 John P. Draper
John P. Draper Julie K. Young
Julie K. Young Eugene W. Schupp
Eugene W. Schupp Noelle G. Beckman
Noelle G. Beckman Trisha B. Atwood
Trisha B. Atwood
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Ecol. Evol., 28 March 2022
Sec. Population, Community, and Ecosystem Dynamics
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.864864
This article is part of the Research TopicSeed Behavior in Response to Extreme EnvironmentsView all 7 articles
Seed dispersal is critical to the ecological performance of sexually reproducing plant species and the communities that they form. The Mammalian order Carnivora provide valuable and effective seed dispersal services but tend to be overlooked in much of the seed dispersal literature. Here we review the literature on the role of Carnivorans in seed dispersal, with a literature search in the Scopus reference database. Overall, we found that Carnivorans are prolific seed dispersers. Carnivorans’ diverse and plastic diets allow them to consume large volumes of over a hundred families of fruit and disperse large quantities of seeds across landscapes. Gut passage by these taxa generally has a neutral effect on seed viability. While the overall effect of Carnivorans on seed dispersal quality is complex, Carnivorans likely increase long-distance dispersal services that may aid the ability of some plant species to persist in the face of climate change.
Seed dispersal is a crucial process that allows populations of sexually reproducing plants to persist locally, improve fitness and gene flow, and shift or expand their geographic ranges (Howe and Smallwood, 1982; Levin et al., 2003). Understanding the dispersal mechanisms in sexually reproducing plants is, therefore, crucial for understanding current plant distributions and community composition, as well as predicting future distributions and compositions (Harper, 1977; Howe and Smallwood, 1982). Natural selection has favored the evolution of fleshy fruits or fleshy accessory structures (hereon referred to as fruits), in turn these fruits encourage the consumption of a plant’s seeds by animals who then disperse the seeds providing a fitness benefit (Tiffney, 1984; Tiffney, 2004; Eriksson, 2016), a process called endozoochory. The pattern of dispersal via endozoochory is influenced by how seed traits interact with disperser traits. Furthermore, the behaviors, life-history traits, and physiology of Carnivora make them distinct from other dispersers. For example, Carnivora often maintain large home ranges (i.e., ursids, Graham and Stenhouse, 2014), gut passage times are long (Escribano-Avila et al., 2014; Draper et al., 2021) which will affect the patterns of seedfall across landscapes and the consequences of those patterns in ways that differ from other frugivores (Schupp and Fuentes, 1995; Jordano et al., 2007; Martínez et al., 2008). Differences in seed disperser gut-passage time and post-consumption movement can influence dispersal distances, while their habitat use and defecation patterns can influence micro-site characteristics and degree of seed clumping that will influence plant recruitment. Therefore, it is important to understand the full complement of seed dispersers on the landscape. In this review, we discuss the unique role of an often-overlooked group of endozoochoric seed dispersers, species in the order Carnivora.
To date, most research on seed dispersal via endozoochory has focused on taxa outside the order Carnivora, primarily birds and other mammals such as primates and bats. For example, a Scopus search on January 5, 2022 of seed disp* AND (bird OR mammal OR carniv*) returned 3,486 studies, of which only 164 focused on taxa in the order Carnivora. This focus on other taxa is in part because of the colloquial view that most Carnivorans are largely meat-eaters, leading to the assumption that they likely play only an indirect and incidental role in seed dispersal through their effects on their frugivorous prey (e.g., Hämäläinen et al., 2017). However, the order Carnivora is a hyperdiverse group of animals that includes 13 families of land dwellers whose diet’s range from nearly obligate meat-eaters (e.g., species in the family Felidae) to omnivores (e.g., black bears, Ursus americanus) to strict herbivores (e.g., pandas, Ailuropoda melanoleuca, Atwood et al., 2020). Although seed dispersal studies have concentrated on other taxa, there is growing evidence that several species of Carnivorans are effective and important seed dispersers (Willson, 1993; Enders and Vander Wall, 2012; Spennemann, 2019). Furthermore, the behaviors, life-history traits, and ecologies of Carnivora make them distinct from other dispersers such as birds and other mammals (e.g., Escribano-Avila et al., 2014; Selwyn et al., 2020). Thus, to obtain a more complete understanding of plant distributions and population and community dynamics across space and time, we must understand Carnivorans role as seed dispersers.
In this paper, we review the role Carnivorans play in seed dispersal. First, we examine the prevalence of observed and potential endozoochoric seed dispersal by species in the order Carnivora. Second, we examine the importance of Carnivoran seed dispersal under two frameworks: seed dispersal effectiveness (SDE) and long-distance dispersal (LDD). The SDE framework measures effective seed dispersal as an interaction of quality (viable seeds, suitable deposition locations etc.) and quantity (the number of seeds dispersed), while LDD is defined as dispersal beyond the seed source stand or beyond the genetic neighborhood that produced the seed (Schupp, 1993; Schupp et al., 2010; Jordano, 2017).
We conducted a review of the literature using the Scopus reference database (November 2020). Our search used 396 pairs of search terms that paired frugivory/seed dispersal terms with a Carnivoran terms including Latin names and common names (Supplementary Table 1). This returned a total of 1,746 papers; 1,406 papers were removed due to an irrelevant double meaning of a search term or a lack of peer review. An additional 62 papers were removed that looked exclusively at seed predation such as the consumption of acorns and other hard mast, or diplochory of seeds in the stomach of prey. Ultimately, 278 papers met our complete search criteria (Supplementary Table 2). In reviewing this literature, we found that Carnivorans are effective and prolific seed dispersers across the globe.
We found that frugivory and seed dispersal are common among taxa in the order Carnivora, occurring in at least 10 families (Table 1 and Supplementary Table 2). Research noting frugivory and seed dispersal by members of the family Canidae (e.g., coyotes, foxes, and wolves) were the most common (33% of studies), while another 28% of studies focused on members of the family Mustelidae (e.g., martens, Martes). Studies on frugivory in Viverridae (e.g., civets, Civettictis), Ursidae (e.g., bears, Ursus), and Procyonidae (e.g., raccoons, Procyon) were also prevalent in the literature, while Mephitidae (e.g., skunks, Mephitis), Herpestidae (e.g., mongoose, Herpestes), Hyaenidae (e.g., hyenas, Hyaena), Ailuridae (red panda, Ailurus), and Nandiniidae (African palm civet, Nandinia) were represented by four or fewer studies each. The limited number of frugivory studies on these particular groups of Carnivora does not necessarily mean that they play only a minor role in seed dispersal, but instead could reflect research or publication biases. However, based on their ecology, some Carnivorans are unlikely to consume fruits intentionally. For example, no studies identified frugivory or seed dispersal in members of Felidae (cats), Prionodontidae (lisang), or Eupleridae (fossa), which are known to consume little to no plant parts (Atwood et al., 2020). These three taxa also lack a functional sweet taste, reducing the likelihood that they would be attracted to consuming fruits (Jiang et al., 2012). Diet studies on Carnivorans also often group all plant parts together or include plant parts in the diet category “other,” likely underestimating the extent of Carnivoran frugivory. Thus, consumption of fruits and dispersal of seeds by Carnivorans is surely higher than documented in the literature.
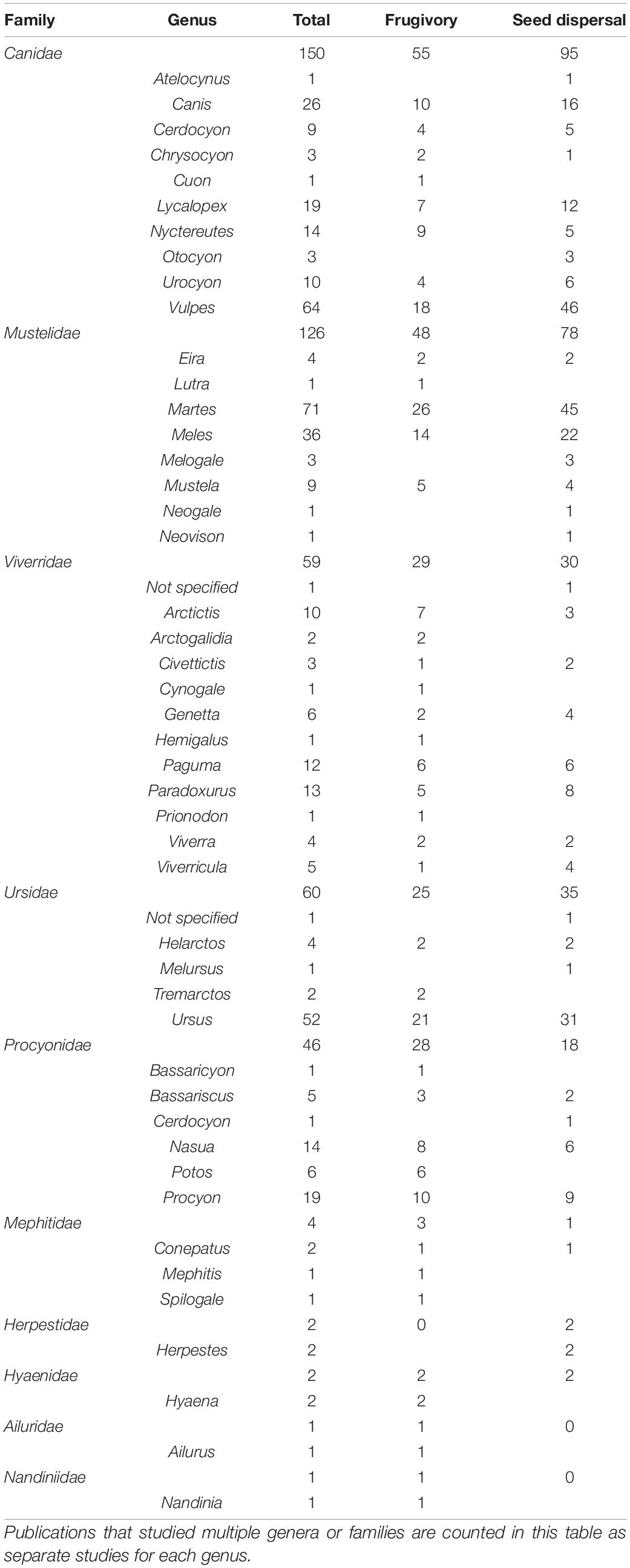
Table 1. Breakdown of the distribution of studies we found in our literature search described in the introduction by family and genus, and whether those studies looked at seed dispersal or frugivory in each taxonomic group.
