 Menghan Qiu
Menghan Qiu Haoran Li
Haoran Li Minxia Lu1
Minxia Lu1 Ruo Li
Ruo Li Lele Ren
Lele Ren
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 13 June 2022
Sec. Paleoecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.908131
This article is part of the Research Topic Effects of Novel Environments on Domesticated Species View all 19 articles
Animal husbandry has been an indispensable part of human subsistence since the origin of agriculture. Along with the prehistoric cultural exchange, several kinds of major domestic animals diffused and gradually got popularized across the Eurasia. The specific geographic setting makes the Hexi Corridor in northwestern China one of the key regions to converge various types of major crops and livestock, and to witness the deep impact of novel species on local subsistence. Archeological evidence reveals an emergence of early oriental-occidental cultural communication at the opening of the local Bronze Age in Hexi Corridor, resulting in a significant shift of local subsistence. However, due to the lack of more detailed archeological evidence, the initial timing and trajectory of the transformation of livestock feeding patterns remain unclear. In this study, we reported systematic stable isotope and precise radiocarbon dating analyses on faunal remains unearthed from Huoshiliang and Ganggangwa, two Early Bronze Age settlements (ca. 4,000–3,700 BP) in middle Hexi Corridor. Our results show distinct diversification in livestock feeding patterns at ∼3,850 cal BP; in contrast with previous periods, some omnivorous livestock appear to have consumed mainly C3 foodstuff and some herbivorous livestock primarily consumed C4 plants. Combined with published stable isotope data and other archeological findings in the neighboring region, a clearer trajectory of the evolution of livestock feeding patterns has been revealed with diversified strategy amid the transformation during the Early Bronze Age in Hexi Corridor. We argued that the alteration of the local livestock feeding pattern reflects the attempt to achieve more efficient economy and sustainable society, in order to withstand the harsh arid environment in Hexi Corridor.
The origin and spread of major crops and livestock markedly changed the pattern of human subsistence during prehistoric times, which generated deep impacts on the development of human society and the interaction between the environment and humans (Diamond and Bellwood, 2003; Liu et al., 2014; Spengler et al., 2017; Revelles et al., 2018; Dong et al., 2021a). During the Early Holocene (∼11,700–8,200 BP, BP for year before present), various types of crops and livestock were domesticated in West Asia (especially wheat, barley, cattle, and sheep/goat; Bradley et al., 1998; Zeder and Hesse, 2000; Zeder, 2008; Riehl et al., 2013) and East Asia (especially foxtail millet, broomcorn millet, rice, pig, and dog; Fuller et al., 2009; Jones and Liu, 2009; Lu et al., 2009; Zhao, 2011; Yuan, 2015; Ren and Dong, 2016), respectively. These botanic and zoic agricultural elements then diffused along with the early phase of trans-continental cultural exchange in the following several millennia (Sherratt, 2006; Anthony, 2010; Dong et al., 2017; Frachetti et al., 2017; Taylor et al., 2021). The spread and popularization of domestic livestock (Ren and Dong, 2016; Hermes et al., 2020; Honeychurch et al., 2021; Wilkin et al., 2021), together with the spread of major crops (also known as food globalization; Jones et al., 2011, 2016; Liu et al., 2019a), deeply transformed the human subsistence worldwide.
The introduction of novel crops and livestock, combined with heterogeneous natural environment across Eurasia, sculpted diverse patterns of human subsistence in different regions during the Neolithic and Bronze Ages. The analysis of stable carbon and nitrogen isotopes has been proved as an effective way to reconstruct paleodiet (van der Merwe and Vogel, 1978) and provide a unique perspective on human subsistence and related behaviors, and has therefore been extensively used in previous research of prehistoric Eurasia (e.g., Hu et al., 2008; Barton et al., 2009; Motuzaite Matuzeviciute et al., 2015; Chen et al., 2016; Ma et al., 2016; Miller et al., 2019; Li et al., 2020; Wilkin et al., 2020). In particular, human behavior related to animal husbandry practices has been frequently discussed via dietary perspective in recent years (e.g., Dai et al., 2016; Hermes et al., 2019; Ma et al., 2021; Vaiglova et al., 2021). These studies of paleodiet have offered valuable insights into human subsistence strategies in the past.
Acting as the passageway between the Yellow River Basin and the Arid Central Asia, the Hexi Corridor in northwestern China is regarded as a key region in cross-Eurasian cultural exchange (Dong et al., 2017; Yang, 2017), and has therefore received attention from an array of multidisciplinary researchers. The evolution and chronology of the prehistoric cultures in Hexi Corridor have been well established (Li, 2011; Yang, 2017; Yang et al., 2019a; Li et al., 2021). In addition, a significant body of Hexi Corridor research has focused on different aspects of the past relationship between humans and the environment (Sun et al., 2010; Li et al., 2011, 2017; Zhou et al., 2012; Yang et al., 2017, 2020; Zhang et al., 2017; Shen et al., 2018; Shi et al., 2019; Dong et al., 2021b). Furthermore, reconstruction of human subsistence has been finely reported via archaeobotanic (Zhou et al., 2016; Liu et al., 2019b,c, 2021; Shi et al., 2022), zooarchaeological (Flad et al., 2007; Song et al., 2016; Yang et al., 2019b), geochemical (Dodson et al., 2009), and genetic perspectives (Xiong et al., 2022).
For dietary perspective, abundant stable isotope data from human and faunal remains in prehistoric Hexi Corridor have been published (Atahan et al., 2011; Liu et al., 2014, 2016; Zhang et al., 2015; Yang et al., 2019b; Ma et al., 2021; Vaiglova et al., 2021). Stable isotope evidence reveals a significant shift in human diet from C4 millet crops to C3 wheat and barley in Hexi Corridor during 4,000–3,000 BP (Yang et al., 2019b). The initial timing of this shift could be traced back to 4,000–3,800 BP (Liu et al., 2014), right after the introduction of these Western crops at the beginning of the local Early Bonze Age (EBA; Dodson et al., 2013). Meanwhile, a similar pattern of dietary shift in domestic omnivores (pigs and dogs) has also been detected (Yang et al., 2019b). As the local domestic omnivore usually possess a C4 dietary habit and the novel herbivorous livestock possess a C3 dietary habit, the appearance of domestic individual showing opposite diet is abnormal and worth an extra attention. Recent studies with abundant stable isotope data from Hexi Corridor faunal remains have shed light on the practice of pastoralism and localized management strategy during EBA (Atahan et al., 2011; Yang et al., 2019b; Ma et al., 2021; Vaiglova et al., 2021). However, in this study, we sought to resolve several outstanding issues regarding livestock feeding patterns, namely, (1) the implications of individual livestock with abnormal δ13C-values and (2) the initial timing and trajectory for the shift in livestock feeding patterns during EBA in the Hexi Corridor. Based on materials acquired from archeological survey, a previous study on stable carbon and nitrogen isotopes from Huoshiliang and Ganggangwa sites has effectively contributed to the investigation of paleodiet and human subsistence during EBA in Hexi Corridor (Atahan et al., 2011). However, systematic excavation is still of necessity to provide a dynamic perspective to reveal the issues mentioned above.
This study reports systematic stable isotope analysis and precise radiocarbon dating results from faunal remains unearthed during the excavation of these two EBA archeological settlements. Carbon and nitrogen isotope values were acquired for a total of 56 specimens from a range of both livestock and wild animals from these two sites. In addition to 6 dates previously reported by Ren et al. (2022), a total of 15 direct radiocarbon dates were obtained to provide precise chronology in this study. We aimed to give clearer insight into the initial timing of the previously observed shift in livestock feeding patterns, as well as the implications of abnormal δ13C-values. By combining these newly acquired data with previously published studies, we were able to use faunal carbon isotope data to more precisely reconstruct the trajectory of the transformation of livestock feeding patterns and give novel insight into the transformation of local subsistence and animal management in the Hexi Corridor.
Being located in northwestern China, the Hexi Corridor is a passageway bordered by the Tibetan Plateau on the southwest and the Gobi Desert on the northeast, which connects the Yellow River Basin at Wushaoling Mountain on the east and the Arid Central Asia at Xingxingxia Gorge on the west (Figure 1). With modern mean annual precipitation lower than 200 mm, the dominant climate type in modern Hexi Corridor involves temperate semi-arid (BSk) and temperate arid (BWk) according to the Köppen classification (Chan et al., 2016). In addition, C3 plants make up the majority of modern flora of Hexi Corridor while a few taxa of C4 plants mainly distributed in the arid zone act as supplementary (Jiang et al., 2019). Rivers originating from the Qilian Mountains of northeastern Tibetan Plateau flow into Hexi Corridor, developing bunches of oases, making the region a relatively decent habitat and transitable passageway among the arid deserts in the communication between the East and the West, especially for the ancient Silk Road. However, restricted by the low precipitation and poor land productivity, this region has always suffered from the harsh natural environment when developing economy. The Heihe River forms its channel across the middle Hexi Corridor and streams toward the hinterland of Gobi Desert, raising a bunch of oases along its valley, which provide a possible avenue linking with the Eurasian Steppes (Jaang, 2015).
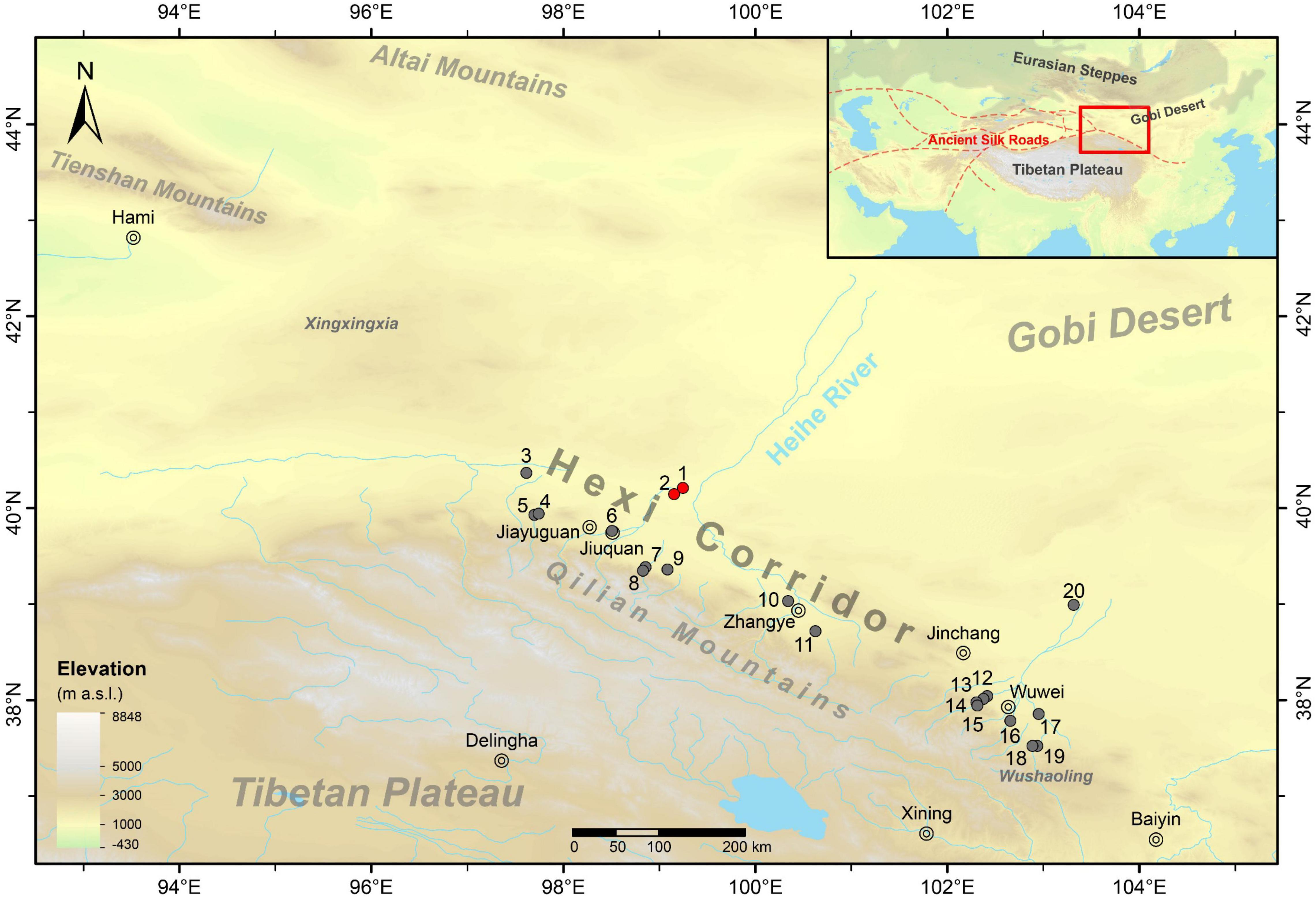
Figure 1. Regional map showing the geographic location of Hexi Corridor and its positional relation with neighboring geographic units. The prehistoric archeological sites mentioned in this paper are labeled in red (this study) and gray (previous studies) dots as (1) Huoshiliang; (2) Ganggangwa; (3) Gudongtan; (4) Huoshaogou; (5) Dadunwan; (6) Zhaojiashuimo; (7) Ganguya; (8) Sanbadongzi; (9) Xihetan; (10) Xichengyi; (11) Xihuishan; (12) Guojiashan; (13) Xinzhai; (14) Qipanshan; (15) Shichengshan; (16) Mozuizi; (17) Tuba; (18) Lijiageleng; (19) Shuikou; and (20) Minqinsanjiaocheng.
