
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 21 April 2022
Sec. Paleontology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.881651
This article is part of the Research TopicHuman Behavior, Cognition, and Environmental Interactions for the Lower PaleolithicView all 10 articles
 Paul Palmqvist1*
Paul Palmqvist1* Guillermo Rodríguez-Gómez1,2,3
Guillermo Rodríguez-Gómez1,2,3 José María Bermúdez de Castro4,5
José María Bermúdez de Castro4,5 José Manuel García-Aguilar1
José Manuel García-Aguilar1 M. Patrocinio Espigares1
M. Patrocinio Espigares1 Borja Figueirido1
Borja Figueirido1 Sergio Ros-Montoya1
Sergio Ros-Montoya1 Alejandro Granados1
Alejandro Granados1 Francisco J. Serrano1,6
Francisco J. Serrano1,6 Bienvenido Martínez-Navarro7,8,9
Bienvenido Martínez-Navarro7,8,9 Antonio Guerra-Merchán1
Antonio Guerra-Merchán1
The chronology and environmental context of the first hominin dispersal in Europe have been subject to debate and controversy. The oldest settlements in Eurasia (e.g., Dmanisi, ∼1.8 Ma) suggest a scenario in which the Caucasus and southern Asia were occupied ∼0.4 Ma before the first peopling of Europe. Barranco León (BL) and Fuente Nueva 3 (FN3), two Early Pleistocene archeological localities dated to ∼1.4 Ma in Orce (Guadix-Baza Depression, SE Spain), provide the oldest evidence of hominin presence in Western Europe. At these sites, huge assemblages of large mammals with evidence of butchery and marrow processing have been unearthed associated to abundant Oldowan tools and a deciduous tooth of Homo sp. in the case of BL. Here, we: (i) review the Early Pleistocene archeological sites of Europe; (ii) discuss on the subsistence strategies of these hominins, including new estimates of resource abundance for the populations of Atapuerca and Orce; (iii) use cartographic data of the sedimentary deposits for reconstructing the landscape habitable in Guadix-Baza; and (iv) calculate the size of the hominin population using an estimate of population density based on resource abundance. Our results indicate that Guadix-Baza could be home for a small hominin population of 350–280 individuals. This basin is surrounded by the highest mountainous reliefs of the Alpine-Betic orogen and shows a limited number of connecting corridors with the surrounding areas, which could have limited gene flow with other hominin populations. Isolation would eventually lead to bottlenecks, genetic drift and inbreeding depression, conditions documented in the wild dog population of the basin, which probably compromised the viability of the hominin population in the medium to long term. This explains the discontinuous nature of the archeological record in Guadix-Baza, a situation that can also be extrapolated to the scarcity of hominin settlements for these ancient chronologies in Europe.
The chronology of the first hominin settlements in Europe has been subject to debate and controversy. Until the mid 1990s most paleoanthropologists believed that there was no significant habitation before 0.6–0.5 Ma (e.g., Aragó, Bilzingsleben, Boxgrove, Ceprano, Mauer, and Verteszöllos), with most sites concentrating on <0.45 Ma (e.g., Atapuerca’s Sima de los Huesos, Petralona, Schöningen, Swanscombe, and Steinheim) (Carbonell and Rodríguez, 1994; Roebroeks and van Kolfschoten, 1994), an increase in the number of occupations that coincided with interglacial MIS 11 (Blain et al., 2021). This “young chronology” was shortly challenged by new findings in Europe (Carbonell et al., 1995; Ascenzi et al., 1996; Bermúdez de Castro et al., 1997; Martínez-Navarro et al., 1997), the Caucasus (Gabunia and Vekua, 1995) and China (Wanpo et al., 1995; Larick and Ciochon, 1996), as well as by the geochronological re-evaluation of the evidence from Java (Swisher et al., 1994; Larick et al., 2001; Hyodo et al., 2011), which all indicated an earlier hominin arrival in Eurasia, during late Early Pleistocene times (Arribas and Palmqvist, 1999). However, although the archeological record in Europe has improved over the last decades, it remains highly fragmentary for these ancient chronologies and many sites lack high-resolution chronostratigraphic frameworks, which raises doubts on the age of the earliest hominin settlements. For example, a re-evaluation of the magnetostratigraphic and radiometric age constraints on several key sites bearing hominin remains and/or lithic tools from southern Europe led Muttoni et al. (2010, 2013) to propose that the first hominin dispersal in Western Europe took place during the Matuyama reverse polarity chron, between the Jaramillo normal polarity subchron and the Brunhes-Matuyama boundary.
Ancient evidence of hominin presence in Western Europe, The Caucasus, the Levantine Corridor, and Northern Africa is now well documented in a number of Early Pleistocene archeological localities (see Figure 1 and references therein). This provides a chronological scenario for the oldest hominin settlements in Eurasia in which the Caucasus (Dmanisi, ∼1.8 Ma) and southern Asia were occupied ∼400 ka before the first peopling of Europe (Arribas and Palmqvist, 1999; Dennell and Roebroeks, 2006; Mosquera et al., 2013). It has been proposed that this delay could result from competition with other carnivores: hominin survival at the middle latitudes, where plant resources are scarce during the cold season, would depend on the regular scavenging of ungulate carcasses (Turner, 1992; Palmqvist et al., 2022a). For this reason, Rodríguez-Gómez et al. (2017a) estimated the level of competition for meat among the members of the carnivore guild of Venta Micena (VM), a site dated by biochronology and paleomagnetism to 1.6–1.5 Ma with no conclusive evidence on hominin presence (Arribas and Palmqvist, 2002; Martínez-Navarro, 2002; Palmqvist et al., 2005). This study provided estimates on meat availability and competition intensity for VM that were higher and lower, respectively, than those obtained for Barranco León (BL) and Fuente Nueva 3 (FN3) (Rodríguez-Gómez et al., 2016a). Given that BL and FN3 are 200–100 ka younger than VM, this suggests that the delay in the colonization of Europe was not a matter of ecological opportunity and other factors (e.g., climatic and/or geographic barriers to dispersal) should be considered (Rodríguez-Gómez et al., 2017a).
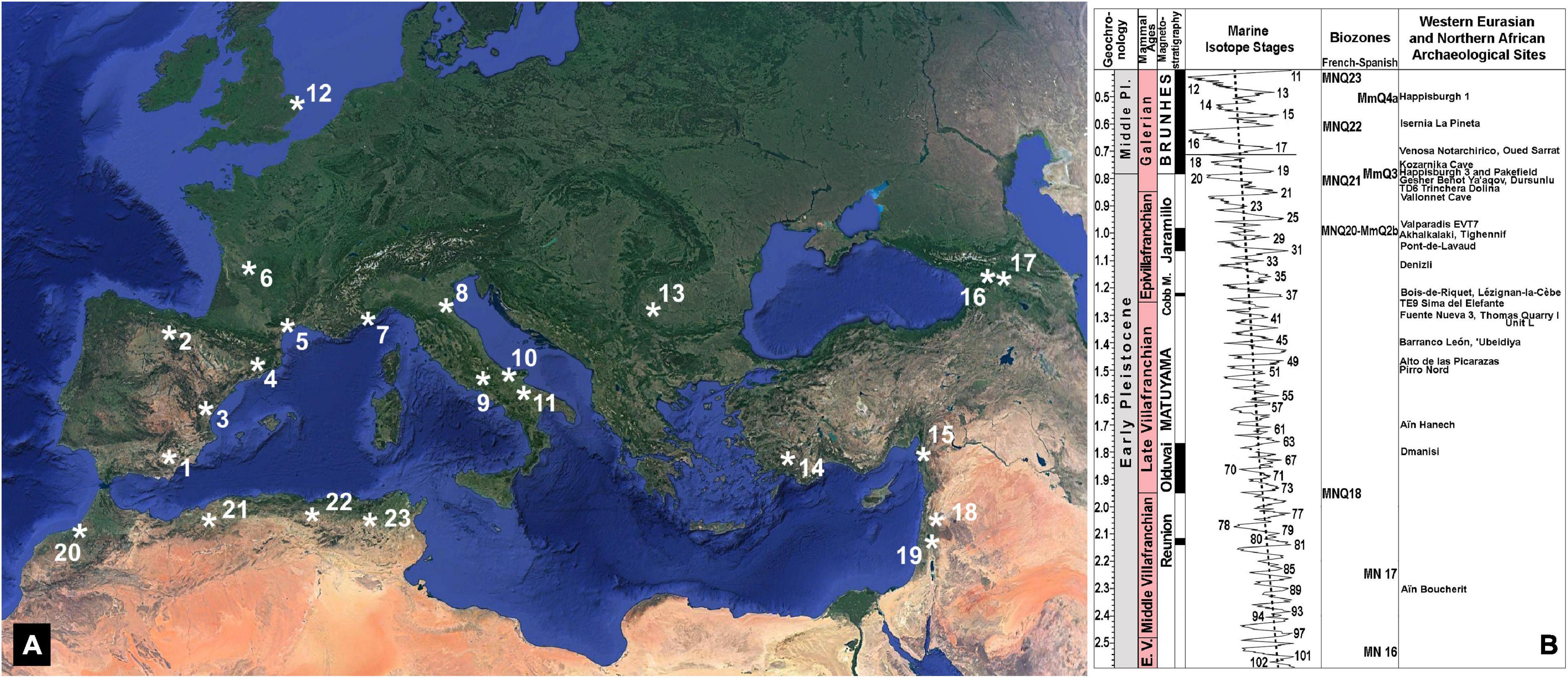
Figure 1. (A) Geographic location of selected Early Pleistocene to early Middle Pleistocene archeological sites of Europe, North Africa, the Levantine Corridor, and the Caucasus. 1: Barranco León (BL) and Fuente Nueva 3 (FN3), two sites of the Guadix-Baza Depression, Spain (1.5–1.2 Ma; Martínez-Navarro et al., 1997; Oms et al., 2000; Espigares et al., 2013, 2019; Toro-Moyano et al., 2013; Titton et al., 2021; Yravedra et al., 2021), which have yielded abundant Oldowan assemblages (∼3,500 artifacts) and evidence of butchery and marrow processing of large mammal bones, as well as a human deciduous tooth in BL. 2: level TE9 of Sima del Elefante (1.3–1.1 Ma; Carbonell et al., 2008) and level TD6 of Trinchera Dolina (∼0.85 Ma, MIS 21; Duval et al., 2018; Parés et al., 2018) in Atapuerca, Spain (a mandibular symphysis and 32 Oldowan artifacts, including four flakes, used for defleshing and marrow extraction have been unearthed in TE9). 3: Alto de las Picarazas, Spain (1.5–1.4 Ma; Vicente-Gabarda et al., 2016), which preserves >2,000 bone remains (several of them with butchery, percussion, and fracture marks) and seven lithic tools (two shapeless flint fragments and some splinters found during the sieving of sediments). 4: Vallparadís, Level EVT7, Spain (0.98–0.95 Ma, MIS 27; Martínez et al., 2015; but see, for a younger chronology of ∼0.85 Ma, Duval et al., 2015), which has yielded small-sized Oldowan tools (notches, becs, scrapers and denticulates on small pebbles, clasts, fragments, and flakes, as well as a large single chopper) elaborated from local raw materials and based on an anvil knapping technique (Garcia et al., 2013). 5: Bois-de-Riquet, Lézignan-la-Cèbe (1.3–1.1 Ma; Bourguignon et al., 2016; but see, for an older chronology, Crochet et al., 2009), which preserves lithic artifacts (177 basalt tools, although anthropically produced artifacts are difficult to differentiate from fragments or blocks detached naturally from the exfoliating surfaces enclosing the sedimentary level). 6: Pont-de-Lavaud (1.1–1.0 Ma; Voinchet et al., 2010), with an Oldowan assemblage that comprises ∼8,000 artifacts made exclusively on quartz pebbles and subangular vein quartz fragments, including ∼4,000 broken pebbles and 1,321 pieces with percussion marks and evidence of flaking with the bipolar-on-anvil technique (De Lombera-Hermida et al., 2016). 7: Vallonnet Cave (1.2–1.1 Ma; Michel et al., 2017; Cauche, 2022), with percussion tools, shaped pebbles, flakes, cores, and cut-marked bones. 8: Monte Poggiolo, Italy (∼0.85 Ma, within the reverse magnetic polarity subchron C1r.1r; Muttoni et al., 2011), which preserves an industry characterized by knapped pebbles and the products derived from their knapping, showing an almost total absence of retouched tools and the presence of some scrapers and denticulates that seem to be incidental and have no distinctive features (Peretto, 2006). 9: Isernia La Pineta, Italy (∼0.6 Ma; Coltorti et al., 2005), with flint and cherty limestone artifacts associated with remains of large mammals, characterized by the use of anvils in an opportunistic and rapid way to produce a large number of flakes and residual cores, usually of very small size. 10: Pirro Nord, Italy (1.6–1.3 Ma; Arzarello et al., 2007, 2015; Cheheb et al., 2019), where 5 out of 340 lithic artifacts preserve use-wear traces and are associated with more than one thousand vertebrate fossil remains, several with cut marks or evidence of intentional breakage. 11: Venosa Notarchirico, Italy (0.695–0.670 Ma, MIS 17; Moncel et al., 2020), which Oldowan industry is made on chert and includes flakes (mostly unretouched), broken flakes, debris, retouched nodules, cores, and limestone pebbles, as well as two bifacial tools and a handaxe. 12: Happisburgh 3 and Pakefield, United Kingdom (∼0.8 Ma; Lewis et al., 2019), which tool assemblages comprise flint flakes, flake tools, cores, and a handaxe in the case of Happisburgh 1 (∼0.5 Ma, MIS 13). 13: Kozarnika Cave, Layers 13a-c, Bulgaria (∼0.75 Ma; Muttoni et al., 2017), which has provided an abundant industry (∼10,000 artifacts, associated to a rich faunal assemblage) that shows a predominance of fragments from primary flaking (the local flint is very fragile), with the flakes obtained by simple unipolar to bipolar debitage (Sirakov et al., 2010). 14: Denizli, Turquey (1.2–1.1 Ma; Lebatard et al., 2014), which has provided the Kocabaş skull fragment, the only known Turkish fossil of Homo erectus. 15: Dursunlu, Turquey (0.99–0.78 Ma; Güleç et al., 2009), where remains of rhino, hippo and horse are found with 135 modified quartz implements. 16: Dmanisi, Georgia (∼1.8 Ma; García et al., 2010; Coil et al., 2020), a site that preserves the oldest evidence of human presence out of Africa (see text), with five hominin skulls and several postcranial bones (Gabunia and Vekua, 1995; Lordkipanidze et al., 2005, 2013), a rich assemblage of Late Villafranchian mammals and large numbers of Oldowan artifacts made of basalt, andesite and tuffs; the tool assemblage includes cores (mostly unifacial), flakes and debris, which shows that all stages of flaking activity took place at the site (Mgeladze et al., 2011). 17: Akhalkalaki, Georgia (1.0–0.9 Ma; Vekua, 1986), with thousands of remains of large mammals, including the giant hippo H. antiquus (not present in Dmanisi), and artifacts associated to the fauna, which could be not contemporaneous with it (Tappen et al., 2002). 18: Gesher Benot Ya’aqov, Israel (∼0.78 Ma, MIS 19; Goren-Inbar et al., 2000), with an Acheulian assemblage composed of bifaces predominantly formed on basalt and core tools mostly made of flint. 19: ‘Ubeidiya, Israel (∼1.4 Ma; Martínez-Navarro et al., 2009, 2012), with rich core-choppers-flake assemblages and also a small but distinctive group of crude Early Acheulian bifaces, trihedrals, and quadrihedrals. 20: Thomas Quarry I-Unit L at Casablanca, Morocco (1.3–0.5 Ma; Gallotti et al., 2021), which preserves Acheulian assemblages made of quartzite and flint resulting from two production systems, one focused on the production of small to medium-sized flakes, the other devoted to the manufacture of large cutting tools. 21: Tighennif (formerly Ternifine), Argelia (∼1.0 Ma; Sahnouni et al., 2018a), with Acheulian tools associated to remains of large mammals accumulated in a primary context. 22: Aïn Boucherit (2.4–1.9; Sahnouni et al., 2018b) and Aïn Hanech, Argelia (∼1.7 Ma; Parés et al., 2014), which preserve Oldowan tools similar to those known at eastern African sites, as well as evidence of cutmarks and use-wear traces that indicate the exploitation by early Homo of animal tissues and marrow. 23: Oued Sarrat, Tunisia (∼0.7 Ma; Martínez-Navarro et al., 2014b), which has yielded the oldest known cranium of Bos primigenius, associated with other small and large vertebrates, and six Acheulian tools. (B) Chronostratigraphic chart of Early Pleistocene to early Middle Pleistocene sites with evidence of hominin presence in Europe, North Africa, the Levantine Corridor, and the Caucasus. This chart shows the geochronological units, the land mammals’ ages, the magnetostratigraphic units, and the biochronological units (French and Spanish biozones based on micromammals). Marine Isotope Stages (MIS) from Lisiecki and Raymo (2005). Biozones from Palombo (2010) and Minwer-Barakat et al. (2012).
Barranco León and FN3 lie in the NE sector of the Baza Basin (Guadix-Baza Depression, SE Spain; Figure 2A), in the vicinity of the town of Orce (Figure 2B). This sedimentary depression is an inland basin that covers an area of ∼4,000 km2 and is surrounded by mountainous reliefs of the Betic Chains, with heights of up to 3,479 m in the Mulhacén peak. The continental Plio-Pleistocene record of the basin is composed of lacustrine and fluvial deposits that show a complex sedimentary architecture (both laterally and vertically) as a result from active tectonics and orbitally induced climatic cycles (García-Aguilar and Palmqvist, 2011). The sediments include limestones, marls, shales, sands, and conglomerates, as well as dark clays and silexites associated to the archeopaleontological sites (García-Aguilar et al., 2014, 2015). The basin was in connection with the Mediterranean Sea during Late Miocene times by the ‘Almanzora Corridor’ and became continental at the end of the Tortonian (Guerra-Merchán, 1990, 1993; Soria et al., 1999; but see also Husing et al. (2010)). Since these times, it was subject to isostatic uplift with an average uplift rate of ∼200 m/Ma, as estimated from Late Neogene coastal marine conglomerates and coral reefs (Braga et al., 2003). As a result, the glacis surface (i.e., the uppermost horizontal infilling level) stands now 1,000 m on average above sea level. During the Plio-Pleistocene, the basin developed a network of endorheic drainage, being subject to: (i) intense tectonic subsidence relative to the surrounding mountains, which facilitated the accumulation of a thick (∼550 m) and relatively continuous sedimentary record; and (ii) hydrothermal activity (Figures 2A,B), which provided a mild and productive environment for the terrestrial fauna (García-Aguilar et al., 2014, 2015). The thermal springs were a major determinant in the establishment of biodiversity ‘hot spots’ for the large mammal fauna, which remains were preserved in many sites like BL and FN3 (Palmqvist et al., 1996, 2005, 2022b; Arribas and Palmqvist, 1998; Viseras et al., 2006; Arribas et al., 2009; Maldonado-Garrido et al., 2017; Ros-Montoya et al., 2017; Martínez-Navarro et al., 2018).

Figure 2. (A) Geological context of the Guadix-Baza Depression in the Betic Cordillera, SE Spain. The box encloses the sedimentary basin. Black dots indicate the thermal springs (N = 122) that are nowadays active in the Betic Cordillera, with water output temperatures between 18 and 60°C. (B) Tectono-sedimentary map of the Guadix-Baza Depression with indication of those points that preserve geochemical, mineralogical or lithological evidence of thermal activity during the Neogene-Quaternary. Geological cartography updated from García-Aguilar et al. (2014, 2015: Figure 1, respectively). 1-External Zones, 2-Internal Zones, 3-Tortonian marine deposits, 4-Turolian alluvial and lacustrine deposits, 5-Plio-Pleistocene alluvial and fluvial deposits, 6-Pliocene lacustrine deposits of the Gorafe-Huélago sector, 7-Late Turolian-Ruscinian lacustrine deposits in the Baza Basin, 8-Middle Villafranchian fluvio-lacustrine deposits, 9-Middle Villafranchian lacustrine marls and evaporites, 10-Late Villafranchian lacustrine deposits, 11-Middle Pleistocene alluvial and lacustrine deposits, 12-Late Pleistocene glacis surface, 13-Holocene fluvial terrace, 14-Modern fluvial sediments, 15-Faults (solid lines; striped lines indicate those faults covered by sediments that have been inferred from aerial photographs), 16-Thermal springs nowadays active, 17-Sulfur deposits, 18-Travertine buildings, 19-Black levels, 20- Magnesium clays, and 21-Silexites. (C) Stratigraphic series of Barranco León (BL) and Fuente Nueva 3 (FN3) sites (adapted from Espigares et al. (2019)). (D) View of the excavation quarry of FN-3. (E) View of the excavation quarry of BL.
The stratigraphy of BL (Figure 2C) spans the middle terrigenous and upper silty calcareous members of the Baza Formation (Vera et al., 1984). The middle member consists of alluvial red clays, sandstones and conglomerates while the upper one is dominated by limestones, sandstones, carbonate silts, and dark mudstones (Turq et al., 1996; Arribas and Palmqvist, 2002) deposited in a shallow lacustrine system with an alternation of oligo- to mesohaline waters (Anadón et al., 1994; Anadón and Gabàs, 2009). The excavated layers show sediments associated with a swampy environment, except level D (formerly BL5; Arribas and Palmqvist, 2002), which shows fluvial features and encases most of the archeological assemblage (Toro-Moyano et al., 2013). The sub-horizontal stratigraphy of FN3 (Figure 2C) shows two sedimentary cycles deposited in a lutitic-carbonate, lacustrine-to-swampy environment, each with limestones at the top of the sequence separated by clays, fine sands and marly lutites, which cluster in two main units, the Lower and Upper Archeological Levels (Turq et al., 1996; Martínez-Navarro et al., 1997; Espigares et al., 2013, 2019).
The age of BL-D (Figures 2C,E) and FN3 (Figures 2C,D) was estimated in 1.43 ± 0.38 and 1.19 ± 0.21 Ma, respectively, using biochronology, magnetostratigraphy and the U-series/electron spin resonance (ESR) dating method applied to optically bleached quartz grains and fossil teeth (Duval et al., 2012; Toro-Moyano et al., 2013). An age of 1.50 ± 0.31 Ma was derived for FN3 based on cosmogenic nuclides (Álvarez et al., 2015). Other age estimates were derived from a biometric approach that considered an orthogenetic, rectilinear pattern of size increase in the lower molar teeth of the arvicolid Mimomys savini (Lozano-Fernández et al., 2013, 2014), but this “vole-clock” was questioned (Martin, 2014; Palmqvist et al., 2014, 2016). The absence of suids from BL and FN3 provides also a useful biochronological inference (Martínez-Navarro et al., 2015): suids are absent from Europe between 1.8 and 1.2 Ma, until the arrival of an evolved form of Sus strozzi during the Epivillafranchian (Cherin et al., 2018, 2020). This species is first recorded in level TE9 (Atapuerca), dated at 1.22 ± 0.16 Ma by cosmogenic nuclides (Carbonell et al., 2008), and later in other sites of Jaramillo age like Untermassfeld, Vallonnet, or Vallparadís (Moullé et al., 2006; Madurell-Malapeira et al., 2010, 2014; Cherin et al., 2018, 2020).
Excavations through the last three decades in BL and FN3 have unearthed huge Oldowan assemblages composed of cores, flakes and debris that represent the whole reduction sequence (Tixier et al., 1995; Turq et al., 1996; Martínez-Navarro et al., 1997; Oms et al., 2000; Palmqvist et al., 2005; Barsky et al., 2010, 2016; Toro-Moyano et al., 2011, 2013; Espigares et al., 2013; Titton et al., 2018, 2021; Yravedra et al., 2021). The tools are associated to skeletal remains of vertebrates, mostly large mammals (>6,500 specimens in BL and >9,000 in FN3). Espigares et al. (2019) showed that of those bones with their cortical surface well preserved (4,249 in BL and 3,852 in FN3), 64 (0.8%) exhibited cut marks and 163 (2%) showed percussion marks resulting from bone fracturing by the hominins for accessing their marrow contents. Similarly, a recent study of 2,857 bone remains from FN3 unearthed during the excavation seasons of the years 2017–2020 has shown that 25 (0.9%) bear cut marks and 16 (0.6%), percussion marks (Yravedra et al., 2021). These frequencies are close to those recorded at Pirro Nord, where 1.1% of the remains are cut-marked and 0.6% show evidence of intentional bone breakage (Cheheb et al., 2019).
The anthropogenic marks of BL and FN3 provides clues on the subsistence strategies of the hominins that first dispersed in Europe (Espigares et al., 2013, 2019; Toro-Moyano et al., 2013; Yravedra et al., 2021). Cut marks are relatively short (length range: 1.8–13.0 mm) and are mostly represented by incisions, although scrapes, sawing marks, and chop marks are also documented. They mostly appear on remains of animals of medium-to-large and very large size, and evidence patterns of skinning, defleshing, disarticulation, evisceration, and periosteum removal. Percussion marks include pits, notches, impact flakes, and negative flake scars generated by hammerstone impact during the butchery of bones for marrow processing. There are also tooth-marked bones, most of them gnawed by the giant, short-faced hyena Pachycrocuta brevirostris, and some by porcupines (Espigares et al., 2019). This is particularly evident in the case of the Upper Archeological level of FN3, in which hyena coprolites are abundantly preserved (Espigares et al., 2013).
During the last years, a wealth of information on the taphonomy of the Orce sites, the technological features of their tool assemblages and the paleoecology of the faunal community has been published. This makes necessary a review of the paleoenvironments inhabited by the large mammals and the hominins in Guadix-Baza during late Early Pleistocene times, as a way of elucidating the ecological context in which the first hominin arrival in Western Europe took place. In this article, our three main goals are: (i) to review the subsistence strategies of the hominins in the archeological sites of BL and FN3; (ii) to evaluate the roles played by the hominins and carnivores in generating the fossil assemblages preserved at both sites; and (iii) to estimate the size of the hominin population that inhabited the basin.
Our review of the evidence on hominin subsistence strategies includes: (i) the inferences on the mammalian fauna (Table 1) and the paleoenvironments of the basin; (ii) the technological features of the tool assemblages and the anthropogenic marks on the bones of large mammals; (iii) the scavenging opportunities provided by sabertooths to the hominins; and (iv) the inferences on the level of competition among the members of the carnivore guild and their consequences on hominin population density.