Many species of Carnivora are opportunistic foragers that switch diets depending on prey availability and vulnerability (Chavez-Ramirez and Slack, 1993; Santos et al., 2003; Takahashi et al., 2008; Zhou et al., 2008). This diet plasticity allows many Carnivora to increase their consumption of fruits when they are readily available or during times when other prey are scarce (Silva et al., 2005; Takahashi et al., 2008). Notably, the diets of some species of Carnivora, such as coyotes and bears, can be dominated by fruits at certain times of the year (Dumond et al., 2001; Takahashi et al., 2008). Additionally, Carnivoran frugivory is influenced by the diverse physical capabilities of taxa within the order. Species capable of climbing commonly consume fruits directly from trees (Viverridae, Ursidae, Mustelidae, Mudappa et al., 2010; García-Rangel, 2012; Takatsuki et al., 2018), while other species are dependent on fruits falling to the ground (Canidae, Mustelidae, Herrera, 1989; Grünewald et al., 2010) or consuming fruits from low bushes that are accessible from the ground (Ursidae, Belant et al., 2010; Harrer and Levi, 2018). This diversity of foraging strategies within and across Carnivoran families is demonstrated in the diversity of plant species and fruit types they consume. We found documentation of Carnivorans consuming the pomes, drupes, berries, arils, or dry cones of at least 118 families of plants, with Rosaceae (e.g., blackberries), Moraceae (e.g., mulberry), and Ericaceae (e.g., heather) being the most prevalent in the literature (Supplementary Table 3). However, we did not identify any plant species that were solely dependent on seed dispersal by a Carnivoran. We also found that Carnivora exploit species of fruits with which they share a long history of co-occurrence, as well as novel fruits (Cypher and Cypher, 1999; Mudappa et al., 2010; Roehm and Moran, 2013). The consumption and dispersal of seeds from familiar and novel fruits suggest that Carnivora could increase the spread of fruit-bearing invasive plant species (Spennemann, 2019), as well as alter the dispersal patterns of native fruit-bearing plants encountered by novel Carnivoran species that are experiencing range expansion (e.g., coyotes, Hody and Kays, 2018). This is particularly important given that defaunation will limit plants’ abilities to track a suitable niche in the changing climate (Fricke et al., 2022); therefore the natural introduction of a novel generalist seed disperser may re-establish climate tracking for some plants.
The geographic locations of studies exploring frugivory in the order Carnivora suggest that they are prolific seed dispersers across the globe (Figure 1). Frugivory or seed dispersal by at least one Carnivoran species has been documented on every continent, except Antarctica. Most studies on Carnivoran frugivory or seed dispersal have taken place in North America, South America, Europe, and East Asia (e.g., Japan and parts of Malaysia), with fewer studies on Carnivoran frugivory or seed dispersal in Africa, Oceania, and western Asia. The lack of studies in Oceania likely stem from there being few native Carnivorans in this region, although two studies from Australia suggest that the introduction of Carnivorans to new regions could result in new mutualistic relationships with native (or invasive) plants (Brunner et al., 1976; O’Connor et al., 2020). The few studies in Africa and Central/Western Asia are somewhat surprising considering that IUCN’s Red List shows 119 and 53 species of Carnivora, respectively across these regions (IUCN, 2021). As a result, the geographic prevalence of Carnivora seed dispersal could be much greater than the literature currently suggests, especially in Africa and parts of Asia.
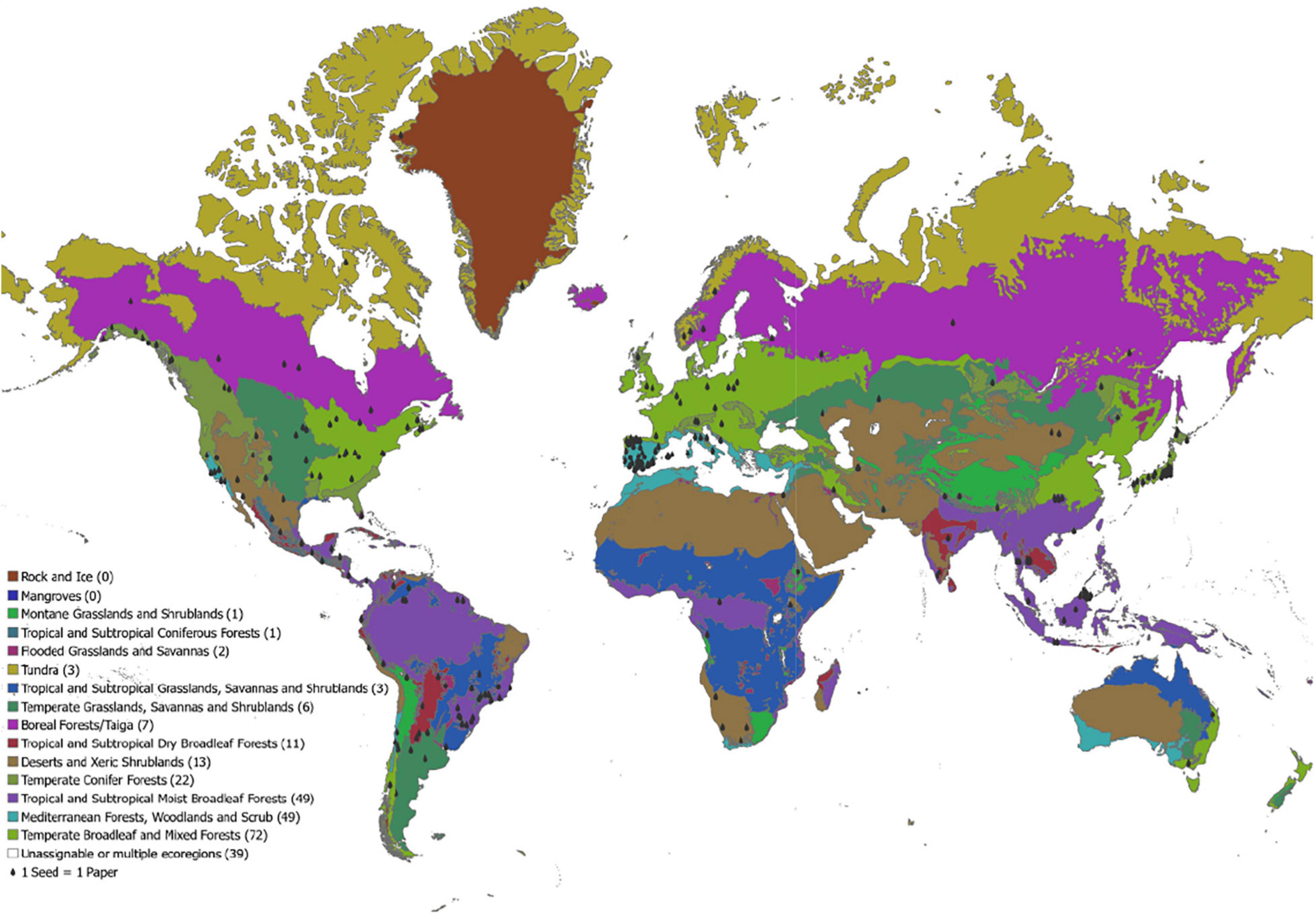
Figure 1. We conducted a literature search as outlined in the introduction. The search yielded 280 peer-reviewed publications that observed or discussed Carnivoran consumption of fruits and seeds. Each seed on this map represents a study that explored or discussed Carnivoran frugivory or seed dispersal. The map is divided by ecoregion as compiled by The Nature Conservancy (Wiken, 1986; Bailey, 1995; Olson and Dinerstein, 2002), included in the legend is the total number of studies identified in that ecoregion. We were unable to assign 39 studies to a specific ecoregion because the studies addressed frugivory and seed dispersal at a wide spatial scale that crossed multiple ecoregions or used species of captive animals that crossed multiple ecoregions in their wild ranges.
