 Enrico Lunghi
Enrico Lunghi Helena Bilandžija
Helena Bilandžija
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Ecol. Evol. , 20 May 2022
Sec. Behavioral and Evolutionary Ecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.874123
This article is part of the Research Topic Adaptations to Subterranean Environments View all 14 articles
An extraordinary longevity has been observed in some cave species, and this raised the hypothesis that a longer lifespan may be considered one of the characteristic traits of these animals. However, only a few cave species have been studied thus far, and a firm conclusion remains to be drawn. Here we review the available knowledge on the longevity of subterranean species, point out the limitations of previous studies, and provide suggestions for future studies to answer important questions regarding the longevity in cave animals, its adaptive value and the related promoting factors. We also argue that studying the longevity in cave animals will contribute to the field of aging, especially to understanding the evolution of this phenomenon.
With their unique ecological setting, subterranean environments are theaters of evolutionary processes that lead to fascinating adaptive traits in the species able to colonize them (Mammola, 2019). All subsurface habitats are characterized by a common feature, the absence of light (Culver and Pipan, 2014, 2019). Light can only penetrate and affect the area surrounding their connections to the surface (Lunghi et al., 2015; Culver and Pipan, 2019), limiting the effects of seasonality and the circadian cycle in deeper zones, a condition that contributes to highly stable microclimate characterized by high humidity and constant temperatures (Biswas, 2009; Lunghi et al., 2015). Without sunlight, photosynthetic processes are absent (Culver and Pipan, 2019), what leads to a strong reduction of the available organic matter, and therefore the subterranean food web is largely dependent on inputs of allochthonous organic material from the surface (Schneider et al., 2011; Barzaghi et al., 2017). This scarcity of food resources results in reduced abundance of species at all levels of the food web (Venarsky et al., 2014; Culver and Pipan, 2019; Manenti et al., 2020).
Colonization of subterranean environments by aboveground species is accompanied by numerous changes in their phenotypic traits (Hervant et al., 2001; Bilandžija et al., 2020; Lunghi and Zhao, 2020). The hallmarks of adaptation to subterranean environments are loss of eyes and pigmentation, which are so common in subterranean taxa that they are used as diagnostic traits to assess the degree of adaptation to these environments (Howarth and Moldovan, 2018; but see Lunghi et al., 2014). Some of the other conspicuous adaptive traits in subterranean species include elongation of body appendages, increase in fat stores, overexpression of mechanosensory and chemosensory systems (Jeffery, 2009; Bilandžija et al., 2012; Gonzalez et al., 2017). Although most obvious, morphological adaptations often represent the final chapter in a story that began with the behavioral and physiological changes that accompanied the early stages of cave colonization (Culver et al., 1995; Pigliucci et al., 2006; Bilandžija et al., 2020).
In this review we explore a poorly known aspect of subterranean species: their longevity. Among subterranean taxa, individuals with exceptional longevity have occasionally been reported (e.g., Voituron et al., 2011; Puljas et al., 2014), raising the idea that the dark passages within the earth harbor animals with extended lifespans. Over the years, most studies have aimed to estimate the age and lifespan of cave species (e.g., Trajano, 1991; Voituron et al., 2011; Puljas et al., 2014), but very few in a comparative framework that included a surface relative (Poulson, 1963). Ultimately, it has yet to be proven whether higher longevity is indeed a characterizing trait of subterranean animals, and eventually whether such trait is adaptive. Here, we review what is known about the longevity of subterranean species and provide recommendations for future studies to experimentally test whether these animals show higher longevity compared to their surface relatives and what the potential promoting factors might be.
The limited information on the lifespan of cave species mostly comes from the few species that have been successfully reared in laboratory facilities (Poulson, 1963; Miaud and Guillaume, 2005; Hinaux et al., 2011; Voituron et al., 2011; Bichuette and Trajano, 2021). Animals kept under controlled conditions enable tracking their growth and development of specific growth rate curves can provide age estimates for wild populations (Brunkow and Collins, 1996; Gallo and Jeffery, 2012). However, controlled conditions may differ from those found in natural environments and may alter growth rates of individuals (Trajano, 1997; Simon et al., 2017). Luckily, growth rates can be estimated also in the wild for species that are marked (e.g., Lunghi and Bruni, 2018; Lunghi et al., 2019) and recaptured over time (Taddei Ruggiero, 2001; Venarsky et al., 2012; Balázs et al., 2020; Lunghi et al., 2022), or when individuals show reliable age marks (Simon et al., 2017; Riddle et al., 2018).
One of the most iconic cave species is the olm, Proteus anguinus, an aquatic salamander distributed in the Dinaric karst in the South-Eastern Europe (Sket, 1997; Gorički et al., 2017). Because it is extremely difficult to study Proteus in the wild (Buzzacott et al., 2009; Balázs et al., 2020), most of the available knowledge comes from captive observations (Juberthie et al., 1996; Ipsen and Knolle, 2017; Aljančič, 2019). This neotenic salamander has a slow development, becoming sexually mature after 14 years and reproducing every 12.5 for at least 50/60 years (Voituron et al., 2011; Ipsen and Knolle, 2017). The oldest known individual has an age between 48 and 58 years (Voituron et al., 2011). Given its slow growth rate and low reproductive activity, the average lifespan of the olm should be around 70 years, although some individuals can live more than 100 years (Bulog et al., 2000; Voituron et al., 2011). In comparison, the closest aboveground relatives are from the North American genus Necturus, and the longevity of Necturus maculosus has been estimated at 34 years (Petranka, 1998). Interestingly, the olm’s small size, metabolic rate comparable to other salamanders, and lack of increased antioxidant activity are in stark contrast to the predictions of aging theory, which foresee higher lifespan in larger species with low metabolic rate and high antioxidant activity (Blanco and Sherman, 2005; Voituron et al., 2011).
