) were involved (Figure 1 and Table 1). The Late Neolithic (2300–1900 BC) settlement sites of Shimao, Zhoujiazhuang, Muzhuzhuliang, and Shengedaliang belonged to the late Longshan Culture, which dates back to around 2300–1900 BC. Recent discoveries revealed a similar pattern of site layout, including large settlements, numerous house foundations, pits, as well as burials (Drennan and Dai, 2010; Shaanxi Provincial Institute of Archaeology, 2015, 2016; Hu et al., 2016). zooarchaeology, archaeobotanical, and isotopic analyses from these sites revealed that intensive agriculture was taking place focusing on millet cultivation and caprids-pig-based herding economy (Zhao and He, 2006; Chen et al., 2012, 2015; Brunson et al., 2015; Hu et al., 2016; Yang et al., 2021). As previous research suggested, mammal species discovered from archeological sites in the Early Neolithic and the Middle Neolithic (−2300 BC) comprise mainly wild boar and domesticated pig [Sus scrofa, 15–85% by number of identified specimens (NISP)], as well as various kinds of deer (up to 80% by NISP), including roe deer (Capreolus pygargus), red deer (Cervus elaphus), Père David’s deer (Elaphurus davidianus), sika deer (Cervus nippon), Chinese water deer (Hydropotes inermis), and muntjak (Muntiacus spp.) (Yu, 2019). Bos remains from these periods were all aurochs and guar (Chen et al., 2020; Yu, 2020). Domestic cattle were introduced in China around 2300 BC; therefore, data from the Late Neolithic represent the circumstance of the earliest stage of cattle husbandry in Chinese civilization (Yu, 2020).
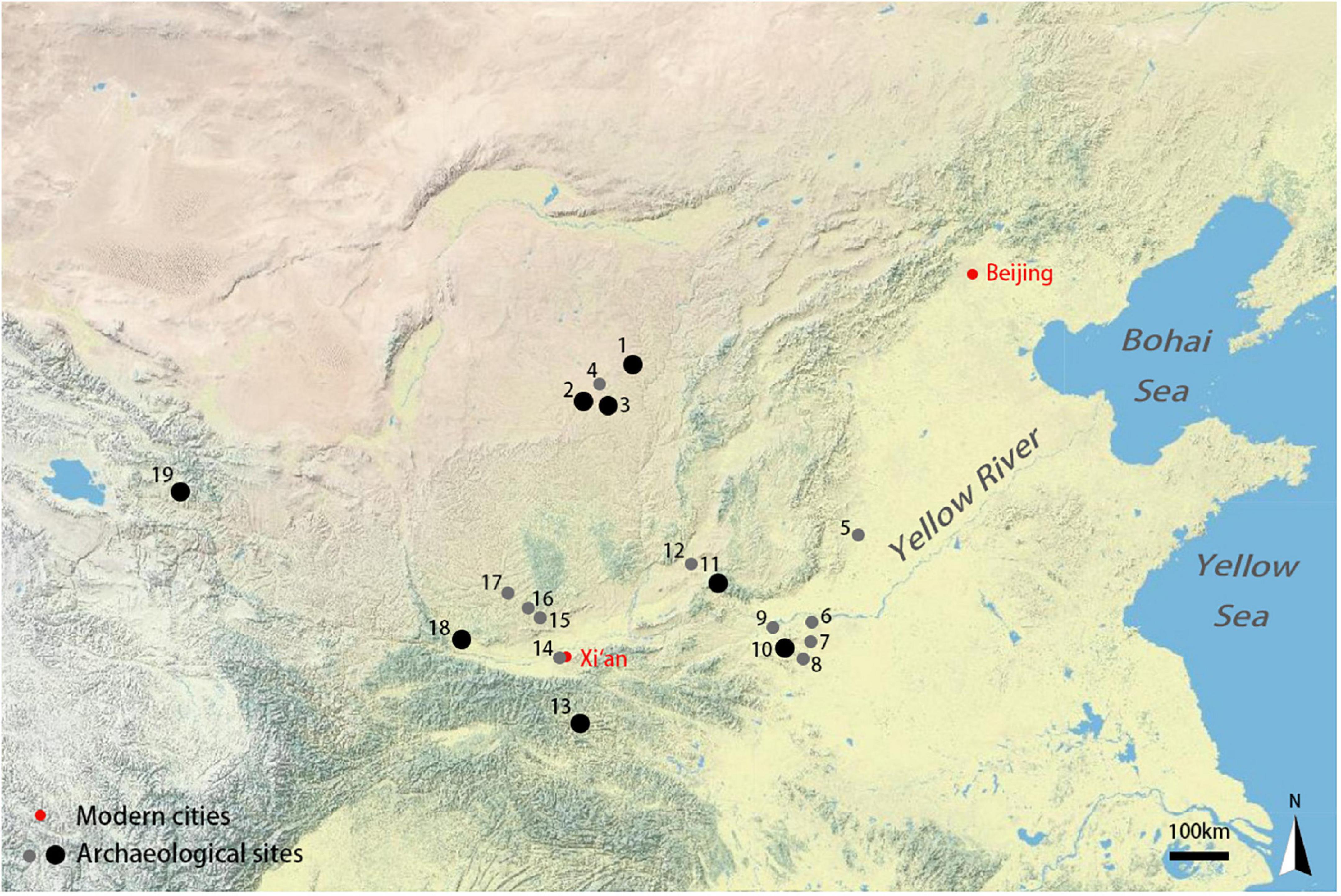
Figure 1. Map of archeological sites included in this study. 1. Shimao; 2. Muzhuzhuliang; 3. Shengedaliang; 4, Huoshiliang; 5, Xiaomintun locus of Anyang Yinxu, Zhangdeng; 6, Guandimiao; 7, Xinzhai; 8, Wadian; 9, Erlitou; 10, Nanwa, Wangchenggang; 11, Zhoujiazhuang; 12, Taosi; 13, Donglongshan; 14, Fengxi; 15, Zaoshugounao; 16, Zaolinhetan; 17, Nianzipo; 18, Yuntang, Qijia, Zhougongmiao locus of Zhouyuan; 19, Changning.
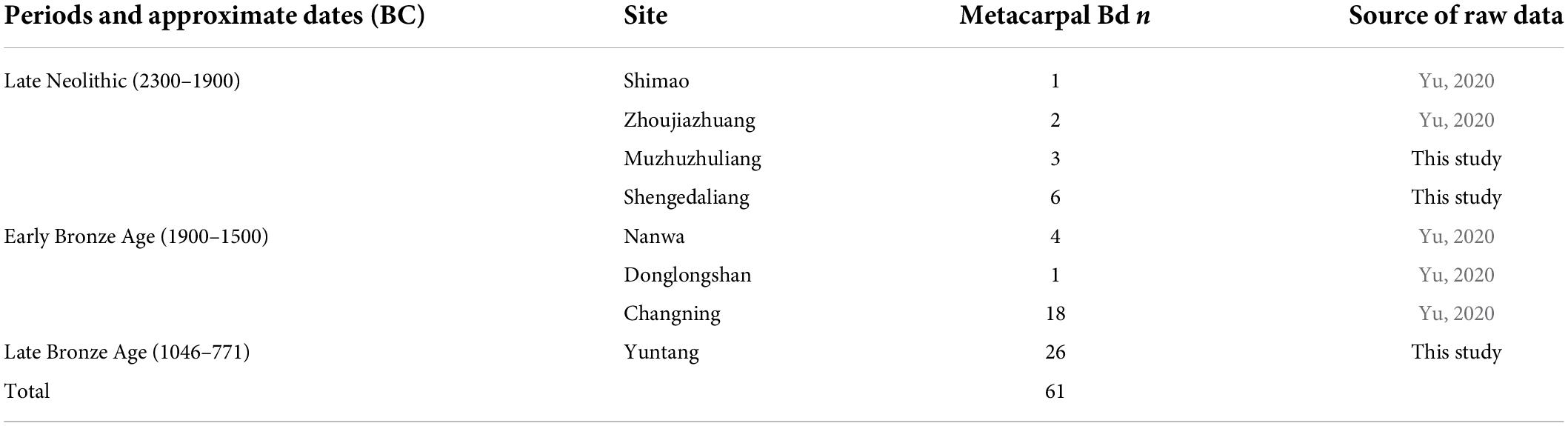
Table 1. Sample sources for the Late Neolithic to the Late Bronze Age cattle.
