 Chloé Vagnon
Chloé Vagnon Rudolf P. Rohr2
Rudolf P. Rohr2- 1University Savoie Mont Blanc, INRAE, CARRTEL, Thonon-les-Bains, France
- 2Department of Biology—Ecology and Evolution, University Fribourg, Fribourg, Switzerland
- 3HES-SO/HEPIA, Route de Presinge, Jussy, Switzerland
The impacts of invasive species on resident communities are driven by a tangle of ecological interactions difficult to quantify empirically. Combining a niche model with a population dynamic model, both allometrically parametrized, may represent a consistent framework to investigate invasive species impacts on resident communities in a food web context when empirical data are scarce. We used this framework to assess the ecological consequences of an invasive apex predator (Silurus glanis) in peri-Alpine lake food webs. Both increases and decreases of resident species abundances were highlighted and differed when accounting for different S. glanis body sizes. Complementarily, the prominence of indirect effects, such as trophic cascades, suggested that common approaches may only capture a restricted fraction of invasion consequences through direct predation or competition. By leveraging widely available biodiversity data, our approach may provide relevant insights for a comprehensive assessment and management of invasive species impacts on aquatic ecosystems.
Introduction
Invasive species represent a major threat to biodiversity through the alterations or extinctions of native populations (Lockwood et al., 2013). Alternatively, invasive species can provide ecological benefits in specific cases possibly mitigating their negative impacts within recipient ecosystems (e.g., resources for native species; Schlaepfer et al., 2011). However, the empirical quantification of both negative (e.g., native population decreasing through predation; Mills et al., 2004) and positive impacts of invasive species (e.g., resource acquisition facilitation for native species; Albertson et al., 2021) toward resident species requires extensive field investigations involving important human and financial costs (Diagne et al., 2020). Moreover, identifying all possible interspecific interactions (direct and indirect) and quantifying their influences on native population abundances are particularly challenging to achieve through field investigations (Crystal-Ornelas and Lockwood, 2020). Consequently, modeling approaches can represent a keystone to circumvent these methodological constraints and forecast repercussions of biological invasions on resident populations (Kamenova et al., 2017).
Few quantitative methods allow predicting invasive species impacts on native populations, and mostly rely on the relationships between resource availability and their intake by consumers (Dick et al., 2014). This relationship has been updated by integrating consumer abundances (Dick et al., 2017), species propagule pressures (i.e., RIR = relative invasion risk; Dickey et al., 2018) or considering functional response ratios (i.e., FRR = attack rate/handling time; Cuthbert et al., 2019). Yet, these methods are not explicitly embedded in a dynamic perspective and have mostly addressed impacts of invaders on reduced communities in experimental conditions (Alexander et al., 2013; Barrios-O’Neill et al., 2014; Cuthbert et al., 2018), leaving a comprehensive assessment of invasive species impact in-situ on wide resident communities out of reach (Frost et al., 2019). However, noticeable invader impacts usually initiate from direct interspecific interactions (i.e., predation; Mills et al., 2004) but can also propagate along food webs, causing indirect repercussions called trophic cascades that can be especially frequent in aquatic ecosystems (Shurin et al., 2002; Carpenter et al., 2010). These ecological interactions are recognized to structure global biodiversity patterns across trophic levels (Zhang et al., 2018) and population dynamics and their disturbance may ultimately affect species assemblages and the whole ecosystem functioning (Terry et al., 2018; Frost et al., 2019).
Identifying direct and indirect ecological interactions between invasive species and resident communities would therefore be a prerequisite for quantifying invader impacts, yet their empirical assessment through traditional methods (e.g., stomach contents) remains challenging when considering the whole food web scale (i.e., primary producers to apex predator). To answer these methodological limitations, species body size is a widely used trait to infer trophic interactions, particularly in aquatic ecosystems (Petchey et al., 2008; Gravel et al., 2013; Pomeranz et al., 2019), as it allometrically relates to most species biological rates (e.g., respiration, reproduction; Brown et al., 2004) and population characteristics (e.g., trophic levels, abundances; Brucet et al., 2017). Trophic interactions could then be scaled up to population dynamic models to investigate species persistence toward destabilizing factors at the whole food web scale (Brose et al., 2006).
In this study, we used an approach combining an allometric niche model with a population dynamic model, both allometrically scaled, to grasp the diversity of impacts (direct/indirect, negative/positive) that invasive species can exert on wide resident communities (from primary producers to the apex predator) in aquatic ecosystems. More precisely, we reconstructed a non-invaded and an invaded food web by inferring trophic interactions among multiple species clusters (S = 58) in presence and in absence of an invader using an allometric niche model (hereafter called aNM; Vagnon et al., 2021). These interactions were then included in a population dynamic model, based on the model from Brose et al. (2006) that we modified by using alternative allometric parameterization, allowing to measure abundance changes of resident species over time in absence/presence of the invader with few empirical data inputs. We computed impact metrics supporting the classification of population abundance modifications and extinctions. Specifically, we distinguished positive or negative impacts on abundances (i.e., increases or decreases) and we characterized the ecological interactions involved in these modifications (i.e., predation, competition, trophic cascades).
We applied this approach to assess the ecological consequences of the recent invasion (∼10 years) of the European catfish (Silurus glanis) in large peri-Alpine lakes that are representative of successful water quality restauration plans and biodiversity management which both could be threatened by new pressures originating from modifications of resident species abundances following invasive species introductions. In order to provide global analyses and results valuable for large peri-Alpine lakes, we applied our approach to a dataset including species co-occurring in Lake Bourget, Lake Geneva and Lake Annecy. By combining food web theory and a population dynamic model, we aimed to answer three major questions regarding the impacts that S. glanis could exert on a resident community typical of large French peri-Alpine lakes. First, we asked which S. glanis body size could cause the highest changes on species abundances. We hypothesized that large S. glanis (>100 cm) could induce the greatest magnitude in species abundance changes due to its highest position in the food web. We then focused at identifying whether direct or indirect negative ecological interactions (i.e., predation, competition and trophic cascade decreasing species abundances) could be balanced by indirect positive interactions (i.e., trophic cascades increasing species abundances). It was expected that the detrimental effects of S. glanis could be balanced by trophic cascades characterized by amplified species abundances at lower trophic levels. Overall, our study highlighted the possible complex consequences of a new invasive species in large peri-Alpine lakes.
Materials and methods
Species inventory
We used species inventories from the three largest peri-Alpine French lakes: Lake Annecy (45°51′ 41.489″ N, 6°10′ 2.364″ E), Lake Bourget (45°43′ 46.842″ N, 5°52′ 10.484″ E) and Lake Geneva (46°26′ 27.213″ N, 6°30′ 38.177″ E) originating from annual monitoring surveys (i.e., recording of environmental parameters and biodiversity samplings) and scientific reports (©SOERE OLA-IS, INRAE Thonon-les-Bains, SILA, CISALB, CIPEL; Rimet et al., 2020). As these lakes are highly similar in terms of biodiversity (Jacquet et al., 2014), species co-occurring in the three lakes (n = 118; from primary producers to large vertebrates) were retained to represent typical species of these ecosystems and were characterized by their taxonomy (i.e., subphylum, group, family, genus, species), their average body size (μm) commonly used in community ecology studies, a habitat trait (i.e., littoral, pelagic/littoral or pelagic) and a feeding trait (i.e., carnivorous, omnivorous, herbivorous or primary producer) referenced in species inventories. Animal species were then clustered to the family level (S = 48) and vegetal species were clustered to the class level (S = 10). This taxonomic aggregation led to 58 species clusters (hereafter called SC; Supplementary Table 1) gathered according to similar ecological functions/requirements and could hence be considered as functional nodes in the reconstructed food webs (Allesina and Pascual, 2009). This clustering procedure allowed to avoid unnecessary food web complexity by taking into account the main SC that may be directly or indirectly impacted by catfish in line with recommendations emerging from recent ecosystem modeling studies (Geary et al., 2020), to promote the computational efficiency and to facilitate ecological interpretations of the processes involved in abundance changes of SC due to S. glanis.
