 Susann Jänig1,2*†
Susann Jänig1,2*† Marlen Kücklich1,2†Lars Kulik1,2Madita Zetzsche1,2
Marlen Kücklich1,2†Lars Kulik1,2Madita Zetzsche1,2 Brigitte M. Weiß1,2‡
Brigitte M. Weiß1,2‡ Anja Widdig1,2‡
Anja Widdig1,2‡- 1Research Group Primate Behavioral Ecology, Department of Human Behavior, Ecology and Culture, Max Planck Institute for Evolutionary Anthropology, Leipzig, Germany
- 2Behavioral Ecology Research Group, Faculty of Life Sciences, Institute of Biology, University of Leipzig, Leipzig, Germany
In mammalian species, olfactory cues are important for within and between species communication. These cues can be part of multimodal signals indicating, for example, female fertility potentially perceived by male conspecifics. However, a large gap exists in our understanding of multimodal signaling in non-human catarrhines. Chimpanzees (Pan troglodytes) live in fission-fusion societies, mate promiscuously and express high levels of male-male competition. Females show a striking visual signal, an exaggerated sexual swelling, known to be a proxy of ovulation, while the maximum swelling is not matching the exact time of ovulation. The question remains if males use additional olfactory cues when being able to approach females closely. This would allow males to pinpoint the exact timing of ovulation and could be one reason why high-ranking males sire offspring more successfully than other males. Here, we present the first systematic test of such multimodal signaling by investigating male sniffing behavior directed to females in relation to their fertility in a group of 13 captive chimpanzees. Our results show that male sniffing behavior significantly increased with female swelling size, with female age as well as when fewer male competitors were present. Hence, odors might be part of a multimodal fertility cue, supporting the idea that males monitor both visual and olfactory cues to gain comprehensive information on female fertility.
Introduction
Throughout the animal kingdom, females and males have evolved different strategies to optimize their own reproductive outcome (Trivers, 1972; Clutton-Brock, 1989). Reproductive strategies, in fact, are often conflicting between the two sexes, leading to an “arms race” between females and males (Engelhardt et al., 2006). Depending on the ecology and mating system of species, various behavioral, anatomical, and physiological traits can be observed with which both sexes try to increase their own fitness (reviewed in Stumpf et al., 2011). For example, in multi-male multi-female groups with male dominance, males compete over access to females by trying, e.g., to monopolize fertile females and females aiming to confuse paternity by mating with several males, while at the same time reproducing with the “best” potential mates (e.g., in terms of high genetic quality or paternal investment).
To achieve their respective reproductive aims, both sexes use cues or signals of reproductive quality and reproductive state in multiple modalities, acting alone or as multimodal cues. While “cues” did not evolve to serve a communicative function, “signals” did evolve to cause a behavioral or physiological change in the recipient (Smith and Harper, 2003). It is well-known that visual and acoustic traits such as sexual swellings or copulation calls can indicate the fertile period in female primates (Cebuella pygmaea: Converse et al., 1995; Microcebus murinus: Buesching et al., 1998). Also olfactory cues can transport a wealth of information about individuals but studies investigating olfactory traits of female fertility are comparatively underrepresented in primates (Drea, 2015). For non-human primates, evidence for olfactory fertility cues predominantly comes from scent-marking strepsirrhine primates and New World monkeys. For instance, when males encountered the odor of fertile females, they reacted with adapted behavior like increased rates of investigation (Galago crassicaudatus: Clark, 1982; Saguinus oedipus: Ziegler et al., 1993; Callithrix jacchus: Smith and Abbott, 1998; Lemur catta: Scordato and Drea, 2007), neuronal activity and increased hormone levels (Callithrix jacchus: Ziegler et al., 2005; Snowdon et al., 2006). In common marmosets, the chemical composition of anogenital odors varied with female fertility and according bioassays provided evidence that males prefer the odor of fertile females compared to non-fertile females (Callithrix jacchus: Kücklich et al., 2019). The few existing studies on Old World monkeys suggest that males can perceive olfactory cues of female fertility and inspect females more often during the fertile period (Papio ursinus: Clarke et al., 2009; Papio anubis: Rigaill et al., 2013; Macaca arctoides: Cerda-Molina et al., 2006a,b; Macaca mulatta: Michael et al., 1971). In humans, men were able to distinguish fertile from non-fertile women using axillary body odors, with the odors from fertile women perceived as most attractive and least intense (e.g., Havliček et al., 2006; Gildersleeve et al., 2012). Furthermore, Michael et al. (1974, 1975) analyzed vaginal secretions of women and discovered a change in volatile aliphatic acids over the menstrual cycle, which may thus underlie men’s odor preferences for fertile women.
To date, our knowledge about the importance of olfactory communication is particularly scarce in great apes. It is known that great apes use their sense of smell to collect information about conspecifics (Matsumoto-Oda et al., 2007; Hepper and Wells, 2010; Klailova and Lee, 2014). A comparative study (Jänig et al., 2018) showed that male chimpanzees sniff more often at conspecifics than females or males of other great ape species (gorillas, orangutans, and bonobos). Analysis of great ape body odor revealed that the richness, intensity, and chemical composition varies between species (bonobos, chimpanzees, gorillas, and orangutans), but also with age (Jänig et al., 2019). However, olfactory fertility cues have not been examined well. It was observed that male chimpanzees inspect females’ genitals (Nishida, 1997; Emery Thompson and Wrangham, 2008) and it was suggested that they might try to collect olfactory cues on the fertility status of the females (Wallis, 1992; Nishida, 1997). Chemical analyses of vaginal secretions of female chimpanzees revealed the presence of substances which were previously found to vary during the menstrual cycle in rhesus macaques and humans (Macaca mulatta: Michael et al., 1971; humans: Michael et al., 1974, 1975), but cycle-related variation could not yet be confirmed in chimpanzees (Fox, 1982; Matsumoto-Oda et al., 2003). Overall, the few existing observations suggest that olfactory cues might be of relevance in great apes and play a particular role for communication in chimpanzees.
Chimpanzees (Pan troglodytes) are particularly interesting to study cues and signals of reproduction involving multiple modalities (or multimodal communication). They form complex fission-fusion groups (Nishida, 1968; Goodall, 1986) and females mate promiscuously during an extended period of receptivity (Tutin, 1980). It has been suggested that the predominant female reproductive strategy is to distribute the probability of paternity across several mating partners (Wrangham, 2002; Kappeler and van Schaik, 2004) to benefit from a reduced risk of infanticide (Stumpf and Boesch, 2006). This may be achieved best when the displayed information on when ovulation actually occurs is imprecise. Indeed, female chimpanzees show several cues and signals with different levels of precision that could potentially be indicative of their fertility, including copulation calls (Townsend et al., 2011), proceptive behavior as well as mating resistance behavior (Stumpf and Boesch, 2006). The most prominent advertisement of fertility in female chimpanzees, however, is an exaggerated anogenital swelling. Sexual swellings depend on estrogen levels and last 10–12 days (Tutin, 1980; Wallis, 1997). However, ovulation may occur within several days of the phase of maximal swelling and importantly, ovulation cannot be determined exactly from swelling size alone (Emery Thompson, 2005). Even though the frequency of copulations is highest when females are fully swollen, mating also occurs outside this period (Wallis, 1982).