Cavefish are within the best-studied subterranean animals, yet only a few studies on their lifespan exist. For example, the Mexican tetra, Astyanax mexicanus, is probably one of the widest used model species in cave biology (Wilkens, 1988; Rétaux and Casane, 2013; Keene et al., 2015; Jeffery, 2020), but reliable information on the potential lifespan of this species is missing. We know from captive breeding that both surface and cave forms of A. mexicanus can reach the age of 15 (Riddle et al., 2018). There is a divergence in growth rate and age distribution between several populations of cave and surface forms (Simon et al., 2017). Specifically, the range of estimated ages of fish was greater in cave populations (2–8 years) than in surface populations (2–5 years), and there is variability in age distribution even between cave populations (Simon et al., 2017). In another comparative study, Poulson (1963) used the scale and otolith to age five amblyopsid fishes, two surface species, Chologaster cornuta and Forbesichthys agassizi, and three subsurface, Typhlichthys subterraneus, Amblyopsis spelaea, and A. rosae. The results showed a higher lifespan in cave fishes, where the age of older individuals ranged from 3 to 7 years, up to three times more than for the surface fishes (Poulson, 1963). A similar age for A. rosae (about 4–5 years) has been estimated in another study (Brown and Johnson, 2001). However, more up to date techniques indicate a potential lifespan of 20–30 years for these cavefish (Poulson, 2001). For the Brazilian Ancistrus cryptophthalmus, an initial estimate determined a lifespan of 8–10 years (Trajano and Bichuette, 2007), while a subsequent study extended it to more than 15 years and provided evidence that the longest-lived individuals can live up to 20–25 years (Secutti and Trajano, 2009). An estimate of lifespan based on growth rate exists also for some other cavefish from the tropics: Pimelodella kronei (10–15 years; Trajano, 1991), Trichomycterus itacarambiensis (7 years; Trajano, 1997), Caecobarbus geertsii (9–15 years; Heuts, 1952; Proudlove and Romero, 2001), and Ituglanis passensis (>10; Secutti and Trajano, 2021). No comparable information on closely related surface species exists for these Brazilian cavefish; however, in comparison with other cyprinids (data from AnAge database; Tacutu et al., 2018) these species are among the longer living ones.
Studies in cave invertebrates also yielded extraordinary estimates of their lifespan. An analysis of the growth line formation in the shells of the cave bivalve Congeria kusceri found that the average lifespan was about 30 years. Importantly, authors also showed that growth lines form annually despite animals residing in stable underground waters (Puljas et al., 2014). The oldest individual in this study, a 14.3 mm long male, had an impressive lifespan of 53 years (Puljas et al., 2014). However, other studies have found much larger individuals, so the lifespan of this and possibly other Congeria species is likely much higher (Morton, 1969; Bilandžija et al., 2013). The average lifespan of other dreissenid species ranges between 2 and 5 years (Puljas et al., 2014), implying that cave bivalves have evolved lifespans 10 times longer than their surface relatives. As with Proteus, this is contrary to predictions which suggest that larger animals have longer lifespans (Blanco and Sherman, 2005), as Congeria is one of the smaller dreissenids. However, its slow growth rate and low investment in reproduction (Morton and Puljas, 2013) are consistent with disposable soma theory of aging (Kirkwood, 1977) which suggests a tradeoff between the investment in the reproduction and somatic maintenance; the latter leading to lifespan increase. Similarly, analysis of growth rate of the brachiopod Neocrania anomala from marine caves estimated that its lifespan may exceed 40 years (Taddei Ruggiero, 2001). The lifespan of N. anomala is about three times higher than Waltonia inconspicua and Terebratalia transversa, brachiopods that can reach a maximum age of 15 and 13 years, respectively, (Paine, 1969; Rickwood, 1977). Age estimation of an extinct brachiopod, Gigantoproductus okensis, indicates that this species could have lived up to 20 years (Angiolini et al., 2012), which is half of the lifespan estimated for the cave N. anomala (Taddei Ruggiero, 2001).
A capture-mark-recapture study conducted over 5 years provided important information on the lifespan of the crayfish Orconectes australis (Venarsky et al., 2012). Using growth rate data, the authors were able to re-evaluate the lifespan for this species, which was incorrectly estimated to be 100 years, and found out that the species can live over 22 years (Venarsky et al., 2012). In addition, these authors compiled a list of crayfish species for which lifespan estimates exist (see Table 2 in Venarsky et al., 2012), including two additional cave species: O. inermis (9–10 years), and Procambarus erythrops (>16 years). Of the 10 surface cambarids included in the list, one has an estimated lifespan of 13 years, while the others ≤7 years. Similar results have been obtained for other cave crustaceans. Based on the distribution of body size, Pacioglu et al. (2020) suggested that the lifespan of Gammarus balcanicus might exceed that of epigean gammarids, which have lifespan estimates of about 1 year. In another study, Magniez (1975) estimated that the cave Stenasellus virei can reach an age of 12–15, a lifespan up to 20 times longer than epigean congeneric species. Furthermore, a 40-year study performed by Carpenter (2021) showed that Bahalana geracei has probably the longest known lifespan (ranging from 24.5 to 35 years) among isopods. Capture mark recapture studies showed that the cave beetle Laemostenus schreibersi can reach the remarkable age of >6.5 (Rusdea, 1994), almost doubling the maximum lifespan observed for epigean ground beetles (Lövei and Sunderland, 1996).
