 Valerio Orazi1†
Valerio Orazi1† Jonas Hagge
Jonas Hagge Martin M. Gossner
Martin M. Gossner- 1Bavarian Forest National Park, Grafenau, Germany
- 2Forest Nature Conservation, Northwest German Forest Research Institute, Hann. Münden, Germany
- 3Forest Nature Conservation, Georg-August-University Göttingen, Göttingen, Germany
- 4Forest Entomology, Swiss Federal Research Institute WSL, Birmensdorf, Switzerland
- 5Department of Environmental Systems Science, Institute of Terrestrial Ecosystems, ETH Zürich, Zurich, Switzerland
- 6Field Station Fabrikschleichach, Department of Animal Ecology and Tropical Biology, Biocenter, University of Würzburg, Rauhenebrach, Germany
- 7Chair of Wildlife Ecology and Management, Faculty of Environment and Natural Resources, University of Freiburg, Freiburg im Breisgau, Germany
- 8Institute for Forest and Wildlife Management, Inland Norway University of Applied Science, Koppang, Norway
Freshwater ecosystems are among the most threatened ecosystems on the planet. Beavers are important engineers in freshwater ecosystems and reintroduction programs have enabled the recovery of beaver populations in several European countries, but the impact on biodiversity conservation is still unclear. We studied the effects of beavers on the terrestrial biodiversity of eight taxonomic groups by comparing beaver ponds with river and forest habitats in a mountain forest ecosystem in Central Europe. Among the 1,166 collected species, 196 occurred exclusively at beaver ponds, 192 in plots at the river, and 156 in the forest plots. More species of conservation concern were found at the beaver ponds (76) than on the river (67) and forest (63) plots. Abundances of bats and birds were higher at the beaver ponds than at the river or forest sites. The number of bird species at the beaver ponds was higher than at the river. The community composition of birds, beetles, and true bugs differed significantly between the beaver ponds and river plots, and for seven taxonomic groups it differed significantly between the beaver ponds and forest plots. An indicator species analysis revealed eight indicator species for the beaver pond but none for the river and forest plots. Our results demonstrate that beavers, as ecological engineers, increase habitat heterogeneity in mountain forests and thereby promote biodiversity. The expansion of beaver populations into these ecosystems should thus be supported, as it may serve as a biotic restoration tool.
Introduction
Global biodiversity is in a severe crisis, with many species, including those of freshwater communities, declining at an alarming rate (Reid et al., 2019). While the crucial role of freshwater and in particular riparian environments for ecosystem services and biodiversity is well established, these habitats are nonetheless widely threatened by habitat degradation and loss (Reid et al., 2019). In fact, freshwater species have suffered an 84% decline in their global abundance since 1970 and include the most threatened species in Central Europe (WWF, 2020). A coordinated effort based on legal protection and the reduction of critical threats is urgently needed to reverse this negative trend. Riparian habitat are, by definition, strictly related to aquatic environments and are therefore likely to face the same threats; a strong relation between the integrity of these habitats and the ecological status of rivers and streams has been suggested (Burdon et al., 2020). However, in addition to halting the loss of freshwater ecosystems, active approaches to restore degraded aquatic environments are needed. The debate on how this can be achieved is developing as knowledge advances (Palmer et al., 2014). As an alternative to direct interventions involving the substantial use of machinery, indirect strategies in which animal species are allowed to create self-regulating ecosystems (nature based solutions) can effectively bridge the gap between restoration ecology and biodiversity conservation (Moss, 2015).
Apex consumers play an essential functional role in ecosystems, through top-down forcing that induces trophic cascades, thus influencing both other species and ecological processes (Estes et al., 2011; Ripple et al., 2015). They also act as ecological engineers by modulating the availability of resources and the creation of new habitats (Jones et al., 1997). However, the engineering effects of these species are complex and vary depending on the spatio-temporal scale as well as the latitude, environment, and species (Romero et al., 2015). In terms of biodiversity, while the impact of ecological engineers is generally positive, it may also be neutral or negative (Jones et al., 1994). Consequently, an ecological engineer’s value in promoting biodiversity conservation within a given ecosystem type can be difficult to determine. Nonetheless, this knowledge can lead to innovative biodiversity conservation strategies (Wright et al., 2002).
The beaver is an ecological engineer par excellence, able to directly create freshwater habitats by raising the water level through dam building (Wright et al., 2002) and to alter existing habitats through foraging on woody and herbaceous plants (Law et al., 2014). Indeed, Eurasian beaver (Castor fiber) and North American beaver (Castor canadensis) are the only known dam builders in the animal kingdom (Butler, 1995). Beaver dams convert terrestrial habitats into aquatic and semiaquatic ecosystems whose specific features radically change the affected landscape’s biotic and abiotic components (Johnston, 2017). Moreover, the ability of beavers to fell large trees is extraordinary and tree felling adds to the heterogeneity of a landscape by increasing the amount of deadwood and by creating gaps in the tree canopy (Donkor and Fryxell, 1999). The consequences of these changes last for decades, even in beaver-modified habitats that have been abandoned for years (Aznar and Desrochers, 2008; Levine and Meyer, 2019). For these reasons, beavers have been suggested as a “restoration tool” to promote the recovery of riparian environments and to increase landscape heterogeneity through creation of new ponds (Johnston and Naiman, 1990; Law et al., 2017). Moreover, a mosaic of old, new and abandoned ponds creates a dynamic mosaic of patches with different ecological features at the landscape scale (Kivinen et al., 2020; Johnson-Bice et al., 2022).
In Europe, the Eurasian beaver was nearly hunted to extinction during the nineteenth century, but some relict populations survived the persecution and were strictly protected. In the twentieth century reintroduction projects were implemented in many countries that helped the species to re-establish large parts of their former range (Halley and Rosell, 2002; Wróbel, 2020). In the areas of their reintroduction in Central Europe, beavers first recolonized the floodplains of large streams and then the woodlands at higher elevations with smaller rivers. In areas such as the latter, which generally lack natural ponds, beaver damming activity mostly occurs at smaller streams, where it is likely to play a major role in enhancing the biodiversity of these ecosystems.
Beaver effects on species abundance and diversity were intensively studied and evaluated in the last decades. In their meta-analysis, Stringer and Gaywood (2016) evaluated the effects of beavers on species abundance and diversity. They found that 73% of the published studies reported a positive, 17% a neutral, and 10% a negative effect on biodiversity. However, the positive and negative effects were not evenly distributed across the different taxonomic groups. For example, the positive effects of beavers on the diversity and productivity of plant species were attributed to flooding and foraging, which create new habitats not commonly associated with forests and riverine areas (Stringer and Gaywood, 2016). Furthermore, the abundance of aquatic invertebrates and the productivity of the respective waters were shown to benefit from beavers because of the larger lentic zones they create (Hering et al., 2001). Most bats species benefit from the increased abundance and availability of food, as the expanded area of open water area provides them with better hunting opportunities (Nummi et al., 2011); however, for some bat species local detrimental effects were observed (Ciechanowski et al., 2011). With the exception of one study (Kuczynski et al., 2012), positive effects on bird species were also found, induced by an increase in food production and habitat heterogeneity (Stringer and Gaywood, 2016). For most terrestrial mammals, the positive effects are relatively small and occur mainly in areas where beavers have been present for decades (e.g., Nummi et al., 2019). In the few studies of the influence of beaver on terrestrial invertebrates, the focus was mostly on saproxylic beetles. For many of those species, flooding and the felling of trees create deadwood habitats and thus an overall positive effect (Zahner et al., 2006; Horak et al., 2010; Mourant et al., 2018).
In this study, we investigated the Eurasian beaver’s effect on the terrestrial species communities of eight taxonomic groups in a typical mixed mountain forest ecosystem in Central Europe after a decade of recolonization (Bavarian Forest National Park, Germany). We compared active beaver ponds and riparian and forest habitats to test the following hypotheses: (a) beaver activity creates habitats for species that do not occur in forests or at rivers showing similar ecological conditions (i.e., tree density, tree species composition); (b) species abundance and species diversity are higher around beaver ponds than at riparian and in forest ecosystems, because the beaver pond comprises also habitat elements of a river (that flows in and out of the pond) and the forest (trees of the same species and density are also at the beaver pond) and therefore more edges; (c) the differences between terrestrial species communities will be larger for beaver ponds vs. forests than for beaver ponds vs. rivers. An additional aim of our study was to identify indicator species significantly related to beaver ponds.