The competition between male chimpanzees over access to females is intense (Goodall, 1986; Muller, 2002) and high-ranking males monopolize most copulations (Klinkova et al., 2005). Several females of a group can ovulate at a time (Goodall, 1986), limiting opportunities of male monopolization. Chimpanzee party size was found to be influenced, amongst other factors, by the presence of swollen females (Tutin, 1979), with males preferably joining parties with fully swollen females (Matsumoto-Oda, 1999), and male sexual behavior changed in correspondence to the size of the sexual swelling (Klinkova et al., 2005). On average, mating frequency closely follows the estrogen profile: it increases with increasing estrogen levels as well as swelling size and decreases with decreasing estrogen levels and swelling size, thus correlating with the most fertile days (Deschner et al., 2004; Emery Thompson, 2005). Interestingly, the rate of genital inspections increases before and with the onset of the maximal sexual swelling, suggesting that males might try to perceive olfactory cues in addition to the visual cue (Wallis, 1992; Nishida, 1997). To optimize their mating efforts and increase the chance of siring offspring, males would benefit from investing into mating with females primarily during their most fertile days, which could be achieved when a more precise indicator of fertility than the visual signal is available.
Indeed, observations suggest that, in addition to the imprecise visual signal of swellings, a more precise cue of ovulation could exist (Emery Thompson and Wrangham, 2008). Some males, particularly high-ranking males, seem to adjust their behavior more precisely to the actual time of ovulation. Alpha males were found to increase copulation frequency 1 day before the start of the peri-ovulatory period and copulation frequency was highest during the fertile days within the phase of maximal swelling (Deschner et al., 2004). Although females mate with several males during their maximal swelling stage, alpha males sire the majority of offspring (Constable et al., 2001; Klinkova et al., 2005). This skewed distribution of mating success suggests that high-ranking males may have better access to females, but possibly also to more detailed information on female fertility. Male chimpanzees may have evolved abilities to pinpoint an approaching ovulation using cues and signals of multiple sensory modalities with varying precision. If this is the case, the visual signal could first be used as an obvious but only approximate advertisement available for all males. High-ranking males, which are able to approach and monopolize females, may then use olfactory cues when inspecting the sexual swelling and gain more precise information. Since male chimpanzees frequently inspect swollen females (Matsumoto-Oda et al., 2007; Jänig et al., 2018), we suggest that males indeed could use olfactory cues. If only high-ranking males get access to these precise information, thus increasing their probability of paternity success, females would also benefit, because their offspring acquires the best genes (Kappeler and van Schaik, 2004) while they still maintain a high level of paternity confusion.
Our general aim of the current study was to provide an insight into olfaction in the reproductive context of chimpanzees in order to fill the existing gap of knowledge. In particular, we intended to test the suggestion that olfactory fertility cues might exist in addition to the exaggerated visual signals by investigating males’ sniffing behavior directed toward females. Accordingly, we examined three properties that might affect the actual fertility of females: their (1) swelling stage, (2) cycle regularity, and (3) age. The swelling stage represents the females’ actual reproductive state within the menstrual cycle, with ovulation most likely occurring within the maximal swelling period (Deschner et al., 2004). The regularity of females’ menstrual cycle can be assumed to be an indicator for receptivity, with non-cycling females (e.g., lactating females or females receiving contraceptives) being less or non-receptive. We further wanted to consider age of the female in our study, because male chimpanzees copulate more often with older than with younger females or preferably join parties with older females (Muller et al., 2006). Age seems to reflect reproductive experience (Muller et al., 2006), with older females being more experienced. We observed the frequency at which male chimpanzees sniff at females and examined whether male sniffing frequency varied with the described reproductive properties of females. Since male chimpanzees compete intensively and the access to females is limited, we furthermore argue that the number of males present at a time influences the rate of olfactory inspection. We suggest that the levels of competition rise when more males are around and that each male thus has fewer opportunities to sniff at a female. Assuming that males try to optimize their mating efforts, we predicted that male chimpanzees sniff more often at females (1) during their maximal swelling period, (2) when they are cycling regularly (are not pregnant or lactating), (3) when they have more reproductive experience (at an advanced age), and (4) when fewer competitors are around.
Materials and Methods
Subjects
We conducted our study on a group of captive chimpanzees (Pan troglodytes verus) at the Wolfgang Köhler Primate Research Centre (WKPRC) in the Leipzig Zoo, Germany. The group composition varied between our two observational periods (2 months in 2013 and 3 months in 2016). In total, we observed sniffing behavior in six males (age 4–41 years) and seven females (age 8–40 years). Four of the females had already reproduced multiple times with one of the males in the group (based on paternity analysis, unpubl. data Zoo Leipzig). None of the females was pregnant during our study period, but in 2016, two females were lactating. Some of the individuals were related (e.g., half-brothers and half-sisters, one mother-son, and one father-daughter-dyad). Since the sample size was low, we did not include relatedness between individuals in our statistical analysis.
The group further comprised regularly cycling females and females on contraception (Implanon® implants). Details on group composition, individuals and cycle regularity can be found in Table 1.
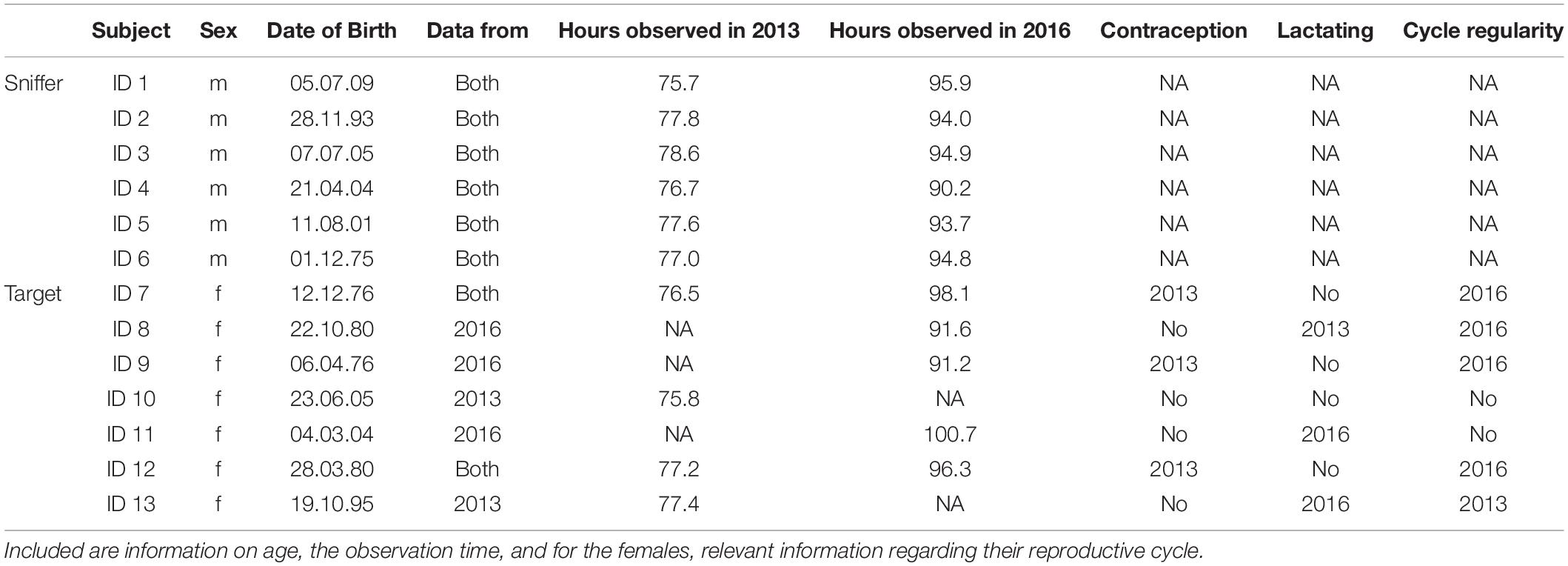
Table 1. Individuals observed as sniffers (males) and targets (females).