All information on cave species lifespan collected in this review are summarized in Table 1.
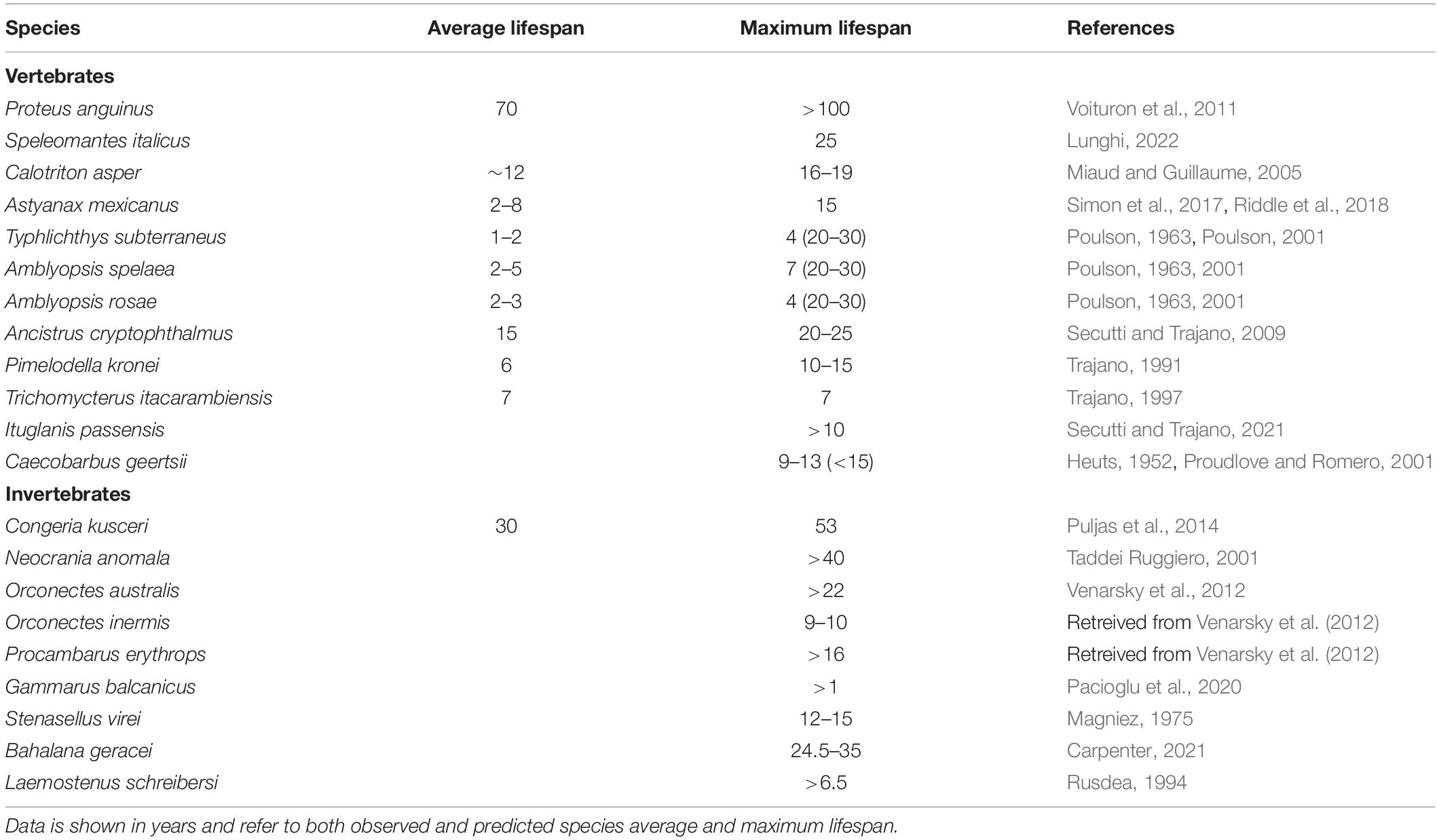
Table 1. Summary of the lifespan data for cave species collected in this review.
Although bats are not strictly bound to subterranean habitats, as they use caves primarily for sheltering purposes and do not exhibit classic cave-related adaptations (Howarth and Moldovan, 2018; Sadier et al., 2020), it is worth mentioning this animal group in the context of longevity, as they are the longest-lived mammals relative to their mass (Podlutsky et al., 2005; Wilkinson and Adams, 2019). Interestingly, cave use predicts the evolution of longevity in several bat lineages (Wilkinson and South, 2002; Salmon et al., 2009).
