
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 11 August 2022
Sec. Population, Community, and Ecosystem Dynamics
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.905309
This article is part of the Research TopicLong-Term Monitoring in Ecology and Evolution: Establishing a Sound Baseline to Help Inform our FutureView all 9 articles
 Eduardo Infantes1*
Eduardo Infantes1* Daire Carroll1
Daire Carroll1 Willian T. A. F. Silva1,2
Willian T. A. F. Silva1,2 Tero Härkönen3
Tero Härkönen3 Scott V. Edwards1,4
Scott V. Edwards1,4 Karin C. Harding1
Karin C. Harding1Detecting changes in population trends depends on the accuracy of estimated mean population growth rates and thus the quality of input data. However, monitoring wildlife populations poses economic and logistic challenges especially in complex and remote habitats. Declines in wildlife populations can remain undetected for years unless effective monitoring techniques are developed, guiding appropriate management actions. We developed an automated survey workflow using unmanned aerial vehicles (drones) to quantify the number and size of individual animals, using the well-studied Scandinavian harbour seal (Phoca vitulina) as a model species. We compared ground-based counts using telescopes with manual flights, using a zoom photo/video, and pre-programmed flights producing orthomosaic photo maps. We used machine learning to identify and count both pups and older seals and we present a new method for measuring body size automatically. We evaluate the population’s reproductive success using drone data, historical counts and predictions from a Leslie matrix population model. The most accurate and time-efficient results were achieved by performing pre-programmed flights where individual seals are identified by machine learning and their body sizes are measured automatically. The accuracy of the machine learning detector was 95–97% and the classification error was 4.6 ± 2.9 for pups and 3.1 ± 2.1 for older seals during good light conditions. There was a clear distinction between the body sizes of pups and older seals during breeding time. We estimated 320 pups in the breeding season 2021 with the drone, which is well beyond the expected number, based on historical data on pup production. The new high quality data from the drone survey confirms earlier indications of a deteriorating reproductive rate in this important harbour seal colony. We show that aerial drones and machine learning are powerful tools for monitoring wildlife in inaccessible areas which can be used to assess annual recruitment and seasonal variations in body condition.
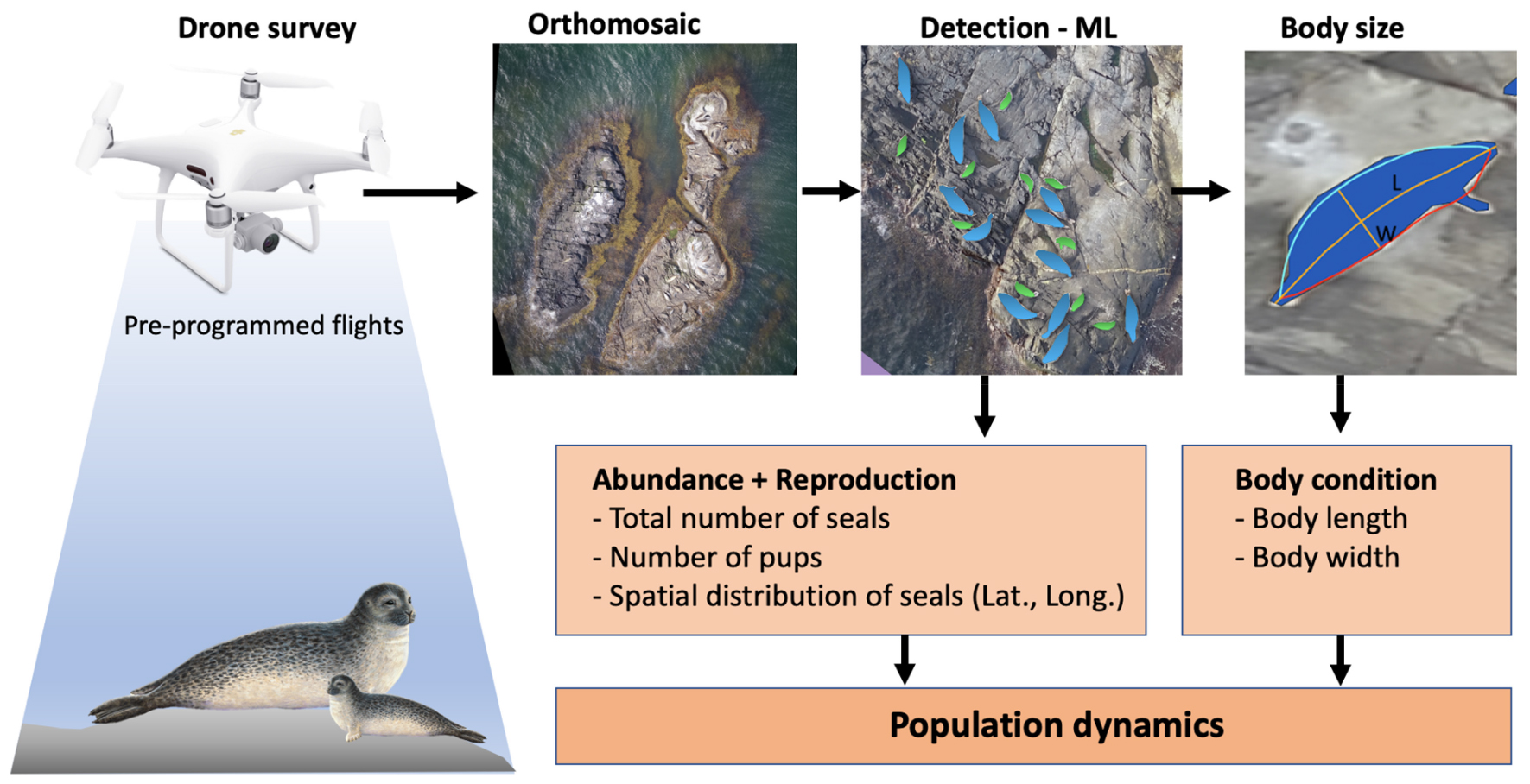
GRAPHICAL ABSTRACT. Overview of the automated work-flow for wildlife surveys, from drone flights to image analysis. Drones are used to collect hundreds of images that are converted into orthomosaics. Thereafter, animals are identified by machine learning (ML) and their body sizes measured.
The rate of population increase is a pivotal parameter in analyses of population dynamics, and monitoring programs are designed to estimate changes in abundance as accurately as possible (Sibly and Hone, 2002; Teilmann et al., 2010). Reliable estimates of population trends are required to implement appropriate management actions, such as hunting or protective measures. Accurate estimates of population parameters are needed for early detection of changes in population growth rate under environmental stress (Harding et al., 2007; Hilde et al., 2020). In vulnerable and declining species, high quality data used to estimate the population rate of increase, or decrease, is vital for informed evaluations of extinction risks by population projection models and for red-list classifications. Another key factor is the length of the time series, where long-term monitoring programs of populations permit assessment of causes and effects and the natural variation in abundance among years (Silva et al., 2021).
Marine mammal populations are sensitive to overexploitation, which can be explained by their limited intrinsic rate of increase and difficulties in assessing changes in population sizes in time (Lotze et al., 2017). As many marine mammal populations today recover from historical low abundances, demands for increased hunting impose an urgent need to develop techniques with shorter lag phases between observation and management actions in order to avoid a new era of overexploitation. The inaccessible habitat and logistic difficulties in monitoring wild marine mammals introduce a sampling error in abundance estimate, which leads to a lag between population decline and detection (Svensson et al., 2011). The Scandinavian harbour seals (Phoca vitulina) have attained historical population sizes after a century of overharvest and epidemics (Silva et al., 2021). They inhabit the Kattegat-Skagerrak coasts and the Baltic Sea (where a smaller population is found), thus they occur in the territorial waters of Sweden, Norway and Denmark. The abundance of harbour seals has been monitored since 1978 by counting the number of seals hauled out on land during peak moult in late August by small aeroplanes.
Different sex and age classes of harbour seals spend different amounts of time on land, for example does subadult seals haul-out more than adult seals, and thus, when the population age structure changes so will the proportion on land (Härkönen et al., 1999). About 65% of the harbour seal population hauls-out on land in the peak moulting season when the population rate of increase is stable (Härkönen et al., 2002; Teilmann et al., 2010). Exponentially growing populations have an inherent “stable age-structure” (Caswell, 2001; Härkönen et al., 2002). However, when the population age structure changes so will the proportion of seals on land. Most populations will eventually be affected by environmental and density-dependent factors, such as decreased food availability and lack of haul-out sites, and the population size will fluctuate (Sibly et al., 2005; Sæther et al., 2016). As the population approaches carrying capacity, population growth declines, age structure shifts to older animals and the average haul-out behaviour may change (Härkönen et al., 1999). In fluctuating populations estimates based on traditional aeroplane surveys during moult, need to be complemented by additional techniques to estimate cohort strength. One limitation of the aeroplane survey is that it mainly monitors cohorts of the population older than 1 year, since pups do not moult the first year and thus spend little time on land during the survey period (Härkönen et al., 2002) thus additional techniques for estimates of annual reproduction are required.
Recent observations show that harbour seal subpopulations in the Kattegat-Skagerrak have different somatic growth rates and population growth rates (Harding et al., 2018; Silva et al., 2021). The causes behind these differences remain unclear but are likely linked to food limitation in some regions. The well documented population of harbour seals in the Kattegat-Skagerrak provides an opportunity to study the mechanisms of population regulation in marine mammals in more detail. Quantifying annual changes in the reproductive rate of females, and the body condition of pups would provide important clues and could disclose cause-effect mechanisms behind observed changes in trends. However, estimating annual pup production and condition of pups is very labour intensive with traditional methods involving catching pups in nets and observing branded seals by telescope in topographically complex archipelagos. Thus, development of new methods are required to complement traditional survey techniques in long-term monitoring, and to increase our understanding of basic processes in the dynamics of populations.
Wildlife surveys using drones are promising techniques currently being developed for monitoring of animal abundance, trends and behaviour for many species (Colefax et al., 2018; Hodgson et al., 2018; Eikelboom et al., 2019; Lyons et al., 2019; Infantes et al., 2020; Gray et al., 2022). Drone observations also provide new possibilities for documenting animal behaviour as well as landscape features, in particular for species sensitive to human activity or aggressive to humans (Inman et al., 2019; Infantes et al., 2020), or at sites that are difficult to access (Dickens et al., 2021; Palomino-González et al., 2021). Although drone surveys collect detailed information rapidly, they still do not solve existing data analysis limitations. In particular, manual counting of animals in imagery is time consuming and inefficient (Linchant et al., 2015; Hodgson et al., 2018). However, novel advances in automated counting using machine learning approaches can help to overcome these inefficiencies (Linchant et al., 2015; Corcoran et al., 2021; Tuia et al., 2022). Automated methods have the potential to reduce the number of animals missed in surveys and the time taken to complete image analysis compared to manual methods, improving both the accuracy and efficiency of detection (Rey et al., 2017; Hodgson et al., 2018). Technological advances in machine-learning automated detection methods, drone platforms and sensors have led to successful detection of individual animals in a wider range of species than previously possible (Linchant et al., 2015; Hollings et al., 2018). Automated detection and counting can speed up population assessments for example in birds (Chabot and Francis, 2016; Hodgson et al., 2018), marine mammals (Seymour et al., 2017; Gorkin et al., 2020) and terrestrial mammals (Chrétien et al., 2016; Kellenberger et al., 2018).