We included adult females in the observational study for which we could identify at least two different swelling stages in a respective observation year irrespective of their reproductive state (i.e., including lactating and contracepted females; for details see Table 1). Two of the females included in this study were quite young (8 and 9 years old, respectively), but studies in captivity showed that the first birth can be as early as 9.8 years of age (Coe et al., 1979; Littleton, 2005). As the two females further showed clear variance in their swelling size, we included them in our analysis.
Two of the males included in this analysis were 8 and 9 years old during sample collection. As studies showed that the onset of puberty can be earlier in male captive chimpanzees (Behringer et al., 2014) and the two males regularly showed mating behavior, we included them in our study. Another male was only 4 years old and thus clearly not being adult, however, this juvenile male regularly sniffed at the females. All adult males were regularly observed to copulate with females; however, older males had more reproductive experience than younger ones. The oldest male had reproduced several times already, while the younger ones had not reproduced yet.
Compliance With Ethical Standards
Statement of the Welfare of Animals
This study is in accordance with the legal requirements of Germany, all national and institutional guidelines for the care and use of animals, and was approved by the ethics commission of the Department of Psychology of the Max Planck Institute for Evolutionary Anthropology and the Leipzig Zoo.
Recording of Swelling Stage and Menstrual Cycle Phases
Sexual swelling size (flat, increasing, maximum and decreasing swelling, as defined in Deschner et al., 2003) of the female chimpanzees was recorded on a daily basis by their regular animal keepers to ensure reliability of categorization. In our study group, mean cycle length was 35.6 ± 9.1 days (from first day in increasing swelling until last day of flat phase) and thus within the ranges previously published for chimpanzees (captive: Nadler et al., 1985; Wallis, 1997; wild: Tutin, 1979; Emery Thompson, 2005). On average, the observed females were detumescent for 16.7 ± 5.4 days, the swelling increased in size for 4.3 ± 5.5 days, remained in maximal tumescence for 12.3 ± 3.4 days and decreased in size for 2.4 ± 1.5 days. Thus, durations of swelling stages resembled data reported from the wild (Deschner et al., 2004). Due to population control, three females included in this study received contraception in one observation period (Implanon® implant for 3 years). For lactating females and those receiving contraceptives we could not observe regular changes in the swelling size, i.e., for example the sexual skin normally remained flat, although some of these females sometimes expressed irregular, comparatively smaller genital swellings. We could not observe all four swelling stages during our observation periods in those females. They were regarded as non-cycling in comparison to females with a regular cycle. Data on swelling size per female per observation are provided in the stored data set.
Behavioral Observation
Sniffing behavior was defined as placing the nose close (≤ 3 cm) to another individual or touching an individual and immediately bringing the hand close to the nose (Zschoke and Thomsen, 2014). As sniffs were relatively rare, we used all-occurrence sampling on the whole group (Altmann, 1974). Observing the whole group was feasible due to elevated observation platforms allowing an optimal overview over the entire group, and because individuals were frequently resting, which made the movement of chimpanzees toward other individual clearly discernible.
We recorded all sniffs in social contexts, i.e., targeted from one individual toward another, and the respective contextual information (group composition, social occurrences such as fights, fusion events etc.), ID of sniffer and recipient (individual at which was sniffed), and body part at which was sniffed. All observers were blind to the specific research questions during data collection. Two observers recorded sniffing behavior for 80 h in April/May 2013 and two others observed the group for 104 h in September/October 2016 (in total 184 h). To achieve consistent observations, SJ trained all observers and conducted inter-observer reliability checks (Cohens Kappa, Hallgren, 2012) with each of them. Cohens Kappa revealed a satisfactory agreement between all observers in both observation periods (Cohens K = 0.86), indicating that observers highly matched in recording the same sniffing event, identifying the participating individuals and all situational information.
We observed sniffing behavior in observation sessions of 60 min each. Each person conducted up to five sessions per day, but a maximum of two sessions in a row. The sessions were evenly distributed over the possible observation time, which ranged from 9:00 a.m. until 7:00 p.m. During morning hours, animals were sometimes temporarily absent due to behavioral studies conducted by researchers of the WKPRC in separate study rooms. The individuals voluntarily chose to leave the group to attend the studies and came back into the groups after the study was finished. We here termed these occasions “fusion events” (as a different set of individuals or male competitors was available), which thus introduced fission-fusion situations similar to wild populations. During each protocol, it was noted which and for how long each group member could be observed during a given observational session.
Statistical Analysis of Male Sniffing Behavior
Our main interest was to compare male sniffing frequency directed toward a given female depending on three reproductive properties of the females (i.e., swelling stage, cycle regularity and age). We determined the number of sniffs a given female received from all six males within the respective 60 min observation session, whereby we used the summed value across males to keep the number of zeros in the response variable within an acceptable range. As we aim at analyzing olfactory inspections in a reproductive context, one might argue that a juvenile male might not intend to achieve information on female fertility when inspecting females. Thus, we also ran an additional model only for the adult males (N = 5) to account for a potential bias in the data caused by the juvenile male. Since the patterns for both models were predominantly the same (full-null model comparison (LRT): χ2 = 19.041, df = 6, P = 0.004; for details see Supplementary Material), we will further discuss the model containing all six males. We modeled the impact of female reproductive properties and group characteristics on the number of sniffs by males per female and session (832 data points obtained from up to six males and seven females in 184 sessions) using a negative-binomial Generalized Linear Mixed Model (GLMM) with the function glmer.nb of the package lme4 (Bates et al., 2015) in R version 3.2.3 (R Core Team, 2013). Swelling stage was included as the first categorical test predictor (flat, increasing, maximal, decreasing). We considered cycle regularity as a second test predictor, taking into account whether the menstrual cycle was regular (defined as all four swelling stages occurring repeatedly) or whether females were not cycling, as we expected males to show an increasing interest in females with a regular cycle rather than non-cycling females (i.e., lactating females or females on contraception). To incorporate reproductive experience, we had originally intended to include parity (how many infants a female has given birth to) and age, respectively. However, the factors parity and age were confounded, as in our study older females had produced several offspring while the young females were mainly nulli- or primiparous. Thus, we could not incorporate both factors and rather included age as the third test predictor as a proxy of both reproductive experience and potential (see also Proctor et al., 2011). In addition to the reproductive properties of females, we included the mean number of males present during a given observation session as a fourth test predictor, because we expected the frequency of sniffing events by any given male to depend on the number of male competitors simultaneously present in the group. The number of males could fluctuate because animals could leave the main enclosure to participate in parallel behavioral tests as mentioned above.
