
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Ecol. Evol., 15 March 2022
Sec. Urban Ecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.850065
This article is part of the Research TopicEcological and Evolutionary Processes in Neotropical Urban EcosystemsView all 8 articles
 Nélida R. Villaseñor1,2*
Nélida R. Villaseñor1,2* Martín A. H. Escobar3
Martín A. H. Escobar3
Billions of wild animals migrate every year between breeding and non-breeding sites, facing new environmental conditions due to anthropogenic change. Although cities engender profound environmental changes, they provide habitat for some migratory birds contributing to conserving biodiversity and restoring human-nature connections. A common issue in Neotropical cities is their socio-environmental segregation, that might result in wealthier people having access to greater biodiversity in their neighborhoods. Although growing evidence shows a relationship between wealth and biodiversity in cities, it remains poorly investigated in relation to animals, particularly migratory birds. We present the first study on the effects of socioeconomic status on a Neotropical austral migrant considered a keystone species. We tested whether neighborhood socioeconomic category and habitat variables relate to the occurrence of the Green-backed Firecrown hummingbird in its overwintering habitat in urban areas. We hypothesized that higher socioeconomic status of neighborhoods would positively influence hummingbird occurrence through mediating effects of woody plant cover. We found that hummingbird occurrence increased with socioeconomic level of neighborhoods and woody cover. Specifically, we found that socioeconomic level influenced hummingbird occurrence indirectly through woody and shrub cover. We also found a direct effect of socioeconomic level on hummingbird occurrence suggesting the need for further research. Our findings show that wealthier neighborhoods exhibit greater woody cover and higher hummingbird occurrence than poorer neighborhoods, providing dissimilar opportunities to experience nature close to home. In neighborhoods where people of lower socioeconomic status live, strategies aiming to increase tree and shrub cover will promote a more environmentally just city.
Billions of wild animals migrate every year between breeding and non-breeding sites (Robinson et al., 2009; Dokter et al., 2018). These migrants commonly travel to reproduce in seasonal environments that offer a large amount of food for a short time (Dingle and Drake, 2007; de Zoeten and Pulido, 2020). Such massive animal movements not only benefit migrating animals but also underpin ecosystem functions along migration routes (e.g., Kauffman et al., 2021).
This adaptation to natural resource fluctuation in space and time can be affected by anthropogenic change. Migrant behavior is rapidly changing due to human impacts on the environment that pose a strong selective pressure on wild populations (de Zoeten and Pulido, 2020). Landscape and climate change can influence species phenology, such as periods of migration and reproduction. For instance, they can cause earlier arrival times (Tryjanowski et al., 2013), decrease distances of migration (Visser et al., 2009), and even lead to the loss of migratory behavior of individuals to become year-round residents (Cubie, 2014; Greig et al., 2017; de Zoeten and Pulido, 2020). Migratory behavior is not changing alone, population sizes are as well. Several long-distance migrants are declining faster than populations of short-distance migrants or resident species (Robinson et al., 2009; Kamp et al., 2015; Koleček et al., 2021).
To preserve migratory birds, their migratory behavior, and ecosystem functioning, it is relevant to understand the ecology of migrants, not only in natural areas, but also in modified environments. Urbanization threatens species with extinction (IPBES, 2019) and commonly affects native species in favor of exotic species (McKinney, 2002; Sol et al., 2020). However, cities offer opportunities to conserve biodiversity and restore human connections with nature (Sanderson and Huron, 2011; Aronson et al., 2014). Although urbanization severely affects Neotropical migratory birds (Stratford and Robinson, 2005), urban vegetation may contribute to support Neotropical birds in cities (MacGregor-Fors and Schondube, 2011; Muñoz-Pacheco and Villaseñor, 2021; Villaseñor et al., 2021) including migratory species (Amaya-Espinel and Hostetler, 2019; Leveau, 2021). In fact, at least fifty-eight species of Neotropical forest migratory birds use urban areas with trees or small forest fragments as stopover and wintering sites, encompassing species that are regarded as interior-forest specialists in their breeding habitats (Amaya-Espinel and Hostetler, 2019).