The Early Bronze Age (1900–1500 BC) sites comprise Erlitou Culture settlements of Nanwa and Donglongshan (Shaanxi Provincial Institute of Archaeology and Shangluo Museum, 2011; Department of Archaeology Zhengzhou University, 2014), and Qijia Culture settlement Changning (Li, 2012). Previous zooarchaeology studies revealed that the faunal assemblages of Nanwa and Donglongshan were dominated by domestic pigs, and Changning was dominant by caprids (Hu, 2011; Li, 2012; Yu, 2014). Analysis of archaeobotanical remains from the Nanwa site suggested a dry-land agriculture tradition heavily relied on millet (Wu et al., 2014).
The Late Bronze Age (1046–771 BC) period was represented by Yuntang bone-working locus belonging to the early and mid-Western Zhou Dynasty. Yuntang locus is located within the core area of the Zhouyuan site, the dynastic capital of the Zhou Dynasty. The total size of Yuntang is approximately six hectares. In the first excavation in 1976, more than 10 tons of bone materials were recovered in an area of 350 m2. In 2013, the Zhouyuan Archeological Team proposed to include the study of the Yuntang bone-working industry as a subproject in the long-term research project “Settlement and Society at Zhouyuan” and decided to conduct a new season of survey and excavation at the Yuntang locus. In 2014, Zhouyuan Archeological Team excavated a trench (14ZYG1) of 20 m2 (2 × 10 m, 3–3.5 m in depth) at the northeast part of the site, approximately 30 m north of the excavation location in 1976. In total, 1.9 tons of bones were recovered. Cattle bones dominate the entire faunal assemblage followed by pigs and caprids (Zhao, 2017).
Materials and methods
Osteometric data used in this study include metrics collected for this study and from published sources (Table 1). This is by far the largest dataset of archeological cattle bone metrics from China. A scatter graph was commonly used to compare the dimensions of archeological specimens (Yu, 2020), but this method requires a relatively large sample size. Besides, two or more dimensions from each specimen will be needed for data plotting, which rarely happens in archeological assemblages. One of the most significant advances to increase the sample size for biometrical methods is the application of the log size index technique. However, the fundamental imperfection of this approach is that the method is based on the assumption that all measurements vary proportionally with the size of the standard animal, ignoring the fact that allometry will have an impact on body proportions, which do vary by age, sex, environment, region, breed, and so on. Meanwhile, by lumping all measurable data together, element-specific information will be weakened (Meadow, 1999).
Therefore, in this study, we used the greatest breadth of the distal end (Bd) of metacarpal to chase the variation of body size and sex ratio among periods, as it has the highest frequency and best preservation condition (Yu, 2020) and relatively high value for size estimation (von den Driesch, 1976) and sex differentiation (Higham, 1969; Svensson et al., 2008; Prummel and Niekus, 2011; Davis et al., 2012; Telldahl et al., 2012) of all measurable body parts. Biometric data were collected on all completely fused metacarpals discovered in the studied sites, following von den Driesch (1976) with a Mitutoyo caliper (precision: tenths of millimeter). As cattle are animals with determinate growth, once fusion takes place further growth in most body dimensions is no longer possible (Reitz and Wing, 2008). The distal metacarpal of cattle fuses around 2–2.5 years of age (Silver, 1969), which said that the age of all individuals involved in this research is subadults and adults (O’Connor, 1991; Jones and Sadler, 2012), older than 2 years of age.
Raw osteometric measurements were first displayed in boxplots showing the variations of the median, inter-quartile range, and maximum and minimum values among the three studied periods. These data were then plotted in histograms for the demonstration of distribution together with kernel analysis, which helps to smooth the distribution of the frequencies of the quantitative variable. Finally, mixture analysis was performed in order to evaluate the number of groups present in a sample and assign each variable to a corresponding group. This method was proved to be a powerful tool for sexing populations. In the meantime, the proportions of each group, mean, and standard deviation were also provided (Monchot et al., 2005). The statistical differences between biometrical data were examined among periods and assigned groups via the Mann–Whitney U test. The level of statistical significance was set at p = 0.05. Statistical tests and analysis for biometric data were performed using PAST 4.07 (Hammer et al., 2001).
Results
The change in general body size
According to Figure 2, the general body size represented by the greatest distal breadth of metacarpals changed through time. The size of Early Bronze Age cattle decreased when compared with Late Neolithic. In contrast, Late Bronze Age cattle showed a marked increase in the median metacarpal Bd value from 60.91 to 66.76 mm. However, the results of the Mann–Whitney U test were not significant (Table 2).
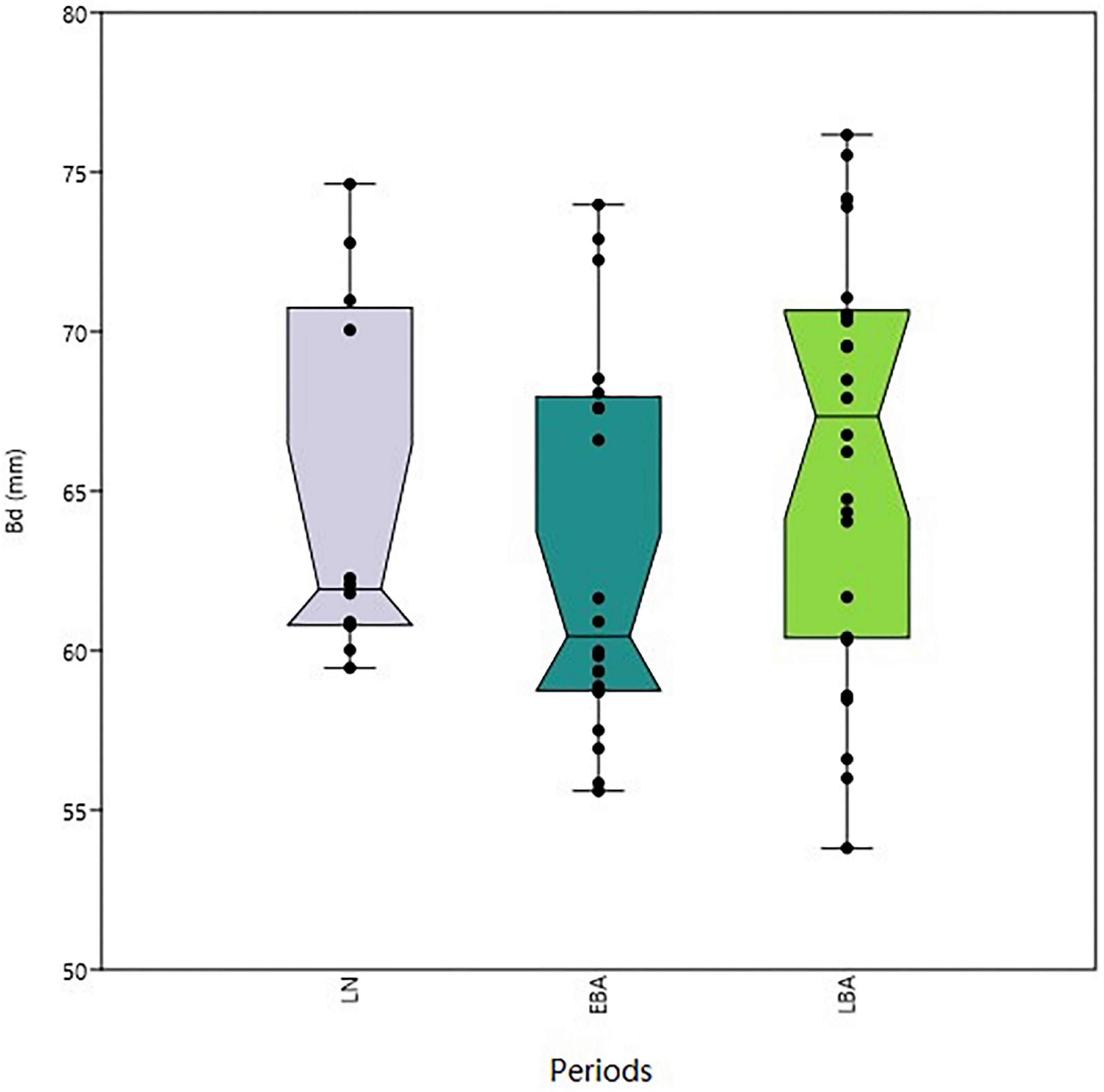
Figure 2. Boxplots showing median, inter-quartile range, and maximum and minimum values for cattle metacarpal Bd.