The information assembled here suggests that longevity can indeed be considered a characteristic of cave animals. However, additional systematic studies encompassing a broader range of animal groups in a comparative framework which includes surface and cave relatives are needed to fortify this conclusion. The main remaining questions are: (1) is longevity adaptive for subterranean lifestyle or did it evolve as a physiological consequence of other adaptive traits, (2) what components of the subterranean environment contribute to the evolution of life extension in resident animals, and (3) what are the molecular and developmental mechanisms that lead to convergent evolution of longevity in cave organisms. Several hypotheses can be made regarding the links of aging with other cave adaptive traits and the characteristics of subterranean environments that may be promoting it. For example, adaptive traits in caves such as slower growth, lower metabolic rate and lower investment in reproduction, have been associated with increased lifespan (Flatt and Schmidt, 2009). Further, many environmental features of caves such as limited food resources (Aspiras et al., 2015), lower extrinsic mortality (lack of predators; Plath and Schlupp, 2008), hypoxia (Boggs and Gross, 2021; van der Weele and Jeffery, 2022) and lack of UV irradiation (Körner et al., 2006), are consistent with known ecological predictors of longevity (Speakman and Selman, 2011; Flament et al., 2013; Omotoso et al., 2021). These nonexclusive hypotheses can be tested using model systems that have closely related species or populations in caves and on the surface, have a reliable method for estimating their age, and can survive in the laboratory to be used for experimental perturbations.
Although important to understand the life history characteristics of subterranean species (e.g., sexual maturity, reproductive potential; Poulson, 1963; Lunghi et al., 2018; Lunghi, 2022), growth rate curves may not be the best method to estimate species ages. First, species generally do not grow indefinitely and their growth rate can only be estimated during a particular phase of their life cycle (Venarsky et al., 2012; Lunghi, 2022). A clear example of this is the aforementioned crayfish Orconectes australis, whose lifespan has been reduced by a factor of four using better-tuned methods (Venarsky et al., 2012). Another limitation to using this method stems from the ecological differences between cave and surface populations. For example, low availability of nutrients in subterranean environments (Huntsman et al., 2011; Culver and Pipan, 2019; Pacheco et al., 2020) strongly influences species growth (Simon et al., 2017). Without rearing cave and surface relatives under the same conditions, it is difficult to distinguish intrinsic from extrinsic effects on lifespan.
The use of molecular biomarkers of aging (e.g., SA-β-Gal, 4-HNE, or lipofuscin staining; Nelson et al., 2012; Dimri et al., 1995) may overcome some of these limitations. In addition to allowing reliable assessment of lifespan differences between subterranean and surface species, the use of markers for relevant physiological and molecular processes will enable following the basic processes underlying aging as well as the assessment of variability in aging on population or species levels (Robins et al., 2017).
Studies on cave animals may also provide important insights into the biology of aging, particularly in the areas of evolution and genetics of aging. One of the greatest challenges in the field of aging is to uncover the genes and processes that cause lifespan differences among species (Partridge and Gems, 2006). Because of the known direction of evolution -from surface to cave- and the ability to compare ancestral form with derived form, cave dwellers provide a rich source of information about mechanisms that lead to slowing rates of aging in nature. Knowledge of these mechanisms could be of great benefit and have implications for human welfare. Because of well-defined environmental characteristics and numerous convergently evolved physiological and life-history adaptations, the study of aging in cave dwellers can shed light on how longevity covaries and co-evolves with other organismal and environmental traits.
Aging is a complex phenotype, so it is important to incorporate research on nontraditional and atypical species and integrate approaches from multiple biological disciplines (Cohen, 2018). How and why senescence evolves in natural populations and how to maintain health in old age is still a major issue in contemporary biology. We propose that studies of aging in cave-adapted animals can provide new and original insights into how nature has solved the problem of extending lifespan.
EL drafted the manuscript. Both authors conceived the study and critically revised the manuscript.
This study was financed within the Tenure Track Pilot Programme of the Croatian Science Foundation and the Ecole Polytechnique Fédérale de Lausanne and the Project TTP-2018-07-9675 EvoDark, with funds of the Croatian-Swiss Research Programme.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aljančič, G. (2019). History of research on Proteus anguinus Laurenti 1768 in Slovenia. Folia Biol. Geol. 60, 39–69. doi: 10.3986/fbg0050
Angiolini, L., Stephenson, M., Leng, M. J., Jadoul, F., Millward, D., Aldridge, A., et al. (2012). Heterogeneity, cyclicity and diagenesis in a Mississippian brachiopod shell of palaeoequatorial Britain. Terra Nova 24, 16–26. doi: 10.1111/j.1365-3121.2011.01032.x