Huoshiliang (99.2446°E, 40.2116°N, 1,198 m a.s.l.) and Ganggangwa (99.1524°E, 40.1469°N, 1,203 m a.s.l.) are located in Jinta County, Jiuquan Region, northwestern Gansu Province (Figure 1). Situated at Jinta Basin in the middle reaches of Heihe River, these two archeological sites are identified as settlements and smelting locations of the EBA in middle Hexi Corridor (Figure 2). Standing on the intersection among the Yellow River Basin, Arid Central Asia, and Eurasian Steppes, Huoshiliang and Ganggangwa sites witnessed the emergence of prehistoric East-West cultural exchange and the opening of EBA in Hexi Corridor. In particular, the river valley of Heihe provides critical connection between Hexi Corridor and the Eurasian Steppes, making these sites pivotal stations during the prehistoric cultural exchange in northwestern China (Jaang, 2015). The geographic settings, landscapes, as well as previous archeological-environmental surveys and related studies of these two sites have been finely reported (Dodson et al., 2009; Sun et al., 2010; Atahan et al., 2011; Li et al., 2011; Zhou et al., 2016; Yang et al., 2017). As indicated by radiocarbon dating results, the age of these two sites spans from ca. 4,100 to 3,700 cal BP (Dodson et al., 2009, 2013; Atahan et al., 2011). The starting age of Ganggangwa site was reported to be slightly earlier than Huoshiliang site, reaching the transition from Late Machang Culture to Xichengyi Culture (Gansu Provincial Institute of Cultural Relics and Archaeology, 2020).

Figure 2. Images of the archeological sites and their cultural relics. (A) Aerial photo of Ganggangwa site. (B) Pottery sherds of Late Machang Culture unearthed from Ganggangwa site (T1911F2). (C) Pottery sherds of Xichengyi Culture unearthed from Ganggangwa site (T1911L3). (D) Aerial photo of Huoshiliang site. (E) Bronze slags unearthed from Huoshiliang site (T1007L3). (F) Pottery sherds of Xichengyi Culture unearthed from Huoshiliang site (T1007H33).
In consideration of achieving more comprehensive understanding on Huoshiliang and Ganggangwa sites, Gansu Provincial Institute of Archaeology and Cultural Relics launched an exploratory excavation in 2017. According to the preliminary archeological prospection, an area of 32 m2 in the core zone of each site was chosen for excavation by archeologists. During the excavation, plenty of pottery sherds, bronze slags, lithic tools, as well as animal bones, were unearthed from different archeological units, including house sites, ash pits, and burials. As indicated by the characters of typical pottery remains, the entirety of Huoshiliang and the majority of Ganggangwa belong to Xichengyi Culture of the local EBA (ca. 4,000–3,600 BP), while the early stage of Ganggangwa belongs to the Late Machang Culture of Late Neolithic Age in Hexi Corridor (ca. 4,100–4,000 BP) (Figure 2).
All unearthed animal bones were collected and cleaned for species identification. Dr. Lele Ren from Lanzhou University performed the species identification; the results were reported in another manuscript in detailed, which was submitted to the Journal of Archaeological Science and has currently been accepted (Ren et al., 2022). On this basis, we sampled bone specimens of different kinds of species from various archeological units. Bones with clearer morphological features were preferentially sampled for stable isotope analysis, which can contribute to lower the risk of misidentification of species. Even so, it is still impossible to 100% avoid such issue. Since Huoshiliang and Ganggangwa are neighboring, homogeneous and almost coetaneous to each other, data from these two sites were reported combinedly. A total of 62 pieces of animal bones from different individuals were sampled from Huoshiliang (n = 34) and Ganggangwa (n = 28) sites, including livestock (pig, n = 11; dog, n = 9; cattle, n = 6; and sheep/goat, n = 16) and wildlife (gazelle, n = 5; deer, n = 7; hare, n = 4; and bird, n = 4). Samples for radiocarbon dating were selected after the completion of stable isotope analysis such that livestock individuals with abnormal isotopic values could be targeted in addition to capturing the full stratigraphic range of the sites.
The method adopted to extract bone collagen in this study referred to Ma et al. (2016), which was modified from Richards and Hedges (1999). Bones were water washed and polished via a handheld electric grinder to remove the sediments adhered on the surface. Dense part on the bone was cut off as small fragment that weighed around 1 g for extraction. Then, the bone fragments were demineralized in 0.5 mol/L hydrochloric (HCl) solution at 5°C for more than 10 days, with refresh of the HCl solution every 1 or 2 days. When the samples were softened and stopped producing bubbles, they were washed with purified water and further reacted in 0.125 mol/L sodium hydroxide (NaOH) solution at 5°C for 20 h. After being neutralized again with purified water, the samples were placed in an acidic solution (HCl, pH = 3) at 75°C for 48 h to gelatinize, and were filtered afterward. For the final step, the samples were frozen in the refrigerator and freeze-dried in a lyophilizer, with concentrated solid collagen remained.
Rate of yield and atomic C/N ratio was utilized to examine the quality of bone collagen. With five samples experienced failure during the collagen extraction, the rest of the samples (n = 57) were weighed to calculate the rate of yield (Table 1). Content of carbon and nitrogen of all collagen samples was measured at the State Key Laboratory of Applied Organic Chemistry in Lanzhou University via an Elemental Analyzer (Vario EL Cube) to calculate atomic C/N ratios as described below.
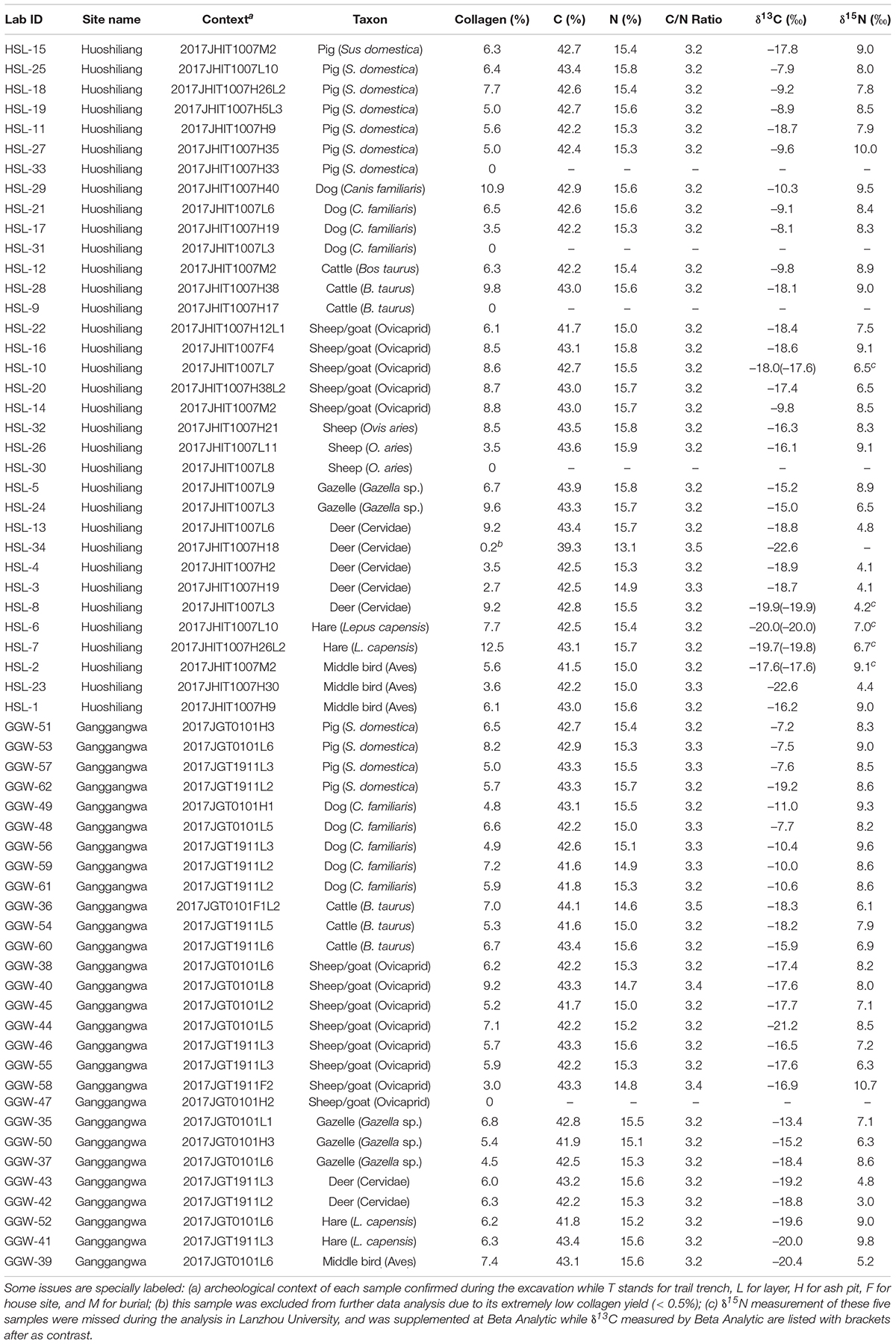
Table 1. Stable carbon and nitrogen isotope and quality indicator measurements of animal bone collagen from Huoshiliang and Ganggangwa sites.
For stable carbon isotope, δ13C-value varied significantly between C3 and C4 plants due to the different CO2 absorption during the photosynthesis. Among the major crops, wheat, barley, and rice are C3 plants while foxtail millet, broomcorn millet, corn, and sorghum are C4 plants. Such signal further transmits to animals and humans through the food chains, thus can be used as detector for diet and human subsistence (van der Merwe and Vogel, 1978), as well as the human behavior about animal husbandry practices (e.g., Dai et al., 2016; Hermes et al., 2019; Ma et al., 2021). Referring to previous study in Hexi Corridor (Yang et al., 2019b), δ13C-values of –18 and –11‰ are adopted as boundary lines for C3 (< –18‰), mixed C3/C4 (–18 to –11‰), and C4 diets (>–11‰) in this study. For stable nitrogen isotope, the proportion of 15N/14N enriches along the food chains, presenting distinct δ15N-values in different trophic levels (Lee-Thorp, 2008). However, the background value of δ15N could be raised by draught stress in arid zones, which results in notable interregional discrepancy (Ambrose and DeNiro, 1986; Schwarcz et al., 1999; Hartman, 2011).
