
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 25 April 2022
Sec. Conservation and Restoration Ecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.850442
 Madeleine Geiger1,2,3*
Madeleine Geiger1,2,3* Claudia Kistler1
Claudia Kistler1 Prisca Mattmann4Lukas Jenni4
Prisca Mattmann4Lukas Jenni4 Daniel Hegglin1,5†
Daniel Hegglin1,5† Fabio Bontadina1,6†
Fabio Bontadina1,6†In many areas, domestic cats are the most abundant predators of small vertebrates. Due to the potential impact on prey populations by cats, there are calls to investigate the effectiveness of visual and acoustic cues as measures to reduce the cat’s hunting efficiency. In this study, we complement previous studies on the efficacy of Birdsbesafe collar-covers (BBScc) in a so far not investigated Continental European setting and explore the effectiveness in combination with a bell. We also evaluate the tolerability of these devices by the cat and the acceptance by their owners. With a randomized and comparative citizen science-based approach we collected data from 26 households with 31 study cats, which were wearing either a BBScc or both a BBScc and a bell. The BBScc reduced the number of birds brought home by 37% (probability of reduction of 88%). The number of mammals brought home was reduced by 54–62%, but only with the additional bell (probability of reduction of >99%). About one fourth of the birds that could be dissected were found to have collided with a hard object prior to having been brought home by the cats. Our results are in line with previous findings from Australia, the United States, and the United Kingdom and highlight the great potential of visual and acoustic cues in reducing hunting success in domestic cats also in Continental Europe. On the other hand, our result show that the number of prey brought home by cats overestimates their hunting bag, if scavenging is not considered. The majority of cat owners reported that their cats habituated quickly to the BBScc. However, frequent scratching in some cats indicates that some individuals may not habituate. Most participating cat owners had a positive attitude toward the BBScc and said that they were willing to use it after the study. However, cat owners reported that their social environment (e.g., neighbors, family, friends) was relatively skeptical, which indicates a need for communication. To conclude, commercially available devices with visual and acoustic stimuli are straightforward and effective ways to mitigate the potentially harmful effect of domestic cats on wildlife.
As an invasive predator in most parts of the world, the domestic cat (Felis catus) has been shown to pose a substantial threat to wildlife, in particular on geographically isolated islands and continents, including Australia (Salo et al., 2007; Medina et al., 2011; Doherty et al., 2016). Here, endemic prey species are often exhibiting degrees of prey naiveté toward these novel predator archetypes (Cox and Lima, 2006). However, in a continental setting, where a multitude of predator-prey relationships have been evolving over extensive periods of time, the magnitude of the impact of domestic cats on native wildlife has been difficult to assess.
Domestic cat populations stand out due to their exceptionally high densities, especially in built up areas, which are usually unmatched by native predators. For example, densities of domestic cats with access to the outdoors in the Swiss city of Zurich have been estimated at 429 individuals/km2 (SWILD, 2011), while densities of the most abundant medium sized wild predator, the red fox (Vulpes vulpes) in the same area were estimated at 7.4–11.2 adult individuals/km2 (Gloor et al., 2001). Wild cats (Felis silvestris) in suitable forest habitats in Switzerland reach densities of 0.23–0.26 individuals/km2 (Maronde et al., 2020; Nussberger and Roth, 2020). Although estimates of the number of birds and small mammals killed by cats per year are in the billions in large areas [see e.g., Loss et al. (2013) and Trouwborst et al. (2020), for a review], studies on the actual impact of domestic cats on different wildlife species and populations on the continents (except Australia, whose fauna evolved without felines for most of its history) are controversial. Several studies came to diverging conclusions, which cannot be extrapolated to other species and transferred into different geographical areas and habitat types without reservation [see e.g., Weggler and Leu (2001), Hawkins et al. (2004), Baker et al. (2005), Loss and Marra (2017), and Mori et al. (2019), for a review].
In a continental setting (again not considering Australia), intensive agricultural practices and urbanization are probably the most important factors resulting in large scale habitat loss and reduction of habitat quality and therefore declining wildlife populations [e.g., in Switzerland: Federal Office for the Environment (2017)]. As fragmentation of increasingly limited natural and semi natural habitats increases, residential gardens, urban parks, and buildings featuring a variety of microhabitats and food resources are important environments and even refuges for various wildlife species, including endangered ones [e.g., Baker et al. (2003), Ineichen et al. (2012), Taucher et al. (2020)]. Residential areas are usually also characterized by a high density of domestic cats [e.g., Liberg et al. (2000)] and the predation rate of pet cats per km2 residential area is many times greater than predation rates by feral cats in natural environments (Legge et al., 2020). Therefore, cat-wildlife conflicts might actually entail a serious conservation issue in the long term. Especially local populations of certain prey species might be negatively affected by domestic cats (Shaw et al., 2008).
Due to the potential negative impact of cats on wildlife populations as well as animal welfare implications of cats injuring wildlife, there is a call driven by some cat owners, animal welfare advocates, and wildlife biologists to investigate the effectiveness of measures to reduce the hunting efficiency of cats. It would be prudent to seek precautionary, feasible, tolerable, and accepted measures that are conforming with animal welfare standards, to reduce cat predation on wildlife and also help to minimize conflicts between cat owners and conservationists (Lynn et al., 2019). Several collar mounted devices have been developed to reduce predation rate of domestic cats, either based on auditory or visual cues to alert and deter potential prey (see below), or to impede free movement of the cats and thus reduce their hunting ability [e.g., catBib; Calver et al. (2007)]. Methods for auditory prey deterrence are bells and the electronic sonic device CatAlert, which in some comparative studies have been shown to reduce the number of prey brought home by cats (Clark and Burton, 1998; Clark, 1999; Ruxton et al., 2002; Nelson et al., 2005; Gordon et al., 2010), while in another study the bell did not reduce the number of birds and mammals brought home by cats (Cecchetti et al., 2021b).
Other measures use visual cues to enhance the detectability of the predator and alert prey early. One of the relatively novel measures is the Birdsbesafe collar-cover (hereafter BBScc), which is available in various patterns. The BBScc’s function is based on its colorful appearance that should act as an anti-camouflage to overcome the stealth ability of the cat. Studies in Australia (Hall et al., 2015), the United States (Willson et al., 2015), and the United Kingdom (Pemberton and Ruxton, 2020; Cecchetti et al., 2021a,b) have shown a substantial reduction of the number of prey brought home (and eaten) by cats wearing the BBScc. Although the results of these studies seem promising, the effect size of the BBScc across these different studies varies, pointing at confounding factors that need to be considered when evaluating the BBScc’s usefulness in reducing predation rates of cats. Notably, different environmental variables might influence the effectiveness of the BBScc’s across geographic areas. These environmental variables include e.g., latitude (i.e., daylength), vegetation zone, human settlement density, and bird species composition, with the different bird species potentially featuring variation in color perception (Price, 2017).
The aim of this study was to test the efficacy of BBScc in a Continental European setting and in combination with a bell, to complement the previous studies. Further, we address a key issue for the successful application in practice: we evaluate the BBScc in terms of its tolerability on the part of the cats and the acceptance on the part of their owners. Specifically, the aims of this study are to (1) quantify the efficiency of the BBScc, and the potential of combining this device with a bell, for reducing the number of prey brought home (interpreted as predation rate, for limitations of this concept see below) in a continental European setting, (2) evaluate the impact of the BBScc on cat behavior and thus potential implications on animal welfare, and (3) evaluate the acceptance of BBScc among the cat owners as well as the precepted acceptance in their surroundings. In light of the previous studies on the BBScc and our own preliminary experience prior to the study, our predictions are: (1) the BBScc and the bell are efficient in reducing the number of prey brought home by cats; (2) there is no substantial animal welfare issue related to the deployment of the BBScc; (3) people are generally reluctant to deploy the BBScc due to its peculiar appearance.
Data collection took place throughout Switzerland, i.e., in an area of temperate to Mediterranean climate with Western European broadleaf forests and Alpine conifer and mixed forests (Supplementary Figure 1). Most households were distributed across the Swiss plateau and situated in rural villages or suburban areas (Supplementary Figure 1 and Supplementary Data Sheet 1). The movement pattern of the study cats could not be tracked and the different habitats could not be accounted for in the analyses.
The main part of the project took place from May to August 2020, with a pilot phase from July to October 2019. We recruited cat owners via newsletters and social media articles hosted by organizations targeting nature lovers and people concerned with animal welfare, as well as via personal networks. Participants were only included in the study if cats were reported to bring home at least one prey item per week prior to the study.
All cats were equipped with a BBScc, which was mounted on a quick-release collar (Figure 1). For the latter we used “cat-life,” which is adjustable to the individual body weight of the cat to ensure animal welfare (Figure 1). For the extended survey (see below), a bell was added to the BBScc (Figure 1). All participants received the same type of bell and BBScc to ensure comparability of results. For the BBScc, we chose a “rainbow” pattern featuring stripes of various colors (Figure 1). Rainbow color BBScc have previously been shown to be more effective in reducing predation rates compared to more uniformly colored BBScc (Hall et al., 2015).

Figure 1. One of the cats participating in our study, wearing a Birdsbesafe collar-cover. The inlet shows a close-up of the Birdsbesafe collar-cover, mounted on a quick-release collar (cat-life), which is also equipped with a bell. This type of collar-cover was used for all cats in the current study. Photograph courtesy by Kathi Märki/swild.ch.
