 Defeng Bai
Defeng Bai Xinru Wan
Xinru Wan Li Zhang
Li Zhang Ahimsa Campos-Arceiz
Ahimsa Campos-Arceiz Fuwen Wei
Fuwen Wei Zhibin Zhang
Zhibin Zhang- 1State Key Laboratory of Integrated Management of Pest Insects and Rodents, Institute of Zoology, Chinese Academy of Sciences, Beijing, China
- 2University of Chinese Academy of Sciences, Beijing, China
- 3Key Laboratory for Biodiversity Science and Ecological Engineering, Ministry of Education, College of Life Sciences, Beijing Normal University, Beijing, China
- 4Southeast Asia Biodiversity Research Institute and Center for Integrative Conservation, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Mengla, Yunnan, China
- 5Key Laboratory of Animal Ecology and Conservation Biology, Institute of Zoology, Chinese Academy of Sciences, Beijing, China
- 6CAS Center for Excellence in Biotic Interactions, University of Chinese Academy of Sciences, Beijing, China
Recently, the northward movement of Asian elephants (Elephas maximus) in Yunnan, China, has attracted international attention. Climate change or human disturbances have been proposed to be the key drivers, but these hypotheses have not been rigorously tested. In this study, we quantified the relationship between climate change and human impacts on the recent range expansion of Asian elephants in southwest China. We found that the first observation probability of this species in a new place during 1959–2021 had a significant and positive association with change in air temperature and human density, resulting in a movement toward a high-latitude region with a warmer climate and higher human density; however, its association with precipitation was scale-dependent in time: positive or negative during the past 10 or 5 years, respectively. Under the enforced protection policy, human-dominated areas became preferred habitats for elephants. Our results indicate that climate change and enforced protection efforts in human-dominated landscapes in the last few decades are significant drivers of the recent range expansion of Asian elephants in Yunnan, China. It is necessary to expand the current protected areas or habitat corridors toward the north or set up new reserves in the north and set up barriers between human settlements and elephant habitats to facilitate elephant movements and minimize human-elephant conflicts under accelerated global change.
Introduction
Many mammal species have become extinct, or their populations have rapidly declined due to accelerated climate change and human disturbances (Cardillo et al., 2005; Lister and Stuart, 2008; Ripple et al., 2015). However, populations of some mammal species have recovered from the brink of extinction and recolonized parts of their former range, such as the giant panda (Ailuropoda melanoleuca) in China (Huang Q. Y. et al., 2020), bison (Bison bison) in the United States (Meagher, 1989) and European bison (Bison bonasus) in Poland (Kowalczyk et al., 2013). The recolonization of these endangered species often brings new challenges to society, such as increased human-wildlife conflicts (Marucco and McIntire, 2010). Therefore, understanding the causes and mechanisms of population expansion of endangered species is crucial for making a better conservation strategy.
Climate change has been shown to affect recent global distribution changes of a broad range of mammalian species (Walther, 2001; Walther et al., 2002; Chen et al., 2011). Climate warming can promote the range expansion of a variety of mammals, such as white-tailed deer (Odocoileus virginianus) in North America (Dawe et al., 2014; Dawe and Boutin, 2016), red fox (Vulpes vulpes) in Canada (Hersteinsson and Macdonald, 1992), and small mammals, including some rodent species in North America (Brown et al., 1997; Moritz et al., 2008; Rubidge et al., 2011). Warm conditions were favorable for the range expansion of Asian elephants (Elephas maximus) in ancient times (Vidya et al., 2009; Girdland-Flink et al., 2018). Human disturbances could shape the geographic range size of terrestrial mammals globally by altering their habitats (Pimm et al., 2014; Di Marco and Santini, 2015; Crees et al., 2016). Anthropogenic activities changed the land use type and degraded the suitable habitat of mammal species (Bintliff, 2005; Scanes, 2018). Human stressors are important drivers causing the local extinction of mammal species (Di Marco et al., 2013; Sandom et al., 2014; Wan et al., 2019). There is evidence that anthropogenic disturbances are partially responsible for the local or total extinction of many large mammals, such as Eurasian steppe bison (Bison priscus) and wild horse (Equus ferus) from Eurasia and North America (Lorenzen et al., 2011), the woolly mammoth (Mammuthus primigenius) in Eurasia and North America (Nogués-Bravo et al., 2008), wooly rhinoceros (Coelodonta antiquitatis) and horses in the east and west Eurasia (Wan and Zhang, 2017), giant pandas in China (Zhao et al., 2013), and Asian elephant, gibbons, macaques, musk deers, rhinoceroses, tiger (Panthera tigris) and water deer (Hydropotes inermis) in China (Wan et al., 2019).
It is essential to disentangle the distinct effects of climate change and human impacts to conserve endangered species (Isaac and Cowlishaw, 2004; Crees et al., 2016; Wan et al., 2019). However, quantitative assessments on the effects of climate change and human impacts (i.e., land use change, habitat destruction, and carbon emission) on species range shift are still rare (Wan et al., 2019). It is still challenging to separate the distinct effects of climate change and human activities, which are often highly correlated during past centuries (Easterling et al., 2000; Christidis et al., 2011; Wan et al., 2019). However, spatio-temporal data on animal distribution could provide an opportunity to solve the collinearity issues (Wan et al., 2019; Bai et al., 2022).
Asian elephants are currently endangered. They are distributed in 13 countries, including China (Jiang et al., 2016; Williams et al., 2020). The northern range of Asian elephants in China reached the Yellow River Basin ∼3,000 years ago (Li et al., 2012), but now they are restricted only to small parts of Yunnan, China, including Xishuangbanna, Lincang, and Pu’er Prefectures (Zhang et al., 2015; Zhao and Jin, 2018). After 1990, government conservation policies for elephants in Yunnan were implemented, such as establishing a routine monitoring program and a compensation mechanism for human-elephant conflicts (He et al., 2020). Under the Chinese government’s effective conservation policies, the population has grown from approximately 150 individuals in 1976 to about 200 individuals in 2006 and around 300 elephants in 2018 (Chen, 2006; Zhang, 2018; Zhao and Jin, 2018). Asian elephants started to recolonize their northern historical range in 1995, likely with an increase in population size (Liu et al., 2017; Huang C. et al., 2020). Human-elephant conflicts have been increasing due to their population expansion as well as land-use expansion by people during the past half-century in Yunnan Province (Xu et al., 2014; Chen et al., 2016; Liu et al., 2017). Recently, a group of 15 Asian elephants moved about 500 km from the Mengyang area in March of 2020 to the suburban areas of Kunming in early June 2021 (Campos-Arceiz et al., 2021; Wang H. et al., 2021). The northward movement of elephants has garnered widespread attention. They caused massive damage to local residents’ crops and buildings when they entered the densely populated urban areas (Supplementary Figure 1). There is an urgent need to understand the key drivers and underlying mechanisms causing the population expansion of Asian elephants in Yunnan and then to take concrete and effective measures for better management and conservation.
There are various speculations on the potential factors causing the recent range expansion (re-occupation of its original distribution area) of the Asian elephant in Yunnan, including population increase, habitat destruction, rubber plantation, climate warming, drought, changes in the behavior of Asian elephants and people toward each other, or dense forest canopy due to better conservation (Huang et al., 2019; Campos-Arceiz et al., 2021; Chen et al., 2021; Wang H. et al., 2021). However, these hypotheses have rarely been rigorously tested using data or experiments. During the past decades, various Asian elephant conservation efforts, such as insurance schemes, cross-border conservation, actions, and corridor construction, have been initiated and implemented in Yunnan (Pan et al., 2009; Chen et al., 2013; Wang L. et al., 2021). There are plenty of presence records (i.e., when and where elephants were noticed) of Asian elephants, such as human-elephant conflict events (Chen et al., 2016), field survey records (Sun et al., 2021), and social questionnaire survey records (Yang, 2017). These data can be edited into spatio-temporal distribution data, allowing us to evaluate the key factors associated with the recent expansion of Asian elephants. In this study, by using historical data extracted from the published literature, we reconstructed the spatio-temporal distribution change of Asian elephants in Yunnan province from 1959 to 2021. We quantified the associations between climate change (air temperature, precipitation) and anthropogenic stressors (represented by human population density) with the recent range expansion of Asian elephants in China. We proposed the following three hypotheses to explain the recent range expansion of Asian elephants in Yunnan:
(1) Hypothesis I: If the recent range expansion of Asian elephants is driven by climate warming, the probability of first observation (the first record of presence) of the Asian elephant in a place at a specific year should be positively associated with the change of air temperature of the place during past 5 or 10 years.
(2) Hypothesis II: If the recent range expansion of Asian elephants is driven by precipitation, the probability of the first observation of an Asian elephant in a place in a specific year should be positively associated with the change in precipitation of the place during the past 5 or 10 years due to habitat or food improvement by precipitation.
(3) Hypothesis III: If human disturbances drive the recent range expansion of Asian elephants due to habitat destruction, the probability of the first observation of Asian elephant in a place at a specific year should be negatively associated with the human population density of the place during the past 5 or 10 years.
Materials and methods
Presence data of Asian elephants
We collected the distribution data of Asian elephants from 1959 to 2021 by searching the published literature on the ISI Web of Knowledge,1 Google Scholar,2 and CNKI.net.3 “Asian elephant,” “Ya Zhou Xiang (亚洲象, in Chinese),” and “Elephas maximus” were used as keywords for literature retrieval in CNKI.net. “Asian elephant,” “China,” and “Elephas maximus” were used as keywords in the ISI Web of Knowledge and Google Scholar. Online databases from the Global Biodiversity Information Facility (GBIF Secretariat, 2021 (4 November 2021) GBIF Occurrence Download)4 and iNaturalist5 with occurrence records of Asian elephants were also searched using “Asian elephant” and “Elephas maximus” as keywords. We also collected news that reported the presence of Asian elephants on Baidu News6 and Google7 using “Ya Zhou Xiang (亚洲象, in Chinese)” as a keyword, and all news reports before October 2021 were recorded for further analysis (Supplementary Table 1, Data 1). For all presence records from the literature and news reports, three types of time resolutions (specific year records, records not designated to a specific year but within one decade, and unclear records without time information) and six spatial resolutions (specific latitude and longitude locations, presence records with a village name recorded, presence records with a presence point map, presence records with detailed small location descriptions but without the name of the village recorded, records with a spatial resolution larger than a village, and unclear records without spatial information) were classified (Supplementary Table 2). The latitude and longitude of records with the name of a village or detailed small location description and presence point map were represented by the coordinates of a point within the village or location and the relative position of each point on the map, respectively. Detail descriptions for reconstructing presence records of Asian elephants can be found in the Supplementary material.