In addition to being globally widespread, Carnivoran frugivory or seed dispersal has been documented in every terrestrial ecoregion (Wiken, 1986; Bailey, 1995; Olson and Dinerstein, 2002), except mangroves and rock/ice (Figure 1). Most studies have been conducted in temperate, tropical, and subtropical regions. In particular, there is a greater number of studies focusing on Carnivoran frugivory or seed dispersal in temperate/broadleaf mixed forests, Mediterranean forests/woodlands/scrubs (mostly in Spain), and tropical and subtropical moist broadleaf ecoregions. In general, the number of studies investigating Carnivoran frugivory was lowest in polar regions. Fewer studies in higher latitude systems could reflect a reduction in the prevalence of omnivory with increasing latitudes, or a reduction in the number of Carnivoran-fruiting plant combinations that could be studied due to reduced species diversity at the poles (Hillebrand, 2004). Regardless, the global expanse of documented Carnivoran frugivory combined with the taxonomic breadth of the plant species consumed suggests that if the seeds consumed by members of Carnivorans are viable, then Carnivora are likely prevalent and important seed dispersers throughout the globe.
Two distinct yet overlapping concepts can help guide our thinking about the consequences of seed dispersal by Carnivorans: SDE and LDD events. These frameworks can be integrated with demographic models (Beckman et al., 2020) to predict how seed dispersal by Carnivorans influences the persistence and spread of plant populations.
Seed dispersal effectiveness has been defined as the contribution a seed disperser makes to the fitness of a plant it disperses. While this is ideally measured in terms of the recruitment of new reproductive adults to the population, empirical studies generally quantify disperser contributions to the performance of some earlier life-history stage, such as successful seedling establishment, rather than new adults (Schupp, 1993; Schupp et al., 2010; Schupp et al., 2017). SDE has a quantity component and a quality component, where SDE = quantity × quality. Quantity is the number of seeds dispersed and quality is the probability that a dispersed seed successfully produces a new adult. In terms of the quality of seed dispersal by Carnivorans, we highlight: (1) the treatment in the mouth and gut, (2) the local distance dispersed, and (3) the seedscape, that is, the local environmental factors that influence recruitment after seed deposition (Beckman and Rogers, 2013).
When thinking about the consequences of seed dispersal by Carnivorans, the quantity of seed dispersal is straightforward – it is simply the number of seeds dispersed and Carnivorans generally consume large quantities of fruit per feeding (Harrer and Levi, 2018; Shakeri et al., 2018). However, quantity is also influenced by the total number of visits by a disperser, and we know much less about this. Carnivoran visitation can be facilitated when other herbivores and frugivores improve the accessibility of fruits by removing thorny vegetative parts or knocking fruits off of trees, thus increasing dispersal quantity (Selwyn et al., 2020). Quality, however, is more complex and can be affected by a variety of outcomes of the Carnivoran-plant interaction as discussed next.
When frugivores feed on fruits there is a continuum of outcomes, from all seeds being treated gently and dispersed physically intact to all being destroyed by chewing or by gut passage itself (Schupp, 1993). For seeds that do pass through the gut intact, seed germinability may be reduced or increased and germination timing may be altered (Traveset, 1998). Gut passage may increase seed germination by scarifying seed coats either chemically or mechanically, potentially reducing physiological or physical seed dormancy (Traveset et al., 2007; Soltani et al., 2018). Furthermore, fecal matter can protect seeds from predators or pathogens and provide fertilizer (Traveset et al., 2007). Consequently, what happens during gut passage can have negative, positive, or neutral consequences for the quality component of SDE.
Consumption of fruits by Carnivorans typically results in intact seeds being passed through the digestive system and deposited in scat (Herrera, 1989; Perea et al., 2013). Destruction of seeds by chewing generally occurs when the seeds are ground in the mouth. However, even highly frugivorous Carnivoran species like bears (Harrer and Levi, 2018) have a dentition that is optimized for a diverse omnivorous diet (Elbroch, 2006). As a result, the molars in Carnivorans are specialized for crushing (i.e., bones and hard mast) rather than grinding (Elbroch, 2006), making the breaking of seeds, especially smaller seeds, less likely in Carnivorans (Koike et al., 2008; Lalleroni et al., 2017). Furthermore, fruits are often consumed whole and processed in the stomach of Carnivorans, allowing for the passing of even large seeds intact (e.g., Prunus; Rogers and Applegate, 1983). Every study within our literature search that quantified the proportion of seeds that were passed found that greater than 90% of seeds were passed intact (Supplementary Table 4). These studies included members of the families Canidae, Mustelidae, Ursidae, and Viverridae, with no clear variation in seed treatment by different Carnivoran families. Studies that included dry fruited or nut species accounted for the highest proportion of broken seeds, while most seeds from fleshy fruited species were defecated with greater than 98% of seeds intact (Supplementary Table 4).
Although most Carnivorans appear to pass seeds intact, the effect of gut passage on seed viability, germination, and dormancy is less straightforward. Carnivoran gut passage of intact seeds generally shows a neutral effect on viability; 70.6% of studies (defined as each seed-Carnivoran pair as some papers explored multiple interactions) showed that viability was maintained at the same proportion as undigested seeds, and more than 167 additional studies showed that seeds were viable after dispersal however without a control comparison to evaluate proportional changes in seed viability (Supplementary Table 4). In some cases, gut-passage can help break seed dormancy or alter germination timing, with examples showing both accelerated and delayed germination as illustrated in the distribution of studies in our review (49% accelerating, 13.7% delaying, and 37.3% resulting in no change in germination timing Supplementary Table 4). However, the effect of gut passage on seed viability and germination can be species-pair-specific (Traveset and Willson, 1997; Rubalcava-Castillo et al., 2021), with the same seed species being affected differently by different Carnivoran species, and vice-a-versa. Altering germination timing and proportions has the potential to create a mismatch between germination and favorable conditions for recruitment while depleting viable seeds from the seed bank and decreasing future recruitment (Traveset et al., 2007; Roehm and Moran, 2013). Alternatively, these alterations in germination timing can also lead to beneficial alignment between germination and recruitment conditions resulting in higher overall recruitment (Traveset et al., 2007). Therefore, changes in germination timing are not necessarily positive or negative, but context-dependent depending on plant species, ecoregion, and past and future climate and weather. Studies to date that look at viability and germination are heavily biased toward Canidae and Ursidae. The variability in seed responses and low representation of Carnivoran taxa studied necessitates further research on Carnivoran seed dispersal to elucidate broad and consistent trends.
The local distance seeds are dispersed from the parent can have a large effect on the quality of dispersal and overall SDE. Dispersal away from the parent plant frequently increases survival and successful recruitment through escape from distance- and density-dependent seed and seedling enemies that concentrate attack beneath and near adult conspecifics (e.g., Janzen, 1970; Connell, 1971; Howe et al., 1985; Schupp, 1988; Comita et al., 2014). Often, the advantage is not simply getting away from the parent, but rather the advantage steadily increases with increasing distance from the parent or other conspecific adults (Howe et al., 1985) provided seeds are deposited in a suitable microsite for germination and establishment. How far a frugivore disperses seeds is dependent on gut-passage time and the distance and directionality of travel by the disperser.
Despite the importance of gut-passage time for understanding the capabilities of an effective seed disperser, few studies have investigated gut-passage time in Carnivorans (Table 2). In our limited dataset, average gut-passage times for Carnivorans that have consumed fruit ranged between 2.5 and 18 h (Table 2), with maximum gut-passage times exceeding 24 h for several Carnivoran species. Of the species studied, Japanese marten (Martes melampus) had the shortest average gut-passage times of 2.5 h, and Asiatic black bear (Ursus thibetanus) had the longest average gut-passage time of 18 h (Koike et al., 2011; Table 2). Gut-passage times roughly scale with body size (p-value = 0.01, r-squared = 0.54, Figure 2). However, one factor that might affect gut-passage times in individual Carnivorans is diet composition; diets containing more fruit likely have shorter gut-passage times. For example, one study found that an increase in fruit in brown bear (Ursus arctos) diets reduced the average gut-passage time from 14.5 to 6.5 h (Elfström et al., 2013). Conversely, there was no effect of the proportion of fruit in the diet on gut-passage times for coyotes (Draper et al., 2021). Though gut-passage time may be altered when consuming fruit for some species (both decreasing and increasing gut-passage time, Cipollini and Levey, 1997), passage times remain on the order of hours suggesting that regardless of how much fruit was consumed, Carnivorans physiology can accommodate long dispersal distances.