The Green-backed Firecrown (Sephanoides sephaniodes) hummingbird is a Neotropical austral migratory bird. It is considered a keystone species because it feeds on nectar and, thus, pollinates a large variety of plant species, having a disproportionately large effect on ecosystems. It is part of the Trochilidae family, a taxonomic group endemic to the Americas, which are the main nectarivorous birds on the continent. This hummingbird is endemic to Chile and Argentina, where it is distributed from the Atacama Desert (27°S) to Cape Horn (55°S), and is the most austral hummingbird in the world (Martínez-Piña and González-Cifuentes, 2017). It is classified as “Least Concern” although its population trend is decreasing1. It is a partial migrant, with several individuals breeding in southern areas that in autumn migrate toward northern wintering sites (Martínez-Piña and González-Cifuentes, 2017). It is the only hummingbird present in the southern forests of South America, and a keystone species that pollinates more than 20% of woody plant genera (Aizen et al., 2002). In this mutualistic interaction, woody plants provide fundamental resources for hummingbird survival and reproduction, including food, resting, and nesting sites. Thus, a positive effect of woody cover on this hummingbird has been found in different environments. In austral regions, the Green-backed Firecrown hummingbird is more abundant in mature native forests than in other vegetation types (Díaz et al., 2005; Silva-Rodríguez et al., 2008; Vergara et al., 2014). In modified landscapes such as those dominated by agriculture and industrial tree plantations, the abundance of the Green-backed Firecrown increases with woody cover (Estades and Temple, 1999; Silva-Rodríguez et al., 2008).
During the non-breeding season, the Green-backed Firecrown is common in Central Chile. It can inhabit urban environments, including Santiago, the country’s capital city. Santiago has large inequalities in the distribution of vegetation, where the area of woody plant cover increases with residents’ wealth (Escobedo et al., 2016; Hernández and Villaseñor, 2018). Although a growing body of research has reported the benefits of vegetation to support Neotropical birds in cities (Villegas et al., 2010; Amaya-Espinel and Hostetler, 2019; Villaseñor et al., 2021), the relationship between socioeconomic status and birds is a key knowledge gap in Latin America (MacGregor-Fors and Schondube, 2011; Muñoz-Pacheco and Villaseñor, 2021), particularly on migratory birds.
Biodiversity research in urban environments has increased globally, but few studies have investigated the relationship between socioeconomics and animals (reviewed by Leong et al., 2018). Most animal research has focused on bird diversity, and have shown a positive relationship between human wealth (income) and bird species richness in urban parks, residential areas and municipal districts (Kinzig et al., 2005; Strohbach et al., 2009; Warren et al., 2019). However, this relationship can change depending on the origin of bird species (e.g., native vs. exotics; Loss et al., 2009), the level of urbanization (Chamberlain et al., 2019), or be of little importance (e.g., when compared to habitat variables; Silva et al., 2015).
To contribute to understanding the relationship between socioeconomics and a keystone species, we investigate whether the occurrence of the Green-backed Firecrown in its urban overwintering habitat varies with the socioeconomic level of neighborhoods. To inform habitat management, we also investigated what habitat variables explain the occurrence of the hummingbird in the city. We expected the occurrence of the species would increase with socioeconomic level and woody cover. In addition, due to previous research reporting a positive relationship between socioeconomic level and woody cover, we hypothesized that socioeconomic level would positively influence the Green-backed Firecrown occurrence through a mediating effect of woody cover. The scientific information derived from this study will provide a better understanding of the ecology of Neotropical migratory birds, particularly linking socioeconomics, habitat attributes and the occurrence of the Green-backed Firecrown hummingbird. It also contributes to inform management practices and policies that promote biodiversity in the city and more equitable opportunities for urban residents to experience nature close to home.
Our study took place in the city of Santiago de Chile (33°27′S – 70°40′O). This capital city is comprised of ca. 80,000 ha and harbors over six million inhabitants (INE, 2019). The area has a semi-arid Mediterranean climate, where January is the warmest month (mean average temperature of 21.5°C) and July the coldest month (mean average temperature of 8.2°C; INE, 2015). Rainfall is less than 300 mm per year and is concentrated in winter (INE, 2015), however, an extended drought has been affecting the area since 2010 with rainfall deficits of 20-40% (Garreaud et al., 2020).