Only records with both high-resolution space and time were designated into grid cells of 5 × 5 km2 in Yunnan and used for statistical analysis. Presence records with places that can be transformed into coordinates were defined as “records with high-resolution space,” such as presence records with latitude and longitude information, a clear name of the village, a presence point map, or other small locations. The coordinates of villages, small detailed locations, and presence point maps were calculated in ArcGIS software (version 10.8). Presence records with location resolution larger than a village, or without spatial information, or other records not being designated into specific latitudes and longitude were excluded (Supplementary Table 2). Presence records with specific year information were defined as “records with high-resolution time.” Records with decades or unclear time resolution were excluded (Supplementary Table 2).
In this study, we proposed a method to evaluate associations of the recent range expansion with climate and environmental variables based on historical records of the first observations of elephants in a new place in a specific year. In order to measure the recent range expansion of Asian elephants in Yunnan, the first recorded time (year) in a grid cell was defined as the first observation time (year) (Figure 1A). The last recorded time (year) with the presence of Asian elephants in a grid cell was defined as the previous observation time (year) (Figure 1B). The first observation time and last observation time in each grid cell were divided into 5-year categories for seeing the trend in both time and space scales and visualized in ArcGIS software (version 10.8) (Figure 1). In this study, the first observation time (year) of Asian elephants in one grid cell was used to approximate the actual time when Asian elephants first appeared in each grid cell. The maximum average temperature in the first observation year was calculated as the threshold of climate pressure for the recent range expansion of Asian elephants from 1959 to 2021.
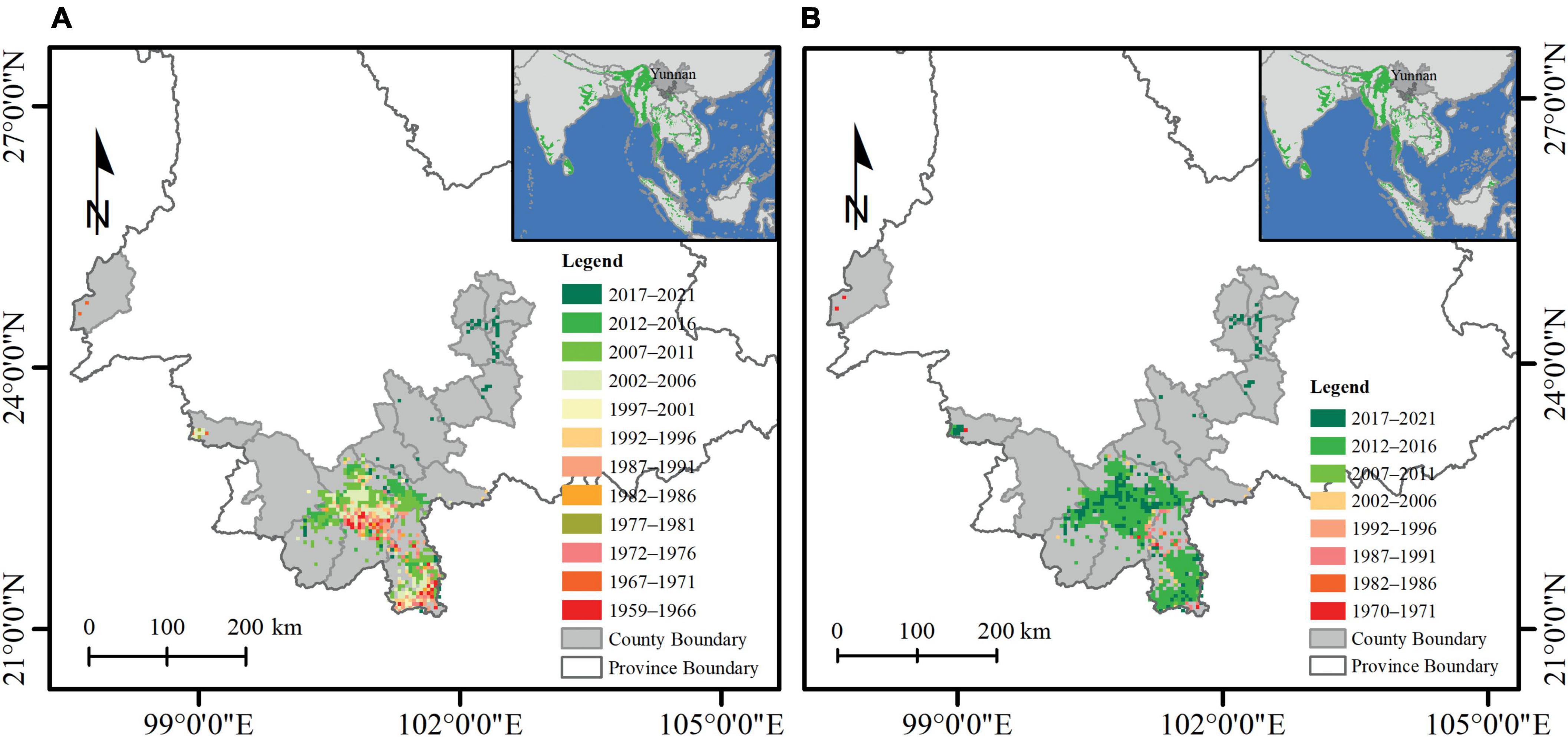
Figure 1. First observation time (year) (A) and last observation time (year) (B) of Asian elephants in Yunnan, China, during 1959–2021. The first observation time was defined as the year with the first presence record of an Asian elephant in the grid cell (5 × 5 km2). The last observation time was defined as the year with the last presence record of an Asian elephant in the grid cell (5 × 5 km2). The first observation time (year) in these grid cells was divided into 12 categories with a 5-year interval, from 1959 to 2021, showing the spatial-temporal change of distribution of elephants. The last observation time (year) in these grid cells was divided into eight categories with a 5-year interval from 1970 to 2021. The red and green colored grid cells represent earlier or later records of this study. The gray-colored background represents the county with the presence of records of Asian elephants in Yunnan province. The green-colored background on the top right panel depicts the recent IUCN distribution of Asian elephants (Williams et al., 2020).
In Figure 1A, the distribution points extracted from the literature were too extensive and overlapped with each other to see the big change in the distributions of elephants in time. Thus, to illustrate the recent range expansion of elephants in latitude and longitude, we divided the whole study period (1959–2021) into two categories (1959–1990 and 1991–2021) because the cumulative number of grid cells of the first observation of Asian elephants in China started to increase sharply around 1990 (Figure 2A). Then, we compared the average latitude or longitude difference between 1959–1990 and 1991–2021. The normality of data was tested using the Shapiro and Wilk, 1965 test. The Mann-Whitney U test was used to examine the latitude or longitude difference between 1959–1990 and 1991–2021 because all data did not conform to normal distribution.
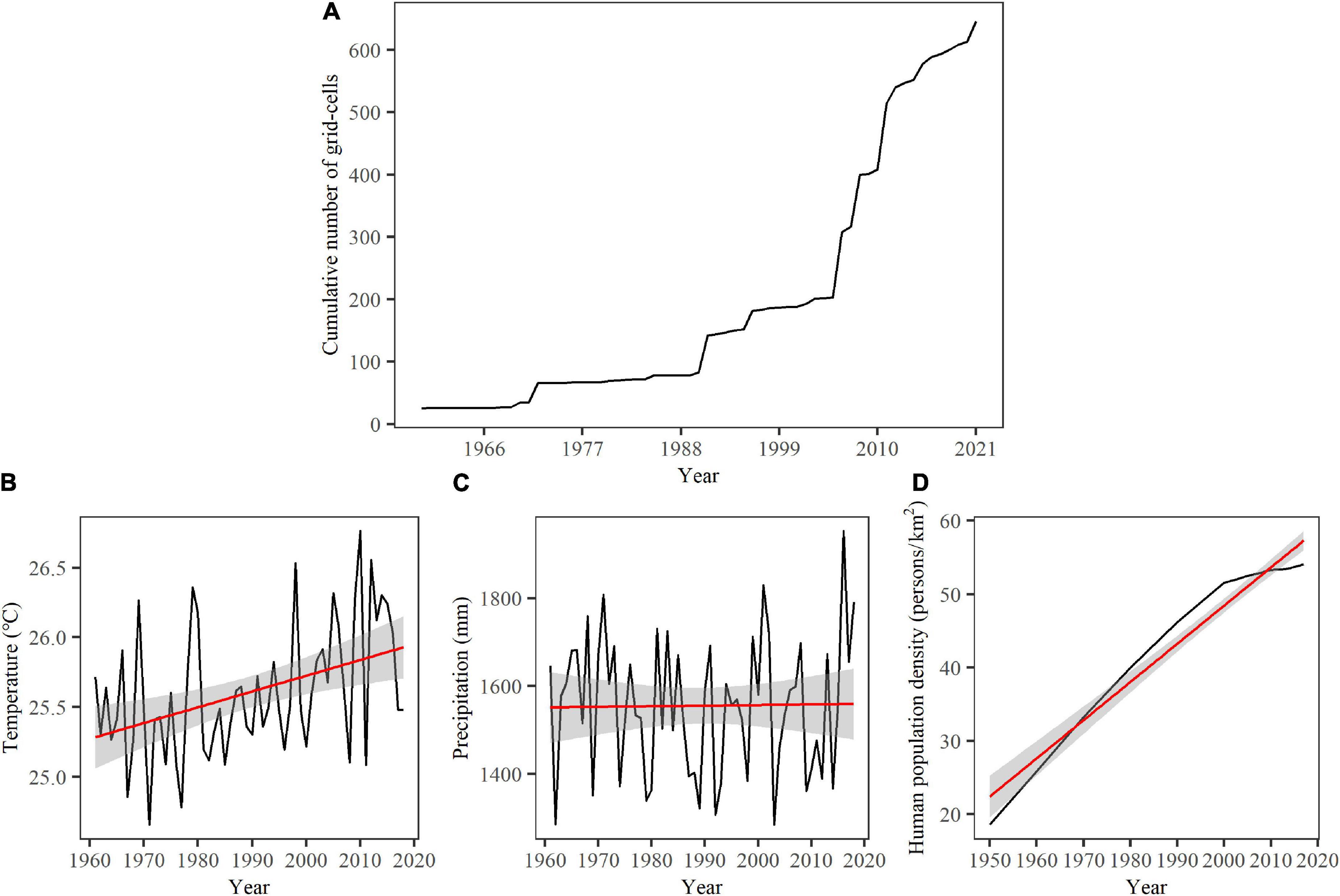
Figure 2. Temporal change of a cumulative number of grid cells of first observation for Asian elephants (A), air temperature (B), precipitation (C), and human population density (D) in Yunnan, China. Gray solid lines represent the temporal change, and the red lines represent their linear regression trend. The linear correlation with time for temperature and human population density is significant (all p < 0.001) while not significant for precipitation (p = 0.911). The gray shadow shows the 95% confidence interval of the regressions.