Table 2. Mann–Whitney U test pairwise comparisons of cattle metacarpal Bd from the Late Neolithic to the Late Bronze Age periods.
Sex ratio diversity among periods
What stands out in Figure 3 is a clear bimodal distribution of metacarpus Bd measurements of cattle bones from three periods, supported by histograms and kernel density curves. Thus, the mixture model was successfully fitted for each period. As males are generally larger than females among most mammals (Higham, 1969); therefore, a lower mean value corresponds to female individuals, and a higher mean value corresponds to male individuals. Previous studies indicated that the greatest length (GL) and the smallest breadth of the diaphysis (SD) of metacarpus were commonly used as indicators for tracing castrates (Albarella, 1997; Minniti and Abatino, 2022), as their limb bones tend to be slender and longer due to the delay of epiphyseal fusion (Davis, 1996, 2000). However, complete metacarpus is extremely rare in our current data set, which enabled the determination of steers. Thus, we used male/castrate cattle to indicate individuals of larger body sizes. The estimation of probable proportions of female and male/castrate cattle, mean and standard deviation, and numbers of assignment of female and male/castrate cattle in each period are reported in Table 3.
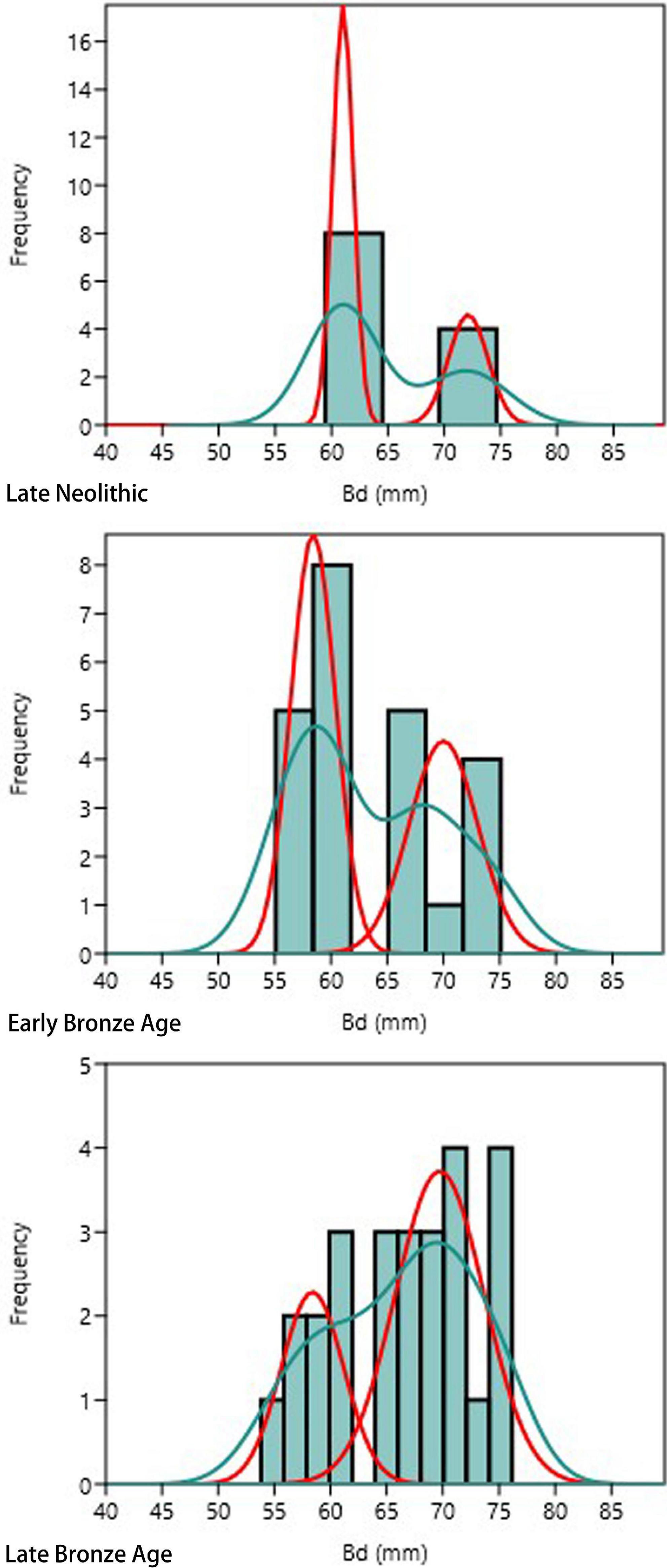
Figure 3. Histograms with Kernel density curves (green) showing the distribution of metacarpal Bd of cattle from the Late Neolithic to the Late Bronze Age and the results of mixture analysis modeling grouping (red).
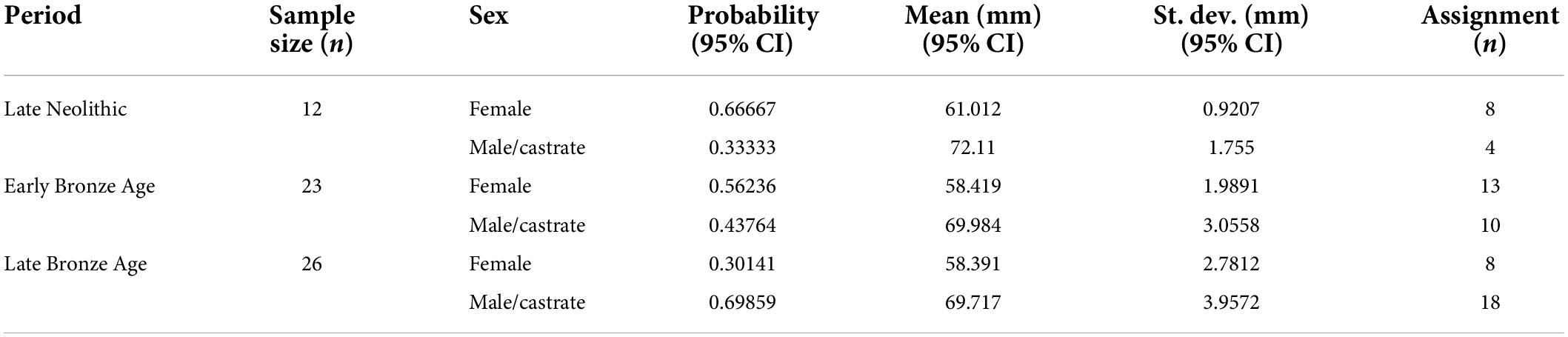
Table 3. Mixture analysis results of metacarpal Bd and numbers of assignment of female and male/castrate cattle in each period.