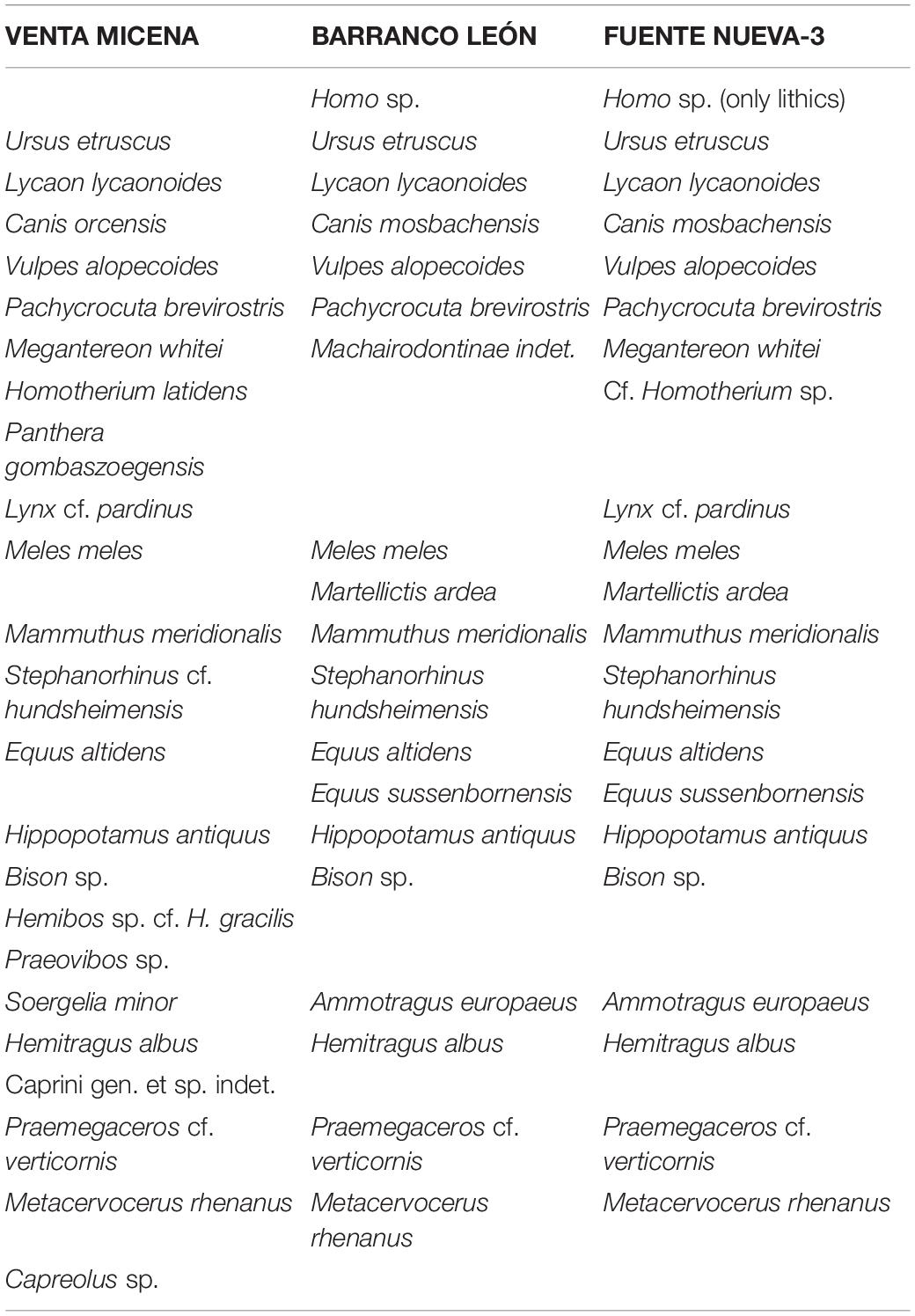
Table 1. Faunal lists (large mammals only) from Venta Micena, Barranco León, and Fuente Nueva-3 (after Alberdi and Ruiz-Bustos, 1985; Moyà-Solà, 1987; Pons-Moyà, 1987; Martínez-Navarro, 1991; Torres Pérez-Hidalgo, 1992; Martínez-Navarro and Palmqvist, 1995; Martínez-Navarro et al., 1997, 2010, 2011, 2021; Cregut-Bonnoire, 1999; Martínez-Navarro and Rook, 2003; Abbazzi, 2010; Alberdi, 2010; Lacombat, 2010; Madurell-Malapeira et al., 2010, 2011; Medin et al., 2017; Bartolini-Lucenti and Madurell-Malapeira, 2020; Ros-Montoya et al., 2021; and references therein).
We use new data for evaluating the roles of hominins and carnivores as bone collecting and modifying agents in the assemblages of BL and FN3. Specifically, we study the abundance of proximal and distal epiphyses of major limb bones of ungulates and their relationship with the mineral density and marrow yields of these bone portions. For doing so, we use modern analogs (Outram and Rowley-Conwy, 1988; Brink, 1997; Lam et al., 1999) and least-squares regression techniques using SPSS Statistics v. 25. Then, we compare the results obtained with those for VM, a site conclusively identified as generated in the surroundings of a denning site of the large bone-destroying hyena P. brevirostris (Palmqvist et al., 1996, 2011, 2022b; Arribas and Palmqvist, 1998; Palmqvist and Arribas, 2001). Thirdly, we analyze with SPSS Statistics v. 25 the relationship between prey biomass availability and rainfall in a set of African Natural Parks and Game Reserves using data from Hatton et al. (2015) and Fick and Hijmans (2017). Then, we compare these results with the estimates available for BL and FN3 (Rodríguez-Gómez et al., 2016a,2017a; Martín-González et al., 2019), to evaluate meat availability in the Orce sites. Finally, we develop a new model of the paleogeography of the Guadix-Baza Depression during the late Early Pleistocene using compiled data on the cartography on the sediments (García-Aguilar et al., 2014). This allows us to estimate with ImageJ v. 1.51 the extent of the paleoenvironments, which provides a reconstruction of the surface area habitable by the large mammals and the hominins. The cartographic surface, which encompasses the area covered by the lake, the outcrop area of alluvial and fluvial deposits as well as the glacis surface, is digitized with ImageJ for calculating the surface habitable by the terrestrial fauna. Based on this estimate, we deliver inferences on the size of the hominin population using the estimates of population density obtained for the hunter-gatherer groups of BL and FN3 (Rodríguez-Gómez et al., 2016a). This in turn leads us to discuss on the long-term viability of this hominin population and the discontinuity of the archeological record in the Guadix-Baza Depression.
The lithic assemblages from BL and FN3 are composed of abundant flakes of small size, cobbles (one third with percussion marks), cores, debris, and flake fragments. The flakes are largely made of flint and, to a lesser extent, of limestones and calcarenites from the surroundings of the sites. Flint was exploited for flake production whereas limestone, although used also for flake production, was mostly employed as percussion instruments (Barsky et al., 2010; Titton et al., 2018). Recently, Titton et al. (2021) indicated that BL is the oldest case reported of knapping and percussive activities on an ancient raw material reservoir deposit, because the analysis of the entire lithic collection of the site describes a tool kit composed of cores, flakes, debris, hammerstones, and other macro-tools like heavy-duty scrapers and sub-spheroidal morphologies. Moreover, the positioning of refitting items in the site points to in situ knapping (Toro-Moyano et al., 2013), which reaffirms the importance of BL as a raw material repository.
Many primates consume animal resources, but only humans regularly exploit animals the same size or larger than themselves (Butynski, 1982; Pobiner, 2020). Large ungulates represent concentrated packages of fat and protein, which are easily digestible and calorically rich macronutrients, and contribute micronutrients that are scarce in plants, such as heme-iron, folic acid and vitamin B12 (Thompson et al., 2019). Internal bone nutrients (e.g., brains and marrow) provide to humans the precursors to docosahexaenoic fatty acids and oleic acids, key for brain development and female reproductive success in hunter-gatherer societies (Plummer, 2004; Pobiner, 2020). This explains why the consumption of large ungulates was a fundamental component of the high-quality diet that allowed the evolutionary transformations that led to the genus Homo (e.g., reduction of postcanine teeth, brain expansion, enhanced cooperation with non-kin, and shorter interbirth intervals). For this reason, the tool-use/meat-eating package has been considered as inherently linked to the definition of Homo (Jiménez-Arenas et al., 2014; Thompson et al., 2019; Pobiner, 2020; however, for a recent criticism of the narrative that links the anatomical and behavioral traits of H. erectus to an increase in meat eating, see Barr et al., 2022).
The technological features of the lithic tools from BL and FN3, including the small dimensions of the flakes, allow discussing on carcass acquisition and processing by these hominins. This relates to the classic debate on Homo as a hunter or as a scavenger. During the eighties and nineties, most researchers interpreted the cut marks found in the Early Pleistocene sites of East Africa as evidence of defleshing activities by the Oldowan hominins of ungulate carcasses obtained through passive scavenging, which implied a secondary access to these resources (Binford, 1981, 1985; Blumenschine, 1986, 1987, 1991, 1995; Blumenschine and Selvaggio, 1988; Blumenschine et al., 1994; Capaldo, 1997; Selvaggio, 1998; Arribas and Palmqvist, 1999). In contrast, from the nineties onward a new scenario was considered, which envisaged the Oldowan hominins as having primary access to fully fleshed carcasses obtained from hunting or through active, confrontational scavenging (Bunn and Ezzo, 1993; Domínguez-Rodrigo, 1999; Bunn, 2001; Domínguez-Rodrigo and Piqueras, 2003; Domínguez-Rodrigo and Barba, 2006; Domínguez-Rodrigo et al., 2007, 2014; Bunn and Pickering, 2010). The reasoning, based on actualistic studies of predation, was as follows: although the large felids do not consume bone marrow contents, which opened to early Homo the opportunity to scavenge these resources, they exploit intensively the prey carcass, which results in a low availability of flesh. The lions of Tarangire National Park (Tanzania) are a good example: they efficiently deflesh small and medium-sized ungulate carcasses and in alluvial environments near water, they even thoroughly deflesh carcasses of prey heavier than 500 kg such as buffalo (Gidna et al., 2014). However, a study on the potential scavenging opportunities in Sweetwaters Game Reserve (Kenya), a conservancy area where lions face a low level of inter-specific competition from hyenas, has shown that lions abandoned 95% of bones of large prey with at least some scavengeable flesh, and over 50% were left with large muscle masses. Moreover, while the scavengeable resources from fresh kills made by lions vary among prey carcasses, a single carcass of a large prey abandoned with only flesh scraps remaining (and all bone marrow contents intact) was usually substantial enough to satisfy the total daily caloric requirements of at least one adult male of Homo erectus, as exprapolated from an estimate for anatomically modern humans (Pobiner, 2015, 2020). This provides a solid argument for interpreting the Oldowan hominins as confrontational kleptoparasites or even as marginal, passive scavengers. However, human impact is high in Sweetwaters, and this could modify the behavior of lions, which deflesh their prey more thoroughly in those ecosystems where no modern humans threaten them (Gidna et al., 2014). In any case, Blumenschine and Pobiner (2007) estimated that the marrow that a hominin could extract from twelve long bones of an adult wildebeest carcass would represent 3,000 kcal, a value close to the daily energetic requirements of an adult early Homo (Aiello and Wells, 2002). In addition, Bickerton and Szathmáry (2011) suggested that the populations of H. erectus would have access to at least one elephant carcass resulting from natural death every month per 1000 km2 foraged.
The hunting vs. scavenging debate was largely focused on the sequence of carcass access initially proposed for the bone assemblage from the FLK Zinjanthropus site at Olduvai, based on the frequency of tooth-marked and percussion-marked long bones of large mammals (Blumenschine, 1995; Capaldo, 1997; Selvaggio, 1998). The model considered a three-stage sequence of site formation: (i) in stage one, flesh-eating felids partially defleshed long bones, as deduced from the high frequency of tooth-marks on midshaft fragments; (ii) in stage two, hominins processed intact long bones for their marrow contents and left percussion marks, with the presence of cut marks indicating that the bones retained at this stage variable amounts of flesh; finally, (iii) in stage three, bone-cracking hyenas consumed long bone epiphyses for grease, as evidenced by the underrepresentation of these elements in the assemblage and the high percentage of tooth marks on the surviving epiphyses and the distal portions of the diaphyses (see discussion in Arribas and Palmqvist, 1999).
Domínguez-Rodrigo and Barba (2006) suggested that Blumenschine (1995) overestimated the number of long-bone midshafts with tooth marks in the FLK 22 assemblage due to the confusion of these marks with those resulting from microbial bioerosion, which would indicate primary access by the hominins to fully fleshed carcasses (but see criticism of their experimental procedure by Blumenschine et al. (2007)). The most recent analysis of the FLK 22 assemblage, using GIS techniques (Parkinson, 2018), has suggested that hominins had early access to largely fleshed carcasses, particularly those of smaller prey —which they may have primarily acquired through hunting— although patterns of bone damage on larger carcasses are consistent with secondary access through aggressive scavenging (Pobiner, 2020). In any case, we must bear in mind that the interpretive context that envisions the Oldowan hominins as primary hunters is based on experimental studies performed on modern felids, which thoroughly exploit the carcasses of their prey. This context does not apply to the Early Pleistocene hominin populations with Oldowan tools of Africa and Europe, where the predator guild was dominated by two sabertooths, Megantereon whitei and Homotherium latidens (Martínez-Navarro and Palmqvist, 1995, 1996; Arribas and Palmqvist, 1999; Palmqvist et al., 2022a).
Sabertooths have no living analogs and dominated the carnivore guild during most of the Cenozoic, filling the niche now occupied by the pantherine felids (Van Valkenburgh, 2001, 2007). Their long and laterally flattened upper canines were an adaptation for killing quickly megafaunal prey with deep wounds onto the prey throat rather than using the prolonged suffocating throat bite typical of the extant felines (Gonyea, 1976; Akersten, 1985; Anyonge, 1996a; Antón et al., 2004; McHenry et al., 2007; Christiansen, 2008; Meachen-Samuels and Van Valkenburgh, 2010; Salesa et al., 2010; Andersson et al., 2011; Meachen-Samuels, 2012). However, canine hypertrophy posed a biomechanical constraint on mandibular gape for delivering the killing bite, which involved a major reorganization of the skull to avoid over-stretching of the temporalis muscle during wide gaping (Figure 3) and led to a reduction of the premolar teeth. This is particularly evident in the African sabertooth M. whitei, which dispersed out of Africa by ∼1.8 Ma (Martínez-Navarro and Palmqvist, 1995, 1996; Palmqvist, 2002): compared to the less specialized M. cultridens, the species replaced in Europe by M. whitei during the late Early Pleistocene, the third lower premolar is reduced in M. whitei to a vestigial peg or even lost, while the fourth premolar and the paraconid of the carnassial are shortened but to a lesser degree, which reflects the greater enlargement of the sabers in the African species (Palmqvist et al., 2007). This resulted in lesser abilities to process the prey carcass and made it available more scavengeable resources for the hyenas and the hominins, which provides the ecological connection between the dispersal of M. whitei out of Africa and the first arrival of Homo in Europe, a continent where the survival of hominins during the cold season —with lowered plant resources compared to East Africa— depended on the regular scavenging of ungulate carcasses (Turner, 1992; Martínez-Navarro and Palmqvist, 1995, 1996; Arribas and Palmqvist, 1999; Martínez-Navarro, 2004, 2010; Palmqvist et al., 2007, 2022a).
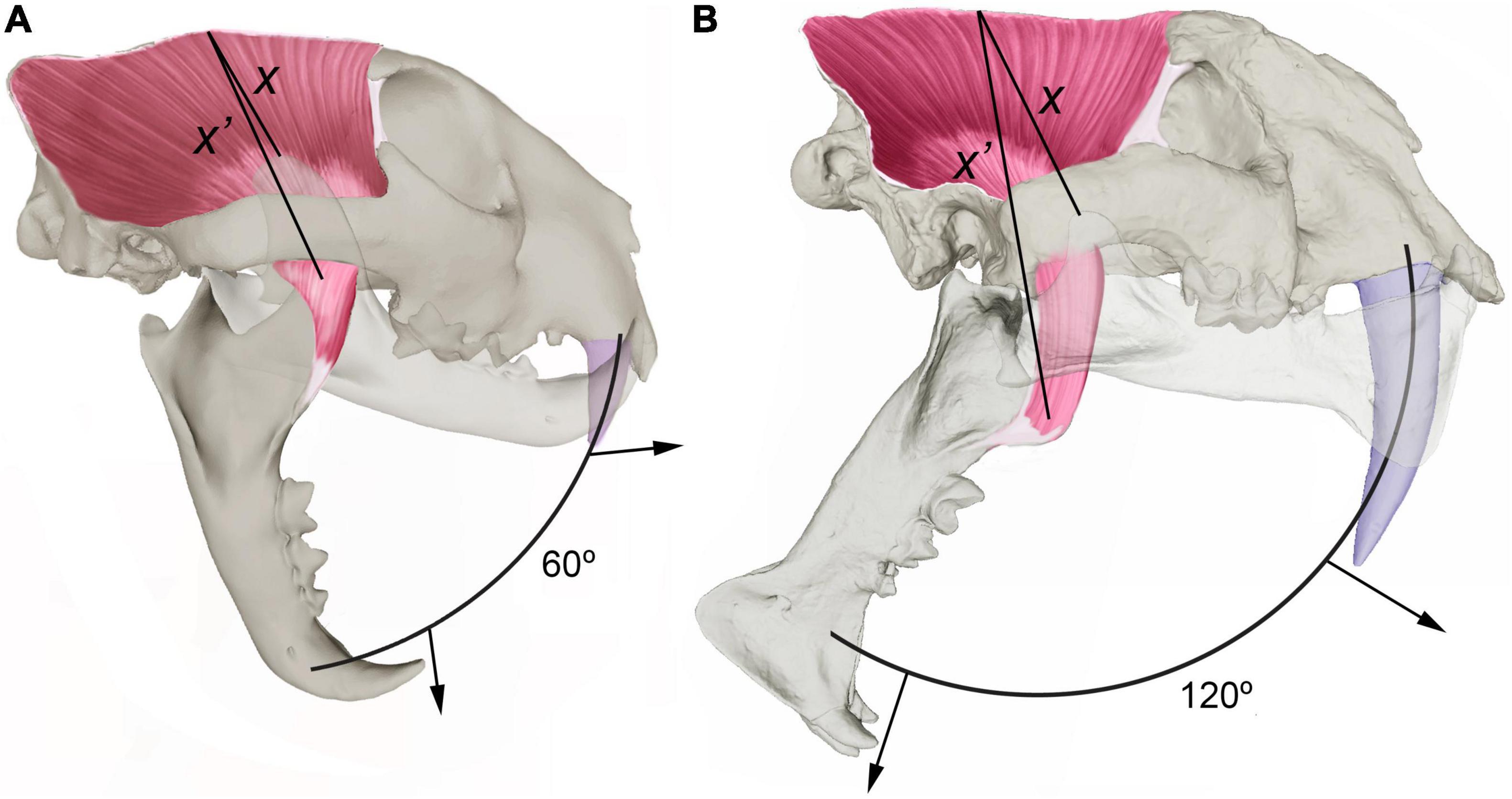
Figure 3. Comparison of the craniodental anatomy of a leopard, Panthera pardus (A), with a sabertooth, Megantereon nihowanensis (B). The lines X and X′ measure the stretching of temporalis muscle from the tip of the coronoid process to the sagittal crest with the jaw closed and open, respectively. The elongation of the upper canines of sabertooths posed a severe biomechanical constraint on mandibular gape for delivering the killing bite, which involved a major reorganization of the temporalis to avoid over-stretching of the muscle fibers during wide gaping. This was achieved by a number of changes in their craniodental anatomy compared to the pantherine felids (Emerson and Radinsky, 1980; Akersten, 1985; Palmqvist et al., 2007; Slater and Van Valkenburgh, 2008; Figueirido et al., 2011; DeSantis et al., 2021): (i) a lowered glenoid fossa; (ii) a shortened coronoid process and a laterally shifted angular process; (iii) a less laterally projected postglenoid process; (iv) an upwardly rotated palate; (v) a shorter and narrower temporal fossa; (vi) a more vertical occiput; and (vi) a protruding incisor arcade, which independized the hypertrophied upper canines from the incisors. Such skull reorganization resulted in a narrowing of the temporalis fibers and their more perpendicular orientation to the tooth row, which allowed to increase jaw gape up to 180° in sabertooths while retaining a degree of muscle stretch like that of pantherine felids. Moreover, the masseter muscle, which exerts its maximum force at smaller gapes, was also reduced. Panel (A) shows that with a jaw gape of 60°, the degree of muscle stretching of the temporalis (measured by the ratio between X′ and X) in the leopard (∼85%) is like in Megantereon (B) with a gape of 120° (∼80%). A result of this major skull reorganization in sabertooths was that the point of maximum bite force exerted at the carnassial was positioned more backwardly, which led to a reduction of the post-canine dentition not related to the slicing function of the carnassial. The protruding incisor arcade of sabertooths (B) helped these predators to avoid canine breakage when feeding on the prey carcass because it independized the function of the incisors, used to tear chunks of flesh from the prey carcass (a task performed in modern felids by their stout, conically shaped canines), from the function of the canines, employed to deliver deep wounds during prey dispatch (Biknevicius et al., 1996). Thus, prevention of canine breakage during prey killing and feeding encounters was a strong selective agent in sabertooths and suggests that a non-scavenging behavior was a clear ecological limitation posed by their hypertrophied upper canines.
Microwear analyses of canines and carnassials provides additional evidence on the lower ability of sabertooths to process the prey carcass. A comparison of the frequency of pits and scratches in the canines of the North American Smilodon fatalis and other living carnivores with disparate feeding and hunting repertoires (e.g., spotted hyenas, lions, cheetahs, and wild dogs) showed that these teeth were used by S. fatalis for prey killing and avoided contact with bone during prey consumption (Anyonge, 1996b). In the case of the wear facet of the carnassials, which are employed exclusively in food processing, the bone-cracking hyenas exhibit few long scratches and a high proportion of pits to scratches, while the cheetah (which feeds exclusively on soft organs and only consumes bones of small prey; Schaller, 1968; Brain, 1981; Skinner and Smithers, 1990) shows a predominance of narrow scratches with very few pits. Strikingly, the microwear pattern of S. fatalis showed relatively narrow and long scratches combined with an extremely low frequency of pits, even lower than in the cheetah (Van Valkenburgh et al., 1990). This suggests that sabertooths probably consumed even less bone than the cheetah to prevent canine breakage, which means that their prey carcasses would retain a significant fraction of scavengeable resources (Van Valkenburgh et al., 1990).
The elongated, laterally flattened canines of sabertooths were optimal for killing large prey, but also were more vulnerable to fracture than the shorter and conical canines of the living felids due to the unpredictable loads generated in prey stabilization during the killing bite (Van Valkenburgh and Hertel, 1993; Van Valkenburgh, 2009). This made the heavily muscled forelimbs of sabertooths imperative for pulling down and immobilizing prey before positioning the killing bite (Gonyea, 1976; Akersten, 1985; Anyonge, 1996a; Antón et al., 2004; Christiansen, 2008; Salesa et al., 2010; Andersson et al., 2011; Meachen-Samuels, 2012). The latter is reflected in the short and robust forelimb bones of S. fatalis, reinforced by cortical thickening, which allowed it to minimize prey struggling, helping to position the killing bite carefully to avoid contact with bone (Meachen-Samuels and Van Valkenburgh, 2010; Martín-Serra et al., 2017). A comparative study of the skulls of S. fatalis and the lion based on finite element analysis showed that the sabertooth skull was less equipped to resist the forces generated by a struggling prey, which pointed to rapid slashing bites during prey killing (McHenry et al., 2007). In contrast, the lion uses its stouter canines to hold a suffocating bite in the snout of large prey such as buffalo, which explains that the forelimbs are less important for subduing prey (Salesa et al., 2010; Meachen-Samuels, 2012; Martín-Serra et al., 2017). Moreover, Figueirido et al. (2018) showed that the rostrum of S. fatalis was almost entirely composed of cortical bone (which supports better directed loads) while the lion skull had a substatinal amount of trabecular bone (which can support unpredicted and multidirectional forces). This indicated that the skull of S. fatalis was well-equipped to deliver a quick killing-shear bite, but it could not withstand the unpredictable forces generated when feeding on bones (Figueirido et al., 2018).
The highly derived craniodental and postcranial anatomy of sabertooths suggests that the pantherine felids cannot be considered as their modern functional analogs, because sabertooths: (i) were able to hunt larger ungulate prey relative to their body size, exerting a higher predation pressure on the juveniles of megafauna; and (ii) exploited their prey to a lesser extent, which would have resulted in greater amounts of flesh abandoned in the prey carcass (Binford, 1980, 1981; Marean and Ehrhardt, 1995; Arribas and Palmqvist, 1999; Palmqvist et al., 2003, 2007, 2011, 2022a,b; Ripple and Van Valkenburgh, 2010; Martínez-Navarro et al., 2014a; Van Valkenburgh et al., 2016; Martín-Serra et al., 2017; Martínez-Navarro, 2018). These resources would in turn be available for the scavengers, including the hominins and hyenas, as documented at FN3 (Espigares et al., 2013, 2019; Yravedra et al., 2021). This interpretation makes sense if we consider the lack of an effective weaponry in the Oldowan hominins for subduing large prey with their small flint flakes and cores, or for driving predators from their kills acting as kleptoparasites (Potts, 1991; Blumenschine and Pobiner, 2007; Treves and Palmqvist, 2007). However, throwing stones for driving away carnivores and stealing their prey would be always a possibility, as suggested by Lordkipanidze (2015) for explaining the abundant allochtonous cobbles found in Dmanisi (Coil et al., 2020). Interestingly, manuports of dolomitic limestone are abundantly represented in FN3 (Espigares et al., 2013).
While it has been argued that expanding group sizes prompted Acheulean hominins to become big game hunters (Martínez-Navarro, 2018), perhaps including elephants as their prey (Agam and Barkai, 2018), it is difficult to conceive that the limited technological skills of the Oldowan hominins allowed them to prey on megafauna. Therefore, it is thus more reasonable to consider that Early Homo initially expanded its diet from the major reliance on plant foods of australopithecines to scavenging bone marrow, brains and meat (Ben-Dor et al., 2021). Therefore, the trophic level of Homo probably evolved from a low base to a high carnivorous position during the Pleistocene, beginning with H. habilis as a scavenger and peaking in H. erectus as a hunter (Ben-Dor et al., 2021; but see Barr et al. (2022)).
Megaherbivores like adult elephants, hippos and rhinos are often regarded as invulnerable to predation, but it has been suggested that lions regularly hunted such large prey during the Pleistocene (Guthrie, 1990). Lion prides have been documented today hunting elephants, mostly juveniles, in high frequencies in those environments where other ungulates are scarce, like the Savuti area of Chobe National Park, Botswana (Joubert, 2006; Power and Shem-Compion, 2009). However, lions preferentially prey on medium-to-large ungulates when they are abundant (Schaller, 1972), within a weight range of 190–550 kg (Hayward and Kerley, 2005), and only occasionally hunt megafauna (Palmqvist et al., 1996). In fact, elephants are rarely preyed upon by lions (Skinner and Smithers, 1990) and when this does occur it involves yearling calves weakened by drought (Loveridge et al., 2006) or older animals wounded by poaching (Ruggiero, 1991).