As random effects control predictors we added female ID (to control for individual variation), observation year (to control for the different observation periods and observers), observation day (to control for deviations between days) and a combination of female ID and observation day, since inspecting a particular female can vary between observation days due to other factors not considered here (e.g., stress or disease). We further included all possible random slopes, i.e., regular cycle, age, and swelling stage within an observation day, as well as swelling stage and number of males present within female ID, but we excluded correlations between random intercept and random slope for the latter because they were unidentifiable. To control for the time females were actually present in the enclosure during a given observation session, we included the amount of time a respective female was observed as a (log-transformed) offset term.
To make model estimates comparable and facilitate convergence of the models, female age, and number of males present were z-transformed. Model stability was established by comparing estimates derived from all data compared to data excluding levels one at a time. No obviously influential cases were revealed. Further, no issues were revealed when checking for potential collinearity using variance inflation factors (VIF) with VIFmax = 1.56 (Quinn and Keough, 2002; Field, 2005) calculated with the function vif of the package car (Fox and Weisberg, 2011). To test the overall effect of the test predictors, we compared the full model with a null model containing only the intercept, random terms and offset term using a Likelihood Ratio Test (Dobson, 2002; Forstmeier and Schielzeth, 2011). To determine p-values, DF, χ2 and Nagelkerke’s R2 for the individual test predictors, we ran reduced models lacking single test predictors and compared them to the full model using Likelihood Ratio Tests.
To control whether cyclic changes are more pronounced for sniffing behavior directed to the females’ genitals, we ran an additional model incorporating only those sniffs. Since the model was less robust due to the decreased number of observed sniffs and since the results are very similar to those of the overall model, we only present the results in Supplementary Material.
Results
In total, we recorded 1817 sniffs directed to conspecifics during 184 h of observation, with an overall mean sniffing frequency of 0.64 sniffs per individual per hour. More than half of these (53.8%) were directed by males toward females. When males sniffed at females, they focused on the genital region in 58.1% of all events, but inspected other body parts in 41.9% of all cases [mainly back (∼18%), head/face/mouth (∼14%), arm (∼7%), hands (∼2%), and chest (∼1%)]. We observed 21 sniffing events (∼2%) where individuals clearly sniffed body fluids (vomit, urine, saliva) or feces. While in 95.2% of the cases individuals directly sniffed at a conspecific, in 4.8% the individual touched the conspecific with its hand and sniffed the own hand afterwards.
In the model containing swelling stage, cycle regularity, female age and male presence we found a clear impact of the test predictors on the number of male sniffs directed toward females (full-null model comparison (LRT): χ2 = 19.174, df = 6, P = 0.0039). Specifically, swelling stage significantly affected sniffing frequencies (Table 2), whereby male chimpanzees preferably sniffed females that had an increasing or maximal swelling rather than females with a flat or decreasing swelling (Figure 1A). Males did not sniff regularly cycling females more often than females without a regular cycle (Table 2), but sniffed older females more often than younger females (Figure 1B and Table 2). Finally, the fewer male competitors were simultaneously present in the group (i.e., when some of them left the group to participate in behavioral studies), the more often males sniffed at females (Figure 1C and Table 2).
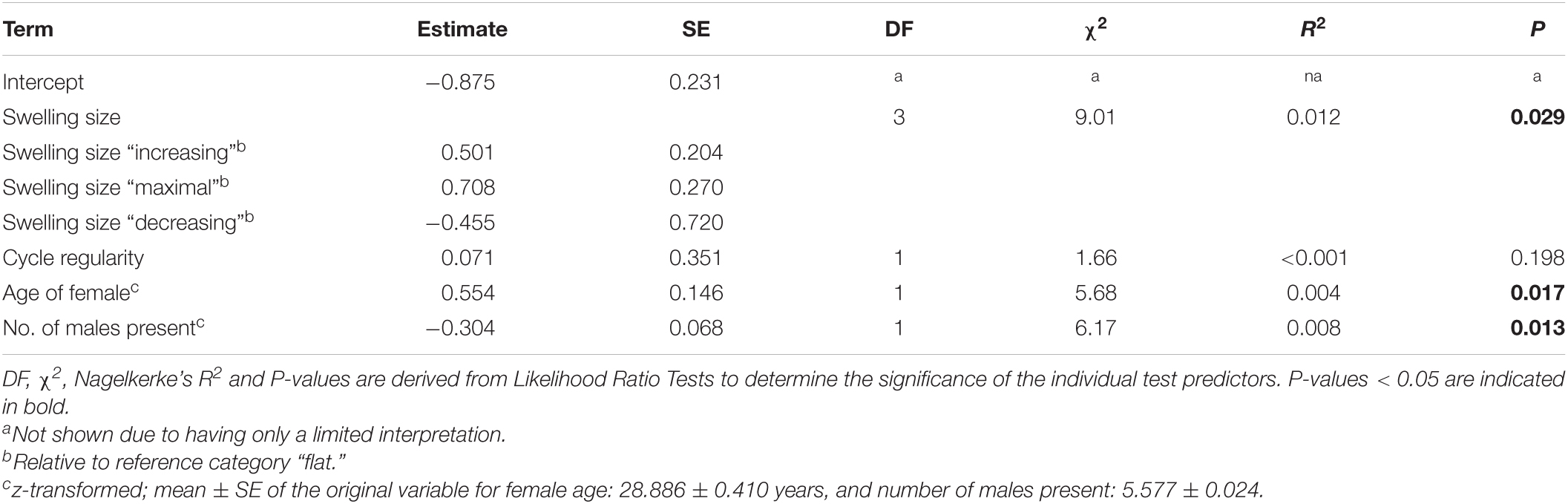
Table 2. Results of the negative-binomial GLMM with the number of sniffs females received by the six males as response variable and swelling size, cycle regularity, age of female, and male presence as test predictors.
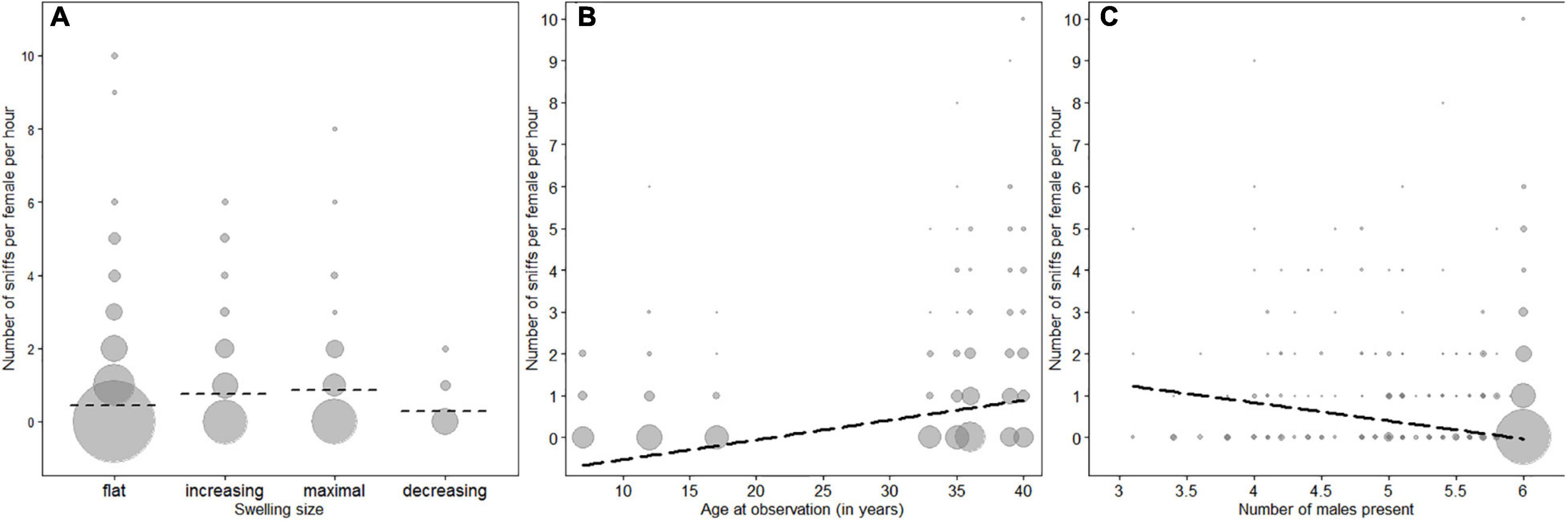
Figure 1. Frequency of sniffs observed per female per session. (A) Influence of the size of the sexual swelling, (B) impact of female age, (C) influence of the number of males present in the group. The size of the circles represents the number of observations; the dashed lines represent the model estimates.