Stable carbon and nitrogen isotope analyses were carried out at the Key Laboratory of Western China’s Environmental Systems (Ministry of Education) in Lanzhou University. Collagen samples were proceeded in an automated carbon and nitrogen analyzer linked to a Thermo Finnigan Flash DELTA-plus XL mass spectrometer. The ratios of carbon and nitrogen isotopes were calculated following the V-PDB and AIR standards using a two-point linear normalization, respectively. Graphite (IAEA code: USGS 24, δ13C = –16.0 ± 0.1‰) and caffeine (IAEA code: IAEA-600, δ15N = 1.0 ± 0.2‰) standards were used to calculate the results, with the precision of final data not exceeding ± 0.2‰. Full information is available in Table 1. Since the δ15N measurement of five samples (specially labeled in Table 1) was missed during the analysis, remedial analysis was carried out by the Beta Analytic Testing Laboratory with instruments of Costech ECS and Thermo Electron DeltaPlus IRMS; and the results were reported with the precision of ± 0.5‰. δ13C of these five samples were also measured as contrast to confirm that data from these two labs are comparable.
In total, 15 collagen samples (including 6 samples first reported in Ren et al., 2022) from various archeological units were selected for radiocarbon dating in different batches to construct the chronology of these two sites precisely. Samples were sent for accelerator mass spectrometry (AMS) radiocarbon dating at Key Laboratory of Western China’s Environmental Systems (Ministry of Education) in Lanzhou University or Beta Analytic Testing Laboratory, respectively. Conventional 14C ages were calibrated with the IntCal20 calibration curve (Reimer et al., 2020) in the OxCal 4.4.4 online program (Bronk Ramsey, 2021).
Excluding one sample that experienced a low collagen yield (<0.5%), the remaining 56 samples yielded between 2.8 and 12.5% collagen. The yield of sample HSL-34 was too low (0.2%); thus, the data of this sample was rejected due to its poor reliability. The quality of bone after the long-term preservation in soil was tested by atomic C/N ratio. As shown in Table 1, these 56 samples (excluded five samples experienced failure during extraction and one sample with too low yield) are well preserved. The carbon content for all samples range between 41.5 and 44.1% while the nitrogen content range between 14.6 and 15.9%, generating atomic C/N ratios from 3.2 to 3.5, which fall into the acceptable range established by previous studies (DeNiro, 1985; Ambrose, 1990; van Klinken, 1999).
The stable carbon and nitrogen isotope data for all the individual animal samples are shown in Table 1 and Figure 3A. Data were further classified and analyzed by species (Table 2 and Figure 3B). The fauna utilized in Huoshiliang and Ganggangwa sites ranges widely in both δ13C and δ15N-values, from –22.7 to –7.2‰ with mean and standard deviation (SD, 1σ) of –15.3 ± 4.4‰ for δ13C, and from 3.0 to 10.8‰ with mean and SD at 7.7 ± 1.7‰ for δ15N, respectively. The δ15N-value of livestock ranges relatively closely, from 6.1 to 10.8‰ with mean and SD at 8.3 ± 1.0‰. However, the δ13C-value of livestock varied significantly between two groups consuming primarily C3 and dominantly C4 foods, respectively. For omnivorous livestock, more than half of the pigs (7/10) and all dogs consumed predominantly C4 diets according to their δ13C-values, while a small group of pigs (3/10) consumed mainly a C3 diet, which was analyzed separately. For herbivorous livestock, most of the cattle and sheep/goat consumed C3 or mixed C3/C4 but primarily C3 diets, with only one individual of each category consumed a C4 diet. The δ13C-value of wildlife mainly fitted with C3 diet, indicating a consuming reliance on the natural ecosystem, except for gazelles. Most of the gazelles (4/5) have mixed C3/C4- δ13C-values, indicating considerable intake of C4 plants. The δ15N-values of deer (4.2 ± 0.6‰) were notably lower than that of other species (Figure 3B).
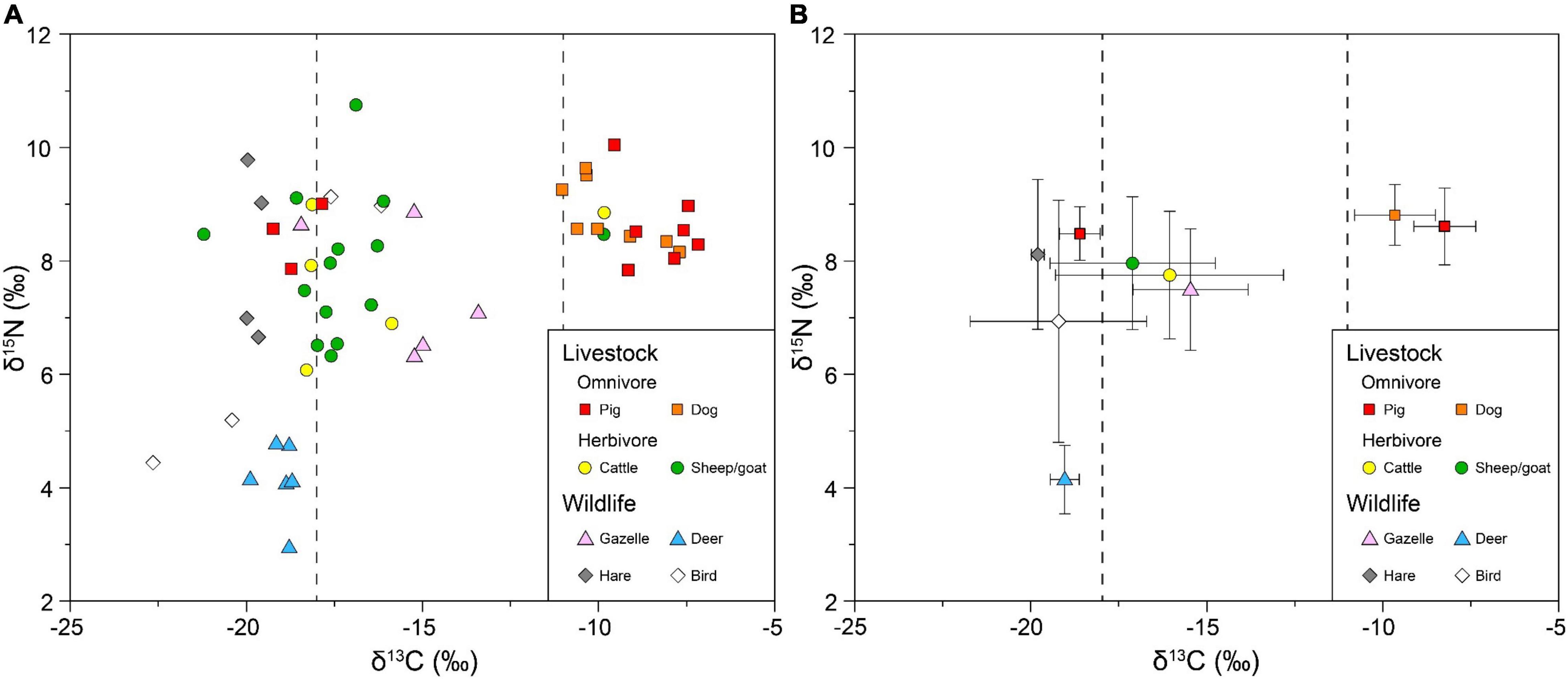
Figure 3. Plot of δ13C and δ15N-values measured from the fauna utilized by Huoshiliang and Ganggangwa sites with gray dotted lines demarcating C3, mixed C3/C4 and C4 diets. (A) Scatterplot of all animal individuals labeled as different species. (B) Scatterplot with X and Y error bars of different species while dot represents the mean value and error bar represents the standard deviation (SD, 1σ); pigs are divided into C3 and C4 groups to sharpen the distinction of their δ13C-values.
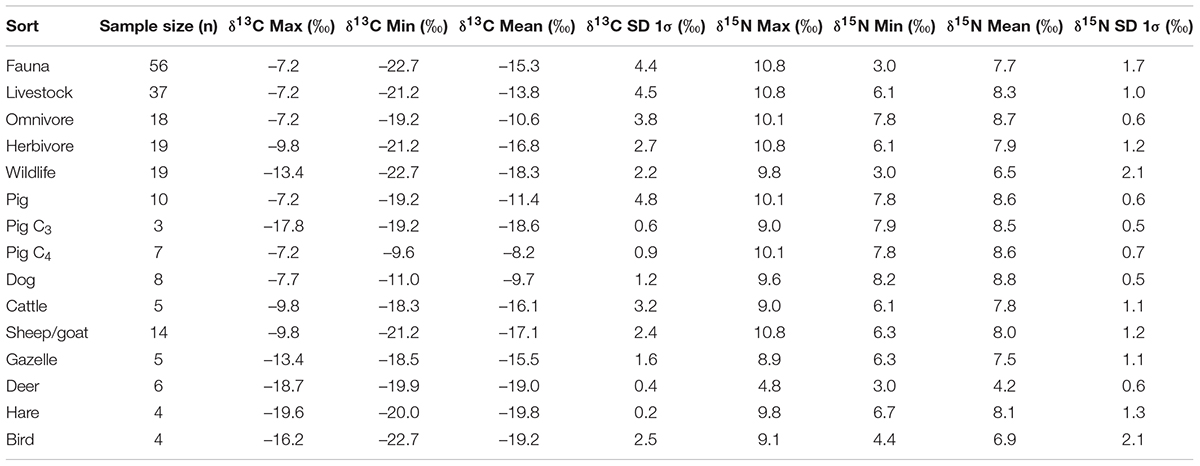
Table 2. Statistics of stable carbon and nitrogen isotope data reported in this study.
The radiocarbon dating results for the 15 directly dated animal collagen samples are listed in Table 3. Among these results, six dates with special labels were first reported in Ren et al. (2022) to represent the integral age of the faunal remains, while the other nine dates were reported for the first time in this study. Bone collagen was regarded as better material than charred wood for radiocarbon dating due to the potential influence of “old wood” on the latter (Gavin, 2001; Dong et al., 2014; Yang et al., 2019a). The conventional age of these 15 samples ranges from 3,610 to 3,480 BP, while the calibrated age 2σ ranges between 4,070 and 3,650 cal BP, basically consistent with previous studies (Dodson et al., 2009; Atahan et al., 2011).