Although automated counting of animal populations is becoming routine, most approaches do not include the measurement of animal body sizes. There are two main methods used to measure body size and body size changes in the wild: capture-mark-recapture of animals or measurements of animal carcasses. Capture-mark-recapture of wild animals is both labour intensive and disruptive to wildlife (Bradshaw et al., 2003) and typically only provides information for a small number of individuals. When measuring carcasses, obviously, more detailed information on body size and condition can be acquired without the risk of causing stress or injury to living animals but samples are not always representative of the population except for during outbreaks of extremely virulent diseases (Harding et al., 2018). Photogrammetry methods for building 2D models of live elephant seals (Shero et al., 2021) or 3D volumetrics of a limited number of free ranging whales (Christiansen et al., 2019) and grey seals, have recently been developed to measure body mass (Alvarado et al., 2020). But simple non-invasive methods for rapidly estimating body size for a large number of individuals have to our knowledge not been performed before.
In this study, we (1) compare the efficiency and accuracy of two drone survey methods using manual and pre-programed flights, relative to land-based telescope surveys to count seal pups; (2) assess the accuracy of aerial drones in combination with automated software for counting and measuring the body sizes of pups and older harbour seals; and (3) evaluate the current pup production in the light of historical monitoring data and discuss the implications of the findings for the dynamics of the population.
The archipelago in Kosterhavet national park, on the west coast of Sweden, is home to a colony of about 3,000 harbour seals (Silva et al., 2021). The population has been surveyed annually during the late summer moult (when a stable and high proportion of the population is on land) by small aeroplanes since 1978 as part of a larger survey of the entire Kattegat-Skagerrak population (Heide-Jørgensen and Härkönen, 1988). These late summer moult surveys provide data for estimates of changes in total population size over time and have recently indicated a lower, but still positive population growth rate than historically observed. To better understand the causes behind this recent decline, we set out to perform a complementary, more detailed survey of the pup production during the early summer reproductive season in 2021. The goal was to critically evaluate several survey methods and to estimate the reproductive output, see (Figure 1).
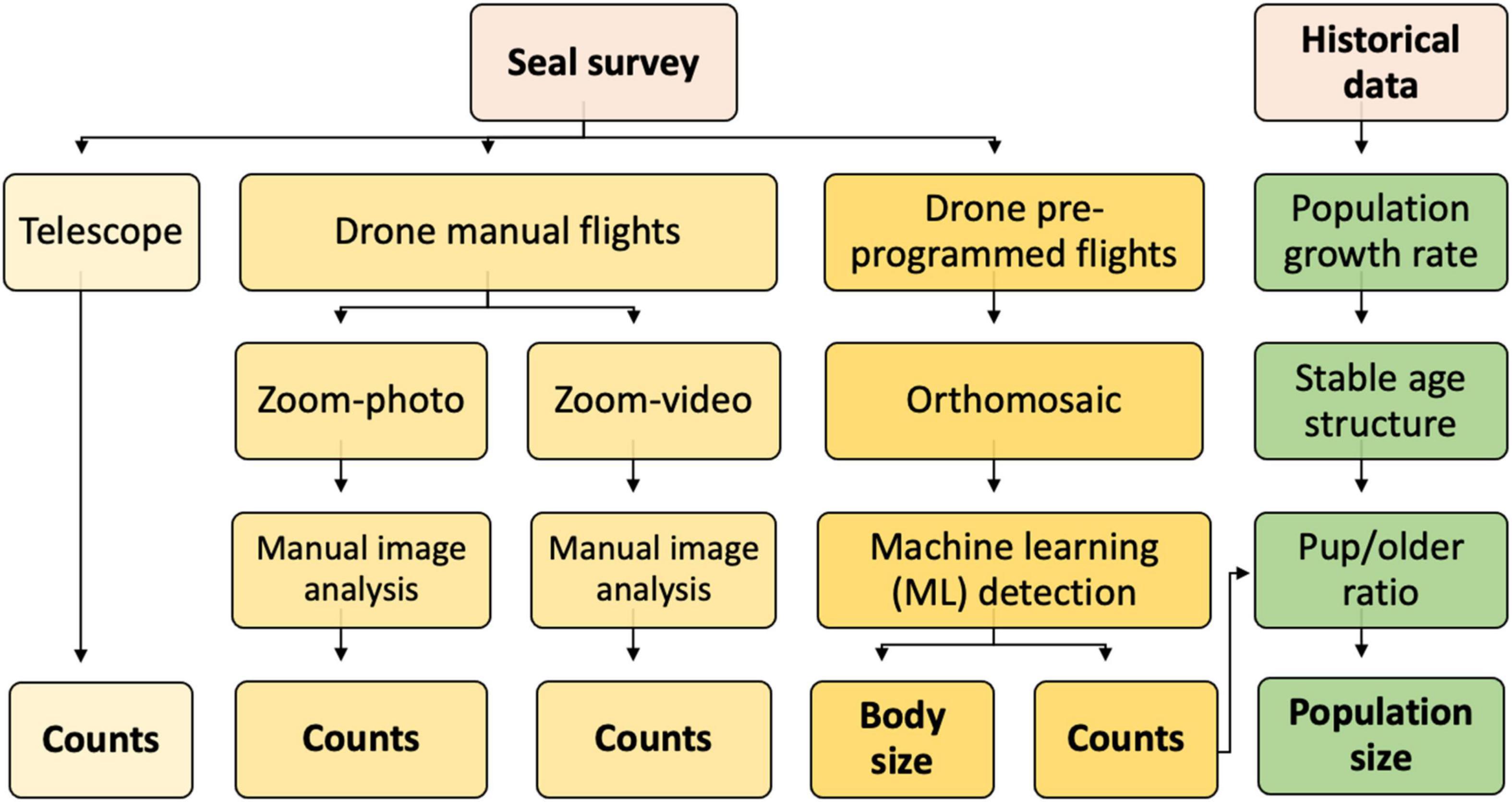
Figure 1. Diagram of the survey methods and workflow tested in the study. Seal surveys were performed both with traditional observations by telescope, and by drones. Drone missions were either manually piloted, or pre-programmed. All methods resulted in estimates of the number of newborn pups, that in turn was evaluated in terms of “pup/older seals ratio” and evaluated in the light of total population size.
We surveyed nine haul-out sites by drones and telescope during the pupping season (June 13–21), and autumn (September 12–15) of 2021, focusing on locations identified as major breeding sites (Figure 2A; Heide-Jørgensen and Härkönen, 1988; Härkönen and Harding, 2001). We visited all nine sites once on each survey day and counted the seals with a telescope and drones, obtaining one sample per site and day with each method. We performed observations between 14:00 and 20:00, because daily seal numbers peak at this time (Heide-Jørgensen and Härkönen, 1988; Härkönen et al., 1999). We identified pups of the year in June by several characteristics, including different colour of pelage compared to older seals, body shape, size and behaviour (Figures 2B,C). In the autumn, we did not attempt to distinguish between pups and older seals.
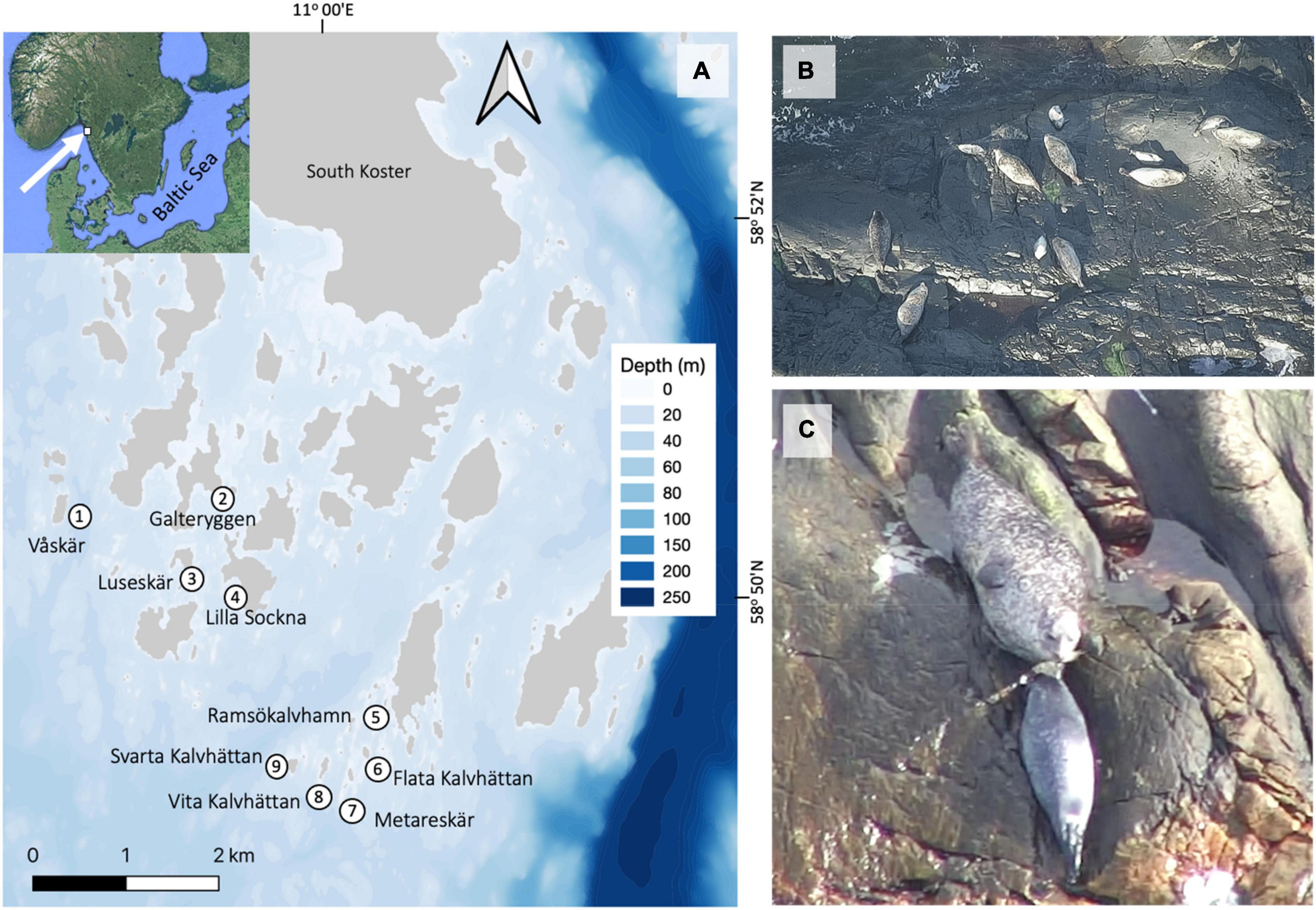
Figure 2. (A) Map of the main haul-out sites for harbour seals in the Kosterhavet archipelago, Sweden. (B) Adult females and pups haul-out during the peak pupping season, and a pregnant female is observed at the bottom left side. (C) Female harbour seals closely watch over their pups during lactation, the first 3 weeks after birth. Mother-pup-pairs can be recognized from drone photos (Images from drone zoom-photo survey of Svarta Kalvhättan, 18th June 2021).
In 2021, we carried out ground-based counts of pups using the same method as in previous surveys, to compare with drone counts. We used a Swarovski ABICHT-AT-80 telescope with flexible magnification up to x60. We observed each site for 20–30 min. and counted pup numbers repeatedly from nearby islets at distances up to 600 m on each survey day. In cases where pups were positioned behind rocks, we attempted to observe the same nursing site from a different location. We surveyed all sites multiple times and took notes of the highest counts for each site. We repeated this method daily until all additional counts were lower than the daily maximum count.
To estimate historical population trends, we provide a new large data set of pup counts by telescope (number of pups) at the nine surveyed sites from the years 1989 to 2013. The intensity of the survey effort using the traditional telescope count was not as high in 2021 as in previous years, since we avoided interfering with the drone survey, and this affected the counts especially at complex habitats and bias the comparison of methods in favour of drone counts. Removing the potential error in historical data by compensating for lower fraction sighted previous years would only strengthen our main finding of a recent decrease in pup production.