Discussion
The results of our study indicate that sniffing behavior of chimpanzee males toward females is influenced by different female reproductive properties. Males sniffed more often at females with increasing swelling size, higher age, and when fewer male competitors were present, while cycle regularity did not influence male sniffing rates. Hence, our study strongly supports the idea that male chimpanzees use both visual and olfactory cues to assess female fertility.
The present study is in line with earlier observations in the wild that male chimpanzees use their sense of smell to inspect females (Matsumoto-Oda et al., 2007). The main finding of our study was that olfactory behavior of male chimpanzees was associated with a visual signal—the female anogenital swelling. Male sniffing frequency rose when female swelling size increased and decreased again after the maximal swelling period ended. Our findings are in line with patterns of male sociosexual behaviors, including olfactory inspection, that became more frequent with increasing sexual swellings of females in chimpanzees (Klinkova et al., 2005). Our result supports the hypothesis that males, in addition to using a visual signal (as indicated in Deschner et al., 2004), sniff at females according to their fertile states. Interestingly, male olfactory inspection was not restricted to the genital area of females, as males in our study sniffed at female body parts others than the genital area in half of the events we observed. This olfactory inspection of body and genital odor could potentially be used to pinpoint the timing of ovulation more precisely than is possible by visual inspection alone. Male chimpanzees preferentially join parties including females with increasing swellings (Matsumoto-Oda et al., 1998; Matsumoto-Oda, 1999; Boesch and Boesch-Achermann, 2000). Our results are supported by the findings of studies investigating the use of olfactory cues to assess female fertility in other primate species (Saguinus oedipus oedipus: Ziegler et al., 1993; Washabaugh and Snowdon, 1998; Macaca arctoides: Cerda-Molina et al., 2006b; Callithix jacchus: Kücklich et al., 2019).
Male chimpanzees altered their olfactory behavior also in relation to the age of females, i.e., males sniffed at older females more frequently than at younger females. Female reproductive potential of chimpanzees has been described to peak between the age of 14 and 24 years (representing the “younger” individuals in our study) and to sharply decline at the age of 25–30 years (Thompson et al., 2007). Based on this age-related decline in fertility, it would be reasonable that males prefer the younger and more fertile females. Nonetheless, in the wild, male chimpanzees seem to favor older over younger females, as suggested by higher copulation rates or greater likelihood to join a party in which older females are present (Muller et al., 2006), probably because they have more reproductive experience. Moreover, Thompson et al. (2007) found that chimpanzees staying healthy seemed to maintain a high fertility until later ages and the observed population-wide decline in fertility with increasing age was based on an increasing number of unhealthy individuals. In our study, age was correlated with parity (younger females were mostly nulliparous or primiparous, while all older females were multiparous) and thus the test predictor age comprises reproductive potential as well as reproductive experience at the same time. Indeed, previous studies found that nulliparous females were less attractive to males (Tutin, 1979; Hasegawa and Hiraiwa-Hasegawa, 1983). Multiparous females might be more interesting to the males because they could have observed these females reproducing and raising offspring successfully. Thus, for males, reproductive experience may be more important than reproductive potential, which remains to be investigated further in populations in which parity and age are less confounded than in our study group.
Relying on more than one sense is likely to be advantageous. Male chimpanzees might use both visual and olfactory cues to inspect females. The visual signal could act as an initial long-distance cue about the fertile state of a female, which may then elicit increased olfactory inspection from a short distance to gain additional and probably more precise information (Matsumoto-Oda et al., 2007). The olfactory information is potentially non-redundant to the visual one, because the information content might differ (see also Higham and Hebets, 2013). In olive baboons (Papio anubis), where females show variation in swelling stage, behavior, and copulation call rates during the fertile phase, males inspected female genitalia through sniffing more frequently in the fertile period as well (Rigaill et al., 2013). Based on these results, it was suggested that male baboons gather olfactory information on the onset of ovulation in addition to visual cues of the sexual swelling. The authors assumed that, while all males can access visual or auditory signals, only male consort partners are able to gain more information on the basis of odor cues obtained from the female in close spatial proximity to the male. Also in other primate species, it has been suggested that males that stay close to females, e.g., through mate-guarding, can obtain more and maybe more precise information about the onset of ovulation by using olfactory cues (Alouatta pigra: Van Belle et al., 2009; Papio anubis: Rigaill et al., 2013). In line with these suggestions, male chimpanzees regularly sniffing females might get more detailed information about the fertile state of the swollen female and thus could gain an advantage in siring offspring. However, it needs to be examined further, whether being close to females per se (triggered by the visual signal or not) elicits an increased rate of olfactory inspections or if males actually sniff more often at fully swollen females, compared to other (not fully swollen) females nearby. Moreover, it is not clarified whether the olfactory changes that males might gather by sniffing at females are “cues” or “signals.” Responses to cues are believed to need learning based on prior experience or conditioning, whereas signals provoke innate behavioral responses, although those innate responses could as well be modified through learning (reviewed in Beny and Kimchi, 2014). However, to unravel the underlying mechanisms, chemical analyses and bioassays would be needed.
Enhanced opportunities to gather olfactory information about females may be one reason why we observed individual males sniffing females more often when fewer male competitors were simultaneously present in the group. During our observations, the number of male competitors was sometimes reduced when the subjects volunteered in experimental studies outside the main enclosure. The resulting situation of leaving from and returning to the group can be regarded to reflect fission-fusion events similar to those in the wild. The temporal absence of certain males may have allowed the remaining males to get closer to females than in the presence of all male competitors. In this case, we would expect to observe increased rates of sniffing in lower-ranking males when higher-ranking males are absent. However, as we had no systematic dominance rank data available, further studies are necessary to examine this hypothesis.
Against our expectation, we found that sniffing frequencies of male chimpanzees did not differ between non-cycling and cycling females. Presumably, the visual signal, i.e., the presence (or absence) of a swelling, may have sufficed to induce high (or low) olfactory interest toward females irrespective of whether they showed full menstrual cycles or not. In fact, this supports the suggestion that the swelling provokes an increase in olfactory inspection, while information on whether a female is actually fertile or not may only become available from the olfactory inspection itself.
