 Andreas Koenig
Andreas Koenig Angela Miles1
Angela Miles1 Durr-E-Ajam Riaz
Durr-E-Ajam Riaz Carola Borries
Carola Borries
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 16 June 2022
Sec. Behavioral and Evolutionary Ecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.860437
This article is part of the Research TopicSex and Gender Effects on Power, Status, Dominance, and Leadership – An Interdisciplinary Look at Human and Other Mammalian SocietiesView all 22 articles
Male-female agonism varies throughout the primate order with males often dominating females, especially in sexually dimorphic species. While intersexual agonism has been attributed to sexually coercive contexts, it can also occur for other reasons and intersexual dominance may be influenced by the adult sex ratio. If the proportion of males is high, certain males will regularly lose against other males. Loser-effects may then pave the way for some females to dominate these males, an effect that has been described in a few primate species. Here we investigated the frequency, general style, and context of agonism among gray langurs (Semnopithecus spp.). Data were collected at two study sites, at Jodhpur, India (one group), and at Ramnagar, Nepal (two groups). The adult sex ratio varied between 0.077 and 1.000 males to females. At both sites, data on agonistic interactions (aggression and submission) were collected in continuous focal animal and ad libitum sampling techniques during 1,945 contact hours (including 1,220 focal animal hours, total). Although aggression intensity was low, high directional consistency and the rare occurrence of counteraggression suggested a despotic dominance style, a likely prerequisite for dominance effects based on adult sex ratio. Aggression by females against males was very rare and mainly occurred in the defense of offspring. We found little evidence for partial female dominance regardless of adult sex ratio. In a few cases in which a female had a higher dominance index than a male, she did not dominate this male in dyadic encounters. Agonism by males directed at females occurred mainly in a feeding context while male policing and a sexual context were both rare. The latter was mostly restricted to females after they had harassed a sexual interaction or after they had behaved proceptively toward a male. Our study suggests that across species the effect of adult sex ratio on female dominance might be more variable than previously suggested. The fact that most agonism between males and females occurred over food identifies intersexual feeding competition as a new research avenue with potentially important consequences for existing ideas on the costs and benefits of group life and composition.
Agonism among the members of group-living animals is often viewed as a sex-specific behavioral strategy. Male mammals are commonly seen as competing with other males for access to females (Darwin, 1871), and females are usually assumed to compete over access to food or safety or both (Wrangham, 1980; van Schaik, 1989). While the reality is much more complex (Clutton-Brock, 2021), these ideas form the basis of so-called socio-ecological models that relate to the number of males in a group and relationships among males (Emlen and Oring, 1977; Carnes et al., 2011; Koenig et al., 2013; Ostner and Schuelke, 2014) as well as to the links between ecology and female social relationships (Wrangham, 1980; van Schaik, 1989; Isbell, 1991; Sterck et al., 1997; Koenig, 2002; Koenig et al., 2013). Over the years, however, it has become clear that these models do not reflect the existing variation in individual behavioral strategies and emerging social systems (van Schaik, 1996; Clutton-Brock and Janson, 2012). Importantly, when focusing on same-sex interactions, the interactions between members of both sexes and their possible influence on the social structure are not being addressed (Kappeler, 2017). As more results have accumulated, it has become particularly evident that male dominance over females and male aggression against females varies tremendously across non-human primates (Hemelrijk et al., 2008; Muller et al., 2009), but its potential impact on social structure and competition in general has remained largely unexplored. While intersexual relationships are certainly also affected by benefits that males and females can provide to each other when residing in a group together, e.g., through predator detection, during intergroup encounters, or as coalitionary support (e.g., Rose, 1994; Perry, 1997; Ostner et al., 2013; Archie et al., 2014), in the following we will focus on intersexual agonism and dominance.
Based on general fighting ability and in the absence of female-female coalitions, it is typically assumed that in sexually dimorphic primate species, males dominate females (Smuts, 1987). While this may be true for a number of primate species, more and more studies have emerged in which females have been shown to exhibit frequent aggression against males or to even dominate some or several males in their group (Smuts, 1987; Hemelrijk et al., 2020; Izar et al., 2021). Generally, such partial female dominance has thus far only been documented for despotic species (despotic sensu de Waal and Luttrell, 1989; Preuschoft and van Schaik, 2000; Thierry, 2000; Flack and de Waal, 2004) and among other factors it seems to be related to the adult sex ratio of a study group (Hemelrijk et al., 2008; Kappeler, 2017). Females are suggested to dominate more males or a larger proportion of the males present in a group possibly because (i) more males can mean more contest competition among males, increasing winner-loser effects among males, allowing females to take advantage and dominate males who frequently lost and/or because of (ii) generally higher and more intense female aggression toward males in groups with more males (Hemelrijk et al., 2008, 2020; Stroebe et al., 2017; Izar et al., 2021). However, larger groups with more males can also mean increased agonism of males toward females (Kappeler, 2017), which begs the question if partial female dominance is possible under increased male aggression. Clearly, more studies on intersexual dominance are needed to examine if the proposed effects of adult sex ratio on partial female dominance in despotic species hold.
Apart from the questions of how and when intersexual dominance varies among non-human primates, it is equally important to determine what primate males may achieve through agonism toward females or conversely, what costs females are facing at the receiving end and what they could thus gain through intersexual dominance (Smuts, 1987; Smuts and Smuts, 1993; Muller et al., 2009). Starting with Smuts (1987) and Smuts and Smuts (1993) groundbreaking studies, the focus primarily has been on sexual coercion particularly sexual harassment and forced copulations (for a discussion of direct and indirect forms of sexual coercion see Muller et al., 2009). In recent years, this focus has been reinforced through detailed, long-term data for chacma baboons, hamadryas baboons, and chimpanzees (Muller et al., 2007, 2009, 2011; Swedell et al., 2014; Baniel et al., 2017; Watts, 2022). From a female perspective, partial female dominance may reduce sexual coercion and increase the extent of female mate choice (Smuts and Smuts, 1993; Hemelrijk et al., 2020). However, while sexual coercion can be common and can have important implications for female and male fitness, it is not the norm among primates (van Schaik et al., 2004). It is especially rare in lemuroids and platyrrhines and occurs in about 50% of catarrhines studied thus far, particularly in species that are prone to infanticide by males (van Schaik et al., 2004; following Muller et al., 2009, we consider infanticide as a third form of sexual coercion in which aggression is not directed at a female herself but her infant). To what extent this variation relates to partial female dominance is unclear at present, making studies of intersexual dominance in species with infanticide an important test case.
Nonetheless, male aggression against females is not solely related to sexual coercion. It has also been suggested to occur in four other general contexts: feeding competition, male policing (i.e., male intervention in female-female conflicts), status competition, and redirected aggression (Muller et al., 2009). While male agonism against females has been well documented in species with strong intergroup feeding competition, such as capuchins and chimpanzees (Muller et al., 2009; Scarry, 2013), only few detailed studies have addressed intersexual within-group feeding competition in primates. In fact, even fewer studies have reported the context of intersexual agonism (e.g., wedge-capped capuchins; O’Brien, 1991; see also Smuts and Smuts, 1993). In general, it can be expected that intersexual feeding competition may be more prominent in species relying on monopolizable food resources (Muller et al., 2009). Similarly, male policing occurs for example in mountain gorillas and Thomas langurs (overview in Watts et al., 2000), but it is unclear if male policing can explain most of the intersexual agonistic interactions in these or other species. Lastly, there is little evidence for status competition and redirected aggression as a major source of intersexual agonism (Muller et al., 2009). Given the major impact that sexual coercion can have on a female’s fitness, intersexual feeding competition may be the only other source of conflict that could have a strong impact. Judging by the effect of scramble and contest competition on female birth rates (Pusey, 2012), males contesting with females over access to food would heighten the already existing feeding competition among females. Consequently, intersexual agonism by females and partial female dominance over males would reduce both sexual coercion and competition over food for females. However, few studies have taken on such a functional perspective.
Here we studied intersexual agonism and dominance among gray langurs (Semnopithecus spp.; taxonomy following Arekar et al., 2021). Gray langurs are suitable subjects for the topic of intersexual agonism for several reasons. First, the study populations in Jodhpur, India (S. entellus) and in Ramnagar, Nepal (S. schistaceus) are characterized by despotic male-male and despotic female-female relationships because agonism is overall frequent and relationships are unidirectional and transitive (Borries et al., 1991; Borries, 1993; Koenig, 2000; Lu et al., 2008; Perlman et al., 2016; Sommer, pers. com.). This may make it likely that male-female relationships are despotic as well. If true, the entire social system would be despotic, a prerequisite for partial female dominance as described by Hemelrijk et al. (2008). Second, while female and male dominance hierarchies can be unstable, dominance has a significant impact on reproductive success of males and females in both populations (Sommer and Rajpurohit, 1989; Borries et al., 1991; Launhardt et al., 2001; de Vries et al., 2016). Thus, in addition to despotic social relationships (despotic sensu de Waal and Luttrell, 1989), reproductive skew is high (despotic sensu Vehrencamp, 1983). Third, females of both populations have been shown to compete primarily over monopolizable food resources (Borries, 1993; Koenig et al., 1998) with most agonism occurring over food (Koenig and Borries, 2006). Fourth, males vigorously fight over access to females (Sommer, 1988; Borries, 2000) and males in both populations regularly commit infanticide with more than 30% of infant mortality attributed to it (Borries and Koenig, 2000). Lastly, at Ramnagar a substantial amount of male-male agonism also occurred over food (a quarter to one-third of the conflicts; Koenig and Borries, unpublished).