Allometric niche model and silurus glanis body size selection
We used the aNM (Vagnon et al., 2021) to infer trophic interactions between SC included in the typical peri-Alpine lake food web and to reconstruct the “non-invaded” (i.e., without S. glanis) and “invaded” food webs (i.e., with S. glanis). This model relies on the niche model principles (Williams and Martinez, 2000) stating that the niche position of consumer j is given by its average body size bsj and that its resources fall within a body size range bs_rj centered on bs_cj. The range bounds were estimated using quantile regressions (i.e., bs_rjmin = QR at 5% and bs_rjmax = QR at 95%) as suggested by Gravel et al. (2013) and are specifically fitted whether consumers are vertebrate or invertebrate (Vagnon et al., 2021).
The aNM allowed obtaining a binary squared matrix (Mb) of trophic link occurrences of the whole food webs. These links were then weighted for each consumer j considering a Gaussian probability density function, similarly to Williams et al. (2010), with μj = bs_cj and σj = standard deviation of 100 points evenly spaced over bs_rj (i.e., scaling of the normal distribution to bs_rj). Weighted links were then normalized by the maximum value of the normal distribution to obtain a maximum weighting (0.95) at bs_cj and a minimal weighting (0.1) corresponding to prey SC with body sizes at bs_rj bounds. The resulting weighted links were finally converted as the proportion of resources i in the diet of consumer j (ωji, eq. 1), so that the total proportion of species in the diet of consumer j sums to 1.
In a first step, trophic interactions between SC were first inferred without S. glanis to reconstruct the food web before invasion. Trophic positions (TP) of resident species were estimated according to the method of Levine (1980) with TPPrimary producers = 1 and TPConsumers = 1 + mean (TPResources).
In a second step, we independently introduced 40 nodes representing S. glanis with increasing body sizes (i.e., 5–200 cm by 5 cm) in the SC inventory to study changes in the food web topology as S. glanis could cause different impacts due to its size-dependent diet (i.e., ontogenetic diet shift from invertebrates to fish; Carol et al., 2009; Copp et al., 2009; Alp, 2017). The directed connectance (i.e., number of actual links over the number of possible links) of each invaded food web was calculated to provide an estimate of interaction variations among resident species and S. glanis of different body sizes (Supplementary Figure 1; Bersier et al., 2002). Three body sizes associated with the highest variations of the directed connectance were selected to simulate three invasion scenarios: small body size 40 cm (S40), medium body size 85 cm (S85) and large body size 150 cm (S150). The trophic interactions inferred for the corresponding three invaded food webs were included in the following steps of the analysis.
Population dynamic model
The simulations of population dynamics were based on an updated version of the allometric population dynamic model proposed by Yodzis and Innes (1992) and extended by Brose et al. (2006). The dynamics of each primary producer i are given by:
The parameters are the intrinsic growth rate ri and biotic capacity Ki. Both are allometrically scaled (Supplementary Table 2) differently from the ones proposed by Brose et al. The functional response (i.e., the per capita consumption rate of consumer j on resource i), Fji follows a Holling Type II and is given by:
The parameters are xj, the metabolic rate body mass dependent, and yj the maximum consumption rate relative to the metabolic rate of consumer j. The former is newly allometrically scaled, while the latter depends on characteristic of the species (Supplementary Table 2). ωji is the proportion of resource i in the diet of consumer j (eq. 1) obtained from the weighting procedure previously described. The handling time hjk of consumer j (i.e., time for handling and consuming the resource k) is also allometrically scaled (Supplementary Table 2), differently from the parameterization of Brose et al. for providing realistic estimations of this parameter. Finally, the dynamics of consumer j is given by:
The parameters are the morality rate mj (allometrically scaled) and the consumption efficiency εj. (depends on species characteristics; Supplementary Table 2). Note that we choose Ni and Nj as the SC abundances (number of individuals in the SC) and not as the biomass for more stable calculations. Therefore, all allometrically scaled parameters are per individual (see Supplementary Table 2).
Simulations
We used 200 simulations of 20,000 time steps for all scenarios (no. S. glanis, S40, S85, and S150) and final results were obtained by averaging simulation outputs at each time step for each SC. Variability in metabolic rates was included in calculations by generating white noise following 𝒩 (μ = 0, σ2 = 0.1% of metabolic rate). For each simulation, random initial abundances (N0) were assumed to be uniformly distributed (Brose et al., 2006) on the interval [0.15–1] and were ranked according to average body sizes of SC (i.e., SC with the largest body size had the lower N0; Peters and Wassenberg, 1983). N0 of S. glanis was fixed at 0.1 in all simulations to be lower than N0 of the resident apex predator SC (Esocidae), to avoid bias resulting from the initial abundance variations of S. glanis.
Quantitative impacts
The impacts of S. glanis on resident species were investigated through two main ecological processes that are SC extinctions and changes in non-extinct SC abundance. Extinctions were assumed to be effective when the abundance of a SC fell below 1.10–6 (Ovaskainen and Hanski, 2003) and extinct SC were not permitted to reintegrate the system. Extinctions were explored both qualitatively (i.e., SC taxonomic category) and quantitatively (i.e., number of extinctions and time lags between time at extinction with and without S. glanis).
The impacts on the abundances of non-extinct SC were investigated by calculating the relative abundance difference (RAD; Eq. 5) at each time step to describe increase/decrease in abundances with S. glanis compared to abundances without S. glanis similarly to Zhang et al. (2019).
The median RAD (MRAD) for the 200 simulations was used to quantify the extent (i.e., amplitude of changes) as well as the type (positive, MRAD > 0, or negative, MRAD < 0) of S. glanis impact on SC abundances (i.e., increase or decrease in abundances).
The model robustness was assessed by including an increasing variability in the metabolic rate using white noise (i.e., 𝒩 (μ = 0, σ2 = 0.1, 0.5, 1, 5, 10, and 20%; Supplementary Figures 3–9) using MRAD as the response variable.
Ecological interactions
Impacts of S. glanis on SC (extinct or with modified abundances) were classified according to three main ecological interactions based on the food web structure inferred with the aNM:
(i) Predation when a SC was a prey for S. glanis;
(ii) Competition when a SC shared common resources but did not directly interact with S. glanis. In this case, Schoener’s overlap index was complementarily calculated to quantify diet similarities between both competitors (Vera-Duarte and Landaeta, 2017) as follows:
where Pxi and Pyi are the proportions of resource i in the inferred diet of competitor x and y;
(iii) Trophic cascades were considered when a SC was not a prey of S. glanis and corresponded to repercussions across multiple trophic levels (Carpenter et al., 2010).
Interactions not corresponding to these three ecological interactions were classified as “Others.”
All statistical and graphical displays were performed using R.3.5.1 (R Core Team, 2018) with the packages ade4 (Dray and Dufour, 2007), deSolve (Soetaert et al., 2010), cheddar (Hudson et al., 2013), igraph (Csárdi, 2019), NetIndices (Kones et al., 2009), foodweb (Perdomo, 2015), ggplot2 (Wickham, 2016). R Markdown files with data and R codes summarizing the conducted analyses are available at https://github.com/chloevagnon/aNM-and-population-dynamics to provide reproducible examples.
Results
General impacts of silurus glanis within the whole food web
Extinctions did not relate to S. glanis body sizes as the same extinctions occurred for the three different invasion scenarios (Figure 1). Extinct species clusters (SC) corresponded to different trophic levels (from primary producers to secondary consumers) and were mainly characterized by a high pelagic reliance (Supplementary Figure 11). In contrast, S. glanis body size was determinant in the amplitude of abundance changes of the non-extinct SC (i.e., MRAD from –0.07 to +0.15). The strongest negative impacts were found for SC with TP (Trophic Position) close to the invader’s TP in the food webs while positive impacts mainly concerned SC with low TP (Figure 1). The lowest and the highest MRAD were observed for S85 and suggested positive impacts on primary littoral and pelagic consumers, and negative ones on SC with TP > 2.5 (Figure 1). Littoral secondary consumers were mainly impacted for S40 and the lowest MRAD were found for S150.