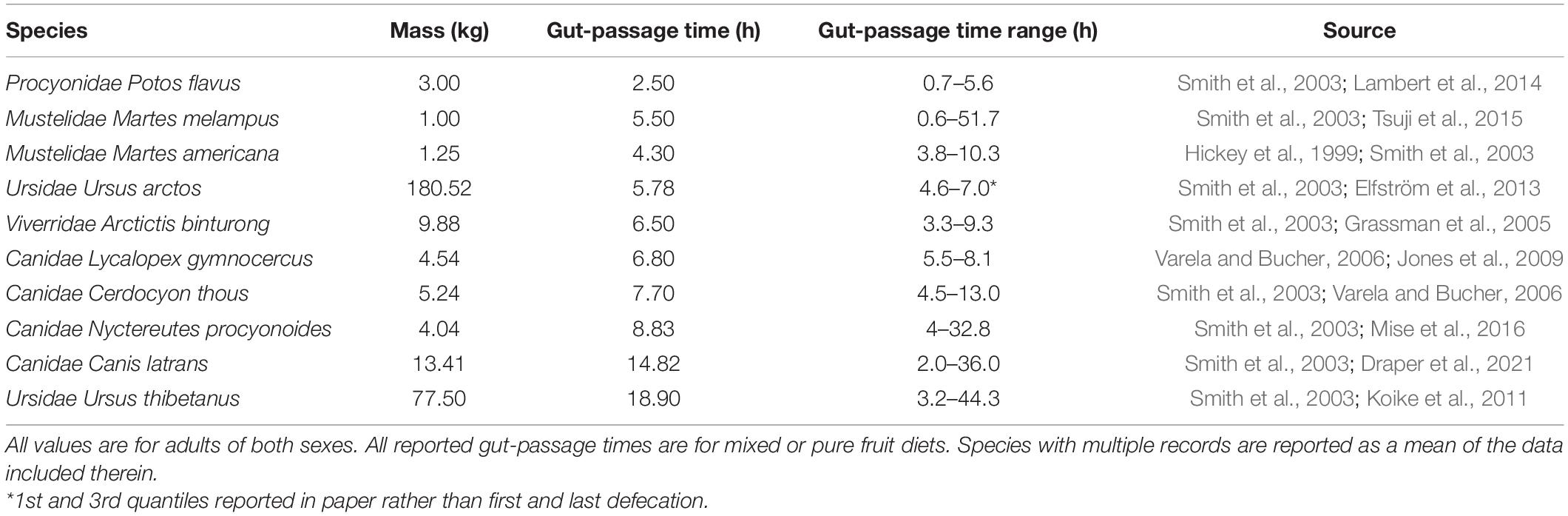
Table 2. Animal body mass, average gut-passage time, and the range of gut-passage time of Carnivoran species identified as frugivores/seed dispersers.
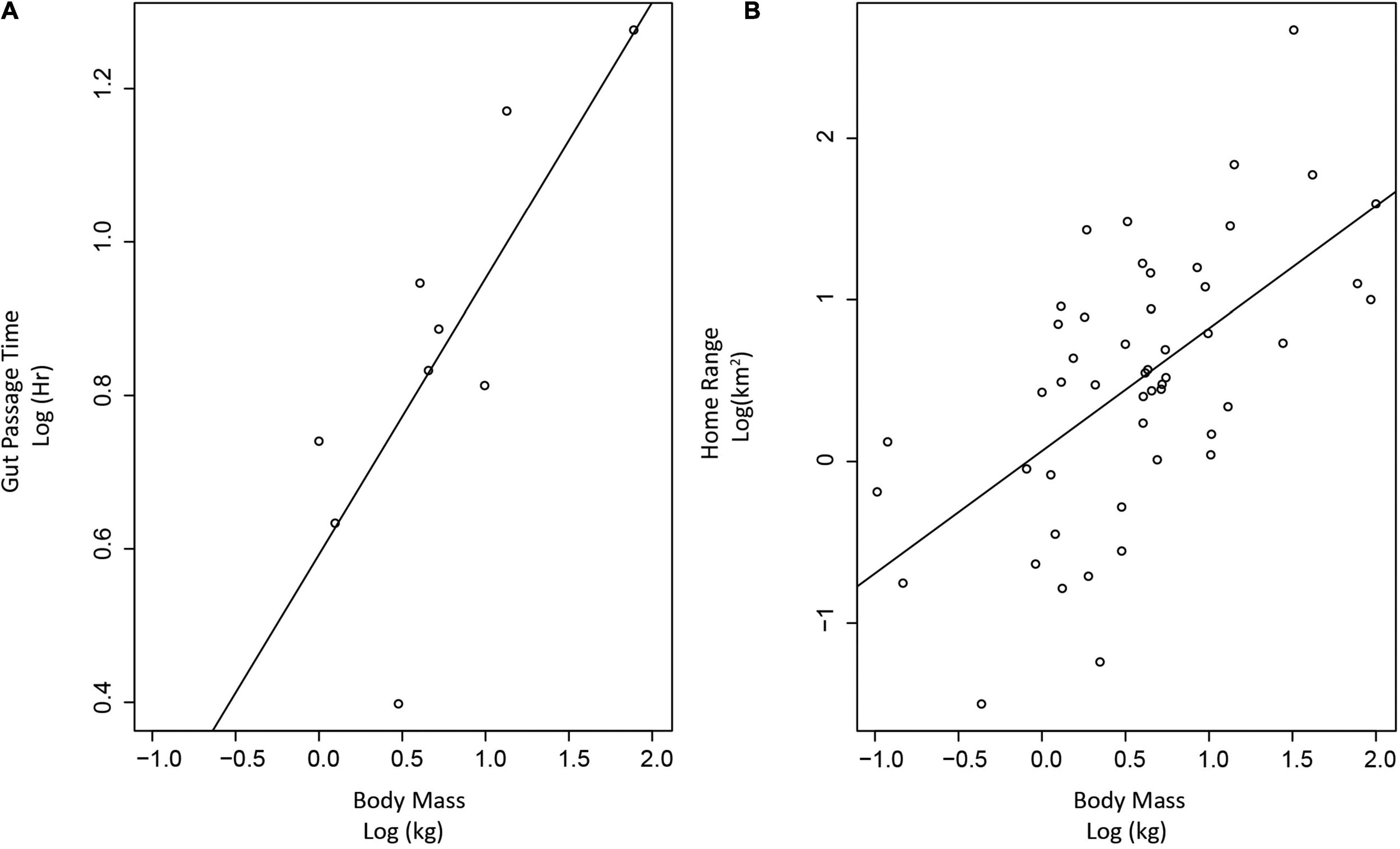
Figure 2. Relationships between gut-passage time, home range, and body mass in Carnivorans. (A) Linear regression between the log body mass (kg) and log gut-passage time (hours) of 10 Carnivoran species (p-value = 0.01, r-squared = 0.54). (B) Linear regression between the log mass (kg) and the log home range (km2) of 51 Carnivoran species (p-value < 0.001, r-squared value = 0.35). Gut-passage time, body mass, and home range values were derived from the available literature for species identified in our literature search as frugivorous Carnivorans (Supplementary Table 5).
Seed dispersal distances provided by Carnivorans have been shown to correlate with the maximum movement possible within the animal’s home range (González-Varo et al., 2013). Movements by Carnivorans are often constrained to an individual’s home range because many species are territorial (Gese and Ruff, 1997; Graham and Stenhouse, 2014). Carnivorans travel extensively within their home range to maximize access to resources, avoid risks from intraspecific and intraguild aggression (Hertel et al., 2019), and engage in territorial marking and patrolling (Gese and Ruff, 1997; Graham and Stenhouse, 2014). While there can be a wide variation in home range size within a species, daily total movement tends to stay the same across a population, with animals with smaller home ranges utilizing the entire space daily, and those with larger home ranges traversing it over multiple days with similar daily travel distances (Goszczyński, 2002). Similar to gut-passage time, home range size also increases with Carnivoran body size (p-value < 0.001, r-squared value = 0.35; Figure 2 and Supplementary Table 5). Therefore, we expect Carnivoran seed dispersal to also scale with body size. Considering that the body size of documented fruit-eating Carnivorans spans three orders of magnitude, from the least weasel (Mustela nivalis; 0.10 kg) to the brown bear (Ursus arctos; 180.5 kg), we would expect that Carnivorans express a multitude of different dispersal distances. Thus, Carnivorans are likely diverse in their abilities to disperse seeds away from the parent plant and other conspecifics that may compete with seedings.