Materials and Methods
Study Area
The study was carried out in the Bavarian Forest National Park (BFNP), located in south-eastern Germany (Figure 1). The national park was established in 1970 and covers a mountainous landscape of 240 km2 with an altitudinal range of 600–1,453 m above sea level. The climate is humid and cold, with an annual mean precipitation of 830–1,820 mm and a mean annual air temperature that ranges between 7.2°C in the valleys and 2.0°C at higher elevations. Snow accounts for 50% of the precipitation and can persist for 5–6 months of the year in the valleys and 7–8 months of the year at higher elevations (Heurich et al., 2010). The area is covered by a mixed-mountain forest, dominated by Norway spruce (Picea abies), European beech (Fagus sylvatica), and silver fir (Abies alba) at low and intermediate elevations (Cailleret et al., 2014). Spruce ecosystems in the park have experienced intense bark beetle (Ips typographus) infestations since the 1980s, resulting in more than 8,000 ha of naturally disturbed spruce forest and large amounts of standing and fallen deadwood, especially at higher elevations (Lausch et al., 2013). The Bavarian Forest is part of the Greater Bohemian Forest Ecosystem, the largest protected contiguous forest expanse in Central Europe. On the German side, freshwater systems mainly consist of oligotrophic mountain streams and ponds in peatbogs with an acid pH; there are only three natural lakes, each of which is characterized by low productivity (Soldán et al., 2012).
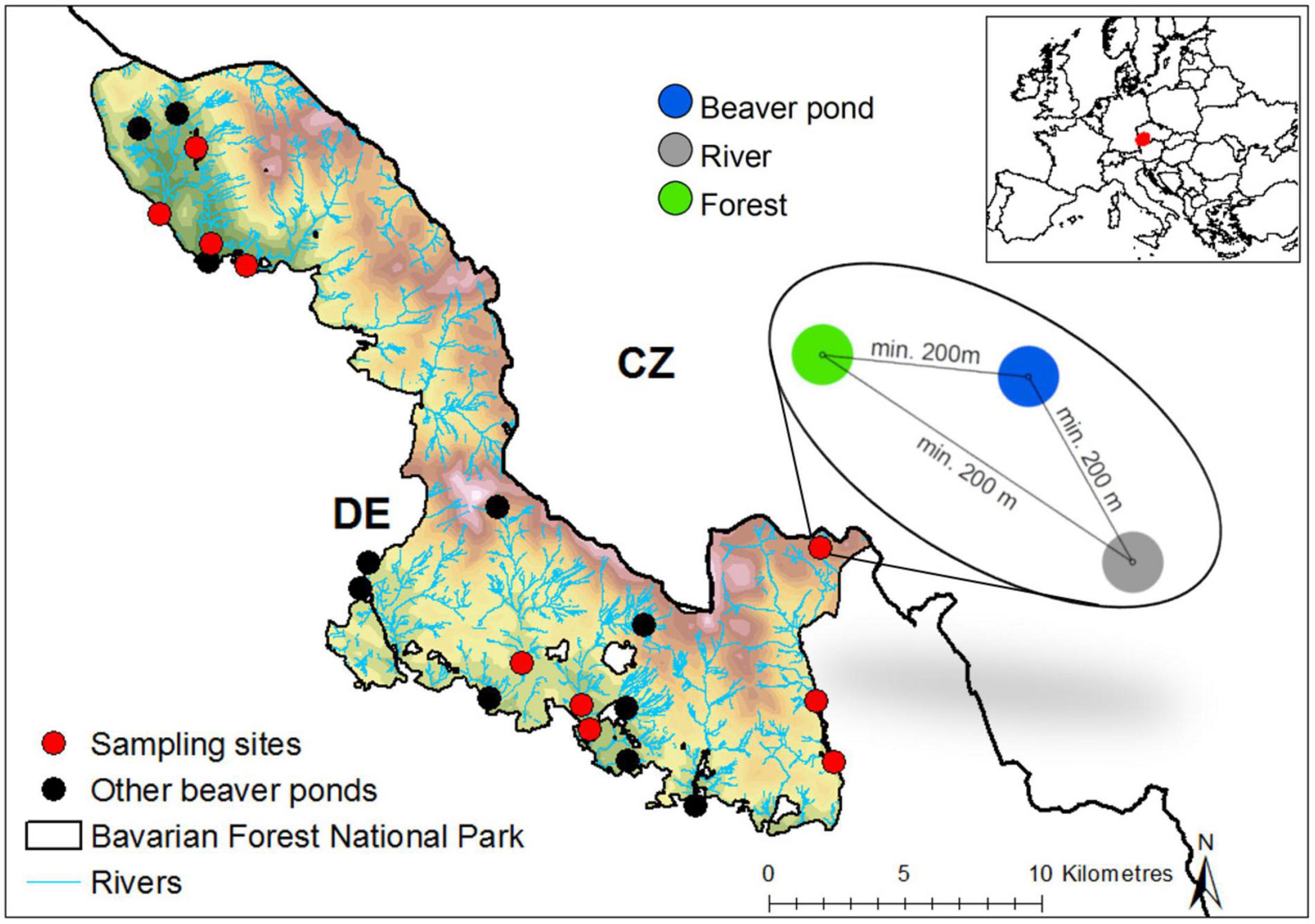
Figure 1. Sampling design and locations of the 10 research sites recoded for eight species groups in 2016. Eleven additional known beaver ponds in the Bavarian Forest National Park (Germany) are shown in the map.
In Bavaria, the last European beaver was killed in 1867. The species became legally protected in 1910 (Nolet and Rosell, 1998) with the first reintroduction in Bavaria occurring in 1966, on the initiative of the non-governmental organization Bund Naturschutz (Schwab, 2009). The success of this reintroduction program in the BFNP was confirmed in 2,000 by the first observation of a beaver colony in the park. Since then the beaver population has continuously increased, with 26 colonies recently identified (Figure 1; Bavarian Forest National Park Administration, 2020).
Sampling Design and Biodiversity Sampling
Field data were collected in a sampling framework composed of 10 blocks, each consisting of a plot at a beaver-made pond, a plot at a river (same river as the beaver pond), and a plot in the adjacent forest. River plots and forest plots (hereafter referred to as “river” and “forest”) had no signs of beaver activity (e.g., debarked trees, footprints, dams). To minimize environmental differences between the three treatments, recent (year 2015) infrared aerial photographs were used to select areas with a minimum difference in their proportions of deadwood and tree coverage (for descriptive boxplots of the environmental features of the plots, see Supplementary Figure 1). To exclude interference between plots, every plot was located at a minimum distance of 200 m from the others (Figure 1).
In 2016, eight taxonomic groups, i.e., plants, spiders, true bugs, beetles, birds, small vertebrates (including amphibians, reptiles, and small mammals), bats and terrestrial mammals were assessed on all study plots using standard methods (Table 1). Abundance was determined as the number of sampled individuals in all groups, with two exceptions: in bats the number of calls at 1-min intervals recorded per night served as a proxy for abundance (see Müller et al., 2012) and for plants the percent coverage according to Londo was used (see Table 1 for the reference and Supplementary Material for a more detailed description on species sampling).
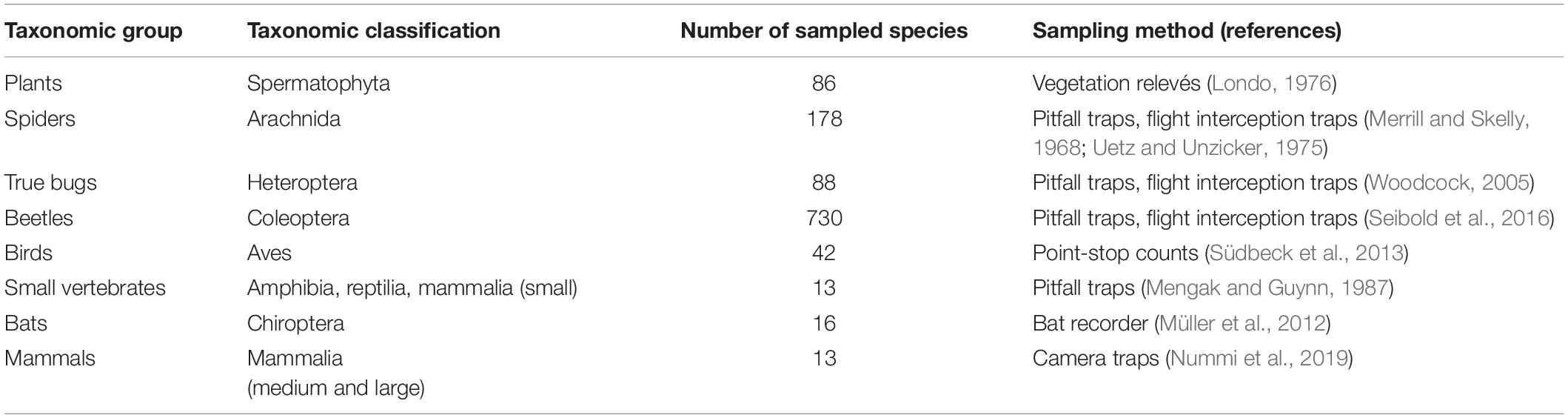
Table 1. Species groups included in the biodiversity monitoring of beaver ponds, rivers, and forests and the respective sampling method (for more details on species sampling see Supplementary Material).