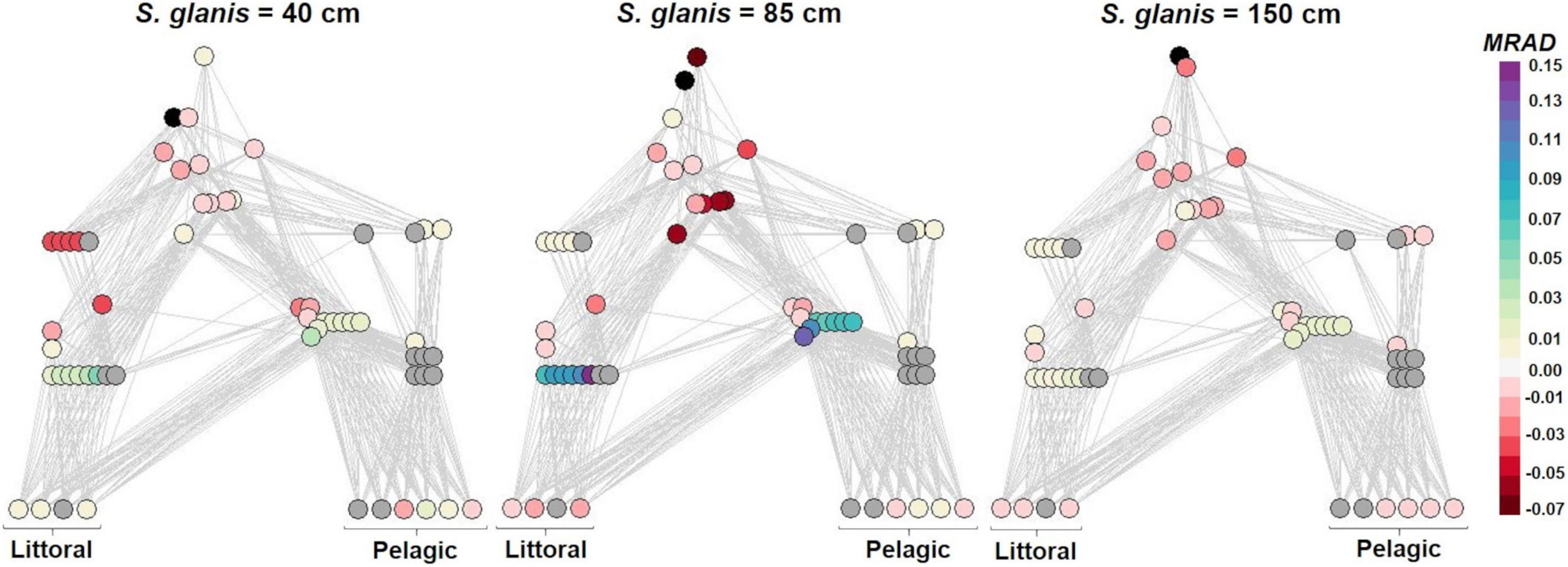
Figure 1. Effects of S. glanis throughout the whole invaded food-web typical of large French peri-Alpine lakes including independently S. glanis measuring 40, 85, and 150 cm. Nodes represent species clusters (SC) vertically arranged by increasing trophic position (TP) and colored according to the median value of the relative abundance differences (MRAD) used as an impact metric. Extinct species clusters are represented in dark gray and S. glanis is in black. The x-axis is defined as the gradient of pelagic reliance calculated from the initial pelagic reliance of primary producers set at 0 for littoral SC and 1 for the strict pelagic. The pelagic reliance of consumers was calculated by averaging the pelagic reliance of their resources considering ascending TP. When consumers shared the same TP and the same pelagic reliance, a delta of 0.02 was added to the pelagic reliance of the involved SC to avoid node overlapping in the plot.
Extinction patterns
Among the 58 SC, 14 were extinct at the end of the simulations in each scenario (Figure 2A). Three major extinction phases were noticeable and concerned in the first place zooplankton (up to 1,043.3 ± 0.5 time steps), followed by invertebrates and phytoplankton (up to 10,475.8 ± 4.3 time steps), and finally one invertebrate (at 17,915.5 ± 35.2 time steps). The same extinctions patterns occurred for all scenarios while time lags between extinctions differed, particularly for the latest extinctions (Figure 2B). Extinctions tented to occur earlier for S85 and S150 while those could appear later for S40 compared to extinctions without S. glanis.
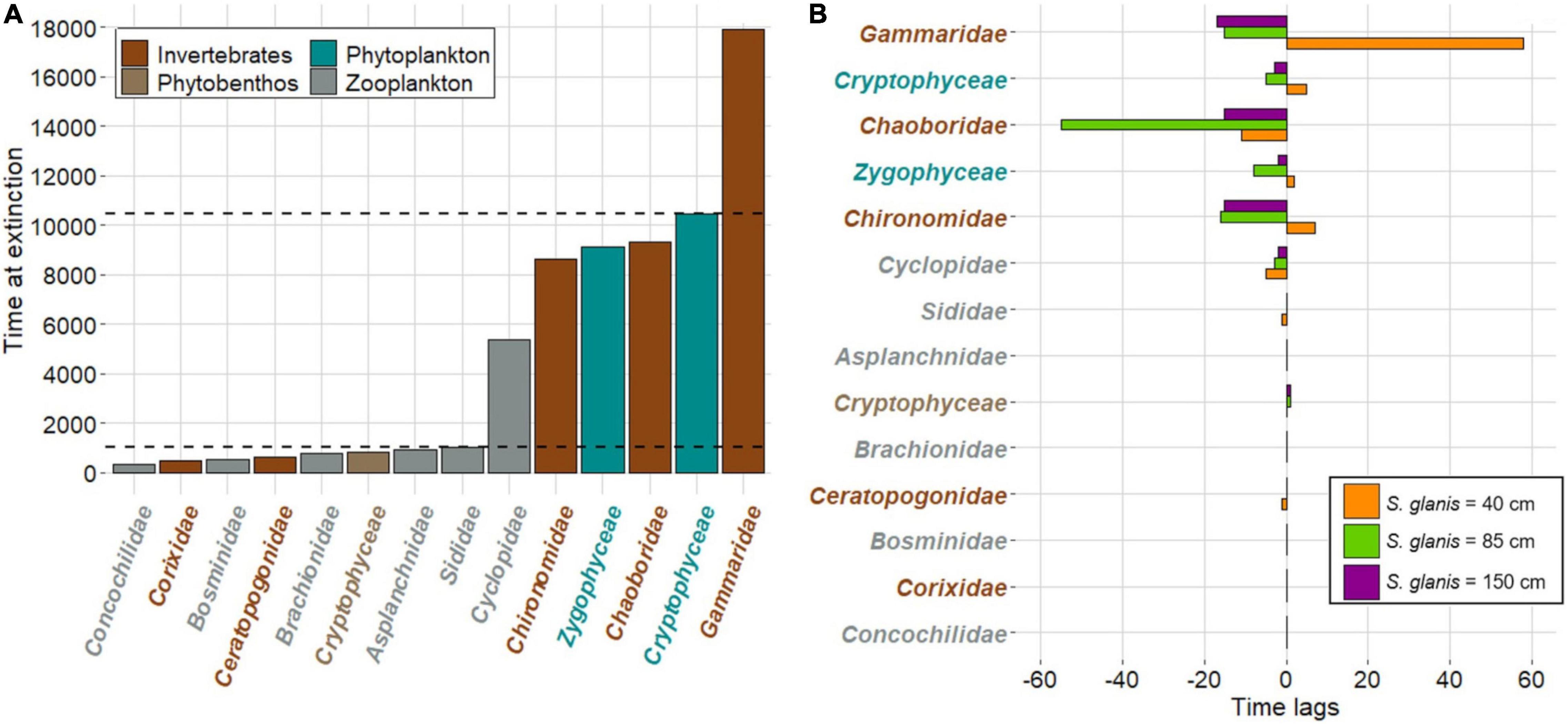
Figure 2. Extinctions of species clusters (SC) in the whole invaded food-web typical of large French peri-Alpine lakes including independently S. glanis measuring 40, 85, and 150 cm. (A) Time at extinction over the 20,000 time steps for the SC are represented (three major extinction phases are delineated by horizontal dashed lines) and correspond to all scenarios. (B) Time lags between time at extinction associated with S. glanis invasion are represented according to the different invasion scenarios. SC are ordered by decreasing time at extinction (from top to bottom). Negative and positive time lags indicate extinctions occurring earlier and latter than without S. glanis invasion, respectively.