Aspiras, A. C., Rohner, N., Martineau, B., Borowsky, R. L., and Tabin, C. J. (2015). Melanocortin 4 receptor mutations contribute to the adaptation of cavefish to nutrient-poor conditions. Proc. Natl. Acad. Sci. U.S.A. 112, 9668–9673. doi: 10.1073/pnas.1510802112
Balázs, G., Lewarne, B., and Herczeg, G. (2020). Extreme site fidelity of the olm (Proteus anguinus) revealed by a long-term capture–mark–recapture study. J. Zool. 311, 99–105. doi: 10.1111/jzo.12760
Barzaghi, B., Ficetola, G. F., Pennati, R., and Manenti, R. (2017). Biphasic predators provide biomass subsidies in small freshwater habitats: a case study of spring and cave pools. Freshw. Biol. 62, 1637–1644. doi: 10.1111/fwb.12975
Bichuette, M. E., and Trajano, E. (2021). Monitoring Brazilian cavefish: Ecology and conservation of four threatened catfish of genus ituglanis (siluriformes: Trichomycteridae) from central brazil. Diversity 13:91. doi: 10.3390/d13020091
Bilandžija, H., Ćetković, H., and Jeffery, W. R. (2012). Evolution of albinism in cave planthoppers by a convergent defect in the first step of melanin biosynthesis. Evol. Dev. 14, 196–203. doi: 10.1111/j.1525-142X.2012.00535.x
Bilandžija, H., Morton, B., Podnar, M., and Ćetković, H. (2013). Evolutionary history of relict Congeria (Bivalvia: Dreissenidae): unearthing the subterranean biodiversity of the Dinaric Karst. Front. Zool. 10:5. doi: 10.1186/1742-9994-10-5
Bilandžija, H., Hollifield, B., Steck, M., Meng, G., Ng, M., Koch, A. D., et al. (2020). Phenotypic plasticity as a mechanism of cave colonization and adaptation. eLife 9:e51830. doi: 10.7554/eLife.51830
Biswas, J. (2009). Kotumsar Cave biodiversity: a review of cavernicoles and their troglobiotic traits. Biodivers. Conserv. 19, 275–289. doi: 10.1007/s10531-009-9710-7
Blanco, M. A., and Sherman, P. W. (2005). Maximum longevities of chemically protected and non-protected fishes, reptiles, and amphibians support evolutionary hypotheses of aging. Mech. Ageing Dev. 126, 794–803. doi: 10.1016/j.mad.2005.02.006
Boggs, T., and Gross, J. (2021). Reduced oxygen as an environmental pressure in the evolution of the blind Mexican cavefish. Diversity 13:26. doi: 10.3390/d13010026
Brown, J. Z., and Johnson, J. E. (2001). Population biology and growth of Ozark cavefish in Logan Cave National Wildlife Refuge, Arkansas. Environ. Biol. Fishes 62, 161–169. doi: 10.1023/a:1011860821932
Brunkow, P. E., and Collins, J. P. (1996). Effects of individual variation in size on growth and development of larval salamanders. Ecology 77, 1483–1492. doi: 10.2307/2265545
Bulog, B., Bizjakmali, L., Kos, M., Mihajl, K., Prelovsek, P.-M., and Aljanaid, G. (2000). Biology and functional morphology of Proteus anguinus (Amphibia. Caudata). Acta Biol. Slov. 43, 85–102.
Buzzacott, P. L., Zeigler, E., Denoble, P., and Vann, R. (2009). American cave diving fatalities 1969-2007. Int. J. Aquat. Res. Educ. 3, 162–177.
Carpenter, J. H. (2021). Forty-year natural history study of Bahalana geracei Carpenter, 1981, an anchialine cave-dwelling isopod (Crustacea, Isopoda, Cirolanidae) from San Salvador Island, Bahamas: reproduction, growth, longevity, and population structure. Subterr. Biol. 37, 105–156. doi: 10.3897/subtbiol.37.60653
Cohen, A. A. (2018). Aging across the tree of life: The importance of a comparative perspective for the use of animal models in aging. Biochim. Biophys. Acta Mol. Basis Dis. 1864, 2680–2689. doi: 10.1016/j.bbadis.2017.05.028
Culver, D. C., Kane, T. C., and Fong, D. W. (1995). Adaptation and Natural Selection in Caves. The Evolution of Gammarus Minus. Cambridge, MA: Harvard University Press.
Culver, D. C., and Pipan, T. (2014). Shallow Subterranean Habitats: Ecology, Evolution, and Conservation. Oxford: Oxford University Press.
Culver, D. C., and Pipan, T. (2019). The Biology of Caves and Other Subterranean Habitats. Oxford: Oxford University Press.
Dimri, G. P., Leet, X., Basile, G., Acosta, M., Scorrt, G., Roskelley, C., et al. (1995). A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc. Natl. Acad. Sci. U.S.A. 92, 9363–9367.
Flament, F., Bazin, R., Laquieze, S., Rubert, V., Simonpietri, E., and Piot, B. (2013). Effect of the sun on visible clinical signs of aging in Caucasian skin. Clin. Cosmet. Investig. Dermatol. 6, 221–232. doi: 10.2147/CCID.S44686
Flatt, T., and Schmidt, P. S. (2009). Integrating evolutionary and molecular genetics of aging. Biochim. Biophys. Acta 1790, 951–962. doi: 10.1016/j.bbagen.2009.07.010
Gallo, N. D., and Jeffery, W. R. (2012). Evolution of space dependent growth in the Teleost Astyanax mexicanus. PLoS One 7:e41443. doi: 10.1371/journal.pone.0041443
Gonzalez, B. C., Worsaae, K., Fontaneto, D., and Martínez, A. (2017). Anophthalmia and elongation of body appendages in cave scale worms (Annelida: Aphroditiformia). Zool. Scr. 47, 106–121. doi: 10.1111/zsc.12258
Gorički, S., Stanković, D., Snoj, A., Kuntner, M., Jeffery, W. R., Trontelj, P., et al. (2017). Environmental DNA in subterranean biology: range extension and taxonomic implications for Proteus. Sci. Rep. 7:45054. doi: 10.1038/srep45054
Hervant, F., Mathieu, J., and Durand, J. P. (2001). Behavioural, physiological and metabolic responses to long-term starvation and refeeding in a blind cave-dwelling (Proteus anguinus) and a surface-dwelling (Euproctus asper) salamander. J. Exp. Biol. 204, 269–281. doi: 10.1242/jeb.204.2.269
Heuts, M. J. (1952). Ecology, variation and adaptation of the blind African cave fish Caecobarbus geertsi Blgr. Ann. Soc. Roy. Zool. Belg. 82, 155–230.