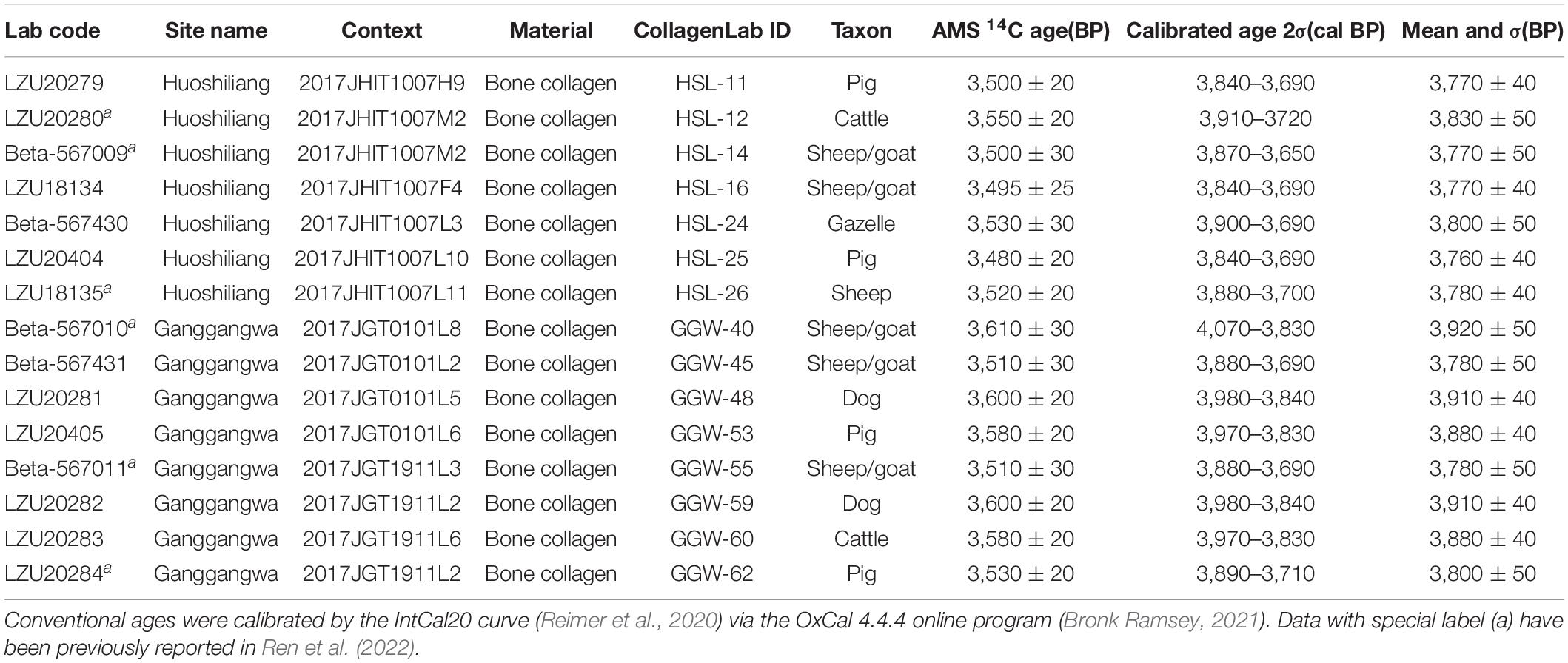
Table 3. Radiocarbon dating results of animal bones excavated from Huoshiliang and Ganggangwa sites.
The radiocarbon dating results reported in Ren et al. (2022) and this study focused on not only the starting and ending ages of the faunal remains, but also the precise age of the individuals with abnormal stable carbon isotope values. The cattle and sheep/goat individuals with C4 values, as well as two of the three pig individuals with C3 values, were chosen for radiocarbon dating. As the result shows, all radiocarbon dates of samples with abnormal δ13C-values fall into the latter half (generally later than ca. 3,850 cal BP) of the sites. Within the context of archeological features, the age of the only C3 pig individual without direct radiocarbon date falls into the latter half of the sites’ occupation, as decided by the two radiocarbon dates derived from the same archeological unit (the back fill of burial M2 in Huoshiliang).
As indicated by radiocarbon dating results and consecutive archeological deposits, animal resources were continuously utilized in Huoshiliang and Ganggangwa sites during 4,070–3,650 cal BP (calibrated age, 2σ range) (Table 3). Although the upper limit of the calibrated age 2σ approaches nearly 4,100 cal BP, the majority of the faunal remains are concentrated between 4,000 and 3,700 cal BP (Table 3), which falls into the time period of Xichengyi Culture as refined by a previous study (Yang et al., 2019a). Moreover, the two earliest conventional ages dated from surficial charcoal in Ganggangwa site reported in Dodson et al. (2009) and Atahan et al. (2011), which are nearly 100 years earlier than other results, may have been influenced significantly by the “old wood” effect (Gavin, 2001; Dong et al., 2014; Yang et al., 2019a), thus should be regarded more cautiously.
For wildlife, the relatively wide distribution in both δ13C and δ15N-values reflects their diverse habitats and diets (Figure 3 and Table 2). Stable isotope values of birds were the most distributed among the wildlife (Figure 3B), consistent with the diversified living space and varied diets of these creatures. Among wild herbivores, hare and deer present consistent diets in consuming prominent C3 plants as revealed by δ13C-values, which can reflect the C3 plant-dominated botanic situation of their habitat. However, the significant distinction in δ15N-values between hare and deer could be attributed to different extents of forest cover in their respective habitats and/or the metabolic variation under different aridity (Ambrose and DeNiro, 1986; Sealy et al., 1987; Schwarcz et al., 1999; Hartman, 2011; Hofman-Kamiñska et al., 2018), since the trophic levels of these species are similar. These hares are more likely to inhabit arid grassland, while the deer are more likely to inhabit moist forest in the piedmont of the Qilian Mountains. The gazelles presented generally mixed C3/C4 signal in δ13C-values (–15.5 ± 1.6‰), which is different from other herbivorous wildlife reported in this study (Figure 3). The relatively positive δ13C-values of these gazelles (about 3.5‰ higher than the deer and slightly higher than the cattle and sheep/goat) demonstrate notable intake of C4 plants and/or ecological influences such as the canopy effect and watering effect (e.g., Drucker et al., 2008; Wallace et al., 2013). Though some scholars argued that wild C4 plants in the arid desert constitute the C4 intake of wild herbivores in Hexi Corridor (Ma et al., 2021), it is impossible to eliminate the potential contribution of C4 crops in these gazelles’ diets. Moreover, the similarity of δ15N-values between gazelles and herbivorous livestock (Figure 3B) may also indicate their overlapped living space. A case study on African grasslands revealed that prehistoric human activity had shaped the savanna landscapes via enriching nutrient for the local ecosystem (Marshall et al., 2018), which indicates that human settlement in arid area could make the perimeter zone more nutritious for wildlife to inhabit. Furthermore, the “inadvertent baiting” from human cultivated crops on wild herbivore was hypothesized and explored in eastern North America (Svizzero, 2016; Bonzani et al., 2021), which has shed light on the relation between agricultural groups and the neighboring wild animals. Thus, we presented a conjecture that the massive cultivation of millet crops during this period around the human settlements attracted the gazelles to forage around and accidently got hunted. This speculation also describes a mutual beneficial relation between human and wildlife, though still requiring further validation.
For livestock, the δ13C-values fall into two distinct groups, while the δ15N-values are more concentrated relative to wildlife (Figure 3). Carbon isotope data suggest that all of the domestic dogs (8/8) and the majority of the domestic pigs (7/10) sampled in Huoshiliang and Ganggangwa consumed C4 diets (Figure 3 and Table 1), indicating the consumption of millet, such as millet grains, millet by-products, and/or human leftovers, which is consistent with the tradition of Neolithic cultures in northwestern China (e.g., Barton et al., 2009; Chen et al., 2016; Ma et al., 2021). For the non-indigenous herbivorous livestock, most of the domestic cattle (4/5) and domestic sheep/goat (13/14) exhibited carbon isotope values indicative of a C3 diet with a minor contribution of C4 plants (Figure 3 and Table 1). With the exception of two individuals exhibiting high δ13C-values (one cattle and one sheep/goat sample), the results are consistent with previous studies in the EBA of the Hexi Corridor and can be attributed to the consumption of wild C3 plants during herding (Atahan et al., 2011; Ma et al., 2021; Vaiglova et al., 2021). Wild C4 plants in arid desert and/or deliberate feeding of millet by-products were pointed out to constitute the intake of C4 foodstuff of herbivorous livestock in Hexi Corridor (Su and Yan, 2008; Ma et al., 2021). The δ15N-values for domestic pigs and dogs were more concentrated and slightly higher (8.7 ± 0.6‰) than values for herbivorous livestock (7.9 ± 1.2‰). The close δ15N-values between omnivorous livestock and herbivores indicate limited protein consumption by these omnivorous livestock. The broader distribution of δ15N-values for herbivorous livestock could result from grazing in different ecological zones (e.g., Pearson et al., 2007; Atahan et al., 2011) since the natural environment along the Heihe River valley as well as the surrounding regions in Hexi Corridor varies significantly from alpine meadow and coniferous forest to desert grassland and desert. The more concentrated δ15N-values of domestic pig and dog indicate a relatively unified living environment and feeding pattern adopted by local people, which might involve captive breeding and deliberate feeding.
Notably, several individuals among the livestock have abnormal δ13C-values; some herbivorous individuals (one cattle and one sheep/goat) appear to have consumed a C4 diet while some omnivorous individuals (three pigs) appear to have consumed a C3 diet (Figure 3). The C3 signal in omnivorous individuals of these two sites has been previously detected and was attributed to the consumption of wild foods (Atahan et al., 2011). The same phenomenon of C3-consuming pigs in coetaneous Xichengyi site was regarded as the result of free-ranging and/or inhabitation site raising with C3 consumption (Ma et al., 2021). However, considering the open arid modern situation and the low productivity of the ecosystem around these two sites, the feasibility of free-ranging pigs remains questionable. Moreover, the abnormal C4 signal of herbivorous livestock has not been well explained yet. It is likely that the existence of these abnormal livestock individuals relates to the transformation of animal husbandry strategies. However, some further investigation of geochemical tracing such as Sr isotope and trace element analyses on these abnormal individuals would be meaningful to examine the issue of local/extra-local provenance since the possibility of livestock’s immigration could not be eliminated.
As indicated by direct radiocarbon dating results supplementary with stratigraphic evidence, a remarkable diversification in δ13C-values of livestock during the latter half of the occupation at these two sites has been revealed. The relation between calibrated radiocarbon ages (shown as mean value and SD) and δ13C-values of the 15 directly dated samples is plotted in Figure 4. The ages of all individuals consuming abnormal diet fall into the latter half of these two sites, basically later than ca. 3,850 cal BP. Radiocarbon dating results supplementary with stratigraphic evidence also reveal that livestock consuming conventional diets existed in both former and latter halves of these two sites’ occupation (Figure 4 and Table 1). Thus, 3,850 BP is possibly the timing for the initial diversification in livestock feeding pattern at Huoshiliang and Ganggangwa sites. According to archaeobotanical findings, the latter half of the occupation at the two sites (ca. 3,850–3,700 BP) witnessed the promotion of wheat and barley cultivation in Hexi Corridor, with the proportion of these non-indigenous crop seeds reached up to roughly a quarter among all crop seeds (Zhou et al., 2016; Yang, 2017). Stable isotope evidence also reveals the consumption of C3 crops by humans since 4,000–3,800 BP (Liu et al., 2014). This evidence proves the potential for humans in the Huoshiliang and Ganggangwa sites to provide their livestock with these non-indigenous C3 crops. Furthermore, pigs consumed a similar diet to humans (Liu and Jones, 2014), and the appearance of C3 consumption in their diets was synchronous with the preliminary shift with increased C3 intake in human diets. Additionally, the individuals with abnormal δ13C-values nonetheless have δ15N-values that are similar to the majority of omnivorous livestock (Figure 3), suggesting similar living environment and feeding patterns. Thus, we speculated that the domestic pigs with C3 signal were more likely to be raised at their inhabitation site, instead of free-ranging, and were deliberately fed with C3 foodstuff, such as wild plants and/or the products or human leftovers of wheat/barley. Similarly, herbivorous individuals with C4 signal are likely to be adopted with the same management as the omnivorous livestock, including captive breeding and deliberate feeding of C4 plants, most likely the products or human leftover of millet crops.