Abundance changes
RAD (Relative Abundance Differences) varied among SC taxonomic categories and for the different S. glanis body sizes. Fish SC were impacted for all scenarios while presenting variable changes as shown by the boxplots representing RAD at each time step (Figure 3A). The three smallest fish SC and the largest one were especially negatively impacted by S85 (lowest RAD and highest variability), followed by S150 especially for the largest fish SC. In contrast, negative effects of S40 were more pronounced for medium-size fish SC and even appeared positive for two fish SC.
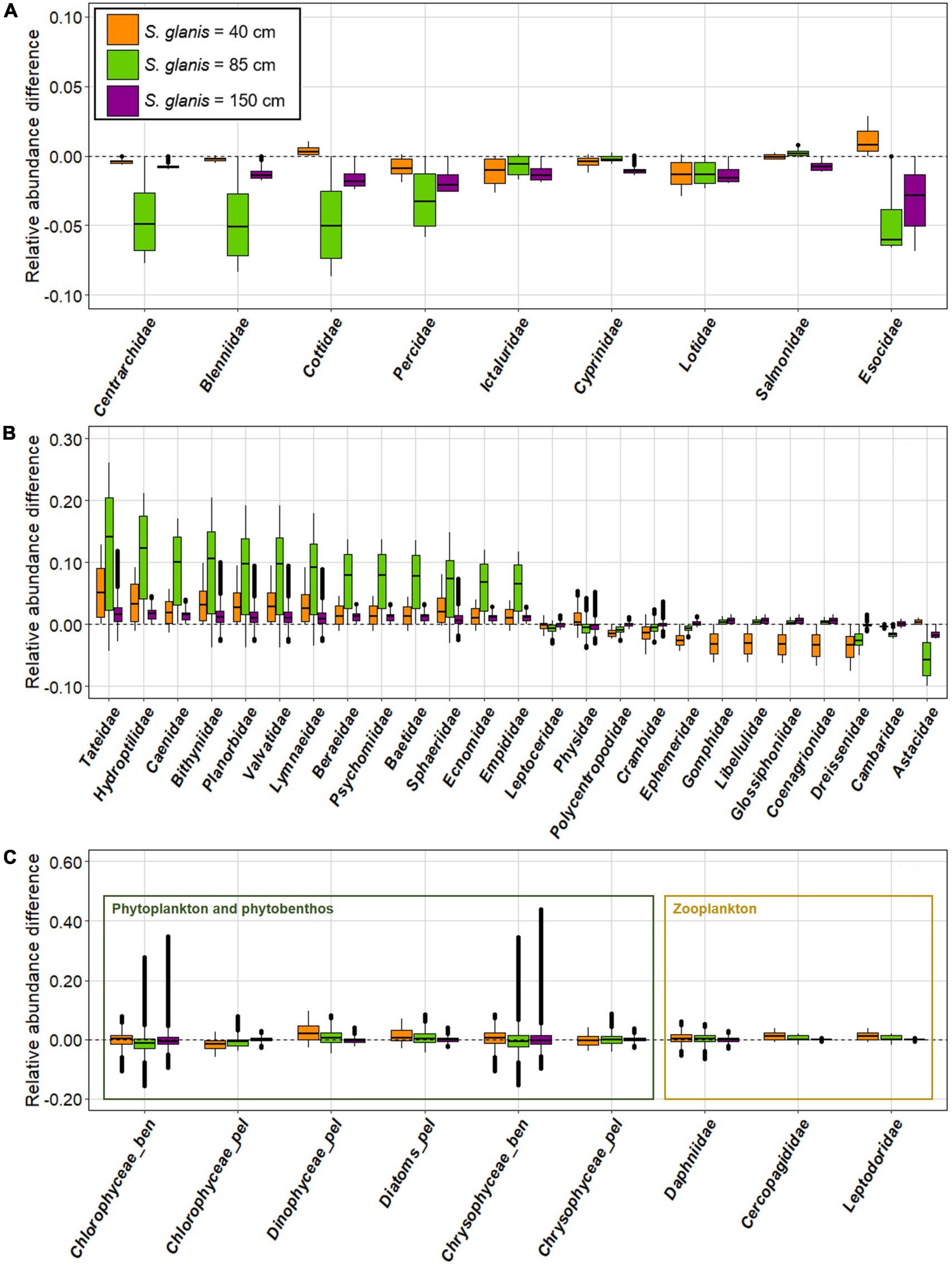
Figure 3. Relative abundance differences (RAD) along the 20,000 time steps for each invasion scenario providing the effect of S. glanis on the species clusters of fish (A), invertebrate (B) and primary producers and zooplankton (C) in the whole invaded food-web typical of large French peri-Alpine lakes. Species clusters are ordered by increasing body sizes from left to right. Boxes comprise 25–75% quantiles and horizontal full lines indicate median values. Median values below 0 correspond to an overall abundance depletion following S. glanis invasion while median values above 0 correspond to an abundance increase.
Half of invertebrate SC had higher abundances in presence of all S. glanis and presented a high variability along time steps, particularly with S85 (i.e., highest RAD; Figure 3B). The other half of invertebrate SC was negatively impacted by S40 for all time steps (i.e., all RAD values < 0) and the three largest invertebrates were negatively affected by S85 and S150.
RAD for phytoplankton, phytobenthos and zooplankton SC were similar among scenarios and more variable in time for phytobenthos SC (Figure 3C). Abundances of zooplankton and phytobenthos SC slightly increased for S40 while they decreased in the scenarios with the two larger S. glanis (S85 and S150). Abundances of half of phytoplankton SC increased with S. glanis body size while the reverse was found for the other half of phytoplankton SC.
Ecological interactions
Extinctions may have been caused by trophic cascades regarding the low trophic position of extinct SC compared to that of the three S. glanis, even for one invertebrate SC predated by S40 (Figure 4A and Supplementary Table 3), but did not seem directly triggered by S. glanis as they also occurred without the predator in the food web.