Given the general characteristics of the study species and the predictions about intersexual agonism and dominance, in the current study we first tested if male-female agonistic relationships were despotic, a prerequisite for partial female dominance. If this would be the case then aggression would be rare, but of high intensity, directional consistency would be high, and counteraggression rare (de Waal and Luttrell, 1989; Preuschoft and van Schaik, 2000; Thierry, 2000; Flack and de Waal, 2004). Next, we examined how often and in what contexts females behaved aggressively toward males. Following Hemelrijk et al. (2008), we then analyzed dominance indices, expecting an effect of adult sex ratio on partial female dominance over males, i.e., females being more often dominant with increasing proportion of males in the group. Lastly, we analyzed the possible functions of male agonism toward females by determining how frequently the context of agonism was related to sexual coercion, feeding competition, or male policing (cf. Muller et al., 2009). We excluded two suggested contexts: (i) status competition because in gray langurs males leave their natal group as immatures and only adult males immigrate (Rajpurohit and Sommer, 1993; Borries, 2000), and (ii) redirected aggression by a male because it did not occur.
Data were collected on one group of wild, provisioned gray langurs (Semnopithecus entellus) near Jodhpur, India around lake Kailana (240 m a.s.l., 26°17’N, 72°58’E; Winkler, 1981) as well as two neighboring groups of wild, unprovisioned gray langurs (Semnopithecus schistaceus) living in a semi-evergreen forest near Ramnagar, Nepal (300 m a.s.l., 27°44’N, 84°27’E; Podzuweit, 1994). Both sites are rather seasonal in weather and food availability (Winkler, 1981; Koenig et al., 1997), although at Jodhpur the effect of seasonality is weakened because of provisioning and reproduction is seasonally restricted only at Ramnagar (Sommer and Rajpurohit, 1989; Koenig et al., 1997).
All individuals in the three groups differed in their stature, coat color, scars, broken fingers, broken tails, or other traits so that they could be distinguished individually. Additionally, at Ramnagar, each female had a hairless skin area ventral to the callosities between the thighs (pubic patch; e.g., Bernstein, 1968) which was uniquely dotted with depigmented (pink) spots (Podzuweit, 1994).
At Jodhpur, the Kailana I group (or B19; hereafter KI group) was regularly provisioned by local people to the extent that provisioned food amounted to one-third of the total feeding time (Winkler, 1984). Throughout the study period, the group had one adult male (who had been in the group for 19 months prior to the beginning of data collection) and 13 adult females resulting in an adult sex ratio of 0.077. No females matured during the study period and group size ranged from 18 to 23 individuals, because of births and infant deaths.
The two groups studied at Ramnagar, A group and O group, varied in size and composition during the study period, but A group was consistently the “small” group, at around 10 individuals in total (range 9–10), and O group the “big,” at approximately 30 individuals (range 26–34; population mean 18.3 individuals; Koenig, 2000). The A group consisted of 1, 2, or 3 adult males and 2 or 3 adult females (4–6 adult individuals in total). The number of adults varied because of emigration (2 adult males) and death (1 adult female) as well as immigration (1 adult female; the only immigration of an adult female ever observed in this population; Koenig et al., 1998) resulting in variable adult sex ratios [1.000 (3:3, 2:2), 0.667 (2:3), and 0.333 (1:3)]. O group consisted of 3 or 5 adult males and 15 adult females (18 or 20 adult individuals). The number of adults varied because two adult males immigrated and assumed the alpha and beta positions. The other three males remained as lower ranking males. Subsequently, three infants likely fell victim to infanticide (Borries, 1997) and one of the respective mothers also disappeared. At the same time, one subadult female entered the adult female hierarchy and was considered an adult. This resulted in two different adult sex ratios of 0.200 (3:15) and 0.333 (5:15) for O group.
At both sites, data were collected via ad libitum and focal animal sampling techniques (Martin and Bateson, 2007). Focal animal samples lasted for 30 min and combined continuous and instantaneous recording (at 30 s intervals). Within an observation period, focal individuals were sampled evenly across all hours of the day from dawn to dusk. As data collection involved the study of focal animals, data were not recorded blindly; however, the sequence of focal animals was randomized for each observation period.
At Jodhpur, CB collected behavioral data on adult females of KI group from January 07 to October 26, 1985 (inclusively) during three distinct periods lasting 45, 97, and 56 days, respectively (see Table I in Borries et al., 1991, p. 236). During the first two periods, eight females were focal animals, and during the last period, six females. The total contact time with KI group was 1,018.5 h of which 569.0 h were spent in focal animal sampling.
At Ramnagar, AK collected behavioral data on all adult individuals (males and females) in A group and O group from January 9, 1994 to January 23, 1995 (inclusively). In A group, data were collected over seven periods (mean 7.9 days/period) and in O group over four periods (mean: 26.3 days/period). Periods were distributed over the course of a year to capture seasonal changes in food availability as well as the mating and the birth season. The total contact time with A group was 257.0 h, of which 152.0 h were spent in focal animal sampling. In group O it was 669.3 h of contact time with 499.0 h of focal animal sampling.
Agonistic behavior was recorded during focal animal continuous sampling as well as via ad libitum sampling during the entire contact time with a group. We defined agonism as all aggression and submission. Following Rowell (1974), we included displacement within submissive behavior, i.e., being displaced: an individual A is giving up its place, food, social partner, etc. in favor of an approaching individual B or following aggression by individual B. Agonistic behaviors included in this study were recorded as behavioral acts, which have been described and defined elsewhere (Dolhinow, 1978; Sommer, 1985; Borries, 1989; see also Supplementary Table 1) and the same ethogram was used at both study locations. Behaviors considered here have previously been shown to occur in an agonistic context (Borries, 1989; Lu et al., 2008). Because this was not always clear for vocalizations, we did not include vocalizations in the analyses.
While recording agonistic behavior, the context was noted as well. The following contexts were distinguished: (i) food, i.e., agonism occurred while one or both individuals were feeding or one of the interactants was replaced at a feeding site or both; (ii) place, i.e., if agonism occurred at a specific location (e.g., regularly used to monitor or to rest) which was initially occupied by one individual and at the end of the interaction it was occupied by the other individual; (iii) after a sexual harassment, i.e., a sexual interaction was interrupted by a third individual usually via approaching and vocalizing, whereupon the male (not the female) of the pair showed agonism toward the harasser; (iv) sexual behavior, i.e., following sexual behavior (which could be mating or solicitation etc.), the male showed agonism toward the female and thus terminated the sexual interaction; (v) policing, i.e., aggressive behavior was directed at one of two individuals already engaged in an agonistic interaction; (vi) social, i.e., preceding an agonistic interaction, one individual was engaged in allogrooming or was in body contact or was close to a (any) group member; (vii) infant; i.e., a female fled, carrying her infant after having been approached by a male who reached for her infant; (viii) unknown; all remaining cases. These context definitions were applied very conservatively in the Jodhpur study, resulting in an overall lower percentage of identified contexts in comparison to the Ramnagar study.
In the analysis, we considered individual behavioral acts as well as bouts. A bout was defined as a temporal cluster of agonistic acts in an interaction between two (or more) individuals (see also Martin and Bateson, 2007). Bouts were considered decided if one individual won because the other individual gave up its place or item or signaled submission (Hausfater, 1975). Bouts were undecided if the interaction ended in a draw, e.g., no individual gave up its place or item after one or more agonistic acts (for coalitions see below). Because of a small sample size for females with infants, we did not distinguish whether a female had an infant clinging to her belly or not during an agonistic bout. For the analysis of rates of behavior, we used data exclusively from focal animal sampling, calculating the number of occurrences per focal observation hour. For all other analyses, we combined data from focal animal and ad libitum sampling.
In the results below, we first generally describe intersexual agonism regardless of the sex of the actor by listing the occurrence and rates of different types of agonism (aggression, submission). Because groups differed in size and composition, we describe the intersexual agonism per group.