Beyond removal from the vicinity of maternal plants, an additional aspect of local dispersal services is the ecological conditions into which seeds are deposited. Carnivorans deposit seeds in different habitats than many other seed dispersers, providing seed dispersal that is complimentary to other dispersers that results in a more diverse seedscape (Beckman and Rogers, 2013). Carnivoran behavior such as patrolling and territorial marking can lead to the deposition of seed-laden scat along travel corridors (Suárez-Esteban et al., 2013; Rubalcava-Castillo et al., 2020) and in open habitats either as a marking or coincidentally as they pass through (Gese and Ruff, 1997; Rost et al., 2012; Fedriani et al., 2018). In contrast, other endozoochoric dispersers (e.g., passerines) and abiotic dispersal generally result in seed deposition within contiguous canopy cover with high fruit occurrence (Jordano and Schupp, 2000; Herrera and García, 2010; Escribano-Avila et al., 2014). The deposition of seeds in travel corridors and open habitats provides seed dispersal to sites that are often either in completely novel habitat or along habitat edges, complementing other dispersal vectors. Furthermore, Carnivorans increase dispersal into open habitats improving colonization and recolonization of disturbed habitats better than their avian counterparts (Escribano-Avila et al., 2014). The complimentary dispersal patterns provided by Carnivorans can improve the fitness of the plants that they disperse (Celedón-Neghme et al., 2013; Escribano-Ávila et al., 2013; Escribano-Avila et al., 2014). However, this diversity of deposition also has the potential to negatively affect the outcome for seeds that are ultimately dispersed to unsuitable habitats.
Carnivorans broadcast seeds widely at a macro scale (i.e., meters to kilometers, González-Varo et al., 2013) by traveling long distances before defecating seeds, but concentrate them at a micro scale (sub 1 m, Shakeri et al., 2018) by concentrating them in a single scat. These patterns can alter their effects on SDE quality (Schupp et al., 2002). The majority of studies in our literature search observing Carnivoran seed consumption and deposition record a large number of seeds per scat (e.g., Matías et al., 2010; Harrer and Levi, 2018; Shakeri et al., 2018). The concentration of many seeds into a single deposition can reduce the quality component of SDE. Seeds suspended within an intact scat have lower rates of emergence (Draper et al., 2021), so the extent to which secondary dispersers remove seeds and distribute them elsewhere will affect the ultimate quality of dispersal (Enders and Vander Wall, 2012). Concentrating seeds in a single location can also attract seed predators (which are often also secondary dispersers) and increase the spread of pathogens, countering some of the many benefits that dispersal provides (LoGiudice and Ostfeld, 2002; Shakeri et al., 2018). Furthermore, the release of many seeds in a single scat can increase the potential for future competition among seedlings and juvenile plants (Loiselle, 1990; Schupp et al., 2002). Some Carnivoran species in the families Canidae, Procyonidae, Nandiniidae, Mustelidae, and Viverridae can further compound the above effects on quality by depositing scats in spatially discrete latrines, potentially resulting in extremely large concentrations of seeds (Clevenger, 1996; Page et al., 1999; Page et al., 2001; Helbig-bonitz et al., 2013; Osugi et al., 2020). Conversely, a clumped rather than even distribution of seeds can result in higher rates of recruitment due to a lower ability of pathogens or predators to track vulnerable seeds (Beckman et al., 2012), and the fertilizing effect of scat can improve germination and recruitment conditions for seeds growing from scats and latrines, helping to overcome the negative effects of high seed and seedling density (O’Farrill et al., 2013; Sugiyama et al., 2018). The potential for both positive and negative effects of Carnivoran dispersal on SDE suggests that future studies should focus on monitoring the number of successful recruits to later life history stages, as opposed to the more common approaches that focus on seed viability and germination.
Long-distance dispersal affects plant genetic exchange and migration (Jordano, 2017). LDD occurs when a seed is moved beyond the geographic limits of its source stand or outside the genetic neighborhood of the source plant (Jordano, 2017). Carnivorans provide dispersal distances that move seeds beyond the threshold defining LDD for many plant species (Lalleroni et al., 2017; Spennemann, 2018; Pereira et al., 2019). With these LDD events Carnivorans aid in maintaining gene flow between disparate stands, improving genetic diversity and resiliency (Harper, 1977). Furthermore, by providing dispersal beyond the confines of a current plant stand, Carnivorans can aid in range shifts to track suitable climate conditions or recolonize disturbed landscapes (Escribano-Avila et al., 2014; Naoe et al., 2016). This will have the most profound effect in areas where Carnivoran species are repatriating former ranges (Draper et al., 2017) and re-establishing LDD services or where they are colonizing new territory and introducing a new LDD service (Hody and Kays, 2018).
The geographic, taxonomic, and ecological breadth of Carnivoran frugivory and seed dispersal supports the hypothesis that Carnivorans are common and likely important seed dispersers worldwide. Given their sizes and gut capacities, many Carnivorans can consume large quantities of fruits and seeds in a feeding bout and thus have the potential to be quantitatively important seed dispersers. However, at this point we lack a full understanding of the quantitative component of SDE provided by Carnivorans due to a paucity of data on the number of visits made to individual plants. Considering treatment in the mouth and gut, Carnivorans commonly pass fruit seeds intact and generally have a neutral effect on seed viability, while gut passage generally has neutral or positive effects on germination, indicating high quality treatment. When seeds are deposited, the scat itself provides fertilizer and protection from some seed predators and pathogens, clearly positive consequences of Carnivoran seed dispersal (Traveset et al., 2007; O’Farrill et al., 2013). In contrast, the concentration of seeds in Carnivoran scat and the latrine behavior of many Carnivorans has the potential to increase density-dependent seed predation (Shakeri et al., 2018) and increase conspecific and heterospecific competition among seedlings and juvenile plants. The long dispersal distances Carnivorans can provide potentially improves seed and seedling survival in the face of distance-dependent enemies. However, the biotic and abiotic characteristics of the sites where seeds are deposited often have a greater impact than distance on the quality of dispersal and this has been poorly addressed. Overall, although there is clear evidence that Carnivoran seed dispersal has the potential to be quantitatively and qualitatively effective, there are substantial gaps in our knowledge of both. Consequently, the overall SDE provided by Carnivorans is uncertain at this time.
The wide range of potential dispersal distances among different Carnivorans combined with other biotic and abiotic dispersal vectors adds to the portfolio of strategies that plants can employ to distribute their seeds across a landscape (Escribano-Avila et al., 2014). Relatively long gut-passage times combined with the potential to move long distances suggest that Carnivorans can provide regular LDD services. While this may sometimes carry seeds outside of quality germination conditions, it can also provide them the potential for range expansion and connect disparate populations of the same species to help maintain genetic diversity (Levine and Murrell, 2003). As a result, LDD by Carnivorans could play an important role in the ability of certain plant species to persist in the face of climate change (Kremer et al., 2012; Corlett and Westcott, 2013).
Perhaps the most intriguing and important aspect of Carnivoran seed dispersal is its potential complementarity with avian seed dispersal. Birds generally defecate or regurgitate seeds while perched, depositing seeds beneath existing vegetation (Jordano and Schupp, 2000; Herrera and García, 2010). By contrast, many Carnivorans defecate and deposit seeds along travel routes and in open habitats, resulting in very different seed shadows than those produced by birds (Suárez-Esteban et al., 2013; Rubalcava-Castillo et al., 2020). Although deposition in open habitats is unlikely to be advantageous for all species, such a pattern of seed dispersal can promote more rapid range expansion and recolonization of disturbed habitats for those species able to recruit in these conditions (Escribano-Avila et al., 2014).
All authors listed have made a substantial, direct, and intellectual contribution to the work, and approved it for publication.
This research was funded by a Department of Watershed Sciences Ph.D. Scholarship, a Utah State University (USU) Ecology Center Scholarship, and the USDA-National Wildlife Research Center. TA was funded by an Early Career Research Fellowship from the Gulf Research Program of the National Academies of Sciences, Engineering, and Medicine. ES was supported by project number 1523 of the Utah Agricultural Experiment Station, USU.
The content is solely the responsibility of the authors and does not necessarily represent the official views of the Gulf Research Program of the National Academies of Sciences, Engineering, and Medicine or the USDA.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We would like to thank E. Hammill and K. Kettenring for comments on an early version of this manuscript.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.864864/full#supplementary-material
Atwood, T. B., Valentine, S. A., Hammill, E., McCauley, D. J., Madin, E. M. P., Beard, K. H., et al. (2020). Herbivores at the Highest Risk of Extinction Among Mammals, Birds, and Reptiles. Sci. Adv. 6:eabb8458. doi: 10.1126/sciadv.abb8458
Bailey, R. G. (1995). Description of the Ecoregions of the United States. United States: United States Forest Service.