Passive scavenging of ungulate carcasses partially defleshed by sabertooths would have enhanced the survival of the hominins of Guadix-Baza during Early Pleistocene times (Martínez-Navarro and Palmqvist, 1996; Arribas and Palmqvist, 1999; Martínez-Navarro, 2004, 2010; Palmqvist et al., 2005, 2007; Espigares et al., 2013, 2019; Martínez-Navarro et al., 2014a; Rodríguez-Gómez et al., 2016a, 2017a). We propose here that they probably exploited a wide repertoire of subsistence strategies, including: (i) the opportunistic hunting of small-sized mammals and other vertebrates (e.g., amphibians and reptiles); (ii) the kleptoparasitism of the prey of primary predators like sabertooths and wild dogs; and (iii) the opportunistic scavenging of carcasses of very large animals not exposed in life to predation that died from other causes, although this is a rather speculative issue. The latter possibility is supported by evidence of competition between Homo sp. and P. brevirostris for the exploitation of a carcass of an old individual of elephant Mammuthus meridionalis in the upper archeological level of FN3: the skeleton of this elephant is dismembered and surrounded in part by flint flakes and coprolites, which suggests a sequential pattern of consumption by the hominins and hyenas (Espigares et al., 2013).
A mathematical model that evaluates the sustainability of the community of secondary consumers, based on the biomass of primary consumers potentially available (Rodríguez-Gómez et al., 2013, 2014a,b,2016b,2020), has provided relevant data on the hominin population that inhabited Guadix-Baza (Rodríguez-Gómez et al., 2016a). The model estimates: (i) the age structure and mortality rate that make the population of each primary consumer stable; (ii) the distribution of individuals among size categories; (iii) the biomass that the secondary consumers can extract from these populations in the long term; and (iv) resource distribution among the members of the carnivore guild. In the case of hominins, the model considered a dietary contribution of 30% of animal resources, in agreement with the diet of modern hunter-gatherer populations at temperate latitudes (Rodríguez-Gómez et al., 2016a). The hunting and scavenging scenarios were both modeled for estimating the population density of Homo sp. in Guadix-Baza. This suggested a scavenging behavior as optimal for the population, which would hold 12 individuals per 100 km2 during a year, a value close to the mean density of modern hunter-gatherers (Binford, 2001; Marlowe, 2005). The density estimated for a strict hunting behavior was slightly lower, 9.5 individuals/100 km2 (Rodríguez-Gómez et al., 2016a). Given that both estimates are similar, to consider a scavenging or a hunting behavior has little effect on the size of the hominin population of the basin, as noted above. The densities estimated for the hominins and carnivores in the Orce sites (Figure 4) agree with the prey/predator biomass ratios derived with the equation of Hatton et al. (2015).
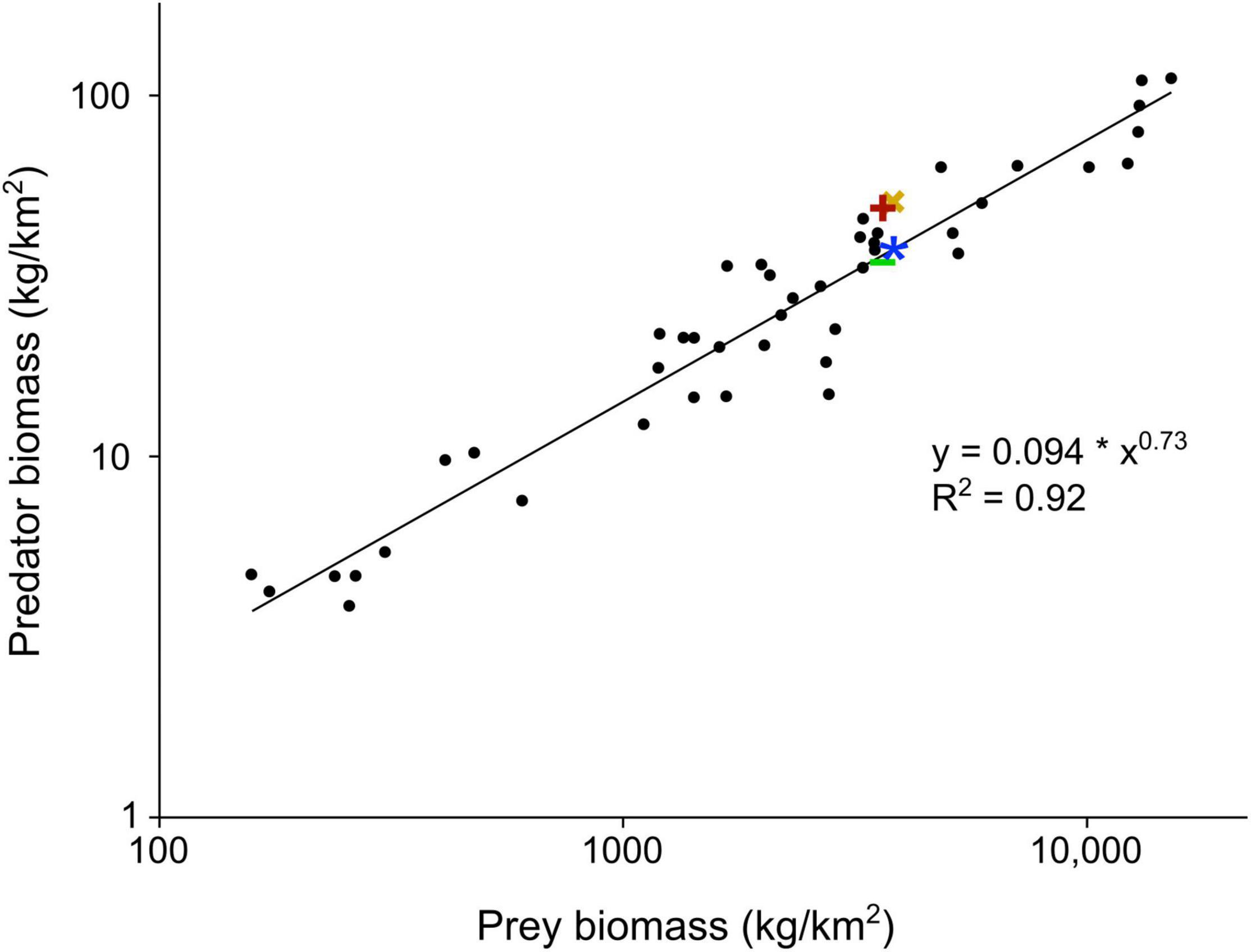
Figure 4. Relationship between the biomass of predators and prey in modern ecosystems (data from Hatton et al. (2015)) and ranges of estimates of predator and prey biomass values obtained for the Orce sites [data from Rodríguez-Gómez et al. (2016a, 2017a)]. Fossil assemblages: Barranco León/Fuente Nueva 3 (maximum: red cross, minimum: green dash) and Venta Micena (maximum: yellow ex, minimum: blue asterisk). Least-squares regression equation derived with software SPSS v. 25.
“Meat made us humans” is a recurrent topic in any debate on the subsistence strategies of hominins (Bunn, 1981, 2007; Stanford, 1999; Bunn et al., 2017). This relates to the evolutionary trend toward increasing encephalization in the genus Homo, because the high maintenance cost of the nervous system involved a reduction of the digestive tract and a shift toward a more carnivore diet compared to the australopithecines (Leonard and Robertson, 1994, 1996; Aiello and Wheeler, 1995; Jiménez-Arenas et al., 2014). However, the subsistence strategies of early Homo probably included a broader spectrum of resources (e.g., small mammals, birds, herpetofauna, invertebrates, eggs, honey, and edible vegetation) that do not leave archeological evidence (Blasco et al., 2011; Hardy et al., 2017; Prado-Nóvoa et al., 2017; Espigares et al., 2019), as happens in the case of modern hunter-gatherers like the !Kung of the Kalahari Desert and the Hadza of northern Tanzania (Woodburn, 1968; Ho et al., 1972; Lee, 1979; Silberbauer, 1981; O’Connell et al., 1988; Hawkes et al., 1991; Cordain et al., 2000; Binford, 2001; Bunn, 2001; Marlowe, 2005). Lee (1968), Cordain et al. (2000), and Marlowe (2005) have shown a negative correlation between gathering and latitude due to the decrease with latitude in the availability of edible plants. In contrast, gathering is the dominant mode of subsistence in latitudes like those of the Orce sites. In the Baza Basin, freshwater fish could have also been a regular source of long-chain polyunsaturated fatty acids (e.g., omega n-3, n-6 and docosahexaenoic acids) that are essential for the early development of the brain, retina and other neural tissues (Uauy et al., 2001; Kuipers et al., 2010).
There is a lack of knowledge on the vegetal resources available at the Orce sites, as all attempts to extract fossil palynomorphs from the sediment were unsuccessful and even the coprolites of hyena analyzed were palynologically sterile (Carrión, 2002; Carrión et al., 2009). Despite this, inferences on the past vegetation of the basin have been derived from δ13C and δ15N isotopes in the bone collagen of the herbivores of VM (Palmqvist et al., 2003, 2008a,b), which allowed to infer their feeding habits. Specifically, the species (Table 1) were classified among: (i) browsers (rhino Stephanorhinus hundsheimensis and deer Praemegaceros verticornis); (ii) mix-feeders (ovibovine Soergelia minor and deer Metacervocerus rhenanus); and (iii) grazers (horse Equus altidens, Bison sp., muskox Praeovibos sp., goat Hemitragus albus, and M. meridionalis). The predominance of taxa adapted to grazing in open habitat agrees with: (i) the synecological reconstruction of this paleocommunity as a plain with bush/forest patches (Mendoza et al., 2005; Saarinen et al., 2021); and (ii) the estimate of ∼780 mm of annual rainfall derived from the range of δ15N values in the ungulates that fed on terrestrial vegetation (García-Aguilar et al., 2014), which is close to the estimate of ∼750 mm obtained from the herpetofauna of BL and FN3 (Blain et al., 2016). According to Blain et al. (2021), the first hominin occupations in Western Europe (e.g., BL/FN3 and TE9) would correspond to warm and humid conditions in an open ‘savanna’ environment with 20–25% wood. A study of the herpetofaunal assemblages of BL and FN3 has indicated different conditions for the levels with the highest density of anthropic evidence at these sites: a humid, wooded biotope for BL and a more open and drier biotope for FN3 (Sánchez-Bandera et al., 2020).
Most ungulate species present at VM are also represented in the faunal assemblages of BL and FN3 (Table 1). The exceptions are the mesodont S. minor, which is replaced by the hypsodont caprine Ammotragus europaeus, and the presence of a second equid species, the large-sized horse Equus sussenbornensis (Moullé et al., 2004; Alberdi, 2010; Martínez-Navarro et al., 2010). Hindgut fermenters process a large volume of food in a short time and can feed on low quality grasses too fibrous for a ruminant to subsist on (Janis, 1976; Janis et al., 1984; Duncan et al., 1990). For this reason, the presence of two hypergrazing equids in BL and FN3 suggests more arid conditions and an herbage of lower quality than in VM (Palmqvist et al., 2008a). This would result in a decrease in the abundance of fruits and other succulent plant stuffs for the hominins, which would make them more dependent on animal resources.
Espigares et al. (2019) reported on the cut marks and percussion marks identified in BL and FN3 (see also Yravedra et al. (2021) for FN3). Tooth marks produced by carnivores, particularly the hyena P. brevirostris, were also found in the bone assemblages of both sites, but at lower frequencies than in the hyena den of VM (Palmqvist et al., 1996, 2005, 2011, 2022b; Luzón et al., 2021). To evaluate the contribution of hominins and carnivores to the site formation process in BL and FN3, we have performed here a study on the abundance of limb bone epiphyses and compared the results with those for VM.
Figure 5 shows the relationship between the abundance of epiphyses of major limb bones and their estimated marrow yields in the three Orce sites. The regressions for BL and FN3 show an inverse, statistically significant relationship between bone survival and marrow contents (r = 0.624, p = 0.05 for BL; r = 0.682, p = 0.03 for FN3), which indicates that the hominins preferentially fractured the anatomical portions with greater nutritional value. However, these regression lines show considerable scatter, as reflected in their wide confidence 95% intervals. There is also a negative relationship in VM, but much closer in statistical terms (r = 0.885, p = 0.001), which shows a greater selectivity in bone fracturing by the hyenas. Figure 6 shows the positive relationship between bone survival and mineral density in the three sites. Again, BL and FN3 show lower levels of statistical significance (r = 0.636, p = 0.003 for BL; r = 0.548, p = 0.015 for FN3) and more scatter around the regression lines than VM (r = 0.745, p < 0.0001 for VM).
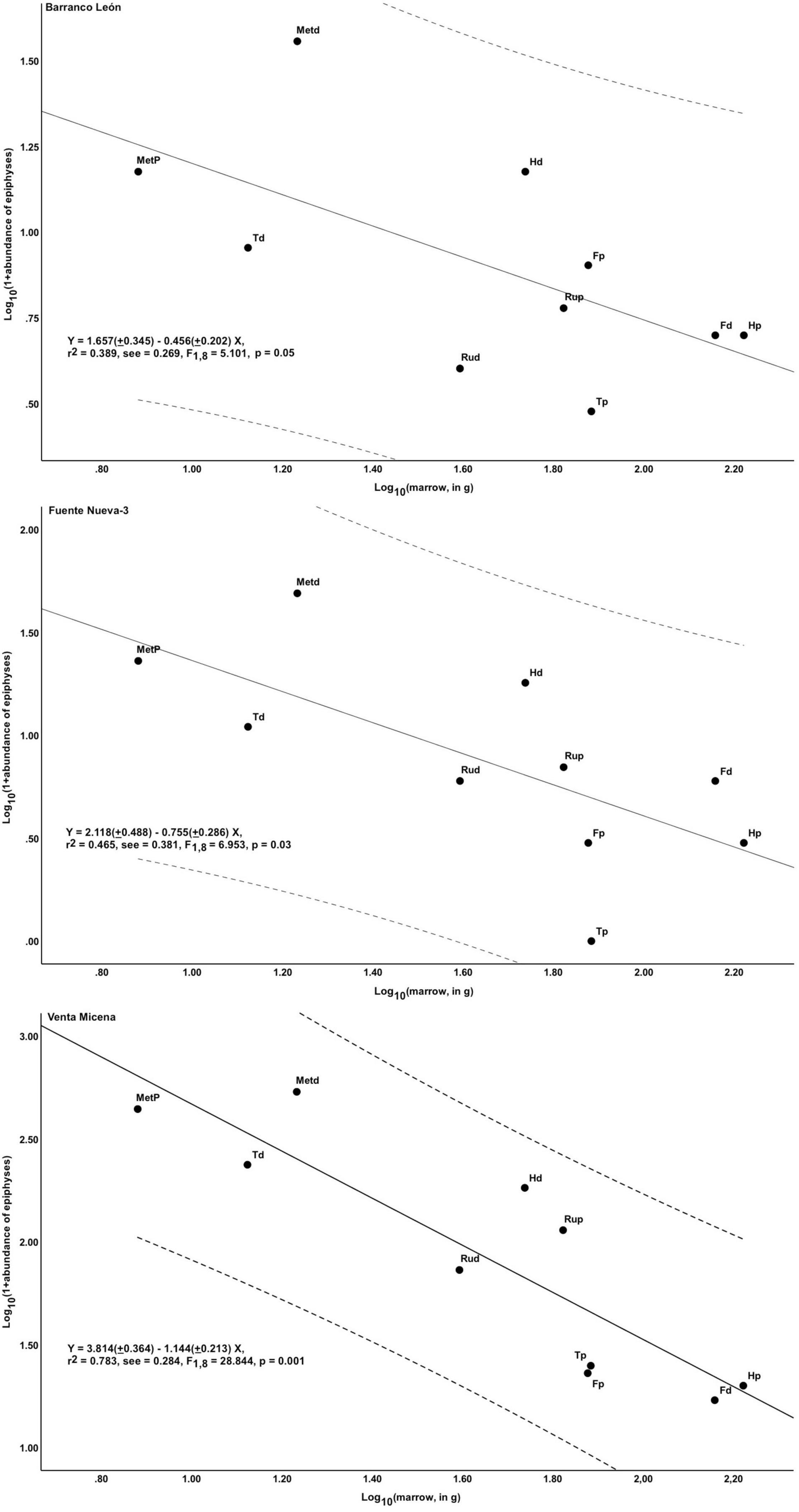
Figure 5. Relationship between the survival of major limb bone epiphyses and their estimated marrow contents (mean for modern horse and bison; Outram and Rowley-Conwy, 1988; Brink, 1997) in BL, FN3, and VM. H, Humerus; Ru, Radius/ulnae; F, Femur; T, Tibia; Met, Metapodials; p, Proximal; d, Distal. Striped lines represent the 95% confidence intervals below and above the regression lines. Least-squares regression equations derived with software SPSS v. 25.
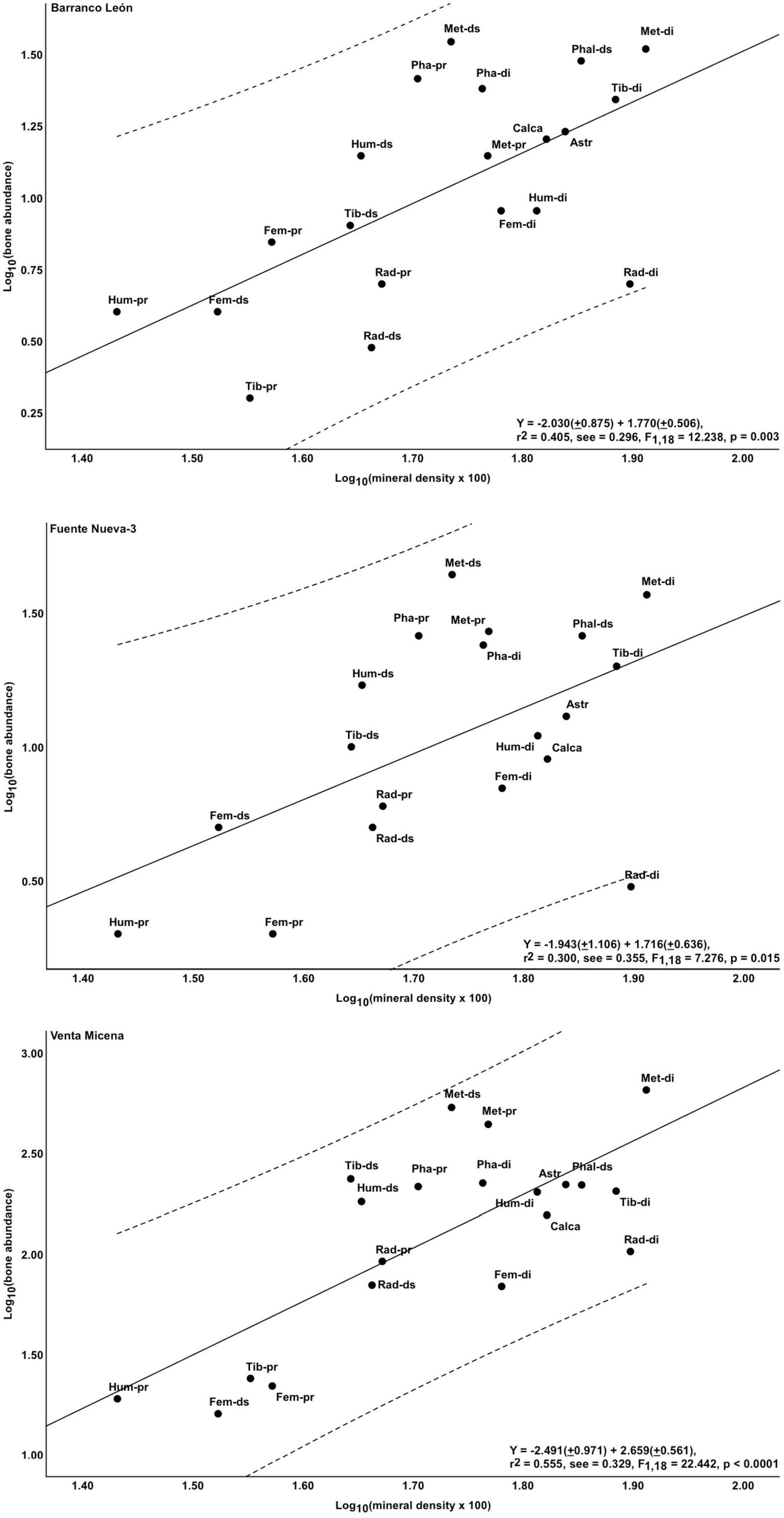
Figure 6. Relationship between the survival of skeletal remains and their estimated mineral density (mean for modern horse, gnu and raindeer; Lam et al., 1999) in BL, FN3, and VM. Hum, Humerus; Rad, Radius; Fem, Femur; Tib, Tibia; Met, Metapodials; Astr, Astragalus; Calca, Calcaneum; Pha, Phalanges; pr, Proximal; di, Diaphysis; ds, Distal. Striped lines represent the 95% confidence intervals below and above the regression lines. Least-squares regression equations derived with software SPSS v. 25.
Marrow determines the interest of the bone collecting and modifying agent on the nutritional contents of the skeletal remains, while mineral density is behind the effort invested in accessing these resources. The hominins fractured the bones in BL and FN3 using stones, a task relatively straightforward that did not represent a major effort, which originated percussion marks (Espigares et al., 2019; Yravedra et al., 2021). Therefore, it is not expected that they were highly selective in their choice of the bones to be fractured, apart from focusing more on the remains that provided more marrow. However, hyenas fractured the bones in VM using their robust premolar teeth assisted by their massive jaws (Palmqvist et al., 2011), which resulted in abundant spiral and longitudinal fractures (Arribas and Palmqvist, 1998). Hyenas were at risk of breaking their teeth or dislocating their jaws while fracturing the densest bones, which forced them to be more selective than the hominins (who risked nothing when fracturing bones with stones). Thus, our results suggest that although BL and FN3 record evidence of hominin and carnivore activity, the main agent involved in the accumulation and modification of the remains preserved at both sites were the hominins.
The hyena P. brevirostris, the jackal-sized Canis mosbachensis and a large-sized crow (unpublished specimen VM-3121, preliminarily classified as Corvus sp.; by the moment, this species has not been identified in BL and FN3) were indeed worthy competitors of the hominins for carrion in BL and FN3. Soaring at high altitude is an energetically efficient mode of locomotion, which allows vultures to cover huge distances and discover carcasses by sight or watching the movements of other neighboring birds such as crows (Palmqvist and Vizcaíno, 2003). Hyenas and jackals often rely upon visual clues such as circling vultures to identify scavengeable resources, and then run long distances to secure the carcass (Bramble and Lieberman, 2004). However, they do not perform well running at long distances, as they need to rest after relatively short distances to breathe and cool down. In contrast, humans are comparatively poor sprinters but are well adapted for endurance running (i.e., running many kilometers at relatively low speed over extended time periods using aerobic metabolism) (Bramble and Lieberman, 2004; Lieberman et al., 2007; Pickering and Bunn, 2007; Liebenberg, 2008; Ruxton and Wilkinson, 2012). This results in an advantage for scavenging in open habitats during the day, particularly during the dry season, when other terrestrial scavengers are prevented from running long distances due to thermoregulatory constraints (Lieberman et al., 2007). Endurance running may have allowed early Homo to reach carcasses before other terrestrial scavengers and to exploit them until surrendering them on the arrival of potentially dangerous hyenas. However, the advantages of endurance running are not unanimously accepted. Pickering and Bunn (2007) argued that endurance running: (i) would have required for hunting that in the absence of visual contact, hominins had the ability to track prey; (ii) is not common among modern foragers except for very open and hot habitats, because it is a physically demanding activity; and (iii) scavenging in modern riparian woodland habitats would result in a low competition intensity for carcasses. These arguments are based on two flawed assumptions, the presumptive link between modern human-like cognition and tracking abilities, as well as the notion that the limited (and biased) modern ethnographic record provides an adequate reflection of past behaviors (Lieberman et al., 2007).
Hominins entered the carnivore guild when they became scavengers (and later hunters), which forced them to compete with other carnivores using a combination of strength, speed, stealth, and cooperation. Modern foragers are no exception in this, as a high percentage of scavenging opportunities observed among Hadza and other hunter-gatherers involve power scavenging for driving off lions or hyenas from their kills using simple weapons like sticks and stones (Potts, 1991; Blumenschine and Pobiner, 2007; Lieberman et al., 2007). Given that early Homo was neither strong nor powerful, and apparently lacked projectile weapons, it is debatable whether they could engage in competition scavenging with dangerous carnivores (Potts, 1991; Lieberman et al., 2007). Therefore, persistence hunting and scavenging may have been more common before the invention of the bow-and-arrow or the domestication of dogs and horses (Liebenberg, 2008).
Domínguez-Rodrigo and Organista (2007) proposed the use of the following ratios for evaluating the degree of ravaging intensity in an assemblage: (i) axial bones to appendicular ones, which would range from 4.25 for a carcass transported complete or died in a setting devoid of competition among carnivores, to 0 for a completely ravaged skeleton; (ii) femur to tibia, which would range from 1 in a ravaging-free assemblage to 0 in one with maximum ravaging intensity; and (iii) proximal humerus plus distal radius to distal humerus plus proximal radius, which relates to the relative abundance of the least dense bone portions (preferentially consumed by the carnivores) to the densest ones, and takes a value between 1 in an undisturbed carcass and 0 in the situation of highest ravaging. The second ratio is only relevant if carnivores had primary access to complete bones and not to bones already broken by hominins (Domínguez-Rodrigo and Organista, 2007). In the latter case, when taphonomic evidence indicates that the hominins broke long limb bones (as in BL and FN3) and only bone portions instead of whole bones were available for ravaging, they recommend the third ratio as the most informative.
Girdle and limb bones are between three and five times more abundant at the hyena den of VM than vertebrae and ribs, depending on the excavation quarry analyzed (García-Aguilar et al., 2015; Luzón et al., 2021; Palmqvist et al., 2022b). Ribs are scarcely represented at VM by small fragments and are even less abundant in BL and FN3 (Espigares, 2010). The ratio of ribs, vertebrae and girdle bones to limb bones is 17.9% (52/290) in BL and 11.8% (35/296) in FN3, figures that compare well with VM, 13.5% (532/3,942). In VM, the overrepresentation of the elements of the appendicular skeleton over those of the axial skeleton indicates the dismemberment by hyenas of the ungulate carcasses scavenged and the preferential transport of the limbs to their denning sites (Palmqvist and Arribas, 2001). Our data suggest that the assemblages of BL and FN3 were also biased by the selective transport of remains by the hominins, who were focused to marrow extraction of the remains in a safe place (Espigares et al., 2013). The third ratio shows similar values in the three sites: 0.37 (7/19) in BL, 0.30 (7/23) in FN3 and 0.32 (89/274) in VM. In contrast, there are differences in the values of the ratio of femur to tibia: 0.63 (20/32) in BL, 0.47 (14/30) in FN3 and 0.23 (107/465) in VM. This suggests that the hominins broke the major limb bones in BL and FN3 for exploiting their marrow contents and this led to the loss of the resource that would make them more attractive to the hyenas, thus explaining their better preservation in BL and FN3 than in VM.