Climate and environmental variables
Temperature, precipitation, and human population density were selected as climate and environmental variables. Historical monthly temperature and precipitation data during 1961–2018 were downloaded from WorldClim.8 The spatial resolution of temperature and precipitation data was approximately 5 × 5 km2. The monthly maximum air temperatures for the 12 months of each year were used to calculate the yearly maximum air temperature by calculating their average values. The maximum annual air temperature was used to analyze the association between the first observation probability (non-presence = 0, presence = 1) of Asian elephants and air temperature from 1959 to 2021 in the grid cells. The annual precipitation was calculated using the summed values of the monthly total precipitation for 12 months each year for each grid cell. Human population density (HPD, the number of persons per km2) was used to represent the pressure of human activities in each grid cell. Because the landscape would be more altered by human activities; thus, the human population density could represent the landscape change to some extent. The human population density data was downloaded from the History Database of the Global Environment (Goldewijk and Dr. ir, 2017). Human population density data had a temporal scale from 10000 B.C. to 2017 and a spatial resolution of approximately 10 × 10 km2.9 We fixed the grid cell size at 5 × 5 km2 to get more temperature and precipitation data used in the model analysis. The center point of each 5 × 5 km2 grid cell was used to extract temperature, precipitation, and human population density data.
To explore the relationship between the recent local range expansion of Asian elephants with the change in climate and human pressures, the yearly maximum temperature, the annual precipitation, and human population density were standardized to remove their regional effects. The zero-mean normalization was used to obtain the standardized values of temperature, precipitation, and human population density by removing the average values of each grid cell during the study period. The collinearity test with the Spearman correlation coefficient was used to quantify the correlations between environmental variables (Supplementary Table 3). We removed one of the variables of air temperature and human population density because their correlation coefficients are high (r > 0.4) to avoid strong collinearity in modeling analysis.
Statistical analysis
The cumulative number of grid cells with the first observation year was used to represent the recent range expansion of Asian elephants. To identify the effects of climate and a human stressor on the local range expansion of Asian elephants, we compared associations of the probability of first observation (presence, 1) or non-observation (absence, 0) of Asian elephants in a grid cell with the average values of temperature, precipitation, and human population density within a time window before the first observation time (year) and a time window before non-observation time (year). It would take some years for elephants to change their distribution site under climate and human stressors. Elephants may tolerate short-term stress through physiological adaptation, while habitat deterioration may take more time to force elephants to leave their original sites. Two-time windows were used to test the scale-dependent effects of stressors in time, including 5- and 10-year time windows, which cover several short periods (about 2–5 years) of climate variation (Supplementary Figure 2). The year that was 5 or 10 years (corresponding to different time window selections) before the first observation year was selected as the non-observation year. Thus, for each grid cell, there is a pair of data of the first observation year (probability of presence = 1) and non-observation year (probability of absence = 0), and the average values of temperature, precipitation, and human density during the 5- or 10-year window. For details of the calculation, please see Supplementary Figure 3.
Following Wan et al. (2019) and Bai et al. (2022), a logistic generalized additive model (GAM) was fitted by using the following equation to reveal the association of the first observation probability (Yit) with the standardized temperature (Tit), standardized precipitation (Pit) and standardized human population density (Hit) in all grid cells:
where Yit represented the first observation probability in the ith grid cell at time t. Tit, Hit, and Pit represent the average values of the standardized air temperature, standardized human population density, and standardized precipitation during a time window after removing the regional difference, respectively. Because correlations between standardized human population density and standardized temperature were relatively high for two time windows (r = 0.45 and 0.66 for 5- and 10-year time windows) (Supplementary Table 3), we also fitted GAM models by including only one of these two correlated variables in the models, separately. s(Loni,Lati) is a 2D smoothing function (k value = 4) for removing the spatial autocorrelation effects (Wan et al., 2019; Bai et al., 2022). Because there is only paired data for each grid cell; thus, the autocorrelation effect in time should be small. at is an intercept. bt, ct, and dt represent the associations of the first observation probability of Asian elephants with the standardized temperature, standardized human population density, and standardized precipitation. εit represents the random error term. A partial autocorrelation function (PACF) and residual semivariogram were used to detect residual temporal correlations and residual spatial correlations (Wan et al., 2019). Generalized additive models can better detect non-linear relationships (Dominici et al., 2002) and were applied using the mgcv library (v 1.8-33) in R software (version 4.0.3). The WaveletComp library (v 1.1) (Roesch and Schmidbauer, 2018) was used to show the periods of temperature and precipitation during 1961–2018.
To project the range expansion of Asian elephants during 2021–2030 under climate change, a GAM model using a 10-year time window was used to model the first observation probability under two future climate warming scenarios. Maximum monthly temperature and monthly total precipitation during 2021–2040 with Shared Socio-economic Pathways 126 (SSP1-2.6) and 245 (SSP2-4.5) from WorldClim (see textfootnote 8) were used as environmental variables. MRI-ESM2-0 (global climate model) from CMIP6 was used because it performs better in major regions of China (Xiang et al., 2021). Half of the absolute temperature and precipitation increment or reduction between 2001–2018 and 2021–2040 were used in the GAM model to project the first observation probability of Asian elephants during 2021–2030. The human population density changes between 2011–2020 and 2021–2030 in each grid cell were also used according to the population growth rate (1.83 persons/km2 per year) during 1950–2017. In this analysis, the probability of the elephant’s arrival at a new grid cell was independent of the cell’s distance to the current elephant range. This is, the results indicate habitat suitability more than the actual probability of recolonization in the time frame considered. The projected range of Asian elephants covers Yunnan Province and its surrounding counties, as well as the neighboring provinces. All selected counties were divided into grid cells of 5 × 5 km2. The first observation probability during 2021–2030 in all grid cells was projected and visualized in ArcGIS software (version 10.8) after classification. The yearly average maximum temperature during 2021–2040 for all presence grid cells from 1959 to 2021 was calculated as future climate pressure with SSP1-2.6 and SSP2-4.5, respectively.
Results
Recent range expansion of Asian elephants
There were 7,117 records (6,996 records from historical literature, 164 records from news reports, 7 from GBIF, and 10 from iNaturalist) of Asian elephants covering the period 1959–2021, and these were designated into 646 grid cells (5 × 5 km2) (Figure 1 and Supplementary Tables 1, 2). The cumulative number of grid cells with the first observation of Asian elephants showed a steady increase since 1991 and then a sharp increase since 2006 (Figure 2A), which corresponds to the big expansion of elephants in space since 1990 in Figure 3. The average latitude of presence grid cells of Asian elephants showed a significant increase between 1959–1990 and 1991–2021, while the average longitude did not show such a significant difference (Figure 3). Both air temperature from 1961 to 2018 and human population density during 1950–2017 showed a significant increasing trend (about 0.5°C increase for temperature and 35 persons per km2 for human population density) (Figures 2B,D, p = 0.001 for temperature and p < 0.001 for HPD), but precipitation during 1961–2018 showed no significant increasing trend (Figures 2C, p = 0.911). Air temperature showed a significant 3-year cycle (p < 0.05) (Figure 2B and Supplementary Figures 2A,B) and precipitation showed significant 2-, 5-, and 16-year (p < 0.05 for 5- and 16-year) cycles (Figure 2C and Supplementary Figures 2C,D) during 1961–2018 in Yunnan. Air temperature showed a greater increase in the north than in the south between 1961–1970 and 2009–2018 in the countries with the presence of Asian elephants (Figure 4); change in air temperature showed a significant and positive correlation with latitude in both 5- and 10-year time windows; a significant and negative correlation with longitude at the 5-year time window (all p < 0.001) (Supplementary Table 4). Precipitation showed more decrease in the north but an increase in the south between 1961–1970 and 2009–2018 (Figure 4); change in precipitation showed a negative correlation with latitude at both time windows (p = 0.012 for 5 years and p = 0.005 for 10 years); a significant and positive association with longitude at both time windows (p < 0.001 for 5 years and p = 0.014 for 10 years) (Supplementary Table 4). Human population density showed an increase between 1960–1970 and 2008–2017 in nearly all counties with the presence of Asian elephants in Yunnan Province; the change in human population density showed a significant and positive correlation with latitude at both time windows (all p < 0.001), not with longitude (Figure 4 and Supplementary Table 4). The threshold of the yearly average maximum air temperature triggering the movement of Asian elephants from 1959 to 2021 was estimated to be 20.46 ± 1.21°C. The predicted yearly average maximum temperature during 2021–2040 was 26.52 ± 1.44°C and 26.67 ± 1.47°C for SSP 1-2.6 and SSP2-4.5, respectively. Thus, the recent northern range expansion of Asian elephants will continue under the expected warming conditions during 2021–2040.

Figure 3. Differences of average latitude (A) and longitude (B) for presence grid cells of Asian elephants between 1959–1990 (C) and 1991–2021 (D) in southwest China. Boxplots indicate average values and variation of latitude and longitude (A,B) for the two periods (NS, non-significant; ***p < 0.001). Blue and pink-red dots in panels (C,D) represent the sites of first observation in the two periods (1959–1990 and 1991–2021). The green colored background on the top right panels in (C,D) represent the recent IUCN distribution of Asian elephants (Williams et al., 2020).