Full waveform LiDAR data across the national park were collected using a Riegl680i laser scanner (350 KHz, nominal point density 30–40 points per m2; altitude 650 m). The data were recorded at a 0.32-m footprint by Milan Flug GmbH during 3 days in June 2012 under leaf-on conditions (Latifi et al., 2015). These data were used to calculate the fractional vegetation cover of three different strata: understory (0.5–2 m above ground), midstory (2–10 m above ground), and overstory vegetation cover (10–60 m above ground), within a 10-m × 10-m grid (Ewald et al., 2014). Based on single tree detections (Yao et al., 2012), the trees were classified as either dead, deciduous, or coniferous (Krzystek et al., 2020).
Data Analysis
Differences in local species diversity (alpha) and abundance among the three habitats were determined using generalized linear mixed models with a quasi-Poisson distribution for over-dispersed count data, employing the glmmPQL function in the MASS package (Ripley, 2015). The spatially nested design was taken into account by using the block (triplets of the three habitats) as a random factor in the model. The function glht in the package multcomp (Hothorn et al., 2008) was used for post hoc comparisons of the three habitats and to adjust the p-values for multiple testing. For each group, a rarefaction-extrapolation method was applied using the iNEXT package (Hsieh et al., 2016) to estimate sample coverage and gamma-diversity with respect to species incidence, based on Hill numbers (0 = species richness, 1 = exponential of Shannon’s entropy index, 2 = inverse of Simpson’s concentration index).
Differences in the community composition of each of the eight taxonomic groups and of the multitaxon community including all sampled species were identified and visualized in a non-metric multidimensional scaling (NMDS) analysis based on the Bray-Curtis distance and presence-absence data (function metaMDS in the package vegan; Oksanen et al., 2016). To test for differences among habitats a permutational multivariate analysis of variance was performed for all taxonomic groups (function adonis in the package vegan; Oksanen et al., 2016). The spatially nested design was taken into account by integrating the block (triplets of the three habitats) as the grouping factor (strata) in the analysis. Species strictly related to one of the three habitats were identified by performing an indicator species analysis using the function multipatt in the package indicspecies (De Cáceres and Legendre, 2009). The resulting p-values were adjusted using the false discovery rate method for multiple testing (Benjamini and Hochberg, 1995). The latest versions of the German Red Lists1 for each group were used to obtain information on the species conservation status. In this study, species of conservation concern were defined as species listed in the German Red Lists as extinct, critically endangered, endangered, vulnerable, threatened of unknown extent, extremely rare, or near threatened. All analyses were performed in R version 3.6.0 (R Core Team, 2019).
Results
Sample coverage within taxonomic groups was generally high (mean 85%, range 73–100%) with 1,166 species sampled in total, including 723 at the beaver pond, 740 at the river, and 637 in the forest.
Beaver Activity Provides Habitats for Unique Species
Among the species of conservation concern, more were detected at the beaver pond (76) than at the river (67) or in the forest (63) (for the number of species in each Red List category, see Supplementary Table 1). Beaver ponds harbored the highest number of unique species (species found only in this habitat) (196) and species of conservation concern (26), followed by river (192, 23) and forest (156, 16) (Figure 2). At the beaver ponds, the sample included an individual of a beetle species (Aegialia sabuleti Panzer, 1797) listed as extinct in the German Red List (see footnote 1). Further, for the beaver ponds exclusively the northern birch mouse (Sicista betulina) listed in Annex IV of Habitats Directive was recoded three times.
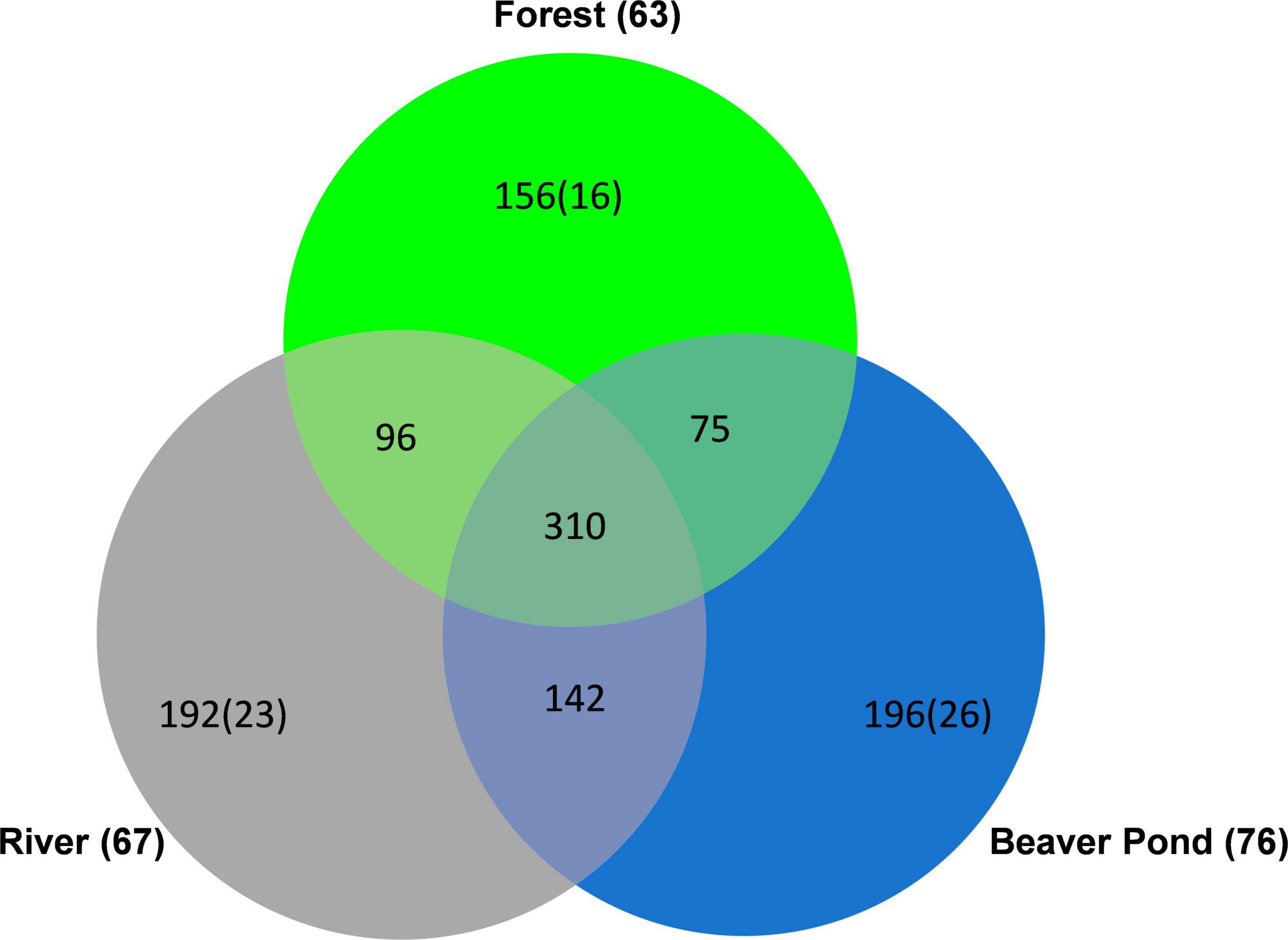
Figure 2. Venn diagram showing the number of shared and unique species at beaver ponds, rivers, and forests. The numbers in parentheses indicate the number of species of conservation concern in each habitat among the total number of species (bold) and among the number of unique species (plain). In total, 1,166 species from the eight taxonomic groups were recorded at 10 research sites in 2016 in the Bavarian Forest National Park (Germany).
Differences Between Species Abundance and Species Diversity
A significantly higher number of bird species was determined at the beaver ponds than at the river (+ 41%, Table 2). In addition, higher numbers of bird (+ 40%; marginally not significant p = 0.06), bat (+ 70%), small vertebrate (+ 45%), and plant (+ 59%) species were found at the beaver ponds than in the forest habitats. The number of species at the river plots was higher than at the forest plots for bats (+ 43%), small vertebrates (+ 33%), and plants (+ 39%) (Tables 2, 3). Abundance at the beaver ponds was significantly higher than on the forest plots for birds (+ 47%) and bats (+ 1394%), but not for beetles (-34%, marginally not significant p = 0.06). The abundances of birds (+ 63%) and bats (+ 140%) were also significantly higher at the beaver ponds than at the river. The abundance of bats (+ 43%) was significantly higher at the river than in the forest whereas the abundance of beetles was lower (-38%) (Tables 2, 3).