The natural landscape has been severely altered by human activities. Original vegetation consisted of sclerophyll forest and shrubland. Now exotic plants dominate in urban forestry and design, where about 86% of city trees are of exotic origin (Hernández and Villaseñor, 2018). The city is in the global biodiversity hotspot of Central Chile, where urgent conservation action is needed because of its high endemism and extensive loss of original vegetation (Myers et al., 2000). Although Central Chile is relevant for global biodiversity conservation, less than 2% of the land is officially protected (INE, 2015) and human activities such as agriculture and urban expansion continue clearing native forests and shrublands.
To select sites, we used a digital layer that represents socioeconomic distribution in the Santiago Metropolitan Area, available at https://observatoriodeciudades.com/. By taking 2012 census data (e.g., level of education of the head of household, occupation, income, access to services, selected goods, number of residents, and number of bedrooms), the layer authors assigned a socioeconomic status to a neighborhood block (polygons) by applying an algorithm to generate a socioeconomic score (Rodríguez et al., 2019). According to these scores, socioeconomic levels can be summarized into three main groups (high, medium, and low). The high socioeconomic group is the wealthier and university educated, with an estimated mean household income greater than US$28,800 per year. The medium socioeconomic group is characterized by households with technical or secondary education and an estimated mean household income greater than US$13,200 per year. The low socioeconomic group are low income and have less education, and an estimated mean household income less than US$8,400 per year (GfK Chile, 2019). To generate a grid of potential sample sites, we rasterized this layer according to their socioeconomic level at a 200 m x 200 m cell size using the “raster” package (Hijmans, 2021) in R 4.1.2 (R Core Team, 2021).
We ensured that different areas were sampled across the city by stratifying in two areas: edge (less than 5 km away from the urban boundary) and interior (more than 5 km away from the urban boundary); where the urban boundary refers to the city’s limits and was defined by a vector layer available from INE - MINVU (2018). This is useful as some species in our study area decrease toward the interior of the city (Villaseñor et al., 2021). It also allows us to reduce confounding socioeconomics and distance to the urban boundary. We focused our sampling on residential areas (land predominantly used for housing) using a layer of land uses from 2014, available from ine.cl that helped us to exclude all non-residential land use types. Then, we used a stratified random selection approach to select 20 sites (raster cells) in residential areas for each combination of socioeconomic level (three levels: high, medium and low) and distance to the urban boundary (two levels: edge and interior), leading to 120 sites (Figure 1).
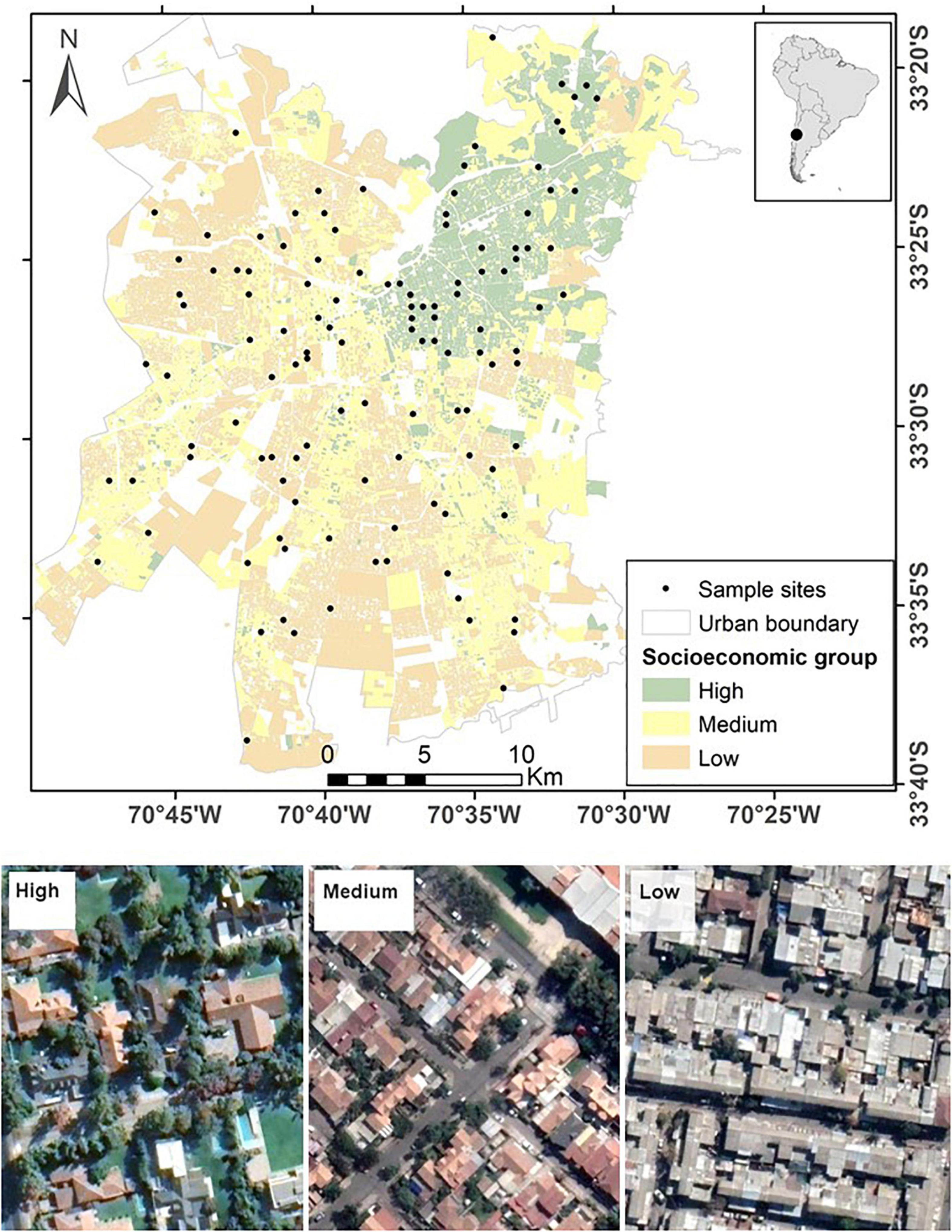
Figure 1. Sample sites and socioeconomic groups in Santiago de Chile. An example of a sample site for each socioeconomic group is provided using satellite imagery from Google Earth. Source of socioeconomic groups https://observatoriodeciudades.com.