Hinaux, H., Pottin, K., Chalhoub, H., Père, S., Elipot, Y., Legendre, L., et al. (2011). A developmental staging table for Astyanax mexicanus surface fish and Pachón cavefish. Zebrafish 8, 155–165. doi: 10.1089/zeb.2011.0713
Howarth, F. G., and Moldovan, O. T. (2018). “The ecological classification of cave animals and their adaptations,” in Cave Ecology, eds Moldovan L. Kováč and S. Halse (Berlin: Springer), 41–67. doi: 10.1007/978-3-319-98852-8_4
Huntsman, B. M., Venarsky, M. P., Benstead, J. P., and Huryn, A. D. (2011). Effects of organic matter availability on the life history and production of a top vertebrate predator (Plethodontidae: Gyrinophilus palleucus) in two cave streams. Freshw. Biol. 56, 1746–1760.
Ipsen, A., and Knolle, F. (2017). The olm of Hermann’s Cave, Harz Mountains, Germany – eggs laid after more than 80 years. Nat. Slov. 19, 51–52.
Jeffery, W. R. (2009). Regressive evolution in Astyanax cavefish. Annu. Rev. Genet. 43, 25–47. doi: 10.1146/annurev-genet-102108-134216
Jeffery, W. R. (2020). Astyanax surface and cave fish morphs. EvoDevo 11:14. doi: 10.1186/s13227-020-00159-6
Juberthie, C., Durand, J., and Dupuy, M. (1996). La reproduction des Protées (Proteus anguinus): bilan de 35 ans d’élevage dans les grottes-laboratoires de Moulis et d’Aulignac. Mém. Biospéol. 23, 53–56.
Keene, A., Yoshizawa, M., and McGaugh, S. E. (2015). Biology and Evolution of the Mexican Cavefish. Cambridge, MA: Academic Press.
Körner, K. E., Schlupp, I., Plath, M., and Loew, E. R. (2006). Spectral sensitivity of mollies: comparing surface- and cave-dwelling Atlantic mollies, Poecilia mexicana. J. Fish Biol. 69, 54–65. doi: 10.1111/j.1095-8649.2006.01056.x
Lövei, G. L., and Sunderland, K. D. (1996). Ecology and behavior of ground beetles (Coleoptera: Carabidae). Annu. Rev. Entomol. 41, 231–256. doi: 10.1146/annurev.en.41.010196.001311
Lunghi, E. (2022). Doubling the lifespan of European plethodontid salamanders. Ecology 103:e03581. doi: 10.1002/ecy.3581
Lunghi, E., and Bruni, G. (2018). Long-term reliability of Visual Implant Elastomers in the Italian cave salamander (Hydromantes italicus). Salamandra 54, 283–286.
Lunghi, E., Corti, C., Biaggini, M., Merilli, S., Manenti, R., Zhao, Y., et al. (2022). Capture-mark-recapture data on the strictly protected Speleomantes italicus. Ecology doi: 10.1002/ecy.3641 [Epub ahead of print].
Lunghi, E., Corti, C., Manenti, R., Barzaghi, B., Buschettu, S., Canedoli, C., et al. (2018). Comparative reproductive biology of European cave salamanders (genus Hydromantes): nesting selection and multiple annual breeding. Salamandra 54, 101–108.
Lunghi, E., Manenti, R., and Ficetola, G. F. (2014). Do cave features affect underground habitat exploitation by non-troglobite species? Acta Oecol. 55, 29–35.
Lunghi, E., Manenti, R., and Ficetola, G. F. (2015). Seasonal variation in microhabitat of salamanders: environmental variation or shift of habitat selection? PeerJ 3:e1122.
Lunghi, E., Romeo, D., Mulargia, M., Cogoni, R., Manenti, R., Corti, C., et al. (2019). On the stability of the dorsal pattern of European cave salamanders (genus Hydromantes). Herpetozoa 32, 249–253. doi: 10.3897/herpetozoa.32.e39030
Lunghi, E., and Zhao, Y. (2020). Do Chinese cavefish show intraspecific variability in morphological traits? Ecol. Evol. 10, 7723–7730. doi: 10.1002/ece3.6495
Magniez, G. (1975). Observations sur la biologie de Stenasellus virei (Crustacea Isopoda Asellota) des eaux souterraines. Int. J. Speleol. 7, 79–228. doi: 10.5038/1827-806x.7.1.8
Mammola, S. (2019). Finding answers in the dark: caves as models in ecology fifty years after Poulson and White. Ecography 42, 1331–1351. doi: 10.1111/ecog.03905
Manenti, R., Melotto, A., Guillaume, O., Ficetola, G. F., and Lunghi, E. (2020). Switching from mesopredator to apex predator: how do responses vary in amphibians adapted to cave living? Behav. Ecol. Sociobiol. 74:126.
Miaud, C., and Guillaume, O. (2005). Variation in age, body size and growth among surface and cave-dwelling populations of the Pyrenean newt, Euproctus asper (Amphibia; Urodela). Herpetologica 61, 241–249. doi: 10.1655/04-29.1
Morton, B. (1969). Studies on the biology of Dreissena polymorpha Pall. (III). Population dynamics. Proc. Malacol. Soc. Lond. 38, 471–482.