In the analysis of despotism, we considered three criteria: intensity of aggression, directional consistency of all agonistic acts, and counteraggression (de Waal and Luttrell, 1989; Preuschoft and van Schaik, 2000; Thierry, 2000; Flack and de Waal, 2004). We categorized intensity of aggression following Lu et al. (2008; see Table 3 in Lu et al., 2008; see also Supplementary Table 1). To ease comparison with other studies, we also provided a breakdown of contact and non-contact aggression (Supplementary Table 1). Using the program MatMan (de Vries et al., 1993), we evaluated directional consistency by calculating the directional consistency index (DCI; van Hooff and Wensing, 1987) for all behavioral acts noted for intersexual agonistic interactions. If a single adult acted against more than one adult simultaneously, all recipients received a score. Counteraggression by females was defined as any aggressive act by a female in response to an aggression received by a male.
To investigate partial female dominance, we first examined how often and in which context females showed spontaneous aggression toward males. This was done to understand if aggression (and winning) by females is confined to certain contexts or is context independent, also referred to as “spheres of dominance” (Hand, 1986; Preuschoft and van Schaik, 2000). Furthermore, to study the proposed relationship between partial female dominance and adult sex ratio, we calculated dominance indices following Hemelrijk et al. (2005, 2008). Using only decided agonistic bouts, we first calculated the proportion of winning for each member of a dyad. If individuals were never observed to interact, the respective dyad was excluded from the analysis. To rank individuals, we used the average dominance index (ADI) for each individual, which was calculated as the average of all its dyadic dominance indices (Hemelrijk et al., 2005). Based on this ranking, we calculated the female dominance index (FDI; Hemelrijk et al., 2008). Here, each male with an ADI lower than a female was counted as one (dyad), and all dyads in which a male ranked below a female were summed up. In the case of a tie between a male and a female, 0.5 was added. The sum for all male-female dyads was then divided by the number of males each female theoretically could dominate summed over all females (i.e., the number of males multiplied by the number of females) to determine the female dominance index. This FDI runs from 0.0 (complete male dominance) to 1.0 (complete female dominance).
To investigate the context of male agonism against females, we considered bouts instead of individual acts, which prevents context inflation because a bout may involve several agonistic acts.
Intersexual agonism, i.e., agonism between males and females regardless of the actor, occurred at a rate of 0.074, 0.174, and 0.112 acts per focal hour for KI group, O group, and A group, respectively. It was primarily characterized by submission (50.3–88.2%; Table 1), which by itself was dominated by displacements (see Supplementary Table 1 for details). This was also reflected in the rates of agonistic acts. With 0.030, 0.044, and 0.013 acts per focal hour (for KI group, O group, and A group, respectively), rates of aggression were less frequent in all three groups compared to submission which occurred at 0.044, 0.130, and 0.100 acts per focal hour.
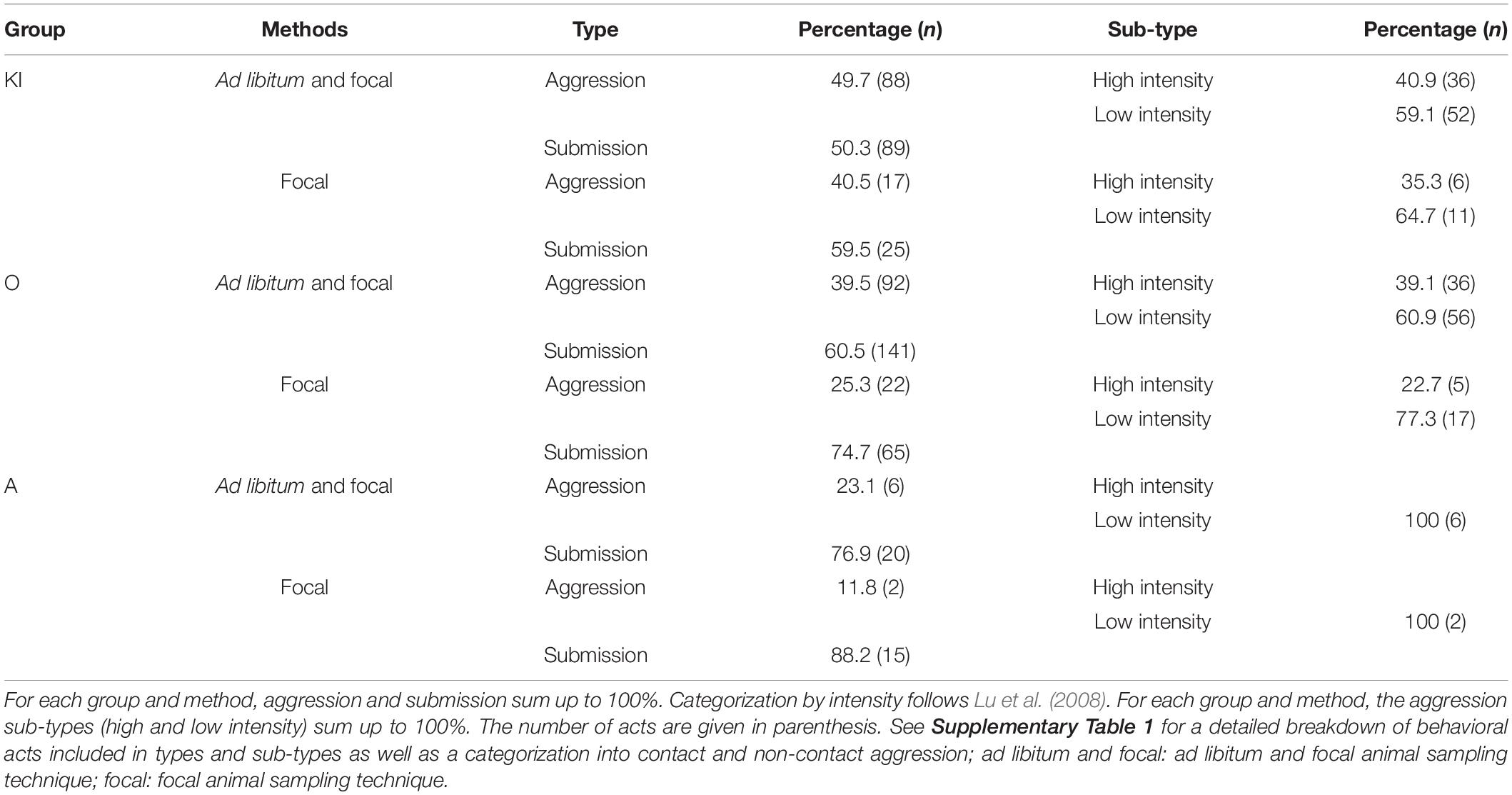
Table 1. Intersexual agonistic behavior and intensity of aggression in gray langurs independent of the sex of the actor.
Of all aggressive behaviors, low intensity ones were more frequent than high intensity ones (59.1–100.0%, Table 1). Unsurprisingly, the percentage of aggression, particularly of high intensity aggression, dropped when only focal animal samples were considered, and consequently the percentage for low intensity aggression and submission increased. This indicates that interactions of high intensity were discovered and thus recorded more frequently during ad libitum sampling (see also Martin and Bateson, 2007). This effect was larger at Ramnagar than at Jodhpur, likely, because of the poorer visibility at Ramnagar, which can bias toward the detection of high intensity behavior such as chases rather than the more subtle submission.
Aggression and submission varied widely across groups (Table 1). Aggression, and specifically high intensity aggression, was most frequent in the single male group KI (35.3–40.9%) but was lower than the percentage of low intensity aggression. In O group with 3 and 5 males, aggression accounted for 25.3–39.5% of the agonistic acts, but the percentage of high intensity aggression out of all aggression was low (under 40%). In the smallest A group with the most even adult sex ratio, aggression was rare, and none was of high intensity.
Agonistic interactions between males and females had a high directional consistency with DC indices between 1.000 and 0.796 (Table 2). The larger groups with a more skewed adult sex ratio and many more interactions (KI and O) had lower directional consistencies (0.796–0.948) than the small A group (DCI 1.000) with a more even sex ratio and much fewer interactions.
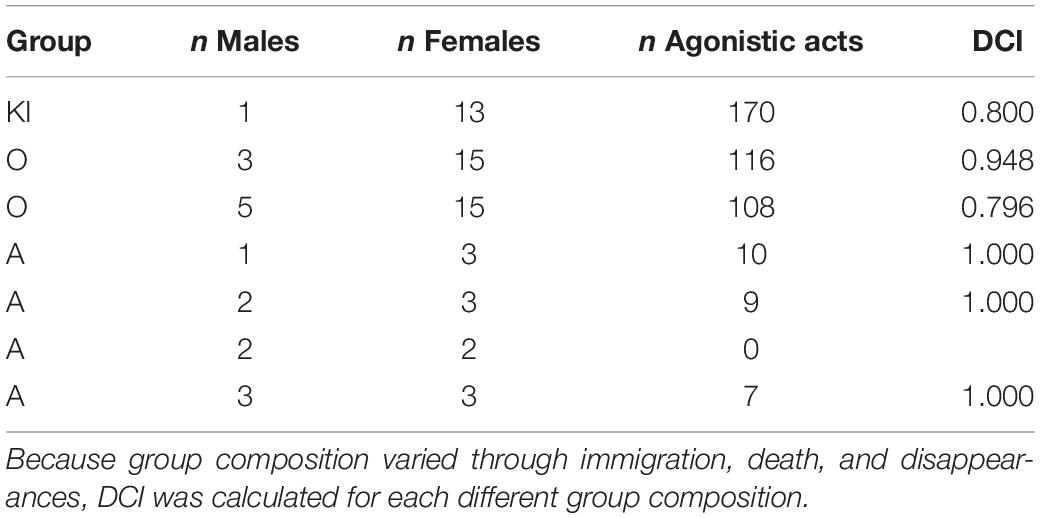
Table 2. Directional consistency index (DCI; van Hooff and Wensing, 1987) for intersexual agonistic acts among gray langurs.