Table 3 shows a marked decrease in the size of the Early Bronze Age individuals. Both female and male/castrate groups were 3 mm smaller when compared with the Late Neolithic ones. However, no statistically significant difference between the means of the Late Bronze Age and the Early Bronze Age was evident. Moreover, the sex ratio also changed over time. The female probability across assemblages reduced from 66 to 56% and then 30%, and the probable proportions of male/castrate cattle increased accordingly. Finally, there was no significant variation between the size difference of the two sexes with time, as male/castrate metacarpal Bd was larger than female by 9 mm at all times.
Table 4 shows a significant Mann–Whitney result (p ≤ 0.05) between the Late Neolithic female and the Early Bronze Age female as well as Late Bronze Age female, indicating the size change among female individuals through time is obvious. In contrast, size variation among male/castrate individuals is not statistically significant.
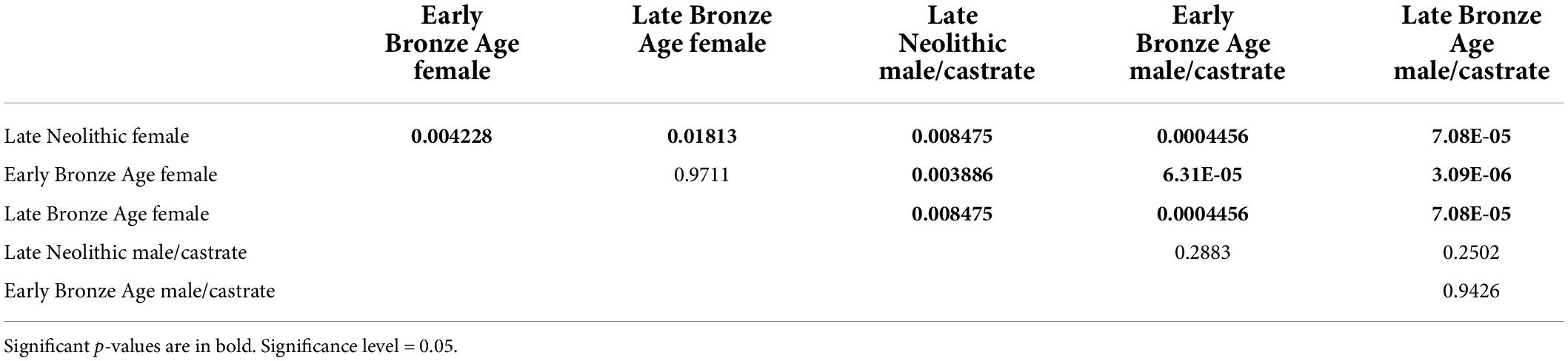
Table 4. Mann–Whitney U test pairwise comparisons of female and male/castrate cattle metacarpal Bd from the Late Neolithic to Late Bronze Age periods.
Discussion and interpretation
Body size variation and natural factors
Previous studies demonstrated that body size reduction was a consequence of domestication, such as pigs, cattle, and caprids in the Near East (Peters et al., 1999; Hongo et al., 2009). After they were introduced to Europe, the body size of cattle decreased from Neolithic to pre-Roman Iron Age (Schibler and Schlumbaumm, 2007; Manning et al., 2015) and then increased from the Iron Age/Roman transition period and beyond (Davis and Beckett, 1999; Albarella et al., 2008; MacKinnon, 2010; Colominas et al., 2014). However, some datasets in Europe addressed that breed improvement indicated by size increase might have happened during the Neolithic and Bronze Age (Gaastra, 2014; Wright, 2021), much earlier than the Roman conquest.
Yet, body size was linked to many factors, including natural and artificial (Reitz and Wing, 2008). In this study, we discussed the possibility of natural factors, which might have caused such variation. Climate and environmental diversity is one significant natural factor that affects animal body size. For instance, biometric data of modern wild goats and ancient aurochs confirmed the presence of geographical variation, with body mass smaller in warmer environments and larger in colder environments (Zeder and Hesse, 2000; Wright and Viner-Daniels, 2015). We would expect that such influence affects all species if this is the main factor related to our dataset. However, pig and sheep biometric data collected from the Late Neolithic to the Late Bronze Age China indicated no distinct variation in animal body size (Luo, 2012; Zuo, 2018). Our data show that the directional size reduction only affected cattle, and therefore, climate and/or environment as causal factors during the post-domestication period can be rejected.
Another natural factor could be hybridization with wild ancestors after domestication. Measurements of cattle postcranial bones revealed a gradual size reduction during the late 9th millennium BC and revealed the early stage of indigenous domestication (Peters et al., 1999; Hongo et al., 2009). The sex-specific analysis also confirmed this trend (Helmer et al., 2005). We can assume that both female and male individuals are smaller than their ancestors of the same sex. Therefore, crossbreeding between female aurochs and male domestic individuals and male aurochs and female domestic individuals would both lead to a size increase. Ancient DNA evidence confirmed the incorporation of wild stock into a domestic cattle herd from a complete metacarpal in Switzerland, which was directly dated to 3,360–3,090 cal. BC. This small and compact adult bovid is morphologically and genetically a female. This cow possessed a novel mtDNA P haplotype variant of the European aurochs, which can be explained as breeding with female aurochs (Schibler et al., 2014). However, the circumstance of hybridization is generally very rare; therefore, we considered that its influence on body size is minor. Taken together, natural factors might not have been the primary cause of cattle body size variation in the post-domestication period.
Cattle body size variation from the Late Neolithic to the Early Bronze Age
From the body size distribution analysis above, we can see that in China, although not significantly supported by the Mann–Whitney U test, the general body size represented by metacarpal Bd decreased from the Late Neolithic to the Early Bronze Age. The decrease in body size was normally explained as the increase in the proportion of smaller individuals, females for instance (Manning et al., 2015). However, the variation of sex ratio over time revealed by mixture analysis suggested that the numbers of female individuals in the Early Bronze Age decreased by 10% (Table 3), thus not supporting the hypothesis of an increase in the proportion of females.
The means of male/castrate and female assigned by mixture analysis proved that both male/castrate and female from the Early Bronze Age were smaller than in earlier periods (Table 3). A possible explanation for this might be that the body weight of reproductive (female) individuals is decreasing. Age at puberty is important as a production trait. The puberty of modern cattle ranged from 10 to 24 months, with an average of 15 months, which means that heifers are bred to calve at the age of 2 years, as the gestation period is approximately 9–10 months (Hafez and Hafez, 2000). However, modern herders make the decision based on weight, not age. Weight is strongly linked to nutritional status, which determines the hormone level of puberty. The traditional rule in heifer development has been to develop heifers to 60–65% of their mature weight by the start of the breeding season. However, the latest animal science research suggested that from an economic standpoint, developing heifers to 50–55% of their mature weight, may yield more economical results as heifers that conceive early in the first breeding season have greater lifetime production. However, smaller heifer development weights may mean lower pregnancy rates and lower birth weights (Funston and Deutscher, 2004).