We performed manually operated flights remotely from islands without haul-out sites to avoid seal disturbance (Krause et al., 2021). The flight altitude was 100 m and the vertical distance to the seals when photographed by this method was between 100 and 150 m. Observations were made using a x30 optical zoom camera (DJI, Zenmuse-Z30) mounted on a quadcopter drone (DJI, Matrice-200). The optical zoom allowed observers to remain unnoticed by the animals while still obtaining highly detailed photos. The drone streamed live images to the pilot and to an assistant observer with FPV goggles (First Person View, DJI Goggles) with a 1080p/30fps screen definition. This arrangement facilitated the location and identification of pups, allowing the pilot to safely navigate the drone while the assistant focused on observations. The survey was carried out by scanning haul out sites while recording photos and videos with zoom camera from left to right and zooming in on seal groups to document the presence of pups while keeping the drone static. The resulting images and videos were taken at 1,920 × 1,080 pixel resolution. Seal counts were first performed manually on all photos and videos by 3–4 persons with different degrees of experience in seal surveys and the average and standard error was calculated from the observations.
We carried out pre-programmed flights using a DJI Phantom-4Pro to evaluate the use of a low-cost commercial drone (1,500 USD) with a digital RGB camera. We captured aerial photographs in nadir position (looking straight downward) following regular transects in pre-programmed flights using Pix4Dcapture app. at an altitude of 40 m, resulting in an image resolution of 1.09 cm/pixel. We designed transects to take a sequence of photos in an exact grid net with each photo having an 80% forward overlap and 80% lateral overlap. This altitude and flight pattern has been found appropriate for optimally capturing estimates of abundance and occupancy in previous studies (Baxter and Hamilton, 2018) and we observed that the harbour seals remained on land with few signs of being disturbed by the drone at this altitude. All images were geolocated by the internal GPS of the drone. For each site, acquired images (∼100) were combined into a high-resolution image with a corrected perspective (orthomosaic) using Pix4Dmapper (v4.7, Pix4D SA). We obtained ground Control Points (GCPs) from a Google Satellite online map to match positioning as closely as possible to satellite data. On each orthomosaic, we selected 10 GCPs to match the coordinates of the satellite image (Topouzelis et al., 2019).
The image quality of mosaics decreased when flights were carried out in the late afternoon, due to lower light levels, which in turn made images more blurry and decreased the probability of automated detection (Sieberth et al., 2014). Also, if seals were moving on land or water when the drone was collecting images, some images became blurry (ghosting effect). Seal counts were performed manually on all orthomosaics by 3–4 persons with variable experience of seal surveys.
To provide an alternative, automated count of seal numbers, we used Picterra, an online machine learning (ML) platform which uses a Convolutional neural network-based (CNN) architecture for the ML object segmentation. CNN’s are Deep Learning architectures that, among other uses, can identify and outline predefined object classes from raster images through patterns in pixel relations (Ren et al., 2017). This approach is well-suited for identifying individual objects, which are not necessarily identical but share a similar representation in images. We trained Deep Learning models to automate object detection with drone imagery. The software uses a version of the U-NET architecture (Ronneberger et al., 2015), a type of CNN, modified to allow instance segmentation, without the need for a complex, data-hungry instance segmentation model like Mask-R-CNN (He et al., 2017).
We developed three detectors focusing on pups and older seals (age 1 + years) separately in June and on all seals together (age 0 + years) in September (Figure 3). This strategy allowed us to focus our training process on a single element per detector, thereby avoiding confusion between classes. We used the drone orthomosaics to train the software with approx. 100 seals for each class by drawing polygons around the individuals. We included a filter to remove smaller and larger objects of 0.07 and 0.55 m2 to improve performance. Each detector was trained using 4,000 steps. The accuracy of the binary classification model, the F1 score, was estimated as in Csurka and Larlus (2013):
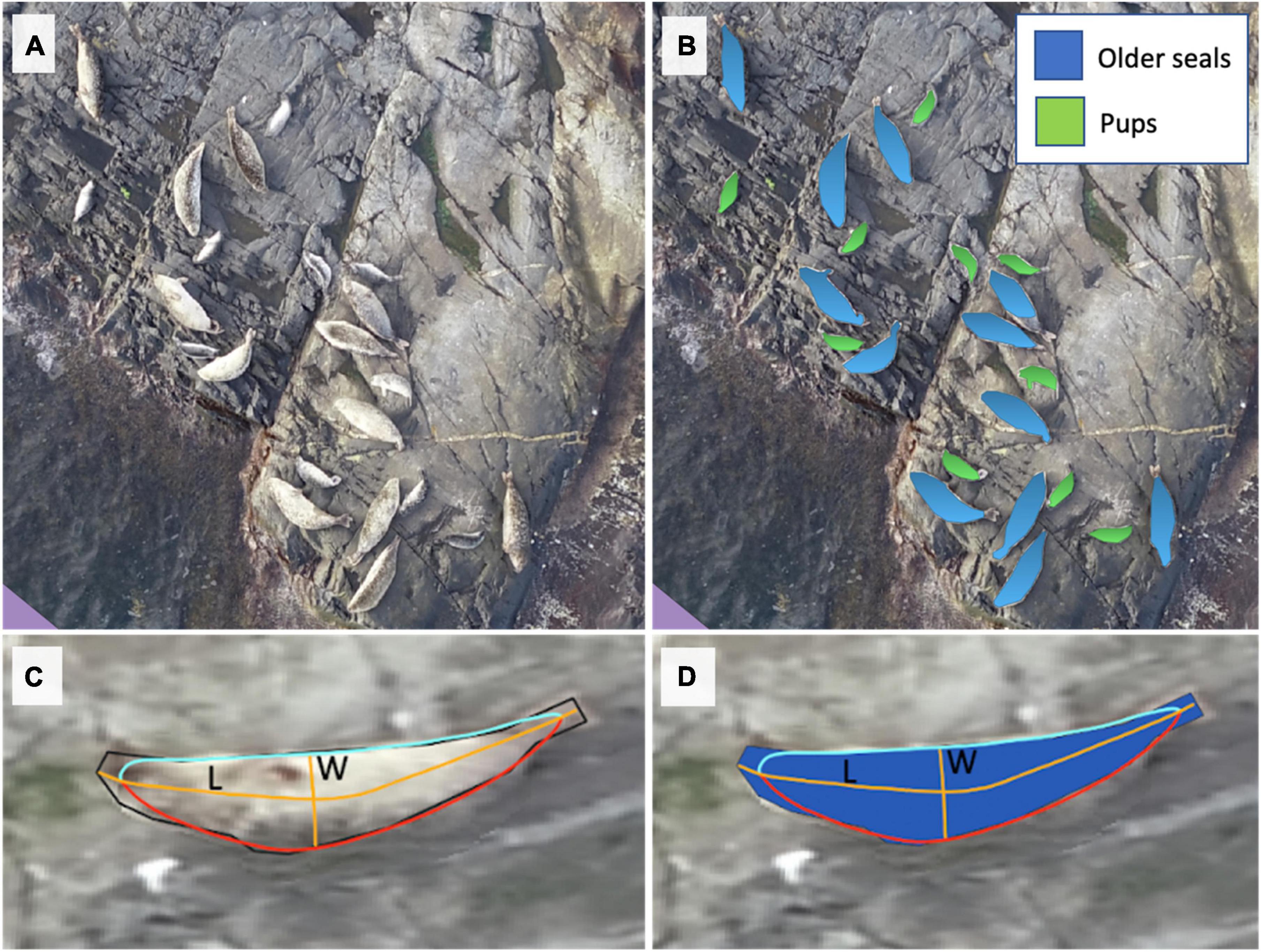
Figure 3. Harbour seal identification using machine learning detectors at the locality “Svarta Kalvhättan,” Sweden, on the 18th of June 2021. (A) A drone orthomosaic map is composed of many overlapping photos. (B) Blue and green polygons indicate pups and older seals identified by machine learning, (C,D) automatic measurement of body lengths (L) and widths (W) describe seal shape. Polygons were smoothed and split into two line segments (red and light blue lines). Coordinate points midway between these segments were used to construct a line representing the position of the spine. To account for the effect of smoothing, the distance between the smooth and unsmoothed polygon was added to the length of this line to calculate seal length (long orange line). Widths were calculated based on the maximum distance between the curved line and the smoothed polygon (short orange line).
Here, “precision” is the ratio between false positives (FP) and false negative (FN) classifications. “Recall” is defined as TP/(TP+FN), where TP is the true positive rate as judged by a human observer. The accuracy of classifications was very high; 97.14% for pups, 97.06% for older seals in June and 95.75% for the group “all seals” in September (Table 1). After running the detectors on the drone mosaics each seal was indicated with a polygon and data on its position (latitude and longitude), body area and perimeter were recorded.

Table 1. Statistics from the automatic seal detecting software.
To calculate seal body sizes we used the polygons classified by the ML for each individual seal. We automated the measurement of seal length and width using a custom written R function (Figure 3C; R Core Team, 2017). Within this function, spatial polygons were plotted and analysed using the packages “sf” and “lwgeom” with further manipulation of the data using package “reshape2” (Wickham, 2007; Pebesma, 2018; Pebesma, 2018). To remove limbs from computation, polygons were smoothed by Gaussian Kernel regression with a bandwidth value of two, using the R-package “smoothr” (Strimas-Mackey, 2020). The two most distant points of the polygon were taken to represent the head and the tip of the tail (n.b. not the tip of hind flippers). Polygons were divided into two-line segments at these points. A set of points midway between corresponding coordinate points in each line segment were used to construct a line which followed the curvature of each polygon. To account for the effect of smoothing, the distance between the farthest points in the smoothed polygon and the original polygon were added to the length of this curved line. This value was taken to correspond to the length of the seal from head to tail following the curvature of its body. Body width was calculated based on the maximum distance between the curved line and the smoothed polygon. All relevant code is available.1
To validate the automatic calculation, we also estimated the standard body length and width of seals manually using classified polygons and mosaics (n = 688). The standard body length of each seal was estimated by measuring the straight-line distance from the head to the tip of the tail (American Society of Mammalogists, 1967). The body width was measured as the longest transversal distance of the seal shaped polygons.
To calculate the error of body size measurements from the drone mosaic, we used a dead harbour seal and 9 panels with known areas of 0.03, 0.12, 0.17, 0.24, 0.29, 0.36, 0.44, 0.58, and 0.79 m2 which were photographed during 3 drone missions. The seal was measured manually at ground level and then from 3 mosaics photographed at 40 m altitude. The manual measurements of the body length and width of the seal were 0.98 and 0.30 m, respectively and the corresponding automatic measurements from the drone were 0.99 ± 0.01 m and 0.32 ± 0.01 m. The area of the panels measured using the drones showed a close match with the manual measurements (F = 2277, p < 0.001, R2 = 0.99, y = 10.5x + 0.0077, see Supplementary Figure 1).
We calculated the population size and rate of increase in the Kosterhavet region retrospectively, based on analysis of annual aerial survey data (2003–2020, data from SHARKweb, n.d.). Following established routines we summed observations from individual Skerries into regions, and calculated the mean of the two highest daily counts from each year, ignoring the lowest count out of three (Teilmann et al., 2010). The proportion of pups in an exponentially growing population is given by the age-dependent demographic rates, and can be analysed by linear algebra in a Leslie matrix framework (Caswell, 2001). The dominant eigenvalue (λ) of the Leslie matrix (L) represents the long term asymptotic growth rate of the population. When a population grows exponentially for many years it assumes a stable age distribution, with a fixed proportion of young born every year (Kendall et al., 2019). Historical data shows that the maximum growth rate for harbour seals is 11% annually (equivalent to: λ = 1.11), and the proportion of newborn pups in a steadily increasing harbour seal population is approximately 20% of the total population size (Härkönen et al., 2002, 2007). These empirically well documented demographic rates are our baseline scenario (Table 2). We also construct an alternative “low growth rate scenario” by manipulating the demographic rates to fit the current population rate of increase.