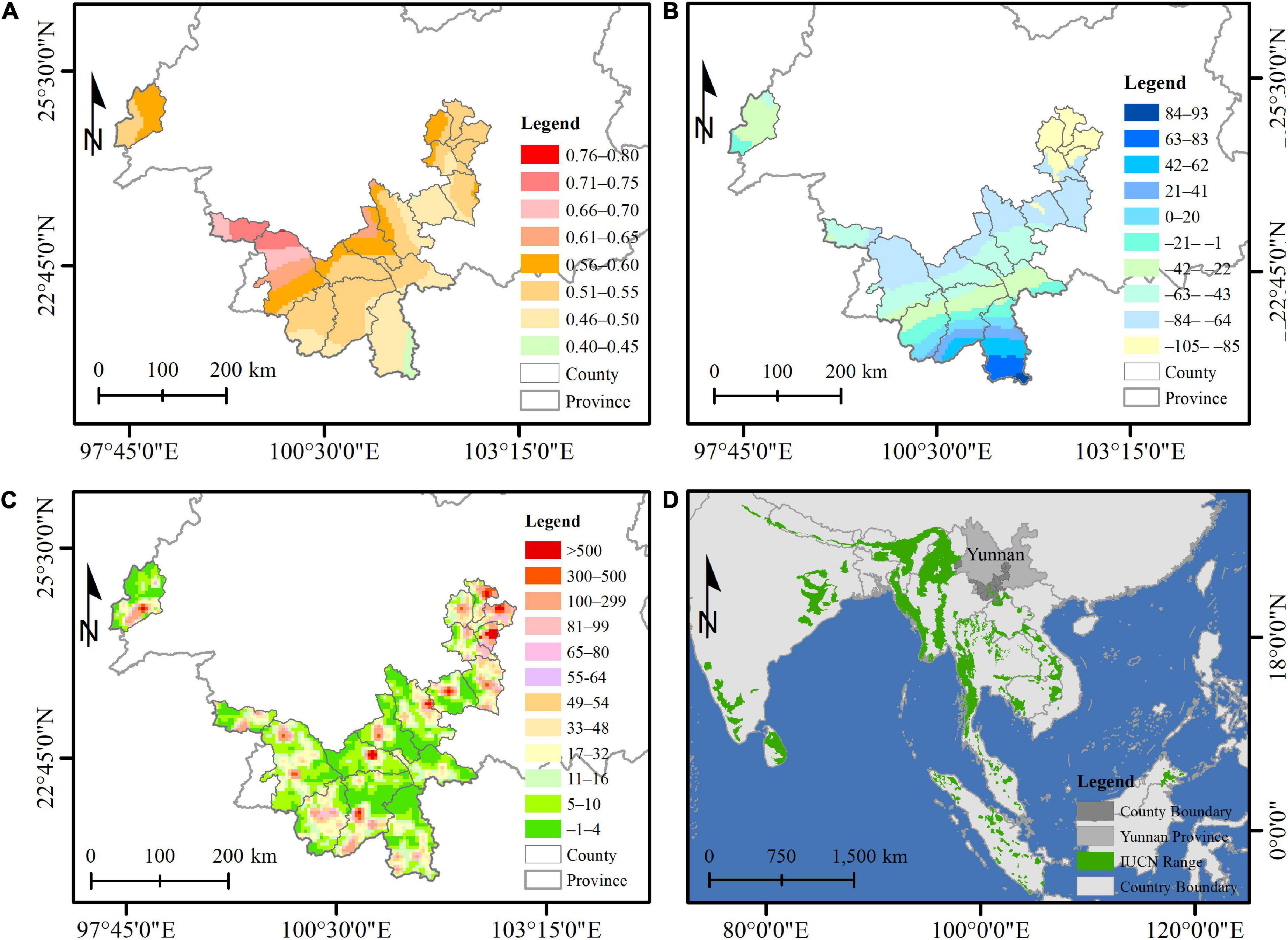
Figure 4. Spatial change of temperature (°C) (A), precipitation (mm) (B), and human population change (persons per km2) (C) in counties with a historical presence of Asian elephants in Yunnan, China (D). Temperature and precipitation changes were calculated by the difference between 1961–1970 and 2009–2018 in each 5 × 5 km2 grid cell. Human population density change was calculated by the difference between 1960–1970 and 2008–2017 in each 5 × 5 km2 grid cell. The dark gray colored background in (D) represents the counties with the presence of Asian elephants in Yunnan, China. The green colored background in (D) represents the recent IUCN distribution of Asian elephants (Williams et al., 2020).
First observation probability of Asian elephants
Based on the significant associations with all environmental variables (we only reported significant associations, see below), we found the first observation probability of Asian elephants showed a positive association with human population density for the 5-year time window (Table 1 and Figures 5A,D); a positive association with temperature for the 10-year time window (Table 1 and Figures 5B,E); a positive association with precipitation for the 10-year time window (Table 1 and Figure 5F), but a negative association with precipitation for the 5-year time window (Table 1 and Figure 5C). The models for 5- and 10-year time windows explained 2.10 and 25.50% of the variance, respectively (Supplementary Table 5).

Table 1. Associations of standardized temperature, standardized precipitation, and standardized human population density (HPD) with the first observation probability of Asian elephants at 5- and 10-year time scales.
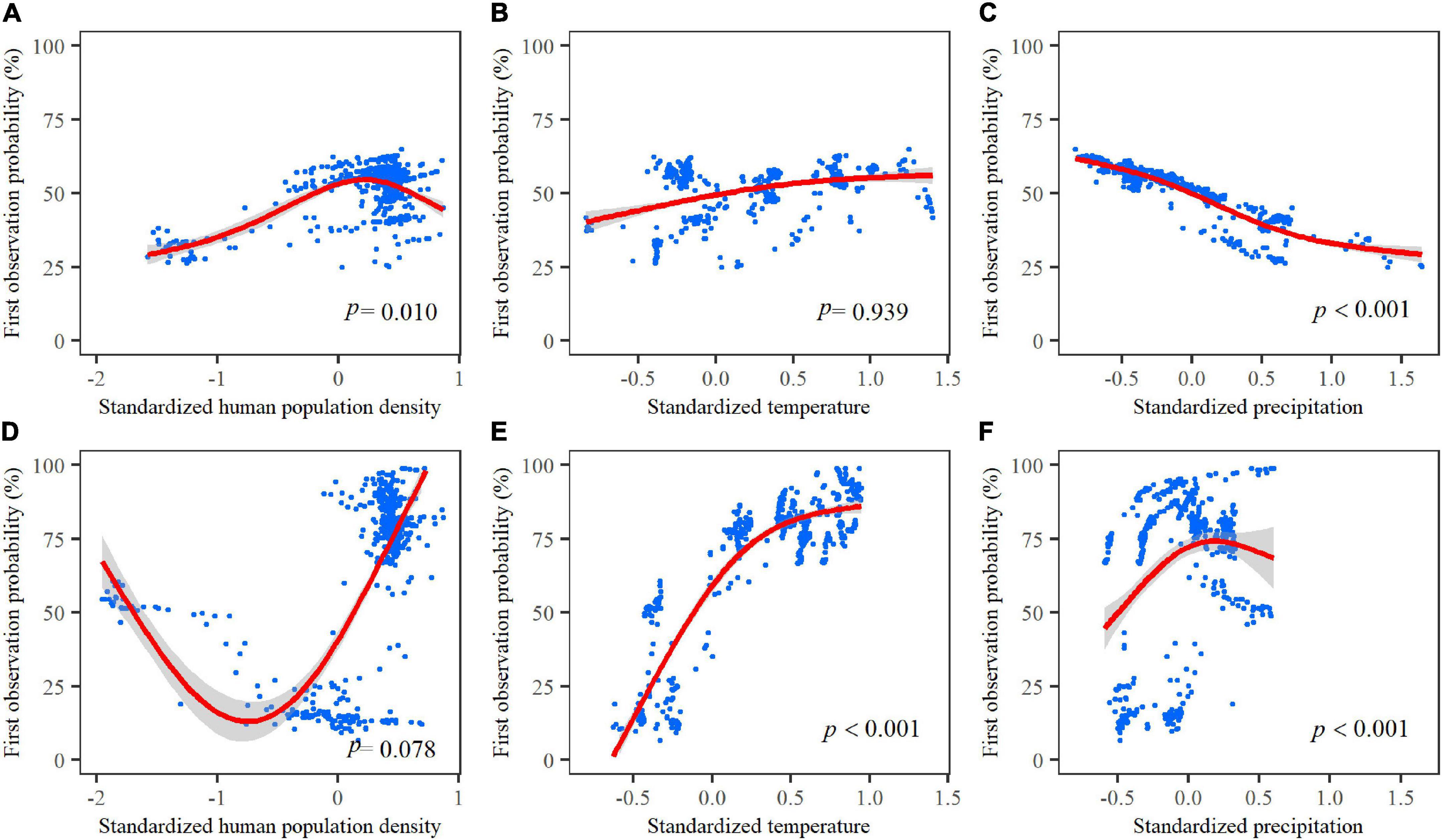
Figure 5. Association of the first observation probability of Asian elephants with the standardized human population density, standardized temperature, and standardized precipitation by using GAM models using different time windows [(A–C) for the 5-year period; (D–F) for 10-year period]. Blue dots represent the first observation probability for each grid cell. Red lines represent the logistic regressions [p < 0.001 for (C,E,F), the confidence interval is 95%].
If we removed temperature from the models to avoid its collinearity with human density, we found the first observation probability of Asian elephants showed a positive association with human population density for the two-time windows (Table 2 and Supplementary Figures 4A,E); a positive association with precipitation for the 10-year time window (Table 2 and Supplementary Figure 4F), but a negative association for the 5-year time window (Table 2 and Supplementary Figure 4B). The models for 5- and 10-year time windows explained 2.11 and 9.42% of the variance, respectively (Supplementary Table 5).
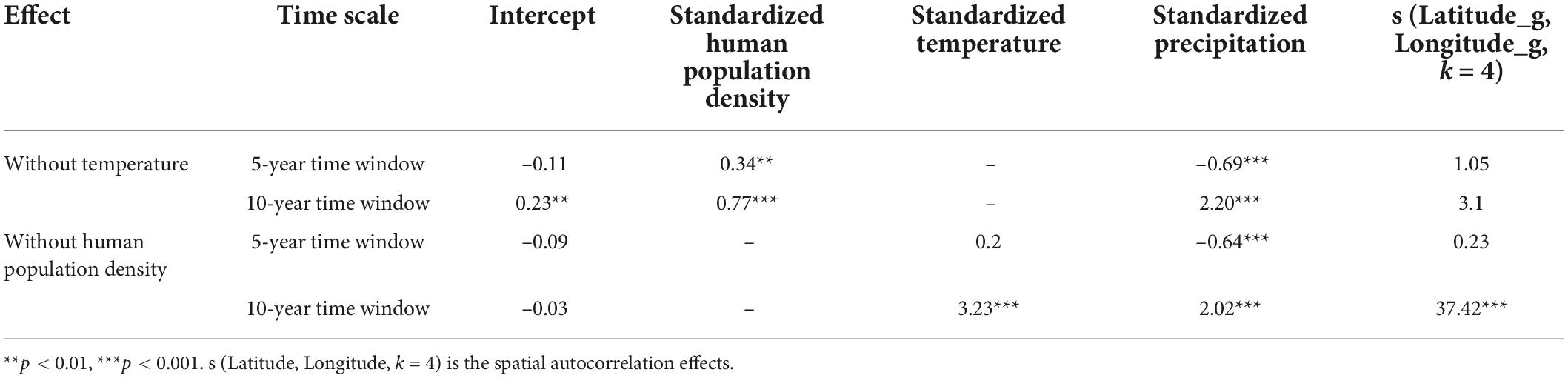
Table 2. Relationships between standardized temperature, standardized precipitation, and standardized human population density (HPD) with the recent range expansion of Asian elephants after excluding highly correlated environment variables.