Beckman, N. G., Aslan, C. E., Rogers, H. S., Kogan, O., Bronstein, J. L., Bullock, J. M., et al. (2020). Advancing an Interdisciplinary Framework to Study Seed Dispersal Ecology. AoB Plants 12, 1–18. doi: 10.1093/aobpla/plz048
Beckman, N. G., Neuhauser, C., and Muller-Landau, H. C. (2012). The Interacting Effects of Clumped Seed Dispersal and Distance and Density-Dependent Mortality on Seedling Recruitment Patterns. J. Ecol. 100, 862–873. doi: 10.1111/j.1365-2745.2012.01978.x
Beckman, N. G., and Rogers, H. S. (2013). Consequences of Seed Dispersal for Plant Recruitment in Tropical Forests: interactions Within the Seedscape. Biotropica 45, 666–681. doi: 10.1111/btp.12071
Belant, J. L., Griffith, B., Zhang, Y., Follmann, E. H., and Adams, L. G. (2010). Population-level resource selection by sympatric brown and American black bears in Alaska. Polar Biol. 33, 31–40. doi: 10.1007/s00300-009-0682-6
Brunner, H., Harris, R. V., and Amor, R. L. (1976). A note on the Dispersal of Seeds of Blackberry (Rubus procerus P.J. Muell.) by Foxes and Emus. Weed Res. 16, 171–173. doi: 10.1111/j.1365-3180.1976.tb00398.x
Celedón-Neghme, C., Traveset, A., and Calviño-Cancela, M. (2013). Contrasting Patterns of Seed Dispersal Between Alien Mammals and Native Lizards in a Declining Plant Species. Plant Ecol. 214, 657–667. doi: 10.1007/s11258-013-0197-7
Chavez-Ramirez, F., and Slack, R. D. (1993). Carnivore Fruit-Use and Seed Dispersal of Two Selected Plant Species of the Edwards Plateau. Texas. Southwest. Nat. 38:141. doi: 10.2307/3672066
Cipollini, M. L., and Levey, D. J. (1997). Secondary Metabolites of Fleshy Vertebrate-Dispersed Fruits: adaptive Hypotheses and Implications for Seed Dispersal. Am. Nat. 150, 346–372. doi: 10.1086/286069
Clevenger, A. P. (1996). Frugivory of Martes martes and Genetta genetta in an Insular Mediterranean Habitat. Rev. Ecol. 51, 19–28.
Comita, L. S., Queenborough, S. A., Murphy, S. J., Eck, J. L., Xu, K., Krishnadas, M., et al. (2014). Testing Predictions of the Janzen-Connell Hypothesis: a Meta-analysis of Experimental Evidence for Distance- and Density-dependent Seed and Seedling Survival. J. Ecol. 102, 845–856. doi: 10.1111/1365-2745.12232
Connell, J. (1971). On the Role of Natural Enemies in Preventing Competitive Exclusion in Some Marine Animals and in Rain Forest Trees. Dyn. Popul. 298, 298–312.
Corlett, R. T., and Westcott, D. A. (2013). Will Plant Movements Keep Up with Climate Change? Trends Ecol. Evol. 28, 482–488. doi: 10.1016/j.tree.2013.04.003
Cypher, B. L., and Cypher, E. A. (1999). Germination Rates of Tree Seeds Ingested by Coyotes and Raccoons. Am. Midl. Nat. 142, 71–76. doi: 10.1674/0003-0031(1999)142[0071:grotsi]2.0.co;2
Draper, J. P., Atwood, T. B., Beckman, N. G., Kettenring, K. M., and Young, J. K. (2021). Mesopredator Frugivory Has No Effect on Seed Viability and Emergence Under Experimental Conditions. Ecosphere 12:e03702. doi: 10.1002/ecs2.3702
Draper, J. P., Waits, L. P., Adams, J. R., Seals, C. L., and Steury, T. D. (2017). Genetic health and population monitoring of two small black bear (Ursus americanus) populations in Alabama, with a regional perspective of genetic diversity and exchange. PLoS One 12:e0186701. doi: 10.1371/journal.pone.0186701
Dumond, M., Villard, M. A., and Tremblay, É (2001). Does Coyote Diet Vary Seasonally Between a Protected and an Unprotected Forest Landscape? Ecoscience 8, 301–310. doi: 10.1080/11956860.2001.11682657
Elbroch, M. (2006). Animal Skulls: a Guide to North American Species. Mechanicsburg: Stackpole Books.
Elfström, M., Støen, O. G., Zedrosser, A., Warrington, I., and Swenson, J. E. (2013). Gut Retention Time in Captive Brown Bears Ursus arctos. Wildlife Biol. 19, 317–324. doi: 10.2981/12-121
Enders, M. S., and Vander Wall, S. B. (2012). Black Bears Ursus americanus Are Effective Seed Dispersers, With a Little Help From Their Friends. Oikos 121, 589–596. doi: 10.1111/j.1600-0706.2011.19710.x
Eriksson, O. (2016). Evolution of angiosperm seed disperser mutualisms: the timing of origins and their consequences for coevolutionary interactions between angiosperms and frugivores. Biol. Rev. 91, 168–186. doi: 10.1111/brv.12164
Escribano-Avila, G., Calviño-Cancela, M., Pías, B., Virgós, E., Valladares, F., and Escudero, A. (2014). Diverse Guilds Provide Complementary Dispersal Services in a Woodland Expansion Process After Land Abandonment. J. Appl. Ecol. 51, 1701–1711. doi: 10.1111/1365-2664.12340
Escribano-Ávila, G., Pías, B., Sanz-Pérez, V., Virgós, E., Escudero, A., and Valladares, F. (2013). Spanish Juniper Gain Expansion Opportunities by Counting on a Functionally Diverse Dispersal Assemblage Community. Ecol. Evol. 3, 3751–3763. doi: 10.1002/ece3.753
Fedriani, J. M., Wiegand, T., Ayllón, D., Palomares, F., Suárez-Esteban, A., and Grimm, V. (2018). Assisting Seed Dispersers to Restore Oldfields: an Individual-Based Model of the Interactions Among Badgers, Foxes and Iberian Pear Trees. J. Appl. Ecol. 55, 600–611. doi: 10.1111/1365-2664.13000
Fricke, E. C., Ordonez, A., Rogers, H. S., and Svenning, J. (2022). The Effects of Defaunation on Plants’ Capacity to Track Climate Change. Science 375, 210–214. doi: 10.1126/science.abk3510
García-Rangel, S. (2012). Andean bear Tremarctos ornatus natural history and conservation. Mamm. Rev. 42, 85–119. doi: 10.1111/j.1365-2907.2011.00207.x
Gese, E. M., and Ruff, R. L. (1997). Scent-Marking by Coyotes, Canis latrans: the Influence of Social and Ecological Factors. Anim. Behav. 54, 1155–1166. doi: 10.1006/anbe.1997.0561
González-Varo, J. P., López-Bao, J. V., and Guitián, J. (2013). Functional Diversity Among Seed Dispersal Kernels Generated by Carnivorous Mammals. J. Anim. Ecol. 82, 562–571. doi: 10.1111/1365-2656.12024
Goszczyński, J. (2002). Home Ranges in Red Fox: territoriality Diminishes With Increasing Area. Acta Theriol. 47, 103–114. doi: 10.1007/bf03192482
Graham, K., and Stenhouse, G. B. (2014). Home Range, Movements, and Denning Chronology of the Grizzly Bear (Ursus arctos) In West-Central Alberta. Can. Field Nat. 128, 223–234. doi: 10.22621/cfn.v128i3.1600
Grassman, L. I., Tewes, M. E., and Silvy, N. J. (2005). Ranging, Habitat Use and Activity Patterns of Binturong Arctictis Binturong and Yellow-Throated Marten Martes flavigula in North-Central Thailand. Wildlife Biol. 11, 49–57. doi: 10.2981/0909-6396(2005)11[49:rhuaap]2.0.co;2
Grünewald, C., Breitbach, N., and Böhning-Gaese, K. (2010). Tree Visitation and Seed Dispersal of Wild Cherries by Terrestrial Mammals Along a Human Land-Use Gradient. Basic Appl. Ecol. 11, 532–541. doi: 10.1016/j.baae.2010.07.007
Hämäläinen, A., Broadley, K., Droghini, A., Haines, J. A., Lamb, C. T., Boutin, S., et al. (2017). The Ecological Significance of Secondary Seed Dispersal by Carnivores. Ecosphere 8:e01685. doi: 10.1002/ecs2.1685
Harrer, L. E. F., and Levi, T. (2018). The Primacy of Bears as Seed Dispersers in Salmon-Bearing Ecosystems. Ecosphere 9:e02076. doi: 10.1002/ecs2.2076
Helbig-bonitz, M., Rutten, G., and Kalko, K. V. (2013). Tree Seed Dispersal by African Civets in the Afromontane Highlands: too Long a Latrine to Be Effective for Tree Population Dynamics. Afr. J. Ecol. 52, 122–125. doi: 10.1111/aje.12090
Herrera, C. M. (1989). Frugivory and Seed Dispersal by Carnivorous Mammals, and Associated Fruit Characteristics, in Undisturbed Mediterranean Habitats. Oikos 55, 250–262. doi: 10.2307/3565429
Herrera, J. M., and García, D. (2010). Effects of Forest Fragmentation on Seed Dispersal and Seedling Establishment in Ornithochorous Trees. Conserv. Biol. 24, 1089–1098. doi: 10.1111/j.1523-1739.2010.01459.x
Hertel, A. G., Leclerc, M., Warren, D., Pelletier, F., Zedrosser, A., and Mueller, T. (2019). Don’t Poke the Bear: using Tracking Data to Quantify Behavioural Syndromes in Elusive Wildlife. Anim. Behav. 147, 91–104. doi: 10.1016/j.anbehav.2018.11.008
Hickey, J. R., Flynn, R. W., Buskirk, S. W., Gerow, K. G., and Willson, M. F. (1999). An Evaluation of a Mammalian Predator, Martes americana, as a Disperser of Seeds. Oikos 87:499. doi: 10.2307/3546814
Hillebrand, H. (2004). On the Generality of the Latitudinal Diversity Gradient. Am. Nat. 163, 192–211. doi: 10.1086/381004
Hody, J. W., and Kays, R. (2018). Mapping the Expansion of Coyotes (Canis latrans) Across North and Central America. Zookeys 759, 81–97. doi: 10.3897/zookeys.759.15149
Howe, H. F., Schupp, E. W., and Westley, L. C. (1985). Early Consequences of Seed Dispersal for a Neotropical Tree (Virola surinamensis). Ecology 66, 781–791. doi: 10.2307/1940539
IUCN (2021). The IUCN Red List of Threatened Species. Version 2021-3. Available online at: https://www.iucnredlist.org (accessed October 15, 2021).