Table 2. The age-specific demographic rates predicted for a population under different growth conditions.
To estimate the rate of increase, λ, given survey data, we used a non-linear least squares method to fit exponential functions to estimated total population size (P) and observed pup numbers, (P = intercept*eln(λ)*Year). To estimate the expected ratio of pups for given demographic conditions, we assumed 50% of observed seals during moult counts were female and that the proportion of mature females can be calculated from the stable age structure of a Leslie matrix. The age structured population (N) took the form:
Age specific birth rates (B) of female offspring were calculated based on the age specific fertility (F) and survival (S) values reported in Silva et al. (2021) according to:
Age specific birth and survival rates were assembled into a Leslie matrix (L) according to:
We modified this matrix to simulate the observed annual increase of 2.8%, and tweaked demographic rates to mimic commonly observed demographic responses to food limitation in pinnipeds (Kjellqwist et al., 1995); we applied a 50% reduction in fertility, simulating year-skipping, and reduction to the age-specific survival of pups (age 0) and subadult seals (ages 1–3) (Table 2). We estimated the expected proportion of pups given these two demographic scenarios (Table 2) and compared them to earlier records of pup production in the Koster archipelago and to the data from the current study.
We used a generalised mixed-effects model assuming a Poisson distribution (function “glmer” of R package “lme4”) of seal pup counts to determine whether seal pup counting methods differed significantly (Bates et al., 2015). The model included seal pup counts as the response variables and method and haul-out site as independent variables, with observer and day of survey modelled as random effects. We fit exponential models to estimated pup and total population sizes using a non-linear least squares method taking year as the explanatory variable and estimated size as response variable.
We measured the length and width of each identified seal automatically. We tested the agreement between manual and automatic measurement by performing linear regressions for each dimension with automatic measurements as input variable and manual measurements as output variable. To correct for differences in measured values between projection systems, the parameter estimates from this linear regression were used to apply a linear transformation to bring automatically measured seal dimensions in line with manual measurements. We performed data analysis and automated measurement of polygons in R.
All methods readily identified harbour seal pups and older seals; however, each method has different strengths and weaknesses. The highest total number of pups (320) on the 9 sites was observed on the 18th of June using drone zoom-video surveys, and the highest number of pups (66) on a single skerry was observed at the island of Våskär. We found significant differences between the estimates depending on survey method (F = 33.137, p < 2.2e-16) (Figure 4A). Specifically, a post-hoc analysis (Tukey’s) indicated significant differences between drone zoom-video surveys and all other methods with drone video surveys generally resulting in higher counts compared to drone zoom-photo and telescope surveys. No significant difference was found between drone zoom-video and orthomosaic+ML counting. As expected, differences between haul-out sites were also significant (F = 103.968, p < 2.2e-16).
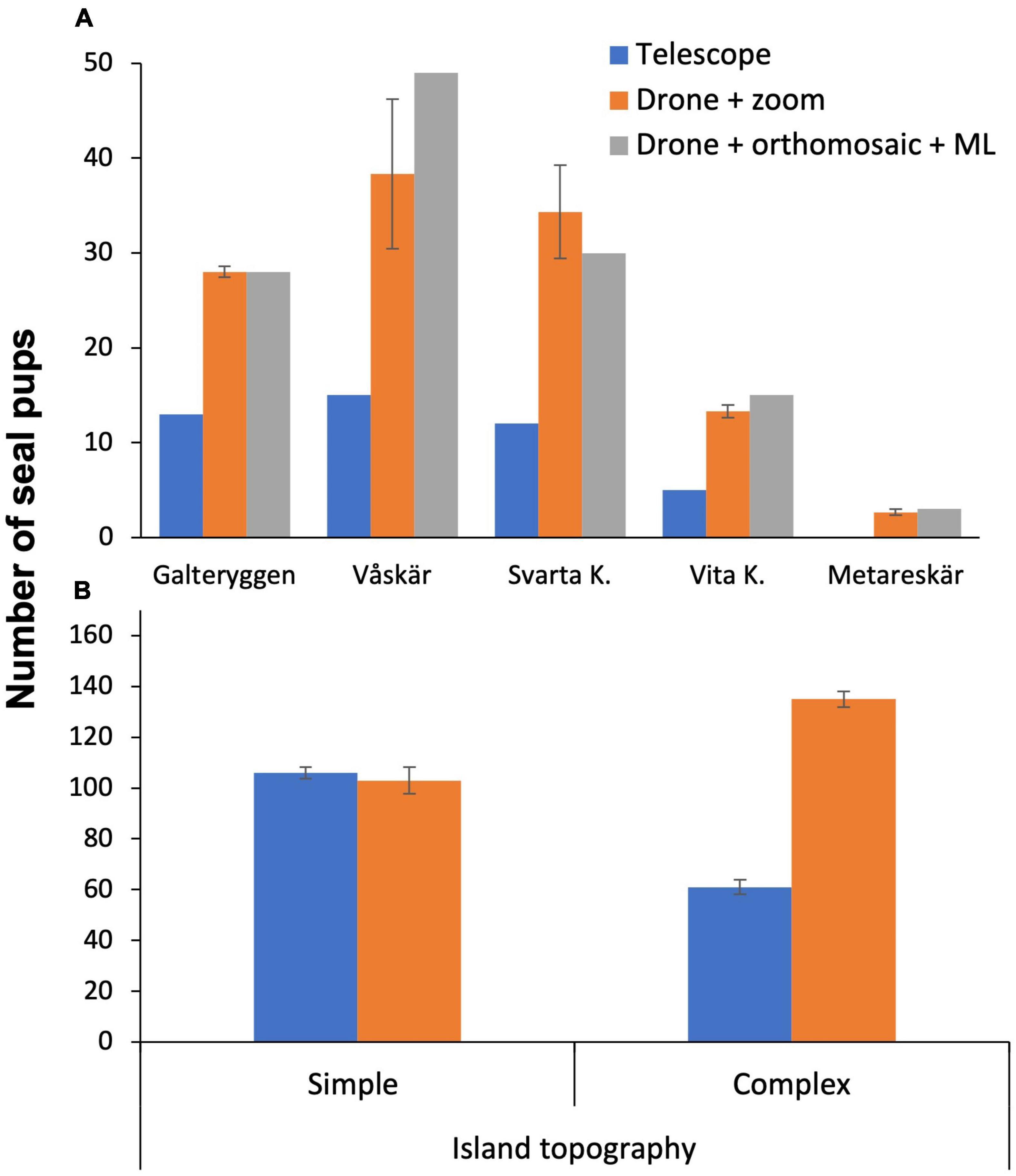
Figure 4. (A) Comparison between survey methods implemented at 5 sites, (B) number of pups counted with telescope and drone at simple and complex island topographies. Seal counts at islands with complex topography showed a significant difference between methods, F = 209.461, p < 0.05. Data collected during 13 and 16 of June 2021.
Counting the total number of pups using drone zoom-photos took 6.4 times longer than zoom-videos, since it was more difficult for the analyst to keep track of the orientation and position within overlapping photos. Counting the total number of pups using zoom-photos took on average 22.4 ± 3 min per site and 201.6 ± 14 min per survey day. On the other hand, a continuous zoom-video across the survey areas took on average 3.5 ± 1 min per site and 31.5 ± 2 min per survey day. In contrast, counting the number of pups using ML was the most efficient approach taking approx. 14 min per site once the detectors were trained (11 min to build the mosaic and 3 min to run the detector). The drone flight time for the zoom-photo, zoom-video and mosaic surveys was similar for all methods (8–10 min per site). Where island topography was simple (sites 2, 3, 4, 5, 7; Figure 2A), there was no difference in the number of pups counted by the drone+zoom and telescopes (Figure 4B). By contrast, when island topography was complex with cracks, large rocks and holes, (sites 1, 6, 8, 9) counts of pups by telescope were 45.5% lower than counts by drone zoom-photo/videos (Figure 4B), although this difference is not expected in the historical telescope surveys that were carried out from more observation points.
On average, seal counting using machine learning failed to identify 13.3 ± 3.8% and 6.4 ± 2.2% of pups and older seals, respectively (Table 3). Orthomosaics were blurry on the 15–16 June due to low daylight late in the evening generating an error of 21.1 ± 6.2 and 7.6 ± 3.6 in the detection of pups and older seals, respectively. However, when orthomosaics were sharp, as on the 18-Jun, the accuracy was higher, with an error of 4.6 ± 2.9 and 3.1 ± 2.1 for pups and older seals, respectively. Swimming seals were not identified by the detector. In addition, seals moving on land appear blurry in the orthomosaic and were not identified by the detector either.
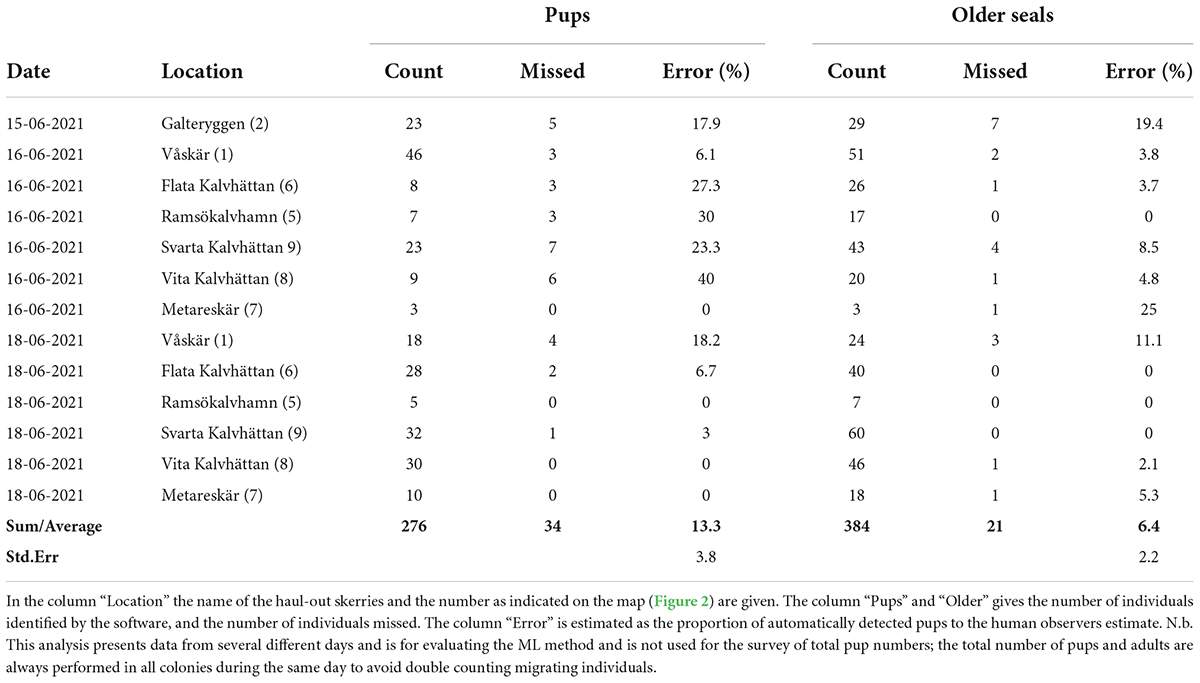
Table 3. Number of pups and older seals identified with machine learning and validated with manual inspection of the digital photos.