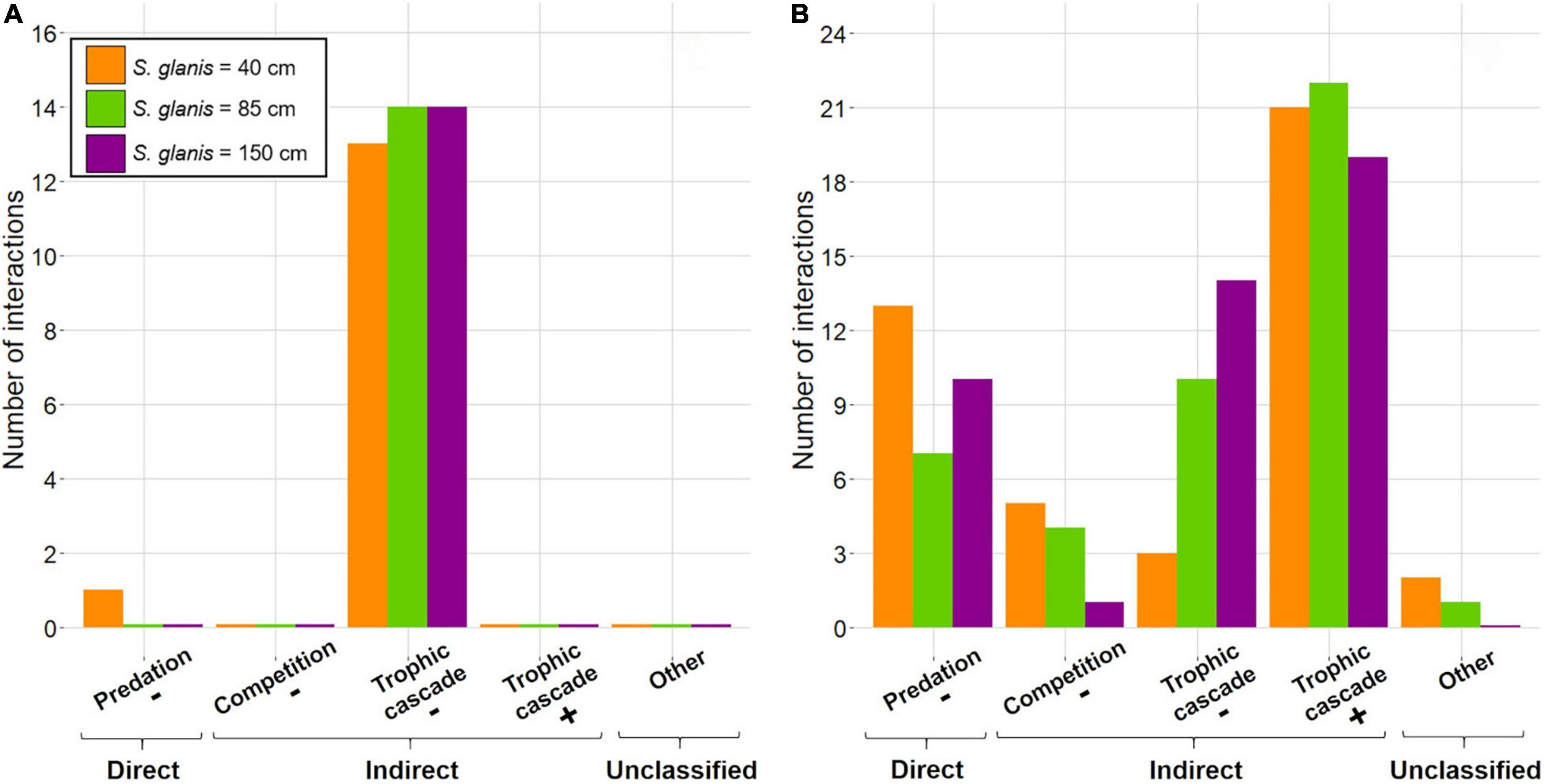
Figure 4. Ecological interactions involved in time lags for extinct species clusters (A) and for species clusters submitted to abundances changes (B) in the whole invaded food-web typical of large French peri-Alpine lakes. The “–” corresponds to interactions with negative impacts on species cluster abundances (i.e., MRAD < 0). Contrary, “+” is used for interactions with positive impacts on species clusters abundances (i.e., MRAD > 0).
Ecological interactions for non-extinct SC were much more diverse among scenarios and corresponded to both negative and positive impacts mediated by direct and indirect interactions, even for three interactions with fish SC for S40 and S85 (Figure 4B and Supplementary Table 3).
Direct negative impacts corresponded to predation with 13 SC for S40 (91% of invertebrate SC), seven SC for S85 (large invertebrates and medium-size fish SC) and 10 SC for S150 (crayfish and all fish SC).
Indirect negative impacts were emphasized through competition identified for different fish SC when considering the invasion of S40 (e.g., Lotidae, Cyprinidae, Ictaluridae; Schoener’s index of 0.78, 0.54 and 0.53, respectively), S85 (e.g., Esocidae; Schoener’s index = 0.91) and S150 (Esocidae; Schoener’s index = 0.71). Negative impacts were also due to trophic cascades, concerning three SC for S40 and increasing with S. glanis body size (i.e., 10 SC for S85 and 14 SC for S150).
Among the different ecological interactions, Indirect positive interactions were preponderant with 21 trophic cascades identified for S40 (composed at 67% of invertebrate SC, 5% of zooplankton and 28% of primary producer SC), 22 trophic cascades for S85 (composed of 82% of invertebrate SC, 9% of zooplankton and 9% of primary producer SC) and 19 trophic cascades for S150 (100 % of invertebrate SC).
Sensitivity analysis
The sensitivity analysis revealed a gradual increase in MRAD variability in response to the increasing metabolic noise. However, MRAD distribution remained consistent among scenarios and SC categories for metabolic noise up to 5% and then started to flatten, traducing a decrease in the consistency of the results with higher metabolic noises (Supplementary Figures 3–9). Especially fish SC seemed more sensitive than other SC to metabolic noise. Therefore, our simulations could be sensitive to high metabolic rate variability while patterns in responses remained robust according to taxonomic categories and among the invasion scenarios.
Discussion
Our approach combines trophic interaction inferences from the aNM with a dynamic population model allowing a thorough investigation of an invasive species impacts at the whole lake food web scale. It supported the consideration of direct and indirect interactions between residents and the invader and thus investigation of both positive and negative impacts on their abundances.
A new predator in peri-alpine lake food webs
The food web topology was only slightly modified by the invasion of S. glanis, which exerted moderate impacts on connectance and abundance modifications (i.e., low connectance changes and restrained amplitude of abundance modifications). This result may be explained by the rather high resolution of the food web, as associated to omnivory that both promote weak interaction strengths (Supplementary Table 4) and stability in the persistence of communities (Emmerson and Yearsley, 2004; Dunne et al., 2005; Landi et al., 2018; Kawatsu et al., 2021). Ecological interactions and abundance dynamics were nevertheless influenced by S. glanis and were modulated according to its body size.
The highest negative impacts of S. glanis were related to direct interactions for resources acquisition. Predation was indeed the main ecological interaction associated with the decrease in SC abundances, in lines with numerous impact studies on invasive fish (Gozlan et al., 2010; Van der Veer and Nentwig, 2015; David et al., 2017). Specifically, the decrease in abundance of invertebrate SC (e.g., Ephemeridae) and small fish SC (e.g., Blenniidae) would be caused by S. glanis of 40 cm while the decrease of larger fish SC (e.g., Percidae) and large invertebrate SC (e.g., crayfish) would mostly result from the invasion of larger S. glanis (85 cm and 150 cm). Percidae and crayfish represented consistent fraction of S. glanis diet in empirical studies (Carol et al., 2009; Ferreira et al., 2019) and the presence of S. glanis was suspected to alter their abundance in few ecosystems (Copp et al., 2009; Guillerault et al., 2015; Vagnon et al., 2022), suggesting plausible predictions from our approach. Competition between S. glanis and large fish SC was also suggested to negatively alter fish abundances and particularly dampened the abundance of the resident apex predator (Esocidae; high diet overlaps for S85 and S150) while fish SC with weak diet overlaps (Schoener index < 0.1) were poorly or not impacted. These results supported both plausible inferences of trophic interactions for predators and species abundance alterations due to both predation and competition. Indeed, invasive predators frequently cause decreases in abundances of their prey (Mills et al., 2004; David et al., 2017), they are expected to have a strong competitive effect in aquatic ecosystems and they are known to induce changes in species assemblages (Gozlan et al., 2010; Allesina and Tang, 2012; David et al., 2017).
Interestingly our results highlighted that considering two trophic levels (i.e., consumer/resource) or two competitors does not allow to identify major impacts of invasive species and their repercussions at the whole food web scale as we underlined main abundance changes for SC through trophic cascades, and particularly we found higher positive impacts than negative ones. A succession of negative and positive impacts was indeed noticeable along the food web and emphasized typical patterns of top-down cascades found in aquatic ecosystems (Carpenter et al., 2010; Heath et al., 2014; Su et al., 2021). In these processes, the negative impacts on abundances of SC such as fish result in a relaxed predation on lower trophic levels (e.g., grazers) and thus in a reduction of primary producers (Carpenter et al., 2008; Heath et al., 2014; Koning and McIntyre, 2021). Consequently, positive and negative impacts were higher for S85 than for S40 and S150, as fish and large invertebrate SC were particularly impacted in this invasion scenario (i.e., competition with fish SC and predation on both fish and large invertebrate SC). Complementarily, trophic cascades were suggested to modulate time at extinctions (earlier for S85 and S150 and later for S40), yet the presence of S. glanis did not qualitatively influence extinction processes (i.e., same extinctions with/without S. glanis). These results underline the ability of S. glanis to cause top-down trophic cascades by regulating mesopredator abundances while suggesting that it may not be a major source of species extinctions, similarly to empirical studies conducted in reservoirs, lakes and rivers (Copp et al., 2009; Vejřík et al., 2017). In fact, the opportunistic feeding behavior of S. glanis (Copp et al., 2009; Cucherousset et al., 2018; Vagnon et al., 2022) could foster its reliance on a diversified prey set limiting strong interaction strengths usually known to induce stronger impacts on resident species populations than weak interactions (Terraube et al., 2011; Wootton and Stouffer, 2016).