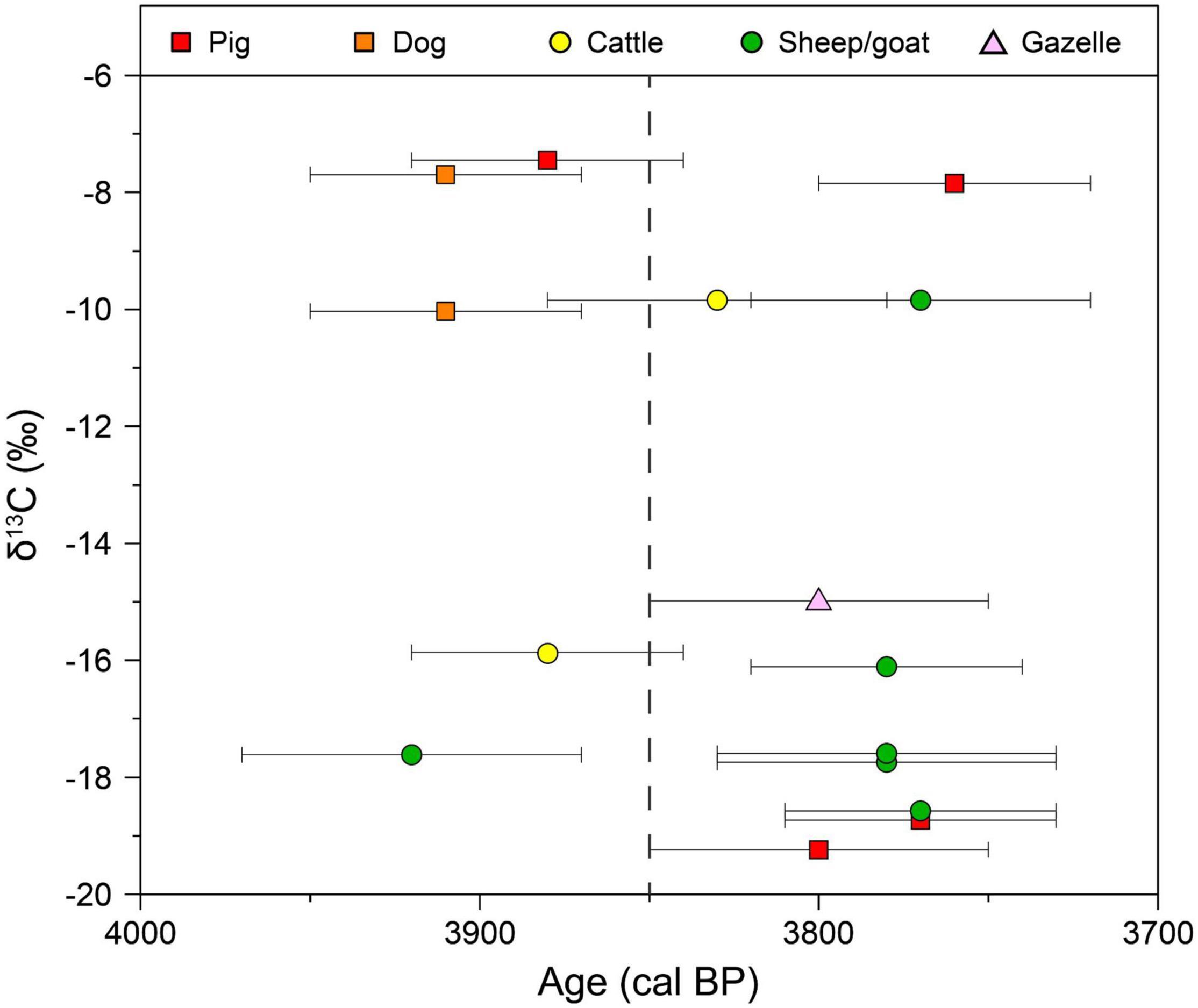
Figure 4. Plot of precise ages and δ13C-values of the 15 directly dated samples. The dot and bar represent the mean value and SD (1σ) of each calibrated age, respectively. The gray dotted line at 3,850 BP roughly divided the ages into early and late phases.
In summary, the absolute age of the faunal remains from Huoshiliang and Ganggangwa sites ranges from 4,070 to 3,650 cal BP (2σ), but are more concentrated between 4,000 and 3,700 cal BP (Table 3 and Figure 4). The appearance of both omnivorous and herbivorous livestock with abnormal δ13C-values during the latter half of the sites’ occupation reveals a diversified livestock feeding pattern adopted by humans. It is likely that some herbivorous livestock were raised at their inhabitation site and deliberately fed with millet products, while a small group of domestic pigs were deliberately fed with C3 foodstuff, involving generally wild C3 plants and/or products or human leftovers of wheat and barley. The initial timing for the diversification in livestock feeding pattern is circa 3,850 BP.
To obtain clearer insight into the temporal variation of livestock feeding pattern in prehistoric Hexi Corridor, published stable isotope data from faunal remains (Atahan et al., 2011; Zhang et al., 2015; Yang, 2017; Yang et al., 2019b; Ma et al., 2021; Vaiglova et al., 2021) were combined with data presented in this study (Supplementary Table 1). Stable carbon isotope data from four categories of major livestock involved in our discussion were charted as box plot and distinguished via cultural affiliations of different site and the material (Figure 5). The cultural affiliations of the materials reported in their original studies involved in Supplementary Table 1 and Figure 5 were reconfirmed through published archeological excavation reports, and the absolute age of each cultural period was determined via their refined chronology (Yang et al., 2019a). According to the report of excavation in Xihetan site (Zhao, 2005) and recent archeological findings in other Xichengyi Culture sites, the cultural affiliation of this site has been refined as Xichengyi Culture (ca. 4,000–3,600 BP). The cultural affiliation of animal stable isotope data from Huoshaogou site (Vaiglova et al., 2021) has been refined as Shanma Culture referring to recently published excavation reports (Wang et al., 2019, 2021). Data collected in Supplementary Table 1 were then divided into four phases in accordance with their cultural affiliations to investigate a more detailed shifting trajectory of livestock δ13C-values. An overlap of time between phase 2 and 3 occurred due to the overlap between Xichengyi/Qijia and Siba Cultures (Yang et al., 2019a).
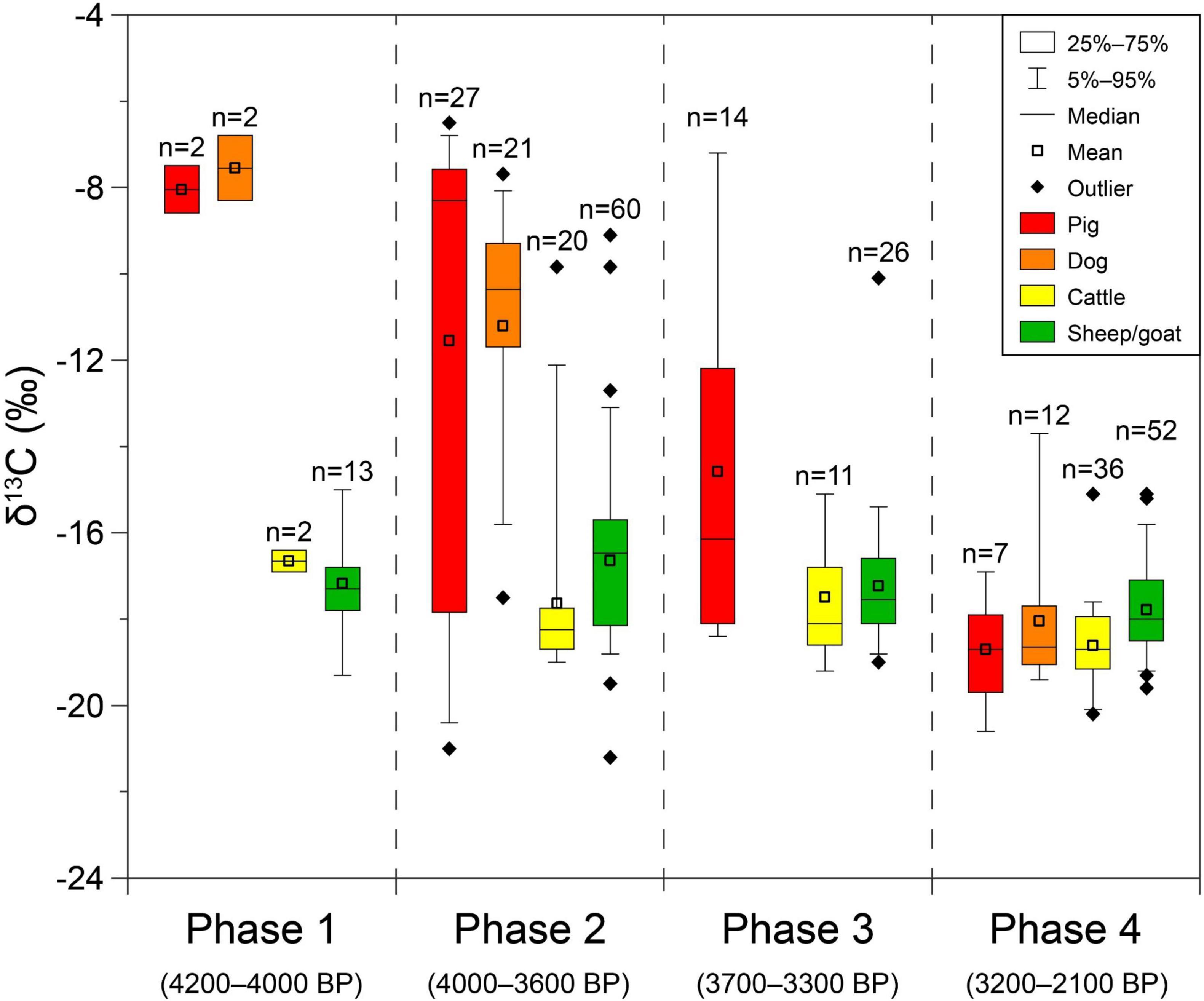
Figure 5. Box plot of stable carbon isotope data of four major omnivorous and herbivorous livestock in prehistoric Hexi Corridor from published works (Atahan et al., 2011; Zhang et al., 2015; Yang, 2017; Yang et al., 2019b; Ma et al., 2021; Vaiglova et al., 2021) and this study (Supplementary Table 1). Data are divided into four phases: (1) Machang Culture; (2) Xichengyi/Qijia Culture; (3) Siba Culture; and (4) Dongjiatai and Shanma/Shajing Cultures.
The population of Majiayao, a type of Late Neolithic culture originated in the upper Yellow River Valley in northwestern China, is currently known as the earliest agricultural immigrants to expand westward and to enter Hexi Corridor at ca. 4,800 BP (Dong et al., 2018), brought in a pattern of subsistence that mainly engaged in millet cultivation and pig-dog feeding (Yang, 2017; Yang et al., 2019b). In the following eight centuries, the Late Neolithic culture in Hexi Corridor evolved into Banshan Culture (ca. 4,450–4,200 BP) and Machang Culture (ca. 4,200–4,000 BP) in succession (Yang et al., 2019a).
During the period of Machang Culture (ca. 4,200–4,000 BP), the eastern-originated foxtail millet and broomcorn millet were major crops and staple foods in the diets of humans and omnivorous livestock, as indicated by archaeobotanical and stable isotope data (Liu et al., 2014; Zhou et al., 2016; Yang et al., 2019b). This pattern of human subsistence was consistent with coetaneous and earlier societies in the neighboring Gansu-Qinghai region (Barton et al., 2009; Jia et al., 2013; Ma et al., 2016, 2021; Ren et al., 2020). Cattle and sheep/goat, the non-indigenous herbivorous livestock, were estimated to be introduced into Hexi Corridor during this period, though direct radiocarbon dating results were slightly younger (Yang, 2017; Yang et al., 2019b; Ren et al., 2022). The δ13C-values of these early cattle and sheep/goat revealed a C3 plant-dominated consumption, which is consistent with the natural vegetation (Yang et al., 2019b; Figure 5). The feeding patterns of both indigenous omnivorous livestock and novel herbivorous livestock during this period in Hexi Corridor are revealed to be consistent with their traditions, respectively.