A similar hypothesis has been raised by Manning et al. (2015) as the size reduction was caused by a younger subadult reproduction age. In fact, weight, size, and age are significantly correlated (Reitz and Wing, 2008), the shift in size suggested changes in weight and/or age. Thus, the fundamental mechanism behind this observation is the lightening of heifer target weights. In modern systems that impose restricted breeding periods, calving by 24 months of age is necessary to obtain maximum lifetime productivity (Patterson et al., 1992). Although we are unable to evaluate the exact age of puberty and calving of ancient cows, the smaller size of metacarpus Bd from the Early Bronze Age is linked to a group of cattle that was delivered by lighter (smaller and younger) heifers, when compared with those from the Late Neolithic.
Cattle body size variation from the Early Bronze to the Late Bronze Age
In contrast, cattle body size increased during the Late Bronze Age compared with the Early Bronze Age. The size increase was usually interpreted as breed improvement, a direct correlation with meat yield and traction (Albarella et al., 2008). However, in our dataset, the general size increase of the assemblage does not coincide with the size increase of individuals. Mixture analysis addressed that the means of assigned female and male/castrate of the Late Bronze Age is almost the same as the Early Bronze Age. The reason for this general gain in size is due to the increased proportion of male/castrate individuals from 44 to 70% (Table 3). So, why did the slaughtering pattern of cattle undergo such a substantial shift in sex ratio?
As cattle is an animal that can be exploited for multiple purposes including meat, milk, and traction. Based on the values one can provide from different age stages and sexes, mortality pattern was widely used to chase herders’ decisions. For example, if meat production is the aim, most individuals are killed when they reach the optimum point in weight gain (Payne, 1973). The high percentage of slaughtered male claves, in contrast, is strongly linked to the intensive exploitation of dairy products (Balasse and Tresset, 2002; Balasse, 2003).
In Yuntang, the survivorship of cattle was generated from epiphyseal fusion order and timings (Silver, 1969), as mandibles were not brought to the site. The result suggested that 100% of the individuals survived through the first year and 99% survived till the age of 2 years. Sizable slaughter started during the 2–3 years old stage; approximately, 31% of the cattle were killed. Notably, 39% more were slaughtered during their 3–4 years old, and only 29% of the cattle were able to survive beyond 4 years. Overall, the intensive slaughter started after 2 years of age, and the majority of cattle were butchered before reaching 4 years old (Zhao, 2017).
Yuntang is a bone workshop, and the source of cattle bones for manufacturing was thought to be originated from settlement sites outside of Yuantang across the Zhouyuan area. Thus, the complete raw materials and offcuts that provided fusion information did not represent the cattle slaughtering pattern well in the Late Bronze Age in general. This is because the bones of immature individuals may not have been transported to Yuntang for manufacturing as they were not robust enough. Therefore, we are unable to rule out the possibility of immature culling in the settlement sites, which is linked to sophisticated milk exploitation, even though nearly no individuals aged younger than 2 years existed in Yuntang.
However, we can still use this pattern to investigate the characteristic of cattle utilization in the West Zhou Dynasty. Most of the cattle in Yuntang were killed within a concentrated age range of 2–4 years. The age of 2–4 years is basically an ideal and reasonable time for butchering cattle for beef. This is because cattle would reach adult size around 2 years old. Thereafter, their size and weight would maintain stability, thus the economic efficiency of keeping cattle older than 2 years would decrease gradually (Greenfield, 1988). Thus, we considered that the majority of cattle (approximately 70%) at Yuntang were butchered mainly for producing beef. Meanwhile, less than 30% of cattle lived longer than 4 years. It seems that not many cattle were kept for long-term secondary production. The Yuntang assemblage, therefore, is considered a good dataset representing the range and distribution of cattle body sizes that were killed for meat consumption in the Late Bronze Age.
The meat provision slaughtering pattern not only demonstrated an age preference but also a sex preference. Since most of the male and castrated individuals were not needed for herd reproduction, only a few being kept for breeding. Although the present dataset did not provide enough information on the identification of oxen, we may still infer that the change of sex ratio with time was driven by intensifying meat production strategies relying on the culling of male or castrated individuals around 2–4 years old. This might be the reason why the general body size of the Late Bronze Age showed a marked increase in the median value (Figure 2).
Although biometrical data from the Middle Bronze Age (Shang Dynasty and the Pre-Zhou period, 1600–1050 BC) is scarce at the moment, the consistency of investigation on cattle husbandry during the Bronze Age is thus limited. Relative proportion, in contrast, may provide direct evidence of herders’ preference. Relative proportions of cattle, pig, and caprids NISP from sites in Northern China during the Late Neolithic and the Late Bronze Age show a marked increase of cattle remains since the Middle Bronze Age (Figure 4; Yuan, 2000; Zhou, 2007; Hu et al., 2008; Hou et al., 2009, 2018, 2019; Shaanxi Provincial Institute of Archaeology et al., 2010; Lin et al., 2013; Li et al., 2014, 2020; Yu, 2019). During the Late Neolithic and the Early Bronze Age, the relative proportion of cattle never exceed one-third of the main domesticates, and the average is 13% in the Late Neolithic and 20% in the Early Bronze Age. In the Middle Bronze Age sites of central China, cattle took up approximately 35–60% of the main three, and the average increased to 43%. During the Late Bronze Age, the average proportion increased to 48% (Yuntang was excluded as other sites were all settlements). When calculated in terms of the contribution to the primary and secondary products per head, the significance of cattle husbandry in the Middle and the Late Bronze Age times is undeniable.
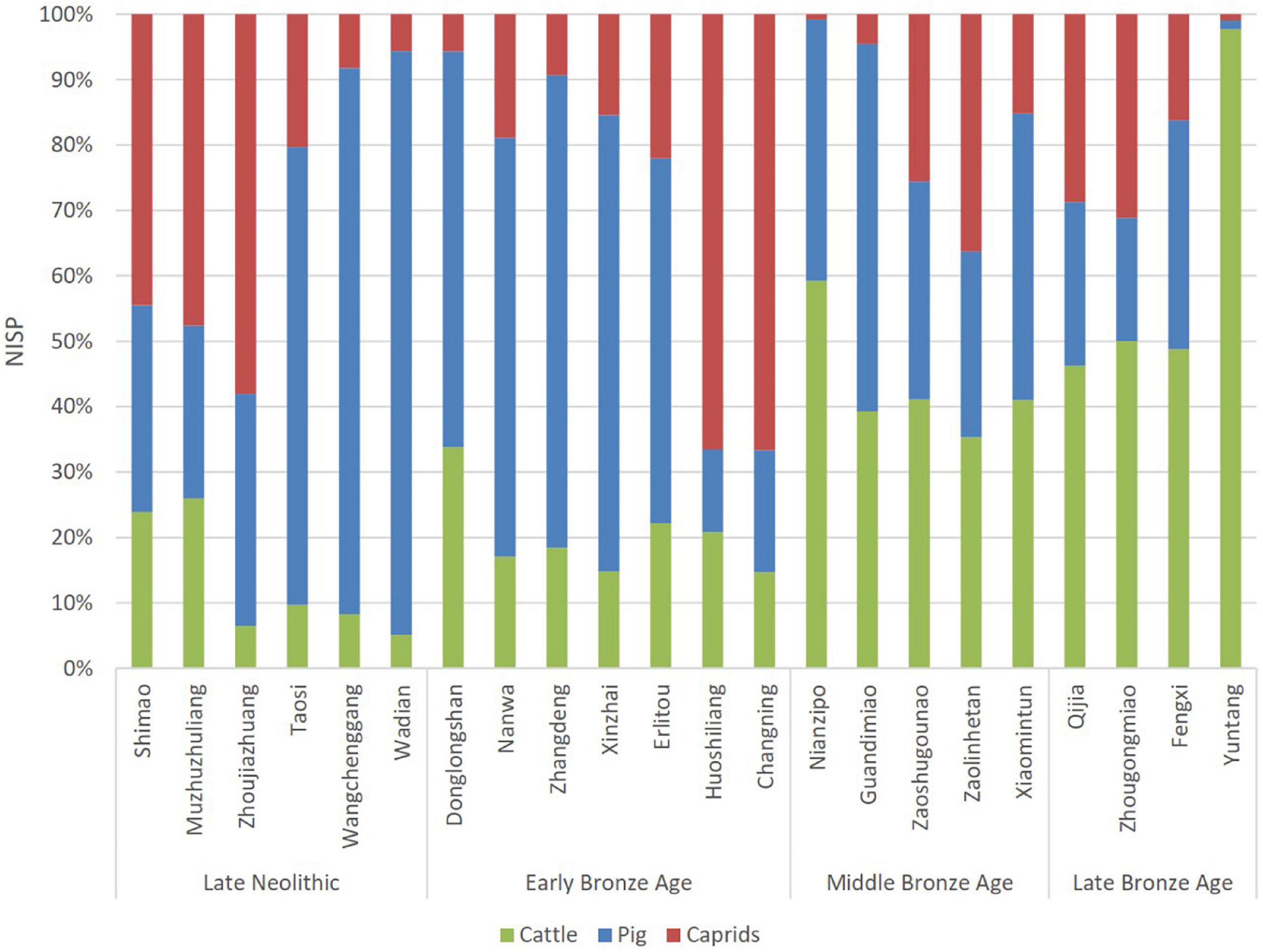
Figure 4. Relative proportions of cattle remains among cattle, pig, and caprids NISP of the studied sites.