At each of the 120 sites, we surveyed birds twice during austral winter season (from 19th July to 6th August 2021). Using the point count method, all birds seen or heard during 5 minutes within a 50-m radius were recorded (Bibby et al., 2000). Birds flying over the site were recorded if using the site (e.g., swallows hunting insects). All sites were surveyed by two different observers, who sampled the same site on different days (nine days average difference between surveys). All counts were performed during mornings (from 7:00 am to 11:57) and on days with favorable weather conditions (no fog, rain, or strong wind; Villaseñor et al., 2021). All points were at distances greater than 300 m from each other.
At each sampling site we measured habitat variables using two sizes of plots: a 50-m radius plot and an 11-m radius plot. These two plot sizes were used because they have proven to be effective in characterizing habitat conditions and their relationship with birds in our study area (Benito et al., 2019; Villaseñor et al., 2021). All plots had their center in the same location as the bird point count. The percent cover of woody vegetation in 50-m radius plots was calculated by interpreting high-resolution satellite imagery (WorldView, DigitalGlobe), digitalizing woody cover within each plot, and calculating the percent cover in ArcMap. Micro-habitat variables were estimated in the field immediately after the bird count using the 11-m radius plot, where the percent cover of shrubs, trees, herbaceous plants, impervious surfaces, and buildings were estimated visually (Benito et al., 2019).
For each site, we pulled together data from the two counts to determine whether the Green-backed Firecrown was present or absent. To investigate whether the occurrence of the species varied with socioeconomic level we fitted a Generalized Linear Model (GLM) with a binomial distribution (link = logit) in R. Then, to investigate the effect of habitat variables on the occurrence of the species we fitted the additive effect of habitat variables in a GLM with a binomial distribution (link = logit) in R. To avoid over parameterizing models, we only fitted habitat variables that exhibited a low correlation with other habitat variables (| r| < 0.4). We interpreted variable estimates and calculated and plotted confidence intervals at 95%.
Due to previous evidence on socioeconomics influencing habitat attributes, and these attributes influencing the occurrence of birds, we investigated whether habitat variables mediated the relationship between socioeconomic level and the occurrence of Green-backed Firecrown. For this, we used a mediation path analysis by fitting general structural equation models with sem function from the lavaan package in R (Rosseel, 2012). We constructed the path model for estimation including as mediators the habitat variables that significantly (P < 0.05) predicted the Green-backed Firecrown occurrence in the GLM. Prior to performing path analysis, we transformed the socioeconomic level (categorical variable with three levels: High, Medium, Low) to numerical (3, 2, 1), and then standardized the new socioeconomic variable and habitat variables (mean = 0, standard deviation = 1). As the response variable was binary (presence, absence) the estimator uses diagonally weighted least squares (DWLS) to estimate model parameters (Rosseel, 2021). We calculated direct and indirect effects (through mediation of habitat variables) of socioeconomic level on the species occurrence. We assessed model fit using Chi-squared (X2) and Standardized root mean square residual (SRMR); and displayed the results of the path analyses if they were satisfactory (Fan et al., 2016; Rosseel, 2021).