The period of time during Xichengyi/Qijia Cultures (ca. 4,000–3,600 BP) witnessed the phenomenal diversification in livestock feeding patterns in Hexi Corridor, with significant expansion of δ13C-values among both herbivorous and omnivorous livestock (Figure 5). As revealed by archaeobotanical evidence, wheat and barley were introduced into Hexi Corridor and widely spread during this period (Dodson et al., 2013; Zhou et al., 2016; Yang, 2017). Charred seeds of these Western-originated crops were commonly unearthed from contemporaneous archeological sites, reaching a proportion up to circa a quarter among all crop remains (Zhou et al., 2016; Yang, 2017). Correspondingly, the contribution of C3 crops in human diet emerged and gradually increased during this period (Liu et al., 2014, 2016). The importance of herbivorous livestock also increased (Yang, 2017; Yang et al., 2019b), together with the increased practice of pastoralism (Atahan et al., 2011; Ma et al., 2021). As shown in Figure 5, the consumption of C3 foodstuff in omnivorous livestock enhanced significantly. The presence of several domestic omnivorous individuals that mainly consumed C3 plants (Supplementary Table 1) is suggestive of deliberate feeding with C3 crop by-products and/or allowing the animals to free range (Ma et al., 2021). In contrast, the general dietary shift of herbivorous livestock was not that prominent (Figure 5). Domestic herbivorous individuals with dominant intake of C4 plants are likely to be raised at inhabitation site and deliberately fed with millet products, similar to the tradition of raising pigs and dogs (e.g., Barton et al., 2009; Chen et al., 2016; Ma et al., 2021). This bilateral transformation of feeding patterns between domestic omnivores and domestic herbivores jointly constructed the diversification in livestock feeding patterns during EBA in Hexi Corridor. Combined with the result shown in this study, the initial timing for this transformation could possibly be ca. 3,850 BP (Figure 3). Moreover, the motivation behind such attempt to partly break with the feeding traditions for both indigenous and novel livestock may involve the exploration of new animal husbandry manners to realize a more efficient economy and better social sustainability, and to withstand the harsh arid living environment in Hexi Corridor.
When it came to the Siba Culture (ca. 3,700–3,300 BP), the diversification in livestock feeding pattern persisted (Figure 5). During this period, wheat and barley became the primary agricultural crops (Zhou et al., 2016; Yang, 2017; Yang et al., 2019b). Meanwhile, cattle and sheep/goat became primary livestock (Yang, 2017; Yang et al., 2019b). The consumption of C3 foodstuff by humans and livestock also increased significantly during this period (Liu et al., 2014; Yang, 2017; Yang et al., 2019b), consistent with the neighboring Gansu-Qinghai region in northwestern China (Ma et al., 2016; Vaiglova et al., 2021). As revealed by Figure 5, a reduction of C4 food consumption in domestic pigs took place. The majority of domestic pigs shifted to C3-based diets, making the C4 consumers the minority. The dietary tendency of domestic dogs during this period remains unknown due to the absence of stable isotope data. The diet of herbivorous livestock at this phase generally returned to the initial status with C3 consumption dominated. Only a single sheep/goat individual remained consuming mainly C4 plants (Figure 5) during this period, indicating the general termination of dietary diversification among herbivorous livestock.
The δ13C-values of phase 4 (ca. 3,300–2,100 BP) demonstrate generally C3-based diets among all major livestock (Figure 5). This situation is consistent with the popularization of wheat and barley, as well as the fall of millet-based agriculture in northern China (e.g., Dodson et al., 2013; Chen et al., 2015, 2020; Liu et al., 2017). The dietary shift from C4-based to C3-based among omnivorous livestock seems to have been accomplished since ca. 3,200 BP (Figure 5). Meanwhile, the diet of herbivorous livestock returned to C3-dominated as well. For the last millennium before the common era, the rise and expansion of nomadism deeply influenced the Eurasian Steppe and northwestern China (Kuz’mina, 2008). The agro-pastoral residents in Hexi Corridor established more developed pastoral economy after ca. 2,800 BP, which are known as Shanma Culture (ca. 2,900–2,100 BP) in western Hexi Corridor and Shajing Culture (ca. 2,700–2,100 BP) in eastern Hexi Corridor (Li, 2011; Yang et al., 2019a).
In summary, the trajectory for the transformation of livestock feeding patterns in the prehistoric Hexi Corridor has been revealed as follows: (1) diets of both omnivorous and herbivorous livestock conformed with their original traditions during the Machang Culture (ca. 4,200–4,000 BP); (2) during the Xichengyi/Qijia Culture (ca. 4,000–3,600 BP), the range of δ13C-values exhibited by both omnivorous and herbivorous livestock expanded dramatically, indicating a diversified livestock feeding pattern during EBA; (3) during the Siba Culture (ca. 3,700–3,300 BP), herbivorous livestock generally returned to a C3 diet, but domestic pigs continued to consume a wide array of C3–C4 food products; and finally (4) the transformation of livestock feeding patterns seems to have fully completed during the Dongjiatai and Shanma/Shajing Cultures (ca. 3,200–2,100 BP), with all four major livestock groups in the Hexi Corridor consuming C3 diets.
In this study, systematic stable carbon and nitrogen isotope analysis and precise radiocarbon dating were performed on faunal remains acquired through archeological excavation of two EBA settlements in middle Hexi Corridor, Huoshiliang, and Ganggangwa sites. In total, stable carbon and nitrogen isotope data were acquired for 56 faunal samples and 15 direct radiocarbon dates were obtained. The stable isotope results of domestic animals reveal a significant diversification in livestock feeding pattern, with some domestic pigs consuming mainly C3 foodstuff and some herbivorous livestock consumed mainly C4 plants, respectively. It seems that humans in these two sites attempted to deliberately feed some of the herbivorous livestock following the tradition in feeding omnivorous livestock; and to deliberately feed some of the domestic pigs with C3 foodstuff, possibly involving wheat/barley products. Supplemented by stratigraphic evidence, the precise radiocarbon dating results indicate that the initial timing for the diversification in livestock feeding pattern lies at ca. 3,850 BP.
Integrated with the summary of published animal stable isotope data, a vivid animation for the transformation of livestock feeding pattern during prehistoric times in Hexi Corridor was exhibited. By adopting more precise phase division, the trajectory of the transformation in livestock feeding pattern was further clarified. A notable diversification in livestock feeding pattern during EBA, which is consistent with the pattern revealed in Huoshiliang and Ganggangwa sites, has been detected. Such diversification existed in the transformation from millet-based agricultural society to wheat/barley-based agro-pastoral society in prehistoric Hexi Corridor, and possibly took place at ca. 3,850 BP. We argued that the motivation behind such attempt to partly break with the feeding traditions involves the exploration to innovate animal husbandry strategies. Such exploration may be effective to improve the economic efficiency and the social sustainability, which can help the local people to withstand the harsh arid living environment in Hexi Corridor.
The original contributions presented in this study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
Ethical review and approval was not required for the animal study because this study only envolves ancient animal skeletal remains (also known as a kind of fossil remain), which is totally distinct from the study of living animals. Thus, we confirm that there is no need of ethical review.
LR and GC designed the study. MQ, SZ, YY, and GC participated in the fieldwork. MQ, HL, ML, RL, and LR conducted the experiment and data analysis. MQ, SZ, YY, HL, and LR wrote the manuscript. All authors contributed to the article and approved the submitted version.
This research was supported by the National Natural Science Foundation of China (Grant No. 41901089), the National Key R&D Program of China (Grant No. 2018YFA0606402), and the Second Tibetan Plateau Scientific Expedition and Research Program (STEP) (Grant No. 2019QZKK0601).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor declared a past co-authorship with one of the authors, GC.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We would like to thank Guanghui Dong, Minmin Ma, Pingyu Zhang, and Linyao Du for the participation of discussions. We would also like to thank Zhilin Shi and Yifu Cui for the participation of field work.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.908131/full#supplementary-material
Ambrose, S. H. (1990). Preparation and characterization of bone and tooth collagen for isotopic analysis. J. Archaeol. Sci. 17, 431–451. doi: 10.1016/0305-4403(90)90007-R
Ambrose, S. H., and DeNiro, M. J. (1986). The isotopic ecology of East Africa mammals. Oecologia 69, 395–406. doi: 10.1007/BF00377062
Anthony, D. W. (2010). The Horse, the Wheel, and Language: How Bronze-Age Riders from the Eurasian Steppes Shaped the Modern World. Princeton: Princeton University Press.
Atahan, P., Dodson, J., Li, X., Zhou, X., Hu, S., Bertuch, F., et al. (2011). Subsistence and the isotopic signature of herding in the Bronze Age Hexi Corridor, NW Gansu, China. J. Archaeol. Sci. 38, 1747–1753. doi: 10.1016/j.jas.2011.03.006
Barton, L., Newsome, S. D., Chen, F., Wang, H., Guilderson, T. P., and Bettinger, R. L. (2009). Agricultural origins and the isotopic identity of domestication in northern China. Proc. Natl. Acad. Sci. U.S.A. 106, 5523–5528. doi: 10.1073/pnas.0809960106
Bonzani, R. M., Alexander, K. V., Metz, A., Munizzi, J. S., Manzano, B. L., Davidson, M. J., et al. (2021). Using deer stable isotope data to test a niche construction hypothesis for an increase in prehistoric human maize consumption in the Eastern Woodlands of the United States. Environ. Archaeol. 27, 258–276. doi: 10.1080/14614103.2021.1921674
Bradley, D. G., Loflus, R. T., Cunningham, P., and Machugh, D. E. (1998). Genetics and domestic cattle origins. Evol. Anthropol. 6, 79–86. doi: 10.1002/(SICI)1520-650519986:3<79:AID-EVAN2<3.0.CO;2-R
Bronk Ramsey, C. (2021). OxCal version 4.4.4. Available online at: https://c14.arch.ox.ac.uk/oxcal.html (accessed February 20, 2022).