The acoustic characteristics of the bells was investigated by comparing five bells with apparently different frequencies (according to human perception). Every bell was recorded manually from one meter distance using an ultrasound recorder (Batlogger M, Elekon AG, Switzerland) with an electret condenser microphone (frequency response 10–130 kHz). The recordings were subsequently analysed using Raven Pro software (Version 1.4, Cornell University). We found that the frequency of the bells was overall similar, with 4–5 peaks with main frequencies between 5 and 45 kHz (Supplementary Figure 2).
Additionally, we provided the participants with a detailed manual with instructions on the exact procedures they had to follow (experimental protocol), on how to familiarize their cats to the BBScc (adjustment period), on how to adjust the quick-release collar, and on how to monitor their cat’s behavior (with a list of criteria to prematurely terminate the treatment in case the cat exhibited unusual behavior, see Supplementary Data Sheet 1).
Participants could choose freely if they wanted to test only the BBScc (standard survey) or the BBScc plus a bell (extended survey). Every study cat was surveyed by their owners during two (standard survey) or three (extended survey) treatment-phases of 2 weeks duration each: treatment 1: wearing a BBScc; treatment 2: wearing a BBScc plus a bell (only for extended survey); control: wearing neither BBScc nor bell. As a result, every cat was either participating in treatment 1 and the control (standard survey, 4 weeks in total), or in treatment 1, treatment 2, and the control (extended survey, 6 weeks in total). There were only two exceptions, where one household participated twice in 1 year (total duration of 12 weeks) and where only the BBScc + bell and the control were tested (without BBScc alone). Most participants held strictly to the study protocol and each scheduled 2-week treatment-phase was on average 13.8 days long (range 11–17 days). In terms of the collar on which the BBScc was mounted, the study cats wore whatever was normal for them (a collar or no collar) prior to this study during the control-phase (Pemberton and Ruxton, 2020). If more than one cat was present in a household, we equipped all cats of that household with a BBScc because prey items may be hard to assign to a specific cat. Further, we attributed all cats in one household to the same treatment-sequence group (see below) and conducted all analyses per household and not per study cat. We randomly assigned every household to a treatment-sequence group. In these groups, the sequence of treatment-phases (treatment 1, treatment 2, and control) was varied in every possible way, with treatment 2 always following or being followed by treatment 1, thus resulting in 6 possible sequences (Supplementary Table 1). As a result, the starting dates of the treatments were equally distributed over the season (Kruskal-Wallis tests for equal medians, p > 0.05) and thus, hunting bags per treatment phase were unlikely to be biased by date, e.g., by the occurrence of many juvenile prey later in summer.
We asked participants to record all prey items of four prey taxa (birds, mammals, “reptiles” [i.e., squamates], insects) brought home by their cats during all treatment-phases (except during an eventual adjustment period prior to the survey), even if they were subsequently eaten by the survey cat or released by the participants if unharmed. Specifically, we asked participants to note the date and observed/suspected time the prey was caught and/or brought home by the cat(s), the treatment-phase during which the prey was brought home (treatment 1, treatment 2, or control), and information on the prey item (taxon, condition, approximate age-stage). Prey items that were not brought home by the cat and killed and/or eaten in the field could not be recorded in this study. We asked participants to take photographs of the prey to determine species and age-stage. If the prey could be seized, we asked participants to store them in a freezer and send them to us after termination of the survey. We assumed that cat owners reported prey unbiased by treatment-phase.
Study cats lost their BBScc 13 times in total during the survey period. In these cases, we provided a replacement as quickly as possible and excluded all the prey items that were brought home by the respective study cat in the period with lost BBScc (and bell) from the analyses. If possible, the lost days were added to the respective treatment-phase so that the treatment-phase lengths could be held as constant among the households as possible. Similarly, we excluded prey items brought home if the study cats by mistake had access to the outdoors without wearing a BBScc (and a bell) during treatment 1 and 2, e.g., if the owners forgot to mount the BBScc before letting the cat out. Because this happened for only a couple of hours, this time could not be added to the respective treatment phase. The actual time the study cats spent outdoors—and thus their potential time of hunting—could not be monitored. The access to the outdoors varied among the households, but we assumed that in the absence of information suggesting otherwise, cat owners did not change the outdoor access of their cats between the control and treatment-phases. Further details about the methodology and the raw data can be obtained from the Supplementary Data Sheets 1, 2.
As we wanted to test the effectiveness of the BBScc and the bell, we excluded birds brought home by study cats from the analyses which were hunted during situations when the BBScc could not have been effective. These were (1) nestlings that were unable to fly; (2) scavenged birds, i.e., which were severely injured or dead before taken by the cat; (3) birds taken during the hours of darkness. [In mammals, only criterion (1) was applied, but early post-natal juvenile mammals with not yet fully developed sensory organs and locomotor system were never brought home during the study.] Nestlings were determined by us or the cat owners (if rescued or eaten by the cat) or from photos sent in.
Since cats are known to feed on carcasses (Welti et al., 2020), we determined the cause of death in the dead birds collected by the cat owners by a veterinarian (P. Mattmann). Specifically, if a bird exhibited fractures of the skull bones or hemorrhage into the brain or air sacs, liver lacerations, or fractured coracoid bones without bite marks being present, we interpreted this bird to have suffered blunt force trauma prior to falling prey to the cat. Blunt force trauma could have occurred e.g., due to collision with a window and was interpreted to have left the bird either already dead or injured, in any case impaired to fly and react on the BBScc and/or the bell. Other causes of death prior to be taken by the cat could not be excluded and hence may lower the effectivity estimates of the BBScc and the bell.
We considered birds that were recorded by the cat owners to have been brought home during the hours of darkness [i.e., during the night or the darker phases of twilight (i.e., astronomical twilight or nautical twilight)] as being hunted during darkness when the BBScc could not have been effective. We assumed that prey was usually promptly brought home by the cats, because caching prey is very rare (Fitzgerald and Turner, 2000; Ruiz-Villar et al., 2020) and cats caching prey prior to bringing it home has not been reported, e.g., in studies investigating prey killed vs. brought home using cameras (Loyd et al., 2013; Seymour et al., 2020) as well as in an extensive review article on cat hunting behavior (Fitzgerald and Turner, 2000). Due to these considerations (excluding nestlings, scavenging, and hunting at night), the here estimated efficacy of the BBScc is indeed a conservative estimate of the efficacy of “preventable” predation. The exclusion of bird prey items hunted during situations when the BBScc could not have been effective was similar for the control-phase and treatment 1 (too few data for treatment 2), as shown by a non-parametric Wilcoxon pairwise signed-rank test applied to the paired numbers of excluded birds in the control and treatment 1 phase of each household (W = 86, p = 0.131).
The number of birds brought home during the first vs. the second week of wearing the BBScc (either during treatment 1 or 2, whichever came first) was similar (n = 10 birds in both weeks; Wilcoxon signed-rank test: W = 12, p = 0.748). Therefore, an influence of the novelty of the device on the cat’s behavior and therefore likeliness of bringing home prey was negligible and we analyzed the 2 weeks together.
To measure the effects of the treatments on the number of prey brought home we used a Bayesian generalized linear mixed model. The number of prey items brought home during each 2-weeks treatment-phase in each household was used as outcome variable. Treatment and prey taxon and the interaction between treatment and prey taxon were used as predictors. The household ID was included as a random factor to account for repeated measures of the same household. We further used the product of the treatment-phase duration (which ranged from 11 to 26.5 days) and the number of cats in the households as an offset. This offset corrects for the fact that the duration of the treatment phases varied somewhat and the number of cats was different among the households. We first used the Poisson and the negative binomial distributions. However, both models overestimated the proportion of zeros and underestimated the variance in the data. Therefore, we used a zero-inflated Poisson model for which the predictive model checking (Gelman et al., 1996; Gabry, 2018) showed a good fit regarding mean, variance and range of the data and proportion of zeros. We allowed for separate proportions of zeros for each treatment and prey taxon. We fitted the model using Hamiltonian Monte Carlo as implemented in Stan (Carpenter et al., 2017). We accessed the Stan software via the interface brms (Bürkner, 2017) within the statistical software R 4.0.5 (R Core Team, 2021). Further, we used the default prior distributions defined by the brms-interface, which are flat priors for the model coefficients and weakly informative prior distributions (Lemoine, 2019) for the intercepts and the variance parameters, i.e., student-t(3, –2.3, 2.5) for the intercept of the count model, logistic (0, 1) for the intercept of the zero-model, and half-student-t(3, 0, 2.5) for the among-household variance (Gelman, 2006). Finally, we assessed convergence of the four Markov chains using the diagnostic plots and statistics given by Gabry (2018). Estimates (medians) and 95% uncertainty intervals (2.5% and 97.5% quantiles) are based on 4,000 simulated values from the posterior distributions of parameters.
Participants were asked to fill in four polls: one prior to the start of the survey, one at the beginning, one during, and one after finishing the survey. We asked for age, neutering status, and coat color and pattern of the survey cats, the cat’s behavior when first wearing the BBScc, and the cat owner’s own and their social environment’s (e.g., neighbors, family, friends) attitude toward the BBScc (Supplementary Table 2).