In June, the body sizes of pups and older seals (seals that belong to age classes 1 +) can be clearly distinguished in terms of body area, body area perimeter, body maximum length and width (Table 4). We found a significant linear relationship between body lengths measured automatically and manually [F(1,465) = 2956, p < 2e-16, R2 = 0.864, slope = 0.81 mm–1, intercept = 0.16 m], with a residual standard error of 0.103 m (Figure 5A). Similarly, we found a significant linear relationship between the two measurement techniques for width [F(1,465) = 2609, p < 2e-16, R2 = 0.849, slope = 0.91 mm–1, intercept = 0.06 m], with a residual standard error of 0.032 m (Figure 5B). The estimated parameter values from linear regressions comparing manual and automated measurements were used to apply a linear transformation to automatic measurements bringing them in line with manual measurements and establishing body dimensions. Automated measurements of body lengths and widths in June showed a clear distinction between pups (average length of 0.69 ± 0.1 m and width of 0.21 ± 0.04 m) and seals older than 1 year (average length of 1.32 ± 0.19 m and width of 0.38 ± 0.05 m, Figure 5). In September, distinct groupings were not possible and the entire population, including all ages, had an average length of 1.19 ± 0.36 m and width of 0.36 ± 0.04 m (Figures 5A,B). Compared to physical measurements of dead seals, average estimates from the digital photos of live seals are several centimetres shorter, likely due to uneven surfaces below the live seals or lifting of their head or tail (Harding et al., 2018).
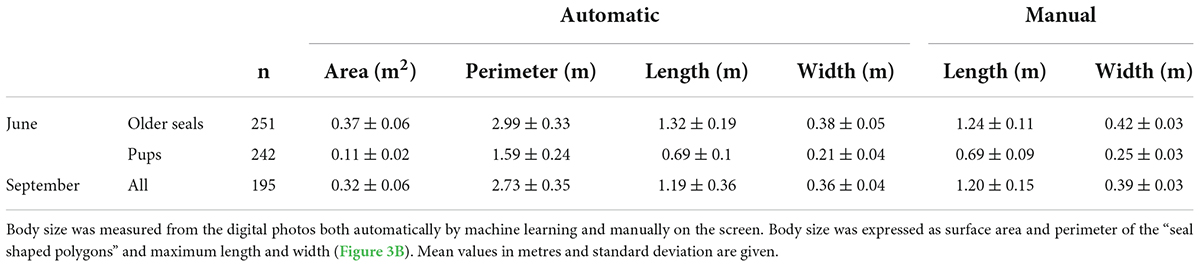
Table 4. Average body sizes of harbour seals hauled out on land calculated from drone orthomosaics during pupping season in June and September 2021.
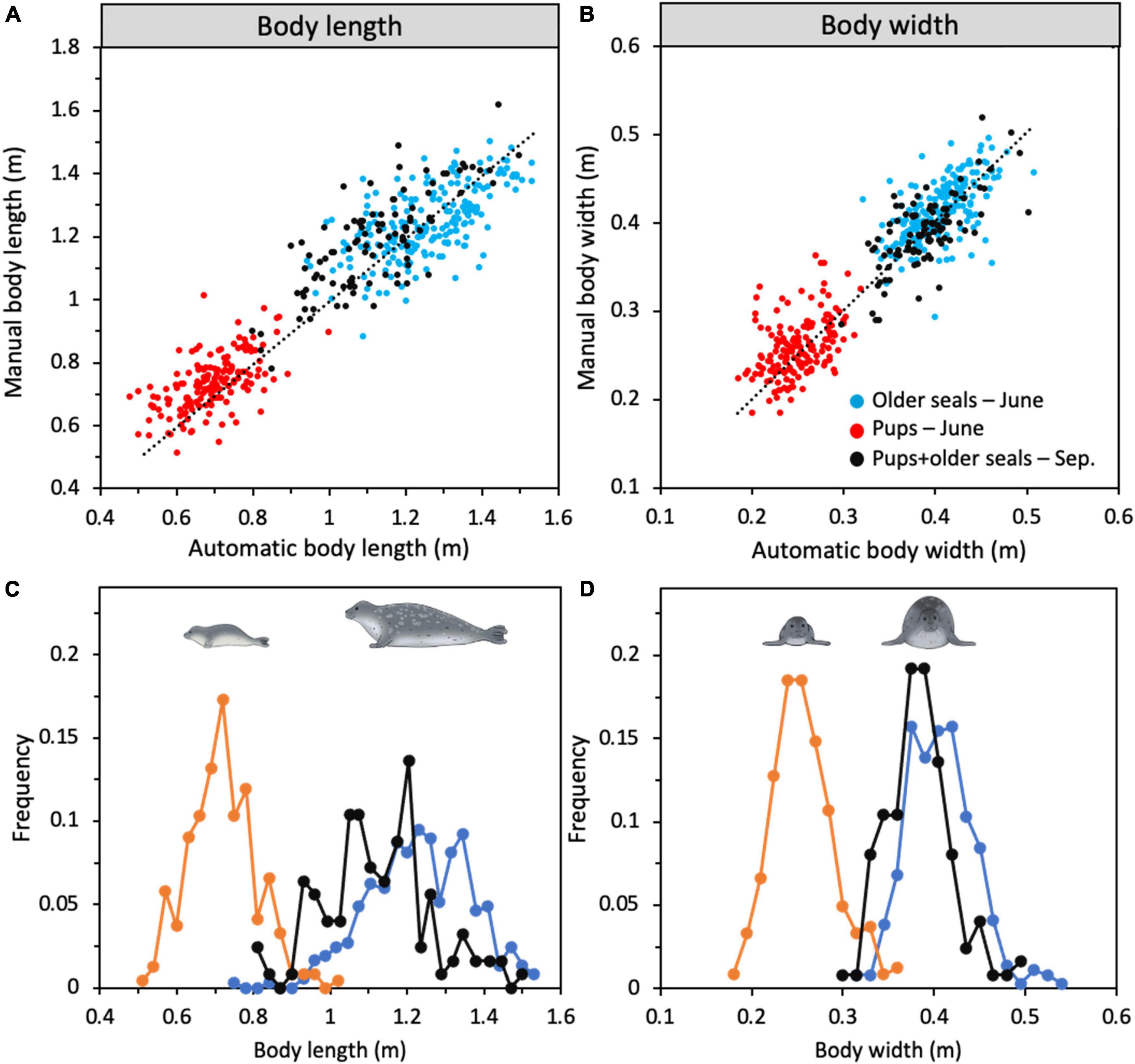
Figure 5. Correlations between manually and automatically performed measurements on digital photos of (A) body length and (B) width (N = 466) of harbour seals. Frequency distribution of (C) body length and (D) body width from the automatic measurements. In June, pups (orange line, N = 242) and older seals (light blue line, N = 251) could be identified as two separate groups on the photos by the machine learning detector (based on e.g., colour and shape) while in September all seals were pooled into the same category (black line, N = 195).
The number of pups in the Kosterhavet archipelago has changed between 1989 and 2021. The maximum pup count has increased from 50 in 1989 up to 320 in 2021 with a maximum of 382 in 2013 (Figure 6A). We assumed that realised growth rate was equal to asymptotic growth rate (λ) and fit separate exponential growth models to survey data for moult counts from 1989 to 2001, and 2002 to 2021 and for pup counts from 1989 to 1997 and from 2012 to 2021 (P = intercept*eln(λ)*Year) (Figure 6 and Table 5). Based on moult surveys, we predicted a λ of 1.028 for the study area between 2002 and 2020 corresponding to an annual increase in population size of 2.8% for the study area (Figure 6). This model was used to estimate a predicted total population of 1,136 in 2021 in the surveyed area. The proportion of pups to the total population estimates, which was increasing prior to 2002, has seen a sharp decrease from 0.43 in 2012 to 0.28 in 2021 (Figure 6B). Historically, the ratio of pups to total counts in the Koster breeding colony has been greater than 0.2. Prior to the last Phocine distemper virus (PDV) outbreak in 2002, this ratio was increasing (Figure 6).
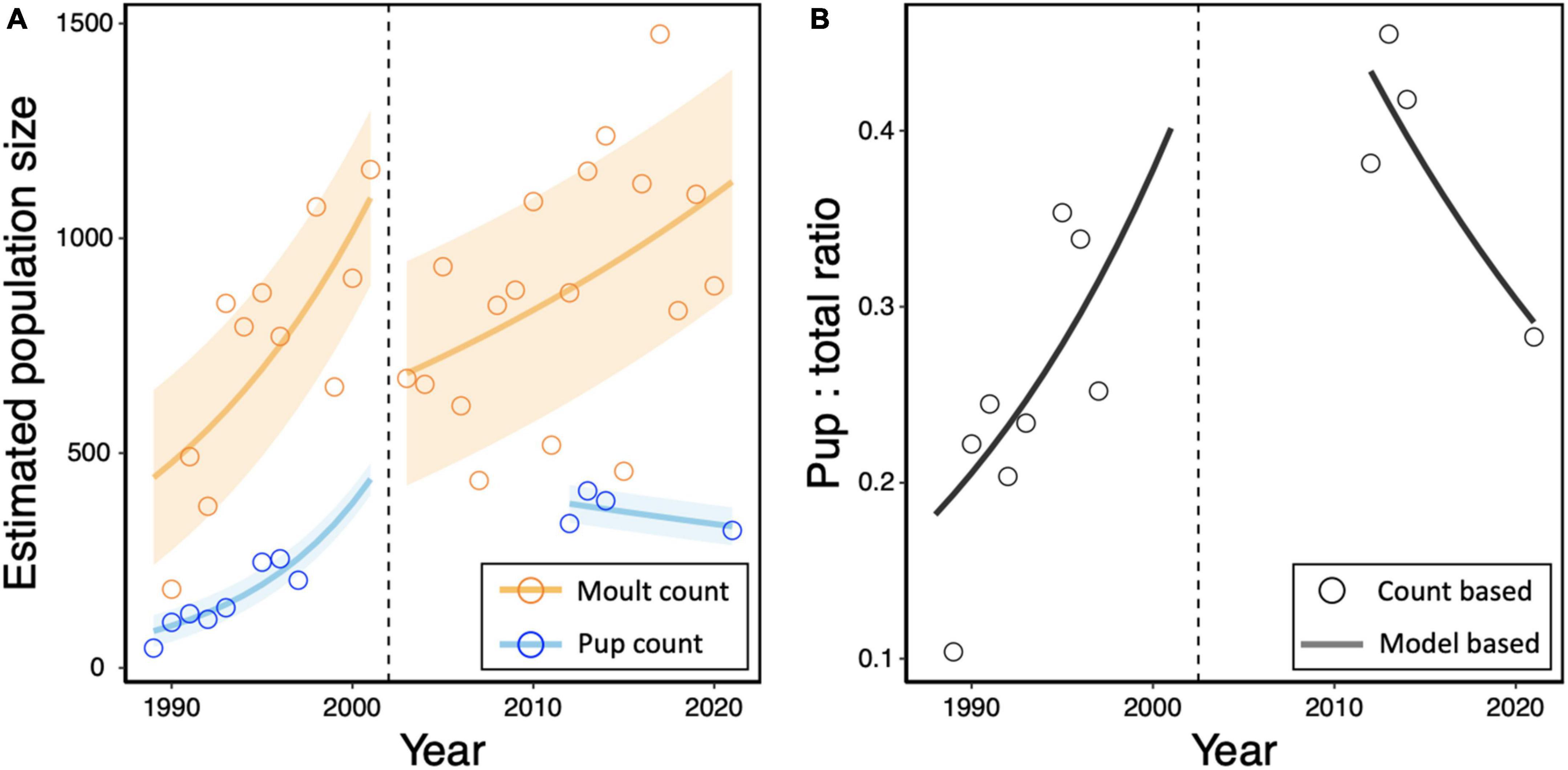
Figure 6. (A) Estimates for total population size of harbour seals in the Koster archipelago are based on August moult counts adjusted for a 65% haul-out rate (orange points). The cumulative maximum number of pups during June pup counts from historical telescope surveys and zoom-video surveys in 2021 were taken to represent 100% of pups (blue points). Separate exponential models were fit to count data for total population size (orange line) and pup numbers (blue line) before and after the year of mass mortality due to Phocine distemper virus (PDV) (dashed line). Shaded regions represent the residual standard error for each model. (B) The ratio of pups to estimated total population is given for annual pup counts (points) and based on the exponential model (line).