Female counteraggression against males was overall very rare. During ad libitum and focal animal sampling, a female never responded to male agonism with aggression in groups KI and A. In O group, female counteraggression occurred three times and only during ad libitum sampling. In two cases, the female slapped a male after being chased/jumped on by him. The context of these two cases could not be determined. In a third case, the female initially harassed a male-female consortship, which resulted in the respective male chasing her. She responded by slapping the male.
In this section, we first describe the occurrence and context of spontaneous aggressive acts of females against males to see if aggression is context dependent or independent. Noteworthy, an aggressive act does not necessarily equal winning an agonistic bout. The winner of a bout will occupy the space or item that the loser gave up or the winner will receive submission. These decided interactions are used in the subsequent analysis of dominance indices and the relationship between partial female dominance and adult sex ratio.
Although rare, spontaneous aggression by females toward males occurred in KI and O groups (details below). It was not observed in A group. Male submission toward a female was even less frequent (in KI and O group), and again not observed in A group.
In KI group we observed 13 bouts in which females behaved aggressively toward the male, 12 of these encounters had a single female aggressor. These 12 bouts also included 5 submissive behaviors by the male. They occurred at a rate of 0.007 per hour or once every 142 focal hours. In O group we observed 23 bouts in which females were the aggressor, 19 of these encounters included a single female. These 19 bouts also included 2 submissive behaviors by males. Bouts with female aggressors (or male submission) occurred at a rate of 0.004 per hour or once every 250 focal hours.
Spontaneous aggression by a single female against the male in KI group did not occur in a particular context (Table 3). For most bouts (66.7%), no specific context could be determined. It did occur twice in a social context, once over food, and once as aggression redirected at the male after a female-female conflict. In O group, aggression by a single female against a male occurred most often when a female harassed a sexual interaction of the male with another female (37.5%). Importantly, all these sexual harassments were performed by the same individual female, but her targets were different males and females. Females were also aggressive when males came close to or approached young infants (25.0%) and one female defended her older, weaned male offspring against aggression by adult males (12.5%). Unknown (12.5%), food (6.4%), and after sexual behavior were the other remaining contexts. It is noteworthy, that aggression by coalitions of females against males (not included in Table 3) mainly occurred when a male approached or came close to a young infant (KI group: 1 of 1; O group: 3 of 4, 1 = unknown context).
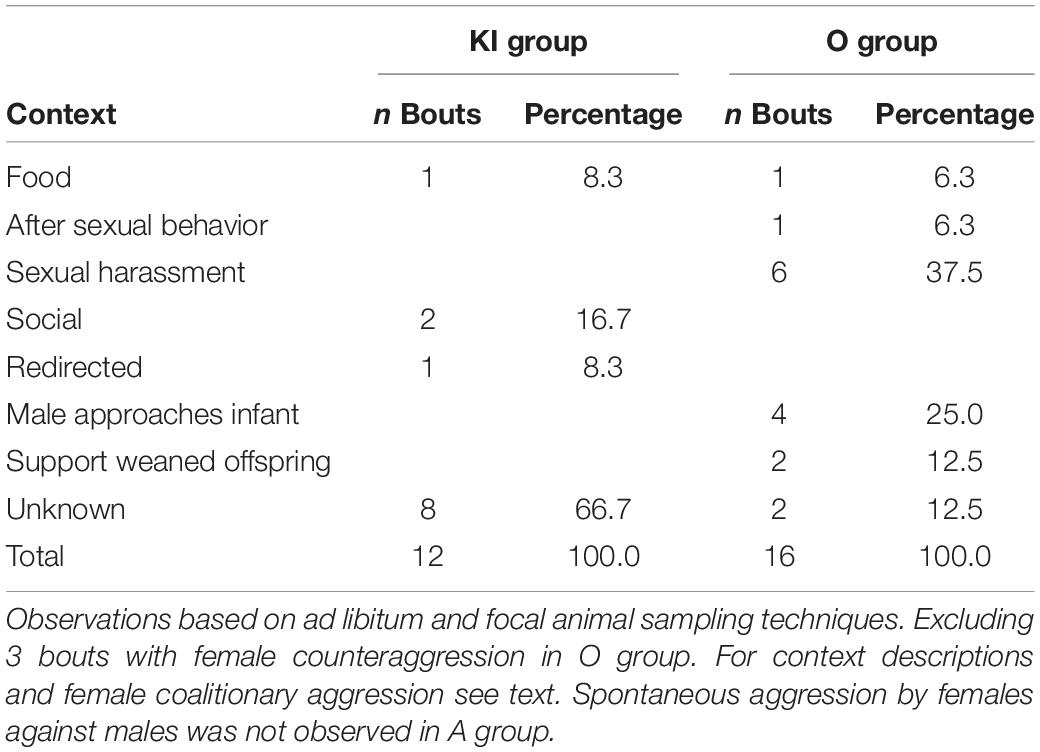
Table 3. Spontaneous aggression by a single female against a male in gray langurs.
To determine the extent to which females dominated males, we compared dominance indices (Table 4; see also Supplementary Tables 2–7 for details). In KI group, females never outranked the male and the FDI was 0.0. In other words, the ADI of the male was higher than the ADIs of every female in the group. In addition, while females occasionally won against the male, never did a female win more often than the male did (Supplementary Table 2). In O group when it had 3 adult males, all males had higher ADIs than all females and the FDI was 0.0. Again, winning by a female occurred, but in all dyads males won more often than females (Supplementary Table 3). In O group when it had 5 adult males, one of the females had a higher ADI than two of the males resulting in an FDI of 0.03. However, this female lost all encounters with the two males who had a lower ADI than she did. In addition, in this 5-male constellation, two females with ADIs lower than all males, nevertheless won against individual males (2 dyads total; Supplementary Table 4). Lastly, in group A the FDI ranged between 0.0 and 0.17. It was 0.0 for the constellation of one male and three females, 0.08 for 2 males and 3 females, and 0.17 for 3 males and 3 females. Noteworthy, in no case did an individual female win against a male (Supplementary Tables 5–7). The FDIs larger than zero are likely an artifact of small sample size and a lack of interactions resulting in several unknown relationships (Supplementary Tables 6, 7).
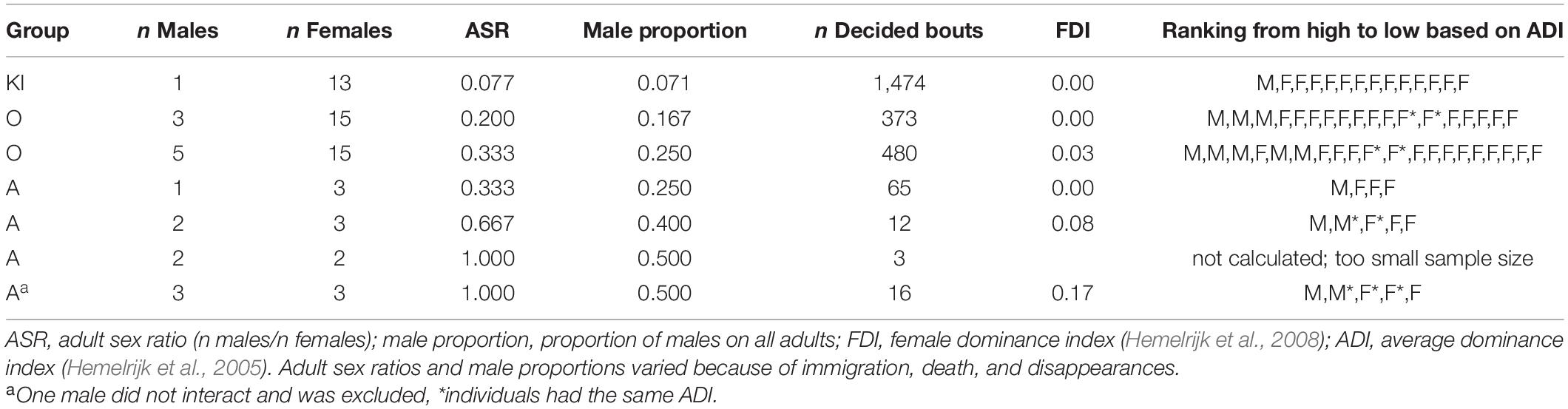
Table 4. Group composition, adult sex ratio, and dominance indices for gray langurs based on decided agonistic bouts of all adult individuals.