In the Early Pleistocene, the environments of the Baza Basin were dominated by shallow lacustrine systems fed by the precipitation of meteoric waters on the lake surface as well as by the contribution of alluvial waters and thermal springs (García-Aguilar and Palmqvist, 2011; García-Aguilar et al., 2014, 2015). This led to deposits of marls, calcilutites, limestones, evaporites, sands, and dark lutites (Figure 2B). The lacustrine systems were dynamic in both time and space, as evidenced by the lateral wedging of the facies linked to these environments, which resulted in the appearance of non-flooded areas inhabited by the mammalian community and the hominins.
The cartography of the Late Villafranchian deposits that correspond to the archeological levels of BL and FN3 allows reconstructing their paleogeographic context (Figure 7). This was characterized by large flood plains crisscrossed by channels in the Guadix Basin as well as in the W and SW sectors of the Baza Basin, with a lacustrine system that spread through the N and NE sectors of the Baza Basin (Cortes de Baza-Huéscar-Orce sector, Figure 2B). The lacustrine deposits represent a stratigraphic unit with a maximum thickness of 40 m (up to 50–60 m in the lake depocenter), which preserves the main sites of Orce. The unit shows an alternation of levels of marls-calcilutites and limestones, each one meter thick on average, with intercalations of thin levels of dark lutites and detritic facies (conglomerates and sands) to the lake borders (García-Aguilar et al., 2014).
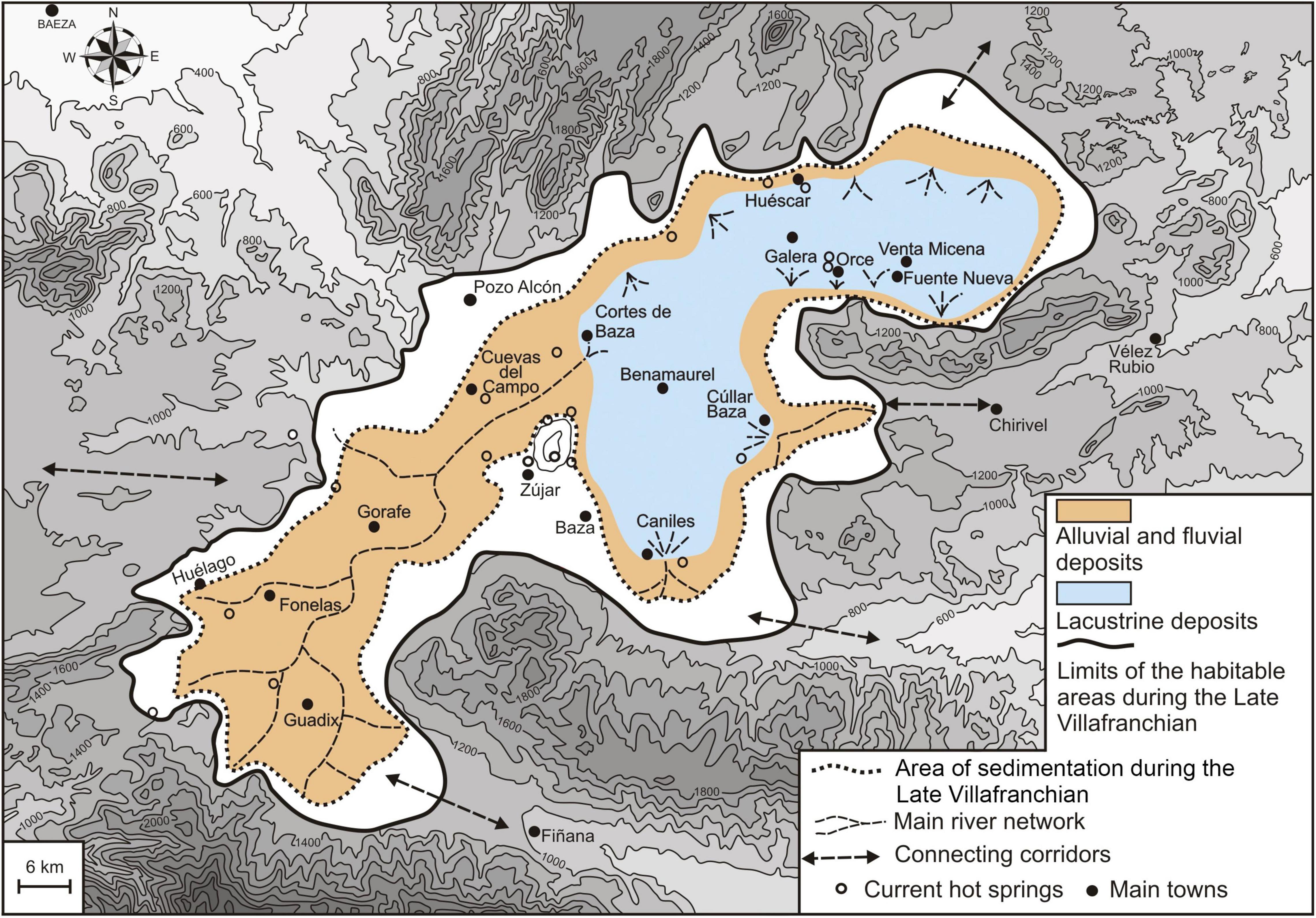
Figure 7. Reconstruction of the paleoenvironments of the Guadix-Baza Depression based on the cartographic extent of the Late Villafranchian sediments of the basin depicted in Figure 1B (for the stratigraphic series sampled, see García-Aguilar et al. (2014): Figure 3). The connecting corridors indicated by arrows are those open during the late Early Pleistocene, when the sedimentary basin was endorheic. The hydrographic network of the basin was captured in the late Middle Pleistocene by the Guadiana Menor River, a tributary of the Guadalquivir River. This led to a stage in which erosion dominated over sedimentation (i.e., a transit from an endorheic regime to an exorheic one). For this reason, the current topography that surrounds the depression shows a corridor to the west of the town of Pozo Alcón, but this connection did not exist during the late Early Pleistocene.
Several mollusks (gastropods Bithynia tentaculata, Hydrobia aff. acuta, Melanoides tuberculata, and Gyraulus cf. laevis; bivalve Pisidium casertanum; Figures 8E–I) indicate the presence of an euryhaline environment in BL (Albesa and Robles, 2020). The lacustrine sediments of BL preserve shells of two ostracods, Candona sp. and Cyprideis torosa (Anadón et al., 1994), which live in oligo- to mesosaline conditions (Figures 8A,B). The limits of tolerance to salinity of these species provide clues on the lake waters: B. tentaculate, H. acuta, and M. tuberculata can live with elevated salinities (12–30‰), but their optimum is lower (0.2–3‰ for M. tuberculata). In contrast, P. casertanum and G. laevis tolerate salinities of only up to 3–5‰ (Albesa and Robles, 2020). This indicates a thermophile lacustrine environment with alternating phases of salinity, from freshwater to moderately brackish conditions in the lake waters and higher values of salinity in the surrounding swampy areas. Such inference agrees with (i) the presence in the sediments of microcrystalline gypsum originated by neoformation (García-Aguilar et al., 2014); (ii) the high δ15N values measured in the bone collagen of Hippopotamus antiquus, a species that fed on the aquatic macrophytes that grew in the oligosaline waters of the lakes instead of consuming terrestrial grasses, as does the living H. amphibius (Palmqvist et al., 2003, 2008a,b,2022a; García-Aguilar et al., 2014); and (iii) the finding of a common Shelduck (Tadorna tadorna) in VM (Figure 8J). This waterfowl dwells in coastal mudflats and lagoons, estuaries, and riverine environments of Europe, generally occurring in salt or brackish water, where it feeds mostly on saltwater snail H. acuta (Carboneras and Kirwan, 2018), a species that can survive at salinities of up to 39‰ (Britton, 1985). In the case of VM, the ostracodes Ilyocypris bradyi and I. gibba (Figures 8C,D) provide additional information on the paleoenvironment: the former lives in high energy streams with abundant underwater vegetation, while the latter evidences shallow lake borders with warm and fresh to oligosaline waters, a sandy substrate and lushy vegetation (Martínez-García et al., 2015, 2017).
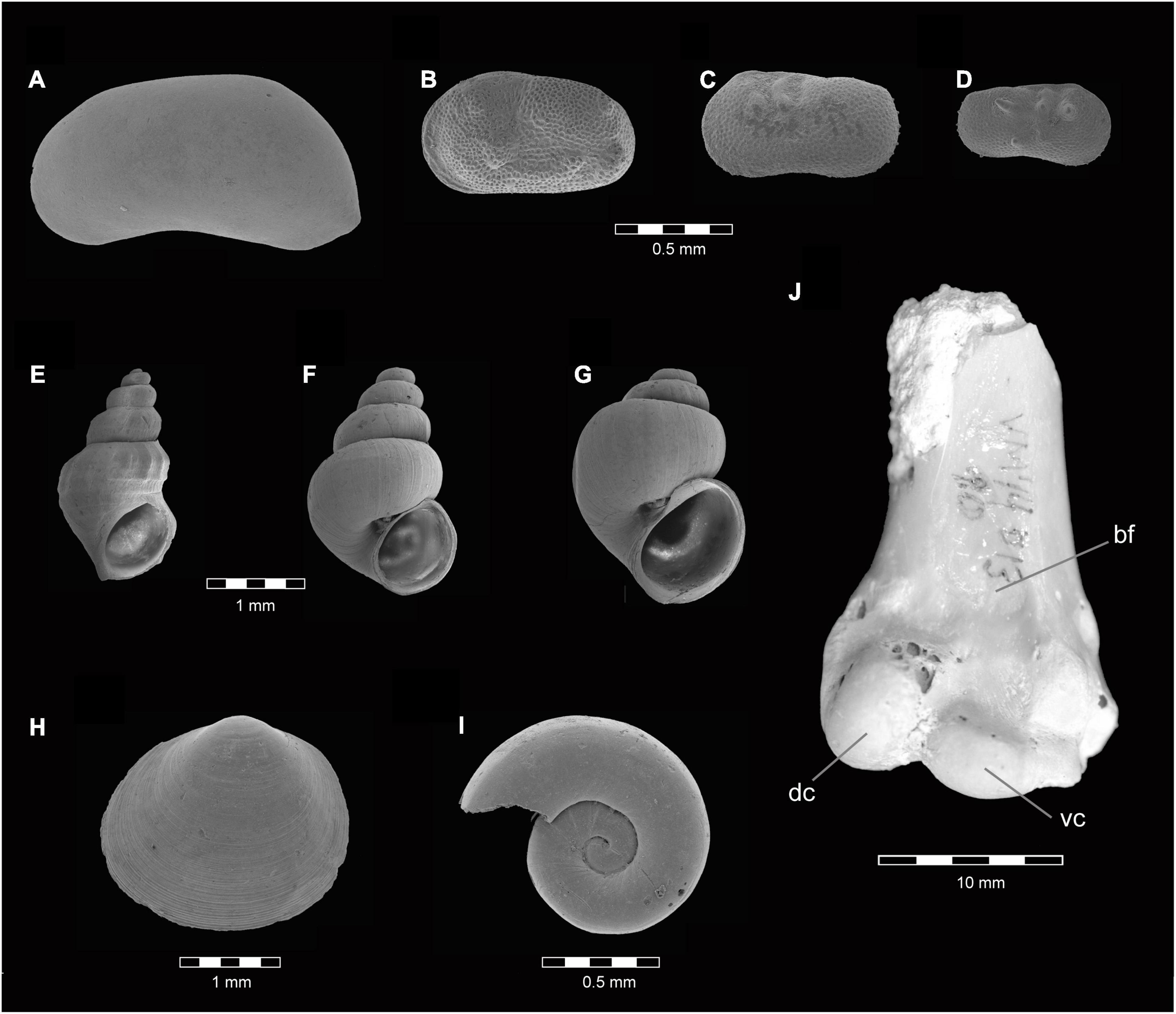
Figure 8. Ostracods, mollusks and avian remains identified in the Late Villafranchian archeological sites of Barranco León (BL) and Fuente Nueva 3 (FN3) in Orce (Guadix-Baza Depression, SE Spain). Scanning electron microscope photographs of ostracod (A–D) and mollusk (E–I) shells. (A) Candona sp. (B) Cyprideis torosa. (C) Ilyocypris bradyi. (D) Ilyocypris gibba. (E) Melanoides tuberculate. (F) Hydrobia aff. acuta. (G) Bithynia tentaculate. (H) Pisidium casertanum. (I) Gyraulus cf. laevis. (J) Distal humeral portion of an aquatic bird from Venta Micena (specimen VM-D13-10) in cranial view, which has been tentatively attributed to a Common Shelduck (Tadorna tadorna), bf-brachial fossa, dc-dorsal condyle, vc-ventral condyle.
The detritic facies of the Baza Basin are composed of sands and silts originated in organic-rich coastal lacustrine environments and emerged plains, although they can also correspond to distal riverine systems (García-Aguilar et al., 2014). Some sandy and silty deposits are associated with conglomerates, which represent flood deposits that penetrated the lakes as highly disorganized deltas (e.g., level D of BL; Arribas and Palmqvist, 2002).
The estimates of prey biomass (kg/km2) for the Orce sites (Figure 9), obtained with the Weibull model following the procedure described by Martín-González et al. (2019) and Rodríguez-Gómez et al. (2022), are lower than those measured in several African Natural Parks and Game Reserves, like Ngorongoro Crater or Amboseli, close to those of Masai Mara, Savuti, Hluhluwe Imfolozi or the Nwaswitshaka River, and higher than in Serengeti, Selous or the Okavango Delta, among many others. This figure shows a direct relationship of prey biomass with annual rainfall (Figure 9A), but not with mean annual temperature (Figure 9B). Prey biomass estimates suggest that meat availability was high in the Orce sites, particularly compared with most sites of Atapuerca. The only exceptions are TD6-1,2 and TD8, in which the estimates are only slightly lower than in BL and FN3. In fact, a comparison of the faunal assemblages of Atapuerca showed that TD6-1,2 and TD8 were the levels with lowest competition intensity among the secondary consumers (Rodríguez-Gómez et al., 2013, 2014a,b), even lower than in VM, BL and FN3. This indicates that, departing from a similar prey biomass, the Orce ecosystems supported a more diverse predatory guild than those recorded at TD6-1,2 and TD8 (Rodríguez-Gómez et al., 2016a,2017a). Based on a comparison between these results and those derived with the equations of Rodríguez et al. (2014) to estimate the maximum carrying capacity of the ecosystems of Orce and Atapuerca, Rodríguez-Gómez et al. (2022) proposed that the differences in secondary production between these sites could result from the exceptional geological conditions of Guadix-Baza, particularly the presence of thermal springs that provided a mild and productive paleoenvironment throughout the year (García-Aguilar and Palmqvist, 2011; García-Aguilar et al., 2013, 2014, 2015).
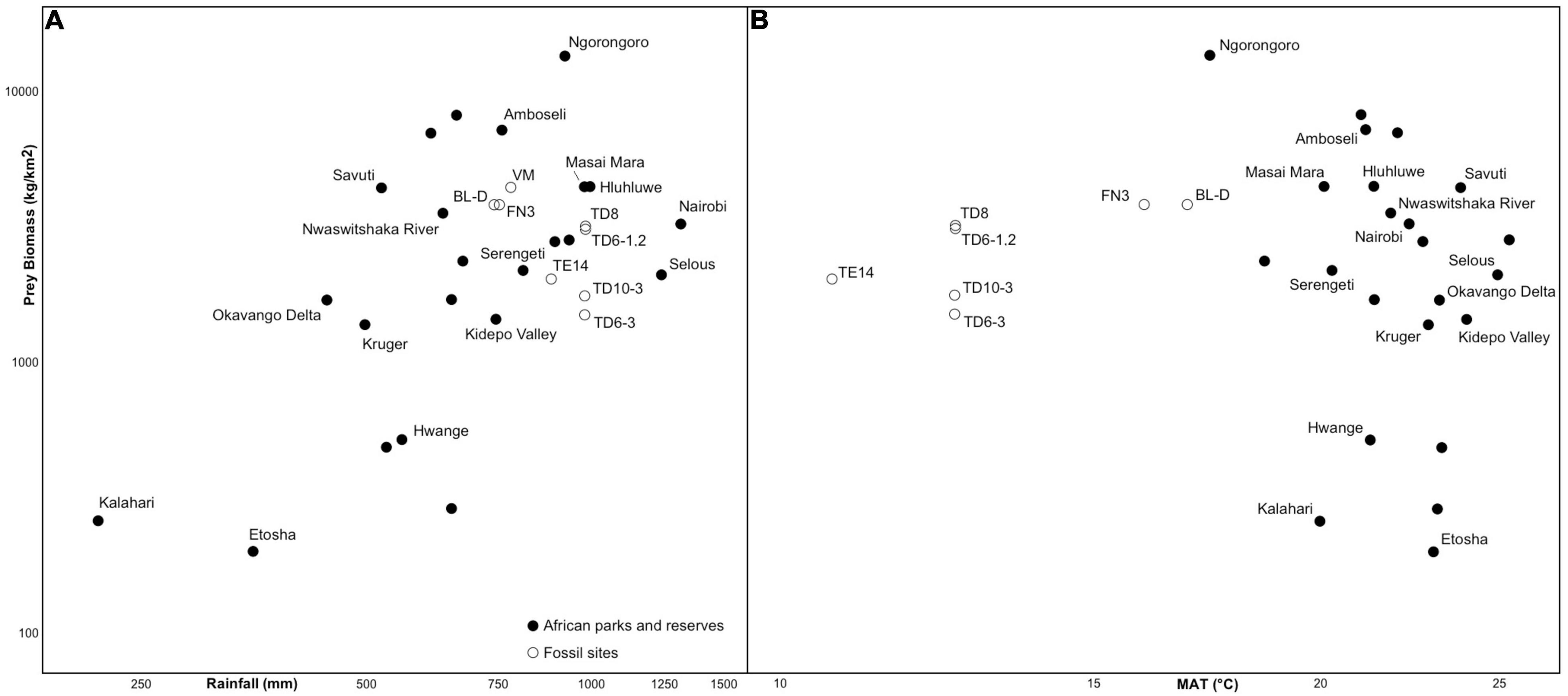
Figure 9. (A) Scatter plot of prey biomass (in kg/km2) and annual rainfall (in mm). (B) Scatter plot of prey biomass and mean annual temperature (MAT, in °C). The points correspond to 23 African Natural Parks and Game Reserves [data on prey biomass compiled by Hatton et al. (2015) from several sources; data on rainfall and MAT from Fick and Hijmans (2017)] and the paleontological sites of Atapuerca and Orce discussed in the text. The estimates on rainfall and MAT are from Blain et al. (2011a,b, 2016), García-Aguilar et al. (2014), and Rodríguez et al. (2014). Prey biomass values for the fossil assemblages were obtained using the Weibull model (see details in Martín-González et al. (2019)). TD, Trinchera Dolina, Atapuerca (NW Spain). TE, Sima del Elefante, Atapuerca. BL-D, Barranco León (Level D), Orce (Guadix-Baza Depression, SE Spain). FN3, Fuente Nueva 3, Orce. VM, Venta Micena, Orce. Scatter plots produced with software SPSS v. 25.
Figure 7 shows the paleogeographic model of Guadix-Baza during the late Early Pleistocene, based on cartographic data of the sedimentary deposits (Figure 2B). The outer perimeter of the basin, which corresponds to the limits of the surface emerged and the one covered by the lake waters, encloses an area of 4,050 km2. This encompasses the outcrop area of alluvial and fluvial deposits (i.e., flood plains, proximal zones of the alluvial systems and alluvial fans, covered by water only during the rainy episodes) and the glacis surface, which includes the area with a gentle slope in the foothills situated up to a height of 20 m over this surface. This represents a living area for the terrestrial fauna and the hominins of 2,925 km2, while the outcrop area of the lacustrine deposits covers 1,125 km2. The extent of the lake would fluctuate between low-stand stages, which correspond to the limestone and marly limestone beds deposited during the drops of the water table, and high-stand stages, which evidence the rising of the water table that resulted in the deposit of marls and calcilutites. The emerged area inhabited by the terrestrial fauna would be greater during the low-stand stages, when the alluvial feeding of the lake originated in the southern and eastern reliefs was scarce and of low energy. Given the limited depth of the lacustrine system (∼2 m on average, with large areas on the swampy environments of the lake margins showing a decametric depth), the lowering of the water table would result in wide emerged areas in the lake surroundings. The opposite situation would apply to the high-stand stages, when higher precipitations and a greater recharge of the lake by alluvial waters originated in the perimeter of the basin took place. This would result in a more restricted extent of the terrestrial ecosystems. Low-stand to high-stand fluctuations would encompass the periodic changes between warm-moist and cold-dry conditions of climatic cycles, which would represent variations in the living area for the terrestrial fauna of 20–25% above (low-stand) and below (high-stand) the cartographic extent of the deposits drawn in Figure 7. Moreover, there would be also yearly oscillations in the extent of the water sheet between the winter and summer seasons. Finally, Figure 7 shows the sedimentary depression during the late Early Pleistocene as a closed, endorheic basin with internal drainage, but with a limited number of connecting corridors for the terrestrial fauna to other surrounding areas, especially to the East. For this reason, the ecological scenario of the Orce sector of the sedimentary depression (a satellite basin of ∼170 km2) resembles the one found in the Ngorongoro Crater, Tanzania, which has a similar extent (∼230 km2) and hydrothermal context (Deocampo and Ashley, 1999; Deocampo, 2005).
The area emerged in the Guadix-Baza Depression (2,925 km2) allows calculating the population size of Homo sp. that could inhabit it. As noted earlier, the estimate of hominin population density for BL and FN-3 is 0.12 individuals per km2 considering a scavenging behavior as the optimal scenario for the procurement of ungulate carcasses (Rodríguez-Gómez et al., 2016a). This translates in a population size of ∼350 individuals, which would result in 7–12 hominin groups based on the estimates of mean local group size for modern hunter-gatherers (Binford, 1980; Marlowe, 2005). If we choose for a strict hunting behavior, which implies that the hominins had not access to the largest prey size classes, the population density would be 0.095 individuals per km2 (Rodríguez-Gómez et al., 2016a). This would result in a population of ∼280 individuals distributed among 5–9 foraging groups. Both estimates are very low and clearly below the minimum viable population size for mammals (including primates), which has been estimated in 3,876 individuals, with a 95% confidence interval of 2,261–5,095 individuals (Traill et al., 2007). This suggests that although the prey biomass estimated in the Guadix-Baza was similar or even higher than in many modern African ecosystems, the viability of the hominin population that inhabited the sedimentary basin could be compromised due to random oscillations in population size such as those resulting from fluctuations in resource availability and epidemics. This could eventually lead to bottlenecks, genetic drift, inbreeding depression, and local extinction.
A skull of the hypercarnivorous wild dog Lycaon lycaonoides (specimen VM-7000) provides evidence of inbreeding depression in the local population of this species. The skull preserves the cranium and mandible in anatomical connection and corresponds to a relatively old individual with moderately worn permanent dentition (Palmqvist et al., 1999). It displays a high degree of cranial fluctuating asymmetry (FA), which is especially marked in the frontal region, and shows dental anomalies, including agenesia of several teeth like the upper right canine (Bartolini-Lucenti et al., 2021). The incidence of FA in a population must be tested at the population level and not based on a single individual (Palmqvist et al., 1999). However, the fossil record of L. lycaonoides is scarce in Guadix-Baza (a few specimens from VM, see details in Palmqvist et al., 2022b) and VM-7000 is the only complete skull. This limitation on sample size precludes the hypothetico-deductive method and forces to use the ‘abductive research method,’ which means that the best explanation at hand for interpreting the anomalies of VM-7000 is to consider that they evidence FA resulting from developmental instabilities caused by inbreeding depression (Palmqvist et al., 1999; Bartolini-Lucenti et al., 2021).
Fluctuating asymmetry results from small, random accidents during ontogeny in morphological traits that would otherwise appear as bilaterally symmetric. These perturbations can also emerge from developmental stress (e.g., induced by parasitic infection), but their incidence in a population uses to correlate with the level of genetic inbreeding, which increases the homozygosity of enzyme polymorphisms and results in developmental instabilities (Wayne et al., 1986; Palmqvist et al., 1999; Leamy and Klingenberg, 2005). Inbreeding increasingly becomes a serious threat to local wildlife populations as habitats shrink and fragment (Spiering et al., 2011). This results in bottlenecks and changes in metapopulation structure, which translate in a quite small effective population size and lead to the loss of genetic diversity and inbreeding depression, as documented in the cheetah (Wayne et al., 1986; Menotti-Raymond and O’Brien, 1993; Hedrick, 1996; however, see also Merola (2011)). The expression of the deleterious effects of increased homozygosity and their consequences for individual fitness have been shown in canids (Ellegren, 1999; Fitzpatrick and Evans, 2009; Spiering et al., 2011). Anodontia and cranial asymmetry are documented in small populations of wolves subject to bottlenecks and inbreeding, as happens in Białowieża Forest (Buchalczyk et al., 1981; Vilà et al., 1993). Edwards et al. (2013) showed an increase in fluctuating asymmetry in museum skulls of African painted dogs that span a period of a hundred years, which parallels the decline in the populations of the species in sub-Saharan Africa during the last century. This suggests that the malformations of the fossil skull of VM would reflect developmental instabilities resulting from a high level of genetic homozygosity in the small population of wild dogs that inhabited Guadix-Baza during Early Pleistocene times. Specifically, the population density of L. lycaonoides expected for optimal ecological conditions, calculated using the equation of Damuth (1993) for African flesh-eaters in open environments, is 0.23 individuals per km2 (Rodríguez-Gómez et al., 2017a). However, the value of sustainable density for this predator in VM is lower, 0.13 ind./km2, which translates in a population of only 380 individuals (i.e., like the one estimated for the hominins). Moreover, the effective population size of modern painted dogs is typically reduced to 20–35% of the censused population by reproductive suppression of subordinates and uneven sex ratios (Creel and Creel, 1998). In the case of VM, this would result in an effective population size for L. lycaonoides of around one hundred individuals, which would have promoted further inbreeding. However, the dog from VM was able to reach adulthood, as shown by its moderately worn permanent dentition, despite severe developmental handicaps, which suggests that cooperative behavior from other members of the hunting pack may have helped it to survive (Palmqvist et al., 1999; Bartolini-Lucenti et al., 2021). Similarly, cooperation among hominins has been reported in Dmanisi. This site documents the earliest human presence out of Africa (∼1.8 Ma) and preserves a human skull that lost all but one of its teeth several years before the time of death (composite skull D 3444/D 3900), as evidenced by extensive bone loss due to resorption of the alveolar processes. Although the etiology of the absence of teeth differs from that described in the wild dog of VM, the survival of this edentulous individual probably relied also on the assistance (e.g., food chewing) from other members of the group (Lordkipanidze et al., 2005, Lordkipanidze et al., 2006).