Chan, D., Wu, Q., Jiang, G., and Dai, X. (2016). Projected shifts in Köppen climate zones over China and their temporal evolution in cmip5 multi-model simulations. Adv. Atmos. Sci. 33, 283–293. doi: 10.1007/s00376-015-5077-8
Chen, F., Dong, G., Zhang, D., Liu, X., Jia, X., An, C., et al. (2015). Agriculture facilitated permanent human occupation of the Tibetan Plateau after 3600 BP. Science 347, 248–250. doi: 10.1126/science.1259172
Chen, T., Qiu, M., Liu, R., Li, H., Hou, H., Howarth, P., et al. (2020). Human responses to climate change in the late prehistoric Western Loess Plateau, northwest China. Radiocarbon 62, 1193–1207. doi: 10.1017/RDC.2020.32
Chen, X., Fang, Y., Hu, Y., Hou, Y., Lü, P., Yuan, J., et al. (2016). Isotopic reconstruction of the late Longshan period (ca. 4200-3900 BP) dietary complexity before the onset of state-level societies at the Wadian site in the Ying River valley, Central Plains, China. Int. J. Osteoarchaeol. 26, 808–817. doi: 10.1002/oa.2482
Dai, L., Li, Z., Zhao, C., Yuan, J., Hou, L., Wang, C., et al. (2016). An isotopic perspective on animal husbandry at the Xinzhai site during the initial stage of the legendary Xia dynasty (2070–1600 BC). Int. J. Osteoarchaeol. 26, 885–896. doi: 10.1002/oa.2503
DeNiro, M. J. (1985). Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317, 806–809. doi: 10.1038/317806a0
Diamond, J., and Bellwood, P. (2003). Farmers and their languages: the first expansions. Science 300, 597–603. doi: 10.1126/science.1078208
Dodson, J., Li, X., Ji, M., Zhao, K., Zhou, X., and Levchenko, V. (2009). Early bronze in two Holocene archaeological sites in Gansu, NW China. Quat. Res. 72, 309–314. doi: 10.1016/j.yqres.2009.07.004
Dodson, J. R., Li, X., Zhou, X., Zhao, K., Sun, N., and Atahan, P. (2013). Origin and spread of wheat in China. Quat. Sci. Rev. 72, 108–111. doi: 10.1016/j.quascirev.2013.04.021
Dong, G., Qiu, M., Li, R., and Chen, F. (2021a). Using the Fulcrum Cognitive Model to explore the mechanism of past human-land co-evolution (in Chinese). Acta Geogr. Sin. 76, 15–29. doi: 10.11821/dlxb202101002
Dong, G., Wang, L., Zhang, D. D., Liu, F., Cui, Y., Li, G., et al. (2021b). Climate-driven desertification and its implications for the ancient Silk Road trade. Clim. Past 17, 1395–1407. doi: 10.5194/cp-17-1395-2021
Dong, G., Wang, Z., Ren, L., Motuzaite Matuzeviciute, G., Wang, H., Ren, X., et al. (2014). A comparative study of 14C dating on charcoal and charred seeds from Late Neolithic and Bronze Age sites in Gansu and Qinghai provinces, NW China. Radiocarbon 56, 157–163. doi: 10.2458/56.16507
Dong, G., Yang, Y., Han, J., Wang, H., and Chen, F. (2017). Exploring the history of cultural exchange in prehistoric Eurasia from the perspectives of crop diffusion and consumption. Sci. China Earth Sci. 60, 1110–1123. doi: 10.1007/s11430-016-9037-x
Dong, G., Yang, Y., Liu, X., Li, H., Cui, Y., Wang, H., et al. (2018). Prehistoric trans-continental cultural exchange in the Hexi Corridor, northwest China. Holocene 28, 621–628. doi: 10.1177/0959683617735585
Drucker, D. G., Bridault, A., Hobson, K. A., Szuma, E., and Bocherens, H. (2008). Can carbon-13 in large herbivores reflect the canopy effect in temperate and boreal ecosystems? Evidence from modern and ancient ungulates. Palaeogeogr. Palaeoclimatol. Palaeoecol. 266, 69–82. doi: 10.1016/j.palaeo.2008.03.020
Flad, R. K., Yuan, J., and Li, S. (2007). Zooarchaeological evidence for animal domestication in northwest China. Dev. Quat. Sci. 9, 167–203. doi: 10.1016/S1571-0866(07)09012-4
Frachetti, M. D., Smith, C. E., Traub, C. M., and Williams, T. (2017). Nomadic ecology shaped the highland geography of Asia’s Silk Roads. Nature 543, 193–198. doi: 10.1038/nature21696
Fuller, D. Q., Qin, L., Zheng, Y., Zhao, Z., Chen, X., Hosoya, L. A., et al. (2009). Domestication rate in rice: spikelet bases from the lower Yangtze. Science 323, 1607–1610. doi: 10.1126/science.1166605
Gansu Provincial Institute of Cultural Relics and Archaeology (2020). Important Archaeological Findings in GANSU (2000–2019) (in Chinese). Beijing: Cultural Relics Publishing House, 168–175.
Gavin, D. G. (2001). Estimation of inbuilt age in radiocarbon ages of soil charcoal for fire history studies. Radiocarbon 43, 27–44. doi: 10.1017/S003382220003160X
Hartman, G. (2011). Are elevated δ15N values in herbivores in hot and arid environments caused by diet or animal physiology? Funct. Ecol. 25, 122–131. doi: 10.1111/j.1365-2435.2010.01782.x
Hermes, T. R., Frachetti, M. D., Doumani Dupuy, P. N., Mar’yashev, A., Nebel, A., and Makarewicz, C. A. (2019). Early integration of pastoralism and millet cultivation in Bronze Age Eurasia. Proc. Biol. Sci. 286:20191273. doi: 10.1098/rspb.2019.1273
Hermes, T. R., Tishkin, A. A., Kosintsev, P. A., Stepanova, N. F., Krause-Kyora, B., and Makarewicz, C. A. (2020). Mitochondrial DNA of domesticated sheep confirms pastoralist component of Afanasievo subsistence economy in the Altai Mountains (3300–2900 cal BC). Archaeol. Res. Asia 24:100232. doi: 10.1016/j.ara.2020.100232
Hofman-Kamiñska, E., Bocherens, H., Borowik, T., Drucker, G. D., and Kowalczyk, R. (2018). Stable isotope signatures of large herbivore foraging habitats across Europe. PLoS One 13:e0190723. doi: 10.1371/journal.pone.0190723
Honeychurch, W., Rogers, L., Amartuvshin, C., Diimaajav, E., Erdene-Ochir, N., Hall, M. E., et al. (2021). The earliest herders of East Asia: examining Afanasievo entry to Central Mongolia. Archaeol. Res. Asia 26:100264. doi: 10.1016/j.ara.2021.100264
Hu, Y., Wang, S., Luan, F., Wang, C., and Richards, M. P. (2008). Stable isotope analysis of humans from Xiaojingshan site: implications for understanding the origin of millet agriculture in China. J. Archaeol. Sci. 35, 2960–2965. doi: 10.1016/j.jas.2008.06.002
Jaang, L. (2015). The landscape of China’s participation in the Bronze Age Eurasian network. J. World Prehist. 28, 179–213. doi: 10.1007/s10963-015-9088-2
Jia, X., Dong, G., Li, H., Brunson, K., Chen, F., Ma, M., et al. (2013). The development of agriculture and its impact on cultural expansion during the late Neolithic in the Western Loess Plateau, China. Holocene 23, 83–90. doi: 10.1177/0959683612450203
Jiang, W., Wu, H., Li, Q., Lin, Y., and Yu, Y. (2019). Spatiotemporal changes in C4 plant abundance in China since the Last Glacial Maximum and their driving factors. Palaeogeogr. Palaeoclimatol. Palaeoecol. 518, 10–21. doi: 10.1016/j.palaeo.2018.12.021
Jones, M., Hunt, H., Kneale, C., Lightfoot, E., Lister, D., Liu, X., et al. (2016). Food globalisation in prehistory: the agrarian foundations of an interconnected continent. J. Brit. Acad. 4, 73–87. doi: 10.5871/jba/004.073
Jones, M., Hunt, H., Lightfoot, E., Lister, D., Liu, X. Y., and Motuzaite-Matuzeviciute, G. (2011). Food globalization in prehistory. World Archaeol. 43, 665–675. doi: 10.1080/00438243.2011.624764
Jones, M. K., and Liu, X. (2009). Origins of agriculture in East Asia. Science 324, 730–731. doi: 10.1126/science.1172082
Kuz’mina, E. E. (2008). The Prehistory of the Silk Road. Philadelphia, PA: University of Pennsylvania Press.
Lee-Thorp, J. A. (2008). On isotopes and old bones. Archaeometry 50, 925–950. doi: 10.1111/j.1475-4754.2008.00441.x
Li, H., Liu, F., Cui, Y., Ren, L., Storozum, M. J., Qin, Z., et al. (2017). Human settlement and its influencing factors during the historical period in an oasis-desert transition zone of Dunhuang, Hexi Corridor, Northwest China. Quat. Int. 458, 113–122. doi: 10.1016/j.quaint.2016.11.044
Li, S. (2011). The Report on the Survey of Prehistoric Archaeology in the Hexi Corridor (in Chinese). Beijing: Cultural Relics Press.
Li, X., Sun, N., Dodson, J., Ji, M., Zhao, K., and Zhou, X. (2011). The impact of early smelting on the environment of Huoshiliang in Hexi Corridor, NW China, as recorded by fossil charcoal and chemical elements. Palaeogeogr. Palaeoclimatol. Palaeoecol. 305, 329–336. doi: 10.1016/j.palaeo.2011.03.015
Li, X., Zhang, S., Lu, M., Qiu, M., Wen, S., and Ma, M. (2020). Dietary shift and social hierarchy from the Proto-Shang to Zhou Dynasty in the Central Plains of China. Environ. Res. Lett. 15:035002. doi: 10.1088/1748-9326/ab6783
Li, Y., Lu, P., Mao, L., Chen, P., Yan, L., and Guo, L. (2021). Mapping spatiotemporal variations of Neolithic and Bronze Age settlements in the Gansu-Qinghai region, China: scale grade, chronological development, and social organization. J. Archaeol. Sci. 129:105357. doi: 10.1016/j.jas.2021.105357
Liu, F., Yang, Y., Chen, G., Zhang, S., and Zhang, H. (2021). Changes in wood utilization due to Iron Age jade mining in the western Hexi Corridor: wood charcoal investigations. Front. Earth Sci. 9:636534. doi: 10.3389/feart.2021.636534
Liu, X., and Jones, M. K. (2014). Food globalisation in prehistory: Top down or bottom up? Antiquity 88, 956–963. doi: 10.1017/S0003598X00050912
Liu, X., Jones, P. J., Motuzaite Matuzeviciute, G., Hunt, H. V., Lister, D. L., An, T., et al. (2019a). From ecological opportunism to multi-cropping: mapping food globalisation in prehistory. Quat. Sci. Rev. 206, 21–28. doi: 10.1016/j.quascirev.2018.12.017
Liu, F., Yang, Y., Shi, Z., Storozum, M. J., and Dong, G. (2019b). Human settlement and wood utilization along the mainstream of Heihe River basin, northwest China in historical period. Quat. Int. 516, 141–148. doi: 10.1016/j.quaint.2018.05.033
Liu, F., Li, H., Cui, Y., Yang, Y., Lee, H. F., Ding, D., et al. (2019c). Chronology and plant utilization from the earliest walled settlement in the Hexi Corridor, northwestern China. Radiocarbon 61, 971–989. doi: 10.1017/RDC.2019.57
Liu, X., Lightfoot, E., O’Connell, T. C., Wang, H., Li, S., Zhou, L., et al. (2014). From necessity to choice: dietary revolutions in west China in the second millennium BC. World Archaeol. 46, 661–680. doi: 10.1080/00438243.2014.953706
Liu, X., Lister, D. L., Zhao, Z., Petrie, C. A., Zeng, X., Jones, P. J., et al. (2017). Journey to the East: diverse routes and variable flowering times for wheat and barley en route to prehistoric China. PLoS One 12:e0187405. doi: 10.1371/journal.pone.0187405
Liu, X., Reid, R. E. B., Lightfoot, E., Motuzaite Matuzeviciute, G., and Jones, M. K. (2016). Radical change and dietary conservatism: mixing model estimates of human diets along the Inner Asia and China’s mountain corridors. Holocene 26, 1556–1565. doi: 10.1177/0959683616646842
Lu, H., Zhang, J., Liu, K., Wu, N., Li, Y., Zhou, K., et al. (2009). Earliest domestication of common millet (Panicum miliaceum) in East Asia extended to 10000 years ago. Proc. Natl. Acad. Sci. U.S.A. 106, 7367–7372. doi: 10.1073/pnas.0900158106
Ma, M., Dong, G., Jia, X., Wang, H., Cui, Y., and Chen, F. (2016). Dietary shift after 3600 cal yr BP and its influencing factors in northwestern China: evidence from stable isotopes. Quat. Sci. Rev. 145, 57–70. doi: 10.1016/j.quascirev.2016.05.041
Ma, M., Ren, L., Li, Z., Wang, Q., Zhao, X., and Li, R. (2021). Early emergence and development of pastoralism in Gan-Qing region from the perspective of isotopes. Archaeol. Anthropol. Sci. 13:93. doi: 10.1007/s12520-021-01331-2
Marshall, F., Reid, R. E. B., Goldstein, S., Storozum, M., Wreschnig, A., Hu, L., et al. (2018). Ancient herders enriched and restructured African grasslands. Nature 561, 387–390. doi: 10.1038/s41586-018-0456-9
Miller, H., Baird, D., Pearson, J., Lamb, A. L., Grove, M., Martin, L., et al. (2019). The origins of nomadic pastoralism in the eastern Jordanian steppe: a combined stable isotope and chipped stone assessment. Levant 50, 281–304. doi: 10.1080/00758914.2019.1651560
Motuzaite Matuzeviciute, G., Lightfoot, E., O’Connell, T. C., Voyakin, D., Liu, X., Loman, V., et al. (2015). The extent of cereal cultivation among the Bronze Age to Turkic period societies of Kazakhstan determined using stable isotope analysis of bone collagen. J. Archaeol. Sci. 59, 23–34. doi: 10.1016/j.jas.2015.03.029
Pearson, J. A., Buitenhuis, H., Hedges, R. E. M., Martin, L., Russell, N., and Twiss, K. C. (2007). New light on early caprine herding strategies from isotope analysis: a case study from Neolithic Anatolia. J. Archaeol. Sci. 34, 2170–2179. doi: 10.1016/j.jas.2007.09.001
Reimer, P. J., Austin, W. E. N., Bard, E., Bayliss, A., Blackwell, P. G., Ramsey, C. B., et al. (2020). The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62, 725–757. doi: 10.1017/RDC.2020.41
Ren, L., and Dong, G. (2016). The history for origin and diffusion of “Six livestock” (in Chinese). Chin. J. Nat. 38, 257–262. doi: 10.3969/j.issn.0253-9608.2016.04.005
Ren, L., Yang, Y., Wang, Q., Zhang, S., Chen, T., Cui, Y., et al. (2020). The transformation of cropping patterns from Late Neolithic to Early Iron Age (5900–2100 BP) in the Gansu–Qinghai region of northwest China. Holocene 31, 183–193. doi: 10.1177/0959683620941137
Ren, L., Yang, Y., Qiu, M., Brunson, K., Chen, G., and Dong, G. (2022). Direct dating of the earliest domesticated cattle and caprines in northwestern China reveals the history of pastoralism in the Gansu-Qinghai region. J. Archaeol. Sci.