The study protocol was approved by the ethics committee of the Veterinary Office of the Canton of Zurich, Switzerland (TV ZH098/20).
Ninety-nine cat-owners were initially interested in participating in this study. Cats in 43 households with 59 study cats started the survey, but 17 household ended it prematurely. Four households (9.3%) with eight cats quit the study due to issues related to the BBScc: two households reported a lack of habituation, one household reported aggressive behavior of cats toward each other, and one household reported continuing and excessive scratching. The other households quit the study due to reasons unrelated to the study itself (Supplementary Data Sheet 1). Finally, 26 households with 31 study cats (17 males, 14 females; all but one neutered prior to this study) completed the survey. 12 of them tested the bell in addition to the BBScc (treatment 2). The 31 study cats were monitored a total of 1,000 days between May and October 2019 (pilot phase) and 2020 (main study).
All birds brought home by the study cats were passerines (for a list see Supplementary Table 3). Of 40 birds brought home that could be aged post-mortem from cadavers or photographs, 90% were full-grown, i.e., able to fly (17 first-year birds, five adults, and 14 of undetermined age, but not nestlings), and 10% were nestlings (n = 4). The majority of identifiable birds were house sparrows (Passer domesticus; n = 18; 50%), followed by great tits (Parus major; n = 8; 22%) (Supplementary Table 3). (All mammals brought home were either muroid rodents or shrews and no early post-natal juveniles with not yet fully developed sensory organs and locomotor system were brought home).
Of 15 non-nestling bird cadavers that were subject to post-mortem dissection, four (27%, Supplementary Table 3) were classified as having collided with a glass window or another hard object prior to having fallen victim to a cat, based on the identified injuries: two birds were brought home by cats wearing neither BBScc nor bell (control), one bird was brought home by a cat wearing a BBScc and no bell (treatment 1), and one bird was brought home by a cat wearing a BBScc and a bell (treatment 2).
Five birds had to be excluded because they were brought home during the hours of darkness: one during the control-phase, three during treatment 1 and one during treatment 2. Four birds had to be excluded because they were nestlings: three during the control-phase, none during treatment 1 and one during treatment 2.
Numbers of prey brought home (excluding nestlings, scavenged birds, and birds brought home during the dark hours) broken up into the three study phases and four prey taxa are given in Table 1. The model’s expected numbers of prey per cat and 2-week treatment-phase for every prey taxon are given in Figure 2. We found evidence that BBScc reduces the number of birds brought home on average by −37% (factor 0.63), with a probability of reduction of 88% (Table 2 and Figures 2, 3). A similar result was found comparing the control-phase with the phase when both BBScc and the bell were worn (Table 2 and Figures 2, 3). In contrast, we found no evidence that the additional bell had an effect on the numbers of birds brought home, compared to the phase with a BBScc alone (Table 2 and Figures 2, 3).
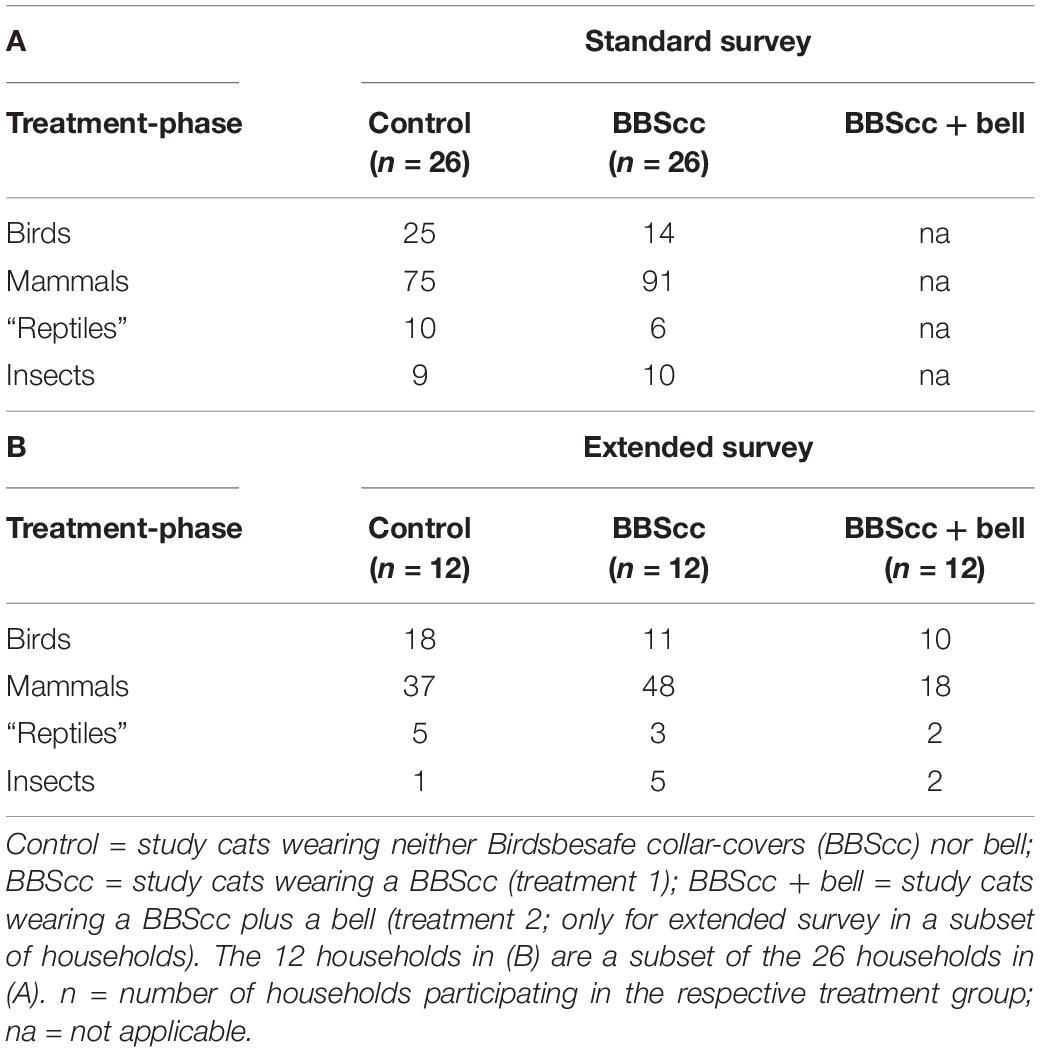
Table 1. Number of prey brought home (excluding nestlings, scavenged birds, and birds brought home during the dark hours), (A) during the phases of the standard survey and (B) the phases of the extended survey.
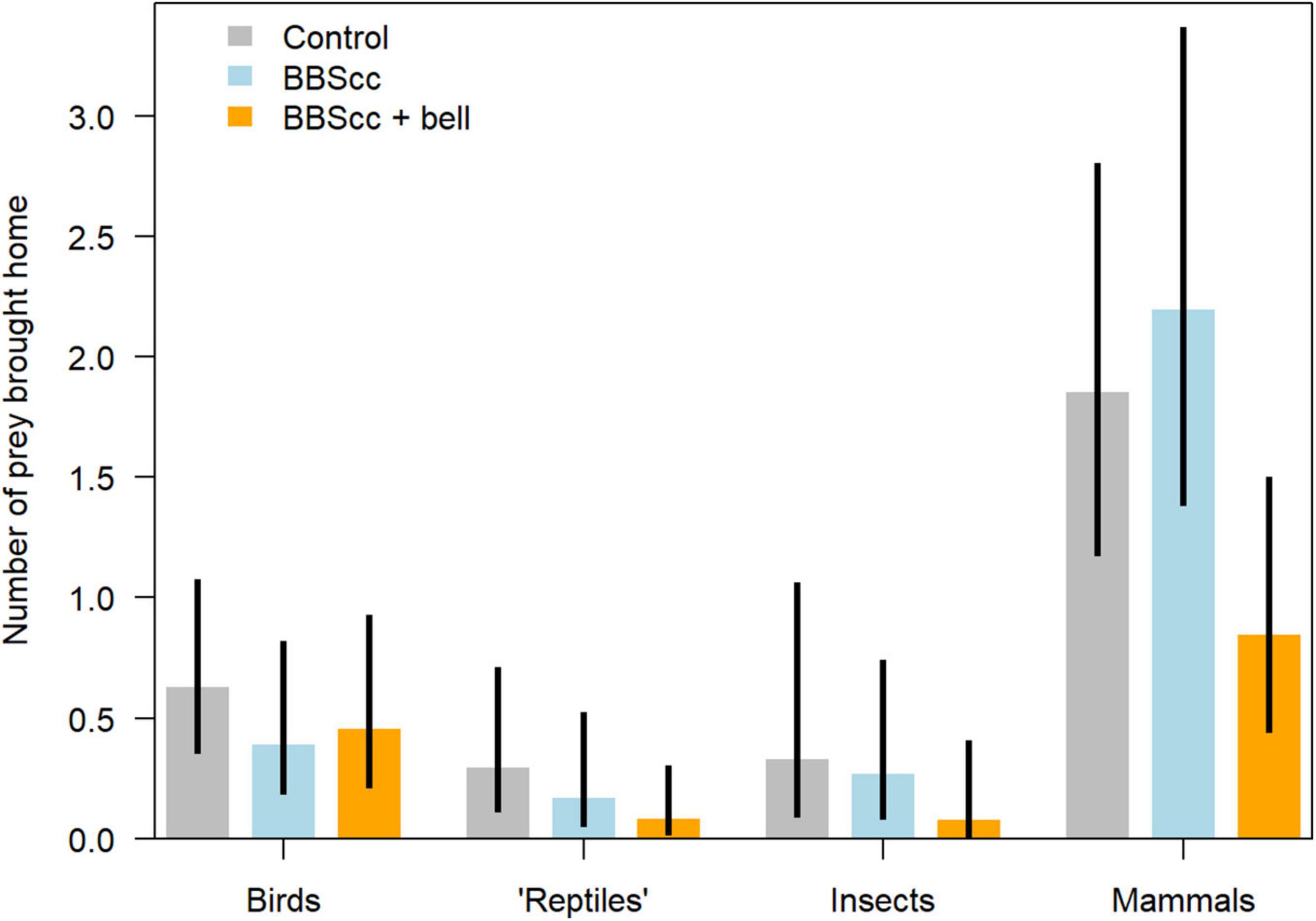
Figure 2. Expected number of prey brought home per household in each 2-week treatment phase and separated per prey taxon (birds, “reptiles” insects, and mammals). Lower and upper limits of the 95% uncertainty interval are indicated with vertical black bars [equals “credible interval” or “confidence interval”; Gelman and Greenland (2019)]. BBScc = wearing a Birdsbesafe collar-cover (BBScc) during treatment 1; BBScc + bell = wearing a BBScc and a bell during treatment 2; control = wearing neither BBScc nor bell.