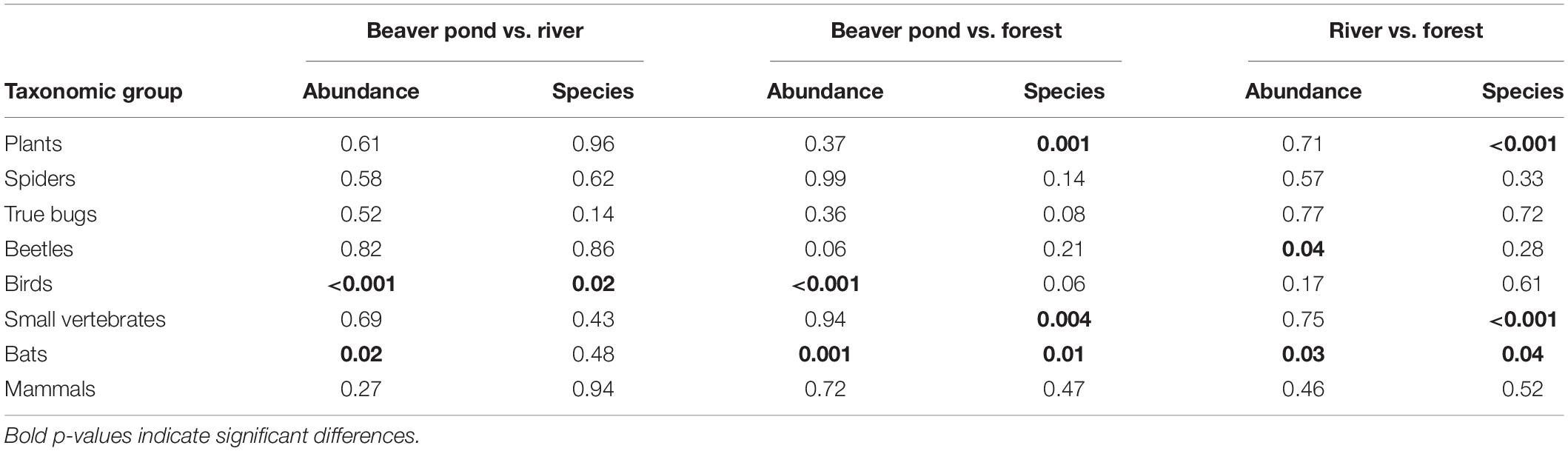
Table 2. Pairwise comparison of abundance and number of species recorded at beaver ponds and river and forest plots for the eight studied species groups at 10 research sites.
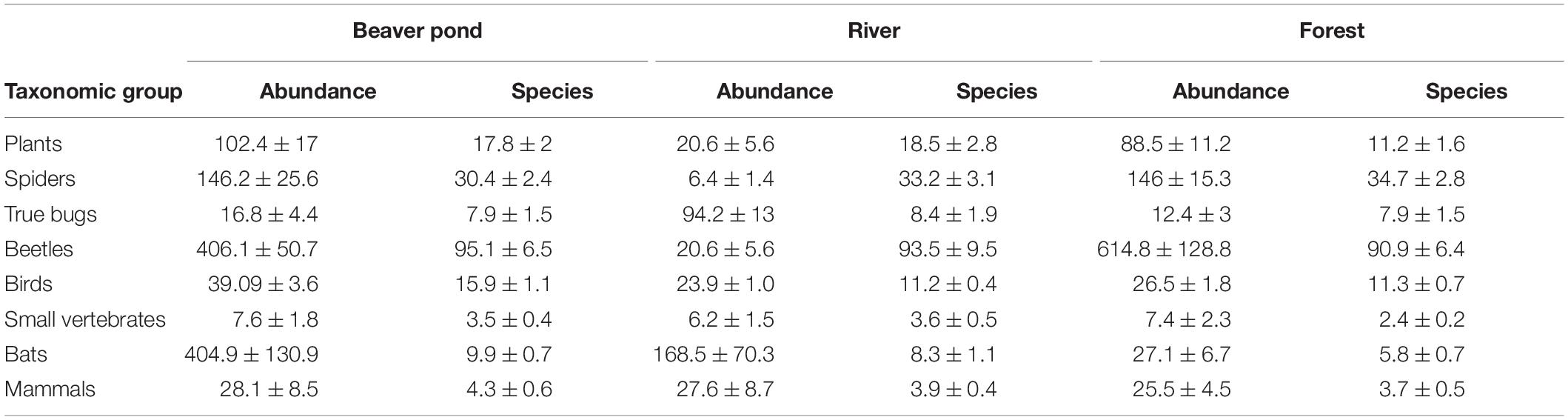
Table 3. Overview of the mean abundance and number of species (± standard error) at the beaver ponds and river and forest plots recoded at 10 research sites, expressed per study plot for the eight studied species groups.
Rarefaction–extrapolation curves (Figure 3, Hill number = 1; graphs for Hill numbers 0 and 2 are presented in Supplementary Figure 2) showed a higher gamma-diversity at the beaver pond than in the forest and river habitats for birds. The gamma-diversity of mammals and small vertebrates showed similar pattern, but with partly overlapping confidence intervals. For bats, beetles, and plants, gamma-diversity at the beaver ponds and river widely overlapped, but in both cases it was well separated from that of the forest, with a higher diversity observed at the latter. The river sites had the highest gamma-diversity for true bugs and spiders, but in the latter group the confidence intervals overlapped between habitats.
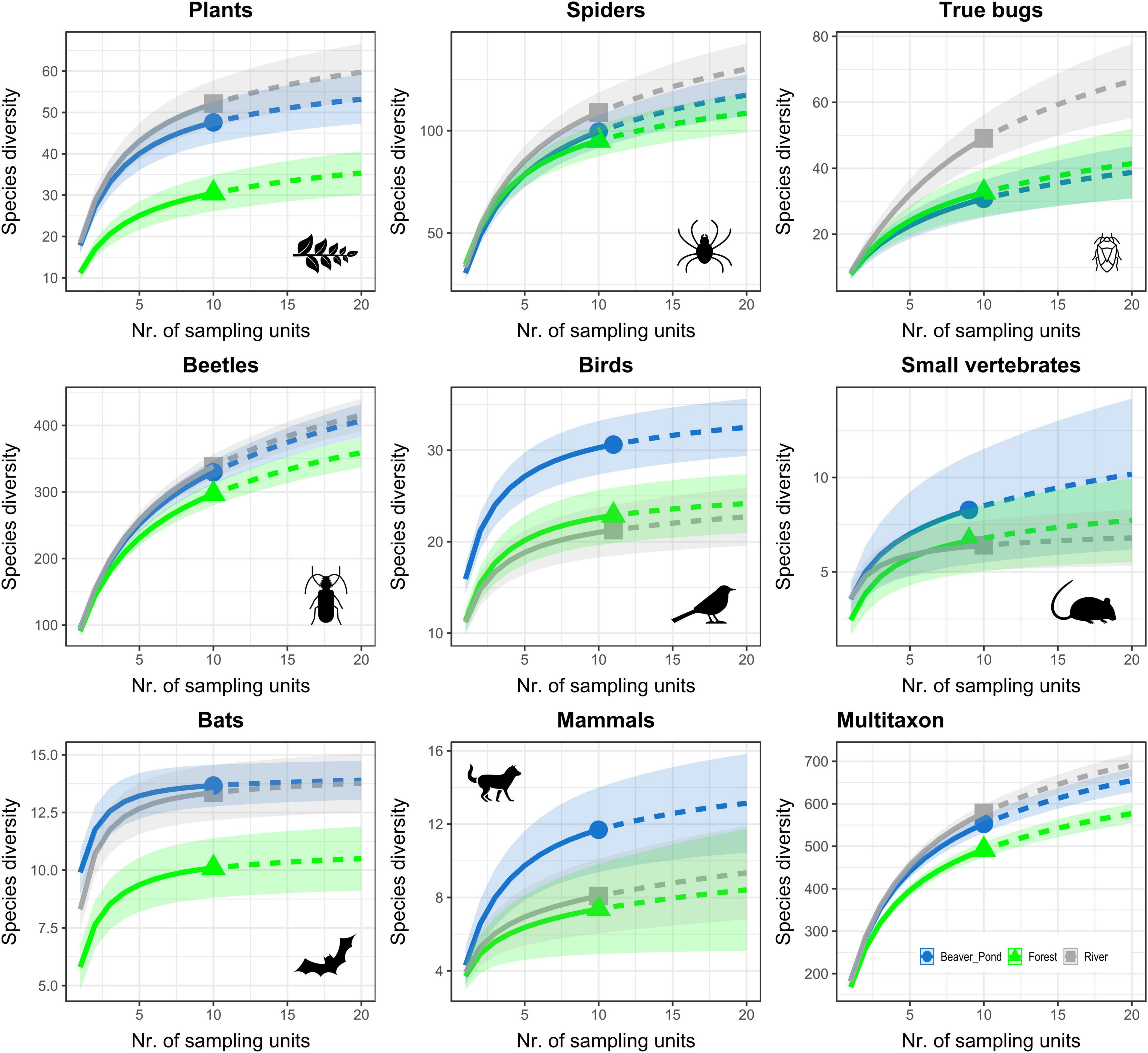
Figure 3. Sample-based rarefaction–extrapolation curves for gamma-diversity (Hill number = 1: exponential of Shannon’s entropy index) up to double the actual maximum sample size for the eight taxonomic groups and the overall community recorded at 10 research sites in 2016 in the Bavarian Forest National Park (Germany). Transparent shading represents the 95% confidence intervals and solid symbols the actual sample size.