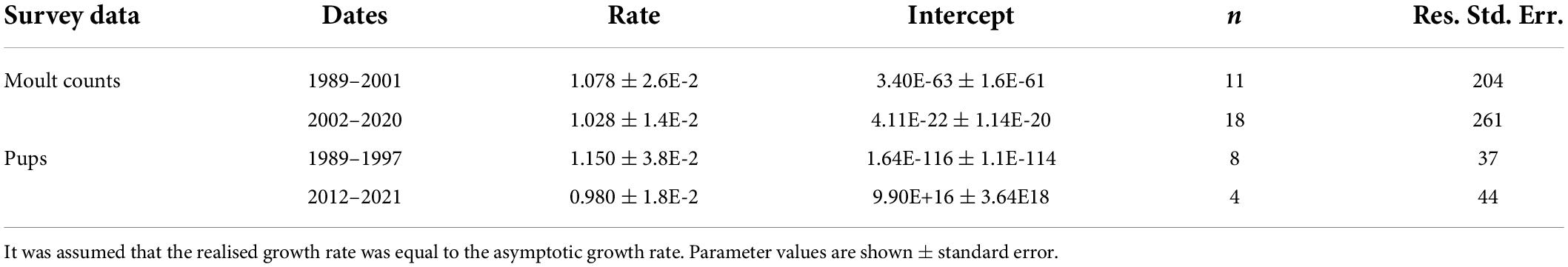
Table 5. Exponential growth models were used to estimate the realised growth rates for moult and pup counts for the time periods before (1989–2001) and after (2002–2021) the most recent Phocine Distemper Virus (PDV) outbreaks.
We investigated scenarios of reduced population growth by parameterising a Leslie matrix for harbour seals. In the baseline scenario, an asymptotic growth rate (λ) of 1.11 is achieved, giving an expected pup/total ratio of 0.20. In the reduced growth scenario, a λ of 1.028 is achieved, giving an expected pup/total ratio of 0.14 (Table 2). Given the observed maximum pup count of 320, we estimated an adult population size of 1,600 and 2,308, assuming λ = 1.11 and λ = 1.028, respectively. It is clear from historical data on age and sex specific haul out patterns that the number of breeding females during pupping is larger than the corresponding population present during moult (Härkönen et al., 1999). Our results indicate that the decreasing proportion of pup to moult count ratio could be accounted for by reductions in fertility and survival of immature seals.
We assess the accuracy and efficiency of different types of telescope and drone surveys at counting and measuring harbour seals during the reproductive season for estimating the annual breeding success. Our results revealed advantages and disadvantages among all methods tested (Figure 7). The most accurate and time-efficient results were provided by performing pre-programmed flights to create orthomosaics, where animals were subsequently identified by machine learning and their body sizes measured automatically. Drone surveys proved to be powerful for monitoring animals in inaccessible rocky archipelagos and could be used to assess annual recruitment and seasonal variations in body condition. This workflow provides high-quality data, permitting early warning signs of changes in population growth and health and could disclose cause-effect mechanisms behind observed population growth trends. Our approach was designed and evaluated for harbour seals but could be applied to other wildlife species or other orthomosaic and machine learning software.
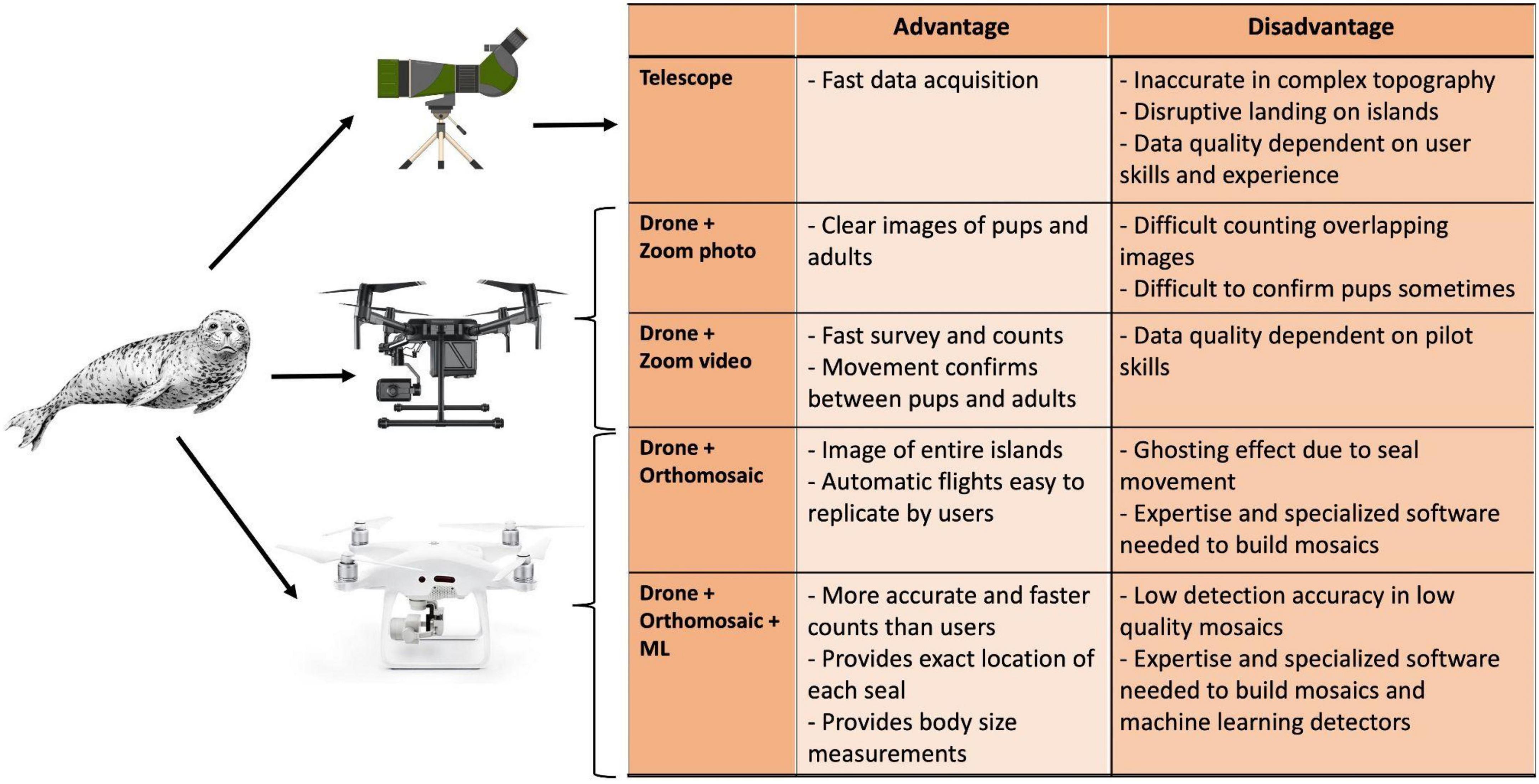
Figure 7. Summary of the advantages and disadvantages for each sampling method to count seal pups by telescope, drone with a photo/video zoom, orthomosaics and machine learning (ML).
This workflow could be easily automated because the software used (e.g., Pix4D, Picterra and RStudio) allows application programming interface (API) integration, which is key to expanding the workflow to operate in a cloud-based service, such as Google Cloud (Bisong, 2019; Shah and Dubaria, 2019). With this approach a user could upload drone images to a cloud server where orthomosaics are built and animals are automatically detected and measured (Malawski et al., 2020). The server can return information on the number of seals and their sizes. Other software with API capabilities can also be used (ArcGISpro, Drone Deploy, DJI-Terra, etc.).
Our results demonstrate that seal pup counts for haul out sites surveyed using drones were 45% higher than when using ground-based telescopes. This difference was influenced by the complexity of the topography, because most Skerries are not flat and seals can be hidden between rocks and thus not easily detected with a land-base telescope. Historical telescope surveys were performed with greater effort including more observation points, however, and can’t be directly compensated by this factor.
Estimating wildlife population size using drones has also been shown to be very effective in other studies. For example, colony nesting bird counts using drones were 43–96% higher (Hodgson et al., 2018), and snow geese counts were 60% higher compared to land-based counts (Chabot and Bird, 2012). Drone counts resulted in 10.6% more hippopotamuses than land-based observations (Inman et al., 2019). In addition, ground-based counting of walruses (pinnipeds) showed a 25% variability between ground-base observers (Udevitz et al., 2005). Nevertheless, telescopes can still provide very important information about the general behaviour of the population because the long observation times (20 min) can be used to identify pups and their behaviour without the need of post-processing. Furthermore, telescope monitoring is less weather-dependent than drones, because land-based counts can be performed even during days with strong winds (>19 m/s) which impede drone flights. A further advantage of telescope surveys is that pups in the water are counted. In summary, four features make counting of harbour seals with drones more accurate and practical than telescope monitoring: (i) access to sites that are not easily visible to the telescope observer; (ii) faster access to target locations; (iii) ability to record images and/or videos that can be used for (re-)counting seals or observe behaviour after surveys, and (iv) the ability to accurately measure seal body sizes.
Drones with zoom-video were shown to be particularly useful for manual estimation of total pup numbers, providing a very fast and effective method both in the field (10 min flight) and during data post-processing (3–5 min per site). However, this method requires a skilled drone pilot to accurately video-scan the colony and a larger and more expensive drone (DJI Matrice+zoom camera, approx. 10,000 USD). Using a drone with zoom-videos can also be useful for documenting marine mammal behaviours (Torres et al., 2018; Inoue et al., 2019; Infantes et al., 2020; Dickens et al., 2021) because it allows longer observations rather than photo snapshots. However, drone photos (orthomosaics) could be used to document population shifts such as changes in haul-out distribution through the day or tidal cycles.
We did not observe any clear disturbance to the seals by the drones at 40 m altitude while collecting orthomosaics or during zoom-photo/video surveys at 100 m altitude. The sizes of the drones used and altitudes of flight have shown to cause low disturbance to marine mammals, such as harbour seals (Duporge et al., 2021; Palomino-González et al., 2021; Krause et al., 2021). Counting pups with telescopes on islands with complex topography requires that observers drive near Skerries by boat and count seals as they escape into the water, which can lead to unnecessary short-term stress both to lactating mothers and pups.
Pre-programmed flights using small consumer drones such as the DJI Phantom-4Pro allow easy replication of survey methods. This trajectory standardisation and low-cost of the drone (1,500 USD) make the method efficient because surveys can be performed by users that are not highly experienced in drone piloting. As a complement to pre-programmed flights, automated detection for animal counting has the advantage of reducing analysis times and can increase the accuracy of wildlife identification in drone images compared to manual counting (Hodgson et al., 2016, 2018).
This study shows that automatic detection using CNN is an efficient technique for seal population surveys. Approximately 100 animals for each class (older seals and pups) were required to train the CNN to automatically detect individuals, instead of larger image datasets (>1000) which are sometimes required for other machine learning methods such as FasterRCNN, YOLO and HOG (Chen and Liu, 2017; Halstead et al., 2018; Corcoran et al., 2021). However, in some surveys the automatic detection method failed to identify a larger fraction of the seals due light conditions (Table 3). Image blur due to low light and animal movement led to a decreased probability of automated detection, as the outlines and features of target species were less clearly distinguishable (Sieberth et al., 2014). In addition, seals swimming in the water or moving on land became blurry (ghosting effect) in the orthomosaic and were not detected by the automated method.