We recorded 66 Green-backed Firecrown hummingbirds in 44 sites of the total 120 sites that were surveyed twice during winter. The greatest number of records were in neighborhoods of the high socioeconomic group (25 sites), followed by medium socioeconomic neighborhoods (13 sites) and least in the low socioeconomic neighborhoods (6 sites).
Sites varied in habitat composition. Woody cover at 50-m radius plots ranged from 0 to 38% cover (mean = 9.7%; Supplementary Table 1). Microhabitat, described by 11-m radius plots, was dominated by impervious surfaces (mean = 50.3%), followed by tree cover (mean = 18.8%), built cover (mean = 17.6%), herbaceous cover (mean = 11.3%), and the least abundant vegetation type was shrub cover (mean = 4.8%; Supplementary Table 1). Woody cover exhibited a strong positive correlation with tree cover (r = 0.6). We fitted woody cover in our Generalized Linear Model as it best represents the surveyed area (50-m radius plot) and data exploration showed these habitat variables exhibited similar association with hummingbird occurrence. Built cover exhibited a moderate positive correlation with impervious surfaces (r = 0.4). Thus, we fit built cover in our Generalized Linear Model as it might represent a barrier for hummingbirds and data exploration showed a greater association between built cover and hummingbird occurrence than impervious surfaces.
The Generalized Linear Model revealed that the occurrence of Green-backed Firecrown significantly varied with neighborhood socioeconomic level. It was highest in sites with high socioeconomic level and lowest in sites with low socioeconomic level (P < 0.001; Supplementary Table 2 and Figure 2). We also found that habitat variables had a significant effect on the occurrence of Green-backed Firecrown at sampling sites. The occurrence of Green-backed Firecrown increased with woody cover (P = 0.002) and shrub cover (P = 0.01). Built cover exhibited a negative effect on Green-Backed Firecrown, although its effect was not statistically significant (P = 0.07; Supplementary Table 3 and Figure 3).
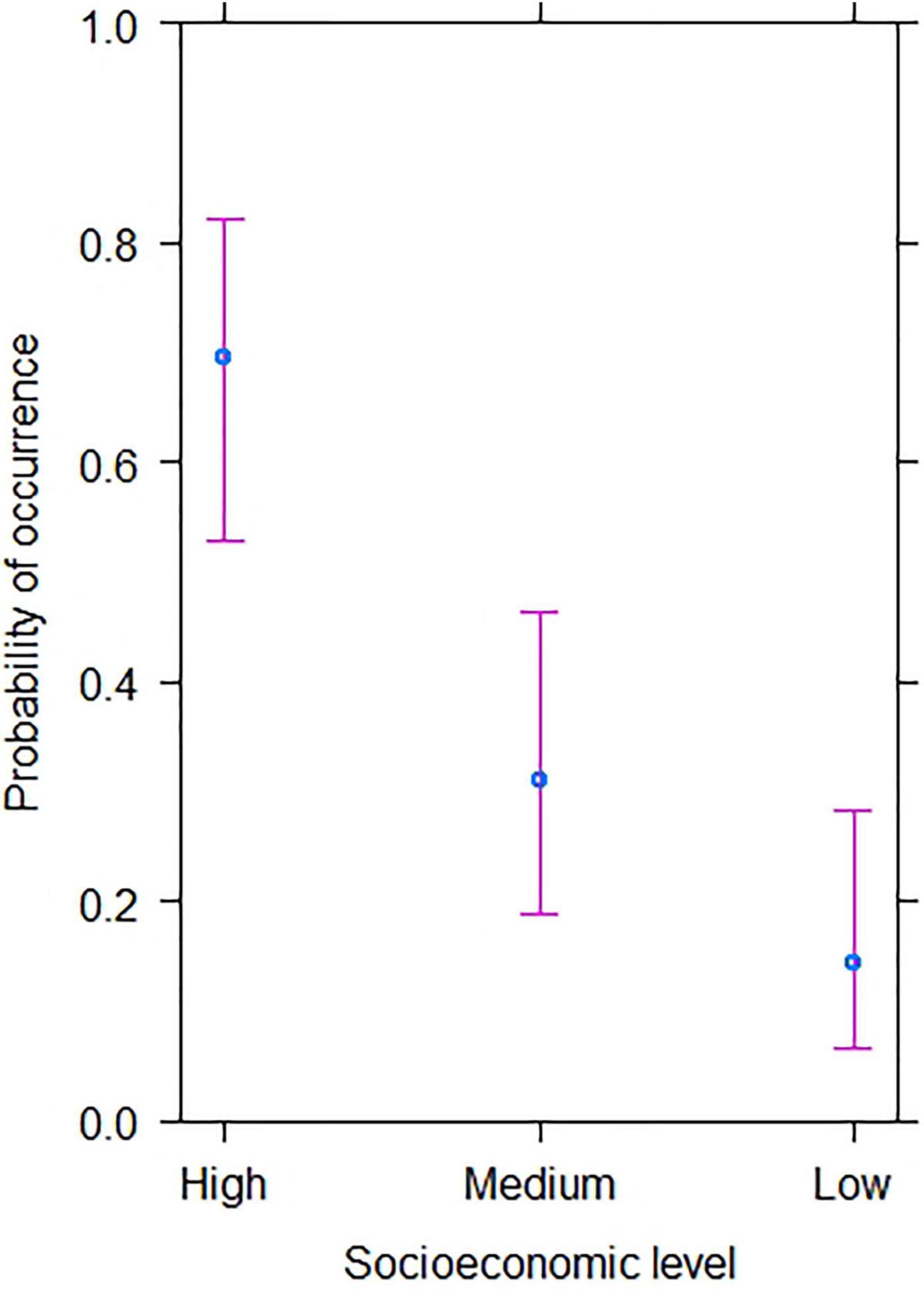
Figure 2. Results from Generalized Linear Model predicting the occurrence of the Green-backed Firecrown hummingbird according to neighborhood socioeconomic level.
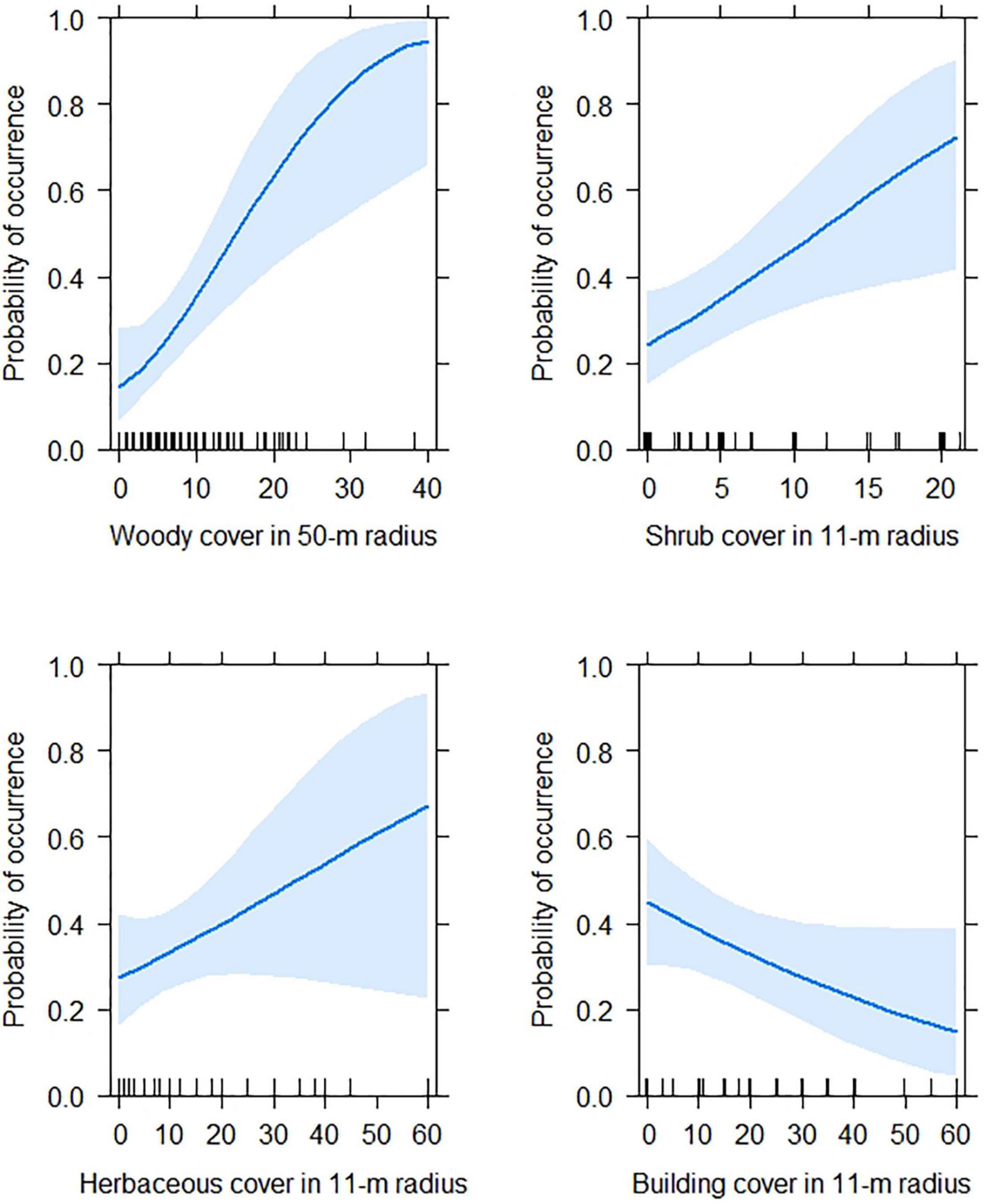
Figure 3. Results from Generalized Linear Models predicting the occurrence the Green-backed Firecrown hummingbird according to habitat variables.
The path analysis built with two habitat variables (woody and shrub cover) as mediators evidenced partial mediation (Figure 4 and Supplementary Table 4). Socioeconomic level positively influenced hummingbird occurrence (total effect = 0.64 ± 0.13, P < 0.001). Indirect effects evidence that woody cover (0.15 ± 0.06, P = 0.01) and shrub cover (0.09 ± 0.04, P = 0.05) mediate the relationship between socioeconomic level and hummingbird occurrence. Regression estimates show that socioeconomic level positively influences woody cover (β = 0.52 ± 0.1, P < 0.001) and shrub cover (β = 0.26 ± 0.09, P = 0.005), and they both positively influence hummingbird occurrence (β = 0.29 ± 0.1, P = 0.004; β = 0.33 ± 0.11, P = 0.004, respectively). Nevertheless, the results also evidence that even after accounting for mediating effects of these habitat variables, socioeconomic level still has a direct positive effect on hummingbird occurrence (β = 0.41 ± 0.14, P < 0.004; Figure 4). Goodness of fit indices mostly indicated a satisfactory fit: X2 = 2.35, P = 0.125, suggesting optimal fitting; SRMR = 0.049 suggesting a good model fit (Fan et al., 2016).