If we removed human population density from the models to avoid its collinearity with temperature, we found the first observation probability of Asian elephants showed a positive association with temperature for 10-year window (Table 2 and Supplementary Figure 4G); and a positive association with precipitation for the 10-year scale (Table 2 and Supplementary Figure 4H) but a negative association for 5-year time window (Table 2 and Supplementary Figure 4D). The models for 5- and 10-year time windows explained 1.64 and 25.30% of the variance, respectively (Supplementary Table 5).
Residual temporal autocorrelation diagnostics showed that partial autocorrelation coefficients gradually converge with an increase in lag number for all selected models, and the temporal autocorrelation decreased with the rise of the time window (5- and 10-year); residual temporal autocorrelation diagnostics results showed a first-order temporal autocorrelation in the 5- and 10-year time windows (Supplementary Figures 5, 6). Semivariogram spatial autocorrelation diagnostics showed a minor spatial autocorrelation for all selected models. The spatial autocorrelation increased with an increase in time windows (5 and 10 years) (Supplementary Figures 5, 6).
Projections of range expansion
The model with the 10-year time window and the largest power to explain the variance was used to project elephant range expansion in the next 10 years. The projected range expansion analysis showed that the first observation probability was much larger in the northeast regions than in the other areas in the next 10 years under two climate scenarios. Asian elephants would expand their recent northern boundary to the northeast part of Yunnan province under climate warming scenarios during 2021–2030 with a rise of 0.50 and 0.47°C under Shared Socio-economic Pathways 126 (SSP1-2.6) and 245 (SSP2-4.5), respectively (Figure 6).
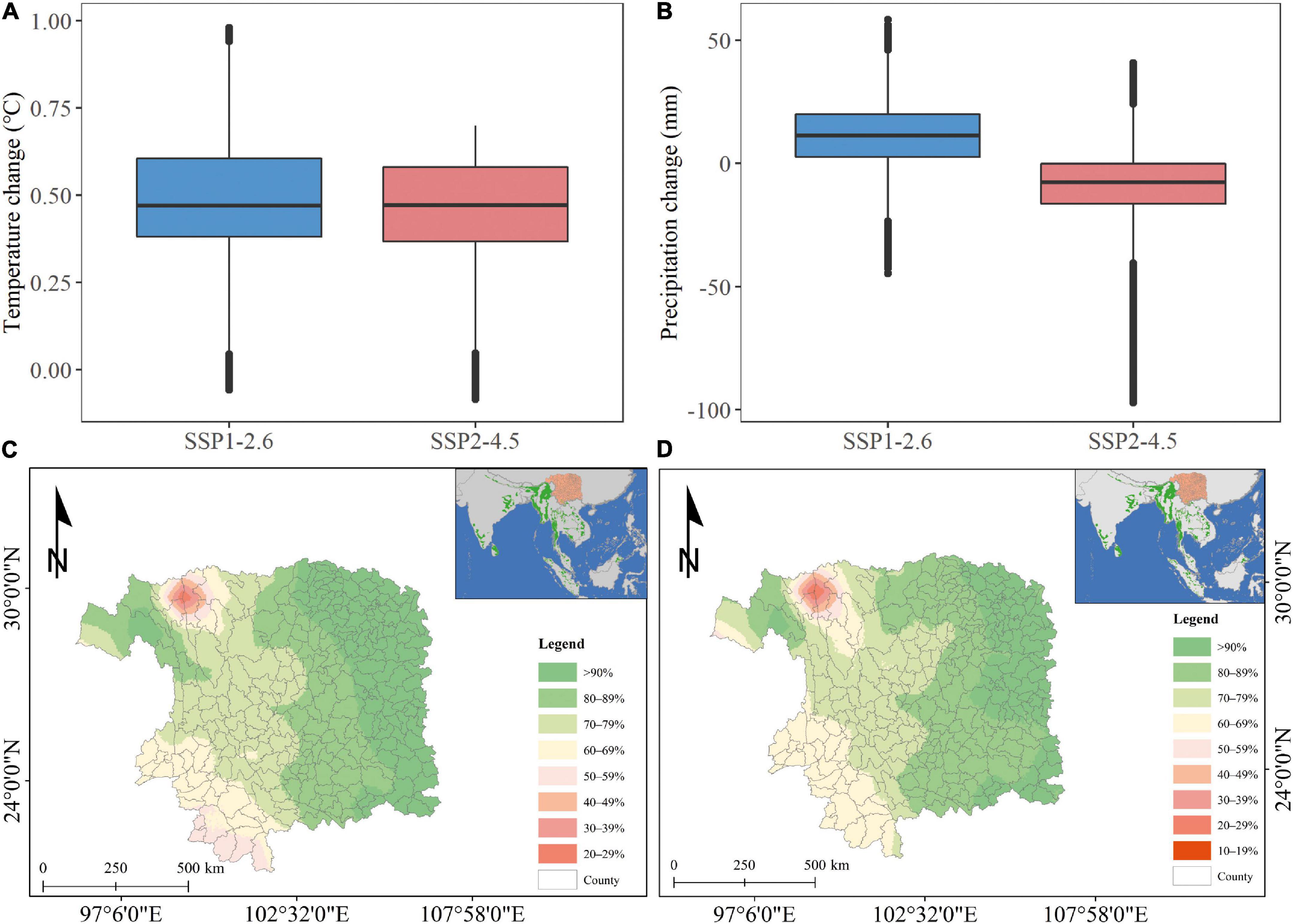
Figure 6. Projected first observation probability of Asian elephants in China under future climate change conditions (A,B) of Shared Socio-economic Pathways 126 (SSP1-2.6) (C) and 245 (SSP2-4.5) (D) during 2021–2030 based on GAM models of 10-year window. The projected range of Asian elephants covers Yunnan Province and its surrounding counties. Half of the absolute temperature and precipitation increment or reduction between 2001–2018 and 2021–2040 were calculated and plotted (A,B) for all projected 5 × 5 km2 grid cells (C,D). SSP1-2.6 and SSP2-4.5 correspond to an annual mean temperature rise of 0.50 and 0.47°C, respectively. The green colored background on the top right panels in (C,D) represents the recent IUCN distribution of Asian elephants (Williams et al., 2020).
Discussion
Our study found Asian elephants showed obvious northward range expansion to areas of higher latitude with a higher increase in human population density but a lower increase in precipitation since 1991 and demonstrated that the recent northward range expansion of Asian elephants in Yunnan as represented by the first observation probability showed close associations with climate change and human settlements. The first observation probability showed a significant positive association with climate warming at 10-year time window and human population density at both 5- and 10-year time windows; a scale-dependent association with precipitation: negative for the 5-year but positive for the 10-year time window. The associations of the recent range expansion were generally consistent among all models, but GAM models with the 10-year time window had a larger explanation power. Our results generally support Hypothesis I and Hypothesis II, but not Hypothesis III. Our study suggests that climate warming was likely associated with the recent range expansion of Asian elephants toward the north and to areas with more human settlements in Yunnan, China; high precipitation would benefit elephants by providing food and habitat resources. Based on projections under climate warming scenarios during 2021–2030, Asian elephants would likely expand their ranges to the northeast part of the current distribution area. The current distribution area of elephants in Yunnan, as revealed by our study area, is much larger than that of IUCN, especially in the northern part of their distribution (Supplementary Figure 7). Our findings could provide some useful cues for managing Asian elephants under accelerated global change.
Association of climate warming with the recent range expansion
There is abundant evidence of the association of climate warming with changes in the distribution range of many mammal species (Hersteinsson and Macdonald, 1992; Peterson et al., 2002; Dawe and Boutin, 2016). Asian elephants are tropical and subtropical mammals. The northern range of elephants is likely limited by their physiological tolerance to cold temperatures. Li et al. (2015) found climate cooling caused historical range contraction of northern and eastern boundaries of Asian elephants in China over the past two millennia. In periods of the warm phase of the Song Dynasty (around A.D. 1500), Asian elephants showed a northern expansion temporally (Li et al., 2015). Asian elephants are more active in warm conditions (Norton-turner, 2020). Disease risks (e.g., foot-and-mouth disease) to Asian elephants are also low in warming environments (Jiang et al., 2020). Favorable plants (such as Musa balbisiana Colla) are abundant in warm climate regions but less in cold climate regions (Feng et al., 2009). In this study, we found that Asian elephants in Yunnan have shown changes in distribution and obvious northward movement since 1990. This demonstrated that this movement is significantly and positively associated with an increase in air temperature, supporting Hypothesis I. Indeed, temperature showed a greater increase in the northern part of Yunnan, and the rise in air temperature was significantly and positively correlated with latitude at both time windows (Figure 4 and Supplementary Table 4), corresponding well to the range movement of Asian elephants toward the north (Figures 1A, 3). The average latitude of places with elephants was significantly higher in the second period of 1991–2021 than that of the first period of 1959–1990 (Figure 3), indicating Asian elephants showed obvious northward expansion during the past decades. Climate warming in the north probably benefits Asian elephants both directly (physiologically) and indirectly (by altering the plant community), which needs further investigation.
Extreme high temperatures may also impose negative effects on an elephant. The mortality of Asian elephants of both sexes and across all ages would increase when the temperature exceeded 24°C (Mumby et al., 2013). It is estimated that more than 45% of the current suitable habitat for Asian elephants in China could be lost by 2050 under climate warming scenarios (Li et al., 2019). Indeed, we found a saturated association of the first observation probability with temperature for the 10-year time window (Figure 5E and Supplementary Figure 4G), suggesting that sustained high temperatures (e.g., on their southern boundary) may impose no extra positive effect on the recent range expansion of elephants. However, by looking at Figure 1B in this study, the range of the southern populations of elephants is still stable during the past decades, suggesting that elephants can comfortably endure the current warmer temperatures. Instead, the rising temperatures in the north provide new favorable environments for Asian elephants, which may facilitate their movement toward the north of Yunnan.