Model limits and strengths
Our study based on the combination of an allometric niche model and an alternative version of the population dynamic model of Brose et al. (2006) succeeded at supporting a comprehensive assessment of resident species abundance modifications and the involved trophic interactions considering a multi-trophic system, often challenging to evaluate only based on experiments or on traditional empirical methods (Crystal-Ornelas and Lockwood, 2020), notwithstanding process simplifications inherent to the elaborated method and to our study objectives.
Firstly, we considered an average body size to represent species nodes in food webs and we integrated independently a unique S. glanis body size node in simulation scenarios. While we recognize that diets of both resident and S. glanis populations can be ontogenetic-dependent, we conserved this approach commonly approved in community ecology to preserve the real significance of each node in the food web (i.e., one S. glanis node = one invertebrate node = one phytoplankton node). Secondly, we assumed environmental and anthropic drivers constant to capture specifically the invasive species impacts in the recipient ecosystem. Although these factors are known to be dynamic in real systems (Brose and Hillebrand, 2016), our approach appeared relevant considering the different time scales involved in species abundances modifications following exposure to various external pressures. Indeed, the catfish impacts could be observed more rapidly than external factors such as climate change (i.e., decades vs. several decades). Our dynamic model could obviously be completed in future studies to account for possible other drivers of species trophic interactions and their dynamics (e.g., metabolism modifications following temperature increase due to climate change) but remained out of the scope of this study.
However, the combination of allometric models would be appropriate for an application to a broad aquatic ecosystem array where body size governs trophic interactions, mainly thanks to the minimal required data inputs often available from monitoring surveys or literature (i.e., species inventory and the average species body sizes). Here we used species clusters for convenience and for limiting computation concerns such as singularities but different taxonomic resolutions can be considered depending on the initial study scope. Moreover, allometric parameterizations in the dynamic model could also be replaced by empirical data (e.g., body mass, metabolic rate) and/or can be completed by other calculation methods, for instance, to infer interaction strengths between consumers and resources (Calizza et al., 2021). When these data are not available, our initial simulation parameterization appears valuable regarding the convergence between our results and literature on predator impacts in food webs and freshwater ecosystems (Cucherousset and Olden, 2011; Cucherousset et al., 2012; Jackson et al., 2017), although empirical validations of our simulations still remain out of reach. Indeed, management plans mostly focus on a part of species compared to the totality of resident species to survey, mainly due to the restricted resources available for management actions (Vander Zanden and Olden, 2008), thus limiting the consideration of all taxa responses at long term that could be underlined in our study.
Overall, our study framework addresses the impact of invasive species and may be relevant regarding the increasing rate of species introductions representing a major threat to ecosystems (Lockwood et al., 2013). We underlined the importance of considering interspecific interactions at the whole food web scale for the assessment of invasive species impacts. The introduction of invaders indeed frequently involves a wide diversity of new ecological interactions, resulting in modifications of species abundances through both direct and indirect interactions and thus causing non-negligible impacts on resident communities. The balance between negative and positive aspects of invasions is also a significant factor to consider as positive effects of invasions can appear non-negligible, and may attenuate a priori expectations (Gozlan, 2008; Tablado et al., 2010; Schlaepfer et al., 2011). Our study can thus participate to the growing corpus of methodologies trying to reach comprehensive assessments and predictions of invader impacts, considering their direct/indirect and positive/negative effects in freshwater ecosystems.
Data availability statement
The original contributions presented in this study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author contributions
CV, RPR, L-FB, FC, JG, and VF designed the study. CV led data analyses and manuscript writing with RPR and VF. All authors provided critical feedbacks.
Funding
This research was funded by the Region Auvergne-Rhône-Alpes (SILAC program).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We are especially grateful for the time spent at the Department of Biology of the University of Fribourg (Switzerland) and the associated fruitful discussions with members of the labs. We thank both two reviewers for their advised comments that improved early draft of the manuscript. We also wanted to thank Vincent Miele (LBBE, Laboratoire de Biométrie et Biologie Evolutive) for discussions during the manuscript elaboration and the Observatory of LAkes OLA for providing full access to lake monitoring data from monitoring surveys financially supported by the CISALB (Comité Intercommunautaire pour l’Assainissement du Lac du Bourget), SILA (Syndicat Mixte du Lac d’Annecy) and CIPEL (Commission internationale pour la protection des eaux du Léman).
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.913954/full#supplementary-material
References
Albertson, L. K., MacDonald, M. J., Tumolo, B. B., Briggs, M. A., Maguire, Z., Quinn, S., et al. (2021). Uncovering patterns of freshwater positive interactions using meta-analysis: identifying the roles of common participants, invasive species and environmental context. Ecol. Lett. 24, 594–607. doi: 10.1111/ele.13664
Alexander, M. E., Dick, J. T. A., and O’Connor, N. E. (2013). Trait-mediated indirect interactions in a marine intertidal system as quantifi ed by functional responses. Oikos 122, 1521–1531.
Allesina, S., and Pascual, M. (2009). Food web models: a plea for groups. Ecol. Lett. 12, 652–662. doi: 10.1111/j.1461-0248.2009.01321.x
Alp, A. (2017). Diet shift and prey selection of the native European catfish, Silurus glanis, in a Turkish reservoir. J. Limnol. Freshw. Fish. Res. 3, 15–23. doi: 10.17216/limnofish.288217
Barrios-O’Neill, D., Dick, J. T., Emmerson, M. C., Ricciardi, A., MacIsaac, H. J., Alexander, M. E., et al. (2014). Fortune favours the bold: a higher predator reduces the impact of a native but not an invasive intermediate predator. J. Anim. Ecol. 83, 693–701. doi: 10.1111/1365-2656.12155
Bersier, L. F., Banasek-Richter, C., and Cattin, M. C. (2002). Quantitative descriptors of food-web matrices. Ecology 83, 2394–2407.
Brose, U., and Hillebrand, H. (2016). Biodiversity and ecosystem functioning in dynamic landscapes. Philos. Trans. R. Soc. B 371:20150267.
Brose, U., Williams, R. J., and Martinez, N. D. (2006). Allometric scaling enhances stability in complex food webs. Ecol. Lett. 9, 1228–1236. doi: 10.1111/j.1461-0248.2006.00978.x
Brown, J. H., Gillooly, J. F., Allen, A. P., Savage, V. M., and West, G. B. (2004). Toward a metabolic theory of ecology. Ecology 85, 1771–1789.
Brucet, S., Tavs̨anoğlu, ÜN., Özen, A., Levi, E. E., Bezirci, G., Çakıroğlu, A. İ, et al. (2017). Size-based interactions across trophic levels in food webs of shallow Mediterranean lakes. Freshw. Biol. 62, 1819–1830. doi: 10.1111/fwb.12997
Calizza, E., Rossi, L., Careddu, G., Sporta Caputi, S., and Costantini, M. L. (2021). A novel approach to quantifying trophic interaction strengths and impact of invasive species in food webs. Biol. Invasions 23, 2093–2107. doi: 10.1007/s10530-021-02490-y
Carol, J., Benejam, L., Benito, J., and García-Berthou, E. (2009). Growth and diet of European catfish (Silurus glanis) in early and late invasion stages. Fundam. Appl. Limnol. 174, 317–328. doi: 10.1127/1863-9135/2009/0174-0317
Carpenter, S., Brock, W., Cole, J., Kitchell, J., and Pace, M. (2008). Leading indicators of trophic cascades. Ecol. Lett. 11, 128–138.