Conclusion
Osteometric data for cattle from eight assemblages from the Late Neolithic to the Late Bronze Age in China were used to evaluate the differences in body size and to explore the effect of sex ratio on size variation, based on mixture analysis using the measurements of the greatest distal breadth (Bd) of the metacarpus. Evidence for time trends shows that the general body size decreased from the Late Neolithic to the Early Bronze Age and then increased from the Early Bronze Age to the Late Bronze Age.
Sex ratio estimated by mixture analysis demonstrated an increase in male/castrate individuals with time. The average mean size of each sex reduced from the Late Neolithic to the Early Bronze Age but showed no differences in size between the Early Bronze Age and the Late Bronze Age. A similar range of size reduction was detected in both sexes, suggesting that the difference between male/castrate and female breeding strategies was minor. An early shift in body size might be correlated to the lightening of heifer target weights, resulting in faster replacement of females and feed cost reduction. Later shift in sex ratio might be correlated to a slaughtering preference for male/castrate individuals. Both were linked to meat production in the Bronze Age times.
Evidenced by biometrical assessments, in conjunction with taxonomic abundances and mortality profiles, this study addressed that although also used for other purposes such as milk, traction, and ritual, the primary motivation for cattle husbandry in Bronze Age China was to fulfill the demands of beef provision. Further investigations with larger sample sizes from the Middle Bronze Age and settlement sites from the Late Bronze Age are required to confirm this cattle utilization pattern in early China.
Data availability statement
The original contributions presented in this study are included in the article/supplementary material, further inquiries can be directed to the corresponding authors.
Author contributions
CY: conceptualization, methodology, formal analysis, data curation, funding acquisition, writing—original draft, and writing—review and editing. HZ, SH, and MY: resources. XG: fieldwork administration and resources. CN: methodology and writing—review and editing. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by the National Social Science Fund of China (Grant No. 18CKG005).
Acknowledgments
We thank Zixian Jiang for her contribution to photo editing. We are also grateful to the reviewers who helped us improve the final version of this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Albarella, U. (1997). Shape variation of cattle metapodials: age, sex or breed? Some examples from medieval and postmedieval sites. Anthropozoologica 2, 37–47.
Albarella, U., Johnstone, C., and Vickers, K. (2008). The development of animal husbandry from the Late Iron Age to the end of the Roman period: a case study from South-East Britain. J. Archaeol. Sci. 35, 1828–1848. doi: 10.1016/j.jas.2007.11.016
Arbuckle, B. S., and Makarewicz, C. A. (2009). The early management of cattle (Bos taurus) in Neolithic central Anatolia. Antiquity 83, 669–686. doi: 10.1017/S0003598X00098902
Balasse, M. (2003). Keeping the young alive to stimulate milk production? Differences between cattle and small stock. Anthropozoologica 37, 3–10.
Balasse, M., and Tresset, A. (2002). Early weaning of Neolithic domestic cattle (Bercy, France) revealed by Intra-tooth variation in nitrogen isotope ratios. J. Archaeol. Sci. 29, 853–859. doi: 10.1006/jasc.2001.0725
Bollongino, R., Burger, J., Powell, A., Mashkour, M., Vigne, J. D., and Thomas, M. G. (2012). Modern taurine cattle descended from small number of Near-Eastern founders. Mole. Biol. Evolut. 29, 2101–2104. doi: 10.1093/molbev/mss092
Bollongino, R., Edwards, C. J., Alt, K. W., Burger, J., and Bradley, D. G. (2006). Early history of European domestic cattle as revealed by ancient DNA. Biol. Lett. 2, 155–159. doi: 10.1098/rsbl.2005.0404
Bollongino, R., Elsner, J., Vigne, J. D., and Burger, J. (2008). Y-SNPs do not indicate hybridisation between European aurochs and domestic cattle. PLoS One 3:e3418. doi: 10.1371/journal.pone.0003418
Brunson, K., He, N., and Dai, X. (2015). Sheep, cattle, and specialization: new zooarchaeological perspectives on the Taosi Longshan. Int. J. Osteoarchaeol. 26, 460–475. doi: 10.1002/oa.2436
Cai, D., Sun, Y., Tang, Z., Hu, S., Li, W., Zhao, X., et al. (2014). The origins of Chinese domestic cattle as revealed by ancient DNA analysis. J. Archaeol. Sci. 41, 423–434. doi: 10.1016/j.jas.2013.09.003
Campbell, R. B., Li, Z., He, Y., and Yuan, J. (2011). Consumption, exchange and production at the Great Settlement Shang: bone-working at Tiesanlu. Anyang. Antiq. 85, 1279–1297. doi: 10.1017/S0003598X00062050
Chen, N., Ren, L., Du, L., Hou, J., Mullin, V. E., Wu, D., et al. (2020). Ancient genomes reveal tropical bovid species in the Tibetan Plateau contributed to the prevalence of hunting game until the late Neolithic. Proc. Natl. Acad. Sci. U.S.A. 117, 28150–28159. doi: 10.1073/pnas.2011696117
Chen, X., Guo, X., Hu, Y., Wang, W., and Wang, C. (2015). Analysis of the subsistence practice at the Muzhuzhuliang site in Shenmu, Shaanxi. Archaeol. Cult. Relics 5, 112–117.
Chen, X., Yuan, J., Hu, Y., He, N., and Wang, C. (2012). Preliminary study on the feeding strategy of domesticated animals in Taosi site: evidences from stable isotope. Archaeology 540, 75–82.