Association of precipitation with the recent range expansion
Low precipitation or drought could change habitat utilization in mammals (Dunham, 1994; Cain et al., 2017). The distribution of Asian elephants was mainly affected by changes in climatic water balance, then by changes in temperature and human impacts; Asian elephants shifted upward in response to different levels of water availability and seasonal droughts in India and Nepal from 2002 to 2017 (Kanagaraj et al., 2019). In the past several decades, a decrease in precipitation has been reported in Yunnan Province (Liu et al., 2007, 2010). Food resource changes in drought conditions might alter the distribution of large mammals (Hale et al., 2016; Gedir et al., 2020). Food was the main factor impacting the ranging patterns of Asian elephants (Lin et al., 2011). The northward movements of Asian elephants in southwestern Yunnan in 2021 were suspected to be connected to food shortages due to the hot and dry weather from 2019 to 2020 (Wang L. et al., 2021).
In this study, we did not find an obvious increasing or decreasing trend in precipitation in Yunnan; rather, precipitation showed large variation across years, ranging from 1,284 to 1,953 mm from 1961 to 2018 (Figure 2). Precipitation showed a time scale-dependent association with the recent range expansion of Asian elephants, positive for the 10-year time window while negative for the 5-year time window. These results, at a 10-year time window scale, provide support for Hypothesis II. As a tropical or subtropical animal, abundant precipitation is essential for the Asian elephants’ food and water resources. However, extreme drought may force them to leave their original habitats to search for food (Bohrer et al., 2014). Thus, this negative relationship between the first observation and precipitation was likely driven by extreme drought events in short-time periods in Yunnan (Figure 2C and Supplementary Figures 2C,D). Extreme episodes, like severe droughts, likely caused food and water limitations and triggered the movement of the northbound elephants to more abundant food and water sources in human settlement areas or farmlands.
The role of precipitation in the driving movement of the surrounding area along longitude may be more important than along latitude. We found that within the distribution area of Asian elephants in Yunnan, precipitation decreased in the north but increased in the south (Figure 4). Changes in precipitation were significantly and negatively associated with the latitude at both time windows (Supplementary Table 4). Change in precipitation showed a positive association with longitude at both time windows (Supplementary Table 4). However, associations between precipitation and latitude or longitude were very weak. Thus, the northward elephant expansion in Yunnan was likely driven by climate warming and human population density, which had a larger correlation with latitude (Supplementary Table 4), which is different from the observations that water balance was more important than temperature and human impacts in affecting the distribution of elephants in India and Nepal (Kanagaraj et al., 2019). The negative association of first observation probability revealed by models at a 5-year time window may be less reliable due to its very low explanation power compared to the models of a 10-year time window. The lower precipitation in the north of our study area may be offset by an increase in cropland, which provides the elephants with better food and water resources.
Relationship between human impacts with the recent range expansion
Human activity (i.e., land use change and habitat fragmentation) is believed to be the key factor shaping the distribution range of large mammals (Real et al., 2010; Polaina et al., 2019; Huang et al., 2021). Human activities, such as land use transformation and artificial facility construction, are the major factors inducing the distribution change of Asian elephants (Lin et al., 2008). Human-related land use change may drive the range expansion of Asian elephants by impacting the availability of food and water (Sukumar, 2006). Approximately 16% of the natural forest area disappeared, tea plantations increased more than 10%, and rubber plantations expanded more than 4,000 km2 between 1975 and 2014 in the eight counties covering the Asian elephant distribution range in Yunnan Province, and the greatest loss occurred in the southern part (mainly in Menghai, Jinghong, and Mengla) of the current range of elephants in China (Liu et al., 2017). Remote sensing data also demonstrates that the area of anthropogenic land cover has been increasing continuously since 1980, especially since 1990 when a large range expansion of elephants started (Supplementary Figure 7). Although large areas of natural land cover were transferred into anthropogenic land cover by 2020 in the southern part of the current range of elephants in Yunnan, the distribution of elephants also increased, likely because they were forced to look for food in the human-disturbed areas. There was a larger range expansion in the northern part of their distribution, where the landscape was less disturbed, likely due to an increased temperature, which provides a favorable climate. Thus, habitat loss would force elephants to expand their distributions. Besides, the population growth of Asian elephants in protected areas might increase intraspecific competition and then contribute to their recent range expansion.
It is widely recognized that human disturbance would impose negative impacts on large animals. High human density contributed to the range contraction of Asian elephants, especially in the eastern part of China, where human activities were more extensive (Li et al., 2015). Fecal cortisol (in part, reflecting general stress) concentrations of Asian elephants increased with intensified human disturbances and the expansion of tea plantations (Tang et al., 2020). Contrasting to traditional knowledge, here we found that the recent range expansion of Asian elephants was positively associated with human population density and a U-shaped association for a 10-year time window in Yunnan, suggesting a distinct mechanism of human activities on Asian elephants between current and old periods. This was likely related to the increase in bold behaviors of elephants in the modern era and their enforced protection. The time inflection point of the U-shaped relationship curve was around 1990. Before 1990, without full conservation, an increase in human population density would have prevented the population expansion of elephants. After 1990, enforcement of policies and laws for conserving elephants in Yunnan was implemented (He et al., 2020). For example, after Xishuangbanna Nature Reserve was upgraded from the province level to the national level to strengthen the protection of wildlife and rainforest in 1986,10 the local government provided more support to the routine monitoring and care service. It established a compensation mechanism for farmers for damage caused by elephants. Thus, elephants are less attacked or disturbed by people. They brazenly invade areas of human settlements with plenty of water and high-quality food resources. Elephants have become accustomed to eating crops and living among people. Thus, they tend to move from the nature reserves to human-inhabited areas, increasing the risk of human-wildlife conflicts. We report the first observation of elephants’ positive association with human density (after 1990), which supports observations that Asian elephants thrive in edge habitats with high human disturbance (Koirala et al., 2016; Wadey et al., 2018; Torre et al., 2021).
Implications for management
During the past decades, the distribution range of Asian elephants in China has expanded steadily toward the north in Yunnan. This has posed a great challenge for managing the increasing human-elephant conflicts. Human-elephant conflicts have caused a massive loss of property and even life to elephants and local residents in Yunnan (Xie et al., 2020). Considering the trend of both climate change and human activities will continue to increase globally, including in Yunnan (IPCC, 2001), it is urgently needed to take more effective measures to conserve Asian elephants and protect the property and lives of local people. Our results suggest that climate warming could promote the recent northward range expansion of Asian elephants. The projection analysis indicated that, in the next decade, Asian elephants would move toward Yunnan’s north and east directions, which is the direction they retreated in ancient times when the climate became cool. Based on our findings, we recommend expanding the existing protected areas or setting up new ones in Yunnan’s north and east to deal with the expected range expansion of the Asian elephant population. In addition, it is necessary to set up barriers between human settlement areas and natural habitats or reserves of elephants to minimize human-elephant conflicts. Local residents in the newly established protected areas should be aided in shifting their agricultural production into ecological tourism with complementary financial support from the government. Our results may have potential implications for protecting Asian elephants in other regions outside China under accelerated climate change and human activities. Enlargement of protection areas, setup of ecological corridors, real-time monitoring and early warning, and elephant damage compensation policy are key solutions to minimize wildlife-human conflicts in the future.
Data availability statement
The original contributions presented in this study are included in the article/Supplementary material, further inquiries can be directed to the corresponding authors.
Ethics statement
Ethical review and approval was not required for the animal study because it’s model analysis not lab experiments.
Author contributions
ZZ and FW designed the study. DB collected, processed, and analyzed the data. DB, XW, LZ, AC-A, and ZZ wrote and revised the manuscript. All authors gave final approval for the manuscript publication.
Funding
This work was partially supported by the IUBS-ISZS program of Biological Consequences of Global Change (BCGC).