The high level of genetic homozygosis deduced for the population of L. lycaonoides that inhabited Guadix-Baza —conditions that would presumably apply also to the hominin population— can be explained as resulting from its small size, as this population was geographically isolated from other similar ecosystems in the surrounding areas (Palmqvist et al., 1999). Interestingly, the lion population that inhabits the Ngorongoro Crater depicts a similar situation: these lions form a small, naturally isolated population of 75–125 individuals, which has been historically subject to severe bottlenecks followed by limited recolonization by lions from the nearby Serengeti ecosystem. This has resulted in high levels of inbreeding and lack of genetic variability in the contemporary Crater lion population compared to the much larger Serengeti population, which correlates with increased levels of sperm abnormality and decreased reproductive performance (Packer et al., 1991).
Any discussion on the level of genetic variability of the population of early Homo that inhabited Guadix-Baza makes it necessary to consider the levels of inbreeding depression withstood by modern hunter-gatherers. Genealogical information showed evidence of inbreeding in 165 out of 931 (∼18%) individuals of the Hadza population (Stevens et al., 1977). Similarly, the !Kung, Khwe and African Pygmies exhibit low genetic diversity coupled with high frequencies of divergent mtDNA haplotypes not found in the surrounding agricultural groups, which suggests small population sizes and long-term isolation, respectively (Vigilant et al., 1991; Chen et al., 2000; Oota et al., 2005). Moreover, Trinkaus (2018) showed an unexpectedly high frequency of abnormalities in the fossil record of Pleistocene Homo, including from minor but rare dental, vertebral, and carpal variants to exceptional systemic disorders. This suggests ubiquitous stress among the Pleistocene foragers and/or high levels of consanguinity.
Although the hominin population of Guadix-Baza was probably small, it would not remain in strict isolation: there are several connecting corridors in the mountainous reliefs that surround the basin (see arrows in Figure 7). Moreover, some species ecologically and climatically constrained like the hippo probably dispersed by these corridors from the coastal environments, which led to their colonization of the lacustrine environments of this endorheic basin. In the case of the hominins, the corridors would result in some gene flow with other populations from adjacent areas, although the amount of gene introgression was probably limited, as happens in modern hunter-gatherers. Moreover, data from Binford (1980: Table 1) for Equatorial and Subequatorial populations of non-equestrian hunters-gatherers provide a mean number of 24.8 residential moves per group and year, a mean distance of 15.3 km between sites and a mean total circuit distance covered annually of 310.6 km. This suggests that each of the few hominin groups that inhabited the Guadix-Baza Depression would travel each year searching for food a linear distance that would cover the entire basin. These changes in residential moves would presumably follow a random motion, as happens in the case of modern hunter-gatherers (Brown et al., 2007; Raichlen et al., 2014). For this reason, the probability of intersecting one of the five connecting corridors of the basin perimeter (Figure 7) in any of these foraging random walks would be low, resulting in a limited amount of gene flow with the surrounding populations and long-term isolation.
According to these data, Guadix-Baza was home for a small hominin population during the late Early Pleistocene, which viability in the medium and long term could be compromised by population bottlenecks and inbreeding depression. This explains the sporadic and discontinuous nature of the archeological record in the sedimentary basin. Although such situation cannot be extrapolated to the whole European record of hominin presence for these ancient chronologies, the scarcity of Early Pleistocene archeological sites points to intermittent dispersal events, which probably reflects hominin incursions that failed to establish a more permanent character (Dennell, 2003; MacDonald et al., 2012).
However, we must introduce here a cautionary note: Flores Island, which has an area of 13,540 km2 (i.e., ∼4.6 times the living area of the Guadix-Baza Depression), has a record of hominin presence since ∼0.7 Ma (van den Bergh et al., 2009, 2016). The island allowed the survival of a local population of H. erectus during more than half a million years and this happened without any evidence of external gene flow having been detected so far. In their adaptation to the insular environment, these hominins evolved in a short time to a dwarfed form, H. floresiensis, which represents a striking case of evolutionary reversal in the trend of body and brain size increase in Homo (van den Bergh et al., 2016). This was facilitated by the relatively small size of the population, although the energy expenditure of the small-bodied H. floresiensis could be half that of H. erectus (Dennell et al., 2014), which probably resulted in a higher population density. Unlike other hominins, H. floresiensis shows a combination of primitive traits in the canine and premolar teeth, which are comparable to those of Early Pleistocene H. erectus, together with a molar morphology more progressive than in modern humans (Kaifu et al., 2015). These features make sense in the context of a highly isolated inbred population with small genetic variation (as expected from the founder effect of the island colonizers), a population that was adaptively constrained. It should be noted, however, that a similar combination of primitive and modern characters is also present in H. luzonensis, the dwarfed hominin of the larger island of Luzon (Philippines), which probably descended also from an initial population of H. erectus (Détroit et al., 2019).
During the last decades, the discovery of a significant number of Early Pleistocene sites that preserve evidence of hominin presence in Western Europe has renewed the debate on: (i) the chronology of the oldest dispersal of Homo out of Africa; (ii) the success of these ancient populations in the colonization of the European landscapes; and (iii) their continuity or discontinuity (Arribas and Palmqvist, 1999; Roebroeks, 2001; Dennell, 2003; MacDonald et al., 2012; Bermúdez de Castro et al., 2013; Rodríguez-Gómez et al., 2014a,2016a,2017a,b,c). Although Dmanisi provides conclusive evidence on the presence of hominins at the gates of Europe by ∼1.8 Ma (García et al., 2010), the overall evidence of hominin settlements in North Africa and Europe is scarce before the Middle Pleistocene and is basically restricted to the circum-Mediterranean realm (Rodríguez-Gómez et al., 2016a,2017a; Butynski, 1982; Capaldo, 1997; Roebroeks et al., 2018). Muttoni et al. (2010, 2013) proposed that the first occurrence of Homo sp. in Southern Europe took place between the Jaramillo subchron and the Brunhes-Matuyama boundary (i.e., in the 0.99–0.78 Ma interval), a time window that encompasses the latest Early Pleistocene global climate transition centered on MIS 22 at ∼0.9 Ma, the first prominent cold stage of the Pleistocene. Under this view, hominin presence in Southern Europe would have been ephemeral from ∼1 Ma onward, with occasional short northward expansions along the western coastal areas when temperate conditions allowed it (Roebroeks, 2001; Muttoni et al., 2010). In contrast, Leroy et al. (2011) argued that there were up to 42 possible narrow windows of 41 ka for hominin dispersal in Europe through the Early Pleistocene (2.58–0.78 Ma). In their model of climate and vegetation change, these windows coincided with transitions from glacial to interglacial cycles forced by obliquity, which resulted in the opening of the landscapes with the appearance of grasslands and forested steppes like those of Orce (Mendoza et al., 2005; Saarinen et al., 2021). This landscape was similar to the recent reconstruction of Dmanisi as an open wooded savannah and grassland by Bartolini-Lucenti et al. (2022). Leroy et al. (2011) considered that the presence of hominins was not possible during the full glacial periods, which would be too cold for them, and neither in the transitions from interglacial to glacial cycles, when the landscapes were densely forested. In any case, the oldest conclusive evidence of human presence in Western Europe is found at BL and FN3 in Orce (∼1.4 Ma) and TE9 level (∼1.2 Ma) in Atapuerca (see review in Palmqvist et al., 2016). In addition, the carnivore guild of VM, a site ∼200 ka older than BL and FN3 that shows no evidence of hominin presence, has a lower level of competition intensity for meat than in the case of BL and FN3 (Rodríguez-Gómez et al., 2016a,2017a). This suggests that the delay of ∼0.4 Ma between hominin arrival in the Caucasus and Western Europe was not a matter of ecological opportunity and other factors (climatic or geographic barriers to dispersal) played a role here (Rodríguez-Gómez et al., 2017a).
The scarcity of Early Pleistocene archeological sites in Europe indicates that the colonizing capabilities of early Homo outside Africa may have been overestimated, as the dispersals were not automatically followed by permanent settlements (Dennell, 2003). Further fieldwork will undoubtedly fill many gaps in the Early Pleistocene hominin record of Europe, but current evidence is consistent with the view that: (i) the oldest populations outside East Africa were spatially and temporally discontinuous; (ii) hominin expansion was strongly constrained by latitude; and (iii) the occupation of temperate regions north of latitude 40° was largely confined to interglacial periods (Dennell, 2003; Leroy et al., 2011; Rodríguez et al., 2011, 2013; MacDonald et al., 2012; Bermúdez de Castro et al., 2013). However, continuity of hominin settlements in the refuge areas of Southern Europe during the cold periods of the Early Pleistocene cannot be discarded: the presence of hominins at Atapuerca during long time periods suggests that they were well-adapted to the hard and seasonal conditions of the northern hemisphere at a latitude of 42° and an altitude of 1000 m (Bermúdez de Castro et al., 2013). Climate conditions in Guadix-Baza were milder than those of Atapuerca (Figure 9), with the presence of open plains with woodland patches and water-edge areas (Mendoza et al., 2005; Saarinen et al., 2021). This difference between the North and South of the Iberian Peninsula is also seen today (Blain et al., 2009, 2011a,b). Moreover, the thermal springs of Guadix-Baza resulted in a milder and more productive environment in the lacustrine systems of the basin compared to present-day conditions (García-Aguilar et al., 2014, 2015). Therefore, the information available suggests continuity for the human settlements in the Iberian Peninsula during a period of at least 300 ka before the Jaramillo subchron (Agustí et al., 2009; Bermúdez de Castro et al., 2013). According to Agustí et al. (2009), after this phase the hominins would disappear from Western Europe during a long period that included part of the Jaramillo subchron and until MIS 22 (0.88–0.87 Ma). This would be caused by an extreme decrease in mean annual temperature and an increase in mean annual precipitation, as deduced from the herpetofauna (Agustí et al., 2009). However, geochronological and paleomagnetic data for levels of Trinchera Dolina of Atapuerca indicate a continued presence of hominins in northern Spain during the time interval of ∼1.0–0.3 Ma (Bermúdez de Castro et al., 2013).
The archeological record of Sima del Elefante and Gran Dolina cave sites in Atapuerca, which preserves relatively continuous stratigraphic sequences that stretch back from ∼1.2 Ma to the Matuyama/Brunhes boundary, suggests that the Iberian Peninsula was occupied by at least two different hominin populations (Bermúdez de Castro et al., 2013). The human remains from level TD6 of Gran Dolina, dated to 0.97–0.79 Ma by a combination of biochronology, magnetostratigraphy and geochronology (Duval et al., 2018; Parés et al., 2018), have been ascribed to Homo antecessor, a species considered to represent the most recent common ancestor of Denisovans, Neandertals and modern humans, or at least very close to the cladogenetic event that preceded the divergence of these species (Bermúdez de Castro et al., 2017; Welker et al., 2020). The dental proteome of H. antecessor indicates that it represents a close sister lineage to subsequent Middle and Late Pleistocene hominins (Welker et al., 2020). In Sima del Elefante, the few fossils from TE9 do not allow to conclude if H. antecessor has deep roots in the Early Pleistocene before the Jaramillo subchron (Bermúdez de Castro et al., 2013, 2017). The hominin population of TE could be different from that recorded at TD6, but the evidence recovered is too scanty to make a strong claim. What seems clear is that the hominins never disappeared from the Atapuerca habitats, as the archeological record at the base of TD4 is of ∼1.0 Ma (Carbonell and Rodríguez, 1994). This argues for the continuity of the hominin settlements in the circum-Mediterranean realm, even if we admit the possibility of discontinuity by the local substitution of the original population, its assimilation or crossbreeding.
Finally, Bermúdez de Castro and Martinón-Torres (2013) hypothesized the existence of a “source population” of hominins in a central area of dispersal, which they tentatively located in the Levantine Corridor. When climatic and ecological conditions were favorable, hominins would colonize from this area the Eastern and Western territories of Eurasia. Following Carrión et al. (2011); Bermúdez de Castro and Martinón-Torres (2013) considered that the key for activating evolutionary change in this hominin population would be the geological instabilities that resulted in an increase in physiographical heterogeneity, biodiversity and ecological interaction. The latter conditions were boosted in the Guadix-Baza Depression by the abundance of thermal springs linked to intense tectonic activity, which was a major determinant in the establishment of biodiversity “hot spots” with high biological productivity (García-Aguilar et al., 2014, 2015; Rodríguez-Gómez et al., 2016a,2017a,2022; Palmqvist et al., 2022a).
Evidence of anthropogenic action from the archeological levels of BL and FN-3 indicates that the hominin population that inhabited the Guadix-Baza Depression during Early Pleistocene times exploited the prey carcasses left abandoned by saber-tooths, which suggests a subsistence strategy based on passive scavenging rather than on active hunting for obtaining meat, fat, and bone marrow. In addition, other animals of small size available in the environment, including rodents, leporids, tortoises and birds, were also presumably consumed, together with eggs, invertebrates, honey, and a wide spectrum of edible vegetation. The population size estimated for the hominins of the basin was low, ∼350–280 individuals distributed in five to twelve groups of hunter-gatherers. This probably compromised the long-term viability of the population due to the effects of bottlenecks and inbreeding depression, as detected in the coeval population of wild dogs, which probably led them to extinction and helps to explain the scarcity and discontinuous nature of the archeological record in the basin.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
PP wrote the manuscript and all co-authors made contributions. SR-M, BM-N, and PP prepared Figure 1. AG-M, JMG-A, AG, and SR-M prepared Figure 2. BF prepared Figure 3. GR-G prepared Figure 4. PP and MPE prepared Figures 5, 6. AG-M and JMG-A prepared Figure 7. AG and FS prepared Figure 8. GR-G and PP prepared Figure 9. BM-N, MPE, and SR-M prepared Table 1. All authors contributed to the article and approved the submitted version.
This work was supported by four projects from the Spanish Ministry of Science, Innovation and University (Grant Nos. CGL-2016-78577-P, CGL-2016-80975-P, PGC2018-093925-B-C31, and PID2019-111185GB-I00), by two projects from Junta de Andalucía (Grant Nos. UMA18-FEDERJA-18 and P18-FR-3193), by one project from Generalitat de Catalunya (Grant No. GENCAT 2017SGR 859), and by Research Group RNM-146 from Junta de Andalucía.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
GR-G was funded by the program IdEx Bordeaux “Investments for the future,” financed by L’Agence Nationale de la Recherche of France (ref. ANR No.-10-IDEX-03-02) with the project Analysis of Kozarnika food webs and comparison with other Pleistocene sites of Western Europe. The Program Juan de la Cierva of the Spanish Ministry of Science, Innovation and University funded GR-G (FJCI-2016-28652) and FS (IJCI-2017-32116). GR-G is supported by an “Atracción de Talento” postdoctoral contract (2019-T2/HUM-13370) from the Comunidad de Madrid and Universidad Complutense de Madrid (Spain). FS is supported by a postdoctoral fellowship of Junta de Andalucía. Last but not least, we gratefully acknowledge the insightful comments and helpful criticism from two reviewers and also from the editor, MA.
Abbazzi, L. (2010). “La fauna de Cérvidos de Barranco León y Fuente Nueva 3,” in Ocupaciones Humanas en el Pleistoceno inferior y medio de la cuenca de Guadix-Baza, Memoria Científica. Junta de Andalucía. Consejería de Cultura. E.P.G. Arqueología Monográfico, eds Toro and A. Martínez-Navarro (Spain: Red de Bibliotecas Universitarias), 273–290.
Agam, A., and Barkai, R. (2018). Elephant and mammoth hunting during the Paleolithic: A review of the relevant archaeological, ethnographic and ethno-historical records. Quaternary 2018:1. doi: 10.3390/quat1010003
Agustí, J., Blain, H.-A., Cuenca-Bescós, G., and Bailon, S. (2009). Climate forcing of first hominid dispersal in Western Europe. J. Hum. Evol. 57, 815–821. doi: 10.1016/j.jhevol.2009.06.005
Aiello, L. C., and Wells, J. C. K. (2002). Energetics and the evolution of the genus Homo. Annu. Rev. Anthropol. 31, 323–338. doi: 10.1146/annurev.anthro.31.040402.085403
Aiello, L. C., and Wheeler, P. (1995). The expensive-tissue hypothesis: the brain and the digestive system in human and primate evolution. Curr. Anthropol. 36, 199–221. doi: 10.1086/204350
Akersten, W. A. (1985). Canine function in Smilodon (Mammalia: Felidae: Machairodontidae), Vol. 356. Los Angeles: Contribution Science Natural History Museum, 1–22.
Alberdi, M. T. (2010). “Estudio de los caballos de los yacimientos de Fuente Nueva-3 y Barranco León-5 (Granada),” in Ocupaciones Humanas en el Pleistoceno inferior y medio de la cuenca de Guadix-Baza, Memoria Científica. Junta de Andalucía. Consejería de Cultura. E.P.G. Arqueología Monográfico, eds I. Toro, B. Martínez-Navarro, and J. Agustí 291–306.
Alberdi, M. T., and Ruiz-Bustos, A. (1985). Descripción y significado bioestratigráfico del Equus e Hipopótamo en el yacimiento de Venta Micena (Granada). Estud. Geol. 41, 251–261. doi: 10.3989/egeol.85413-4708
Albesa, J., and Robles, F. (2020). Contribución al conocimiento de los moluscos del Pleistoceno Inferior del yacimiento de Barranco León (Cuenca de Guadix-Baza). Spanish J. Palaeontol. 35, 125–146. doi: 10.7203/sjp.35.1.17324
Álvarez, C., Parés, J. M., Granger, D., Duval, M., Sala, R., and Toro, I. (2015). New magnetostratigraphic and numerical age of the Fuente Nueva-3 site (Guadix-Baza basin Spain). Quat. Int. 389, 224–234. doi: 10.1016/j.quaint.2015.04.044
Anadón, P., and Gabàs, M. (2009). Paleoenvironmental evolution of the early Pleistocene lacustrine sequence at Barranco León archeological site (Orce, Baza Basin, Southern Spain) from stable isotopes and Sr and Mg chemistry of ostracod shells. J. Paleolimnol. 42, 261–279. doi: 10.1007/s10933-008-9275-6
Anadón, P., Utrilla, R., and Julià, R. (1994). Palaeoenvironmental reconstruction of a Pleistocene lacustrine sequence from faunal assemblages and ostracode shell geochemistry. Baza Basin, SE Spain. Palaeogeogr. Palaeoclimatol. Palaeoecol. 111, 191–205. doi: 10.1016/0031-0182(94)90062-0
Andersson, K., Norman, D., and Werdelin, L. (2011). Sabretoothed carnivores and the killing of large prey. PLoS One 6:e24971. doi: 10.1371/journal.pone.0024971
Antón, M., Salesa, M. J., Pastor, J. F., Sánchez, I. M., Fraile, S., and Morales, J. (2004). Implications of the mastoid anatomy of larger extant felids for the evolution and predatory behaviour of sabretoothed cats (Mammalia, Carnivora, Felidae). Zool. J. Linn. Soc. 140, 207–221. doi: 10.1111/j.1096-3642.2003.00093.x
Anyonge, W. (1996a). Locomotor behaviour in Plio-Pleistocene sabre-tooth cats: a biomechanical analysis. J. Zool. 238, 395–413. doi: 10.1111/j.1469-7998.1996.tb05402.x
Anyonge, W. (1996b). Microwear on canines and killing behavior in large carnivores: saber function in Smilodon fatalis. J. Mammal. 77, 1059–1067. doi: 10.2307/1382786
Arribas, A., Garrido, G., Viseras, C., Soria, J. M., Pla, S., Solano, J. G., et al. (2009). A mammalian lost world in Southwest Europe during the Late Pliocene. PLoS One 4:e7127. doi: 10.1371/journal.pone.0007127
Arribas, A., and Palmqvist, P. (1998). Taphonomy and paleoecology of an assemblage of large mammals: hyaenid activity in the lower Pleistocene site at Venta Micena (Orce, Guadix-Baza Basin, Granada, Spain). Geobios 31(Suppl.), 3–47. doi: 10.1016/s0016-6995(98)80056-9
Arribas, A., and Palmqvist, P. (1999). On the ecological connection between sabre-tooths and hominids: faunal dispersal events in the lower Pleistocene and a review of the evidence for the first human arrival in Europe. J. Archaeol. Sci. 26, 571–585. doi: 10.1006/jasc.1998.0346
Arribas, A., and Palmqvist, P. (2002). The first human dispersal to Europe: remarks on the archaeological and palaeoanthropological record from Orce (Guadix-Baza basin, southeastern Spain). Hum. Evol. 17, 55–78. doi: 10.1007/bf02436429
Arzarello, M., Marcolini, F., Pavia, G., Pavia, M., Petronio, C., Petrucci, M., et al. (2007). Evidence of earliest human occurrence in Europe: the site of Pirro Nord (southern Italy). Naturwissenschaften 94, 107–112. doi: 10.1007/s00114-006-0173-3
Arzarello, M., Peretto, C., and Moncel, M. H. (2015). The Pirro Nord site (Apricena, Fg, Southern Italy) in the context of the first European peopling: Convergences and divergences. Quat. Int. 389, 255–263. doi: 10.1016/j.quaint.2014.08.051
Ascenzi, A., Biddittu, I., Cassoli, P. F., Segre, A. G., and Segre-Naldini, E. (1996). A calvarium of late Homo erectus from Ceprano, Italy. J. Hum. Evol. 31, 409–423. doi: 10.1006/jhev.1996.0069
Barr, W. A., Pobiner, B., Rowan, J., Dud, A., and Faith, J. T. (2022). No sustained increase in zooarchaeological evidence for carnivory after the appearance of Homo erectus. Proc. Nat. Acad. Sci. USA 119:e2115540119. doi: 10.1073/pnas.2115540119
Barsky, D., Celiberti, V., Cauche, D., Grégire, S., Lebègue, F., Lumley, H., et al. (2010). Raw material discernment and technological aspects of the Barranco León and Fuente Nueva 3 stone assemblages (Orce southern Spain). Quat. Int. 223-224, 201–219. doi: 10.1016/j.quaint.2009.12.004
Barsky, D., Vergès, J. M., Sala, R., Menéndez, L., and Toro, I. (2016). Limestone percussion tools from the late Early Pleistocene sites of Barranco León and Fuente Nueva 3 (Orce. Spain). Phil. Trans. R. Soc. B 370:20140352. doi: 10.1098/rstb.2014.0352
Bartolini-Lucenti, S., Cirilli, O., Pandolfi, L., Bernor, R. L., Bukhsianidze, M., Carotenuto, F., et al. (2022). Zoogeographic significance of Dmanisi large mammal assemblage. J. Hum. Evol. 163:103125. doi: 10.1016/j.jhevol.2021.103125
Bartolini-Lucenti, S., and Madurell-Malapeira, J. (2020). Unraveling the fossil record of foxes: an updated review on the Plio-Pleistocene Vulpes spp. from Europe. Quat. Sci. Rev. 236:106296. doi: 10.1016/j.quascirev.2020.106296
Bartolini-Lucenti, S., Madurell-Malapeira, J., Martínez-Navarro, B., Palmqvist, P., Lordkinapidze, D., and Rook, D. (2021). The early hunting dog from Dmanisi with comments on the social behaviour in Canidae and hominins. Sci. Rep. 11:13501. doi: 10.1038/s41598-021-92818-4
Ben-Dor, M., Sirtoli, R., and Barkai, R. (2021) The evolution of the human trophic level during the Pleistocene. Yearbook Phys. Anthropol. 175, 27–56. doi: 10.1002/ajpa.24247
Bermúdez de Castro, J. M., Arsuaga, J. L., Carbonell, E., Rosas, A., Martínez, I., et al. (1997). A hominid from the Lower Pleistocene of Atapuerca, Spain: possible ancestor to Neandertals and modern humans. Science 276, 1392–1395. doi: 10.1126/science.276.5317.1392
Bermúdez de Castro, J. M., and Martinón-Torres, M. (2013). A new model for the evolution of the human Pleistocene populations of Europe. Quat. Int. 295, 102–112. doi: 10.1016/j.quaint.2012.02.036
Bermúdez de Castro, J. M., Martinón-Torres, M., Arsuaga, J. L., and Carbonel, E. (2017). Twentieth anniversary of Homo antecessor (1997-2017): a review. Evol. Anthropol. 26, 157–171. doi: 10.1002/evan.21540
Bermúdez de Castro, J. M., Martinón-Torres, M., Blasco, R., Rosell, J., and Carbonell, E. (2013). Continuity or discontinuity in the European Early Pleistocene human settlement: The Atapuerca evidence. Quat. Sci. Rev. 76, 53–65. doi: 10.1016/j.quascirev.2013.06.023
Bickerton, D., and Szathmáry, E. (2011). Confrontational scavenging as a possible source for language and cooperation. BMC Evol. Biol. 11:261. doi: 10.1186/1471-2148-11-261
Biknevicius, A. R., Van Valkenburgh, B., and Walker, J. (1996). Incisor size and shape: implications for feeding behaviors in saber-toothed “cats”. J. Vertebr Paleontol. 16, 510–521. doi: 10.1080/02724634.1996.10011336
Binford, L. R. (1980). Willow smoke and dogs’ tails: hunter-gatherer settlement systems and archaeological site formation. Am. Antiq. 45, 4–20. doi: 10.2307/279653
Binford, L. R. (2001). Constructing frames of reference. An analytical method for archaeological theory building using ethnographic and environmental data sets. Berkeley: University of California Press, 563.
Blain, H.-A., Bailón, S., Agustí, J., Martínez-Navarro, B., and Toro, I. (2011a). Paleoenvironmental and paleoclimatic proxies to the Early Pleistocene hominids of Barranco León D and Fuente Nueva 3 (Granada, Spain) by means of their amphibian and reptile assemblages. Quat. Int. 243, 44–53. doi: 10.1016/j.quaint.2010.12.031
Blain, H.-A., Bailón, S., Cuenca-Bescós, G., Bennàsa, M., Rofes, J., López-García, J. M., et al. (2011b). Climate and environment of the earliest amphibian and squamata reptile assemblages: Sima del Elefante Lower Red Unit, Atapuerca, Spain. Quat. Sci. Rev. 29, 3034–3044. doi: 10.1016/j.quascirev.2010.07.006
Blain, H.-A., Bailon, S., Cuenca-Bescós, G., Arsuaga, J. L., Bermúdez, de Castro, J. M., et al. (2009). Long-term climate record inferred from early-middle Pleistocene amphibian and squamate reptile assemblages at the Gran Dolina Cave, Atapuerca, Spain. J. Hum. Evol. 56, 55–65. doi: 10.1016/j.jhevol.2008.08.020
Blain, H.-A., Fagoaga, A., Ruiz-Sánchez, F. J., García-Medrano, P., Ollé, A., and Jiménez-Arenas, J. M. (2021). Coping with arid environments: a critical threshold for human expansion in Europe at the Marine Isotope Stage 12/11 transition? The case of the Iberian Peninsula. J. Hum. Evol. 153:102950. doi: 10.1016/j.jhevol.2021.102950
Blain, H.-A., Lozano-Fernández, I., Agustí, J., Bailón, S., Menéndez, L., Espigares, M. P., et al. (2016). Refining upon the climatic background of the Early Pleistocene hominid settlement in Western Europe: barranco León and Fuente Nueva-3 (Guadix-Baza Basin, SE Spain). Quat. Sci. Rev. 144, 132–144. doi: 10.1016/j.quascirev.2016.05.020
Blasco, R., Blain, H.-A., Rosell, J., Díez, J. C., Huguet, R., Rodríguez, J., et al. (2011). Earliest evidence for human consumtion of tortoises in the European Early Pleistocene from Sima del Elefante, Sierra de Atapuerca, Spain. J. Hum. Evol. 61, 503–509. doi: 10.1016/j.jhevol.2011.06.002
Blumenschine, R. J. (1986). Early hominid scavenging opportunities. In: Implications of Carcass Availability in the Serengeti and Ngorongoro Ecosystems. BAR Int. Ser. 1986:283.