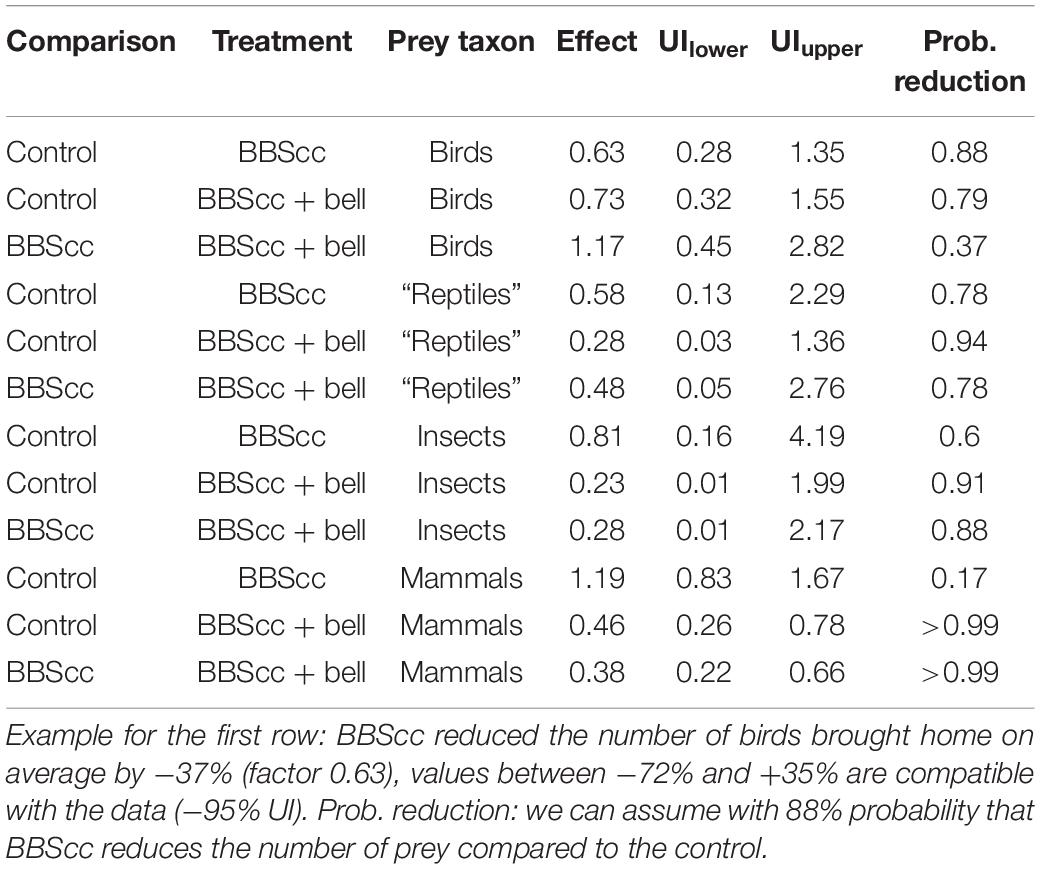
Table 2. Multiplicative effects of the Birdsbesafe collar-cover (BBScc) and BBScc + bell treatments compared to the control (without BBScc and bell); or of BBScc + bell also compared to BBScc, with upper (UIupper) and lower (UIlower) limits of the 95% uncertainty interval (UI).
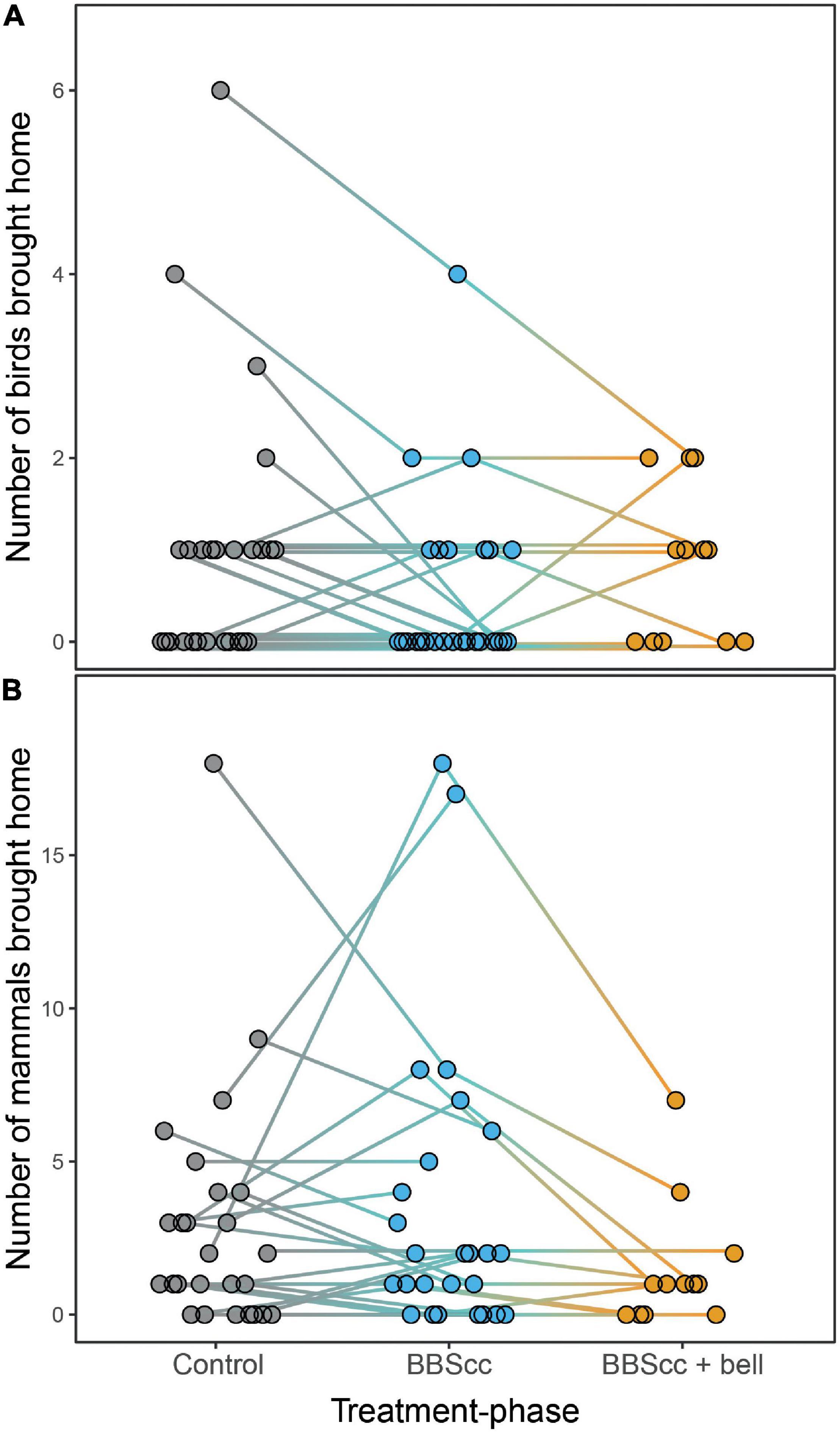
Figure 3. Number of birds (A) and mammals (B) brought home by 2-week treatment-phase. Dots are individual prey numbers per household. Lines between treatment phases connect data of the same household. Control = study cats wearing neither Birdsbesafe collar-covers (BBScc) nor bell; BBScc = study cats wearing a BBScc (treatment 1); BBScc + bell = study cats wearing a BBScc plus a bell (treatment 2; only for extended survey in a subset of households). For sample sizes see Table 1. For individual prey numbers per cat, see Supplementary Data Sheet 2.