Acknowledgments
We are grateful for the photos provided by the Yunnan Forest Fire Department.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.889077/full#supplementary-material
Footnotes
- ^ https://www.webofscience.com/
- ^ https://www.scholar.google.com
- ^ http://www.cnki.net/
- ^ https://doi.org/10.15468/dl.7jduxy
- ^ https://www.inaturalist.org/
- ^ https://www.baidu.com
- ^ https://www.google.com.hk/
- ^ https://www.worldclim.org
- ^ https://doi.org/10.17026/dans-25g-gez3
- ^ https://www.xsbn.gov.cn
References
Bai, D. F., Wan, X. R., Li, G. L., Guo, Y. W., Shi, D. Z., and Zhang, Z. B. (2022). Factors influencing range contraction of a rodent herbivore in a steppe grassland over the past decades. Ecol. Evol. 12:e8546. doi: 10.1002/ece3.8546
Bintliff, J. (2005). Human impact, land-use history, and the surface archaeological record: A case study from Greece. Geoarchaeology 20, 135–147. doi: 10.1002/gea.20040
Bohrer, G., Beck, P. S., Ngene, S. M., Skidmore, A. K., and Douglas-Hamilton, I. (2014). Elephant movement closely tracks precipitation-driven vegetation dynamics in a Kenyan forest-savanna landscape. Mov. Ecol. 2:2. doi: 10.1186/2051-3933-2-2
Brown, J. H., Valone, T. J., and Curtin, C. G. (1997). Reorganization of an arid ecosystem in response to recent climate change. Proc. Natl. Acad. Sci. U.S.A. 94, 9729–9733. doi: 10.1073/pnas.94.18.9729
Cain, J. W., Jay, V. G., Jason, P. M., Paul, R. K., Jamison, D. A., Glenn, C. D., et al. (2017). Extreme precipitation variability, forage quality and large herbivore diet selection in arid environments. Oikos 126, 1459–1471. doi: 10.1111/oik.04282
Campos-Arceiz, A., de la Torre, J. A., Wei, K., Wu, X. O., Zhu, Y., Zhao, M., et al. (2021). The return of the elephants: How two groups of dispersing elephants attracted the attention of billions and what can we learn from their behavior. Conserv. Lett. 14:e12836. doi: 10.1111/conl.12836
Cardillo, M., Georgina, M. M., Jones, K. E., Bielby, J., Bininda-Emonds, O. R., Sechrest, W., et al. (2005). Multiple causes of high extinction risk in large mammal species. Science 309, 1239–1241. doi: 10.1126/science.1116030
Chen, I., Hill, J. K., Ohlemüller, R., Roy, D. B., and Thomas, C. D. (2011). Rapid range shifts of species associated with high levels of climate warming. Science 1024, 17–20. doi: 10.1126/science.1206432
Chen, S., Yi, Z. F., Campos-Arceiz, A., Chen, M. Y., and Webb, E. L. (2013). Developing a spatially-explicit, sustainable and risk-based insurance scheme to mitigate human–wildlife conflict. Biol. Conserv. 168, 31–39. doi: 10.1016/j.biocon.2013.09.017
Chen, Y., Jorgelina, M., Chen, Y., Tao, Q., Sullivan, C. D., Shi, K., et al. (2016). Predicting hotspots of human-elephant conflict to inform mitigation strategies in Xishuangbanna, southwest China. PLoS One 11:e0162035. doi: 10.1371/journal.pone.0162035
Chen, Y., Sun, Y. K., Atzeni, L., Gibson, L., Hua, M., Li, K. Y., et al. (2021). Anthropogenic pressures increase extinction risk of an isolated Asian elephant (Elephas maximus) population in Southwestern China, as revealed by a combination of molecular- and landscape-scale approaches. Integr. Zool. [Epub ahead of print]. doi: 10.1111/1749-4877.12534
Christidis, N., Stott, P. A., and Brown, S. J. (2011). The role of human activity in the recent warming of extremely warm daytime temperatures. J. Clim. 24, 1922–1930. doi: 10.1175/2011JCLI4150.1
Crees, J. J., Carbone, C., Sommer, R. S., Benecke, N., and Turvey, S. T. (2016). Millennial-scale faunal record reveals differential resilience of European large mammals to human impacts across the Holocene. Proc. R. Soc. B Biol. Sci. 283:20152152. doi: 10.1098/rspb.2015.2152
Dawe, K. L., Bayne, E. M., and Boutin, S. (2014). Influence of climate and human land use on the distribution of white-tailed deer (Odocoileus virginianus) in the western boreal forest. Can. J. Zool. 92, 353–363. doi: 10.1139/cjz-2013-0262
Dawe, K. L., and Boutin, S. (2016). Climate change is the primary driver of white-tailed deer (Odocoileus virginianus) range expansion at the northern extent of its range; land use is secondary. Ecol. Evol. 6, 6435–6451. doi: 10.1002/ece3.2316
Di Marco, M., Rondinini, C., Boitani, L., and Murray, K. A. (2013). Comparing multiple species distribution proxies and different quantifications of the human footprint map, implications for conservation. Biol. Conserv. 165, 203–211. doi: 10.1016/j.biocon.2013.05.030
Di Marco, M., and Santini, L. (2015). Human pressures predict species’ geographic range size better than biological traits. Glob. Change Biol. 21, 2169–2178. doi: 10.1111/gcb.12834
Dominici, F., McDermott, A., Zeger, S. L., and Samet, J. M. (2002). On the use of generalized additive models in time-series studies of air pollution and health. Am. J. Epidemiol. 156, 193–203. doi: 10.1093/aje/kwf062
Dunham, K. M. (1994). The effect of drought on the large mammal populations of Zambezi riverine woodlands. J. Zool. 234, 489–526. doi: 10.1111/j.1469-7998.1994.tb04862.x
Easterling, D. R., Meehl, G. A., Parmesan, C., Changnon, S. A., Karl, T. R., and Mearns, L. O. (2000). Climate extremes: Observations, modeling, and impacts. Science 289, 2068–2074. doi: 10.1126/science.289.5487.2068
Feng, H. M., Chen, Y., Deng, C. J., and Wu, Y. T. (2009). Distribution of wild Musa species. J. Fruit Sci. 26, 361–368. doi: 10.13925/j.cnki.gsxb.2009.03.020
GBIF Secretariat (2021). Elephas maximus Linnaeus, 1758 in GBIF Secretariat. GBIF Backbone Taxonomy. Available Online at: https://doi.org/10.15468/39omei [accessed November 14, 2021].
Gedir, J. V., Cain, J. W., Swetnam, T. L., Krausman, P. R., and Morgart, J. R. (2020). Extreme drought and adaptive resource selection by a desert mammal. Ecosphere 11:e03175. doi: 10.1002/ecs2.3175
Girdland-Flink, L., Albayrak, E., and Lister, A. M. (2018). Genetic insight into an extinct population of Asian elephants (Elephas maximus) in the Near East. Open Q. 4, 1–9. doi: 10.5334/oq.36
Goldewijk, K., and Dr. ir, C. G. M. (2017). Anthropogenic land-use estimates for the Holocene; HYDE 3.2. DANS. Utrecht: Utrecht University. doi: 10.17026/dans-25g-gez3
Hale, S., Nimmo, D. G., Cooke, R., Holland, G., James, S., Stevens, M., et al. (2016). Fire and climatic extremes shape mammal distributions in a fire-prone landscape. Divers. Distrib. 22, 1127–1138. doi: 10.1111/ddi.12471
He, C. H., Du, J. J., Zhu, D., and Zhang, L. (2020). Population viability analysis of small population: A case study for Asian elephant in China. Integr. Zool. 15, 350–362. doi: 10.1111/1749-4877.12432
Hersteinsson, P., and Macdonald, D. W. (1992). Interspecific competition and the geographical distribution of red and foxes Vulpes vulpes and Alopex lagopus. Oikos 64, 505–515. doi: 10.2307/3545168
Huang, C., Li, X. Y., Hu, W. Q., and Jiang, X. L. (2020). Predicting indirect effects of transportation network expansion on Asian elephants: Implications for environmental impact assessments. Biotropica 52, 196–202. doi: 10.1111/btp.12726
Huang, C., Li, X. Y., Khanal, L., and Jiang, X. L. (2019). Habitat suitability and connectivity inform a co-management policy of protected area network for Asian elephants in China. PeerJ 7:e6791. doi: 10.7717/peerj.6791
Huang, P. Z., Bian, K., Huang, Z. P., Li, Q., Dunn, D. W., Fang, G., et al. (2021). Human activities and elevational constraints restrict ranging patterns of snub-nosed monkeys in a mountainous refuge. Integr. Zool. 16, 202–213. doi: 10.1111/1749-4877.12490
Huang, Q. Y., Lothspeich, A., Hernández-Yáñez, H., Mertes, K., Liu, X. H., and Songer, M. (2020). What drove giant panda Ailuropoda melanoleuca expansion in the Qinling Mountains? An analysis comparing the influence of climate, bamboo, and various landscape variables in the past decade. Environ. Res. Lett. 15:084036. doi: 10.1088/1748-9326/ab86f3
Isaac, N. J. B., and Cowlishaw, G. (2004). How species respond to multiple extinction threats. Proc. R. Soc. B Biol. Sci. 271, 1135–1141. doi: 10.1098/rspb.2004.2724
Jiang, F., Song, P. F., Zhang, J. J., Cai, Z. Y., Chi, X. W., Gao, H. M., et al. (2020). Assessing the impact of climate change on the spatio-temporal distribution of foot-and-mouth disease risk for elephants. Glob. Ecol. Conserv. 23:e01176. doi: 10.1016/j.gecco.2020.e01176
Jiang, Z., Jiang, J., Wang, Y., Zhang, E., Zhang, Y., Li, L., et al. (2016). Red list of China’s vertebrates. Biodivers. Sci. 24, 500–551. doi: 10.17520/biods.2016076
Kanagaraj, R., Araujo, M. B., Barman, R., Davidar, P., De, R., Digal, D. K., et al. (2019). Predicting range shifts of Asian elephants under global change. Divers. Distrib. 25, 822–838. doi: 10.1111/ddi.12898
Koirala, R. K., Ji, W. H., Aryal, A., Rothman, J., and Raubenheimer, D. (2016). Dispersal and ranging patterns of the Asian Elephant (Elephas maximus) in relation to their interactions with humans in Nepal. Ethol. Ecol. Ecol. 28, 221–231. doi: 10.1080/03949370.2015.1066872
Kowalczyk, R., Krasińska, M., Kamiński, T., Górny, M., Struś, P., Hofman-Kamińska, E., et al. (2013). Movements of European bison (Bison bonasus) beyond the Białowieża Forest (NE Poland): Range expansion or partial migrations? Acta Theriol. 58, 391–401. doi: 10.1007/s13364-013-0136-y
Li, J., Hou, Y. J., Li, Y. X., and Zhang, J. (2012). The latest straight-tusked elephants (Palaeoloxodon)? “Wild elephants” lived 3000 years ago in North China. Quat. Int. 281, 84–88. doi: 10.1016/j.quaint.2011.10.039
Li, W. W., Yu, Y., Liu, P., Tang, R. C., Dai, Y. C., Li, L., et al. (2019). Identifying climate refugia and its potential impact on small population of Asian elephant (Elephas maximus) in China. Glob. Ecol. Conserv. 19:e00664. doi: 10.1016/j.gecco.2019.e00664
Li, X. H., Jiang, G. S., Tian, H. D., Xu, L., Yan, C., Wang, Z. W., et al. (2015). Human impact and climate cooling caused range contraction of large mammals in China over the past two Millennia. Ecography 38, 74–82. doi: 10.1111/ecog.00795
Lin, L., Feng, L. M., Pan, W. J., Guo, X. M., Zhao, J. W., Luo, A. D., et al. (2008). Habitat selection and the change in distribution of Asian elephants in Mengyang Protected Area, Yunnan, China. Acta Theriol. 53, 365–374. doi: 10.1089/aid.2007.0229
Lin, L., Zhang, L. T., Luo, A. D., Wang, L. F., and Zhang, L. (2011). Population dynamics, structure and seasonal distribution pattern of Asian elephant (Elephas maximus) in Shangyong Protected Area, Yunnan, China. Acta Theriol. Sin. 31, 226–234. doi: 10.16829/j.slxb.2011.03.002
Lister, A. M., and Stuart, A. J. (2008). The impact of climate change on large mammal distribution and extinction: Evidence from the last glacial/interglacial transition. C. R. Geosci. 340, 615–620. doi: 10.1016/j.crte.2008.04.001
Liu, S. L., Dong, Y. J., Cheng, F. Y., Zhang, Y. Q., Hou, X. Y., Dong, S. K., et al. (2017). Effects of road network on Asian elephant habitat and connectivity between the nature reserves in Xishuangbanna, Southwest China. J. Nat. Conserv. 38, 11–20. doi: 10.1016/j.jnc.2017.05.001
Liu, X. R., Li, G. P., Fan, G. Z., Gheng, B. Y., and Li, H. Q. (2007). Spatial and temporal characteristics of precipitation resource in southwest China during 1961–2000. J. Nat. Resour. 22, 783–792.
Liu, Y., Zhao, R. X., Huang, W., Zhou, J. Q., and Ju, J. H. (2010). Characteristic analysis of precipitation and temperature trend in Yunnan Province in recent 46 years. J. Catastrophol. 25, 39–44, 63.