Blumenschine, R. J. (1987). Characteristics of an early hominid scavenging niche. Curr. Anthropol. 28, 383–407. doi: 10.1086/203544
Blumenschine, R. J. (1991). Hominid carnivory and foraging strategies, and the socioeconomic function of early archaeological sites. Phil.Trans. R.Soc. 334, 211–221. doi: 10.1098/rstb.1991.0110
Blumenschine, R. J. (1995). Percussion marks, tooth marks, and experimental determinations of the timing of hominid and carnivore access to long bones at FLK Zinjanthropus, Olduvai Gorge, Tanzania. J. Hum. Evol. 29, 21–25. doi: 10.1006/jhev.1995.1046
Blumenschine, R. J., Cavallo, J. A., and Capaldo, S. D. (1994). Competition for carcasses and early hominid behavioral ecology: a case study and a conceptual framework. J. Hum. Evol. 27, 197–213. doi: 10.1006/jhev.1994.1042
Blumenschine, R. J., and Pobiner, B. L. (2007). “Zooarchaeology and the ecology of Oldowan hominin carnivory,” in Evolution of the Human Diet, ed. P. Ungar (Oxford: University Press), 167–190.
Blumenschine, R. J., Prassack, K. A., Kreger, C. D., and Pante, M. C. (2007). Carnivore tooth marks, microbial bioerosion, and the invalidation of Domínguez-Rodrigo and Barba’s (2006) test of Oldowan hominin scavenging behavior. J. Hum. Evol. 53, 420–426. doi: 10.1016/j.jhevol.2007.01.011
Blumenschine, R. J., and Selvaggio, M. M. (1988). Percussion marks on bone surfaces as a new diagnostic of hominid behaviour. Nature 333, 763–765. doi: 10.1038/333763a0
Bourguignon, L., Crochet, J. Y., Capdevila, R., Ivorra, J., Antoine, P. O., Agustí, J., et al. (2016). Bois-de-Riquet (Lézignan-la-Cèbe, Hérault): a late Early Pleistocene archeological occurrence in southern France. Quat. Int. 393, 24–40. doi: 10.1016/j.quaint.2015.06.037
Braga, J. C., Martín, J. M., and Quesada, C. (2003). Patterns and average rates of late Neogene-Recent uplift of the Betic Cordillera, SE Spain. Geomorphology 50, 3–26. doi: 10.1016/s0169-555x(02)00205-2
Brain, C. K. (1981). The Hunters or the Hunted: An introduction to African Cave Taphonomy. Chicago: The University of Chicago Press.
Bramble, D., and Lieberman, D. (2004). Endurance running and the evolution of Homo. Nature 432, 345–352. doi: 10.1038/nature03052
Brink, J. W. (1997). Fat content in leg bones of Bison bison, and applications to archaeoloy. J. Archaeolog.Sci. 24, 259–274. doi: 10.1006/jasc.1996.0109
Britton, R. H. (1985). Life cycle and production of Hydrobia acuta Drap. (Gastropoda: Prosobranchia) in a hypersaline coastal lagoon. Hydrobiologia 122, 219–230. doi: 10.1007/bf00018282
Brown, C. T., Liebovitch, L. S., and Glendon, R. (2007). Leìvy flights in Dobe Ju/’hoansi foraging patterns. Hum. Ecol. 35, 129–138. doi: 10.1007/s10745-006-9083-4
Buchalczyk, T., Dynowski, J., and Szteyn, S. (1981). Variations in number of teeth and asymmetry of the skull in the wolf. Acta Theriolog. 26, 23–30. doi: 10.4098/at.arch.81-2
Bunn, H. T. (1981). Archaeological evidence for meat-eating by Plio-Pleistocene hominids from Koobi Fora and Olduvai Gorge. Nature 291, 574–576. doi: 10.1038/291574a0
Bunn, H. T. (2001). “Hunting, power scavenging, and butchering by Hadza foragers and by Plio-Pleistocene Homo,” in Meat-eating and Human Evolution, eds C. B. Stanford and H. T. Bunn (Oxford: Oxford University Press), 199–218.
Bunn, H. T. (2007). “Meat made us human,” in Evolution of the Human Diet, ed. P. Ungar (Oxford: Oxford University Press), 191–211.
Bunn, H. T., and Ezzo, J. A. (1993). Hunting and scavenging by Plio-pleistocene hominids: nutritional contraints, archaeological patterns, and behavioural implications. J. Archaeolog. Sci. 20, 365–398. doi: 10.1006/jasc.1993.1023
Bunn, H. T., and Pickering, T. R. (2010). Bovid mortality profiles in paleoecological context falsify hypotheses of endurance running-hunting and passive scavenging by early Pleistocene hominins. Quat. Res. 74, 395–404. doi: 10.1016/j.yqres.2010.07.012
Bunn, H. T., Pickering, T. R., and Domínguez-Rodrigo, M. (2017). “How meat made us human: Archaeological evidence of the diet and foraging capabilities of early Pleistocene Homo in East Africa,” in The Oxford Handbook of the Archaeology of Diet, eds J. Lee-Thorp and M. A. Katzenberg (Oxford: Oxford University Press), 1330.
Butynski, T. M. (1982). Vertebrate predation by primates: a review of hunting patterns and prey. J. Hum. Evol. 11, 421–430. doi: 10.1016/s0047-2484(82)80095-x
Capaldo, S. D. (1997). Experimental determinations of carcass processing by Plio-Pleistocene hominids and carnivores at FLK 22 (Zinjanthropus), Olduvai Gorge, Tanzania. J. Hum. Evol. 33, 555–597. doi: 10.1006/jhev.1997.0150
Carbonell, E., Bermúdez, de Castro, J. M., Parés, J. M., Pérez-González, A., Cuenca-Bescós, G., et al. (2008). The first hominin of Europe. Nature 452, 465–470.
Carbonell, E., Mosquera, M., Rodríguez, X. P., and Sala, R. (1995). The first human settlement of Europe. J. Anthropol. Res. 51, 107–114. doi: 10.1086/jar.52.1.3630238
Carbonell, E., and Rodríguez, X. P. (1994). Early Middle Pleistocene deposits and artefacts in the Gran Dolina site (TD4) of the “Sierra de Atapuerca” (Burgos, Spain). J. Hum. Evol. 26, 291–311. doi: 10.1006/jhev.1994.1018
Carboneras, C., and Kirwan, G. M. (2018). “Common Shelduck (Tadorna tadorna),” in Handbook of the Birds of the World Alive, eds J. del Hoyo, A. Elliott, J. Sargatal, D. A. Christie, and E. de Juana (Barcelona: Lynx Edicions).
Carrión, J. S. (2002). A taphonomic study of modern pollen assemblages from dung and surface sediments in arid environments of Spain. Rev. Palaeobot. Palynol. 120, 217–232. doi: 10.1016/s0034-6667(02)00073-8
Carrión, J. S., Fernández, S., González-Sampériz, P., Leroy, S. A. G., Bailey, G. N., López-Sáez, J. A., et al. (2009). Quaternary pollen analysis in the Iberian Peninsula: the value of negative results. Internet Archaeol. 25, 1–53. doi: 10.1016/j.quascirev.2017.11.016
Carrión, J. S., Rose, J., and Stringer, C. B. (2011). Early human evolution in the western Palaeartic: ecological scenarios. Quat. Sci. Rev. 30, 1281–1295. doi: 10.1016/j.quascirev.2011.04.003
Cauche, D. (2022). The Vallonnet cave on the northern Mediterranean border: A record of one of the oldest human presences in Europe. L’Anthropologie 126:102974. doi: 10.1016/j.anthro.2021.102974
Cheheb, R. C., Arzarello, M., Arnaud, J., Berto, C., Cáceres, I., Caracausi, S., et al. (2019). Human behavior and Homo-mammal interactions at the first European peopling: new evidence from the Pirro Nord site (Apricena, Southern Italy). Sci. Nat. 106:16. doi: 10.1007/s00114-019-1610-4
Chen, Y. S., Olckers, A., Schurr, T. G., Kogelnik, A., Huoponen, K., and Wallace, D. C. (2000). mtDNA variation in the South African Kung and Khwe—and their genetic relationships to other African populations. Am. J. Hum. Genet. 66, 1362–1383. doi: 10.1086/302848
Cherin, M., Alba, D. M., Crotti, M., Menconero, S., Moullé, P.-E., Sorbelli, L., et al. (2020). The post-Jaramillo persistence of Sus strozzii (Suidae, Mammalia) in Europe: New evidence from the Vallparadís Section (NE Iberian Peninsula) and other coeval sites. Quat. Sci. Rev. 233:106234. doi: 10.1016/j.quascirev.2020.106234
Cherin, M., Sorbelli, L., Crotti, M., Iurino, D. A., Sardella, R., and Souron, A. (2018). New material of Sus strozzii (Suidae, Mammalia) from the Early Pleistocene of Italy and a phylogenetic analysis of suines. Quat. Sci. Rev. 194, 94–115. doi: 10.1016/j.quascirev.2018.06.029
Christiansen, P. (2008). Evolution of skull and mandible shape in cats (Carnivora: Felidae). PLoS One 3:e2807. doi: 10.1371/journal.pone.0002807
Coil, R., Tappen, M., Ferring, R., Bukhsianidze, M., Nioradze, M., and Lordkipanidze, D. (2020). Spatial patterning of the archaeological and paleontological assemblage at Dmanisi, Georgia: an analysis of site formation and carnivore-hominin interaction in Block 2. J. Hum. Evol. 143, 102773. doi: 10.1016/j.jhevol.2020.102773
Coltorti, M., Feraud, G., Marzolic, A., Peretto, C., Ton-Thate, T., Voinchet, P., et al. (2005). New 40Ar/39Ar, stratigraphic and palaeoclimatic data on the Isernia La Pineta Lower Palaeolithic site, Molise, Italy. Quat. Int. 131, 11–22. doi: 10.1016/j.quaint.2004.07.004
Cordain, L., Miller, J. B., Eaton, S. B., Mann, N., Holt, S. H. A., and Speth, J. D. (2000). Plant-animal subsistence ratios and macronutrient energy estimations in worldwide hunter-gatherer diets. Am. J. Clin. Nutr. 71, 682–692. doi: 10.1093/ajcn/71.3.682
Creel, S., and Creel, N. M. (1998). Six ecological factors that may limit African wild dogs. Lycaon pictus. An. Cons. 1, 1–9. doi: 10.1111/j.1469-1795.1998.tb00220.x
Cregut-Bonnoire, E. (1999). “Les petits Bovidae de Venta Micena (Andalousie) et de Cueva Victoria (Murcia),”. in The Hominids and their environment during the Lower and Middle Pleistocene of Eurasia eds J, Gibert F, Sánchez L, Gibert and F, Ribot (Orce: Proc. Int. Conf. Human Paleont.) 1995, 191–228.
Crochet, J. Y., Welcomme, J. L., Ivorra, J., Ruffet, G., Boulbes, N., Capdevila, R., et al. (2009). Une nouvelle faune de vertebras continentaux, associée à des artefacts dans le Pléistocène inférieur de l’Hérault (Sud de la France), vers 1,57 Ma. C. R. Palevol 8, 725–736. doi: 10.1016/j.crpv.2009.06.004
Damuth, J. (1993). Cope’s rule, the island rule and the scaling of mammalian population density. Nature 365, 748–750. doi: 10.1038/365748a0
De Lombera-Hermida, A., Rodríguez-Alvarez, X. P., and Sala, R. (2016). The lithic assemblage from Pont-de-Lavaud (Indre, France) and the role of the bipolar-on-anvil technique in the Lower and Early Middle Pleistocene technology. J. Anthropol. Archaeol. 41, 159–184. doi: 10.1016/j.jaa.2015.12.002
Dennell, R. (2003). Dispersal and colonisation, long and short chronologies: how continuous is the Early Pleistocene record for hominins outside East Africa? J. Hum. Evol. 45, 421–440. doi: 10.1016/j.jhevol.2003.09.006
Dennell, R., and Roebroeks, W. (2006). An Asian perspective on early human dispersal from Africa. Nature 438, 1099–1104. doi: 10.1038/nature04259
Dennell, R. W., Louys, J., O’Regan, H. J., and Wilkinson, D. M. (2014). The origins and persistence of Homo floresiensis on Flores: biogeographical and ecological perspectives. Quat. Sci.Rev. 96, 98–107. doi: 10.1016/j.quascirev.2013.06.031
Deocampo, D., and Ashley, G. M. (1999). Siliceous islands in a carbonate sea: modern and Pleistocene records of spring-fed wetlands in Ngorongoro Crater and Olduvai Gorge, Tanzania. J. Sed. Res. 69, 974–979. doi: 10.2110/jsr.69.974
Deocampo, D. M. (2005). Evaporative evolution of surface waters and the role of aqueous CO2 in magnesium silicate precipitation: lake Eyasi and Ngorongoro Crater, northern Tanzania. S. Afr. J. Geol. 108, 493–504. doi: 10.2113/108.4.493
DeSantis, L. R. G., Feranec, R. S., Antón, M., and Lundelius, E. L. (2021). Dietary ecology of the scimitar-toothed cat Homotherium serum. Curr. Biol. 2021:61. doi: 10.1016/j.cub.2021.03.061
Détroit, F., Mijares, A. S., Corny, J., Daver, G., Zanolli, C., Dizon, E., et al. (2019). A new species of Homo from the Late Pleistocene of the Philippines. Nature 568, 181–186. doi: 10.1038/s41586-019-1067-9
Domínguez-Rodrigo, M. (1999). Flesh availability and bone modifications in carcasses consumed by lions: palaeoecological relevance in hominid foraging patterns. Palaeogeogr., Palaeoclimatol. Palaeoecol. 149, 373–388. doi: 10.1016/s0031-0182(98)00213-2
Domínguez-Rodrigo, M., and Barba, R. (2006). New estimates of tooth mark and percussion mark frequencies at the FLK Zinj site: the carnivore-hominid-carnivore hypothesis falsified. J. Hum. Evol. 50, 170–194. doi: 10.1016/j.jhevol.2005.09.005
Domínguez-Rodrigo, M., Bunn, H. T., and Yravedra, J. (2014). A critical re-evaluation of bone surface modification models for inferring fossil hominin and carnivore interactions through a multivariate approach: application to the FLK Zinj archaeofaunal assemblage (Olduvai Gorge. Tanzania). Quat. Int. 32, 32–43. doi: 10.1016/j.quaint.2013.09.042
Domínguez-Rodrigo, M., Egeland, C. P., and Barba, R. (2007). “The hunting-versus scavenging debate,” in Deconstructing Olduvai: a Taphonomic Study of the Bed I Sites, eds M. Domínguez-Rodrigo, R. Barba, and C. P. Egeland (New York, NY: Springer), 11–22. doi: 10.1371/journal.pone.0194324
Domínguez-Rodrigo, M., and Organista, E. (2007). “Natural background bone assemblages and their ravaging stages in Olduvai Bed I,” in Deconstructing Olduvai: A Taphonomic Study of the Bed I Sites, eds M. Domínguez-Rodrigo, R. Barba, and C. P. Egeland (The Netherlands: Springer), 201–215. doi: 10.1007/978-1-4020-6152-3_11
Domínguez-Rodrigo, M., and Piqueras, A. (2003). The use of tooth pits to identify carnivore taxa in tooth-marked archaofaunas and their relevance to reconstruct hominid carcass processing behaviours. J. Archaol.Sci. 30, 1385–1391. doi: 10.1016/s0305-4403(03)00027-x
Duncan, P., Foose, T. J., Gordon, I. J., Gakahu, C. G., and Loyd, M. (1990). Comparative nutrient extraction from forages by grazing bovids and equids: a test of the nutritional model of equid-bovid competition and coexistence. Oecologia 84, 411–418. doi: 10.1007/BF00329768
Duval, M., Bahain, J.-J., Falguères, C., Garcia, J., Guilarte, V., Grün, R., et al. (2015). Revisiting the ESR chronology of the Early Pleistocene hominin occupation at Vallparadís (Barcelona, Spain). Quat. Int. 389, 213–223. doi: 10.1016/j.quaint.2014.08.054
Duval, M., Falguères, C., Bahain, J. J., Grun, R., Shao, Q., Aubert, M., et al. (2012). On the limits of using combined U-series/ESR method to date fossil teeth from two Early Pleistocene archaeological sites of the Orce area (Guadix-Baza basin, Spain). Quat. Res. 77, 481–482.
Duval, M., Grün, R., Parés, J. M., Martín-Francés, L., Campaña, I., Rosell, J., et al. (2018). The first direct ESR dating of a hominin tooth from Atapuerca Gran Dolina TD-6 (Spain) supports the antiquity of Homo antecessor. Quat. Geochronol. 47, 120–137. doi: 10.1016/j.quageo.2018.05.001
Edwards, C. T. T., Rasmussen, G. S. A., Riordan, P., Courchamp, F., and Macdonald, D. W. (2013). Non-adaptive phenotypic evolution of the endangered carnivore Lycaon pictus. PLoS One 8:e73856. doi: 10.1371/journal.pone.0073856
Ellegren, H. (1999). Inbreeding and relatedness in Scandinavian grey wolves Canis lupus. Hereditas 130, 239–244. doi: 10.1111/j.1601-5223.1999.00239.x
Emerson, S. B., and Radinsky, L. (1980). Functional analysis of sabertooth cranial morphology. Paleobiology 6, 295–312. doi: 10.1017/s0094837300006813
Espigares, M. P. (2010). Análisis y modelización del contexto sedimentario y los atributos tafonómicos de los yacimientos pleistocénicos del borde nororiental de la cuenca de Guadix-Baza. Ph. D. Dissertation. Spain: University of Granada, 533.
Espigares, M. P., Martínez-Navarro, B., Palmqvist, P., Ros-Montoya, S., Toro, I., Agustí, J., et al. (2013). Homo vs. Pachycrocuta: Earliest evidence of competition for an elephant carcass between scavengers at Fuente Nueva-3 (Orce, Spain). Quat. Int. 295, 113–125. doi: 10.1016/j.quaint.2012.09.032
Espigares, M. P., Palmqvist, P., Guerra-Merchán, A., Ros-Montoya, S., García-Aguilar, J. M., Rodríguez-Gómez, G., et al. (2019). The earliest cut marks of Europe: a discussion on hominin subsistence patterns in the Orce sites (Baza basin, SE Spain). Sci. Rep. 9:15408. doi: 10.1038/s41598-019-51957-5
Fick, S. E., and Hijmans, R. J. (2017). Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315. doi: 10.1002/joc.5086
Figueirido, B., Lautenschlager, S., Pérez-Ramos, A., and Van Valkenburgh, B. (2018). Distinct predatory behaviors in scimitar-and dirk-toothed sabertooth cats. Curr. Biol. 28, 3260–3266. doi: 10.1016/j.cub.2018.08.012
Figueirido, B., MacLeod, N., Krieger, J., de Renzi, M., Pérez-Claros, J. A., and Palmqvist, P. (2011). Constraint and adaptation in the evolution of carnivoran skull shape. Paleobiology 37, 490–518. doi: 10.1666/09062.1
Fitzpatrick, J. L., and Evans, J. P. (2009). Reduced heterozygosity impairs sperm quality in endangered mammals. Biol. Let. 5, 320–323. doi: 10.1098/rsbl.2008.0734
Gabunia, L., and Vekua, A. (1995). A Plio-Pleistocene hominid from Dmanisi, East Georgia, Caucasus. Nature 373, 509–512. doi: 10.1038/373509a0
Gallotti, R., Muttoni, G., Lefèvre, D., Degeai, J. P., Geraads, D., Zerboni, A., et al. (2021). First high resolution chronostratigraphy for the early North African Acheulean at Casablanca (Morocco). Sci. Rep. 11:15340. doi: 10.1038/s41598-021-94695-3
Garcia, J., Martínez, K., and Carbonell, E. (2013). The Early Pleistocene stone tools from Vallparadís (Barcelona, Spain): rethinking the European mode 1. Quat. Int. 316, 94–114. doi: 10.1016/j.quaint.2013.09.038
García, T., Féraud, G., Falguères, C., Lumley, H., Perrenoud, C., and Lordkipanidze, D. (2010). Earliest human remains in Eurasia: New 40Ar/39Ar dating of the Dmanisi hominid-bearing levels, Georgia. Quat. Geochronol. 5, 443–451. doi: 10.1016/j.quageo.2009.09.012
García-Aguilar, J. M., Guerra-Merchán, A., Serrano, F., Flores-Moya, A., Delgado-Huertas, A., Espigares, M. P., et al. (2015). A reassessment of the evidence for hydrothermal activity in the Neogene-Quaternary lacustrine environments of the Baza basin (Betic Cordillera, SE Spain) and its paleoecological implications. Quat. Sci. Rev. 112, 226–235. doi: 10.1016/j.quascirev.2015.01.014
García-Aguilar, J. M., Guerra-Merchán, A., Serrano, F., and Palmqvist, P. (2013). Sedimentary cyclicity in early Pleistocene, evaporitic, playa-lake lacustrine deposits in the Guadix-Baza basin (Betic Cordillera, Spain). Bol. Geol. Min. 124, 239–251.
García-Aguilar, J. M., Guerra-Merchán, A., Serrano, F., Palmqvist, P., Flores-Moya, A., and Martínez-Navarro, B. (2014). Hydrothermal activity and its paleoecological implications in the latest Miocene to Middle Pleistocene lacustrine environments of the Baza Basin (Betic Cordillera, SE Spain). Quat. Sci. Rev. 96, 204–221. doi: 10.1016/j.quascirev.2013.07.009
García-Aguilar, J. M., and Palmqvist, P. (2011). A model of lacustrine sedimentation for the Lower Pleistocene deposits of Guadix-Baza basin (southeast Spain). Quat. Int. 243, 3–15. doi: 10.1016/j.quaint.2011.02.008
Gidna, A. O., Kisui, B., Mabulla, A., Musiba, C., and Domínguez-Rodrigo, M. (2014). An ecological neo-taphonomic study of carcass consumption by lions in Tarangire National Park (Tanzania) and its relevance for human evolutionary biology. Quat. Int. 322-323, 167–180. doi: 10.1016/j.quaint.2013.08.059
Gonyea, W. J. (1976). Behavioral implications of saber-toothed felid morphology. Paleobiology 2, 332–342. doi: 10.1371/journal.pone.0052453
Goren-Inbar, N., Feibel, C. S., Verosub, K. L., Melamed, Y., Kislev, M. E., Tchernov, E., et al. (2000). Pleistocene milestones on the out-of-Africa corridor at Gesher Benot Ya‘aqov, Israel. Science 289, 944–947. doi: 10.1126/science.289.5481.944
Guerra-Merchán, A. (1990). Sobre la conexión entre la Depresión de Guadix-Baza y el Corredor del Almanzora. (Cordilleras Béticas, Andalucía oriental). Geogaceta 8, 97–99.
Guerra-Merchán, A. (1993). La cuenca neógena del corredor del Almanzora. Ph.D. Thesis Dissertation, ed. S. A. Edinford (Spain: University of Grenade).
Güleç, E., White, T. D., Kuhn, S., Özer, I., Sagir, M., Yilmaz, H., et al. (2009). The Lower Pleistocene lithic assemblage from Dursunlu (Konya), central Anatolia, Turkey. Antiquity 83, 11–22. doi: 10.1017/s0003598x00098057
Guthrie, D. (1990). Frozen Fauna of Mammoth Steppe: The Story of Blue Babe. Chicago: The University Chicago Press, 342.
Hardy, K., Radini, A., Buckley, S., Blasco, R., Copeland, L., Burjachs, F., et al. (2017). Diet and environment 1.2 million years ago revealed through analysis of dental calculus from Europe’s oldest hominin at Sima del Elefante, Spain. Sci. Nat. 104:2. doi: 10.1007/s00114-016-1420-x
Hatton, I. A., McCann, K. S., Fryxell, J. M., Davies, T. J., Smerlak, M., Sinclair, A. R. E., et al. (2015). The predator-prey power law: Biomass scaling across terrestrial and aquatic biomes. Science 349:aac6284. doi: 10.1126/science.aac6284
Hawkes, K., O’connell, J. F., Blurton Jones, G., Oftedal, O. T., and Blumenshchine, R. J. (1991). Hadza: Big game, common goods, foraging goals, and the evolution of the human diet. Phil.Trans. R.Soc. Lond. B 334, 243–251. doi: 10.1098/rstb.1991.0113
Hayward, M. W., and Kerley, G. I. H. (2005). Prey preferences of the lion (Panthera leo). J. Zool. 267, 309–322. doi: 10.1017/s0952836905007508
Hedrick, P. (1996). Bottleneck(s) or metapopulation in cheetahs. Conserv. Biol. 10, 897–899. doi: 10.1046/j.1523-1739.1996.10030897.x
Ho, K. J., Mikkelson, B., Lewis, L. A., Feldman, S. A., and Taylor, C. B. (1972). Alaskan Arctic Eskimos: responses to a customary high fat diet. Am. J. Clin. Nutr. 25, 737–745. doi: 10.1093/ajcn/25.8.737
Husing, S. K., Oms, O., Agustí, J., Garcés, M., Kouwenhoven, T. J., Krijgsman, W., et al. (2010). On the late Miocene closure of the Mediterranean-Atlantic gateway through the Guadix basin (southern Spain). Palaeogeograp., Palaeoclimatol., Palaeoecol. 291, 167–179. doi: 10.1016/j.palaeo.2010.02.005
Hyodo, M., Matsu’ura, S., Kamishima, Y., Kondo, M., Takeshita, Y., Kitaba, I., et al. (2011). High-resolution record of the Matuyama-Brunhes transition constrains the age of Javanese Homo erectus in the Sangiran dome, Indonesia. Proc. Nat. Acad. Sci. USA 108, 19563–19568. doi: 10.1073/pnas.1113106108
Janis, C. M. (1976). The evolutionary strategy of the Equidae and the origins of rumen and caecal fermentation. Evolution 30, 757–774. doi: 10.1111/j.1558-5646.1976.tb00957.x
Janis, C. M., Gordon, I. J., and Illius, A. W. (1984). Modelling equid/ruminant competition in the fossil record. Hist. Biol. 8, 15–29. doi: 10.1080/10292389409380469
Jiménez-Arenas, J. M., Pérez-Claros, J. A., Aledo, J. C., and Palmqvist, P. (2014). On the relationships of postcanine tooth size with dietary quality and brain volume in Primates: Implications for hominin evolution. BioMed. Res. Int. 406507:11. doi: 10.1155/2014/406507
Joubert, D. (2006). Hunting behaviour of lions (Panthera leo) on elephants (Loxodonta africana) in the Chobe National Park, Botswana. Afr. J. Ecol. 44, 279–281. doi: 10.1111/j.1365-2028.2006.00626.x
Kaifu, Y., Kono, R. T., Sutikna, T., Saptomo, E. W., and Jatmiko Due Awe, R. (2015). Unique Dental morphology of Homo floresiensis and its evolutionary implications. PLoS One 10:e0141614. doi: 10.1371/journal.pone.0141614
Kuipers, R. S., Luxwolda, M. F., Dijck-Brouwer, D. A. J., Eaton, S. B., Crawford, M. A., Cordain, L., et al. (2010). Estimated macronutrient and fatty acid intakes from an East African Paleolithic diet. Br. J. Nutr. 104, 1666–1687. doi: 10.1017/S0007114510002679
Lacombat, F. (2010). “Estudio paleontológico de Stephanorhinus hundsheimensis de Fuente Nueva 3 y de Barranco León 5,” in Ocupaciones Humanas en el Pleistoceno inferior y medio de la Cuenca de Guadix-Baza. Arqueología Monografías, eds I. Toro, B. Martínez-Navarro, and J. Agustí (Sevilla: Junta de Andalucía, Consejería de Cultura), 237–246.