Overall, our study provides evidence that male olfactory behavior is associated with visual signs of female fertility in chimpanzees and therewith brings new insights into the field of great ape olfactory communication. However, certain aspects could not be addressed in the present study and these limitations should be investigated in future studies. First and foremost, this relates to hormone measurements which would allow us to pinpoint the timing of ovulation and thus refine our analysis. During the data collection for this study, we were not able to accomplish the extensive training of the individuals required for the collection of urine or saliva samples to investigate hormones. Second, sniffing data were collected from a single chimpanzee group with only six adult males. Given this limited sample size and scarce knowledge about olfactory fertility cues in catarrhine primates, further observations of other chimpanzee groups would help to compare to and confirm our findings. Additionally, systematic data on male and female dominance ranks or spatial proximity would allow differentiating between possible explanations for the results obtained here, and to widen the scope beyond the reproductive context. Despite these limitations, the data indicate that in a great ape species with an obvious visual signal of female fertility, males additionally appear to aim at collecting olfactory information, which sets a starting point for ongoing research and raises further questions about great ape olfactory communication in general and chimpanzee reproduction in specific. Finally, the chemical composition of female chimpanzees’ body odor, specifically perceivable chemical cues of female fertility, would complement the limited knowledge about great ape body odor.
Conclusion
In conclusion, our study associates olfactory behavior with female reproductive properties of a great ape species. Our results provide evidence that male chimpanzees sniffing behavior is closely linked to visual information on female fertility, which may allow males to collect information about female reproductive status using multiple modalities. Males in promiscuous species with varying group composition, where females provide only imprecise visual signals of fertility, could use additional olfactory cues to pinpoint the timing of ovulation and thus maximize their mating efforts to ultimately increase their reproductive success.
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics Statement
The animal study was reviewed and approved by the Ethics Commission of the Department of Psychology of the Max Planck Institute for Evolutionary Anthropology and the Leipzig Zoo.
Author Contributions
SJ, MK, BW, and AW conceived and planned the study. SJ and MZ carried out the observations and prepared the data. SJ, MK, LK, and BW planned and carried out the statistical analyses. SJ, MK, LK, BW, and AW contributed to the interpretation of the results. SJ and MK took the lead in writing the manuscript. All authors provided critical feedback and helped shape the research, analyses and manuscript.
Funding
This work was supported by the German Research Foundation within the Emmy-Noether Program (Grant #: WI 1808/3-1 awarded to AW) and the University of Leipzig. In addition, graduate funding by the University of Leipzig was awarded to SJ (DFPL R00061) and MK (DFPL R00017).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Josep Call and the curators of the Leipzig Zoo for the permission to sample the chimpanzees at the Wolfgang Köhler Primate Research Center (WKPRC) and are most grateful for the help of the keepers in the Leipzig Zoo and the staff at the WKPRC. We thank Ruth Thomsen for logistic support during sample collection, Katja Rudolph and Peggy Weist for help with behavioral observations and several reviewers for comments on earlier versions of the manuscript. Finally, we thank the Max Planck Institute for Evolutionary Anthropology and the University of Leipzig for their logistic support.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.884661/full#supplementary-material
References
Altmann, J. (1974). Observational Study of Behavior: Sampling Methods. Behaviour 49, 227–267. doi: 10.1163/156853974X00534
Bates, D., Maechler, M., Bolker, B., and Walker, S. (2015). Lme4: Linear Mixed-Effects Models Using Eigen and S4, R Package (version version 1.0-8). http://CRAN.R-project.org/package=lme4 (accessed May 19, 2022).
Behringer, V., Deschner, T., Deimel, C., Stevens, J. M. G., and Hohmann, G. (2014). Age-Related Changes in Urinary Testosterone Levels Suggest Differences in Puberty Onset and Divergent Life History Strategies in Bonobos and Chimpanzees. Horm. Behav. 66, 525–533. doi: 10.1016/j.yhbeh.2014.07.011
Beny, Y., and Kimchi, T. (2014). Innate and Learned Aspects of Pheromone-Mediated Social Behaviours. Anim. Behav. 97, 301–311. doi: 10.1016/j.anbehav.2014.09.014
Boesch, C., and Boesch-Achermann, H. (2000). The Chimpanzees of the Taï Forest: Behavioural Ecology and Evolution. Oxford: Oxford University Press.
Buesching, C. D., Heistermann, M., Hodges, J. K., and Zimmermann, E. (1998). Multimodal Oestrus Advertisement in a Small Nocturnal Prosimian, Microcebus Murinus. Folia Primatologica 69, 295–308. doi: 10.1159/000052718
Cerda-Molina, A. L., Hernandez-Lopez, L., Chavira, R., Cardenas, M., Paez-Ponce, D., and Cervantes-De Luz, H. La, et al. (2006a). Endocrine Changes in Male Stumptailed Macaques (Macaca Arctoides) as a Response to Odor Stimulation with Vaginal Secretions. Horm. Behav. 49, 81–87. doi: 10.1016/j.yhbeh.2005.04.014
Cerda-Molina, A. L., Hernandez-Lopez, L., Rojas-Maya, S., Murcia-Mejia, C., and Mondragon-Ceballos, R. (2006b). Male-Induced Sociosexual Behavior by Vaginal Secretions in Macaca Arctoides. Int. J. Primatol. 27, 791–807. doi: 10.1007/s10764-006-9045-0
Clark, A. B. (1982). Scent Marks as Social Signals in Galago Crassicaudatus I. Sex and Reproductive Status as Factors in Signals and Responses. J. Chem. Ecol. 8, 1133–1151. doi: 10.1007/BF00986984
Clarke, P. M. R., Barrett, L., and Henzi, S. P. (2009). What Role Do Olfactory Cues Play in Chacma Baboon Mating? Am. J. Primatol. 71, 493–502. doi: 10.1002/ajp.20678
Coe, C. L., Connolly, A. C., Kraemer, H. C., and Levine, S. (1979). Reproductive Development and Behavior of Captive Female Chimpanzees. Primates 20, 571–582. doi: 10.1007/BF02373439
Constable, J. L., Ashley, M. V., Goodall, J., and Pusey, A. E. (2001). Noninvasive Paternity Assignment in Gombe Chimpanzees. Mol. Ecol. 10, 1279–1300.
Converse, L. J., Carlson, A. A., Ziegler, T. E., and Snowdon, C. T. (1995). Communication of Ovulatory State to Mates by Female Pygmy Marmosets, Cebuella Pygmaea. Anim. Behav. 49, 615–621. doi: 10.1016/0003-3472(95)80194-4
Deschner, T., Heistermann, M., Hodges, K., and Boesch, C. (2003). Timing and Probability of Ovulation in Relation to Sex Skin Swelling in Wild West African Chimpanzees, Pan Troglodytes Verus. Anim. Behav. 66, 551–560. doi: 10.1006/anbe.2003.2210
Deschner, T., Heistermann, M., Hodges, K., and Boesch, C. (2004). Female Sexual Swelling Size, Timing of Ovulation, and Male Behavior in Wild West African Chimpanzees. Horm. Behav. 46, 204–215. doi: 10.1016/j.yhbeh.2004.03.013
Dobson, A. J. (2002). An Introduction to Generalized Linear Models. 2nd ed. Texts in Statistical Science. Boca Raton: CRC Press.