Janzen, D. H. (1970). Herbivores and the Number of Tree Species in Tropical Forests. Am. Nat. 104, 501–528. doi: 10.1086/282687
Jiang, P., Josue, J., Li, X., Glaser, D., Li, W., Brand, J. G., et al. (2012). Major taste loss in carnivorous mammals. Proc. Natl. Acad. Sci. U. S. A. 109, 4956–4961. doi: 10.1073/pnas.1118360109
Jones, K. E., Bielby, J., Cardillo, M., Fritz, S. A., O’Dell, J., Orme, C. D. L., et al. (2009). PanTHERIA: a Species-Level Database of Life History, Ecology, and Geography of Extant and Recently Extinct Mammals. Ecology 90, 2648–2648. doi: 10.1890/08-1494.1
Jordano, P. (2017). What Is Long-Distance Dispersal? And a Taxonomy of Dispersal Events. J. Ecol. 105, 75–84. doi: 10.1111/1365-2745.12690
Jordano, P., Garcia, C., Godoy, J. A., Garcia-Castano, J. L., and García-Castaño, J. L. (2007). Differential Contribution of Frugivores to Complex Seed Dispersal Patterns. Proc. Natl. Acad. Sci. U. S. A. 104, 3278–3282. doi: 10.1073/pnas.0606793104
Jordano, P., and Schupp, E. W. (2000). Seed Disperser Effectiveness: the Quantity Component and Patterns of Seed Rain for Prunus mahaleb. Ecol. Monogr. 70, 591–615. doi: 10.2307/2657187
Koike, S., Masaki, T., Nemoto, Y., Kozakai, C., Yamazaki, K., Kasai, S., et al. (2011). Estimate of the Seed Shadow Created by the Asiatic Black Bear Ursus thibetanus and Its Characteristics as a Seed Disperser in Japanese Cool-Temperate Forest. Oikos 120, 280–290. doi: 10.1111/j.1600-0706.2010.18626.x
Koike, S., Morimoto, H., Goto, Y., Kozakai, C., and Yamazaki, K. (2008). Frugivory of Carnivores and Seed Dispersal of Fleshy Fruits in Cool-Temperate Deciduous Forests. J. For. Res. 13, 215–222. doi: 10.1007/s10310-008-0069-5
Kremer, A., Ronce, O., Robledo-Arnuncio, J. J., Guillaume, F., Bohrer, G., Nathan, R., et al. (2012). Long-Distance Gene Flow and Adaptation of Forest Trees to Rapid Climate Change. Ecol. Lett. 15, 378–392. doi: 10.1111/j.1461-0248.2012.01746.x
Lalleroni, A., Quenette, P. Y., Daufresne, T., Pellerin, M., and Baltzinger, C. (2017). Exploring the Potential of Brown Bear (Ursus arctos arctos) As a Long-Distance Seed Disperser: a Pilot Study in South-Western Europe. Mammalia 81, 1–9. doi: 10.1515/mammalia-2015-0092
Lambert, J. E., Fellner, V., McKenney, E., and Hartstone-Rose, A. (2014). Binturong (Arctictis binturong) And Kinkajou (Potos flavus) Digestive Strategy: implications for Interpreting Frugivory in Carnivora and Primates. PLoS One 9:e105415. doi: 10.1371/journal.pone.0105415
Levin, S. A., Muller-Landau, H. C., Nathan, R., and Chave, J. (2003). The Ecology and Evolution of Seed Dispersal: a Theoretical Perspective. Annu. Rev. Ecol. Evol. Syst. 34, 575–604. doi: 10.1146/annurev.ecolsys.34.011802.132428
Levine, J. M., and Murrell, D. J. (2003). The Community-Level Consequences of Seed Dispersal Patterns. Annu. Rev. Ecol. Evol. Syst. 34, 549–574. doi: 10.1146/annurev.ecolsys.34.011802.132400
LoGiudice, K., and Ostfeld, R. (2002). Interactions between mammals and trees: predation on mammal-dispersed seeds and the effect of ambient food. Oecologia 130, 420–425. doi: 10.1007/s004420100810
Loiselle, B. A. (1990). Seeds in Droppings of Tropical Fruit-Eating Birds: importance of Considering Seed Composition. Oecologia 82, 494–500. doi: 10.1007/BF00319792
Martínez, I., García, D., and Obeso, J. R. (2008). Differential Seed Dispersal Patterns Generated by a Common Assemblage of Vertebrate Frugivores in Three Fleshy-Fruited Trees. Ecoscience 15, 189–199. doi: 10.2980/15-2-3096
Matías, L., Zamora, R., Mendoza, I., and Hódar, J. A. (2010). Seed Dispersal Patterns by Large Frugivorous Mammals in a Degraded Mosaic Landscape. Restor. Ecol. 18, 619–627. doi: 10.1111/j.1526-100X.2008.00475.x
Mise, Y., Yamazaki, K., Soga, M., and Koike, S. (2016). Comparing Methods of Acquiring Mammalian Endozoochorous Seed Dispersal Distance Distributions. Ecol. Res. 31, 881–889. doi: 10.1007/s11284-016-1397-0
Mudappa, D., Kumar, A., and Chellam, R. (2010). Diet and Fruit Choice of the Brown Palm Civet Paradoxurus jerdoni, a Viverrid Endemic to the Western Ghats Rainforest. India. Trop. Conserv. Sci. 3, 282–300. doi: 10.1177/194008291000300304
Naoe, S., Tayasu, I., Sakai, Y., Masaki, T., Kobayashi, K., Nakajima, A., et al. (2016). Mountain-Climbing Bears Protect Cherry Species From Global Warming Through Vertical Seed Dispersal. Curr. Biol. 26, R315–R316. doi: 10.1016/j.cub.2016.03.002
O’Connor, J. M., Burrows, D. M., Allen, B. L., and Burnett, S. E. (2020). Is the European Red Fox a Vector of the Invasive Basket Asparagus (Asparagus aethiopicus) In Eastern Australia? Aust. Mammal. 42, 204–210. doi: 10.1071/AM19001
O’Farrill, G., Galetti, M., and Campos-Arceiz, A. (2013). Frugivory and Seed Dispersal by Tapirs: an Insight on Their Ecological Role. Integr. Zool. 8, 4–17. doi: 10.1111/j.1749-4877.2012.00316.x
Olson, D. M., and Dinerstein, E. (2002). The Global 200: priority Ecoregions for Global Conservation. Ann. Miss. Bot. Gard. 89, 199–224. doi: 10.2307/3298564
Osugi, S., Trentin, B. E., and Koike, S. (2020). What Determines the Seedling Viability of Different Tree Species in Raccoon Dog Latrines? Acta Oecol. 106:103604. doi: 10.1016/j.actao.2020.103604
Page, L. K., Swihart, R. K., and Kazacos, K. R. (1999). Implications of Raccoon Latrines in the Epizootiology of Baylisascariasis. J. Wildl. Dis. 35, 474–480. doi: 10.7589/0090-3558-35.3.474
Page, L. K., Swihart, R. K., and Kazacos, K. R. (2001). Seed Preferences and Foraging by Granivores at Raccoon Latrines in the Transmission Dynamics of the Raccoon Roundworm (Baylisascaris procyonis). Can. J. Zool. 79:4.