Revelles, J., Burjachs, F., Palomo, A., Pique, R., Iriarte, E., and Perez-Obiol, R. (2018). Human-environment interaction during the Mesolithic- Neolithic transition in the NE Iberian Peninsula. Vegetation history, climate change and human impact during the Early-Middle Holocene in the Eastern Pre-Pyrenees. Quat. Sci. Rev. 184, 183–200. doi: 10.1016/j.quascirev.2017.08.025
Richards, M. P., and Hedges, R. E. M. (1999). Stable isotope evidence for similarities in the types of marine foods used by late Mesolithic humans at sites along the Atlantic coast of Europe. J. Archaeol. Sci. 26, 717–722. doi: 10.1006/jasc.1998.0387
Riehl, S., Zeidi, M., and Conard, N. J. (2013). Emergence of agriculture in the foothills of the Zagros Mountains of Iran. Science 341, 65–67. doi: 10.1126/science.1236743
Schwarcz, H. P., Dupras, T. L., and Fairgrieve, S. I. (1999). 15N enrichment in the Sahara: in search of a global relationship. J. Archaeol. Sci. 26, 629–636. doi: 10.1006/jasc.1998.0380
Sealy, J. C., van der Merwe, N. J., Lee Thorp, J. A., and Lanham, J. L. (1987). Nitrogen isotopic ecology in southern Africa: implications for environmental and dietary tracing. Geochim. Cosmochim. Acta 51, 2707–2717. doi: 10.1016/0016-7037(87)90151-7
Shen, H., Zhou, X., Zhao, K., Betts, A., Jia, P. W., and Li, X. (2018). Wood types and human impact between 4300 and 2400 yr BP in the Hexi Corridor, NW China, inferred from charcoal records. Holocene 28, 629–639. doi: 10.1177/0959683617735586
Sherratt, A. (2006). “The Trans-Eurasian exchange: the prehistory of Chinese relations with the West,” in Contact and Exchange in the Ancient World, ed. V. H. Mair (Honolulu: Hawaii University Press), 30–61.
Shi, Z., Chen, T., Storozum, M. J., and Liu, F. (2019). Environmental and social factors influencing the spatiotemporal variation of archaeological sites during the historical period in the Heihe River basin, northwest China. Quat. Int. 507, 34–42. doi: 10.1016/j.quaint.2018.12.016
Shi, Z., Liu, F., Yang, Y., Li, H., Wang, G., Dong, G., et al. (2022). Multiple factors affecting the historical development of agriculture in the Hei River Basin, northwestern China. Environ. Archaeol. doi: 10.1080/14614103.2022.2031837
Song, Y., Chen, G., Wang, H., Fan, X., and Jin, G. (2016). Faunal remains analysis of Xichengyi site of Zhangye City in 2014 (in Chinese). East Asia Archaeol. 13, 233–242.
Spengler, R. N., Miller, N. F., Neef, R., Tourtellotte, P. A., and Chang, C. (2017). Linking agriculture and exchange to social developments of the Central Asian Iron Age. J. Anthropol. Archaeol. 48, 295–308. doi: 10.1016/j.jaa.2017.09.002
Su, P., and Yan, Q. (2008). Stable carbon isotope variation in plants and their indicating significances along the inland Heihe River basin of northwestern China (in Chinese). Acta Ecol. Sin. 28, 1616–1624. doi: 10.3321/j.issn:1000-0933.2008.04.032
Sun, N., Li, X., Zhou, X., Zhao, K., and Yang, Q. (2010). Early smelting recorded by charcoal fossils in Hexi Corridor, Gansu Province–A environmental influence factor (in Chinese). Quat. Sci. 30, 319–325. doi: 10.3969/j.issn.1001-7410.2010.02.08
Svizzero, S. (2016). Hunting strategies with cultivated plants as bait and the prey pathway to animal domestication. Int. J. Res. Soc. Anthropol. 2, 53–68. doi: 10.20431/2454-8677.0202007
Taylor, W. T. T., Pruvost, M., Posth, C., Rendu, W., Krajcarz, M. T., Abdykanova, A., et al. (2021). Evidence for early dispersal of domestic sheep into Central Asia. Nat. Hum. Behav. 5, 1169–1179. doi: 10.1038/s41562-021-01083-y
Vaiglova, P., Reid, R. E. B., Lightfoot, E., Pilaar Birch, S. E., Wang, H., Chen, G., et al. (2021). Localized management of non-indigenous animal domesticates in Northwestern China during the Bronze Age. Sci. Rep. 11:15764. doi: 10.1038/s41598-021-95233-x
van der Merwe, N. J., and Vogel, J. C. (1978). 13C content of human collagen as a measure of prehistoric diet in woodland North America. Nature 276, 815–816. doi: 10.1038/276815a0
van Klinken, G. J. (1999). Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J. Archaeol. Sci. 26, 687–695. doi: 10.1006/jasc.1998.0385
Wallace, M., Jones, G., Charles, M., Fraser, R., Halstead, P., Heaton, T. H. E., et al. (2013). Stable carbon isotope analysis as a direct means of inferring crop water status and water management practices. World Archaeol. 45, 388–409. doi: 10.1080/00438243.2013.821671
Wang, H., Wen, C., and Ai, W. (2021). Brief report on the excavation of Huoshaogou Siba Culture Cemetery in Yumen, Gansu (in Chinese). Archaeol. Cult. Relics 5, 3–21. doi: 10.3969/j.issn.1000-7830.2021.05.002
Wang, H., Zhang, X., Wen, C., and Ai, W. (2019). Brief report on the excavation in 2005 of Huoshaogou site in Yumen, Gansu (in Chinese). Cult. Relics 3, 4–18. doi: 10.13619/j.cnki.cn11-1532/k.2019.03.001
Wilkin, S., Miller, A. V., Fernandes, R., Spengler, R., Taylor, W. T. T., Brown, D. R., et al. (2021). Dairying enabled Early Bronze Age Yamnaya steppe expansions. Nature 598, 629–633. doi: 10.1038/s41586-021-03798-4
Wilkin, S., Miller, A. V., Miller, B. K., Spengler, R. N., Taylor, W. T. T., Fernandes, R., et al. (2020). Economic diversification supported the growth of Mongolia’s nomadic empires. Sci. Rep. 10:3916. doi: 10.1038/s41598-020-60194-0
Xiong, J., Du, P., Chen, G., Tao, Y., Zhou, B., Yang, Y., et al. (2022). Sex-biased population admixture mediated subsistence strategy transition of Heishuiguo people in Han Dynasty Hexi Corridor. Front. Genet. 13:827277. doi: 10.3389/fgene.2022.827277
Yang, L., Shi, Z., Zhang, S., and Lee, H. F. (2020). Climate change, geopolitics, and human settlements in the Hexi Corridor over the last 5,000 years (in Chinese). Acta Geol. Sin. 94, 612–623. doi: 10.1111/1755-6724.14529
Yang, Y. (2017). The transition of human subsistence strategy and its influencing factors during prehistoric times in the Hexi Corridor, northwest China (in Chinese). Ph.D. thesis. Lanzhou: Lanzhou University.
Yang, Y., Dong, G., Zhang, S., Cui, Y., Li, H., Chen, G., et al. (2017). Copper content in anthropogenic sediments as a tracer for detecting smelting activities and its impact on environment during prehistoric period in Hexi Corridor, Northwest China. Holocene 27, 282–291. doi: 10.1177/0959683616658531
Yang, Y., Zhang, S., Oldknow, C., Qiu, M., Chen, T., Li, H., et al. (2019a). Refined chronology of prehistoric cultures and its implication for re-evaluating human-environment relations in the Hexi Corridor, northwest China. Sci. China Earth Sci. 62, 1578–1590. doi: 10.1007/s11430-018-9375-4
Yang, Y., Ren, L., Dong, G., Cui, Y., Liu, R., Chen, G., et al. (2019b). Economic change in the prehistoric Hexi Corridor (4800–2200 BP), north-west China. Archaeometry 61, 957–976. doi: 10.1111/arcm.12464
Zeder, M. A. (2008). Domestication and early agriculture in the Mediterranean Basin: origins, diffusion, and impact. Proc. Natl. Acad. Sci. U.S.A. 105, 11597–11604. doi: 10.1073/pnas.0801317105
Zeder, M. A., and Hesse, B. (2000). The initial domestication of goats (Capra hircus) in the Zagros Mountains 10000 years ago. Science 287, 2254–2257. doi: 10.1126/science.287.5461.2254
Zhang, S., Yang, Y., Storozum, M. J., Li, H., Cui, Y., and Dong, G. (2017). Copper smelting and sediment pollution in Bronze Age China: a case study in the Hexi corrido, Northwest China. Catena 156, 92–101. doi: 10.1016/j.catena.2017.04.001
Zhang, X., Zhang, J., Li, Z., Zhang, L., Chen, G., Wang, P., et al. (2015). A preliminary analysis to the diet of the primitive residents of the Xichengyi site in Zhangye, Gansu (in Chinese). Archaeology 7, 110–120.
Zhao, C. (2005). The main gains and meaning of disinterment of Xihetan archaeological sites (in Chinese). J. NW Univ. 35, 50–51.
Zhao, Z. (2011). New archaeobotanic data for the study of the origins of agriculture in China. Curr. Anthropol. 52, S295–S306. doi: 10.1086/659308
Zhou, X., Li, X., Dodson, J., and Zhao, K. (2016). Rapid agricultural transformation in the prehistoric Hexi corridor, China. Quat. Int. 426, 33–41. doi: 10.1016/j.quaint.2016.04.021
Keywords: Early Bronze Age, Hexi Corridor, stable isotopes, paleodiet, human subsistence, animal husbandry, diversified management
Citation: Qiu M, Li H, Lu M, Yang Y, Zhang S, Li R, Chen G and Ren L (2022) Diversification in Feeding Pattern of Livestock in Early Bronze Age Northwestern China. Front. Ecol. Evol. 10:908131. doi: 10.3389/fevo.2022.908131
Received: 30 March 2022; Accepted: 17 May 2022;
Published: 13 June 2022.
Edited by:
Xinyi Liu, Washington University in St. Louis, United StatesReviewed by:
Rachel Reid, Virginia Tech, United StatesCopyright © 2022 Qiu, Li, Lu, Yang, Zhang, Li, Chen and Ren. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guoke Chen, Y2hlbmd1b2tlMTk4MEBzaW5hLmNvbQ==; Lele Ren, cmVubGVsZUBsenUuZWR1LmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.