Colominas, L., Schlumbaum, A., and Saña, M. (2014). The impact of the Roman Empire on animal husbandry practices: study of the changes in cattle morphology in the north-east of the Iberian Peninsula through osteometric and ancient DNA analyses. Archaeol. Anthropol. Sci. 6, 1–16. doi: 10.1007/s12520-013-0116-9
Davis, S. J. M. (1996). Measurements of a group of adult female Shetland sheep skeletons from a single flock: a baseline for zooarchaeologists. J. Archaeol. Sci. 23, 593–612. doi: 10.1006/jasc.1996.0056
Davis, S. J. M. (2000). The effect of castration and age on the development of the Shetland sheep skeleton and a metric comparison between bones of males, females and castrate. J. Archaeol. Sci. 27, 373–390. doi: 10.1006/jasc.1999.045
Davis, S. J. M. (2008). Zooarchaeological evidence for Moslem and Christian improvements of sheep and cattle in Portugal. J. Archaeol. Sci. 35, 991–1010. doi: 10.1016/j.jas.2007.07.001
Davis, S. J. M., and Beckett, J. (1999). Animal husbandry and agricultural improvement: the archaeological evidence from animal bones and teeth. Rural Hist. 10, 1–17.
Davis, S. J. M., Svensson, E. M., Albarella, U., Detry, C., Götherström, A., Pires, A. E., et al. (2012). Molecular and osteometric sexing of cattle metacarpals: A case study from 15th century AD Beja. Portugal. J. Archaeol. Sci. 39, 1445–1454. doi: 10.1016/j.jas.2011.12.003
Drennan, R. D., and Dai, X. (2010). Chiefdoms and states in the Yuncheng Basin and the Chifeng region: A comparative analysis of settlement systems in North China. J. Anthropol. Archaeol. 29, 455–468. doi: 10.1016/j.jaa.2010.09.001
Edwards, C. J., Bollongino, R., Scheu, A., Chamberlain, A., Tresset, A., and Vigne, J. D. (2007). Mitochondrial DNA analysis shows a Near Eastern Neolithic origin for domestic cattle and no indication of domestication of European aurochs. Proc. R. Soc. B. 274, 1377–1385. doi: 10.1098/rspb.2007.0020
Evershed, R. P., Payne, S., Sherratt, A. G., Copley, M. S., Coolidge, J., and Urem-Kotsu, D. (2008). Earliest date for milk use in the Near East and southeastern Europe linked to cattle herding. Nature 455, 528–531. doi: 10.1038/nature07180
Funston, R. N., and Deutscher, G. H. (2004). Comparison of target breeding weight and breeding date for replacement beef heifers and effects on subsequent reproduction and calf performance. J. Anim. Sci. 82, 3094–3099. doi: 10.2527/2004.82103094x
Gaastra, J. S. (2014). Shipping sheep or creating cattle: domesticate size changes with Greek colonisation in Magna Graecia. J. Archaeol. Sci. 52, 483–496. doi: 10.1016/j.jas.2014.08.030
Greenfield, H. J. (1988). The origins of milk and wool production in the Old World: a zooarchaeological perspective from Central Balkans. Curr. Anthropol. 29, 573–593.
Grigson, C. (1989). Size and Sex: Evidence for the Domestication of Cattle in the Near East. Oxford: British Archaeological Reports.
Hafez, B., and Hafez, E. (2000). Reproduction in Farm Animals, 7th Edn. Philadelphia, USA: Lippincott Williams & Wilkins.
Hammer, Ø, Harper, D. A. T., and Ryan, P. D. (2001). PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electr. 4:9.
Helmer, D., Gourichon, L., Monchot, H., Peters, J., and Segui, M. S. (2005). “Identifying early domestic cattle from Pre-Pottery Neolithic sites on the Middle Euphrates using sexual dimorphism,” in The First Steps of Animal Domestication, eds J.-D. Vigne, J. Peters, and D. Helmer (Oxford: Oxbow Books), 86–95.
Higham, C. (1969). The metrical attributes of two samples of bovine limb bones. J. Zool. 157, 63–74. doi: 10.1111/j.1469-7998.1969.tb01689.x
Hongo, H., Pearson, J., Öksüz, B., and Ilgezdi, G. (2009). The process of ungulate domestication at Cayönü, Southeastern Turkey: a multidisciplinary approach focusing on Bos sp. and Cervus elaphus. Anthropozoologica 44, 63–78.
Hou, Y., Campbell, R., Li, Z., Zhang, Y., Li, S., and He, Y. (2018). The Guandimiao bone assemblage (and what it says about the Shang economy). Asian Perspect. 57, 281–310. doi: 10.1353/asi.2018.0018
Hou, Y., Campbell, R., Zhang, Y., and Li, S. (2019). Animal use in a Shang village: The Guandimiao zooarchaeological assemblage. Int. J. Osteoarchaeol. 29, 335–345. doi: 10.1002/oa.2745
Hou, Y., Li, S., Ma, X., and Sun, L. (2009). The exploitation of animal resources at Zhangdeng site in Anyang. Cult. Relics Central China 5, 38–47.
Hu, S. (2011). “Analysis on the faunal remains from the Donglongshan site, Shangluo,” in Donglongshan in Shangluo, eds Shaanxi Provincial Institute of Archaeology and Shangluo Museum (Beijing: Science Press), 312–430.
Hu, S., Yang, M., Sun, Z., and Shao, J. (2016). Research on faunal remains from the 2012 – 2013 season excavation at the Shimao site in Shenmu, Shaanxi. Archaeol. Cult. Relics 4, 109–121.
Hu, S., Zhang, P., and Yuan, M. (2008). A study on the faunal remains from the Huoshiliang site in Yulin, Shaanxi. Acta Anthropol. Sin. 27, 232–248.
Jones, G., and Sadler, P. (2012). Age at death in cattle: methods, older cattle and known-age reference material. Environ. Archaeol. 17, 11–28. doi: 10.1179/1461410312Z.0000000002
Li, L. (2012). Study on the animal remains from the site of changning. Master thesis. Changchun: Jilin University.
Li, Y., Zhang, C., Wang, Z., Dou, H., Liu, H., Hou, F., et al. (2020). Animal use in the late second millennium BCE in northern China: Evidence from Zaoshugounao and Zaolinhetan in the Jing River valley. Int. J. Osteoarchaeol. 30, 318–329. doi: 10.1002/oa.2860
Li, Z., Campbell, R., Brunson, K., Yang, J., and Tao, Y. (2014). “The exploitation of domestic animal products from the late Neolithic Age to the early Bronze Age in the heartland of ancient China,” in Animal Secondary Products: Domestic Animal Exploitation in Prehistoric Europe, the Near East and the Far East, ed. H. J. Greenfield (Oxford: Oxbow Books), 56–79.
Lin, Y., Chong, J., and Lei, X. (2013). Preliminary analysis on the utilization of animal resources during the Shang-Zhou period at the Zhougongmiao site. Archaeol. Cult. Relics 3, 39–47.
Luo, Y. (2012). The Domestication, Raising and Ritual Use of Pig in Ancient China. Beijing: Science Press.
MacKinnon, M. (2010). Cattle “breed” variation and improvement in Roman Italy: connecting the zooarchaeological and ancient textual evidence. World Archaeol. 42, 55–73. doi: 10.1080/00438240903429730
Manning, K., Timpson, A., Shennan, S., and Crema, E. (2015). Size reduction in early European domestic cattle relates to intensification of Neolithic herding strategies. PLoS One 10:e0141873. doi: 10.1371/journal.pone.0141873
Meadow, R. (1999). “The use of size index scaling techniques for research on archaeozoological collections from the Middle East,” in Historia Animalium ex Ossibus Festschrift für Angela von den Driesch, eds C. Becker, H. Manhart, J. Peters, and J. Schibler (Rahden: Westf), 285–300.