Carpenter, S., Cole, J., Kitchell, J., and Pace, M. (2010). “Chapter 4. Trophic Cascades in Lakes: Lessons and Prospects,” in Trophic Cascades: Predators, Prey, and the Changing Dynamics of Nature, eds J. Terborgh and J. A. Este (Washington D.C: Island Press).
Copp, G. H., Robert Britton, J., Cucherousset, J., García-Berthou, E., Kirk, R., Peeler, E., et al. (2009). Voracious invader or benign feline? A review of the environmental biology of European catfish Silurus glanis in its native and introduced ranges. Fish Fish. 10, 252–282. doi: 10.1111/j.1467-2979.2008.00321.x
Crystal-Ornelas, R., and Lockwood, J. L. (2020). The ‘known unknowns’ of invasive species impact measurement. Biol. Invasions 22, 1513–1525. doi: 10.1007/s10530-020-02200-0
Cucherousset, J., and Olden, J. (2011). Ecological Impacts of Non-native Freshwater Fishes. Fisheries 36, 215–230.
Cucherousset, J., Bouletreau, S., Martino, A., Roussel, J. M., and Santoul, F. (2012). Using stable isotope analyses to determine the ecological effects of non-native fishes. Fish. Manag. Ecol. 19, 111–119. doi: 10.1111/j.1365-2400.2011.00824.x
Cucherousset, J., Horky, P., Slavík, O., Ovidio, M., Arlinghaus, R., Boulêtreau, S., et al. (2018). Ecology, behaviour and management of the European catfish. Rev. Fish Biol. Fish. 28, 177–190. doi: 10.1007/s11160-017-9507-9
Cuthbert, R. N., Dalu, T., Wasserman, R. J., Dick, J. T. A., Mofu, L., Callaghan, A., et al. (2018). Intermediate predator naivete and sex-skewed vulnerability predict the impact of an invasive higher predator. Sci. Rep. 8:14282. doi: 10.1038/s41598-018-32728-0
Cuthbert, R. N., Dickey, J. W. E., Coughlan, N. E., Joyce, P. W. S., and Dick, J. T. A. (2019). The Functional Response Ratio (FRR): advancing comparative metrics for predicting the ecological impacts of invasive alien species. Biol. Invasions 21, 2543–2547. doi: 10.1007/s10530-019-02002-z
David, P., Thébault, E., Anneville, O., Duyck, P. F., Chapuis, E., and Loeuille, N. (2017). “Impacts of Invasive Species on Food Webs,” in Networks of Invasion: A Synthesis of Concepts, eds D. Bohan, A. Dumbrell, and F. Massol (Cambridge: Academic Press), 1–60.
Diagne, C., Leroy, B., Gozlan, R. E., Vaissiere, A. C., Assailly, C., Nuninger, L., et al. (2020). InvaCost, a public database of the economic costs of biological invasions worldwide. Sci. Data 7:277. doi: 10.1038/s41597-020-00586-z
Dick, J. T. A., Alexander, M. E., Jeschke, J. M., Ricciardi, A., MacIsaac, H. J., Robinson, T. B., et al. (2014). Advancing impact prediction and hypothesis testing in invasion ecology using a comparative functional response approach. Biol. Invasions 16, 735–753. doi: 10.1007/s10530-013-0550-8
Dick, J. T. A., Laverty, C., Lennon, J. J., Barrios-O’Neill, D., Mensink, P. J., Robert Britton, J., et al. (2017). Invader Relative Impact Potential: a new metric to understand and predict the ecological impacts of existing, emerging and future invasive alien species. J. Appl. Ecol. 54, 1259–1267. doi: 10.1111/1365-2664.12849
Dickey, J. W. E., Cuthbert, R. N., Rea, M., Laverty, C., Crane, K., South, J., et al. (2018). Assessing the relative potential ecological impacts and invasion risks of emerging and future invasive alien species. NeoBiota 40, 1–24. doi: 10.3897/neobiota.40.28519
Dray, S., and Dufour, A. B. (2007). The ade4 package: implementing the duality diagram for ecologists. J. Stat. Softw. 22, 1–20.
Dunne, J. A., Brose, U., Williams, R. J., and Martinez, N. D. (2005). “Modeling food-web dynamics: complexity–stability implications,” in Aquatic Food Webs: An Ecosystem Approach, eds A. Belgrano, U. M. Scharler, J. Dunne, and R. E. Ulanowicz (Oxford: Oxford Univeristy Press), 117–129.
Emmerson, M., and Yearsley, J. M. (2004). Weak interactions, omnivory and emergent food-web properties. Proc. R. Soc. B: Biol. Sci. 271, 397–405. doi: 10.1098/rspb.2003.2592
Ferreira, M., Gago, J., and Ribeiro, F. (2019). Diet of European catfish in a newly invaded region. Fishes 4:58. doi: 10.3390/fishes4040058
Frost, C. M., Allen, W. J., Courchamp, F., Jeschke, J. M., Saul, W. C., and Wardle, D. A. (2019). Using Network Theory to Understand and Predict Biological Invasions. Trends Ecol. Evol. 34, 831–843. doi: 10.1016/j.tree.2019.04.012
Geary, W. L., Bode, M., Doherty, T. S., Fulton, E. A., Nimmo, D. G., Tulloch, A. I. T., et al. (2020). A guide to ecosystem models and their environmental applications. Nat. Ecol. Evol. 4, 1459–1471. doi: 10.1038/s41559-020-01298-8
Gozlan, R. E. (2008). Introduction of non-native freshwater fish: is it all bad? Fish Fish. 9, 106–115. doi: 10.1111/j.1467-2979.2007.00267.x
Gozlan, R. E., Britton, J. R., Cowx, I., and Copp, G. H. (2010). Current knowledge on non-native freshwater fish introductions. J. Fish Biol. 76, 751–786. doi: 10.1111/j.1095-8649.2010.02566.x
Gravel, D., Poisot, T., Albouy, C., Velez, L., Mouillot, D., and Freckleton, R. (2013). Inferring food web structure from predator-prey body size relationships. Methods Ecol. Evol. 4, 1083–1090. doi: 10.1111/2041-210x.12103
Guillerault, N., Delmotte, S., Boulêtreau, S., Lauzeral, C., Poulet, N., and Santoul, F. (2015). Does the non-native European catfish Silurus glanis threaten French river fish populations? Freshw. Biol. 60, 922–928. doi: 10.1111/fwb.12545
Heath, M. R., Speirs, D. C., and Steele, J. H. (2014). Understanding patterns and processes in models of trophic cascades. Ecol. Lett. 17, 101–114.
Hudson, L. N., Emerson, R., Jenkins, G. B., Layer, K., Ledger, M. E., Pichler, D. E., et al. (2013). Cheddar: analysis and visualisation of ecological communities in R. Methods Ecol. Evol. 4, 99–104. doi: 10.1111/2041-210x.12005
Jackson, M. C., Wasserman, R. J., Grey, J., Ricciardi, A., Dick, J. T. A., and Alexander, M. E. (2017). “Novel and Disrupted Trophic Links Following Invasion in Freshwater Ecosystems,” in Networks of Invasion: Empirical Evidence and Case Studies, eds D. Bohan, A. Dumbrell, and F. Massol (Cambridge: Academic Press), 55–97.
Jacquet, S., Domaizon, I., and Anneville, O. (2014). The need for ecological monitoring of freshwaters in a changing world: a case study of Lakes Annecy, Bourget, and Geneva. Environ. Monit. Assess. 186, 3455–3476. doi: 10.1007/s10661-014-3630-z
Kamenova, S., Bartley, T. J., Bohan, D. A., Boutain, J. R., Colautti, R. I., Domaizon, I., et al. (2017). “Invasions Toolkit : Current methods for tracking the spread and impact of invasive species,” in Networks of Invasion: A Synthesis of Concepts, eds D. Bohan, A. Dumbrell, F. Massol, D. Bohan, A. Dumbrell, and F. Massol (Cambridge: Academic Press), 85–182.