Differences Between Species Communities
Community composition differed significantly between the beaver ponds and the river plots for birds (Adonis p-value = 0.002), beetles (0.002), and true bugs (0.02) (Figure 4 and Supplementary Table 2). In comparisons of the beaver ponds and forest plots, the community composition was significantly different for all groups, with the exception of small vertebrates (Adonis p-values: bats = 0.008, birds = 0.001, beetles = 0.001, spiders = 0.002, true bugs = 0.002, mammals = 0.01, plants = 0.002; Figure 4 and Supplementary Table 2). In the forest vs. river comparison, significant differences were determined for beetles (Adonis p-value = 0.01), spiders (0.009), true bugs (0.009), and plants (0.02) (Figure 4 and Supplementary Table 2).
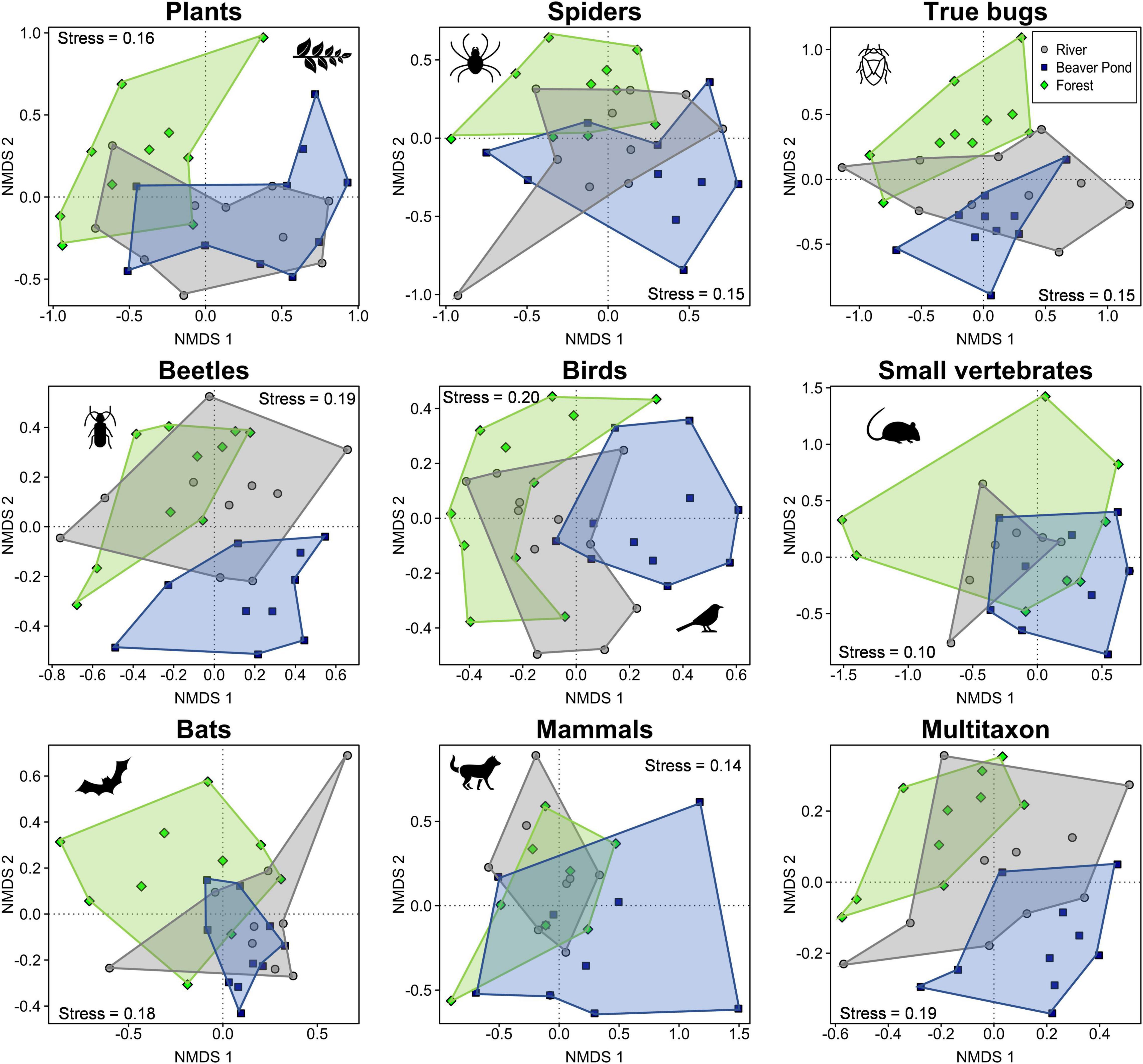
Figure 4. Non-metric multidimensional scaling (NMDS) analysis of community composition for the eight taxonomic groups and the overall community of all 1,166 sampled species recorded at 10 research sites in 2016 in the Bavarian Forest National Park (Germany). Distance between single points of samples represents community dissimilarity. Blue squares: beaver pond; gray circles: river; green rhombi: forest. Colored convex hulls group study plots according to the three habitats. Stress values for each group are shown.
The indicator analysis showed eight indicator species for the beaver pond, but none was found for either river or forest (Table 4).

Table 4. List of indicator species detected by the indicator analysis at the beaver ponds for the 10 recoded research sites.
Discussion
Our analysis showed that beavers were able to provide new habitats for a high number of terrestrial species, including unique species and species of conservation concern, thus supporting our first hypothesis. The second hypothesis was partially sustained, as we found higher diversity and abundance in some specific taxonomic group (bats and birds) at beaver ponds than on river and forest plots. The community composition of the beaver ponds differed significantly from that of the surrounding area with more pronounced differences in the comparison with forest plots (significance in seven taxonomic groups) than with river plots (significance in three taxonomic groups), thus confirming the third hypothesis. In addition, indicator species were only found for beaver ponds. Overall, our findings suggest that beavers are able to increase the alpha- and gamma-diversity of a mountain forest ecosystem in Central Europe, by creating ponds that in turn provide habitats for complementary species communities that includes rare and threatened species.
In mountainous forest landscapes few if any natural ponds exist such that aquatic ecosystems are mainly represented by small rivers and a few artificial ponds. At the ponds created by beaver, we identified almost 200 species that were either detected in the forest nor along the river. This finding suggests that the establishment of these ponds promoted the appearance of species were rare or absent in the landscape before the beaver’s arrival. Moreover, it demonstrated that some terrestrial species groups, the focus of our study, benefit from newly created ponds. Among the rare species uniquely found at the beaver ponds were the European polecat (Mustela putorius) and the Alpine newt (Ichthyosaura alpestris), thus indicating a potential role of these environments in the conservation of rare species (Bartel et al., 2010). While the total number of species was slightly higher at the river than at the beaver ponds, the number of species of conservation concern was higher in the latter (67 and 76 species, respectively) and included a beetle species considered extinct in Germany (Aegialia sabuleti) and one of the rarest mammals in central Europe, the northern birch mouse (Sicista betulina) listed in Annex IV of the Habitats Directive. Our results indicate that beaver ponds promote the development of communities of birds, beetles, and true bugs that are distinct from those found associated with river and forest habitats. Thus, preserving these ponds also preserves entire communities that hardly occur, if at all, in forested landscapes and increase landscape level diversity (Wright et al., 2002). This contribution of beavers to conservation efforts is especially valuable considering the increasing threats to riparian biodiversity (Reid et al., 2019).
Birds benefited most from the presence of beaver, evidenced by both a higher number of species and a higher abundance at the beaver ponds than on the river and forest plots. The presence of a specific bird community and the higher gamma-diversity of birds at the beaver ponds together suggest that beaver activity adds components to the riparian ecosystem that can be exploited by birds, such as an expansion of water and wetlands coverage (Grover and Baldassarre, 1995) and a wider variety of micro-habitats that increase food resources, which are both determinants of bird community composition (Bulluck and Rowe, 2006). Previous studies reported that, at beaver ponds, waterbirds such as mallard (Anas platyrhynchos) and Eurasian teal (A. crecca) can achieve both a higher diversity at the community level (Nummi and Holopainen, 2014) and higher brood densities (Longcore et al., 2006). In addition to the wide variety of riparian and non-riparian species [including, as in this study, gray wagtail (Motacilla cinerea) and Eurasian siskin (Carduelis spinus)] that benefit from beaver-induced habitat modifications (Cooke and Zack, 2008), positive effects have been demonstrated for species that are much less dependent on wetlands, such as woodpeckers, due to the increased variety and amount of deadwood (Lochmiller, 1979) and possibly a high number of emerging insects (Nummi et al., 2011). Moreover, in the BFNP, park administrators have confirmed the breeding of the common snipe (Gallinago gallinago), the first such observation in 50 years, and the common crane (Grus grus), thus further highlighting the importance of beaver activity in promoting bird diversity (Longcore et al., 2006). Given the alarming rate of decline of several European bird species (Inger et al., 2015), the presence of a well-structured, undisturbed beaver pond system can act as a shelter, thereby allowing the recovery and conservation of avian populations.