Minniti, C., and Abatino, C. (2022). Biometric variation of domestic animals in Rome from the Orientalizing/Archaic period to the Middle Ages. Quater. Int. [preprint] doi: 10.1016/j.quaint.2022.06.010
Monchot, H., Mashkour, M., and Vigne, J.-D. (2005). “Kernel smoothing and mixture analyses for the determination of the sex ratios at death, at the beginning of the domestication of ungulates,” in The First Steps of Animal Domestication, eds J.-D. Vigne, J. Peters, and D. Helmer (Oxford: Oxbow Books), 55–60.
O’Connor, T. (1991). Bones from 46–54 Fishergate, The Archaeology of York 15/4. London: Council for British Archaeology.
Patterson, D. J., Perry, R. C., Kiracofe, G. H., Bellows, R. A., Staigmiller, R. B., and Corah, L. R. (1992). Management considerations in heifer development and puberty. J. Anim. Sci. 70, 4018–4035. doi: 10.2527/1992.70124018x
Payne, S. (1973). Kill-off patterns in sheep and goats: The mandibles from Aşvan Kale. Anatol. Stud. 23, 281–303. doi: 10.2307/3642547
Peters, J., Helmer, D., von, den Driesch, A., and Sana Segui, M. (1999). Early animal husbandry in the northern Levant. Paléorient 25, 27–48.
Prummel, W., and Niekus, M. (2011). Late Mesolithic hunting of a small female aurochs in the valley of the River Tjonger (the Netherlands) in the light of Mesolithic aurochs hunting in NW Europe. J. Archaeol. Sci. 38, 1456–1467. doi: 10.1016/j.jas.2011.02.009
Schibler, J., Elsner, J., and Schlumbaum, A. (2014). Incorporation of aurochs into a cattle herd in Neolithic Europe: single event or breeding? Sci. Rep. 4:5798. doi: 10.1038/srep05798
Schibler, J., and Schlumbaumm, A. (2007). Geschichte und wirtschaftliche Bedeutung des Hausrindes (Bos taurus L.) in der Schweiz von der Jungsteinzeit bis ins fruhe Mittrlalter. Schweiz. Arch. Tierh. 149, 23–29. doi: 10.1024/0036-7281.149.1.23
Shaanxi Provincial Institute of Archaeology (2015). Preliminary report on the excavation of the Shengedaliang site in Shenmu, Shaanxi. Archaeol. Cult. Relics 5, 3–11.
Shaanxi Provincial Institute of Archaeology (2016). Preliminary report on the excavation of the Shengedaliang site in Shenmu, Shaanxi. Archaeol. Cult. Relics 4, 34–44.
Shaanxi Provincial Institute of Archaeology, School of Archaeology and Museology of Peking University, and The Institute of Archaeology Chinese Academy of Social Sciences, and Zhouyuan Archaeological Team (2010). Zhouyuan: Excavation of Qijia and Licun in 2002. Beijing: Science Press.
Shaanxi Provincial Institute of Archaeology, and Shangluo Museum (2011). Donglongshan in Shangluo. Beijing: Science Press.
Sherratt, A. (1981). “Plough and pastoralism: aspects of the secondary products revolution,” in Pattern of the Past: Studies in Honour of David Clarke, eds I. Hodder, G. Isaac, and N. Hammond (Cambridge: Cambridge University Press), 261–305.
Silver, I. A. (1969). “The ageing of domestic animals,” in Science in Archaeology: A Comprehensive Survey of Progress and Research, eds D. R. Brothwell and E. S. Higgs (London: Thames and Hudson), 283–302.
Svensson, E. M., Götherström, A., and Vretemark, M. (2008). A DNA test for sex identification in cattle confirms osteometric results. J. Archaeol. Sci. 35, 942–946. doi: 10.1016/j.jas.2007.06.021
Telldahl, Y., Götherström, A., and Storå, J. (2012). Osteometric and molecular sexing of cattle metapodial. J. Archaeological Sci. 39, 121–127. doi: 10.1016/j.jas.2011.09.009
von den Driesch, A. (1976). A Guide to Measurement of Animal Bones from Archaeological Sites. Cambridge: Peabody museum Bulletin.
Wright, E. (2021). Investigating cattle husbandry in the Swiss Late Neolithic using different scales of temporal precision: potential early evidence for deliberate livestock “improvement” in Europe. Archaeol. Anthropol. Sci. 13:36. doi: 10.1007/s12520-020-01252-6
Wright, E., and Viner-Daniels, S. (2015). Geographical variation in the size and shape of the European aurochs (Bos primigenius). J. Archaeol. Sci. 54, 8–22. doi: 10.1016/j.jas.2014.11.021
Wu, W., Zhang, J., and Jin, G. (2014). Archaeobotanical analysis on Nanwa site, Dengfeng, Henan. Cult. Relic. Central China 1, 109–117.
Yang, M., Hu, S., Guo, X., Wang, W., and Yang, T. (2021). Research on the faunal remains from Muzhuzhuliang site in Shenmu city Shaanxi Province. Acta Anthropol. Sin. 40, 394–405. doi: 10.16359/j.1000-3193/AAS.2021.0050
Yu, C. (2014). “The identification and the research on the faunal remains from Nanwa site,” in Nanwa in Dengfeng, ed. Department of Archaeology Zhengzhou University (Beijing: Science Press), 788–795.
Yu, C. (2019). “The application of big data in zooarchaeology: subsistence patterns of Neolithic and Early Bronze Age China using published mammal records,” in Zooarchaeology 3, ed. Henan Provincial Institute of Cultural Heritage and Archaeology (Beijing: Cultural Relics Press), 127–141.
Yu, C. (2020). The Origin of Cattle in China from the Neolithic to the Early Bronze Age. Oxford: British Archaeological Reports. doi: 10.30861/9781407316871
Zeder, M. A., and Hesse, B. (2000). The initial domestication of goats (Capra hircus) in the Zagros Mountains 10,000 years ago. Science 287, 2254–2257. doi: 10.1126/science.287.5461.2254
Zhang, Y., Sun, G., Wang, Y., Huang, Y., Kikuchi, H., and Yang, X. (2022). Sustainable hunting strategy of Sika Deer (Cervus nippon) in the Neolithic Lower Yangtze River Region China. Front. Earth Sci. 9:812910. doi: 10.3389/feart.2021.812910
Zhao, H. (2017). Mass Bone-Working Industry in the Western Zhou period (1046-771 BC). Ph.D thesis, Stanford: Stanford University.
Zhao, Z., and He, N. (2006). Result and analysis of the flotation of Taosi site 2002. Archaeology 5, 77–86.
Zhou, B. (2007). “Research on the faunal remains from Nianzipo site,” in Nanbinzhou and Nianzipo, ed. The Institute of Archaeology Chinese Academy of Social Sciences (Beijing: World Publishing Corporation), 490–492.
Keywords: cattle husbandry, body size, sex ratio, Late Neolithic, Bronze Age, China
Citation: Yu C, Zhao H, Hu S, Yang M, Guo X and Ng C (2022) A passion for beef: Post-domestication changes in cattle body size in China from the Late Neolithic to the Late Bronze Age. Front. Ecol. Evol. 10:949860. doi: 10.3389/fevo.2022.949860
Received: 21 May 2022; Accepted: 29 July 2022;
Published: 22 August 2022.
Edited by:
Xinyi Liu, Washington University in St. Louis, United StatesReviewed by:
Li Liu, Stanford University, United StatesHelina Woldekiros, Washington University in St. Louis, United States
Copyright © 2022 Yu, Zhao, Hu, Yang, Guo and Ng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chong Yu, eXVjaG9uZzNAbWFpbC5zeXN1LmVkdS5jbg==; Chuenyan Ng, bXJuY3lAb3V0bG9vay5jb20=