Kawatsu, K., Ushio, M., van Veen, F. J. F., and Kondoh, M. (2021). Are networks of trophic interactions sufficient for understanding the dynamics of multi-trophic communities? Analysis of a tri-trophic insect food-web time-series. Ecol. Lett. 24, 543–552. doi: 10.1111/ele.13672
Kones, J. K., Soetaert, K., van Oevelen, D., and Owino, J. O. (2009). Are network indices robust indicators of food web functioning? A Monte Carlo approach. Ecol. Model. 220, 370–382. doi: 10.1016/j.ecolmodel.2008.10.012
Koning, A. A., and McIntyre, P. B. (2021). Grassroots reserves rescue a river food web from cascading impacts of overharvest. Front. Ecol. Environ. 19:152–158. doi: 10.1002/fee.2293
Landi, P., Minoarivelo, H. O., Brännström, Å, Hui, C., and Dieckmann, U. (2018). Complexity and stability of ecological networks: a review of the theory. Popul. Ecol. 60, 319–345. doi: 10.1007/s10144-018-0628-3
Levine, S. (1980). Several measures of trophic structure applicable to complex food webs. J. Theor. Biol. 83, 195–207. doi: 10.1007/s00442-014-2936-4
Lockwood, J. L., Hoopes, M. F., and Marchetti, M. P. (2013). Invasion Ecology (Second Edition). Hoboken: Wiley–Blackwell.
Mills, M. D., Rader, R. B., and Belk, M. C. (2004). Complex interactions between native and invasive fish: the simultaneous effects of multiple negative interactions. Oecologia 141, 713–721. doi: 10.1007/s00442-004-1695-z
Ovaskainen, O., and Hanski, I. (2003). Extinction threshold in metapopulation models. Ann. Zool. Fenn. 40, 81–97.
Perdomo, G. (2015). Package ‘Foodweb’. Version 1.0. Available Online at: https://CRAN.R-project.org/package=foodweb (accessed April 5, 2020).
Petchey, O. L., Beckerman, A. P., Riede, J. O., and Warren, P. H. (2008). Size, foraging, and food web structure. Proc. Natl. Acad. Sci. U.S.A. 105, 4191–4196.
Peters, R. H., and Wassenberg, K. (1983). The effect of body size on animal abundance. Oecologia 60, 89–96.
Pomeranz, J. P. F., Thompson, R. M., Poisot, T., Harding, J. S., and Lecomte, N. (2019). Inferring predator–prey interactions in food webs. Methods Ecol. Evol. 10, 356–367. doi: 10.1111/2041-210x.13125
R Core Team (2018). R: A Language And Environment For Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rimet, F., Anneville, O., Barbet, D., Chardon, C., Crépin, L., Domaizon, I., et al. (2020). The Observatory on LAkes (OLA) database: sixty years of environmental data accessible to the public. J. Limnol. 79, 164–178. doi: 10.4081/jlimnol.2020.1944
Schlaepfer, M., Sax, D., and Olden, J. (2011). The Potential Conservation Value of Non-Native Species. Conserv. Biol. 25, 428–437.
Shurin, J. B., Borer, E. T., Seabloom, E. W., Anderson, K., Blanchette, C. A., Broitman, B., et al. (2002). A cross-ecosystem comparison of the strength of trophic cascades. Ecol. Lett. 5, 785–791. doi: 10.1046/j.1461-0248.2002.00381.x
Soetaert, K., Petzoldt, T., and Setzer, W. R. (2010). Solving Differential Equations in R: package deSolve. J. Stat. Softw. 33, 1–25.
Su, H., Feng, Y., Chen, J., Chen, J., Ma, S., Fang, J., et al. (2021). Determinants of trophic cascade strength in freshwater ecosystems: a global analysis. Ecology 102:e03370. doi: 10.1002/ecy.3370
Tablado, Z., Tella, J. L., Sanchez-Zapata, J. A., and Hiraldo, F. (2010). The paradox of the long-term positive effects of a North American crayfish on a European community of predators. Conserv. Biol. 24, 1230–1238. doi: 10.1111/j.1523-1739.2010.01483.x
Terraube, J., Arroyo, B., Madders, M., and Mougeot, F. (2011). Diet specialisation and foraging efficiency under fluctuating vole abundance: a comparison between generalist and specialist avian predators. Oikos 120, 234–244.
Terry, J. C. D., Morris, R. J., and Bonsall, M. B. (2018). Trophic Interaction Modifications Disrupt The Structure and Stability of Food Webs. bioRxiv [Preprint] doi: 10.1101/345280
Vagnon, C., Bazin, S., Cattanéo, F., Goulon, C., Guillard, J., and Frossard, V. (2022). The opportunistic trophic behaviour of the European catfish (Silurus glanis) in a recently colonised large peri-alpine lake. Ecol. Freshw. Fish doi: 10.1111/eff.12659
Vagnon, C., Cattanéo, F., Goulon, C., Grimardias, D., Guillard, J., and Frossard, V. (2021). An allometric niche model for species interactions in temperate freshwater ecosystems. Ecosphere 12:e03420.
Van der Veer, G., and Nentwig, W. (2015). Environmental and economic impact assessment of alien and invasive fish species in Europe using the generic impact scoring system. Ecol. Freshw. Fish 24, 646–656. doi: 10.1111/eff.12181
Vander Zanden, M. J., and Olden, J. D. (2008). A management framework for preventing the secondary spread of aquatic invasive species. Can. J. Fish. Aquat. Sci. 65, 1512–1522.
Vejřík, L., Vejříková, I., Blabolil, P., Eloranta, A. P., Kočvara, L., Peterka, J., et al. (2017). European catfish (Silurus glanis) as a freshwater apex predator drives ecosystem via its diet adaptability. Sci. Rep. 7:15970. doi: 10.1038/s41598-017-16169-9
Vera-Duarte, J. A., and Landaeta, M. F. (2017). Feeding habits and dietary overlap during the larval development of two sandperches (Pisces: pinguipedidae). Sci. Mar. 81, 195–204. doi: 10.3989/scimar.04544.06A
Williams, R. J., Anandanadesan, A., and Purves, D. (2010). The probabilistic niche model reveals the niche structure and role of body size in a complex food web. PLoS One 5:e12092. doi: 10.1371/journal.pone.0012092
Williams, R. J., and Martinez, N. D. (2000). Simple rules yield complex food webs. Nature 404, 180–183. doi: 10.1038/35004572
Wootton, K. L., and Stouffer, D. B. (2016). Many weak interactions and few strong; food-web feasibility depends on the combination of the strength of species’ interactions and their correct arrangement. Theor. Ecol. 9, 185–195. doi: 10.1007/s12080-015-0279-3
Yodzis, P., and Innes, S. (1992). Body Size and Consumer-Resource Dynamics. .Am. Nat. 139, 1151–1175.
Zhang, H., Rutherford, E. S., Mason, D. M., Wittmann, M. E., Lodge, D. M., Zhu, X., et al. (2019). Modeling potential impacts of three benthic invasive species on the Lake Erie food web. Biol. Invasions 21, 1697–1719. doi: 10.1007/s10530-019-01929-7
Keywords: biological invasions, trophic interactions, trophic cascade, impact assessment, Silurus glanis
Citation: Vagnon C, Rohr RP, Bersier L-F, Cattanéo F, Guillard J and Frossard V (2022) Combining food web theory and population dynamics to assess the impact of invasive species. Front. Ecol. Evol. 10:913954. doi: 10.3389/fevo.2022.913954
Received: 06 April 2022; Accepted: 29 June 2022;
Published: 15 July 2022.
Edited by:
Lawrence Hurd, Washington and Lee University, United StatesReviewed by:
Franco Leandro de Souza, Federal University of Mato Grosso do Sul, BrazilQuan-Xing Liu, East China Normal University, China
Copyright © 2022 Vagnon, Rohr, Bersier, Cattanéo, Guillard and Frossard. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chloé Vagnon, Y2hsb2UudmFnbm9uQGdtYWlsLmNvbQ==