Lam, Y. M., Chen, X., and Pearson, O. M. (1999). Intertaxonomic variability in patterns of bone density and the differential representation of bovid, cervid, and equid elements in the archaeological record. Am. Antiq. 64, 343–362. doi: 10.2307/2694283
Larick, R., and Ciochon, R. L. (1996). The African emergence and early Asian dispersals of the genus Homo. Am. Sci. 84, 538–551.
Larick, R., Ciochon, R. L., Zaim, Y., Sudijono Suminto, Rizal, Y., et al. (2001). Early Pleistocene 40Ar/39Ar ages for Bapang Formation hominins, Central Jawa, Indonesia. Proc. Nat. Acad. Sci. USA 98, 4866–4871. doi: 10.1073/pnas.081077298
Leamy, L. J., and Klingenberg, C. P. (2005). The genetics and evolution of fluctuating asymmetry. An. Rev. Ecol., Evol. Syst. 36, 1–21.
Lebatard, A.-E., Alçiçek, M. C., Rochette, P., Khatib, S., Vialet, A., Boulbes, N., et al. (2014). Dating the Homo erectus bearing travertine from Kocabas (Denizli, Turkey) at least 1.1 Ma. Earth Planet. Sci. Let. 390, 8–18. doi: 10.1016/j.epsl.2013.12.031
Lee, R. B. (1968). “What hunters do for a living, or how to make out on scarce resources,” in Man the hunter, eds R. B. Lee and I. DeVore (Chicago: Aldine Publishing Co), 30–48. doi: 10.4324/9780203786567-6
Lee, R. B. (1979). The !Kung San: Men, Women and Work in a Foraging Society. Cambridge, MA: Cambridge University Press, 556.
Leonard, W., and Robertson, M. L. (1994). Evolutionary perspectives on human nutrition: The influence of brain and body size on diet and metabolism. Am. J. Hum. Biol. 6, 77–88. doi: 10.1002/ajhb.1310060111
Leonard, W. R., and Robertson, M. L. (1996). On diet, energy metabolism, and brain size in human evolution. Curr. Anthropol. 37, 125–129. doi: 10.1086/204476
Leroy, S. A. G., Arpe, K., and Mikolajewicz, U. (2011). Vegetation context and climatic limits of the Early Pleistocene hominin dispersal in Europe. Quat. Sci. Rev. 30, 1448–1463. doi: 10.1016/j.quascirev.2010.01.017
Lewis, S. G., Ashton, N., Field, M. H., Hoare, P. G., Kamermans, H., Knul, M., et al. (2019). Human occupation of northern Europe in MIS 13: Happisburgh Site 1 (Norfolk, UK) and its European context. Quat. Sci. Rev. 211, 34–58. doi: 10.1016/j.quascirev.2019.02.028
Liebenberg, L. (2008). The relevance of persistence hunting to human evolution. J. Hum. Evol. 55, 1156–1159. doi: 10.1016/j.jhevol.2008.07.004
Lieberman, D. E., Bramble, D. M., Raichlen, D. A., and Shea, J. J. (2007). The evolution of endurance running and the tyranny of ethnography: a reply to Pickering and Bunn J. Hum. Evol. 53, 439–442. doi: 10.1016/j.jhevol.2007.07.002
Lisiecki, L. E., and Raymo, M. E. (2005). A Pliocene-Pleistocene stack of 57 globally distributed benthic d18O records. Paleoceanography 20:A1003. doi: 10.1029/2004PA001071
Lordkipanidze, D. (2015). “The traces of the first humans in Eurasia,” in Human Origin Sites and the World Heritage Convention in Eurasia, Vol. I, ed. N. Sanz (Paris: United Nations Educational, Scientific and Cultural Organization), 153–162.
Lordkipanidze, D., Ponce, de León, M. S., Margvelashvili, A., Rak, Y., Rightmire, G. P., et al. (2013). A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo. Science 342, 326–331. doi: 10.1126/science.1238484
Lordkipanidze, D., Vekua, A., Ferring, R., Rightmire, G. P., Agustí, J., Kiladze, G., et al. (2005). The earliest toothless hominin skull. Nature 434, 717–718. doi: 10.1038/434717b
Lordkipanidze, D., Vekua, A., Ferring, R., Rightmire, G. P., Zollikofer, C. P., and Ponce, et al. (2006). A fourth hominin skull from Dmanisi, Georgia. Anat. Rec. A 288, 1146–1157. doi: 10.1002/ar.a.20379
Loveridge, A. J., Hun, J. E., Murindagom, F., and Macdonal, D. W. (2006). Influence of drought on predation of elephant (Loxodonta africana) calves by lions (Panthera leo) in an African wooded savannah. J. Zool. 270, 523–530. doi: 10.1111/j.1469-7998.2006.00181.x
Lozano-Fernández, I., Blain, H.-A., López-García, J. M., and Agustí, J. (2014). Biochronology of the first hominid remains in Europe using the vole Mimomys savini: Fuente Nueva 3 and Barranco León D, Guadix-Baza Basin, south-eastern Spain. Hist. Biol. 27, 1021–1028. doi: 10.1080/08912963.2014.920015
Lozano-Fernández, I., Cuenca-Bescós, G., Blain, H.-A., López-García, J. M., and Agustí, J. (2013). Mimomys savini size evolution in the Early Pleistocene of south-western Europe and possible biochronological implications. Quat. Sci. Revi. 76, 96–101. doi: 10.1016/j.quascirev.2013.06.029
Luzón, C., Yravedra, J., Courtenay, L. A., Saarinen, J., Blain, H. A., DeMiguel, D., et al. (2021). Taphonomical and spatial analyses from the Early Pleistocene site of Venta Micena 4 (Orce, Guadix-Baza, Basin, southern Spain). Sci. Rep. 11:13977. doi: 10.1038/s41598-021-93261-1
MacDonald, K., Martinón-Torres, M., Dennell, R., and Bermúdez de Castro, J. M. (2012). Discontinuity in the record for hominin occupation in south-western Europe: implications for occupation of the middle latitudes of Europe. Quat. Int. 271, 84–97. doi: 10.1016/j.quaint.2011.10.009
Madurell-Malapeira, J., Martínez-Navarro, B., Ros-Montoya, S., Espigares, M. P., Toro, I., and Palmqvist, P. (2011). The earliest European badger (Meles meles), from the Late Villafranchian site of Fuente Nueva 3 (Orce. Granada, SE Iberian Peninsula). C. R. Palevol 10, 609–615. doi: 10.1016/j.crpv.2011.06.001
Madurell-Malapeira, J., Minwer-Barakat, R., Alba, D. M., Garcés, M., Gómez, M., Aurell-Garrido, J., et al. (2010). The Vallparadís section (Terrassa, Iberian Peninsula) and the latest Villafranchian faunas of Europe. Quat. Sci. Rev. 29, 3972–3982. doi: 10.1016/j.quascirev.2010.09.020
Madurell-Malapeira, J., Ros-Montoya, S., Espigares, M. P., Alba, D. M., and Aurell-Garrido, J. (2014). Villafranchian large mammals from the Iberian Peninsula: paleogeography, paleoecology and dispersal events. J. Iber. Geol. 40, 167–178.
Maldonado-Garrido, E., Piñero, P., and Agustí, J. (2017). A catalogue of the vertebrate fossil record from the Guadix-Baza Basin (SE Spain). Spanish J. Paleontol. 32, 207–236. doi: 10.7203/sjp.32.1.17040
Marean, C. W., and Ehrhardt, C. L. (1995). Paleoanthropological and paleoecological implications of the taphonomy of a sabertooth’s den. J. Hum. Evol. 29, 515–547. doi: 10.1006/jhev.1995.1074
Martin, R. A. (2014). A critique of vole clocks. Quat. Sci.Rev. 94, 1–6. doi: 10.1016/j.quascirev.2014.03.004
Martínez, K., Garcia, J., Burjachs, F., Yll, R., and Carbonell, E. (2015). Early human occupation of Iberia: the chronological and palaeoclimatic inferences from Vallparadís (Barcelona, Spain). Quat. Sci. Rev. 85, 136–146. doi: 10.1016/j.quascirev.2013.12.002
Martínez-García, B., Ordiales, A., Pérez, A., Muñoz, A., Luzón, A., and Murelaga, X. (2017). Palaeoenvironmental evolution of the Holocene record of the Añavieja lake (Soria, NE Iberian Peninsula) based on the study of the ostracod assemblages. Geogaceta 62, 43–46.
Martínez-García, B., Suárez-Hernando, O., Hernández, J. M., Suárez-Bilbao, A., and Murelaga, X. (2015). Palaeoenvironmental analysis based on alluvial ostracod assemblages of the Cenicero section (lower Miocene, NW Ebro Basin). Estud. Geol. 71:e024. doi: 10.3989/egeol.41729.320
Martínez-Navarro, B. (1991). Revisión sistemática y estudio cuantitativo de la fauna de macromamíferos del yacimiento de Venta Micena (Orce, Granada). Ph. D. dissertation. Spain: Univ. Autónoma de Barcelona, 264.
Martínez-Navarro, B. (2002). The skull of Orce: Parietal bones or frontal bones? J. Hum. Evol. 43, 265–270. doi: 10.1006/jhev.2002.0566
Martínez-Navarro, B. (2004). “Hippos, pigs, bovids, sabertoothed tigers, monkeys and hominids: dispersals during Late Pliocene and Early Pleistocene times through the Levantine Corridor,” in Human Paleoecology in the Levantine Corridor, eds N. Goren-Inbar and J. D. Speth (Oxford: Oxbow Books), 37–51. doi: 10.2307/j.ctvh1dtct.9
Martínez-Navarro, B. (2010). “Early Pleistocene faunas of Eurasia and hominid dispersals,” in The First Hominin Colonization of Eurasia, Contributions from the Second Stony Brook Human Evolution Symposium and Workshop, eds J. G. Fleagle, J. J. Shea, F. E. Grine, A. L. Baden, and R. E. Leakey (New York, NY: Springer), 207–224. doi: 10.1007/978-90-481-9036-2_13
Martínez-Navarro, B. (2018). Oldowan scavengers vs. Acheulian hunters: What does the faunal record say? Glob. J. Arch. Anthropol. 6:555679.
Martínez-Navarro, B., Bartolini Lucenti, S., Palmqvist, P., Ros-Montoya, S., Madurell-Malapeira, J., and Espigares, M. P. (2021). A new species of dog from the Early Pleistocene site of Venta Micena (Orce, Baza Basin, Spain). C. R. Palevol 20, 297–314.
Martínez-Navarro, B., Belmaker, M., and Bar-Yosef, B. (2012). The Bovid assemblage (Bovidae, Mammalia) from the Early Pleistocene site of ’Ubeidiya, Israel: biochronological and paleoecological implications for the fossil and lithic bearing strata. Quat. Int. 267, 78–97. doi: 10.1016/j.quaint.2012.02.041
Martínez-Navarro, B., Belmaker, M., and Bar-Yosef, O. (2009). The large carnivores from ’Ubeidiya (early Pleistocene, Israel): biochronological and biogeographical implications. J. Hum. Evol. 56, 514–524. doi: 10.1016/j.jhevol.2009.02.004
Martínez-Navarro, B., Karoui-Yaakoub, N., Oms, O., Amri, L., López-García, J. M., Zeräi, K., et al. (2014b). The early Middle Pleistocene archeopaleontological site of Wadi Sarrat (Tunisia) and the earliest record of Bos primigenius. Quat. Sci. Rev. 90, 37s–46.
Martínez-Navarro, B., Espigares, M. P., Pastó, I., Ros-Montoya, S., and Palmqvist, P. (2014a). Early Homo fossil records of Europe. Encyclop. Glob. Archaeol. 2014, 2561–2570. doi: 10.1007/978-1-4419-0465-2_646
Martínez-Navarro, B., Madurell-Malapeira, J., Ros-Montoya, S., Espigares, M. P., Medin, T., Hortolà, P., et al. (2015). The Epivillafranchian and the arrival of pigs into Europe. Qua. Int. 389, 131–138. doi: 10.1016/j.quaint.2015.09.039
Martínez-Navarro, B., and Palmqvist, P. (1995). Presence of the African machairodont Megantereon whitei (Broom, 1937) (Felidae, Carnivora, Mammalia) in the Lower Pleistocene site of Venta Micena (Orce, Granada, Spain), with some considerations on the origin, evolution and dispersal of the genus. J. Archaeolog. Sci. 22, 569–582. doi: 10.1006/jasc.1994.0054
Martínez-Navarro, B., and Palmqvist, P. (1996). Presence of the African saber-toothed felid Megantereon whitei (Broom, 1937) (Mammalia, Carnivora, Machairodontinae) in Apollonia-1 (Migdonia Basin, Macedonia, Greece). J. Archeol. Sci. 23, 869–872. doi: 10.1006/jasc.1996.0081
Martínez-Navarro, B., Palmqvist, P., Madurell-Malapeira, J., Ros-Montoya, S., Espigares, M. P., Torregrosa, V., et al. (2010). “La fauna de grandes mamíferos de Fuente Nueva-3 y Barranco León-5: Estado de la cuestión,” in Ocupaciones humanas en el Pleistoceno inferior y medio de la Cuenca de Guadix-Baza, eds I. Toro, B. Martínez-Navarro, and J. Agustí (Sevilla: Junta de Andalucía, Consejería de Cultura, 197–236.
Martínez-Navarro, B., and Rook, L. (2003). Gradual evolution in the African hunting dog lineage. Systematic implications. C. R. Palevol 2, 695–702. doi: 10.1016/j.crpv.2003.06.002
Martínez-Navarro, B., Ros-Montoya, S., Espigares, M. P., Madurell-Malapeira, J., and Palmqvist, P. (2018). Los mamíferos del Plioceno y Pleistoceno de la Península Ibérica. Revista PH 94, 206–249. doi: 10.33349/2018.0.4203
Martínez-Navarro, B., Ros-Montoya, S., Espigares, M. P., and Palmqvist, P. (2011). Presence of Asian origin Bovini, Hemibos sp. aff. H. gracilis and Bison sp., at the Early Pleistocene site of Venta Micena (Orce, Spain). Quat. Int. 243, 54–60. doi: 10.1016/j.quaint.2011.05.016
Martínez-Navarro, B., Turq, A., Agustí, J., and Oms, O. (1997). Fuente Nueva-3 (Orce, Granada, Spain) and the first human occupation of Europe. J. Hum. Evol. 33, 611–620. doi: 10.1006/jhev.1997.0158
Martín-González, J. A., Rodríguez-Gómez, G., and Palmqvist, P. (2019). Survival profiles from linear models versus Weibull models: Estimating stable and stationary population structures for Pleistocene large mammals. J. Archaeol. Sci. Rep. 25, 370–386. doi: 10.1016/j.jasrep.2019.03.031
Martín-Serra, A., Figueirido, B., and Palmqvist, P. (2017). Non-decoupled morphological evolution of the fore- and hindlimb of sabretooth predators. J. Anat. 231, 532–542. doi: 10.1111/joa.12654
McHenry, C., Wroe, S., Clausen, P. D., Moreno, K., and Cunningham, E. (2007). Supermodeled sabercat, predatory behavior in Smilodon fatalis revealed by high-resolution 3D computer simulation. Proc. Nat. Acad. Sci. USA 104, 16010–16015. doi: 10.1073/pnas.0706086104
Meachen-Samuels, J. A. (2012). Morphological convergence of the prey-killing arsenal of sabertooth predators. Paleobiology 38, 715–728.
Meachen-Samuels, J. A., and Van Valkenburgh, B. (2010). Radiographs reveal exceptional forelimb strength in the sabertooth cat, Smilodon fatalis. PLoS One 5:e11412. doi: 10.1371/journal.pone.0011412
Medin, T., Martínez-Navarro, B., Rivals, F., Madurell-Malapeira, J., Ros-Montoya, S., Espigares, M. P., et al. (2017). Late Villafranchian Ursus etruscus and other large carnivorans from the Orce sites (Guadix-Baza basin, Andalusia, Southern Spain): Taxonomy, biochronology paleobiolgy and ecogeographical context. Quat. Int. 431, 20–41. doi: 10.1016/j.quaint.2015.10.053
Mendoza, M., Janis, C. M., and Palmqvist, P. (2005). Ecological patterns in the trophic-size structure of large mammal communities: a ‘taxon-free’ characterization. Evol. Ecol. Res. 7, 505–530.
Menotti-Raymond, M., and O’Brien, S. J. (1993). Dating the genetic bottleneck of the African cheetah. Proc. Nat. Acad. Sci. USA 90, 3172–3176. doi: 10.1073/pnas.90.8.3172
Merola, M. (2011). A reassessment of homozygosity and the case for inbreeding depression in the cheetah, Acinonyx jubatus: Implications for conservation. Conserv. Biol. 8, 961–971. doi: 10.1046/j.1523-1739.1994.08040961.x
Mgeladze, A., Lordkipanidze, D., Moncel, M.-H., Despriee, J., Chagelishvili, R., Nioradze, M., et al. (2011). Hominin occupations at the Dmanisi site, Georgia, Southern Caucasus: Raw materials and technical behaviours of Europe’s first hominins. J. Hum. Evol. 60, 571–596. doi: 10.1016/j.jhevol.2010.10.008
Michel, V., Shen, C. C., Woodhead, J., Hu, H. M., Wu, C. C., and Moullé, P. E. (2017). New dating evidence of the early presence of hominins in Southern Europe. Sci. Rep. 7:10074. doi: 10.1038/s41598-017-10178-4
Minwer-Barakat, R., García-Alix, A., Suárez, E. M., Freudenthal, M., and Viseras, C. (2012). Micromammal biostratigraphy of the Upper Miocene to lowest Pleistocene continental deposits of the Guadix basin, southern Spain. Lethaia 45, 594–614. doi: 10.1111/j.1502-3931.2012.00324.x
Moncel, M. H., Santagata, C., Pereira, A., Nomade, S., Voinchet, P., Bahain, J.-J., et al. (2020). The origin of early Acheulean expansion in Europe 700 ka ago: new findings at Notarchirico (Italy). Sci. Rep. 10:13802. doi: 10.1038/s41598-020-68617-8
Mosquera, M., Ollé, A., and Rodríguez, X. P. (2013). From Atapuerca to Europe: Tracing the earliest peopling of Europe. Quat. Int. 295, 130–137. doi: 10.1016/j.quaint.2012.01.031
Moullé, P.-E., Echassoux, A., and Martínez-Navarro, B. (2004). Ammotragus europaeus: une nouvelle espéce de Caprini (Bovidae, Mammalia) du Pléistocéne inférieur á la grotte du Vallonnet (France). C. R. Palevol 3, 663–673. doi: 10.1016/j.crpv.2004.09.001
Moullé, P.-E., Lacombat, F., and Echassoux, A. (2006). Contribution of the large mammals of Vallonnet cave (Roquebrune-Cap-Martin, Alpes-Maritimes, France) to the knowledge of biochronological frame of the second half of the Lower Pleistocene in Europe. L’Anthropologie 110, 837–849. doi: 10.1016/j.anthro.2006.10.006
Moyà-Solà, S. (1987). Los bóvidos (Artiodactyla, Mammalia) del yacimiento del Pleistoceno inferior de Venta Micena (Orce, Granada, España). Paleontologia i Evolució, Mem. Espec. 1, 181–236.
Muttoni, G., Scardia, G., and Kent, D. V. (2010). Human migration into Europe during the late Early Pleistocene climate transition. Palaeogeogr. Palaeoclimatol. Palaeoecol. 296, 79–93. doi: 10.1016/j.palaeo.2010.06.016
Muttoni, G., Scardia, G., and Kent, D. V. (2013). A critique of evidence for human occupation of Europe older than the Jaramillo subchron (~1 Ma): Comment on ‘The oldest human fossil in Europe from Orce (Spain)’ by Toro-Moyano, et al. (2013). J. Hum. Evol. 65, 746–749. doi: 10.1016/j.jhevol.2013.08.005
Muttoni, G., Scardia, G., Kent, D. V., Morsiani, E., Tremolada, F., Cremaschi, M., et al. (2011). First dated human occupation of Italy at ~0.85 Ma during the late Early Pleistocene climate transition. Earth Planet. Sci. Let. 307, 241–252. doi: 10.1016/j.epsl.2011.05.025
Muttoni, G., Sirakov, N., Guadelli, J.-L., Kent, D. V., Scardia, G., Monesi, E., et al. (2017). An early Brunhes (<0.78 Ma) age for the Lower Paleolithic tool-bearing Kozarnika cave sediments. Bulgaria. Quat. Sci. Rev. 178, 1–13. doi: 10.1016/j.quascirev.2017.10.034
O’Connell, J. F., Hawkes, K., and Blurton Jones, N. (1988). Hadza scavenging: implications for Plio/Pleistocene hominid subsistence. Curr. Anthrop. 29, 356–363. doi: 10.1086/203648
Oms, O., Parés, J. M., Martínez-Navarro, B., Agustí, J., Toro, I., Martínez-Fernández, G., et al. (2000). Early human occupation of Western Europe: paleomagnetic dates for two Paleolithic sites in Spain. Proc. Nat. Acad. Sci. USA 97, 10666–10670. doi: 10.1073/pnas.180319797
Oota, H., Pakendorf, B., Weiss, G., Von Haeseler, A., Pookajorn, S., Settheetham-Ishida, W., et al. (2005). Recent origin and cultural reversion of a hunter-gatherer group. PLoS Biol. 3:e71. doi: 10.1371/journal.pbio.0030071
Outram, A., and Rowley-Conwy, P. (1988). Meat and marrow utility indices for horse (Equus). J. Archaeol. Sci. 25, 839–849. doi: 10.1006/jasc.1997.0229
Packer, C., Pusey, A. E., and Rowley, H. (1991). Case study of a population bottleneck: lions of the Ngorongoro Crater. Conserv. Biol. 5, 219–230. doi: 10.1111/j.1523-1739.1991.tb00127.x
Palmqvist, P. (2002). On the presence of Megantereon whitei at the South Turkwel hominid site, northern Kenya. J. Paleontol. 76, 928–930. doi: 10.1666/0022-3360(2002)076<0928:otpomw>2.0.co;2
Palmqvist, P., and Arribas, A. (2001). Taphonomic decoding of the paleobiological information locked in a lower Pleistocene assemblage of large mammals. Paleobiology 27, 512–530. doi: 10.1666/0094-8373(2001)027<0512:tdotpi>2.0.co;2
Palmqvist, P., Arribas, A., and Martínez-Navarro, B. (1999). Ecomorphological study of large canids from the lower Pleistocene of southeastern Spain. Lethaia 32, 75–88. doi: 10.1111/j.1502-3931.1999.tb00583.x
Palmqvist, P., Duval, M., Diéguez, A., Ros-Montoya, S., and Espigares, M. P. (2016). On the fallacy of using orthogenetic models of rectilinear change in arvicolid teeth for estimating the age of the first human settlements in Western Europe. Hist. Biol. 28, 734–752. doi: 10.1080/08912963.2015.1025390
Palmqvist, P., González-Donoso, J. M., and De Renzi, M. (2014). Rectilinear evolution in arvicoline rodents and numerical dating of Iberian Early Pleistocene sites. Quat. Sci. Rev. 98, 100–109. doi: 10.1016/j.quascirev.2014.06.002
Palmqvist, P., Gröcke, D. R., Arribas, A., and Fariña, R. (2003). Paleoecological reconstruction of a lower Pleistocene large mammals community using biogeochemical (δ13C, δ15N, δ18O, Sr: Zn) and ecomorphological approaches. Paleobiology 29, 205–229. doi: 10.1666/0094-8373(2003)029<0205:proalp>2.0.co;2
Palmqvist, P., Martínez-Navarro, B., and Arribas, A. (1996). Prey selection by terrestrial carnivores in a lower Pleistocene paleocommunity. Paleobiology 22, 514–534. doi: 10.1017/s009483730001650x
Palmqvist, P., Martínez-Navarro, B., Pérez-Claros, J. A., Torregrosa, V., Figueirido, B., Jiménez-Arenas, J. M., et al. (2011). The giant hyena Pachycrocuta brevirostris: modelling the bone-cracking behavior of an extinct carnivore. Quat. Int. 243, 61–79. doi: 10.1016/j.quaint.2010.12.035
Palmqvist, P., Martínez-Navarro, B., Toro, I., Espigares, M. P., Ros-Montoya, S., Torregrosa, V., et al. (2005). Réévaluation de la présence humaine au Pléistocène inférieur dans le Sud de l’Espagne. L’Anthropologie 109, 411–450.