The high bat abundance observed at the studied beaver ponds can be explained by the richness and abundance of aquatic invertebrate taxa in these highly productive habitats (Hood and Larson, 2014; Bush and Wissinger, 2016), which would improve the foraging success of insectivorous taxa. Ponds in mountain areas also provide an important source of drinking water for bats (Seibold et al., 2013). The essential role of ponds in the ecology of many bat species is consistent with our identification of Bechstein’s bat (Myotis bechsteinii) and common pipistrelle (Pipistrellus pipistrellus) as indicator species of beaver ponds, including those in the mountainous forest ecosystem of the BFNP. Similar results were reported from boreal forests and suggest a positive impact of beaver-mediated pond generation on the conservation of bats in different biomes (Mickleburgh et al., 2002).
True bugs, beetles, and spiders were selected as representative invertebrate communities in this study because of their ecological and functional roles in the microhabitats of both terrestrial and aquatic ecosystems. Although beaver-related effects on macroinvertebrate taxa have been widely studied (see Bush and Wissinger, 2016 for a complete review), most of that research focused on water-dwelling species whereas terrestrial species have been largely ignored. Our study contributes to closing this gap in that it revealed a specific community composition for true bugs and beetles at beaver ponds. In the beetles’ group, the community composition was specific to the studied habitats. While the number of species did not differ between those habitats, beetle abundance tended to be higher in the forest than at the river or at the beaver ponds (although barely significantly different in the latter). In Germany, the total number of forest-dwelling species is estimated to be 13,600 (Schmidt, 2006; Dorow et al., 2019), with saproxylic organisms, mostly fungi and beetles, accounting for 34% (Müller et al., 2008). Among the important consequences of beaver activity is the creation of deadwood of several different types in areas that rarely experience other disturbance events (Thompson et al., 2016). In the BFNP, an intense bark beetle (I. typographus) outbreak has resulted in the generation of large amounts of Norway spruce (P. abies) deadwood, such that it is not a limited resource (Müller et al., 2010). However, in the park’s buffer zone (the location of some of the analyzed beaver ponds) dead spruce trees have been removed to prevent bark beetle spread to the adjacent managed forest. This might have limited the beaver’s effect on saproxylic organisms (especially beetles) at the respective plots, such that neither the species diversity nor the abundance of beetles was significantly higher at the beaver ponds than in the forest or river plots. Nonetheless, distinct communities of beetle species were detected in each of the studied habitats, which in the case of beaver ponds demonstrated their ability to create biological diversity in a forested landscape.
Our analysis showed no difference in the number of species, abundance, or community composition of vascular plants between the beaver ponds and river plots. Beavers use vegetation as a source of food and as materials for dam building, thereby increasing the openness and moisture level of the surrounding forest (Johnston, 2017). While these changes can influence plant communities in various ways (Stringer and Gaywood, 2016), previous studies have shown that in high-quality habitats the effect of beavers on the vegetation may be neutral (Willby et al., 2014). In some environments, aquatic plants constitute 60–80% of the beaver diet (Milligan and Humphries, 2010), such that foraging activity on terrestrial plants is minimal. However, aquatic plants were not considered in our study. Follow up studies should thus include cluster sampling (Brown and Manly, 1998). It should also be noted that our vegetation recordings were obtained from the immediate vicinity of the pond and river and because of the limited size of the plots (10 m × 10 m) they probably did not fully capture the heterogeneous effects of beaver activities.
The ponds surveyed in this study were relatively new (median: 4 years; range: 1–12 years), and the taxonomic groups that seemed to benefit most from their presence were mainly bats and birds. The high mobility of these animals allows them to reach recently created ponds faster than other, less mobile taxa (e.g., invertebrates and small mammals). It can therefore be expected that the ecological differences identified in this study will become more pronounced over time, as observed at other study sites (Sferra et al., 2017), and that the ponds will eventually be colonized by species not previously found in the area (Fairchild et al., 2000; Stevens et al., 2006; Bonner et al., 2009; Ecke et al., 2017). This is especially likely considering that before the arrival of beavers the mountainous study site was largely free of natural ponds. At the landscape scale, the presence of different successional stages of beaver patches can result in increased species diversity (Bush et al., 2019; Nummi et al., 2021). Particular long-term beaver presence can add a further level of complexity to the landscape, as the strength of its engineering effects seems to be time and density dependent (Johnson-Bice et al., 2022).
Conclusion
In mountainous forest ecosystems, aquatic communities are often underrepresented because even riverine areas are dominated by forest cover. By cutting trees, beavers open the forests and create ponds and wetlands, thus increasing the extent of these ecosystems as well as habitat availability for many species. Our study showed that, by positively impacting both abundance and species diversity, beavers are essential to the restoration of river systems, even in mountainous forest ecosystems. However, most beaver territories included in this study were relatively new, as beaver recolonization is a recent event and the number of colonies is still increasing. Therefore, even more pronounced effects of the presence of beaver can be expected in the near future, following increases in the number and heterogeneity of ponds in the landscape (Hill et al., 2018).
Our findings provide a basis for long-term research, using the same methodology in the same beaver territories, to observe the effects of beavers throughout the evolution of their territories, including old ponds and beaver meadows (presently not available). This will be a valuable opportunity to observe and quantify the changes induced by the presence of beavers and thus increase our knowledge of related ecosystem processes and community responses over time. The resulting insights will lead to more effective measures aimed at conserving biodiversity within and outside protected areas. While beaver conservation is not a universal solution to the biodiversity crisis, it can certainly be part of a larger strategy to improve the health and conservation status of whole communities, especially those in threatened riparian environments.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Ethics Statement
Ethical review and approval was not required for the animal study because we used non-invasive passive species sampling.
Author Contributions
MH, JH, and JM: conception of the study. MH and JH: data collection. MG: species identification of true bugs. VO and JH: statistical analysis. MH, VO, and JH: writing the first draft. MG and JM: revisions. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the Bavarian Forest National Park Administration.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Maik Henrich for creating the map and Karina Anders, Leo Christoph, Mathias Haunstetter, Jan Hemmerling, Paul Hertlein, Julia Hetzl, Johannes Jakob, Sophie Kirberg, Karolin Prott, Katrin Schäfer, and Moritz Waas for performing the fieldwork and sorting the insects. We are also grateful for the language revision by Wendy Ran.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.873307/full#supplementary-material
Footnotes
References
Aznar, J.-C., and Desrochers, A. (2008). Building for the future: abandoned beaver ponds promote bird diversity. Écoscience 15, 250–257. doi: 10.2980/15-2-3107
Bartel, R. A., Haddad, N. M., and Wright, J. P. (2010). Ecosystem engineers maintain a rare species of butterfly and increase plant diversity. Oikos 119, 883–890. doi: 10.1111/j.1600-0706.2009.18080.x
Bavarian Forest National Park Administration (2020). (Grafenau: Nationalparkverwaltung Bayerischer Wald), 18.
Benjamini, Y., and Hochberg, Y. (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 57, 289–300. doi: 10.2307/2346101
Bonner, J. L., Anderson, J. T., Rentch, J. S., and Grafton, W. N. (2009). Vegetative composition and community structure associated with beaver ponds in Canaan valley, West Virginia, USA. Wetl. Ecol. Manag. 17, 543–554. doi: 10.1007/s11273-009-9131-0
Brown, J. A., and Manly, B. J. F. (1998). Restricted adaptive cluster sampling. Environ. Ecol. Stat. 5, 49–63. doi: 10.1023/A:1009607403647
Bulluck, J. F., and Rowe, M. P. (2006). The use of Southern Appalachian wetlands by breeding birds, with a focus on Neotropical migratory species. Wilson J. Ornithol. 118, 399–410. doi: 10.1676/04-116.1
Burdon, F. J., Ramberg, E., Sargac, J., Forio, M. A. E., de Saeyer, N., Mutinova, P. T., et al. (2020). Assessing the benefits of forested riparian zones: a qualitative index of riparian integrity is positively associated with ecological status in european streams. Water 12:1178. doi: 10.3390/w12041178
Bush, B. M., Stenert, C., Maltchik, L., and Batzer, D. P. (2019). Beaver-created successional gradients increase β−diversity of invertebrates by turnover in stream-wetland complexes. Freshw. Biol. 64, 1265–1274. doi: 10.1111/fwb.13302
Bush, B. M., and Wissinger, S. A. (2016). “Invertebrates in beaver-created wetlands and ponds,” in Invertebrates in Freshwater Wetlands, (Manhattan, NY: Springer International Publishing). 411–449. doi: 10.1007/978-3-319-24978-0_12
D. R. Butler (ed.) (1995). “The geomorphic influence of beavers,” in Zoogeomorphology, (Cambridge, MA: Cambridge University Press). 148–183. doi: 10.1017/CBO9780511529900.008
Cailleret, M., Heurich, M., and Bugmann, H. (2014). Reduction in browsing intensity may not compensate climate change effects on tree species composition in the Bavarian Forest National Park. For. Ecol. Manag. 328, 179–192. doi: 10.1016/j.foreco.2014.05.030
Ciechanowski, M., Kubic, W., Rynkiewicz, A., and Zwolicki, A. (2011). Reintroduction of beavers Castor fiber may improve habitat quality for vespertilionid bats foraging in small river valleys. Eur. J. Wildl. Res. 57, 737–747. doi: 10.1007/s10344-010-0481-y
Cooke, H. A., and Zack, S. (2008). Influence of beaver dam density on riparian areas and riparian birds in shrubsteppe of Wyoming. West. North Am. Nat. 68, 365–373. doi: 10.3398/1527-0904(2008)68[365:iobddo]2.0.co;2
De Cáceres, M., and Legendre, P. (2009). Associations between species and groups of sites: indices and statistical inference. Ecology 90, 3566–3574. doi: 10.1890/08-1823.1
Donkor, N. T., and Fryxell, J. M. (1999). Impact of beaver foraging on structure of lowland boreal forests of Algonquin Provincial Park. Ontario. For. Ecol. Manag. 118, 83–92. doi: 10.1016/S0378-1127(98)00487-3
Dorow, W., Blick, T., Pauls, S., and Schneider, A. (2019). Waldbindung Ausgewählter Tiergruppen Deutschlands: Lumbricidae, Araneae, Opiliones, Pseudoscorpiones, Heteroptera, Coleoptera, Aculeata, Macrolepidoptera, Aves. Bonn: Bundesamt für Naturschutz. 544.