Most of the agonism by males directed at females occurred in a feeding context (Table 5). This could be a simple approach-retreat interaction at a feeding location or could include aggression. Food related agonism by males occurred most often at Ramnagar (A group: 94.1%; O group: 57.3%) and was less common at Jodhpur (21.5%). But given that in the latter population most contexts (60.7%) were not determined, feeding was also the most frequently determined context. If only known contexts are considered, food makes up more than 50% of the contexts in all three groups (KI group: 54.8%, out of n = 42; O group: 73.5%, out of n = 117; A group: 100.0%, out of n = 16).
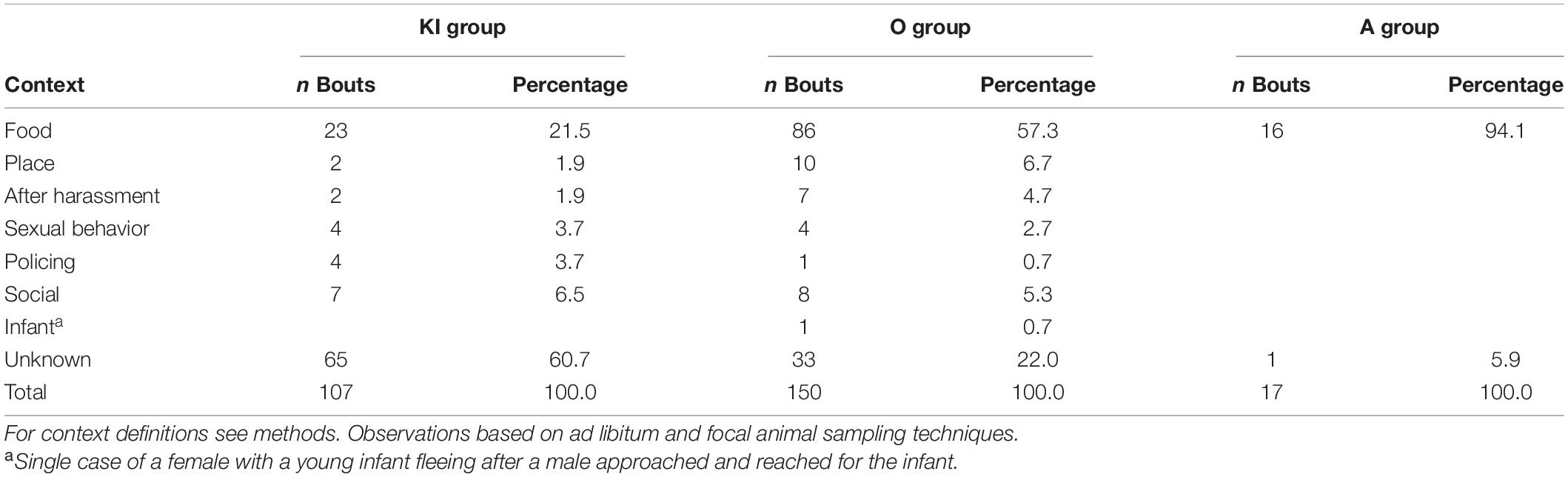
Table 5. Male aggression and displacements directed at females in gray langurs.
Male intervention in conflicts occurred but was rare (see “policing” Table 5). In KI group, the male always intervened on behalf of the loser. In three cases those were conflicts between adult females, and in one case he supported a juvenile male in a conflict with its mother. In O group a male intervened once on behalf of a winner in a female-female conflict. Male interventions were not observed in A group.
Agonism by males related to a sexual context was also rare, occurring in two main situations: (i) directed at a female who had harassed a sexual interaction, or (ii) after a sexual interaction with the very female. In both KI group (1.9%) and O group (4.7%) males behaved aggressively toward females who approached or actively harassed a sexual interaction between the male and another female. In the same two groups (KI group: 3.7%, O group: 1.3%), males behaved aggressively after females had directed proceptive behavior at them. Only in the remaining two cases in O group was a male aggressive during a sexual interaction. In one case this happened following a harassment by another female. In the other case, a male chased a female after a copulation attempt.
We found that in gray langurs intersexual agonistic relationships were characterized primarily by female submission toward males. If aggression by males occurred, it was predominantly of low intensity. Overall, the directionality of interactions was very consistent and counteraggression by females against males was rare. Using the criteria developed for male-male as well as for female-female relationships, these results match to some extent the definition of a despotic dominance style (de Waal and Luttrell, 1989; Preuschoft and van Schaik, 2000; Thierry, 2000; Flack and de Waal, 2004). The notion of despotism is also supported by the fact that female gray langurs use bare-teeth displays in agonistic interactions. Such signals of submission are only expected in despotic species (Preuschoft and van Schaik, 2000).
Contrasting with the finding of despotic intersexual relationships is the relatively low amount of high intensity aggression, perhaps indicating a less despotic dominance style. Alternatively, it may suggest a weaker correlation between directional consistency, counteraggression, and high intensity aggression than theoretically predicted (de Waal and Luttrell, 1989; Thierry, 2000). This would fit to the idea that intersexual relationships in gray langurs are less despotic than, for example, in rhesus macaque females but more despotic than in stump-tailed macaque females, supporting the notion that the despotic-egalitarian dominance distinction is less categorical and more gradual (Preuschoft and van Schaik, 2000; Thierry, 2000).
The low amount of high intensity aggression does not, however, mean that females do not potentially suffer from aggression by males. At both study sites, the frequency of aggression by gray langur males directed toward females was clearly lower than what has been found, for example, in chimpanzees, chacma baboons, or hamadryas baboons (Muller et al., 2007, 2009, 2011; Swedell et al., 2014; Baniel et al., 2017; Watts, 2022). Nonetheless, male aggression can reduce female fitness because incoming males (both at Jodhpur and Ramnagar) may attack and kill infants (Sommer, 1987; Borries, 1997). In both populations, more than 30% of infant mortality could be attributed to infanticide committed by males (Borries and Koenig, 2000). Thus, the mainly low intensity of male aggression directed at females in gray langurs does not equal low impact. The sexual conflict between males and females in this species is expressed indirectly via attacks on infants and infanticide rather than directly through the sexual coercion of females themselves (see in addition the discussion under section “Contexts of Male Agonism Against Females”).
It seems noteworthy that the characterization of despotism extends beyond male-female relationships in gray langurs. At both sites, all agonistic relationships have been characterized as despotic. This is true for male-male relationships at both Jodhpur (Sommer pers. com.) and Ramnagar (Perlman et al., 2016). And it is also true for female-female relationships at Jodhpur (Borries et al., 1991; Lu et al., 2008) and at Ramnagar (Lu et al., 2013; Riaz et al., in preparation). Additionally, the relationship between rank and reproduction for males and females can be characterized as despotic (sensu Vehrencamp, 1983), i.e., reproductive skew is high (males Jodhpur: Sommer and Rajpurohit, 1989; males: Ramnagar: Launhardt et al., 2001; females Jodhpur: Borries et al., 1991; females Ramnagar: de Vries et al., 2016). This general despotism in both agonistic relationships and reproductive skew is, however, not the norm among non-human primates. Male and female reproductive skew varies widely in primates (Muller and Emery Thompson, 2012; Pusey, 2012) and may or may not match the dominance style. Even within a single species, male and female dominance styles can differ (Preuschoft et al., 1998).
We found that aggression by females against males was extremely rare (once every 142–250 h). Furthermore, adult females rarely dominated adult males and the FDI was close to zero. In the few cases when a female had an average dominance index similar to or higher than a male, the respective female never won a single encounter with this male. To some extent this relates to the small sample size in A group, but it also shows that the dominance indices overall are measures of “power” (Hemelrijk et al., 2008) and may not necessarily reflect dyadic relationships.
The finding of rare female aggression against males and few dyads of female dominance over males stands in contrast to the theoretical prediction that depending on the adult sex ratio, females show frequent aggression toward males or females may even dominate some or several males in their group (Hemelrijk et al., 2008, 2020; Izar et al., 2021). So far, such partial female dominance has been shown within some non-human primate species and humans (Stroebe et al., 2017; Hemelrijk et al., 2020; Izar et al., 2021), as well as in a comparative analysis of male-female agonism across a larger sample of non-human primates (Hemelrijk et al., 2008).
An absence of the suggested relationship between partial female dominance and adult sex ratio has previously been found in species with more egalitarian relationships, as in some macaques (Hemelrijk et al., 2008). However, as shown above, gray langurs must be considered despotic in their agonistic relationships and their reproductive skew, making this explanation unlikely.