In this study, we also provide a simple, efficient, and non-invasive method to estimate body size by measuring both their structural size (body length) and relative body condition (body width). Measuring the body size of 737 seals took only 5.8 min of computer processing after the seals were identified using machine learning. In comparison, manually measuring body sizes took 13 h. Drones have also been used to estimate the body mass of free living whales by measuring their body length and body width (Christiansen et al., 2019) and to calculate the volume of grey seals (Shero et al., 2021). Here we report a simplification of previous methods; our approach only requires average computing times between 60 and 90 min per surveyed site (orthomosaic construction, running ML detectors and measuring body sizes), while still providing key information on the population.
We tested our method for estimating seal body sizes using a dead seal and a set of panels of known size and found a very good correlation between true and estimated sizes. However, this approach requires further development because the body lengths given in Figure 5 should be treated as proxies given that body lengths for both pups and older seals are underestimated compared with values given in the literature (Härkönen and Heide-Jørgensen, 1990). One possible source of error likely involves orientation of seals while basking: whereas the objects in our test were flat on the ground, seals basking on rocky shores mostly lie on slopes. Because images are taken in nadir any deviation from the horizontal of seals will lead to underestimation of their sizes. This problem will not occur in habitats dominated by sand or mudflats. An additional way to improve future surveys is to change the position where body width is measured from the widest part to instead measure 10 cm below the front flipper, as is performed on captured seals. Such an approach would permit a more stable and repeatable point of measurement.
This study shows that the trend in harbour seal pup numbers in Kosterhavet was positive between 1989 and 1997 and negative between 2012 and 2021. In June of 2021, a maximum daily count of 320 pups was recorded using drones. This suggests that the proportion of pups in the population has declined despite improved methodology (Figure 6A). This result suggests a population under increased stress, resulting in altered demographic parameters (Kjellqwist et al., 1995; Harding et al., 2018). The proposed scenario of reduced population growth explains this trend as the outcome of reproductive skipping and reduced pup and subadult survival, simulating a population under food limitation. This is one of many potential explanations for the observed counts and illustrates how historical monitoring data can be supplemented with advanced survey techniques, providing more informative means of assessing population health at lower costs than traditionally laborious survey methods. Such data can be used as a source of information for integrated population modelling, improving the ability of models to describe past trends and make predictions about future population development (Plard et al., 2019). Additional surveys are required to establish if this trend is consistent over time.
Changes in growth rate in pinniped populations not directly affected by human activities are often caused by variations in access to resources such as food or breeding habitat. Food limitation leads to a sequence of negative effects on vital population parameters, with reproductive output being the first to be affected, followed by increased juvenile mortality, with adult mortality affected last. Somatic growth is also hampered, leading to higher age at first reproduction and year skipping (Bowen et al., 2003). Therefore, pairing a time-series of well-designed surveys assessing breeding success with total population counts can provide a much earlier assessment of the population status than total population counts alone. Where previously the high cost would have prohibited multiple surveys of the same population within a single year, automated methods as presented in this study present a viable option.
The International Council for the Exploration of the Sea (ICES) and the Convention for the Protection of the Marine Environment of the North-East Atlantic (OSPAR) frameworks propose that Good Environmental Status (GES) is achieved for populations at carrying capacity when there is ‘No decline in population size or pup production exceeding 10% occurred over a period up to 10 years’. Our data indicate a dramatic drop in pup production, which is an early warning sign, but the time series needs to be extended to obtain conclusive evidence.
Overall, the use of drones allowed observations that are informative for the management of seal populations and could be integrated into current monitoring practices. Orthomosaic collection can be easily replicated with low-cost drones, by providing high-resolution mosaics (1 cm/pixel) to count pups and older seals as well as documenting their spatial location, which could be important when revisiting colonies. In addition to population surveys, drones with zoom-video cameras can provide new information on seal behaviours, such as mother and pup interactions during lactation, or response to disturbance (Krause et al., 2021). Orthomosaics with the location of each individual older seal and pup can be used to understand how groups are distributed across skerries and islands and the impact of human presence, weather patterns, spread of infectious diseases, etc. This automated workflow can be used to assess the demography and condition of large mammal populations (e.g., pinnipeds, sirenians, cetaceans, etc.) by increasing the statistical power, and provides a new tool for research and population management.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
EI and KCH conceived the ideas, designed methodology, and led the writing of the manuscript. EI, WTAFS, TH, SVE, and KCH collected the data. EI, WTAFS, and DC analysed the data. DC developed the algorithm for automatic estimation of seal body size. All authors contributed critically to the drafts and gave final approval for publication.
We would like to acknowledge Naturvårdsverket (Viltanslaget Dnr 2021-00028), Carl Tryggers foundation, and Swedish Research Council (FORMAS) Dnr:2019-01192. SVE was supported by a King XVI Carl Gustafs Professorship in Environmental Science. We are thankful for the assistance by Collin Wheatley, Chiau Yu Chen, Anders Tysklind, and Anita Tullrot at Kosterhavet National Park. Permit number to work and drone fly on seal colonies at Kosterhavet Dnr:521-14883-2021, County Council of Västra Götaland.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.905309/full#supplementary-material
Alvarado, D. C., Robinson, P. W., Frasson, N. C., Costa, D. P., and Beltran, R. S. (2020). Calibration of aerial photogrammetry to estimate elephant seal mass. Mar. Mammal. Sci. 36, 1347–1355. doi: 10.1111/mms.12714
American Society of Mammalogists (1967). Standard measurements of seals. J. Mammal. 48, 459–462. doi: 10.2307/1377778
Bates, D., Mächler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Baxter, P. W. J., and Hamilton, G. (2018). Learning to fly: Integrating spatial ecology with unmanned aerial vehicle surveys. Ecosphere 9:e02194. doi: 10.1002/ecs2.2194
Bisong, E. (2019). Building machine learning and deep learning models on google cloud platform: A comprehensive guide for beginners. New York, NY: Apress.
Bowen, W. D., Ellis, S. L., Iverson, S. J., and Boness, D. J. (2003). Maternal and newborn life-history traits during periods of contrasting population trends: Implications for explaining the decline of harbour seals (Phoca vitulina), on Sable Island. J. Zool. 261, 155–163. doi: 10.1017/S0952836903004047
Bradshaw, C., Barker, R., Harcourt, R., and Davis, L. (2003). Estimating survival and capture probability of fur seal pup using multistate mark–recapture models. J. Mammal. 84, 65–80. doi: 10.1644/1545-15422003084<0065:ESACPO<2.0.CO;2
Caswell, H. (2001). Matrix population models: Construction, analysis, and interpretation, 2nd Edn. Sunderland, MA: Sinauer Associates Inc.
Chabot, D., and Bird, D. M. (2012). Evaluation of an off-the-shelf unmanned aircraft system for surveying flocks of geese. Waterbirds 35, 170–174. doi: 10.1675/063.035.0119
Chabot, D., and Francis, C. M. (2016). Computer-automated bird detection and counts in high-resolution aerial images: A review. J. Field Ornithol. 87, 343–359. doi: 10.1111/jofo.12171
Chen, C.-H., and Liu, K.-H. (2017). “Stingray detection of aerial images with region-based convolution neural network,” in Proceedings of the 2017 IEEE International Conference on Consumer Electronics – Taiwan (ICCE-TW), Taiwan, 175–176. doi: 10.1109/ICCE-China.2017.7991052
Chrétien, L.-P., Théau, J., and Ménard, P. (2016). Visible and thermal infrared remote sensing for the detection of white-tailed deer using an unmanned aerial system. Wildl. Soc. Bull. 40, 181–191. doi: 10.1002/wsb.629
Christiansen, F., Sironi, M., Moore, M. J., Di Martino, M., Ricciardi, M., Warick, H. A., et al. (2019). Estimating body mass of free-living whales using aerial photogrammetry and 3D volumetrics. Methods Ecol. Evol. 10, 2034–2044. doi: 10.1111/2041-210X.13298
Colefax, A., Butcher, P., and Kelaher, B. (2018). The potential for unmanned aerial vehicles (UAVs) to conduct marine fauna surveys in place of manned aircraft. ICES J. Mar. Sci. 75, 1–8. doi: 10.1093/icesjms/fsx100
Corcoran, E., Winsen, M., Sudholz, A., and Hamilton, G. (2021). Automated detection of wildlife using drones: Synthesis, opportunities and constraints. Methods Ecol. Evol. 12, 1103–1114. doi: 10.1111/2041-210X.13581
Csurka, G., and Larlus, D. (2013). “What is a good evaluation measure for semantic segmentation?,” in Proceedings of the IEEE Transaction Pattern Analysis Machine Intelligent, Meylan. doi: 10.5244/C.27.32
Dickens, J., Hollyman, P. R., Hart, T., Clucas, G. V., Murphy, E. J., Poncet, S., et al. (2021). Developing UAV monitoring of South Georgia and the South Sandwich Islands’ iconic land-based marine predators. Front. Mar. Sci. 8:654215. doi: 10.3389/fmars.2021.654215
Duporge, I., Spiegel, M. P., Thomson, E. R., Chapman, T., Lamberth, C., Pond, C., et al. (2021). Determination of optimal flight altitude to minimise acoustic drone disturbance to wildlife using species audiograms. Methods Ecol. Evol. 12, 2196–2207. doi: 10.1111/2041-210X.13691
Eikelboom, J. A. J., Wind, J., van de Ven, E., Kenana, L. M., Schroder, B., de Knegt, H. J., et al. (2019). Improving the precision and accuracy of animal population estimates with aerial image object detection. Methods Ecol. Evol. 10, 1875–1887. doi: 10.1111/2041-210X.13277
Gorkin, R., Adams, K., Berryman, M. J., Aubin, S., Li, W., Davis, A. R., et al. (2020). Sharkeye: Real-time autonomous personal shark alerting via aerial surveillance. Drones 4:18. doi: 10.3390/drones4020018
Gray, P. C., Larsen, G. D., and Johnston, D. W. (2022). Drones address an observational blind spot for biological oceanography. Front. Ecol. Environ. doi: 10.1002/fee.2472
Halstead, M., McCool, C., Denman, S., Perez, T., and Fookes, C. (2018). Fruit quantity and quality estimation using a robotic vision system. arXiv [Preprint]. arXiv180105560 Cs, doi: 10.3389/frobt.2021.627067
Harding, K. C., Härkönen, T., Helander, B., and Karlsson, O. (2007). Status of Baltic grey seals: Population assessment and extinction risk. NAMMCO Sci. Publ. 6, 33–56.
Harding, K. C., Salmon, M., Teilmann, J., Dietz, R., and Härkönen, T. (2018). Population wide decline in somatic growth in harbor seals—early signs of density dependence. Front. Ecol. Evol. 6:59. doi: 10.3389/fevo.2018.00059
Härkönen, T., and Harding, K. (2001). Spatial structure of harbour seal populations and the implications thereof. Can. J. Zool. 79, 2115–2127. doi: 10.1139/cjz-79-12-2115
Härkönen, T., and Heide-Jørgensen, M.-P. (1990). Comparative life histories of East Atlantic and other harbour seal populations. Ophelia 32, 211–235.
Härkönen, T., Harding, K., Rasmussen, T. D., Teilmann, J., and Dietz, R. (2007). Age- and sex-specific mortality patterns in an emerging wildlife epidemic: The phocine distemper in european harbour seals. PLoS One 2:e887. doi: 10.1371/journal.pone.0000887
Härkönen, T., Harding, K. C., and Lunneryd, S. G. (1999). Age- and sex-specific behaviour in harbour seals Phoca vitulina leads to biased estimates of vital population parameters. J. Appl. Ecol. 36, 825–841.