Perea, R., Delibes, M., Polko, M., Suárez-Esteban, A., and Fedriani, J. M. (2013). Context-Dependent Fruit-Frugivore Interactions: partner Identities and Spatio-Temporal Variations. Oikos 122, 943–951. doi: 10.1111/j.1600-0706.2012.20940.x
Pereira, L., Vasques, A., Maia, P., Ramos Pereira, M. J., Fonseca, C., and Matos, M. (2019). Native and Exotic Seed Dispersal by the Stone Marten (Martes foina): implications for the Regeneration of a Relict Climactic Forest in Central Portugal. Integr. Zool. 14, 280–292. doi: 10.1111/1749-4877.12362
Roehm, K., and Moran, M. D. (2013). Is the Coyote (Canis latrans) A Potential Seed Disperser for the American Persimmon (Diospyros virginiana)? Am. Midl. Nat. 169, 416–421. doi: 10.1674/0003-0031-169.2.416
Rogers, L. L., and Applegate, R. D. (1983). Dispersal of Fruit Seeds by Black Bears. J. Mammal. 64, 310–311. doi: 10.2307/1380564
Rost, J., Pons, P., and Bas, J. M. (2012). Seed Dispersal by Carnivorous Mammals Into Burnt Forests: an Opportunity for Non-indigenous and Cultivated Plant Species. Basic Appl. Ecol. 13, 623–630. doi: 10.1016/j.baae.2012.08.016
Rubalcava-Castillo, F. A., Sosa-Ramírez, J., Luna-Ruíz, J. J., Valdivia-Flores, A. G., Díaz-Núñez, V., and Íñiguez-Dávalos, L. I. (2020). Endozoochorous Dispersal of Forest Seeds by Carnivorous Mammals in Sierra Fría. Aguascalientes, Mexico. Ecol. Evol. 10, 2991–3003. doi: 10.1002/ece3.6113
Rubalcava-Castillo, F. A., Sosa-Ramírez, J., Luna-Ruíz, J. D. J., Valdivia-Flores, A. G., and Íñiguez-Dávalos, L. I. (2021). Seed Dispersal by Carnivores in Temperate and Tropical Dry Forests. Ecol. Evol. 11, 3794–3807. doi: 10.1002/ece3.7201
Santos, E. F., Setz, E. Z. F., and Gobbi, N. (2003). Diet of the Maned Wolf (Chrysocyon brachyurus) And Its Role in Seed Dispersal on a Cattle Ranch in Brazil. J. Zool. 260, 203–208. doi: 10.1017/S0952836903003650
Schupp, E. W. (1988). Seed and Early Seedling Predation in the Forest Understory and in Treefall Gaps. Oikos 51, 71–78. doi: 10.2307/3565808
Schupp, E. W. (1993). Quantity, Quality and the Effectiveness of Seed Dispersal. Vegetatio 107, 15–29. doi: 10.1007/978-94-011-1749-4_2
Schupp, E. W., and Fuentes, M. (1995). Spatial Patterns of Seed Dispersal and the Unification of Plant Population Ecology. Ecoscience 2, 267–275. doi: 10.1080/11956860.1995.11682293
Schupp, E. W., Jordano, P., and Gómez, J. M. (2010). Seed Dispersal Effectiveness Revisited: a Conceptual Review. New Phytol. 188, 333–353. doi: 10.1111/j.1136-8137.2010.03402.x
Schupp, E. W., Jordano, P., and Gómez, J. M. (2017). A General Framework for Effectiveness Concepts in Mutualisms. Ecol. Lett. 20, 577–590. doi: 10.1111/ele.12764
Schupp, E. W. E., Milleron, T., and Russo, S. S. E. (2002). ““Dissemination Limitation and the Origin and Maintenance of Species-Rich Tropical Forests” in Seed Dispersal and Frugivory: ecology, Evolution Andconservation. eds D. Levey, W. Silva, and M. Galetti (Wallingford: CABI). 19–33. doi: 10.1079/9780851995250.0019
Selwyn, M., Garrote, P. J., Castilla, A. R., and Fedriani, J. M. (2020). Interspecific Interactions Among Functionally Diverse Frugivores and Their Outcomes for Plant Reproduction: a New Approach Based on Camera-Trap Data and Tailored Null Models. PLoS One 15:e0240614. doi: 10.1371/journal.pone.0240614
Shakeri, Y. N., White, K. S., and Levi, T. (2018). Salmon-Supported Bears, Seed Dispersal, and Extensive Resource Subsidies to Granivores. Ecosphere 9, 1–13. doi: 10.1002/ecs2.2297
Silva, S. I., Bozinovic, F., and Jaksic, F. M. (2005). Frugivory and Seed Dispersal by Foxes in Relation to Mammalian Prey Abundance in a Semiarid Thornscrub. Austral Ecol. 30, 739–746. doi: 10.1111/j.1442-9993.2005.01449.x
Smith, F. A., Lyons, S. K., Ernest, S. K. M., Jones, K. E., Kaufman, D. M., Dayan, T., et al. (2003). Body Mass of Late Quaternary Mammals. Ecology 84, 3403–3403. doi: 10.1890/02-9003
Soltani, E., Baskin, C. C., Baskin, J. M., Heshmati, S., and Mirfazeli, M. S. (2018). A Meta-Analysis of the Effects of Frugivory (Endozoochory) On Seed Germination: role of Seed Size and Kind of Dormancy. Plant Ecol. 219, 1283–1294. doi: 10.1007/s11258-018-0878-3
Spennemann, D. H. R. (2018). Review of the Vertebrate-Mediated Dispersal of the Date Palm, Phoenix dactylifera. Zool. Middle East 64, 283–296. doi: 10.1080/09397140.2018.1514785
Spennemann, D. H. R. (2019). The Connective Potential of Vertebrate Vectors Responsible for the Dispersal of the Canary Island Date Palm (Phoenix canariensis). Flora Morphol. Distrib. Funct. Ecol. Plants 259:151468. doi: 10.1016/j.flora.2019.151468
Suárez-Esteban, A., Delibes, M., and Fedriani, J. M. (2013). Barriers or Corridors? The Overlooked Role of Unpaved Roads in Endozoochorous Seed Dispersal. J. Appl. Ecol. 50, 767–774. doi: 10.1111/1365-2664.12080
Sugiyama, A., Comita, L. S., Masaki, T., Condit, R., and Hubbell, S. P. (2018). Resolving the Paradox of Clumped Seed Dispersal: positive Density and Distance Dependence in a Bat-Dispersed Species. Ecology 99, 2583–2591. doi: 10.1002/ecy.2512
Takahashi, K., Shiota, T., Tamatani, H., Koyama, M., Washitani, I., Takahashi, N. K., et al. (2008). Seasonal Variation in Fleshy Fruit Use and Seed Dispersal by the Japanese Black Bear (Ursus thibetanus japonicus). Ecol. Res. 23, 471–478. doi: 10.1007/s11284-007-0382-z
Takatsuki, S., Miyaoka, R., and Sugaya, K. (2018). A Comparison of Food Habits between Japanese Marten and Raccoon Dog in Western Tokyo with Reference to Fruit Use. Zoolog. Sci. 35, 68–74. doi: 10.2108/zs170116
Tiffney, B. H. (1984). Seed Size, Dispersal Syndromes, and the Rise of the Angiosperms: evidence and Hypothesis. Ann. Missouri Bot. Gard. 71, 551–576.
Tiffney, B. H. (2004). Vertebrate dispersal of seed plants through time. Annu. Rev. Ecol. Evol. Syst. 35, 1–29. doi: 10.1146/ANNUREV.ECOLSYS.34.011802.132535
Traveset, A. (1998). Effect of Seed Passage Through Vertebrate Frugivores’ Guts on Germination: a Review. Perspect. Plant Ecol. Evol. Syst. 1, 151–190. doi: 10.1078/1433-8319-00057
Traveset, A., Robertson, A. W., and Rodríguez-Pérez, J. (2007). “A Review on the Role of Endozoochory in Seed Germination” in Seed Dispersal: theory and its Application in a Changing World. eds A. J. Dennis, R. J. Green, E. W. Schupp, and D. A. Westcott (Wallingford: CABI). 78–103. doi: 10.1079/9781845931650.0078
Traveset, A., and Willson, M. F. (1997). Effect of Birds and Bears on Seed Germination of Fleshy-Fruited Plants in Temperate Rainforests of Southeast Alaska. Oikos 80:89. doi: 10.2307/3546519
Tsuji, Y., Miura, S., Kotoge, T., Shiraishi, T., and Murai, H. (2015). Effects of Food Intake on Digesta Passage Time in Captive Japanese Martens (Martes melampus) and Implications for Endozoochorous Seed Dispersal. Mamm. Study 40, 13–18. doi: 10.3106/041.040.0103
Varela, O., and Bucher, E. H. (2006). Passage Time, Viability, and Germination of Seeds Ingested by Foxes. J. Arid Environ. 67, 566–578. doi: 10.1016/j.jaridenv.2006.03.013
Willson, M. F. (1993). Mammals as Seed Dispersal Mutualist in North America. Oikos 67, 159–176. doi: 10.2307/3545106
Keywords: carnivore, seed dispersal, carnivoran, frugivory, endozoochory
Citation: Draper JP, Young JK, Schupp EW, Beckman NG and Atwood TB (2022) Frugivory and Seed Dispersal by Carnivorans. Front. Ecol. Evol. 10:864864. doi: 10.3389/fevo.2022.864864
Received: 28 January 2022; Accepted: 07 March 2022;
Published: 28 March 2022.
Edited by:
Mathew Samuel Crowther, The University of Sydney, AustraliaReviewed by:
Lisieux Fuzessy, Ecological and Forestry Applications Research Center (CREAF), SpainCopyright © 2022 Draper, Young, Schupp, Beckman and Atwood. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: John P. Draper, am9obi5kcmFwZXJAdXN1LmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.