Another potential reason for the lack of a relationship between partial female dominance and adult sex ratio might relate to prior attributes of fighting ability overriding the suggested winner-loser effects in adult males assumed to facilitate partial female dominance. While the suggested relationship between partial female dominance and adult sex ratio seems to hold across primates and within species such as vervets and capuchins, it might be absent in a single species with very strong sexual size dimorphism. For example, mandrills and gorillas with a body mass dimorphism greater than two have so far not been shown to have partial female dominance (Table 4 in Hemelrijk et al., 2008). However, gray langurs have a body mass dimorphism of about 1.3 (Smith and Jungers, 1997), which is much lower than in gorillas and mandrills. In fact, the dimorphism in gray langurs is even lower than in vervets and capuchins thus also rendering this explanation unlikely.
In addition, a relationship between adult sex ratio and partial female dominance is predicted to occur when conflict levels among males are high. In this situation, the winner-loser effects among males may be stronger and females can benefit from exploiting these effects (Stroebe et al., 2017). In gray langur males at Ramnagar agonism consists of about equal proportions of aggression and submission, and most aggression is of low intensity (75%; Koenig and Borries, unpublished), perhaps indicating a low level of conflict. This contrasts with the finding that injuries among males are twice as frequent as among females (Feder et al., 2019). Thus, we are not in the position to test the idea of winner-loser effects but contend that a low level of agonistic conflict among males could relate to the absence of an adult sex ratio effect.
It seems noteworthy that, while female aggression against males was rare, it occurred in specific contexts. Interestingly, females (in O group) behaved aggressively most frequently during high stakes interventions, when infants or older offspring were perceived as at risk. During these situations females even formed coalitions with each other, a behavior that is otherwise rare among gray langur females (Borries, 1993). While speculative, because of the few events observed, this might indicate that females may dominate males in specific contexts only, i.e., context-dependent dominance or spheres of dominance (Hand, 1986; Preuschoft and van Schaik, 2000). If true, this could explain why a general dominance index would not yield partial female dominance.
Overall, we note that the proposed relationship between female aggression, partial female dominance, and adult sex ratio does not seem to hold in gray langurs. Our study suggests more variation in these relationships than previously thought (Hemelrijk et al., 2008, 2020). More studies with many more species are clearly needed to determine how general the predicted relationships are and if the expected criteria are being met.
We found that agonistic behavior of males against females occurred primarily over food but was very rarely observed in a policing or sexual context.
The near absence of male policing as a context of male agonism seems to fit the general primate pattern (Watts et al., 2000). Male interventions are expected to be more common when female dispersal is common. Here, conflicts between females may lead to the loser leaving the group and male interventions may act to calm conflicts, facilitate female integration, and prevent dispersal (Watts et al., 2000). In support of this idea, male interventions seem to be more common in species with female dispersal such as gorillas, hamadryas baboons, and some Asian colobines (Watts et al., 2000). In contrast, female dispersal in gray langurs is extremely rare (Borries et al., 1991; Koenig et al., 1998) and, hence, frequent male intervention is neither expected nor found.
We observed only two cases (in O group) regarding direct sexual coercion that might link sexual behavior and agonism. This does not necessarily mean that male aggression cannot affect the behavior of receptive females, because aggression and mating may be decoupled (Baniel et al., 2017). Gray langur males, at least in Ramnagar, have been shown to vary in their aggressiveness, but high aggressiveness did not lead to higher reproductive success (Borries et al., 2017). In addition, the overall low level of aggression, particularly high intensity aggression, makes it unlikely that direct sexual coercion is a tactic used by gray langur males. These results stand in contrast to the findings of frequent sexual coercion that have accumulated over the past years particularly for some baboons and chimpanzees (Muller et al., 2007, 2009, 2011; Swedell et al., 2014; Baniel et al., 2017; Watts, 2022). Importantly, in these species male aggression not only occurs frequently, but is a tactic that increases mating and reproductive success (Muller et al., 2007, 2011; Feldblum et al., 2014; Baniel et al., 2017; Watts, 2022). However, our results emphasize the need for a broader look at the causes underlying this variation in male reproductive tactics.
Overall, most of the agonistic behavior of males directed at females occurred over access to food. This is insofar surprising as the potential effect of feeding competition is almost never explicitly considered in male-male or male-female relationships (but see Perry, 1997; Pereira et al., 2000; Schuelke, 2001; Muller et al., 2009; Perlman et al., 2016). However, males need appropriate food to stay healthy and to compete with other males (Muller et al., 2009). Importantly, in the study population at Ramnagar, male physical condition is an important aspect relating to male dominance rank; males in better physical condition are often higher ranking (Perlman et al., 2016). It is thus not very surprising that male agonism even toward females centers on food. As male reproductive success is primarily affected by dominance rank (Launhardt et al., 2001) and dominance rank is affected by access to food (Perlman et al., 2016), males may compete with females over food. The importance of feeding competition is further underscored by the fact that agonism over food for male-male dyads was the second most common context with more than 25% (most common was the context unknown; Koenig and Borries, unpublished). In addition, the proportion of intersexual conflicts over food is very similar to female-female dyads, where it ranged between 50 and 80% (Koenig and Borries, 2006). Unfortunately, there are very little comparative data available for male-female agonism. In some primate species, food context may make up around 20% of male-female conflicts (Smuts, 1987; Muller et al., 2009) and only some species of capuchins have more frequent encounters over food (Janson, 1985; O’Brien, 1991). The latter fits with other observations of capuchins, in which foraging of females was frequently interrupted by males and females formed coalitions to supplant males from feeding sites (Rose, 1994; Perry, 1997). Thus, while this topic has received little attention, the few available results suggest that competition for food can be an important factor impacting male-female relationships.
Overall, our results indicate that female gray langurs may face not one but two major costs of living in groups with males. They face the threat of infanticide, and also compete with males over access to food. While we know that infanticide has a significant impact on female fitness, the effect of intersexual feeding competition is less clear and requires further study.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Ethical review and approval was not required for the animal study because the two non-invasive studies were conducted in 1985 (in India) and 1994-95 (in Nepal), respectively.
AK and CB collected and analyzed the data from Ramnagar, Nepal, and Jodhpur, India, respectively, and designed this study. AM and DR transcribed and proofread the data, ran first analyses, and produced first tables, with assisted by AK and CB. AK drafted the manuscript. All authors contributed to and edited the content, structure, and language.
Data collection in India was financially supported by the German Academic Exchange Service (DAAD, Germany) and the Ministry of Education, Culture, and Social Welfare (Government of India). Data collection in Nepal was financially supported by the German Research Council (DFG; Vo 124/19-1 + 2) and the Alexander von Humboldt-Foundation (V-3-FLF-1014527). Analysis, write-up, and publication were supported through funds from the Dean of the College of Arts and Sciences, Stony Brook University.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank Marcy Ekanayake-Weber, Rachel Perlman, and Samantha Vee for discussion and Charlotte Hemelrijk, Elise Huchard, and two reviewers for their helpful comments. For permission to conduct the study in Jodhpur, we thank the Ministry of Education, Culture, and Social Welfare (Government of India). For permission to conduct the study in Ramnagar, we also thank the Ministry of Education, Culture, and Social Welfare (HMG, Katmandu) as well as the Research Division of the Tribhuvan University (Katmandu). The research was non-invasive, adhered to the ASAB/ABS guidelines for the use of animals in research (Association for the Study of Animal Behaviour and Animal Behavior Society, 1992), and was in compliance with the laws and regulations of India, Nepal, and Germany.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.860437/full#supplementary-material
Archie, E. A., Tung, J., Clark, M., Altmann, J., and Alberts, S. C. (2014). Social affiliation matters: both same-sex and opposite-sex relationships predict survival in wild female baboons. Proc. R. Soc. Lond. B. 281:20141261. doi: 10.1098/rspb.2014.126
Arekar, K., Sathyakumar, S., and Karanth, P. K. (2021). Integrative taxonomy confirms the species status of the Himalayan langurs, Semnopithecus schistaceus Hodgson, 1840. J. Zool. Syst. Evol. Res. 59, 543–556. doi: 10.1111/jzs.12437
Association for the Study of Animal Behaviour, and Animal Behavior Society (1992). Guidelines for the use of animals in research. Anim. Behav. 43, 185–188. doi: 10.1016/S0003-3472(05)80094-6
Baniel, A., Cowlishaw, G., and Huchard, E. (2017). Male violence and sexual intimidation in a wild primate society. Curr. Biol. 27, 2163–2168. doi: 10.1016/j.cub.2017.06.013
Borries, C. (1989). Konkurrenz unter freilebenden Langurenweibchen (Presbytis entellus). Ph.D. thesis. Goettingen: Georg-August University.
Borries, C. (1993). Ecology of female social relationships – Hanuman langurs (Presbytis entellus) and the van Schaik model. Folia Primatol. 61, 21–30. doi: 10.1159/000156723
Borries, C. (1997). Infanticide in seasonally breeding multimale groups of Hanuman langurs (Presbytis entellus) in Ramnagar (South Nepal). Behav. Ecol. Sociobiol. 41, 139–150. doi: 10.1007/s002650050373
Borries, C. (2000). “Male dispersal and mating season influxes in Hanuman langurs living in multi-male groups,” in Primate Males: Causes and Consequences of Variation in Group Composition, ed. P. M. Kappeler (Cambridge: Cambridge University Press)), 146–158.