Morton, B., and Puljas, S. (2013). Life-history strategy, with ctenidial and pallial larval brooding, of the troglodytic ‘living fossil’ Congeria kusceri (Bivalvia: Dreissenidae) from the subterranean Dinaric Alpine karst of Croatia. Biol. J. Linn. Soc. 108, 294–314. doi: 10.1111/j.1095-8312.2012.02020.x
Nelson, P. T., Alafuzoff, I., Bigio, E. H., Bouras, C., Braak, H., Cairns, N. J., et al. (2012). Correlation of alzheimer disease neuropathologic changes with cognitive status: A review of the literature. J. Neuropathol. Exp. Neurol. 71, 362–381. doi: 10.1097/NEN.0b013e31825018f7
Omotoso, A. O., Reyer, H., Oster, M., Ponsuksili, S., Trakooljul, N., Muráni, E., et al. (2021). Jejunal transcriptomic profiling of two layer strains throughout the entire production period. Sci. Rep. 11:20086. doi: 10.1038/s41598-021-99566-5
Pacheco, G. S. M., Souza Silva, M., Cano, E., and Ferreira, R. L. (2020). The role of microhabitats in structuring cave invertebrate communities in Guatemala. Int. J. Speleol. 49, 161–169. doi: 10.5038/1827-806x.49.2.2333
Pacioglu, O., Strungaru, S.-A., Ianovici, N., Filimon, M. N., Sinitean, A., Iacob, G., et al. (2020). Ecophysiological and life-history adaptations of Gammarus balcanicus (Schäferna, 1922) in a sinking-cave stream from Western Carpathians (Romania). Zoology 139:125754. doi: 10.1016/j.zool.2020.125754
Paine, R. T. (1969). Growth and size distribution of the brachiopod Terebratalia transversa Sowerby. Pac. Sci. 23, 337–343.
Partridge, L., and Gems, D. (2006). Beyond the evolutionary theory of ageing, from functional genomics to evo-gero. Trends Ecol. Evol. 21, 334–340. doi: 10.1016/j.tree.2006.02.008
Petranka, J. W. (1998). Salamanders of the United States and Canada. Washington, DC: Smithsonian press.
Pigliucci, M., Murren, C. J., and Schlichting, C. D. (2006). Phenotypic plasticity and evolution by genetic assimilation. J. Exp. Biol. 209, 2362–2367. doi: 10.1242/jeb.02070
Plath, M., and Schlupp, I. (2008). Parallel evolution leads to reduced shoaling behavior in two cave dwelling populations of Atlantic mollies (Poecilia mexicana, Poeciliidae, Teleostei). Environ. Biol. Fish. 82, 289–297. doi: 10.1007/s10641-007-9291-9
Podlutsky, A., Khritankov, A. M., Ovodov, N. D., and Austad, S. N. (2005). A new field record for bat longevity. J. Gerontol. A Biol. Sci. Med. Sci. 60, 1366–1368. doi: 10.1093/gerona/60.11.1366
Poulson, T. L. (1963). Cave adaptation in Amblyopsid fishes. Amer. Midl. Nat. 70, 257–290. doi: 10.2307/2423056
Poulson, T. L. (2001). Adaptations of cave fishes with some comparisons to deep-sea fishes. Environ. Biol. Fishes 62, 345–364. doi: 10.1007/978-94-015-9795-1_28
Proudlove, G. S., and Romero, A. (2001). Threatened fishes of the world: Caecobarbus geertsii Boulenger, 1921 (Cyprinidae). Environ. Biol. Fishes 62, 238–238. doi: 10.1023/a:1011803519256
Puljas, S., Peharda, M., Morton, B., Štambuk Giljanović, N., and Jurić, I. (2014). Growth and longevity of the “living fossil” Congeria kusceri (Bivalvia: Dreissenidae) from the subterranean Dinaric karst of Croatia. Malacologia 57, 353–364.
Rétaux, S., and Casane, D. (2013). Evolution of eye development in the darkness of caves: adaptation, drift, or both? EvoDevo 4:26. doi: 10.1186/2041-9139-4-26
Rickwood, A. E. (1977). Age, growth and shape of the intertidal brachiopod Waltonia inconspicua Sowerby, from New Zealand. Am. Zool. 17, 63–73. doi: 10.1093/icb/17.1.63
Riddle, M. R., Aspiras, A. C., Gaudenz, K., Peuß, R., Sung, J. Y., Martineau, B., et al. (2018). Insulin resistance in cavefish as an adaptation to a nutrient-limited environment. Nature 555, 647–651. doi: 10.1038/nature26136
Robins, C., McRae, A. F., Powell, J. E., Wiener, H. W., Aslibekyan, S., Kennedy, E. M., et al. (2017). Testing two evolutionary theories of human aging with DNA methylation data. Genetics 207, 1547–1560. doi: 10.1534/genetics.117.300217
Rusdea, E. (1994). “Population dynamics of Laemostenus schreibersi (Carabidae) in a cave in Carinthia (Austria),” in Carabid Beetles: Ecology and Evolution (Series Entomologica 51), eds K. Desender et al. (Dordrecht: Springer Science), 219–225.