In mammals, we found no evidence that the BBScc had an effect on the number of mammals brought home compared to the control-phase without BBScc (Table 2 and Figures 2, 3). However, an additional bell did reduce the number of mammals brought home by −54 or −62% compared to the control-phase or the phase with a BBScc alone, respectively (Table 2 and Figures 2, 3). The probability for this reduction was >99% in both cases (Table 2). For “reptiles” and insects, reductions of prey brought home due to the BBScc and the bell was on average about—55%, with a probability of reduction of on average 81% (Table 2 and Figures 2).
From the 26 households that successfully finished the study (27 answers due to two answers in one of the households with >1 cat), the majority (n = 19, 70.4%) reported that their cats habituated to the BBScc within a couple of hours up to 1 day after starting the study. In seven cases (25.9%) the habituation period took 1 day to 1 week, and in one household (3.7%) the habituation took up to 2 weeks.
In the households that finished the study successfully (n = 26, >1 statement per household possible), eight households (30.8%) reported no behavioral problems related to the BBScc whatsoever and 11 (42.3%) reported more frequent scratching incidents of the cat(s) in the neck region due to the BBScc (Figure 4). Less frequent behavioral issues are reported in Figure 4 and the Supplementary Data Sheet 1 (Supplementary Table 4). These issues were not severe enough for the cat owners to stop the survey prematurely and were mostly related to the initial habituation phase.
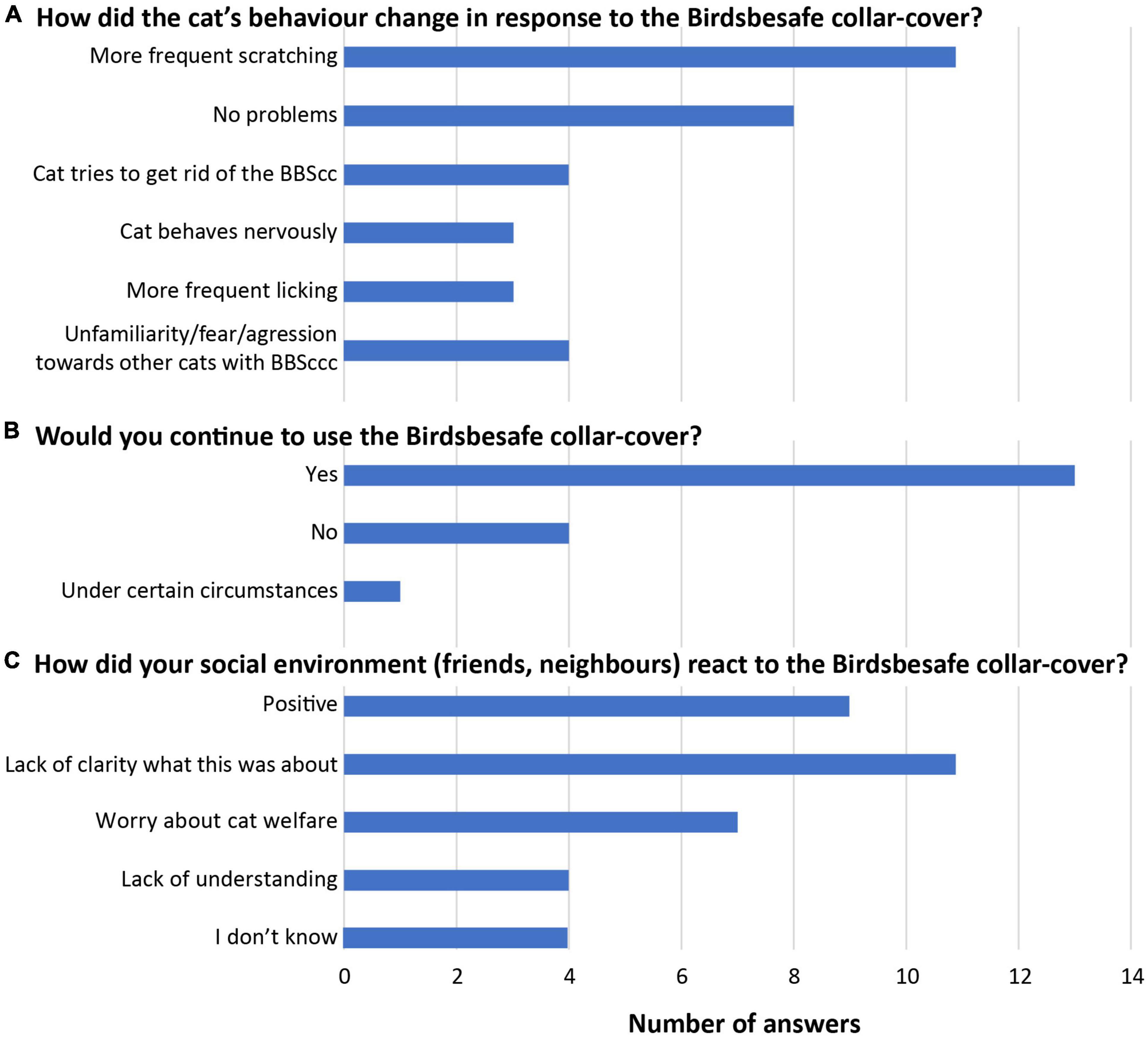
Figure 4. Results of cat behavior (A) and cat owner (B,C) polls. (A) Answers from the 26 households that successfully finished the study (more than 1 answer per household possible). (B,C) Answers from a final survey that was filled in by 18 participants, including some which terminated the survey prematurely, and with multiple answers possible.
From a final survey (which was filled in by 18 of the participants, including some which terminated the survey prematurely, and with multiple answers possible), 44.4% of households thought that the BBScc was working, while 11.1% thought it did not work (44.4% were not sure or thought the BBScc would work in certain circumstances).
The majority of households stated that they were ready to pay the customary price for the BBScc (65% of households) and that they would be willing to continue to use the BBScc after the survey, either during the entire year or through part or the entire breeding season of birds (72% of households) (Figure 4). For households stating that they would not use the BBScc after the survey (22% of households), the main reason was the unusual behavior of their cat(s) toward the BBScc (including frequent scratching).
A large fraction of households (33%) reported their social environment (e.g., neighbors, family, and friends) to be puzzled at first sight by the colorful collar (Figure 4). 27% of households reported positive reactions from their surroundings, while 21% reported also negative reactions (e.g., worry concerning animal welfare) (Figure 4). 12% of households further reported a lack of understanding in their surroundings (Figure 4).
Our study set out to quantify the efficacy of Birdsbesafe collar-covers (BBScc, Figure 1) and bells for reducing domestic cats’ predation rate (measured via prey brought home) in a Continental European setting. Our results suggest that the BBScc reduced the number of birds brought home by cats (Table 2 and Figures 2, 3). There was no evidence that the additional bell had an effect. On the other hand, we found that the BBScc alone has probably no effect on mammals, while the additional bell reduced the number of mammals brought home by cats (Table 2 and Figures 2, 3). For “reptiles” and insects, the sample size was too small and the uncertainty interval too large to allow conclusions. This pattern might be explained by the different sensory abilities of birds and small mammals, respectively [e.g., Portfors (2007); Westheide and Rieger (2009)]: color vision is more differentiated in birds compared to mammals, while the hearing ability in the ultrasonic spectrum—which is emitted by the bells (see Materials and Methods and Supplementary Figure 2)—is better in mammals than birds. Also, birds may be more often in direct visual contact with an approaching cat than small mammals in the ground or in ground vegetation.
Our study sample is limited—also due to the difficulties to acquire participants—but is within the range of previously published reports on the efficacy of the BBScc. Our results correspond with these previous studies in that all of them reported a probably substantial reduction in the number of birds brought home when the BBScc was worn (Hall et al., 2015; Willson et al., 2015; Pemberton and Ruxton, 2020; Cecchetti et al., 2021a,b). On the other hand, the magnitude of the effect of the BBScc was estimated to be quite different among the studies. The reasons for this disparity are unclear, but might be related to methodological differences (including applied statistics), as well as differences in surveyed habitats and species communities, which might influence the general effectiveness of the BBScc. However, overall, the results of these studies suggest that across continents and habitats, the BBScc has the potential to reduce the number of birds (but not mammals) brought home by −37–95%.
The expected number of birds brought home per household in each 2-week treatment phase was estimated to be 0.63 without the BBScc and 0.39 while wearing the BBScc (Figure 2). In Switzerland, it has been estimated that in every spring month 0.1 to 0.3 million birds are killed by owned domestic cats based on a sample of average hunters (not particularly active hunters, such as in the present study) and taking into account that not all cats are hunting birds and have access to the outdoors (Tschanz et al., 2011). We could now assume that the −37% reduction in birds brought home (“preventable” predation, Table 2) due to the BBScc directly translates into a reduction of −37% in birds killed by cats due to the BBScc. If all owned cats in Switzerland with access to the outdoors would wear a BBScc, this would ideally translate into 74’000 less birds being killed by cats per spring-month in Switzerland, or 296’000 in the course of the 4 months of the main breeding season. This is of course a too optimistic estimate, because not all cats will habituate to the collar and/or the collar-cover, and not all cat owners will be willing to deploy these measures (Figure 4). Further, stray cats without an owner will not be accessible for collars. However, the estimate still facilitates to grasp the potential of these measures.