Borries, C., and Koenig, A. (2000). “Infanticide in Hanuman langurs: social organization, male migration, and weaning age,” in Infanticide by Males and its Implications, eds C. P. van Schaik and C. H. Janson (Cambridge: Cambridge University Press), 99–122.
Borries, C., Perlman, R. F., and Koenig, A. (2017). Characteristics of alpha males in Nepal gray langurs. Am. J. Primatol. 79:e22437. doi: 10.1002/ajp.22437
Borries, C., Sommer, V., and Srivastava, A. (1991). Dominance, age, and reproductive success in free-ranging female Hanuman langurs (Presbytis entellus). Int. J. Primatol. 12, 231–257. doi: 10.1007/BF02547586
Carnes, L. M., Nunn, C. L., and Lewis, R. J. (2011). Effects of the distribution of female primates on the number of males. PLoS One 6:e19853. doi: 10.1371/journal.pone.0019853
Clutton-Brock, T. H. (2021). Social evolution in mammals. Science 373, eabc9699. doi: 10.1126/science.abc9699
Clutton-Brock, T. H., and Janson, C. H. (2012). Primate socioecology at the crossroads: past, present, and future. Evol. Anthropol. 21, 136–150. doi: 10.1002/evan.21316
de Vries, D., Koenig, A., and Borries, C. (2016). Female reproductive success in a species with an age-inversed hierarchy. Integr. Zool. 11, 433–446. doi: 10.1111/1749-4877.12201
de Vries, H., Netto, W. J., and Hanegraaf, P. L. H. (1993). MatMan: a program for the analysis of sociometric matrices and behavioral transition matrices. Behaviour 125, 157–175.
de Waal, F. B. M., and Luttrell, L. M. (1989). Toward a comparative socioecology of the genus Macaca: different dominance styles in rhesus and stumptail monkeys. Am. J. Primatol. 19, 83–109. doi: 10.1002/ajp.1350190203
Dolhinow, P. J. (1978). A behavior repertoire for the Indian langur monkey (Presbytis entellus). Primates 19, 449–472.
Emlen, S. T., and Oring, L. W. (1977). Ecology, sexual selection, and the evolution of mating systems. Science 197, 215–223. doi: 10.1126/science.327542
Feder, J. A., Lu, A., Koenig, A., and Borries, C. (2019). The costs of competition: injury patterns in 2 Asian colobine monkeys. Behav. Ecol. 30, 1242–1253. doi: 10.1093/beheco/arz070
Feldblum, J. T., Wroblewski, E. E., Rudicell, R. S., Hahn, B. H., Paiva, T., Cetinkaya-Rundel, M., et al. (2014). Sexually coercive male chimpanzees sire more offspring. Curr. Biol. 24, 2855–2860. doi: 10.1016/j.cub.2014.10.039
Flack, J. C., and de Waal, F. B. M. (2004). “Dominance style, social power, and conflict management in macaque societies: a conceptual framework,” in Macaque Societies: A Model for the Study of Social Organization, eds B. Thierry, M. Singh, and W. Kaumanns (Cambridge: Cambridge University Press), 157–181.
Hand, J. L. (1986). Resolutions of social conflict: dominance, egalitarianism, spheres of dominance and game theory. Q. Rev. Biol. 61, 201–220. doi: 10.1086/414899
Hemelrijk, C. K., Wantia, J., and Gygax, L. (2005). The construction of dominance order: comparing performance of five methods using an individual-based model. Behaviour 142, 1037–1058. doi: 10.1163/156853905774405290
Hemelrijk, C. K., Wantia, J., and Isler, K. (2008). Female dominance over males in primates: self-organisation and sexual dimorphism. PLoS One 3:e2678. doi: 10.1371/journal.pone.0002678
Hemelrijk, C. K., Wubs, M., Gort, G., Botting, J., and van de Waal, E. (2020). Dynamics of intersexual dominance and adult sex-ratio in wild vervet monkeys. Front. Psychol. 11:839. doi: 10.3389/fpsyg.2020.00839
Isbell, L. A. (1991). Contest and scramble competition: patterns of female aggression and ranging behavior among primates. Behav. Ecol. 2, 143–155. doi: 10.1093/beheco/2.2.143
Izar, P., Fernandez-Bolanos, M., Seex, L., Gort, G., Suscke, P., Tokuda, M., et al. (2021). Female emancipation in a male dominant, sexually dimorphic primate under natural conditions. PLoS One 16:e0249039. doi: 10.1371/journal.pone.0249039
Janson, C. H. (1985). Aggressive competition and individual food consumption in wild brown capuchin monkeys (Cebus apella). Behav. Ecol. Sociobiol. 18, 125–138.
Kappeler, P. M. (2017). Sex roles and adult sex ratios: insights from mammalian biology and consequences for primate behaviour. Phil. Trans. R. Soc. B. 372:20160321. doi: 10.1098/rstb.2016.0321
Koenig, A. (2000). Competitive regimes in forest-dwelling Hanuman langur females (Semnopithecus entellus). Behav. Ecol. Sociobiol. 48, 93–109. doi: 10.1007/s002650000198
Koenig, A. (2002). Competition for resources and its behavioral consequences among female primates. Int. J. Primatol. 23, 759–783.
Koenig, A., Beise, J., Chalise, M. K., and Ganzhorn, J. U. (1998). When females should contest for food - testing hypotheses about resource density, distribution, size, and quality with Hanuman langurs (Presbytis entellus). Behav. Ecol. Sociobiol. 42, 225–237. doi: 10.1007/s002650050434
Koenig, A., and Borries, C. (2006). “The predictive power of socioecological models: a reconsideration of resource characteristics, agonism, and dominance hierarchies,” in Feeding Ecology in Apes and Other Primates: Ecological, Physiological, and Behavioral Aspects, eds G. Hohmann, M. M. Robbins, and C. Boesch (Cambridge: Cambridge University Press), 263–284.
Koenig, A., Borries, C., Chalise, M. K., and Winkler, P. (1997). Ecology, nutrition, and timing of reproductive events in an Asian primate, the Hanuman langur (Presbytis entellus). J. Zool. 243, 215–235. doi: 10.1111/j.1469-7998.1997.tb02778.x
Koenig, A., Scarry, C. J., Wheeler, B. C., and Borries, C. (2013). Variation in grouping patterns, mating systems and social structure: what socio-ecological models attempt to explain. Phil. Trans. R. Soc. B. 368:9. doi: 10.1098/rstb.2012.0348
Launhardt, K., Borries, C., Hardt, C., Epplen, J. T., and Winkler, P. (2001). Paternity analysis of alternative male reproductive routes among the langurs (Semnopithecus entellus) of Ramnagar. Anim. Behav. 61, 53–64. doi: 10.1006/anbe.2000.1590
Lu, A., Borries, C., Caselli, A., and Koenig, A. (2013). Effects of age, reproductive state, and the number of competitors on the dominance dynamics of wild female Hanuman langurs. Behaviour 150, 485–523. doi: 10.1163/1568539X-00003064
Lu, A., Koenig, A., and Borries, C. (2008). Formal submission, tolerance and socioecological models: a test with female Hanuman langurs. Anim. Behav. 76, 415–428. doi: 10.1016/j.anbehav.2008.04.006
Martin, P., and Bateson, P. (2007). Measuring Behaviour: An Introductory Guide. Cambridge: Cambridge University Press.
Muller, M. N., and Emery Thompson, M. (2012). “Mating, parenting, and male reproductive strategies,” in The Evolution of Primate Societies, eds J. C. Mitani, J. Call, P. M. Kappeler, R. A. Palombit, and J. B. Silk (Chicago: The University of Chicago Press)), 387–411.
Muller, M. N., Emery Thompson, M., Kahlenberg, S. M., and Wrangham, R. W. (2011). Sexual coercion by male chimpanzees shows that female choice may be more apparent than real. Behav. Ecol. Sociobiol. 65, 921–933. doi: 10.1007/s00265-010-1093-y
Muller, M. N., Kahlenberg, S. M., Emery Thompson, M., and Wrangham, R. W. (2007). Male coercion and the costs of promiscuous mating for female chimpanzees. Proc. R. Soc. Lond. B 274, 1009–1014. doi: 10.1098/rspb.2006.0206
Muller, M. N., Kahlenberg, S. M., and Wrangham, R. W. (2009). “Male aggression and sexual coercion of females in primates,” in Sexual Coercion in Primates and Humans: An Evolutionary Perspective on Male Aggression Against Females, eds M. N. Muller and R. W. Wrangham (Cambridge: Harvard University Press)), 3–22.