Ecke, F., Levanoni, O., Audet, J., Carlson, P., Eklöf, K., Hartman, G., et al. (2017). Meta-analysis of environmental effects of beaver in relation to artificial dams. Environ. Res. Lett. 12:113002. doi: 10.1088/1748-9326/aa8979
Estes, J. A., Terborgh, J., Brashares, J. S., Power, M. E., Berger, J., Bond, W. J., et al. (2011). Trophic downgrading of planet earth. Science 333, 301–306. doi: 10.1126/science.1205106
Ewald, M., Dupke, C., Heurich, M., Müller, J., and Reineking, B. (2014). LiDAR remote sensing of forest structure and GPS telemetry data provide insights on winter habitat selection of european roe deer. Forests 5, 1374–1390. doi: 10.3390/f5061374
Fairchild, G. W., Faulds, A. M., and Matta, J. F. (2000). Beetle assemblages in ponds: effects of habitat and site age. Freshw. Biol. 44, 523–534. doi: 10.1046/j.1365-2427.2000.00601.x
Grover, A. M., and Baldassarre, G. A. (1995). Bird species richness within beaver ponds in south-central New York. Wetlands 15, 108–118. doi: 10.1007/BF03160664
Halley, D. J., and Rosell, F. (2002). The beaver’s reconquest of Eurasia: status, population development and management of a conservation success. Mamm. Rev. 32, 153–178. doi: 10.1046/j.1365-2907.2002.00106.x
Hering, D., Gerhard, M., Kiel, E., Ehlert, T., and Pottgiesser, T. (2001). Review study on near-natural conditions of central european mountain streams, with particular reference to debris and beaver dams: results of the “REG meeting” 2000. Limnologica 31, 81–92. doi: 10.1016/S0075-9511(01)80001-3
Heurich, M., Beudert, B., Rall, H., and Křenová, Z. (2010). “National parks as model regions for interdisciplinary long-term ecological research: the bavarian forest and ??umav?? national parks underway to transboundary ecosystem research,” in Long-term Ecological Research. Between Theory and Application, eds F. Muller, C. Baessler, H. Schubert, and S. Klotz (Berlin: Springer). 327–344. doi: 10.1007/978-90-481-8782-9_23
Hill, M. J., Hassall, C., Oertli, B., Fahrig, L., Robson, B. J., Biggs, J., et al. (2018). New policy directions for global pond conservation. Conserv. Lett. 11:e12447. doi: 10.1111/conl.12447
Hood, G. A., and Larson, D. G. (2014). Beaver-created habitat heterogeneity influences aquatic invertebrate assemblages in boreal Canada. Wetlands 34, 19–29. doi: 10.1007/s13157-013-0476-z
Horak, J., Vavrova, E., and Chobot, K. (2010). Habitat preferences influencing populations, distribution and conservation of the endangered saproxylic beetle Cucujus cinnaberinus (Coleoptera: Cucujidae) at the landscape level. Eur. J. Entomol. 107, 81–88. doi: 10.14411/eje.2010.011
Hothorn, T., Bretz, F., and Westfall, P. (2008). Simultaneous inference in general parametric models. Biom. J. 50, 346–363. doi: 10.1002/bimj.200810425
Hsieh, T. C., Ma, K. H., and Chao, A. (2016). iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 6, 1451–1456. doi: 10.1111/2041-210X.12613
Inger, R., Gregory, R., Duffy, J. P., Stott, I., Voříšek, P., and Gaston, K. J. (2015). Common European birds are declining rapidly while less abundant species’ numbers are rising. Ecol. Lett. 18, 28–36. doi: 10.1111/ele.12387
Johnson-Bice, S. M., Gable, T. D., Windels, S. K., and Host, G. E. (2022). Relics of beavers past: time and population density drive scale-dependent patterns of ecosystem engineering. Ecography 2022:e05814. doi: 10.1111/ecog.05814
Johnston, C. A. (2017). Beavers: Boreal Ecosystem Engineers. New York, NY: Springer International Publishing. doi: 10.1007/978-3-319-61533-2
Johnston, C. A., and Naiman, R. J. (1990). Aquatic patch creation in relation to beaver population trends. Ecology 71, 1617–1621. doi: 10.2307/1938297
Jones, C. G., Lawton, J. H., and Shachak, M. (1994). Organisms as ecosystem engineers. Oikos 69:373. doi: 10.2307/3545850
Jones, C. G., Lawton, J. H., and Shachak, M. (1997). Positive and negative effects of organisms as physical ecosystem engineers. Ecology 78, 1946–1957. doi: 10.1890/0012-9658(1997)078[1946:paneoo]2.0.co;2
Kivinen, S., Nummi, P., and Kumpula, T. (2020). Beaver-induced spatiotemporal patch dynamics affect landscape-level environmental heterogeneity. Environ. Res. Lett. 15:094065. doi: 10.1088/1748-9326/ab9924
Krzystek, P., Serebryanyk, A., Schnörr, C., Červenka, J., and Heurich, M. (2020). Large-scale mapping of tree species and dead trees in šumava national park and bavarian forest national park using lidar and multispectral imagery. Remote Sens. 12:661. doi: 10.3390/rs12040661
Kuczynski, E. C., Paszkowski, C. A., and Gingras, B. A. (2012). Horned grebe habitat use of constructed wetlands in Alberta, Canada. J. Wildlife Manag. 76, 1694–1702. doi: 10.1002/jwmg.421
Latifi, H., Fassnacht, F. E., Müller, J., Tharani, A., Dech, S., and Heurich, M. (2015). Forest inventories by LiDAR data: a comparison of single tree segmentation and metric-based methods for inventories of a heterogeneous temperate forest. Int. J. Appl. Earth Obs. Geoinf. 42, 162–174. doi: 10.1016/j.jag.2015.06.008
Lausch, A., Heurich, M., and Fahse, L. (2013). Spatio-temporal infestation patterns of Ips typographus (L.) in the Bavarian Forest National Park. Germany. Ecol. Indic. 31, 73–81. doi: 10.1016/j.ecolind.2012.07.026
Law, A., Gaywood, M. J., Jones, K. C., Ramsay, P., and Willby, N. J. (2017). Using ecosystem engineers as tools in habitat restoration and rewilding: beaver and wetlands. Sci. Total Environ. 605-606, 1021–1030. doi: 10.1016/j.scitotenv.2017.06.173
Law, A., Jones, K. C., and Willby, N. J. (2014). Medium vs. short-term effects of herbivory by Eurasian beaver on aquatic vegetation. Aquat. Bot. 116, 27–34. doi: 10.1016/j.aquabot.2014.01.004
Levine, R., and Meyer, G. A. (2019). Beaver-generated disturbance extends beyond active dam sites to enhance stream morphodynamics and riparian plant recruitment. Sci. Rep. 9:8124. doi: 10.1038/s41598-019-44381-2
Lochmiller, R. L. (1979). Use of Beaver Ponds by Southeastern Woodpeckers in Winter. J. Wildl. Manage. 43, 263–266. doi: 10.2307/3800670
Londo, G. (1976). The decimal scale for releves of permanent quadrats. Vegetatio 33, 61–64. doi: 10.1007/BF00055300
Longcore, J. R., McAuley, D. G., Pendelton, G. W., Bennatti, C. R., Mingo, T. M., and Stromborg, K. L. (2006). Macroinvertebrate abundance, water chemistry, and wetland characteristics affect use of wetlands by avian species in Maine. Hydrobiologia 567, 143–167. doi: 10.1007/s10750-006-0055-x
Mengak, M. T., and Guynn, D. C. (1987). Pitfalls and snap traps for sampling small mammals and Herpetofauna. Am. Midl. Nat. 118:284. doi: 10.2307/2425786
Merrill, W., and Skelly, J. M. (1968). A window trap for collection of insects above the forest canopy. J. Econ. Entomol. 61, 1461–1462. doi: 10.1093/jee/61.5.1461
Mickleburgh, S. P., Hutson, A. M., and Racey, P. A. (2002). A review of the global conservation status of bats. Oryx 36, 18–34. doi: 10.1017/S0030605302000054
Milligan, H. E., and Humphries, M. M. (2010). The importance of aquatic vegetation in beaver diets and the seasonal and habitat specificity of aquatic-terrestrial ecosystem linkages in a subarctic environment. Oikos 119, 1877–1886. doi: 10.1111/j.1600-0706.2010.18160.x