Such extrapolations should be interpreted with caution due to uncertainties related to the initial estimation of birds killed by cats (Tschanz et al., 2011), the non-inclusion of nestlings and injured birds, as well as the lack of knowledge about the cats’ actual impact on wildlife at the population level. Simultaneous examinations of demographic variation of the wildlife populations would be warranted, also considering confounding variables such as source-sink dynamics, (apparent) competition, diseases, food availability, habitat quality and availability, and pollution (Barratt, 1997, 1998; Crooks and Soulé, 1999; Fitzgerald and Turner, 2000; Kays and DeWan, 2004; Grendelmeier et al., 2018), as well as the non-lethal impacts of cats on prey populations [landscape of fear; Brown et al. (1999), Beckerman et al. (2007), and Bonnington et al. (2013)]. Further, it should be determined if the effect of predation by domestic cats is additive or compensatory to other sources of mortality (Møller and Erritzøe, 2000; Baker et al., 2008; van Heezik et al., 2010). We did not examine the physical condition of birds brought home by study cats, apart from the cause of death. As a high fraction of the birds brought home by study cats consisted of first-year birds, which generally suffer from relatively high mortality rates [e.g., Naef-Daenzer et al. (2001) and Weggler and Leu (2001)], the cats’ effect on the breeding populations might be indicative of a compensatory form of mortality. However, additive mortality would most likely increase when predation pressure increases (Baker et al., 2008; van Heezik et al., 2010). The high relative number of prey incidents recorded among first-year birds indicate that it would be most recommendable to deploy BBScc at least during the main breeding season of birds.
An additional aspect that requires further study is potential (long-term) habituation effects. Wearing a novel collar-cover may change the behavior of a cat and make it less likely to hunt (although we found no evidence for this here, see above). Further, it has been speculated that with time, cats that are regularly equipped with bells might adapt their hunting strategies in order to overcome the adverse effects of the bells on their stealth. This has not been tested yet. Similarly, cats regularly wearing a BBScc might learn that hunting during twilight or the night (if given the chance by their owners), or the selection of hunting habitats that provide better visual hiding, might enhance their hunting success.
Most of the bird species brought home by cats in this study (except the green finch, which is categorized as “Near Threatened” (NT); Supplementary Table 3) are currently listed as least concern (LC) according to the red list of breeding birds in Switzerland (Knaus et al., 2021). All of the species are internationally categorized as LC (IUCN, 2021). Most birds brought home were house sparrows (Passer domesticus) and great tits (Parus major). Birds feeding on the ground and in low vegetation—as the house sparrow and the great tit–have been reported to often be the victims of domestic cats (Mead, 1982; Churcher and Lawton, 1987; Pavisse et al., 2019), although the year-round gregarious house sparrow might be less prone to cat predation compared to solitary species (Mead, 1982). These species’ frequency as cat victims might also be related to their relatively high abundance in settlement areas (Ineichen et al., 2012). Further aspects that might play a role in the unequal frequency of bird species as prey are body size and related feeding habits, with larger ground feeding birds—like the blackbird (Turdus merula)—being less likely to feed close to cover and therefore less likely to be depredated by stalking cats (Mead, 1982). Lastly, different bird species have been speculated to vary in taste, with less tasty birds—such as the house sparrow—being more likely to be brought home and not be eaten in situ (Fitzgerald and Turner, 2000).
It is important to note that the number of prey brought home by cats almost certainly underestimates the total number of prey killed by cats. Studies applying collar-mounted cameras found that only about 20% of prey killed by cats were actually brought home (Loyd et al., 2013; Seymour et al., 2020), while the rest was eaten or left behind in situ. However, there is no reason to suspect that the BBScc or the bell change the rate of prey brought home [Cecchetti et al. (2021b); also because—contrary to the main trend—the number of prey brought home in a few households increased during the phase when BBScc (and the bell) was worn; Figure 3]. Additionally, a recent study investigating nutrition specific nitrogen stable isotope ratios in whiskers of owned cats showed that wearing a BBScc also reduced the actual consumption of wild prey (Cecchetti et al., 2021a). Thus, we consider the estimate of “prey brought home” to be stable across study phases and an accurate index of hunting activity and success.
On the other hand, cats are known to scavenge (Welti et al., 2020) and our results show that a considerable fraction of birds brought home by cats (27% of the birds that could be dissected) was probably already seriously injured or dead prior to being brought home by the cat (e.g., because they collided with a window). These birds could not have reacted on the BBScc (and the bell) and the number of birds brought home is subsequently overestimated. Our sample size regarding these results was small (four birds out of 15 dissected probably did collide with a window before being brought home by cats). However, our findings highlight that the measure of “prey brought home” at the same time overestimates somewhat the actual predation rate of cats, although the overall rate might still be underestimated (see previous paragraph). Considerations on cat predation rates based on prey brought home should consider this. Further investigations into the likeliness of cats bringing home prey depending on the state of the prey (alive, dead) would also be worthwhile in this respect.
No cats were injured due to the quick-release collar or the BBScc during the study phase. The BBScc appears to be a welfare issue for only a small number of study cats, while most were habituating quickly to it. High rates of tolerance have also been reported by a similar study (Willson et al., 2015). Our experience suggests that many of these issues may be overcome if the cat is slowly familiarized with the BBScc and/or the quick-release collar underneath prior to their first use. In our experimental design we could not determine whether behavioral salience was due to the BBScc or the quick-release collar underneath (see Materials and Methods section, Figure 1), especially in cats which do not usually wear a collar. However, some issues related to the BBScc, such as frequent scratching (especially when wet), might, together with the clownish appearance of the BBScc to many humans, limit its widespread use among cat owners.
The unusual appearance of the BBScc might be the reason why it often meets with incomprehension among people. However, most cat owners are interested in reducing predation of their pet cats by some means (Crowley et al., 2020), although perceived risks to their pets might reduce the owners’ acceptance of collar mounted devices in general (Thomas et al., 2012). Therefore, widespread public awareness campaigns to make the function of BBScc better known, as well as alternative and/or supplementing actions compliant with high animal welfare standards, might pose a productive and constructive avenue to decrease hunting activity and predation rates by domestic cats, without fueling conflicts among cat owners and conservationists (Crowley et al., 2020). For example, a recent study suggests that frequent playing with the cat and high protein content of the cat diet can decrease the predation rate (Cecchetti et al., 2020, 2021b). Although this somewhat contrasts with previously presented evidence that preying takes precedence over food consumption (Adamec, 1976), such measures could be considered as complements to the use of collar mounted devices, if protein rich cat food can be produced sustainably (Okin, 2017). It would be particularly valuable to apply such measures in areas that are habitats of endangered species, including urban areas (Ineichen et al., 2012; Legge et al., 2020). As an—yet to study—asset for the promotion of the BBScc might be its fluorescent rim, which might increase the visibility of the cats for automobilists.
Although one of the studies investigating the BBScc found no influence of the cat’s sex, age, and home location on variation of prey mortality with and without BBScc (Willson et al., 2015), it is still to be investigated whether other confounding variables such as e.g., body weight and coat length might influence the effect of the BBScc. Specifically, the BBScc might be less effective in large cats with long fur, where the BBScc stands out less conspicuously. Our sample size does not allow to account for such confounding variables and further studies with an increased sample size might better inform about their potential influence. Further studies might also investigate an improved BBScc design. This could include an improved consideration of the visual and auditory capabilities of different prey taxa in relation to the taxon-specific hunting strategy of cats, as well as the habitat where the cats are hunting (e.g., more light and therefore higher likeliness of the BBScc functioning also by night in more urbanized areas). Effects of habituation (also long term) should be considered. This could be accomplished via a longer habituation period to the collar and the collar-cover, as well as the usage of plain black collars during the control-phase. Combinations of different, improved collar mounted devices, together with easy to implement measures at home (e.g., playing with the cat, feeding high-protein cat food) would be avenues of research worthwhile pursuing to explore all possibilities to optimize protective measures to reduce the impact of domestic cat predation.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
The animal study was reviewed and approved by the Veterinary Office of the Canton of Zurich, Switzerland (TV ZH098/20). Written informed consent was obtained from the owners for the participation of their animals in this study.
CK, DH, FB, LJ, and MG conceptualized the study. MG collected the data with the cat owners and wrote the manuscript. PM analyzed the bird cadavers and photographs. All authors revised the manuscript and gave final approval for publication.
This work was financially supported by the Swiss Ornithological Institute, Sempach (Switzerland) and the Wolfermann-Nägeli-Foundation (Switzerland).