O’Brien, T. G. (1991). Female-male social interactions in wedge-capped capuchin monkeys: benefits and costs of group living. Anim. Behav. 41, 555–567. doi: 10.1016/S0003-3472(05)80896-6
Ostner, J., and Schuelke, O. (2014). The evolution of social bonds in primate males. Behaviour 151, 871–906. doi: 10.1163/1568539X-00003191
Ostner, J., Vigilant, L., Bhagavatula, J., Franz, M., and Schuelke, O. (2013). Stable heterosexual associations in a promiscuous primate. Anim. Behav. 86, 623–631. doi: 10.1016/j.anbehav.2013.07.004
Pereira, M. E., Clutton-Brock, T. H., and Kappeler, P. M. (2000). “Understanding male primates,” in Primate Males: Causes and Consequences of Variation in Group Composition, ed. P. M. Kappeler (Cambridge: Cambridge University Press), 271–277.
Perlman, R. F., Borries, C., and Koenig, A. (2016). Dominance relationships in male Nepal gray langurs (Semnopithecus schistaceus). Am. J. Phys. Anthropol. 160, 208–219. doi: 10.1002/ajpa.22958
Perry, S. (1997). Male-female social relationships in wild white-faced capuchins (Cebus capucinus). Behaviour 134, 477–510. doi: 10.1163/156853997X00494
Podzuweit, D. (1994). Sozio-Oekologie weiblicher Hanuman Languren (Presbytis entellus) in Ramnagar, Suednepal. Goettingen: Cuvillier Verlag.
Preuschoft, S., Paul, A., and Kuester, J. (1998). Dominance styles of female and male Barbary macaques (Macaca sylvanus). Behaviour 135, 731–755.
Preuschoft, S., and van Schaik, C. P. (2000). “Dominance and communication: conflict management in various social settings,” in Natural Conflict Resolution, eds F. Aurel and F. B. M. de Waal (Berkeley: University of California Press), 77–105.
Pusey, A. E. (2012). “Magnitude and sources of variation in female reproductive performance,” in The Evolution of Primate Societies, eds J. C. Mitani, J. Call, P. M. Kappeler, R. A. Palombit, and J. B. Silk (Chicago: The University of Chicago Press), 343–366.
Rajpurohit, L. S., and Sommer, V. (1993). “Juvenile male emigration from natal one-male troops in Hanuman langurs,” in Juvenile Primates – Life History, Development, and Behavior, eds M. E. Pereira and L. A. Fairbanks (New York: Oxford University Press), 86–103.
Rose, L. M. (1994). Benefits and costs of resident males to females in white-faced capuchins, Cebus capucinus. Am. J. Primatol. 32, 235–248. doi: 10.1002/ajp.1350320402
Rowell, T. E. (1974). The concept of dominance. Behav. Biol. 11, 131–154. doi: 10.1016/S0091-6773(74)90289-2
Scarry, C. J. (2013). Between-group contest competition among tufted capuchin monkeys, Sapajus nigritus, and the role of male resource defence. Anim. Behav. 85, 931–939. doi: 10.1016/j.anbehav.2013.02.013
Schuelke, O. (2001). Differential energy budget and monopolization potential of harem holders and bachelors in Hanuman langurs (Semnopithecus entellus): preliminary results. Am. J. Primatol. 55, 57–63. doi: 10.1002/ajp.1039
Smith, R. J., and Jungers, W. L. (1997). Body mass in comparative primatology. J. Hum. Evol. 32, 523–559. doi: 10.1006/jhev.1996.0122
Smuts, B. B. (1987). “Gender, aggression, and influence,” in Primate Societies, eds B. B. Smuts, D. L. Cheney, R. M. Seyfarth, R. W. Wrangham, and T. T. Struhsaker (Chicago: University of Chicago Press), 400–412.
Smuts, B. B., and Smuts, R. W. (1993). Male aggression and sexual coercion of females in nonhuman primates and other mammals: evidence and theoretical implications. Adv. Study Behav. 22, 1–63. doi: 10.1016/S0065-3454(08)60404-0
Sommer, V. (1985). Weibliche und maennliche Reproduktionsstrategien der Hanuman-Languren (Presbytis entellus) von Jodhpur, Rajasthan/India. Ph.D. thesis. Goettingen: Georg-August University.
Sommer, V. (1987). Infanticide among free-ranging langurs (Presbytis entellus) at Jodhpur (Rajasthan/India): recent observations and a reconsideration of hypotheses. Primates 28, 163–197.
Sommer, V. (1988). Male competition and coalitions in langurs (Presbytis entellus) at Jodhpur, Rajasthan, India. Hum. Evol. 3, 261–278.
Sommer, V., and Rajpurohit, L. S. (1989). Male reproductive success in harem troops of Hanuman langurs (Presbytis entellus). Int. J. Primatol. 10, 293–317.
Sterck, E. H. M., Watts, D. P., and van Schaik, C. P. (1997). The evolution of female social relationships in nonhuman primates. Behav. Ecol. Sociobiol. 41, 291–309.
Stroebe, K., Nijstad, B. A., and Hemelrijk, C. K. (2017). Female dominance in human groups: effects of sex ratio and conflict level. Soc. Psychol. Personal. Sci. 8, 209–218. doi: 10.1177/1948550616664956
Swedell, L., Leedom, L., Saunders, J., and Pines, M. (2014). Sexual conflict in a polygynous primate: costs and benefits of a male-imposed mating system. Behav. Ecol. Sociobiol. 68, 263–273. doi: 10.1007/s00265-013-1641-3
Thierry, B. (2000). “Covariation of conflict management patterns across macaque species,” in Natural Conflict Resolution, eds F. Aureli and F. B. M. de Waal (Berkeley, CA: University of California Press), 106–128.
van Hooff, J. A. R. A. M., and Wensing, J. A. B. (1987). “Dominance and its behavioral measures in a captive wolf pack,” in Man and Wolf, ed. H. Frank (Dordrecht: W Junk), 219–252.
van Schaik, C. P. (1989). “The ecology of social relationships amongst female primates,” in Comparative Socioecology: The Behavioural Ecology of Humans and Other Mammals, eds V. Standen and R. A. Foley (Oxford: Blackwell Scientific Publications), 195–218.
van Schaik, C. P. (1996). Social evolution in primates: the role of ecological factors and male behaviour. Proc. Brit. Acad. 88, 9–31.
van Schaik, C. P., Pradhan, G. R., and van Noordwijk, M. A. (2004). “Mating conflict in primates: infanticide, sexual harassment and female sexuality,” in Sexual Selection in Primates: New and Comparative Perspectives, eds P. M. Kappeler and C. P. van Schaik (Cambridge: Cambridge University Press), 131–150.
Vehrencamp, S. L. (1983). A model for the evolution of despotic versus egalitarian societies. Anim. Behav. 31, 667–682. doi: 10.1016/S0003-3472(83)80222-X
Watts, D. P. (2022). Male chimpanzee sexual coercion and mating success at Ngogo. Am. J. Primatol. 84:e23361. doi: 10.1002/ajp.23361
Watts, D. P., Colmenares, F., and Arnold, K. (2000). “Redirection, consolation, and male policing - how targets of aggression interact with bystanders,” in Natural Conflict Resolution, eds F. Aureli and F. B. M. de Waal (Berkeley, CA: University of California Press), 281–300.
Winkler, P. (1981). Zur Oeko-Ethologie freilebender Hanuman-Languren (Presbytis entellus entellus Dufresne, 1797) in Jodhpur (Rajasthan), Indien. Ph.D. thesis. Goettingen: Georg-August University.
Winkler, P. (1984). “The adaptive capacities of the Hanuman langur and the categorizing of diet,” in Food Acquisition and Processing in Primates, eds D. J. Chivers, B. A. Wood, and A. Bilsborough (New York, NY: Plenum Press), 161–166.
Keywords: adult sex ratio, aggression, counteraggression, despotism, dominance index, offspring defense, sexual coercion, submission
Citation: Koenig A, Miles A, Riaz D and Borries C (2022) Intersexual Agonism in Gray Langurs Reflects Male Dominance and Feeding Competition. Front. Ecol. Evol. 10:860437. doi: 10.3389/fevo.2022.860437
Received: 22 January 2022; Accepted: 30 May 2022;
Published: 16 June 2022.
Edited by:
Elise Huchard, UMR 5554 Institut des Sciences de l’Evolution de Montpellier (ISEM), FranceReviewed by:
Dieter Lukas, Max Planck Institute for Evolutionary Anthropology, GermanyCopyright © 2022 Koenig, Miles, Riaz and Borries. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andreas Koenig, YW5kcmVhcy5rb2VuaWdAc3Rvbnlicm9vay5lZHU=
†Present addresses: Durr-E-Ajam Riaz, Master’s Program in Developmental, Neural, and Behavioral Biology, Georg-August University, Göttingen, Germany; Cognitive Ethology Laboratory, German Primate Center, Göttingen, Germany
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.