Moss, B. (2015). Mammals, freshwater reference states, and the mitigation of climate change. Freshw. Biol. 60, 1964–1976. doi: 10.1111/fwb.12614
Mourant, A., Lecomte, N., and Moreau, G. (2018). Indirect effects of an ecosystem engineer: how the Canadian beaver can drive the reproduction of saproxylic beetles. J. Zool. 304, 90–97. doi: 10.1111/jzo.12506
Müller, J., Bußler, H., and Kneib, T. (2008). Saproxylic beetle assemblages related to silvicultural management intensity and stand structures in a beech forest in Southern Germany. J. Insect Conserv. 12, 107–124. doi: 10.1007/s10841-006-9065-2
Müller, J., Mehr, M., Bässler, C., Fenton, M. B., Hothorn, T., Pretzsch, H., et al. (2012). Aggregative response in bats: prey abundance versus habitat. Oecologia 169, 673–684. doi: 10.1007/s00442-011-2247-y
Müller, J., Noss, R. F., Bussler, H., and Brandl, R. (2010). Learning from a “benign neglect strategy” in a national park: response of saproxylic beetles to dead wood accumulation. Biol. Conserv. 143, 2559–2569. doi: 10.1016/j.biocon.2010.06.024
Nolet, B. A., and Rosell, F. (1998). Comeback of the beaver Castor fiber: an overview of old and new conservation problems. Biol. Conserv. 83, 165–173. doi: 10.1016/S0006-3207(97)00066-9
Nummi, P., and Holopainen, S. (2014). Whole-community facilitation by beaver: ecosystem engineer increases waterbird diversity. Aquat. Conserv. 24, 623–633. doi: 10.1002/aqc.2437
Nummi, P., Kattainen, S., Ulander, P., and Hahtola, A. (2011). Bats benefit from beavers: a facilitative link between aquatic and terrestrial food webs. Biodivers. Conserv. 20, 851–859. doi: 10.1007/s10531-010-9986-7
Nummi, P., Liao, W., Huet, O., Scarpulla, E., and Sundell, J. (2019). The beaver facilitates species richness and abundance of terrestrial and semi-aquatic mammals. Glob. Ecol. Conserv. 20:e00701. doi: 10.1016/j.gecco.2019.e00701
Nummi, P., Liao, W., van der Schoor, J., and Loehr, J. (2021). Beaver creates early successional hotspots for water beetles. Biodivers. Conserv. 30, 2655–2670. doi: 10.1007/s10531-021-02213-8
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O’Hara, R. B., et al. (2016). vegan: community ecology package. R package. doi: 10.1016/j.dib.2020.105500
Palmer, M. A., Hondula, K. L., and Koch, B. J. (2014). Ecological restoration of streams and rivers: shifting strategies and shifting goals. Annu. Rev. Ecol. Evol. Syst. 45, 247–269. doi: 10.1146/annurev-ecolsys-120213-091935
Reid, A. J., Carlson, A. K., Creed, I. F., Eliason, E. J., Gell, P. A., Johnson, P. T. J., et al. (2019). Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 94, 849–873. doi: 10.1111/brv.12480
Ripley, B. (2015). Package MASS: Support Functions and Datasets for Venables and Ripley’s Mass, Version: 7.3–56. Available online at: https//cran.r-project.org/web/packages/MASS/index.Html (accessed January 20, 2022).
Ripple, W. J., Newsome, T. M., Wolf, C., Dirzo, R., Everatt, K. T., Galetti, M., et al. (2015). Collapse of the world’s largest herbivores. Sci. Adv. 1:e1400103. doi: 10.1126/sciadv.1400103
Romero, G. Q. Gonçalves-Souza, T., Vieira, C., and Koricheva, J. (2015). Ecosystem engineering effects on species diversity across ecosystems: a meta-analysis. Biol. Rev. 90, 877–890. doi: 10.1111/brv.12138
Schwab, G. (2009). Biber in Bayern – Biologie und Management. Augsburg: Bayerisches Landesamt für Umwelt (LfU), 49.
Seibold, S., Bässler, C., Brandl, R., Büche, B., Szallies, A., Thorn, S., et al. (2016). Microclimate and habitat heterogeneity as the major drivers of beetle diversity in dead wood. J. Appl. Ecol. 53, 934–943. doi: 10.1111/1365-2664.12607
Seibold, S., and Buchner, J. Bässler, C.Müller, J. (2013). Ponds in acidic mountains are more important for bats in providing drinking water than insect prey. J. Zool. 290, 302–308. doi: 10.1111/jzo.12041
Sferra, C. O., Hart, J. L., and Howeth, J. G. (2017). Habitat age influences metacommunity assembly and species richness in successional pond ecosystems. Ecosphere 8:e01871. doi: 10.1002/ecs2.1871
Soldán, T., Bojková, J., Vrba, J., Bitušík, P., Chvojka, P., Papáček, M., et al. (2012). Aquatic insects of the Bohemian Forest glacial lakes: diversity, long-term changes, and influence of acidification. Silva Gabreta 18, 123–283.
Stevens, C. E., Paszkowski, C. A., and Scrimgeour, G. J. (2006). Older is better: beaver ponds on boreal streams as breeding habitat for the wood frog. J. Wildl. Manage. 70, 1360–1371. doi: 10.2193/0022-541x(2006)70[1360:oibbpo]2.0.co;2
Stringer, A. P., and Gaywood, M. J. (2016). The impacts of beavers Castor spp. on biodiversity and the ecological basis for their reintroduction to Scotland, UK. Mamm. Rev. 46, 270–283. doi: 10.1111/mam.12068
Südbeck, P., Andretzke, H., Fischer, S., Gedeon, K., Schikore, T., Schröder, K., et al. (2013). Methodenstandards zur Erfassung der Brutvögel Deutschlands. Naturschutz im L. Sachsen-Anhalt 42, 60–61.
Thompson, S., Vehkaoja, M., and Nummi, P. (2016). Beaver-created deadwood dynamics in the boreal forest. For. Ecol. Manag. 360, 1–8. doi: 10.1016/j.foreco.2015.10.019
Uetz, G. W., and Unzicker, J. D. (1975). Pitfall trapping in ecological studies of wandering spiders. J. Arachnol. 3, 101–111.
Willby, N., Perfect, C., and Law, A. (2014). The scottish beaver trial: monitoring of aquatic vegetation and associated features of the knapdale lochs 2008-2013, final report. Scottish Nat. Herit. Comm. Rep. 688, 1–127.
Woodcock, B. A. (2005). “Pitfall Trapping in Ecological Studies,” in Insect Sampling in Forest Ecosystems, (Hoboken, NJ: Wiley-Blackwell). 37–57. doi: 10.1002/9780470750513.ch3
Wright, J. P., Jones, C. G., and Flecker, A. S. (2002). An ecosystem engineer, the beaver, increases species richness at the landscape scale. Oecologia 132, 96–101. doi: 10.1007/s00442-002-0929-1
Wróbel, M. (2020). Population of Eurasian beaver (Castor fiber) in Europe. Glob. Ecol. Conserv. 23:e01046. doi: 10.1016/j.gecco.2020.e01046
WWF (2020). “Living Planet Report 2020,” in Bending the Curve of Biodiversity Loss, eds R. E. A. Almond, M. Grooten, and T. Petersen (Switzerland: WWF). 43–47.
Yao, W., Krzystek, P., and Heurich, M. (2012). Tree species classification and estimation of stem volume and DBH based on single tree extraction by exploiting airborne full-waveform LiDAR data. Remote Sens. Environ. 123, 368–380. doi: 10.1016/j.rse.2012.03.027
Keywords: multitaxon biodiversity, ecosystem engineer, habitat restoration, Eurasian beaver (Castor fiber), Bavarian Forest National Park
Citation: Orazi V, Hagge J, Gossner MM, Müller J and Heurich M (2022) A Biodiversity Boost From the Eurasian Beaver (Castor fiber) in Germany’s Oldest National Park. Front. Ecol. Evol. 10:873307. doi: 10.3389/fevo.2022.873307
Received: 10 February 2022; Accepted: 31 March 2022;
Published: 13 May 2022.
Edited by:
Hideyuki Doi, University of Hyogo, JapanReviewed by:
Petri Nummi, University of Helsinki, FinlandBruce Alexander Schulte, Western Kentucky University, United States
Copyright © 2022 Orazi, Hagge, Gossner, Müller and Heurich. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jonas Hagge, am9uYXNoYWdnZUBwb3N0ZW8uZGU=
†These authors have contributed equally to this work and share first authorship