Sadier, A., Urban, D. J., Anthwal, N., Howenstine, A. O., Sinha, I., and Sears, K. E. (2020). Making a bat: The developmental basis of bat evolution. Genet. Mol. Biol. 43:e20190146. doi: 10.1590/1678-4685-GMB-2019-0146
Salmon, A. B., Leonard, S., Masamsetti, V., Pierce, A., Podlutsky, A. J., Podlutskaya, N., et al. (2009). The long lifespan of two bat species is correlated with resistance to protein oxidation and enhanced protein homeostasis. FASEB J. 23, 2317–2326. doi: 10.1096/fj.08-122523
Schneider, K., Christman, M. C., and Fagan, W. F. (2011). The influence of resource subsidies on cave invertebrates: results from an ecosystem-level manipulation experiment. Ecology 92, 765–776. doi: 10.1890/10-0157.1
Secutti, S., and Trajano, E. (2009). Reproductive behavior, development and eye regression in the cave armored catfish, Ancistrus cryptophthalmus Reis, 1987 (Siluriformes: Loricariidae), breed in laboratory. Neotrop. Ichthyol. 7, 479–490. doi: 10.1590/s1679-62252009000300016
Secutti, S., and Trajano, E. (2021). Reproduction, development, asymmetry and late eye regression in the Brazilian cave catfish Ituglanis passensis (Siluriformes, Trichomycteridae): evidence contributing to the neutral mutation theory. Subterr. Biol. 38, 91–112. doi: 10.3897/subtbiol.31.60691
Simon, V., Elleboode, R., Mahé, K., Legendre, L., OrnelasGarcia, P., Espinasa, L., et al. (2017). Comparing growth in surface and cave morphs of the species Astyanax mexicanus: insights from scales. EvoDevo 8:23. doi: 10.1186/s13227-017-0086-6
Sket, B. (1997). Distribution of Proteus (Amphibia: Urodela: Proteidae) and its possible explanation. J. Biogeogr. 24, 263–280. doi: 10.1046/j.1365-2699.1997.00103.x
Speakman, J. R., and Selman, C. (2011). The free-radical damage theory: Accumulating evidence against a simple link of oxidative stress to ageing and lifespan. Bioessays 33, 255–259. doi: 10.1002/bies.201000132
Tacutu, R., Thornton, D., Johnson, E., Budovsky, A., Barardo, D., Craig, T., et al. (2018). Human ageing genomic resources: new and updated databases. Nucleic Acids Res. 46, D1083–D1090. doi: 10.1093/nar/gkx1042
Taddei Ruggiero, E. (2001). “Brachiopods of the Isca submarine cave: observations during ten years,” in Brachiopods, eds H. Brunton, L. R. M. Cocks, and S. L. Long (Milton Park: Taylor & Francis), 261–267. doi: 10.1201/9780203210437.pt4
Trajano, E. (1991). Population ecology of Pimelodella kronei, troglobitic catfish from Southeastern Brazil (Siluriformes, Pimelodidae). Environ. Biol. Fishes 30, 407–421. doi: 10.1007/bf02027984
Trajano, E. (1997). Population ecology of Trichomycterus itacarambiensis, a cave catfish from eastern Brazil (Siluriformes, Trichomycteridae). Environ. Biol. Fishes 50, 357–369. doi: 10.1023/a:1007366119261
Trajano, E., and Bichuette, M. E. (2007). Population ecology of cave armoured catfish, Ancistrus cryptophthalmus Reis 1987, from central Brazil (Siluriformes: Loricariidae). Ecol. Freshw. Fish 16, 105–115. doi: 10.1111/j.1600-0633.2006.00196.x
van der Weele, C. M., and Jeffery, W. R. (2022). Cavefish cope with environmental hypoxia by developing more erythrocytes and overexpression of hypoxiainducible genes. eLife 11:e69109. doi: 10.7554/eLife.69109
Venarsky, M. P., Huntsman, B. M., Huryn, A. D., Benstead, J. P., and Kuhajda, B. R. (2014). Quantitative food web analysis supports the energy limitation hypothesis in cave stream ecosystems. Oecologia 176, 859–869. doi: 10.1007/s00442-014-3042-3
Venarsky, M. P., Huryn, A. D., and Benstead, J. P. (2012). Re-examining extreme longevity of the cave crayfish Orconectes australis using new mark–recapture data: a lesson on the limitations of iterative size-at-age models. Freshw. Biol. 57, 1471–1481. doi: 10.1111/j.1365-2427.2012.02812.x
Voituron, Y., de Fraipont, M., Issartel, J., Guillaume, O., and Clobert, J. (2011). Extreme lifespan of the human fish (Proteus anguinus): a challenge for ageing mechanisms. Biol. Lett. 7, 105–107. doi: 10.1098/rsbl.2010.0539
Wilkens, H. (1988). Evolution and genetics of epigean and cave Astyanax fasciatus (Characidae, Pisces) support for the neutral mutation theory. Evol. Biol. 23, 271–367. doi: 10.1007/978-1-4613-1043-3_8
Wilkinson, G. S., and Adams, D. M. (2019). Recurrent evolution of extreme longevity in bats. Biol. Lett. 15:20180860. doi: 10.1098/rsbl.2018.0860
Keywords: adaptation, biospeleology, cave biology, lifespan, senescence, subterranean, troglobite, convergent evolution
Citation: Lunghi E and Bilandžija H (2022) Longevity in Cave Animals. Front. Ecol. Evol. 10:874123. doi: 10.3389/fevo.2022.874123
Received: 11 February 2022; Accepted: 12 April 2022;
Published: 20 May 2022.
Edited by:
Hope Klug, University of Tennessee at Chattanooga, United StatesReviewed by:
Oana Moldovan, Emil Racovita Institute of Speleology, Romanian Academy, RomaniaCopyright © 2022 Lunghi and Bilandžija. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Helena Bilandžija, aGVsZW5hLmJpbGFuZHppamFAaXJiLmhy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.