Palmqvist, P., Torregrosa, V., Pérez-Claros, J. A., Martínez-Navarro, B., and Turner, A. (2007). A re-evaluation of the diversity of Megantereon (Mammalia, Carnivora, Machairodontinae) and the problem of species identification in extinct carnivores. J. Vert. Paleontol. 27, 160–175. doi: 10.1671/0272-4634(2007)27[160:arotdo]2.0.co;2
Palmqvist, P., Pérez-Claros, J. A., Gröcke, D. R., and Janis, C. M. (2008a). Tracing the ecophysiology of ungulates and predator-prey relationships in an early Pleistocene large mammal community. Palaeogeogr., Palaeoclimatol., Palaeoecol. 266, 95–111. doi: 10.1016/j.palaeo.2008.03.015
Palmqvist, P., Pérez-Claros, J. A., Janis, C. M., Figueirido, B., Torregrosa, V., and Gröcke, D. R. (2008b). Biogeochemical and ecomorphological inferences on prey selection and resource partitioning among mammalian carnivores in an early Pleistocene community. Palaios 23, 724–737. doi: 10.2110/palo.2007.p07-073r
Palmqvist, P., Rodríguez-Gómez, G., Figueirido, B., García-Aguilar, J. M., and Pérez-Claros, J. A. (2022a). On the ecological scenario of the first hominin dispersal out of Africa. L’Anthropologie 126:102998. doi: 10.1016/j.anthro.2022.102998
Palmqvist, P., Espigares, M. P., Perez−Claros, J. A., Figueirido, B., Guerra−Merchan, A., Ros−Montoya, S., et al. (2022b). Déjà vu: a reappraisal of the taphonomy of quarry VM4 of the Early Pleistocene site of Venta Micena (Baza Basin, SE Spain). Sci. Rep. 12:705. doi: 10.1038/s41598-021-04725-3
Palmqvist, P., and Vizcaíno, S. F. (2003). Ecological and reproductive constraints of body size in the gigantic Argentavis magnificens (Aves, Theratornithidae) from the Miocene of Argentina. Ameghiniana 40, 379–385.
Palombo, M. R. (2010). A scenario of human dispersal in the northwestern Mediterranean throughout the Early to Middle Pleistocene. Quat. Int. 223-224, 179–194. doi: 10.1016/j.quaint.2009.11.016
Parés, J. M., Álvarez, C., Sier, M., Moreno, D., Duval, M., Woodhead, J. D., et al. (2018). Chronology of the cave interior sediments at Gran Dolina archaeological site, Atapuerca (Spain). Quat. Sci. Rev. 186, 1–16. doi: 10.1016/j.quascirev.2018.02.004
Parés, J. M., Sahnouni, M., van der Made, J., Pérez-González, A., Harichane, Z., Derradji, A., et al. (2014). Early human settlements in Northern Africa: paleomagnetic evidence from the Ain Hanech Formation (northeastern Algeria). Quat. Sci. Rev. 99, 203–209. doi: 10.1016/j.quascirev.2014.06.020
Parkinson, J. (2018). Revisiting the hunting-versus-scavenging debate at FLK Zinj: a GIS spatial analysis of bone surface modifications produced by hominins and carnivores in the FLK 22 assemblage, Olduvai Gorge, Tanzania. Palaeogeogr. Palaeoclimatol. Palaeoecol. 511, 29–51. doi: 10.1016/j.palaeo.2018.06.044
Peretto, C. (2006). The first peopling of southern Europe: the Italian case. C. R. Palevol 5, 283–290. doi: 10.1016/j.crpv.2005.11.006
Pickering, T. R., and Bunn, H. (2007). The endurance running hypothesis and hunting and scavenging in savanna-woodlands. J. Hum.Evol. 53, 434–438. doi: 10.1016/j.jhevol.2007.01.012
Plummer, T. (2004). Flaked stones and old bones: biological and cultural evolution at the dawn of technology. Yearbook Phys. Anthropol. 47, 118–164. doi: 10.1002/ajpa.20157
Pobiner, B. L. (2015). New actualistic data on the ecology and energetics of hominin scavenging opportunities. J. Hum. Evol. 80, 1–15. doi: 10.1016/j.jhevol.2014.06.020
Pobiner, B. L. (2020). The zooarchaeology and paleoecology of early hominin scavenging. Evol. Anthropol. 2020:21824. doi: 10.1002/evan.21824
Pons-Moyà, J. (1987). Los carnívoros (Mammalia) de Venta Micena (Granada, España). Paleontologia i Evolució Mem. Espec. 1, 10–127.
Potts, R. (1991). Why the Oldowan? Plio-Pleistocene toolmaking and the transport of resources. J. Anthropol. Res. 47, 153–176. doi: 10.1086/jar.47.2.3630323
Power, R. J., and Shem-Compion, R. X. (2009). Lion predation on elephants in the Savuti, Chobe National Park, Botswana. A. Zool. 44, 36–44. doi: 10.3377/004.044.0104
Prado-Nóvoa, O., Mateos, A., Zorrilla-Revilla, G., Vidal-Cordasco, M., and Rodríguez, J. (2017). Efficiency of gathering and its archaeological implications for a European Early Paleolithic population. J. Anthropol. Archaeol. 45, 131–141. doi: 10.1016/j.jaa.2016.12.002
Raichlen, D. A., Wood, B. M., Gordon, A. D., Mabullad, A. Z. P., Marlowe, F. W., and Pontzerf, H. (2014). Evidence of Leìvy walk foraging patterns in human hunter-gatherers. Proc. Nat. Acad. Sci. USA 111, 728–733. doi: 10.1073/pnas.1318616111
Ripple, W. J., and Van Valkenburgh, B. (2010). Linking top-down forces to the Pleistocene megafaunal extinctions. BioScience 60, 516–526. doi: 10.1525/bio.2010.60.7.7
Rodríguez, J., Blain, H.-A., Mateos, A., Martín-González, J. A., Cuenca-Bescós, G., and Rodríguez-Gómez, G. (2014). Ungulate carrying capacity in Pleistocene Mediterranean ecosystems: Evidence from the Atapuerca sites. Palaeogeogr., Palaeoclimatol., Palaeoecol. 393, 122–134. doi: 10.1016/j.palaeo.2013.11.011
Rodríguez, J., Burjachs, F., Cuenca-Bescós, G., García, N., Made, J., Pérez-González, A., et al. (2011). One million years of cultural evolution in a stable environment at Atapuerca (Burgos, Spain). Quat. Sci. Rev. 30, 1396–1412. doi: 10.1016/j.quascirev.2010.02.021
Rodríguez, J., Martín-González, J. A., Goikoetxea, I., Rodríguez-Gómez, G., and Mateos, A. (2013). Mammalian paleobiogeography and the distribution of Homo in early Pleistocene Europe. Quat.Int. 295, 48–58. doi: 10.1016/j.quaint.2011.09.023
Rodríguez-Gómez, G., Cassini, G. H., Palmqvist, P., Bargo, M. S., Toledo, N., Martín-González, J. A., et al. (2020). Testing the hypothesis of an impoverished predator guild in the Early Miocene ecosystems of Patagonia: An analysis of meat availability and competition intensity among carnivores. Palaeogeogr. Palaeoclimatol. Palaeoecol. 554:109805. doi: 10.1016/j.palaeo.2020.109805
Rodríguez-Gómez, G., Rodríguez, J., Martín-González, J. A., Goikoetxea, I., and Mateos, A. (2013). Modeling trophic resource availability for the first human settlers of Europe: the case of Atapuerca-TD6. J. Hum. Evol. 64, 645–657. doi: 10.1016/j.jhevol.2013.02.007
Rodríguez-Gómez, G., Mateos, A., Martín-González, J. A., Blasco, R., Rosell, J., and Rodríguez, J. (2014a). Discontinuity of human presence at Atapuerca during the early Middle Pleistocene: A matter of ecological competition? PLoS One 9:e101938. doi: 10.1371/journal.pone.0101938
Rodríguez-Gómez, G., Martín-González, J. A., Goikoetxea, I., Mateos, A., and Rodríguez, J. (2014b). “A new mathematical approach to model trophic dynamics of mammalian palaeocommunities. The case of Atapuerca-TD6,” in Mathematics of Planet Earth, 15th Annual Conference of the International Association for Mathematical Geosciences, eds E. Pardo-Igúzquiza, C. Guardiola-Albert, J. Heredia, L. Moreno-Merino, J. J. Durán, and J. A. Vargas-Guzmán (Madrid: Lectures Notes in Earth System Sciences - Springer), 739–745. doi: 10.1007/978-3-642-32408-6_160
Rodríguez-Gómez, G., Palmqvist, P., Rodríguez, J., Mateos, A., Martín-González, J. A., Espigares, M. P., et al. (2016a). On the ecological context of the earliest human settlements in Europe: Resource availability and competition intensity in the carnivore guild of Barranco León-D and Fuente Nueva-3 (Orce, Baza Basin, SE Spain). Quat. Sci. Rev. 134, 69–83. doi: 10.1016/j.quascirev.2016.05.018
Rodríguez-Gómez, G., Mateos, A., Martín-González, J. A., and Rodríguez, J. (2016b). Measuring intraguild competition from faunal assemblages to compare environmental conditions among paleocommunities. Quat. Int. 413, 55–68. doi: 10.1016/j.quaint.2015.11.087
Rodríguez-Gómez, G., Palmqvist, P., Martínez-Navarro, B., Martín-González, J. A., and Bermúdez de Castro, J. M. (2022). Mean body size estimation in large mammals and the computation of biomass in past ecosystems: An application to the Pleistocene sites of Orce and Sierra de Atapuerca (Spain). C.R. Palevol 21, 1–27. doi: 10.5852/cr-palevol2022v21a10
Rodríguez-Gómez, G., Palmqvist, P., Ros-Montoya, S., Espigares, M. P., and Martínez-Navarro, B. (2017a). Resource availability and competition intensity in the carnivore guild of the Early Pleistocene site of Venta Micena (Orce, Baza Basin, SE Spain). Quat. Sci. Rev. 164, 154–167. doi: 10.1016/j.quascirev.2017.04.006
Rodríguez-Gómez, G., Rodríguez, J., Martín-González, J. A., and Mateos, A. (2017b). Carnivores and humans during the Early and Middle Pleistocene at Sierra de Atapuerca. Quat. Int. 433, 402–414. doi: 10.1016/j.quaint.2015.12.082
Rodríguez-Gómez, G., Rodríguez, J., Martín-González, J. A., and Mateos, A. (2017c). Evaluating the impact of Homo-carnivore competition in European human settlements during the Early to Middle Pleistocene. Quat. Res. 88, 129–151. doi: 10.1017/qua.2017.20
Roebroeks, W. (2001). Hominid behaviour and the earliest occupation of Europe: an exploration. J. Hum. Evol. 41, 437–461. doi: 10.1006/jhev.2001.0499
Roebroeks, W., Gaudzinski-Windheuser, S., Baales, M., and Kahlke, R.-D. (2018). Uneven data quality and the earliest occupation of Europe—the case of Untermassfeld (Germany). J. Paleolit. Archaeol. 1, 5–31. doi: 10.1007/s41982-017-0003-5
Roebroeks, W., and van Kolfschoten, T. (1994). The earliest occupation of Europe: a short chronology. Antiquity 68, 489–503. doi: 10.1017/s0003598x00047001
Ros-Montoya, S., Bartolini-Lucenti, S., Espigares, M. P., Palmqvist, P., and Martínez-Navarro, B. (2021). First review of Lyncodontini material (Mustelidae, Carnivora, Mammalia) from the lower Pleistocene archaeo-palaeontological sites of Orce (southeastern Spain). Riv. Ital. Paleontol. Stratigr. 127, 33–47.
Ros-Montoya, S., Martínez-Navarro, B., Espigares, M. P., Guerra-Merchán, A., García-Aguilar, J. M., Piñero, P., et al. (2017). A new Ruscinian site in Europe: Baza-1 (Baza Basin, Andalusia, Spain). C. R. Palevol 16, 746–761. doi: 10.1016/j.crpv.2017.05.005
Ruggiero, R. G. (1991). Opportunistic predation on elephant calves. Af. J. Ecol. 29, 86–89. doi: 10.1098/rsbl.2013.0518
Ruxton, G. D., and Wilkinson, D. M. (2012). Endurance running and its relevance to scavenging by early hominins. Evolution 67, 861–867. doi: 10.1111/j.1558-5646.2012.01815.x
Saarinen, J., Oksanen, O., Žliobaitë, I., Fortelius, M., DeMiguel, D., Azanza, B., et al. (2021). Pliocene to Middle Pleistocene climate history in the Guadix-Baza Basin, and the environmental conditions of early Homo dispersal in Europe. Quat. Sci. Rev. 268:107132. doi: 10.1016/j.quascirev.2021.107132
Sahnouni, M., Parés, J. M., Duval, M., van der Made, J., Harichane, Z., Pérez-González, A., et al. (2018a). “Early Homo adaptive and behavioral patterns in North Africa: perspectives from Ain Hanech and Tighennif (formerly Ternifine) sites in Northern Algeria,” in 15th Congress of PanAfrican Archaeological Association for Prehistory and Related Studies, (PanAf), 5–6.
Sahnouni, M., Parés, J. M., Duval, M., Cáceres, I., Harichane, Z., van der Made, J., et al. (2018b). 1.9-million- and 2.4-million-year-old artifacts and stone tool–cutmarked bones from Ain Boucherit, Algeria. Science 362, 1297–1130. doi: 10.1126/science.aau0008
Salesa, M., Antón, M., Turner, A., and Morales, J. (2010). Functional anatomy of the forelimb in Promegantereon ogygia (Felidae, Machairodontinae, Smilodontini) from the Late Miocene of Spain and the origins of the sabre-toothed felid model. J. Anat. 216, 381–396. doi: 10.1111/j.1469-7580.2009.01178.x
Sánchez-Bandera, C., Oms, O., Blain, H.-A., Lozano-Fernández, I., Bisbal-Chinesta, J. F., Agustí, J., et al. (2020). New stratigraphically constrained palaeoenvironmental reconstructions for the first human settlement in Western Europe: the Early Pleistocene herpetofaunal assemblages from Barranco Leon and Fuente Nueva 3 (Granada, SE Spain). Quat. Sci. Rev. 243, 106466. doi: 10.1016/j.quascirev.2020.106466
Schaller, G. B. (1968). Hunting behaviour of the cheetah in the Serengeti National Park, Tanzania. Afr. J. Ecol. 6, 95–100. doi: 10.1111/j.1365-2028.1968.tb00906.x
Schaller, G. B. (1972). The Serengeti Lion. A Study of Predator-Prey Relations. Chicago: The University of Chicago Press, 480.
Selvaggio, M. M. (1998). Evidence for a three-stage sequence of hominid and carnivore involvement with long bones at FLK Zinjanthropus, Olduvai Gorge, Tanzania. J. Archaeol. Sci. 25, 191–202. doi: 10.1006/jasc.1997.0281
Silberbauer, G. B. (1981). Hunter and Habitat in the Central Kalahari Desert. Cambridge, MA: Cambridge University Press, 388.
Sirakov, N., Guadelli, J. L., Ivanova, S., Sirakova, S., Boudadi-Maligne, M., Dimitrova, I., et al. (2010). An ancient continuous human presence in the Balkans and the beginnings of human settlement in western Eurasia: a Lower Pleistocene example of the Lower Paleolithic levels in Kozarnika cave (North-western Bulgaria). Quat. Int. 223, 94–106. doi: 10.1016/j.quaint.2010.02.023
Skinner, J. D., and Smithers, R. H. N. (1990). The Mammals of the Southern African Subregion. Cambridge, MA: Cambridge University Press, 814.
Slater, G. J., and Van Valkenburgh, B. (2008). Long in the tooth: evolution of sabertooth cat cranial shape. Paleobiology 34, 403–419. doi: 10.1666/07061.1
Soria, J. M., Fernández, J., and Viseras, C. (1999). Late Miocene stratigraphy and palaeogeographic evolution of the intramontane Guadix Basin (Central Betic Cordillera, Spain): implications for an Atlantic-Mediterranean connection. Palaeogeogr., Palaeoclimatol. Palaeoecol. 151, 255–266. doi: 10.1016/s0031-0182(99)00019-x
Spiering, P., Szykman Gunther, M., Somers, M. J., Wildt, D. E., Walters, M., Wilson, A. S., et al. (2011). Inbreeding, heterozygosity and fitness in a reintroduced population of endangered African wild dogs (Lycaon pictus). Cons. Gen. 12, 401–412. doi: 10.1007/s10592-010-0147-z
Stanford, C. B. (1999). Meat eating and the origins of human behaviour. Princeton: Princeton University Press, 263.
Stevens, A., Morissette, J., Woodburn, J. C., and Bennett, F. J. (1977). The inbreeding coefficients of the Hadza. Ann. Hum. Biol. 4, 219–223. doi: 10.1080/03014467700007102
Swisher, C. C., Curtis, G. H., Jacob, T., Getty, A. G., and Suprijo, A. (1994). Age of the earliest known hominids in Java. Indonesia. Sci. 263, 1118–1121. doi: 10.1126/science.8108729
Tappen, M., Adler, D. S., Ferring, C. R., Gabunia, M., Vekua, A., and Swisher, C. C. (2002). Akhalkalaki: the taphonomy of an Early Pleistocene locality in the Republic of Georgia. J. Archaeol. Sci. 29, 1367–1391. doi: 10.1006/jasc.2001.0797
Thompson, J. C., Carvalho, S., Marean, C. W., and Alemseged, Z. (2019). Origins of the human predatory pattern: the transition to large animal exploitation by early hominins. Curr. Anthropol. 60, 1–23. doi: 10.1086/701477
Titton, S., Barsky, D., Bargalló, A., Vergès, J. M., Guardiola, M., García-Solano, J., et al. (2018). Active percussion tools from the Oldowan site of Barranco León (Orce, Andalusia, Spain): The fundamental role of pounding activities in hominin lifeways. J. Archaeol. Sci. 96, 131–147. doi: 10.1016/j.jas.2018.06.004
Titton, S., Oms, O., Barsky, D., Bargalló, A., Serrano-Ramos, A., García-Solano, J., et al. (2021). Oldowan stone knapping and percussive activities on a raw material reservoir deposit 1.4 million years ago at Barranco León (Orce, Spain). Archaeol. Anthropol. Sci. 13:108. doi: 10.1007/s12520-021-01353-w
Tixier, J., Roe, D., Turq, A., Gibert, J., Martínez-Navarro, B., Arribas, A., et al. (1995). Presence d’industries lithiques dans le Pléistocène inférieur de la région d’Orce (Grenade, Espagne): etat de la question. C. R. Acad. Sci. Paris 321, 71–78.
Toro-Moyano, I., Barsky, D., Cauche, D., Celiberti, V., Grégoire, S., Lebegue, F., et al. (2011). The archaic Stone tool industry from Barranco León and Fuente Nueva 3 (Orce, Spain): Evidence of the earliest hominin presence in southern Europe. Quat. Int. 243, 80–91. doi: 10.1016/j.quaint.2010.12.011
Toro-Moyano, I., Martínez-Navarro, B., Agustí, J., Souday, C., Bermúdez, de Castro, J. M., et al. (2013). The oldest human fossil in Europe, from Orce (Spain). J. Hum. Evol. 65, 1–9. doi: 10.1016/j.jhevol.2013.01.012
Torres Pérez-Hidalgo, T. (1992). “Los restos de oso del yacimiento de Venta Micena (Orce, Granada) y el material de Ursus etruscus G. Cuvier del Villafranquiense europeo,” in Proyecto Orce-Cueva Victoria (1988-1992): Presencia humana en el Pleistoceno inferior de Granada y Murcia. Museo de Prehistoria Josep Gibert-Ayuntamiento de Orce, ed. J. Gibert (Orce), 87–106.
Traill, L. W., Bradshaw, C. J., and Brook, B. W. (2007). Minimum viable population size: a meta-analysis of 30 years of published estimates. Biol. Conserv. 139, 159–166. doi: 10.1016/j.biocon.2007.06.011
Treves, A., and Palmqvist, P. (2007). “Reconstructing hominin interactions with mammalian carnivores (6.0-1.8 Ma),” in Primate Anti-Predator Strategies. Developments in Primatology Series: Progress and Prospects, eds S. L. Gursky and K. A. I. Nekaris (Springer, NY), 355–381. doi: 10.1007/978-0-387-34810-0_17
Trinkaus, E. (2018). An abundance of developmental anomalies and abnormalities in Pleistocene people. Proc. Nat. Acad. Sci. USA 115, 11941–11946. doi: 10.1073/pnas.1814989115
Turner, A. (1992). Large carnivores and earliest European hominids: changing determinants of resource availability during the lower and middle Pleistocene. J. Hum. Evol. 22, 109–126. doi: 10.1016/0047-2484(92)90033-6
Turq, A., Martínez-Navarro, B., Palmqvist, P., Arribas, A., Agustí, J., and Rodríguez-Vidal, J. (1996). Le Plio-Pléistocène de la région d’Orce, province de Grenada, Espagne: Bilan et perspectivas de Recherche. Paléo 8, 161–204. doi: 10.3406/pal.1996.911
Uauy, R., Hoffman, D. R., Peirano, P., Birch, D. G., and Birch, E. E. (2001). Essential fatty acids in visual and brain development. Lipids 36, 885–895. doi: 10.1007/s11745-001-0798-1
van den Bergh, G. D., Aziz, F., Kurniawan, I., Morwood, M. J., Lentfer, C. J., and Suyone Setiawan, R. (2009). “Environmental reconstruction of the Middle Pleistocene archaeological and palaeontological site of Mata Menge, Central Flores, Indonesia,” in Pleistocene geology, palaeontology and archaeology of the Soa Basin, Central Flores, Indonesia. Special publication no. 36. Pusat Survei Geologi, Bandung, eds F. Aziz, M. J. Morwood, and G. D. van den Bergh 58–94.
van den Bergh, G. D., Kaifu, Y., Kurniawan, I., Kono, R. T., Brumm, A., Setiyabudi, E., et al. (2016). Homo floresiensis-like fossils from the early Middle Pleistocene of Flores. Nature 534, 245–248. doi: 10.1038/nature17999
Van Valkenburgh, B. (2001). “Predation in sabre-tooth cats,” in Paleobiology II, eds D. E. G. Briggs and P. R. Crowther (Oxford: Blackwell), 420–423. doi: 10.1002/9780470999295.ch101
Van Valkenburgh, B. (2007). Déjà vu: the evolution of feeding morphologies in the Carnivora. Integr.Comp. Biol. 47, 147–163. doi: 10.1093/icb/icm016
Van Valkenburgh, B. (2009). Costs of carnivory: tooth fracture in Pleistocene and Recent carnivorans. Biol. J. Linnean Soc. 96, 68–81. doi: 10.1111/j.1095-8312.2008.01108.x
Van Valkenburgh, B., Hayward, M. W., Ripple, W. J., Meloro, C., and Roth, V. L. (2016). The impact of large terrestrial carnivores on Pleistocene ecosystems. Proc. Nat. Acad. Sci. USA 113, 862–867. doi: 10.1073/pnas.1502554112
Van Valkenburgh, B., and Hertel, F. (1993). Tough times at La Brea: tooth breakage in large carnivores of the late Pleistocene. Science 261, 456–459. doi: 10.1126/science.261.5120.456
Van Valkenburgh, B., Teaford, M. F., and Walker, A. (1990). Molar microwear and diet in large carnivores: inferences concerning diet in the sabretooth cat, Smilodon fatalis. J. Zool. 222, 319–340. doi: 10.1111/j.1469-7998.1990.tb05680.x
Vekua, A. (1986). The lower pleistocene mammalian fauna of Akhalkalaki (southern Georgia, USSR). Palaeontogr. Ital. 74, 63–96.
Vera, J. A., Fernández, J., López-Garrido, A. C., and Rodríguez-Fernández, J. (1984). Geología y estratigrafía de los materiales plioceno-pleistocenos de del sector Orce - Venta Micena (Prov. Granada). Paleontología i Evolució 18, 3–11.
Vicente-Gabarda, M., Martínez-Valle, R., Guillem-Calatayud, M., Garay-Martí, P., Pueyo, E., and Casabó, J. (2016). The Lower Palaeolithic site Alto de las Picarazas (Andilla-Chelva, Valencia). Quat. Int. 393, 83–94. doi: 10.1016/j.quaint.2015.04.049
Vigilant, L., Stoneking, M., Harpending, H., Hawkes, K., and Wilson, A. C. (1991). African populations and the evolution of Human mitochondrial DNA. Science 253, 1503–1507. doi: 10.1126/science.1840702
Vilà, C., Urios, V., and Castroviejo, J. (1993). Tooth losses and anomalies in the wolf (Canis lupus). Canadian J. Zool. 71, 968–971. doi: 10.1139/z93-127
Viseras, C., Soria, J. M., Durán, J. J., Pla, S., Garrido, G., García-García, F., et al. (2006). A large-mammal site in a meandering fluvial context (Fonelas P-1. Late Pliocene, Guadix basin, Spain): sedimentological keys for the paleoenvironmental reconstruction. Palaeogeogr., Palaeoclimatol., Palaeoecol. 242, 139–168. doi: 10.1016/j.palaeo.2006.05.013
Voinchet, P., Despriée, J., Tissoux, H., Falguères, C., Bahain, J. J., Gageonnet, R., et al. (2010). ESR chronology of alluvial deposits and first human settlements of the Middle Loire Basin (Region Centre, France). Quatern. Geochronol. 5, 381–384. doi: 10.1016/j.quageo.2009.03.005
Wanpo, H., Ciochon, R., Yumin, G., Larick, R., Qiren, F., Schwarcz, H., et al. (1995). Early Homo and associated artefacts from Asia. Nature 378, 275–278. doi: 10.1038/378275a0
Wayne, R. K., Modi, W. S., and O’Brien, S. J. (1986). Morphological variability and asymmetry in the cheetah (Acinonyx jubatus), a genetically uniform species. Evolution 40, 78–85. doi: 10.1111/j.1558-5646.1986.tb05719.x
Welker, F., Ramos-Madrigal, J., Gutenbrunner, P., Mackie, T. S., Jersie-Christensen, R. R., Chiva, C., et al. (2020). A Pleistocene hominin proteome from Homo antecessor. Nature 580, 235–238.
Woodburn, J. (1968). “An introduction to Hazda ecology,” in Man the hunter, eds R. B. Lee and I. DeVore (Chicago: Aldine Publishing Co), 49–55.
Keywords: early Homo, Western Europe, subsistence strategies, Barranco León, Fuente Nueva 3, population size
Citation: Palmqvist P, Rodríguez-Gómez G, Bermúdez de Castro JM, García-Aguilar JM, Espigares MP, Figueirido B, Ros-Montoya S, Granados A, Serrano FJ, Martínez-Navarro B and Guerra-Merchán A (2022) Insights on the Early Pleistocene Hominin Population of the Guadix-Baza Depression (SE Spain) and a Review on the Ecology of the First Peopling of Europe. Front. Ecol. Evol. 10:881651. doi: 10.3389/fevo.2022.881651
Received: 22 February 2022; Accepted: 14 March 2022;
Published: 21 April 2022.
Edited by:
Marta Arzarello, University of Ferrara, ItalyReviewed by:
Juan Rofes, University of the Philippines Diliman, PhilippinesCopyright © 2022 Palmqvist, Rodríguez-Gómez, Bermúdez de Castro, García-Aguilar, Espigares, Figueirido, Ros-Montoya, Granados, Serrano, Martínez-Navarro and Guerra-Merchán. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paul Palmqvist, cHBiQHVtYS5lcw==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.