Drea, C. M. (2015). D’scent of Man: A Comparative Survey of Primate Chemosignaling in Relation to Sex. Horm. Behav. 68, 117–133. doi: 10.1016/j.yhbeh.2014.08.001
Emery Thompson, M. (2005). Reproductive Endocrinology of Wild Female Chimpanzees (Pan Troglodytes Schweinfurthii): Methodological Considerations and the Role of Hormones in Sex and Conception. Am. J. Primatol. 67, 137–158. doi: 10.1002/ajp.20174
Emery Thompson, M., and Wrangham, R. W. (2008). Male Mating Interest Varies with Female Fecundity in Pan Troglodytes Schweinfurthii of Kanyawara, Kibale National Park. Int. J. Primatol. 29, 885–905. doi: 10.1007/s10764-008-9286-1
Engelhardt, A., Heistermann, M., Hodges, J. K., Nuernberg, P., and Niemitz, C. (2006). Determinants of Male Reproductive Success in Wild Long-Tailed Macaques (Macaca Fascicularis)—Male Monopolisation, Female Mate Choice or Post-Copulatory Mechanisms? Behav. Ecol. Sociobiol. 59, 740–752. doi: 10.1007/s00265-005-0104-x
Forstmeier, W., and Schielzeth, H. (2011). Cryptic Multiple Hypotheses Testing in Linear Models: Overestimated Effect Sizes and the Winner’s Curse. Behav. Ecol. Sociobiol. 65, 47–55. doi: 10.1007/s00265-010-1038-5
Fox, G. J. (1982). Potentials for Pheromones in Chimpanzee Vaginal Fatty Acids. Folia Primatologica 37, 255–266. doi: 10.1159/000156036
Fox, J., and Weisberg, S. (2011). An {R} Companion to Applied Regression (version Second Edition). Thousand Oaks CA: Sage.
Gildersleeve, K. A., Haselton, M. G., Larson, C. M., and Pillsworth, E. G. (2012). Body Odor Attractiveness as a Cue of Impending Ovulation in Women: Evidence from a Study Using Hormone-Confirmed Ovulation. Horm. Behav. 61, 157–166. doi: 10.1016/j.yhbeh.2011.11.005
Goodall, J. (1986). The Chimpanzees of Gombe. Patterns of Behavior. Cambridge, MA: Belknap Press of Harvard University.
Hallgren, K. A. (2012). Computing Inter-Rater Reliability for Observational Data: An Overview and Tutorial. Tutor. Quant. Methods Psychol. 8, 23–34.
Hasegawa, T., and Hiraiwa-Hasegawa, M. (1983). Opportunistic and Restrictive Matings among Wild Chimpanzees in the Mahale Mountains, Tanzania. J. Ethol. 1, 75–85. doi: 10.1007/BF02347833
Havliček, J., Dvorakova, R., Bartoš, L., and Flegr, J. (2006). Non-Advertized Does Not Mean Concealed: Body Odour Changes across the Human Menstrual Cycle. Ethology 112, 81–90. doi: 10.1111/j.1439-0310.2006.01125.x
Hepper, P. G., and Wells, D. L. (2010). Individually Identifiable Body Odors Are Produced by the Gorilla and Discriminated by Humans. Chem. Senses 35, 263–268. doi: 10.1093/chemse/bjq015
Higham, J. P., and Hebets, E. A. (2013). An Introduction to Multimodal Communication. Behav. Ecol. Sociobiol. 67, 1381–1388. doi: 10.1007/s00265-013-1590-x
Jänig, S., Weiß, B. M., and Widdig, A. (2018). Comparing the Sniffing Behavior of Great Apes. Am. J. Primatol. 80:e22872. doi: 10.1002/ajp.22872
Jänig, S., Weiß, B. M., Birkemeyer, C., and Widdig, A. (2019). Comparative Chemical Analysis of Body Odor in Great Apes. Am. J. Primatol. 0:e22976. doi: 10.1002/ajp.22976
Kappeler, P. M., and van Schaik, C. (2004). Sexual Selection in Primates - New and Comparative Perspectives. Cambridge: Cambridge University Press.
Klailova, M., and Lee, P. C. (2014). Wild Western Lowland Gorillas Signal Selectively Using Odor. PLoS One 9:e99554. doi: 10.1371/journal.pone.0099554
Klinkova, E., Hodges, J. K., Fuhrmann, K., Jong, T., and Heistermann, M. (2005). Male Dominance Rank, Female Mate Choice and Male Mating and Reproductive Success in Captive Chimpanzees. Int. J. Primatol. 26, 357–484. doi: 10.1007/s10764-005-2929-6
Kücklich, M., Weiß, B. M., Birkemeyer, C., Einspanier, A., and Widdig, A. (2019). Chemical Cues of Female Fertility States in a Non-Human Primate. Sci. Rep. 9:13716. doi: 10.1038/s41598-019-50063-w
Littleton, J. (2005). Fifty Years of Chimpanzee Demography at Taronga Park Zoo. Am. J. Primatol. 67, 281–298. doi: 10.1002/ajp.20185
Matsumoto-Oda, A. (1999). Mahale Chimpanzees: Grouping Patterns and Cycling Females. Am. J. Primatol. 47, 197–207. doi: 10.1002/(SICI)1098-2345199947:3<197::AID-AJP2<3.0.CO;2-3
Matsumoto-Oda, A., Hosaka, K., Huffman, M. A., and Kawanaka, K. (1998). Factors Affecting Party Size in Chimpanzees of the Mahale Mountains. Int. J. Primatol. 19, 999–1011. doi: 10.1023/A:1020322203166
Matsumoto-Oda, A., Kutsukake, N., Hosaka, K., and Matsusaka, T. (2007). Sniffing Behaviors in Mahale Chimpanzees. Primates 48, 81–85. doi: 10.1007/s10329-006-0006-1
Matsumoto-Oda, A., Oda, R., Hayashi, Y., Murakami, H., Maeda, N., Kumazaki, K., et al. (2003). Vaginal Fatty Acids Produced by Chimpanzees during Menstrual Cycles. Folia Primatologica 74, 75–79. doi: 10.1159/000070000
Michael, R. P., Bonsall, R. W., and Kutner, M. (1975). Volatile Fatty Acids, ‘Copulins’, in Human Vaginal Secretions. Psychoneuroendocrinology 1, 153–163. doi: 10.1016/0306-4530(75)90007-4
Michael, R. P., Bonsall, R. W., and Warner, P. (1974). Human Vaginal Secretions: Volatile Fatty Acid Content. Science 186, 1217–1219.