We purchased Birdsbesafe collar-covers from the official Swiss distributor Pik Pik (pikpik.ch) with a kindly granted discount. However, the authors have no financial ties to these companies. Neither developer nor distributer of the Birdsbesafe collar-covers and the bells were involved in the study design, data analysis, and manuscript writing.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank the participating cat owners and their cats for taking part in this study, naturschutz.ch and the Zürcher Tierschutz for help with the search for participants, the Zürcher Tierschutz and the Malou-Foundation for support during the preparation of this project, the Swiss Ornithological Institute, Sempach for the contribution of conceptual, logistical, and financial support, the Wolfermann-Nägeli-Foundation for financial support, Pik Pik (pikpik.ch) and the Schweizerische Tiermeldezentrale (stmz.ch) for help and support with the acquisition of materials, Valeria Renna and Timia Sanchez Alcocer for help with data gathering, Fränzi Korner-Nievergelt for help with the analyses, Mirco Lauper and Kathi Märki for help with the frequency-analysis of the bells, Fränzi Korner-Nievergelt, Gilberto Pasinelli, Hans Schmid, Stefan Werner, Michael Schaad, and Daniela Heynen for helpful comments to improve the manuscript, Susan Willson, Martin Weggler, Martina Cecchetti, and Robbie A. McDonald for discussion, Mathew Samuel Crowther for editorial work, and John Read, Emiliano Mori, and Jenny Mattisson for valuable input to improve the manuscript.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.850442/full#supplementary-material
Adamec, R. E. (1976). The interaction of hunger and preying in the domestic cat (Felis catus): an adaptive hierarchy? Behav. Biol. 18, 263–272. doi: 10.1016/S0091-6773(76)92166-0
Baker, P. J., Ansell, R. J., Dodds, P. A. A., Webber, C. E., and Harris, S. (2003). Factors affecting the distribution of small mammals in an urban area. Mamm. Rev. 33, 95–100. doi: 10.1046/j.1365-907.2003.00003.x
Baker, P. J., Bentley, A. J., Ansell, R. J., and Harris, S. (2005). Impact of predation by domestic cats Felis catus in an urban area. Mamm. Rev. 35, 302–312.
Baker, P. J., Molony, S. E., Stone, E., Cuthill, I. C., and Harris, S. (2008). Cats about town: is predation by free-ranging pet cats Felis catus likely to affect urban bird populations? Ibis Int. J. Avian Sci. 150, 86–99. doi: 10.1111/j.1474-919X.2008.00836.x
Barratt, D. G. (1997). Predation by house cats. Felis catus (L.), in Canberra, Australia. I. Prey composition and preference. Wildl. Res. 24, 263–277. doi: 10.1071/WR96020
Barratt, D. G. (1998). Predation by house cats, Felis catus (L.), in Canberra, Australia. II. Factors affecting the amount of prey caught and estimates of the impact on wildlife. Wildl. Res. 25, 475–487. doi: 10.1071/WR97026
Beckerman, A. P., Boots, M., and Gaston, K. J. (2007). Urban bird declines and the fear of cats. Anim. Conserv. 10, 320–325. doi: 10.1111/j.1469-1795.2007.00115.x
Bonnington, C., Gaston, K. J., and Evans, K. L. (2013). Fearing the feline: domestic cats reduce avian fecundity through trait-mediated indirect effects that increase nest predation by other species. J. Appl. Ecol. 50, 15–24. doi: 10.1111/1365-2664.12025
Brown, J. S., Laundré, J. W., and Gurung, M. (1999). The ecology of fear: optimal foraging, game theory, and trophic interactions. J. Mammal. 80, 385–399. doi: 10.2307/1383287
Bürkner, P.-C. (2017). Brms: an R package for Bayesian multilevel models using Stan. J. Stat. Softw. 80, 1–28. doi: 10.18637/jss.v080.i01
Calver, M., Thomas, S., Bradley, S., and McCutcheon, H. (2007). Reducing the rate of predation on wildlife by pet cats: the efficacy and practicability of collar-mounted pounce protectors. Biol. Conserv. 137, 341–348. doi: 10.1016/j.biocon.2007.02.015
Carpenter, B., Gelman, A., Hoffman, M. D., Lee, D., Goodrich, B., Betancourt, M., et al. (2017). Stan: a probabilistic programming language. J. Stat. Softw. 76, 1–32. doi: 10.18637/jss.v076.i01
Cecchetti, M., Crowley, S. L., Goodwin, C. E. D., McDonald, J., Bearhop, S., McDonald, R. A., et al. (2021a). Contributions of wild and provisioned foods to the diets of domestic cats that depredate wild animals. Ecosphere 12:e03737. doi: 10.1002/ECS2.3737
Cecchetti, M., Crowley, S. L., Goodwin, C. E. D., and McDonald, R. A. (2021b). Provision of high meat content food and object play reduce predation of wild animals by domestic cats Felis catus. Curr. Biol. 31, 1107–1111.e5. doi: 10.1016/j.cub.2020.12.044
Cecchetti, M., Crowley, S. L., and Mcdonald, R. A. (2020). Drivers and facilitators of hunting behaviour in domestic cats and options for management. Mamm. Rev. 51, 307–322. doi: 10.1111/mam.12230
Churcher, P. B., and Lawton, J. H. (1987). Predation by domestic cats in an English village. J. Zool. 212, 439–455. doi: 10.1111/j.1469-7998.1987.tb02915.x
Clark, N. A. (1999). Progress Report on the Effectiveness of the Mark II CatAlert(TM) Collar at Reducing Predation by Domestic Cats. Thetford: British Trust for Ornithology.
Clark, N. A., and Burton, N. H. K. (1998). A Pilot Field Trial into the Effectiveness of the CatAlert(TM) Collar at Reducing Predation by Domestic Cats. Thetford: British Trust for Ornithology.
Cox, J. G., and Lima, S. L. (2006). Naiveté and an aquatic-terrestrial dichotomy in the effects of introduced predators. Trends Ecol. Evol. 21, 674–680. doi: 10.1016/j.tree.2006.07.011
Crooks, K. R., and Soulé, M. E. (1999). Mesopredator release and avifaunal extinctions in a fragmented system. Nature 400, 563–566. doi: 10.1038/23028
Crowley, S. L., Cecchetti, M., and McDonald, R. A. (2020). Diverse perspectives of cat owners indicate barriers to and opportunities for managing cat predation of wildlife. Front. Ecol. Environ. 18:544–549. doi: 10.1002/fee.2254
Doherty, T. S., Glen, A. S., Nimmo, D. G., Ritchie, E. G., and Dickman, C. R. (2016). Invasive predators and global biodiversity loss. Proc. Natl. Acad. Sci. U.S.A. 113, 11261–11265. doi: 10.1073/pnas.1602480113
Federal Office for the Environment (2017). Biodiversität in der Schweiz: Zustand und Entwicklung. Ergebnisse des Überwachungssystems im Bereich Biodiversität, Stand 2016. Bundesamt für. Umwelt, Bern: Umwelt-Zustand Nr 60.
Fitzgerald, B. M., and Turner, D. C. (2000). “Hunting behaviour of domestic cats and their impact on prey populations,” in The Domestic cat: the Biology of its Behaviour, Second Edi Edn, eds D. C. Turner and P. Bateson (Cambridge: Cambridge University Press), 151–175.
Gabry, J. (2018). Shinystan: Interactive Visual and Numerical Diagnostics and Posterior Analysis for Bayesian Models. Available online at: https://cran.r-project.org/package=shinystan (accessed December 9, 2021).
Gelman, A. (2006). Prior distributions for variance parameters in hierarchical models. Bayesian Anal. 1, 515–534.
Gelman, A., and Greenland, S. (2019). Are confidence intervals better termed “uncertainty intervals”? BMJ 366:15381. doi: 10.1136/bmj.l5381
Gelman, A., Meng, X. L., and Stern, H. (1996). Posterior predictive assessment of model fitness via realized discrepancies. Stat. Sin. 6, 733–807.
Gloor, S., Bontadina, F., Hegglin, D., Deplaze, P., and Breitenmoser, U. (2001). The rise of urban fox populations in Switzerland. Mamm. Biol. 66, 155–164. doi: 10.1016/j.vetpar.2015.07.027
Gordon, J. K., Matthaei, C., and Van Heezik, Y. (2010). Belled collars reduce catch of domestic cats in New Zealand by half. Wildl. Res. 37, 372–378. doi: 10.1071/wr09127
Grendelmeier, A., Arlettaz, R., and Pasinelli, G. (2018). Numerical response of mammalian carnivores to rodents affects bird reproduction in temperate forests: a case of apparent competition? Ecol. Evol. 8, 11596–11608. doi: 10.1002/ece3.4608
Hall, C. M., Fontaine, J. B., Bryant, K. A., and Calver, M. C. (2015). Assessing the effectiveness of the Birdsbesafe® anti-predation collar cover in reducing predation on wildlife by pet cats in Western Australia. Appl. Anim. Behav. Sci. 173, 40–51. doi: 10.1016/j.applanim.2015.01.004
Hawkins, C. C., Grant, W. E., and Longnecker, M. T. (2004). “Effect of house cats, being fed in parks, on California birds and rodents”. in Proceedings 4th International Urban Wildlife Symposium (Eds). K. Lisa and W. William Arizona: ShawLarry VanDruff.164–170.
Ineichen, S., Ruckstuhl, M., and Klausnitzer, B. (2012). Stadtfauna: 600 Tierarten Unserer Städte. Bern: Haupt
IUCN (2021). The IUCN Red List of Threatened Species. Version 2021-3. Available online at: https://www.iucnredlist.org. [Accessed on Jan 25, 2022]
Kays, R. W., and DeWan, A. A. (2004). Ecological impact of inside/outside house cats around a suburban nature preserve. Anim. Conserv. 7, 273–283. doi: 10.1017/S1367943004001489
Knaus, P., Antoniazza, S., Keller, V., Sattler, T., Schmid, H., and Strebel, N. (2021). Rote Liste der Brutvögel. Gefährdete Arten der Schweiz. Bundesamt für Umwelt (BAFU); Schweizerische Vogelwarte. Umwelt-Vollzug Nr. 2124.