Lorenzen, E. D., Nogués-Bravo, D., Orlando, L., Weinstock, J., Binladen, J., Marske, K. A., et al. (2011). Species-specific responses of Late Quaternary megafauna to climate and humans. Nature 479, 359–364. doi: 10.1038/nature10574
Marucco, F., and McIntire, E. J. B. (2010). Predicting spatio-temporal recolonization of large carnivore populations and livestock depredation risk: Wolves in the Italian Alps. J. Appl. Ecol. 47, 789–798. doi: 10.1111/j.1365-2664.2010.01831.x
Meagher, M. (1989). Range expansion by bison of Yellowstone National Park. J. Mammal. 70, 670–675. doi: 10.2307/1381449
Moritz, C., Patton, J. L., Conroy, C. J., Parra, J. L., White, G. C., and Beissinger, S. R. (2008). Impact of a century of climate change on small-mammal communities in Yosemite National Park, USA. Science 322, 261–264. doi: 10.1126/science.1163428
Mumby, H., Courtiol, A., Mar, K. U., and Lummaa, V. (2013). Climatic variation and age-specific survival in Asian elephants from Myanmar. Ecology 94, 1131–1141. doi: 10.1890/12-0834.1
Nogués-Bravo, D., Rodríguez, J., Hortal, J., Batra, P., and Araújo, M. B. (2008). Climate change, humans, and the extinction of the woolly mammoth. PLoS Biol. 6:e79. doi: 10.1371/journal.pbio.0060079
Norton-turner, A. J. (2020). A camera trap study on the spatio-temporal behaviour of Asian elephant (Elephas maximus) to mitigate human-elephant conflicts in the Dong Phayayen-Khao Yai Forest Complex, Thailand. Master thesis. Uppsala: Swedish University of Agricultural Sciences.
Pan, W. J., Lin, L., Luo, A. D., and Zhang, L. (2009). Corridor use by Asian elephants. Integr. Zool. 4, 220–231. doi: 10.1111/j.1749-4877.2009.00154.x
Peterson, A. T., Ortega-Huerta, M. A., Bartley, J., Sánchez-Cordero, V., Soberón, J., Buddemeier, R. H., et al. (2002). Future projections for Mexican faunas under global climate change scenarios. Nature 416, 626–629. doi: 10.1038/416626a
Pimm, S. L., Jenkins, C. N., Abell, R., Brooks, T. M., Gittleman, J. L., Joppa, L. N., et al. (2014). The biodiversity of species and their rates of extinction, distribution, and protection. Science 344:1246752. doi: 10.1126/science.1246752
Polaina, E., González-Suárez, M., and Revilla, E. (2019). The legacy of past human land use in current patterns of mammal distribution. Ecography 42, 1623–1635. doi: 10.1111/ecog.04406
Real, R., Barbosa, A. M., Porras, D., Kin, M. S., Márquez, A. L., Guerrero, J. C., et al. (2010). Relative importance of environment, human activity and spatial situation in determining the distribution of terrestrial mammal diversity in Argentina. J. Biogeogr. 30, 939–947. doi: 10.1046/j.1365-2699.2003.00871.x
Ripple, W. J., Newsome, T. M., Wolf, C., Dirzo, R., Everatt, K. T., Galetti, M., et al. (2015). Collapse of the world’s largest herbivores. Sci. Adv. 1:e1400103. doi: 10.1126/sciadv.1400103
Roesch, A., and Schmidbauer, H. (2018). WaveletComp: Computational wavelet analysis. R package version 1.1. Available Online at: https://CRAN.R-project.org/package=WaveletComp (assessed November 13, 2021).
Rubidge, E. M., Monahan, W. B., Parra, J. L., Cameron, S. E., and Brashares, J. S. (2011). The role of climate, habitat, and species co-occurrence as drivers of change in small mammal distributions over the past century. Glob. Change Biol. 17, 696–708. doi: 10.1111/j.1365-2486.2010.02297.x
Sandom, C., Faurby, S., Sandel, B., and Svenning, J. C. (2014). Global late Quaternary megafauna extinctions linked to humans, not climate change. Proc. R. Soc. B Biol. Sci. 281:20133254. doi: 10.1098/rspb.2013.3254
Scanes, C. G. (2018). “Human activity and habitat loss: Destruction, fragmentation, and degradation,” in Animals and human society, eds C. G. Scanes and S. R. Toukhsati (Cambridge, MA: Academic Press), 451–482. doi: 10.1016/B978-0-12-805247-1.00026-5
Shapiro, S. S., and Wilk, M. B. (1965). An analysis of variance test for normality (complete samples). Biometrika 52, 591–611. doi: 10.2307/2333709
Sukumar, R. (2006). A brief review of the status, distribution and biology of wild Asian elephants. Int. Year Book 40, 1–8. doi: 10.1111/j.1748-1090.2006.00001.x
Sun, Y., Chen, Y., Jose, J., and Shi, K. (2021). Assessing population structure and body condition to inform conservation strategies for a small isolated Asian elephant (Elephas maximus) population in southwest China. PLoS One 16:e0248210. doi: 10.1371/journal.pone.0248210
Tang, R., Li, W., Zhu, D., Shang, X., Guo, X., and Zhang, L. (2020). Raging elephants: Effects of human disturbance on physiological stress and reproductive potential in wild Asian elephants. Conserv. Physiol. 8:coz106. doi: 10.1093/conphys/coz106
Torre, J. A., Wong, E. P., Lechner, A. M., Zulaikha, N., Zawawi, A., Abdul-Patah, P., et al. (2021). There will be conflict – agricultural landscapes are prime, rather than marginal, habitats for Asian elephants. Anim. Conserv. 24, 720–732. doi: 10.1111/acv.12668
Vidya, T. N. C., Sukumar, R., and Melnick, D. J. (2009). Range-wide mtDNA phylogeography yields insights into the origins of Asian elephants. Proc. R. Soc. B Biol. Sci. 276, 893–902. doi: 10.1098/rspb.2008.1494
Wadey, J., Beyer, H. L., Saaban, S., Othman, N., and Leimgruber, P. (2018). Why did the elephant cross the road? The complex response of wild elephants to a major road in Peninsular Malaysia. Biol. Conserv. 218, 91–98. doi: 10.1016/j.biocon.2017.11.036
Walther, G. (2001). “Adapted behaviour and shifting ranges of species-a result of recent climate warming,” in “Fingerprints” of climate change, eds G.-R. Walther, C. A. Burga, and P. J. Edwards (New York, NY: Kluwer Academic/Plenum), 1–16.
Walther, G., Post, E., Convey, P., Menzel, A., Parmesank, C., Beebee, T. J. C., et al. (2002). Ecological response to recent climate change. Nature 416, 389–395. doi: 10.1038/416389a
Wan, X. R., Jiang, G. S., Yan, C., He, F. L., Wen, R. S., Gu, J. Y., et al. (2019). Historical records reveal the distinctive associations of human disturbance and extreme climate change with local extinction of mammals. Proc. Natl. Acad. Sci. U.S.A. 116, 19001–19008. doi: 10.1073/pnas.1818019116
Wan, X. R., and Zhang, Z. B. (2017). Climate warming and humans played different roles in triggering Late Quaternary extinctions in east and west Eurasia. Proc. R. Soc. B Biol. Sci. 284:20162438. doi: 10.1098/rspb.2016.2438
Wang, H., Wang, P., Zhao, X., Zhang, W., Li, J., Xu, C., et al. (2021). What triggered the Asian elephant’s northward migration across Southwestern Yunnan? Innovation 2:100142. doi: 10.1016/j.xinn.2021.100124
Wang, L., Yang, B., Bai, Y., Lu, X., Zhu, J., Liu, Y., et al. (2021). Conservation planning on China’s borders with Myanmar, Laos, and Vietnam. Conserv. Biol. 35:1797–1808. doi: 10.1111/cobi.13733
Williams, C., Tiwari, S. K., Goswami, V. R., de Silva, S., Kumar, A., Baskaran, N., et al. (2020). Elephas maximus. The IUCN Red List of Threatened Species 2020: e.T7140A45818198. Available Online at: [accessed May 21, 2021]. doi: 10.2305/IUCN.UK.2020-3.RLTS.T7140A45818198.en
Xiang, J. W., Zhang, L. P., Deng, Y., She, D. X., and Zhang, Q. (2021). Projection and evaluation of extreme temperature and precipitation in major regions of China by CMIP6 models. Eng. J. Wuhan Univ. 54, 46–57, 81. doi: 10.1016/j.scitotenv.2021.148915
Xie, W. F., Song, J. P., Su, H. P., Yan, B., and Wang, Q. Y. (2020). Compensation status and measures of wildlife accident in Xishuangbanna national nature reserve. For. Inventory Plan. 45, 182–186.
Xu, J., Grumbine, R. E., and Beckschäfer, P. (2014). Landscape transformation through the use of ecological and socioeconomic indicators in Xishuangbanna, Southwest China, Mekong Region. Ecol. Indic. 36, 749–756. doi: 10.1016/j.ecolind.2012.08.023
Yang, Z. C. (2017). Distribution in entire range and assessment on local populations of Asian elephant in China. Master thesis. Beijing: Beijing Forestry University.
Zhang, L., Dong, L., Lin, L., Feng, L., Yan, F., Wang, L., et al. (2015). Asian elephants in China: Estimating population size and evaluating habitat suitability. PLoS One 10:e0124834. doi: 10.1371/journal.pone.0124834
Zhao, S., Zheng, P., Dong, S., Zhan, X., Wu, Q., Guo, X., et al. (2013). Whole-genome sequencing of giant pandas provides insights into demographic history and local adaptation. Nat. Genet. 45, 67–71. doi: 10.1038/ng.2494
Keywords: range shift, Asian elephant, climate warming, human impacts, human-wildlife conflict
Citation: Bai D, Wan X, Zhang L, Campos-Arceiz A, Wei F and Zhang Z (2022) The recent Asian elephant range expansion in Yunnan, China, is associated with climate change and enforced protection efforts in human-dominated landscapes. Front. Ecol. Evol. 10:889077. doi: 10.3389/fevo.2022.889077
Received: 03 March 2022; Accepted: 17 August 2022;
Published: 07 September 2022.
Edited by:
Ricardo Dobrovolski, Federal University of Bahia, BrazilReviewed by:
Xuelong Jiang, Kunming Institute of Zoology (CAS), ChinaChase A. LaDue, George Mason University, United States
Daniella Chusyd, Indiana University, United States
Copyright © 2022 Bai, Wan, Zhang, Campos-Arceiz, Wei and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fuwen Wei, d2VpZndAaW96LmFjLmNu; Zhibin Zhang, emhhbmd6YkBpb3ouYWMuY24=