Michael, R. P., Keverne, E. B., and Bonsall, R. W. (1971). Pheromones: Isolation of Male Sex Attractants from a Female Primate. Science 172, 964–966. doi: 10.1126/science.172.3986.964
Muller, M. N. (2002). “Agonistic Relations among Kanyawara Chimpanzees,” in Behavioural Diversity in Chimpanzees and Bonobos, eds C. Boesch, G. Hohmann, and L. Marchant (Cambridge: Cambridge University Press), 112–124. doi: 10.1017/CBO9780511606397.012
Muller, M. N., Emery Thompson, M., and Wrangham, R. W. (2006). Male Chimpanzees Prefer Mating with Old Females. Curr. Biol. 16, 2234–2238. doi: 10.1016/j.cub.2006.09.042
Nadler, R. D., Graham, C. E., Gosselin, R. E., and Collins, D. C. (1985). Serum Levels of Gonadotropins and Gonadal Steroids, Including Testosterone, during the Menstrual Cycle of the Chimpanzee (Pan Troglodytes). Am. J. Primatol. 9, 273–284. doi: 10.1002/ajp.1350090404
Nishida, T. (1968). The Social Group of Wild Chimpanzees in the Mahali Mountains. Primates 9, 167–224. doi: 10.1007/BF01730971
Nishida, T. (1997). Sexual Behavior of Adult Male Chimpanzees of the Mahale Mountains National Park, Tanzania. Primates 38, 379–398. doi: 10.1007/BF02381879
Proctor, D. P., Lambeth, S. P., Schapiro, S. J., and Brosnan, S. F. (2011). “Male Chimpanzees’ Grooming Rates Vary by Female Age, Parity, and Fertility Status. Am. J. Primatol. 73, 989–996. doi: 10.1002/ajp.20964
Quinn, G. P., and Keough, M. J. (2002). Experimental Design and Data Analysis for Biologists. 1st ed. Cambridge, UK: Cambridge University Press.
R Core Team (2013). R: A Language and Environment for Statistical Computing (version Version 2.15.3). Vienna: R Foundation for Statistical Computing.
Rigaill, L., Higham, J. P., Lee, P. C., Blin, A., and Garcia, C. (2013). Multimodal Sexual Signaling and Mating Behavior in Olive Baboons (Papio Anubis). Am. J. Primatol. 75, 774–787. doi: 10.1002/ajp.22154
Scordato, E., and Drea, C. M. (2007). Scents and Sensibility: Information Content of Olfactory Signals in the Ringtailed Lemur, Lemur Catta. Anim. Behav. 73, 301–314. doi: 10.1016/j.anbehav.2006.08.006
Smith, T. E., and Abbott, D. H. (1998). Behavioral Discrimination between Circumgenital Odor from Peri-Ovulatory Dominant and Anovulatory Female Common Marmosets (Callithrix Jacchus). Am. J. Primatol. 46, 265–284. doi: 10.1002/(SICI)1098-2345199846:4<265::AID-AJP1<3.0.CO;2-D
Snowdon, C. T., Ziegler, T. E., Schultz-Darken, N. J., and Ferris, C. F. (2006). Social Odours, Sexual Arousal and Pairbonding in Primates. Philos. Trans. R. Soc. B 361, 2079–2089. doi: 10.1098/rstb.2006.1932
Stumpf, R. M., and Boesch, C. (2006). The Efficacy of Female Choice in Chimpanzees of the Tai Forest, Cote d’Ivoire. Behav. Ecol. Sociobiol. 60, 749–765. doi: 10.1007/s00265-006-0219-8
Stumpf, R. M., Martinez-Mota, R., Milich, K. M., Righini, N., and Shattuck, M. R. (2011). Sexual Conflict in Primates. Evol. Anthropol. 20, 62–75. doi: 10.1002/evan.20297
Thompson, E. M., Jones, J. H., Pusey, A. E., Brewer-Marsden, S., Goodall, J., Marsden, D., et al. (2007). Aging and Fertility Patterns in Wild Chimpanzees Provide Insights into the Evolution of Menopause. Curr. Biol. 17, 2150–2156.
Townsend, S. W., Deschner, T., and Zuberbühler, K. (2011). Copulation Calls in Female Chimpanzees (Pan Troglodytes Schweinfurthii) Convey Identity but Do Not Accurately Reflect Fertility. Int. J. Primatol. 32, 914–923. doi: 10.1007/s10764-011-9510-2
Trivers, R. L. (1972). “Parental Investment and Sexual Selection,” in Sexual Selection and the Descent of Man, ed. B. Campbell (Chicago: Aldine), 139–179.
Tutin, C. E. (1980). Reproductive Behaviour of Wild Chimpanzees in the Gombe National Park, Tanzania. J. Reprod. Infertil. 28, 43–57.
Tutin, C. E. G. (1979). Mating Patterns and Reproductive Strategies in a Community of Wild Chimpanzees (Pan Troglodytes Schweinfurthii). Behav. Ecol. Sociobiol. 6, 29–38. doi: 10.1007/BF00293242
Van Belle, S., Estrada, A., Ziegler, T. E., and Strier, K. B. (2009). Sexual Behavior across Ovarian Cycles in Wild Black Howler Monkeys (Alouatta Pigra): Male Mate Guarding and Female Mate Choice. Am. J. Primatol. 71, 153–164. doi: 10.1002/ajp.20635
Wallis, J. (1982). Sexual Behavior of Captive Chimpanzees (Pan Troglodytes): Pregnant versus Cycling Females. Am. J. Primatol. 3, 77–88. doi: 10.1002/ajp.1350030107
Wallis, J. (1992). Chimpanzee Genital Swelling and Its Role in the Pattern of Sociosexual Behavior. Am. J. Primatol. 28, 101–113. doi: 10.1002/ajp.1350280203
Wallis, J. (1997). A Survey of Reproductive Parameters in the Free-Ranging Chimpanzees of Gombe National Park. J. Reprod. Infertil. 109, 297–307.
Washabaugh, K., and Snowdon, C. T. (1998). Chemical Communication of Reproductive Status in Female Cotton-Top Tamarins (Saguinus Oedipus Oedipus). Am. J. Primatol. 45, 337–349. doi: 10.1002/(SICI)1098-2345199845:4<337::AID-AJP2<3.0.CO;2-X
Wrangham, R. W. (2002). “The Cost of Sexual Attraction: Is There a Tradeoff in Female Pan between Sex Appeal and Received Coercion?,” in Behavioural Diversity in Chimpanzees and Bonobos, eds C. Boesch, G. Hohmann, and L. Marchant (Cambridge, UK: Cambridge University Press), 204–215.
Ziegler, T. E., Epple, G., Snowdon, C. T., Porter, T. A., Belcher, A. M., and Küderling, I. (1993). Detection of the Chemical Signals of Ovulation in the Cotton-Top Tamarin, Saguinus Oedipus. Anim. Behav. 45, 313–322. doi: 10.1006/anbe.1993.1036
Ziegler, T. E., Schultz-Darken, N. J., Scott, J. J., Snowdon, C. T., and Ferris, C. F. (2005). Neuroendocrine Response to Female Ovulatory Odors Depends upon Social Condition in Male Common Marmosets, Callithrix Jacchus. Horm. Behav. 47, 56–64. doi: 10.1016/j.yhbeh.2004.08.009
Keywords: olfactory communication, odor, sniffing, signaling, menstrual cycle, anogenital swelling
Citation: Jänig S, Kücklich M, Kulik L, Zetzsche M, Weiß BM and Widdig A (2022) Olfactory Inspection of Female Reproductive States in Chimpanzees. Front. Ecol. Evol. 10:884661. doi: 10.3389/fevo.2022.884661
Received: 26 February 2022; Accepted: 16 June 2022;
Published: 05 July 2022.
Edited by:
Luisa Amo, Universidad Rey Juan Carlos, SpainReviewed by:
Jan Havlicek, Charles University, CzechiaMaría Abellán Álvaro, University of Valencia, Spain
Copyright © 2022 Jänig, Kücklich, Kulik, Zetzsche, Weiß and Widdig. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Susann Jänig, c3VzYW5uX2phZW5pZ0BldmEubXBnLmRl
†These authors share first authorship
‡These authors share last authorship