Legge, S., Woinarski, J. C., Dickman, C. R., Murphy, B. P., Woolley, L. A., and Calver, M. C. (2020). We need to worry about Bella and Charlie: the impacts of pet cats on Australian wildlife. Wildl. Res. 47, 523–539. doi: 10.1071/WR19174
Lemoine, N. P. (2019). Moving beyond noninformative priors: why and how to choose weakly informative priors in Bayesian analyses. Oikos 128, 912–928. doi: 10.1111/oik.05985
Liberg, O., Sandell, M., Pontier, D., and Natoli, E. (2000). Density, Spatial Organisation and Reproductive Tactics in the Domestic Cat and other Felid. in: The Domestic Cat: the Biology of its Behaviour. Cambridge: Cambridge University Press, 119–147.
Loss, S. R., and Marra, P. P. (2017). Population impacts of free-ranging domestic cats on mainland vertebrates. Front. Ecol. Environ. 15:502–509. doi: 10.1002/fee.1633
Loss, S. R., Will, T., and Marra, P. P. (2013). The impact of free-ranging domestic cats on wildlife of the United States. Nat. Commun. 4:1396. doi: 10.1038/ncomms2380
Loyd, K. A. T., Hernandez, S. M., Carroll, J. P., Abernathy, K., and Marshall, G. J. (2013). Quantifying free-roaming domestic cat predation using animal-borne video cameras. Biol. Conserv. 160, 183–189. doi: 10.1016/j.biocon.2013.01.008
Lynn, W. S., Santiago-Ávila, F., Lindenmayer, J., Hadidian, J., Wallach, A., King, B. J., et al. (2019). A moral panic over cats. Conserv. Biol. 33, 769–776. doi: 10.1111/cobi.13346
Maronde, L., McClintock, B. T., Breitenmoser, U., and Zimmermann, F. (2020). Spatial capture–recapture with multiple noninvasive marks: an application to camera-trapping data of the European wildcat (Felis silvestris) using R package multimark. Ecol. Evol. 10, 13968–13979. doi: 10.1002/ece3.6990
Mead, C. J. (1982). Ringed birds killed by cats. Mamm. Rev. 12, 183–186. doi: 10.1111/j.1365-2907.1982.tb00014.x
Medina, F. M., Bonnaud, E., Vidal, E., Tershy, B. R., Zavaleta, E. S., Donlan, C. J., et al. (2011). A global review of the impacts of invasive cats on island endangered vertebrates. Glob. Chang Biol. 17, 3503–3510. doi: 10.1111/j.1365-2486.2011.02464.x
Møller, A. P., and Erritzøe, J. (2000). Predation against birds with low immunocompetence. Oecologia 122, 500–504. doi: 10.1007/s004420050972
Mori, E., Menchetti, M., Camporesi, A., Cavigioli, L., De Fatis, K. T., Girardello, M., et al. (2019). License to kill? Domestic cats affect a wide range of native fauna in a highly biodiverse mediterranean country. Front. Ecol. Evol. 7:477. doi: 10.3389/fevo.2019.00477
Naef-Daenzer, B., Widmer, F., and Nuber, M. (2001). Differential post-fledging survival of great and coal tits in relation to their condition and fledging date. J. Anim. Ecol. 70, 730–738. doi: 10.1046/J.0021-8790.2001.00533.X
Nelson, S. H., Evans, A. D., and Bradbury, R. B. (2005). The efficacy of collar-mounted devices in reducing the rate of predation of wildlife by domestic cats. Appl. Anim. Behav. Sci. 94, 273–285. doi: 10.1016/j.applanim.2005.04.003
Nussberger, B., and Roth, T. (2020). Bericht Wildkatzenmonitoring Schweiz: Verbreitung, Dichte und Hybridisierung der Wildkatze in der Schweiz. Ergebnisse der zweiten Erhebung 2018/20. Switzerland: Wildtier Schweiz.
Okin, G. S. (2017). Environmental impacts of food consumption by dogs and cats. PLoS One 12:e0181301. doi: 10.1371/journal.pone.0181301
Pavisse, R., Vangeluwe, D., and Clergeau, P. (2019). Domestic cat predation on garden birds: an analysis from European ringing programmes. Ardea 107, 103–109. doi: 10.5253/arde.v107i1.a6
Pemberton, C., and Ruxton, G. D. (2020). Birdsbesafe® collar cover reduces bird predation by domestic cats (Felis catus). J. Zool. 310, 106–109. doi: 10.1111/jzo.12739
Portfors, C. V. (2007). Types and functions of ultrasonic vocalizations in laboratory rats and mice. J. Am. Assoc. Lab Anim. Sci. 46, 28–34.
Price, T. D. (2017). Sensory drive, color, and color vision. Am. Nat. 190, 157–170. doi: 10.1086/692535
R Core Team. (2021). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing
Ruiz-Villar, H., López-Bao, J. V., and Palomares, F. (2020). A small cat saving food for later: caching behavior in the European wildcat (Felis silvestris silvestris). Eur. J. Wildl. Res. 66:76.
Ruxton, G. D., Thomas, S., and Wright, J. W. (2002). Bells reduce predation of wildlife by domestic cats (Felis catus). J. Zool. 256, 81–83. doi: 10.1017/s0952836902000109
Salo, P., Korpimäki, E., Banks, P. B., Nordström, M., and Dickman, C. R. (2007). Alien predators are more dangerous than native predators to prey populations. Proc. R. Soc. B Biol. Sci. 274, 1237–1243. doi: 10.1098/rspb.2006.0444
Seymour, C. L., Simmons, R. E., Morling, F., George, S. T., O’Riain, J. M., Peters, K., et al. (2020). Caught on camera: the impacts of urban domestic cats on wild prey in an African city and neighbouring protected areas. Glob. Ecol. Conserv. 23:e01198. doi: 10.1016/j.gecco.2020.e01198
Shaw, L. M., Chamberlain, D., and Evans, M. (2008). The House Sparrow Passer domesticus in urban areas: reviewing a possible link between post-decline distribution and human socioeconomic status. J. Ornithol. 149, 293–299. doi: 10.1007/s10336-008-0285-y
SWILD (2011). Bericht zur Pilotphase des Projekts, Katzenspur - Auswirkungen der Prädation von Katzen und Wege zur Konfliktminimierung. Interner Schlussbericht an die Malou-Stiftung für Tierschutz. Zürich : Zürcher Tierschutz.
Taucher, A. L., Gloor, S., Dietrich, A., Geiger, M., Hegglin, D., Bontadina, F., et al. (2020). Decline in distribution and abundance: urban hedgehogs under pressure. Animals 10:1606. doi: 10.3390/ani10091606
Thomas, R. L., Fellowes, M. D. E., and Baker, P. J. (2012). Spatio-temporal variation in predation by urban Domestic cats (Felis catus) and the acceptability of possible management actions in the UK. PLoS One 7:e49369. doi: 10.1371/journal.pone.0049369
Trouwborst, A., McCormack, P. C., and Martínez Camacho, E. (2020). Domestic cats and their impacts on biodiversity: a blind spot in the application of nature conservation law. People Nat. 2, 235–250. doi: 10.1002/pan3.10073
Tschanz, B., Hegglin, D., Gloor, S., and Bontadina, F. (2011). Hunters and non-hunters: skewed predation rate by domestic cats in a rural village. Eur. J. Wildl. Res. 57, 597–602. doi: 10.1007/s10344-010-0470-1
van Heezik, Y., Smyth, A., Adams, A., and Gordon, J. (2010). Do domestic cats impose an unsustainable harvest on urban bird populations? Biol. Conserv. 143, 121–130. doi: 10.1016/j.biocon.2009.09.013
Weggler, M., and Leu, B. (2001). Eine Überschuss produzierende Population des Hausrotschwanzes (Phoenicurus ochruros) in Ortschaften mit hoher Hauskatzendichte (Felis catus). J. Ornithol. 142, 273–283. doi: 10.1007/bf01651366
Welti, N., Scherler, P., and Grüebler, M. U. (2020). Carcass predictability but not domestic pet introduction affects functional response of scavenger assemblage in urbanized habitats. Funct. Ecol. 34, 265–275. doi: 10.1111/1365-2435.13469
Westheide, W., and Rieger, G. (eds) (2009). Spezielle Zoologie. Teil 2 Wirbel-oder Schädeltiere. Heidelberg: Springer.
Keywords: Felis catus, Birdsbesafe, scavenging, animal welfare, conservation, predation
Citation: Geiger M, Kistler C, Mattmann P, Jenni L, Hegglin D and Bontadina F (2022) Colorful Collar-Covers and Bells Reduce Wildlife Predation by Domestic Cats in a Continental European Setting. Front. Ecol. Evol. 10:850442. doi: 10.3389/fevo.2022.850442
Received: 07 January 2022; Accepted: 17 March 2022;
Published: 25 April 2022.
Edited by:
Mathew Samuel Crowther, The University of Sydney, AustraliaReviewed by:
John Read, University of Adelaide, AustraliaCopyright © 2022 Geiger, Kistler, Mattmann, Jenni, Hegglin and Bontadina. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Madeleine Geiger, bWFkZWxlaW5lLmdlaWdlckBzd2lsZC5jaA==
†These authors have contributed equally to this work and share senior authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.