Härkönen, T., Harding, K., and Heide-Jørgensen, M. P. (2002). Rates of increase in age-structured populations: A lesson from the European harbour seals. Can. J. Zool. 80, 1498–1510. doi: 10.1139/z02-141
He, K., Gkioxari, G., Dollar, P., and Girshick, R. (2017). Mask R-CNN in 2961–2969. Available online at: https://openaccess.thecvf.com/content_iccv_2017/html/He_Mask_R-CNN_ICCV_2017_paper.html (accessed August 28, 2021).
Heide-Jørgensen, M.-P., and Härkönen, T. J. (1988). Rebuilding seal stocks in the Kattegat-Skagerrak. Mar. Mammal Sci. 4, 231–246. doi: 10.1111/j.1748-7692.1988.tb00204.x
Hilde, C. H., Gamelon, M., Sæther, B.-E., Gaillard, J.-M., Yoccoz, N. G., and Pélabon, C. (2020). The demographic buffering hypothesis: Evidence and challenges. Trends Ecol. Evol. 35, 523–538. doi: 10.1016/j.tree.2020.02.004
Hodgson, J. C., Baylis, S. M., Mott, R., Herrod, A., and Clarke, R. H. (2016). Precision wildlife monitoring using unmanned aerial vehicles. Sci. Rep. 6:22574. doi: 10.1038/srep22574
Hodgson, J. C., Mott, R., Baylis, S. M., Pham, T. T., Wotherspoon, S., Kilpatrick, A. D., et al. (2018). Drones count wildlife more accurately and precisely than humans. Methods Ecol. Evol. 9, 1160–1167. doi: 10.1111/2041-210X.12974
Hollings, T., Burgman, M., van Andel, M., Gilbert, M., Robinson, T., and Robinson, A. (2018). How do you find the green sheep? A critical review of the use of remotely sensed imagery to detect and count animals. Methods Ecol. Evol. 9, 881–892. doi: 10.1111/2041-210X.12973
Infantes, E., Cossa, D., Stankovic, M., Panyawai, J., Tuntiprapas, P., Daochai, C., et al. (2020). Dugong (Dugong dugon) reproductive behaviour in Koh Libong, Thailand: Observations using drones. Aquat. Mammal. 46, 603–608. doi: 10.1578/AM.46.6.2020.603
Inman, V. L., Kingsford, R. T., Chase, M. J., and Leggett, K. E. A. (2019). Drone-based effective counting and ageing of hippopotamus (Hippopotamus amphibius) in the Okavango Delta in Botswana. PLoS One 14:e0219652. doi: 10.1371/journal.pone.0219652
Inoue, S., Yamamoto, S., Ringhofer, M., Mendonça, R. S., Pereira, C., and Hirata, S. (2019). Spatial positioning of individuals in a group of feral horses: A case study using drone technology. Mammal. Res. 64, 249–259. doi: 10.1007/s13364-018-0400-2
Kellenberger, B., Marcos, D., and Tuia, D. (2018). Detecting mammals in UAV images: Best practices to address a substantially imbalanced dataset with deep learning. Remote Sens. Environ. 216, 139–153. doi: 10.1016/j.rse.2018.06.028
Kendall, B. E., Fujiwara, M., Diaz-Lopez, J., Schneider, S., Voigt, J., and Wiesner, S. (2019). Persistent problems in the construction of matrix population models. Ecol. Model. 406, 33–43. doi: 10.1016/j.ecolmodel.2019.03.011
Kjellqwist, S. A., Haug, T., and Øritsland, T. (1995) Trends in age-composition, growth and reproductive parameters of barents sea harp seals, Phoca groenlandica’. ICES J. Mar. Sci. 52, 197–208. doi: 10.1016/1054-3139(95)80035-2
Krause, D. J., Hinke, J. T., Goebel, M. E., and Perryman, W. L. (2021). Drones minimize antarctic predator responses relative to ground survey methods: An appeal for context in policy advice. Front. Mar. Sci. 8:648772. doi: 10.3389/fmars.2021.648772
Linchant, J., Lisein, J., Semeki, J., Lejeune, P., and Vermeulen, C. (2015). Are unmanned aircraft systems (UASs) the future of wildlife monitoring? A review of accomplishments and challenges. Mammal. Rev. 45, 239–252. doi: 10.1111/mam.12046
Lotze, H. K., Flemming, J. M., and Magera, A. M. (2017). Critical factors for the recovery of marine mammals. Conserv. Biol. J. Soc. Conserv. Biol. 31, 1301–1311. doi: 10.1111/cobi.12957
Lyons, M. B., Brandis, K. J., Murray, N. J., Wilshire, J. H., McCann, J. A., Kingsford, R. T., et al. (2019). Monitoring large and complex wildlife aggregations with drones. Methods Ecol. Evol. 10, 1024–1035. doi: 10.1111/2041-210X.13194
Malawski, M., Gajek, A., Zima, A., Balis, B., and Figiela, K. (2020). Serverless execution of scientific workflows: Experiments with HyperFlow, AWS Lambda and google cloud functions. Fut. Gen. Comput. Syst. 110, 502–514.
Palomino-González, A., Kovacs, K. M., Lydersen, C., Ims, R. A., and Lowther, A. D. (2021). Drones and marine mammals in Svalbard, Norway. Mar. Mammal. Sci. 37, 1212–1229. doi: 10.1111/mms.12802
Pebesma, E. (2018). Simple features for R: Standardized support for spatial vector data. R J. 10:439. doi: 10.32614/RJ-2018-009
Plard, F., Fay, R., Kéry, M., Cohas, A., and Schaub, M. (2019). Integrated population models: powerful methods to embed individual processes in population dynamics models. Ecology 100:e02715. doi: 10.1002/ecy.2715
R Core Team (2017). R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
Ren, S., He, K., Girshick, R., and Sun, J. (2017). Faster R-CNN: Towards real-time object detection with region proposal networks. IEEE Trans. Pattern Anal. Mach. Intell. 39, 1137–1149. doi: 10.1109/TPAMI.2016.2577031
Rey, N., Volpi, M., Joost, S., and Tuia, D. (2017). Detecting animals in African Savanna with UAVs and the crowds. Remote Sens. Environ. 200, 341–351. doi: 10.1016/j.rse.2017.08.026
Ronneberger, O., Fischer, P., and Brox, T. (2015). “U-Net: Convolutional networks for biomedical image segmentation,” in Medical image computing and computer-assisted intervention – MICCAI 2015 lecture notes in computer science, eds N. Navab, J. Hornegger, W. M. Wells, and A. F. Frangi (Cham: Springer International Publishing), 234–241. doi: 10.1007/978-3-319-24574-4_28
Sæther, B.-E., Grøtan, V., Engen, S., Coulson, T., Grant, P. R., Visser, M. E., et al. (2016). Demographic routes to variability and regulation in bird populations. Nat. Commun. 7:12001. doi: 10.1038/ncomms12001
Seymour, A. C., Dale, J., Hammill, M., Halpin, P. N., and Johnston, D. W. (2017). Automated detection and enumeration of marine wildlife using unmanned aircraft systems (UAS) and thermal imagery. Sci. Rep. 7:45127. doi: 10.1038/srep45127
Shah, J., and Dubaria, D. (2019). “Building Modern Clouds: Using Docker, Kubernetes amp; Google Cloud Platform,” in Proceedings of the 2019 IEEE 9th Annual Computing and Communication Workshop and Conference (CCWC), Las Vegas, NV, 0184–0189. doi: 10.1109/CCWC.2019.8666479
SHARKweb (n.d.). Välj Datatyp och Parameter. Available online at: https://sharkweb.smhi.se/hamta-data/ (accessed December 20, 2021).
Shero, M. R., Dale, J., Seymour, A. C., Hammill, M. O., Mosnier, A., Mongrain, S., et al. (2021). Tracking wildlife energy dynamics with unoccupied aircraft systems and three-dimensional photogrammetry. Methods Ecol. Evol. 12, 2458–2472. doi: 10.1111/2041-210X.13719
Sibly, R. M., and Hone, J. (2002). Population growth rate and its determinants: An overview. Philos. Trans. R. Soc. B Biol. Sci. 357, 1153–1170. doi: 10.1098/rstb.2002.1117
Sibly, R. M., Barker, D., Denham, M. C., Hone, J., and Pagel, M. (2005). On the regulation of populations of mammals, birds, fish, and insects. Science 309, 607–610. doi: 10.1126/science.1110760
Sieberth, T., Wackrow, R., and Chandler, J. H. (2014). Motion blur disturbs – The influence of motion-blurred images in photogrammetry. Photogramm. Rec. 29, 434–453. doi: 10.1111/phor.12082
Silva, W. T. A. F., Bottagisio, E., Härkönen, T., Galatius, A., Olsen, M. T., and Harding, K. C. (2021). Risk for overexploiting a seemingly stable seal population: Influence of multiple stressors and hunting. Ecosphere 12:e03343. doi: 10.1002/ecs2.3343
Strimas-Mackey, M. (2020). smoothr: Smooth and tidy spatial features. Available online at: https://github.com/mstrimas/smoothr
Svensson, C. J., Eriksson, A., Harkonen, T., and Harding, K. C. (2011). Detecting density dependence in recovering seal populations. Ambio 40, 52–59. doi: 10.1007/s13280-010-0091-7
Teilmann, J., Rigét, F., and Harkonen, T. (2010). Optimizing survey design for Scandinavian harbour seals: Population trend as an ecological quality element. ICES J. Mar. Sci. 67, 952–958. doi: 10.1093/icesjms/fsq001
Topouzelis, K., Papakonstantinou, A., and Garaba, S. P. (2019). Detection of floating plastics from satellite and unmanned aerial systems (Plastic Litter Project 2018). Int. J. Appl. Earth Obs. Geoinformation 79, 175–183. doi: 10.1016/j.jag.2019.03.011
Torres, L. G., Nieukirk, S. L., Lemos, L., and Chandler, T. E. (2018). Drone up! quantifying whale behavior from a new perspective improves observational capacity. Front. Mar. Sci. 5:319. doi: 10.3389/fmars.2018.00319
Tuia, D., Kellenberger, B., Beery, S., Costelloe, B. R., Zuffi, S., Risse, B., et al. (2022). Perspectives in machine learning for wildlife conservation. Nat. Commun. 13:792. doi: 10.1038/s41467-022-27980-y
Udevitz, M. S., Jay, C. V., and Cody, M. B. (2005). Observer variability in pinniped counts: Ground-based enumeration of walruses at haul-out sites. Mar. Mammal. Sci. 21, 108–120. doi: 10.1111/j.1748-7692.2005.tb01211.x
Keywords: harbour seals (Phoca vitulina), machine learning (ML), wildlife conservation, population dynamics, somatic growth, marine mammal monitoring, drone monitoring, wildlife management
Citation: Infantes E, Carroll D, Silva WTAF, Härkönen T, Edwards SV and Harding KC (2022) An automated work-flow for pinniped surveys: A new tool for monitoring population dynamics. Front. Ecol. Evol. 10:905309. doi: 10.3389/fevo.2022.905309
Received: 26 March 2022; Accepted: 18 July 2022;
Published: 11 August 2022.
Edited by:
Brett K. Sandercock, Norwegian Institute for Nature Research (NINA), NorwayReviewed by:
Chloé Rebecca Nater, Norwegian Institute for Nature Research (NINA), NorwayCopyright © 2022 Infantes, Carroll, Silva, Härkönen, Edwards and Harding. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eduardo Infantes, ZWR1YXJkby5pbmZhbnRlc0BndS5zZQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.