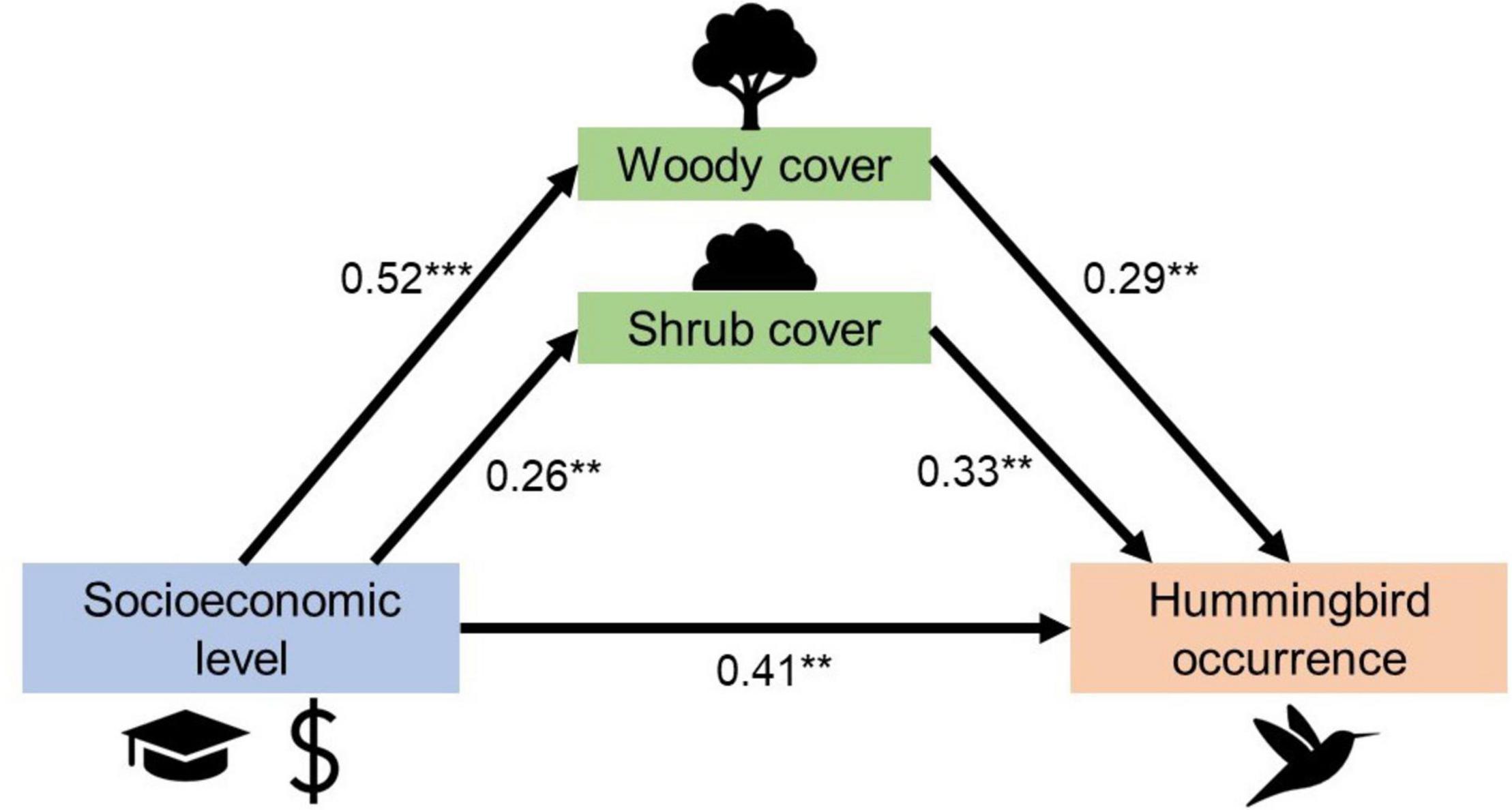
Figure 4. Results from path analysis evidences a partial mediation of woody and shrub cover on the observed relationship between neighborhood socioeconomic level and the Green-backed Firecrown hummingbird occurrence. Arrows represent a regression (causal) relationship between independent and dependent variables. The numbers on the arrows are standardized regression coefficients that indicate the strength and direction of the effects between variables. Statistical significance: **P < 0.01, ***P < 0.001. The proportion of variance of the Green-backed Firecrown hummingbird occurrence explained by the predictors: R2 = 0.41.
To better direct conservation efforts it is essential to understand species responses to landscape change. This knowledge is particularly relevant for migratory species that are important pollinators in their habitats and can be affected by environmental modification, such as the most austral Neotropical migratory hummingbird. In its urban overwintering habitat, Green-backed Firecrown occurrence increased with socioeconomic level and woody cover. Specifically, we found that socioeconomic level influenced the Green-backed Firecrown occurrence indirectly (mediated by woody and shrub cover), as well as through direct effects. Wealthier neighborhoods have greater vegetation cover and higher hummingbird occurrence than poorer neighborhoods, providing dissimilar opportunities for urban residents to experience nature close to home.
Socioeconomics is related to the distribution of vegetation and hummingbirds in the city. As revealed in our path analyses, woody cover, that comprises trees and shrubs, increases with neighborhood socioeconomic level and positively influence hummingbird occurrence. Such uneven distribution of urban vegetation has been documented in different countries. For instance, vegetation cover relates to wealth in Colombia (Escobedo et al., 2015), education level in Australia (Luck et al., 2009), and socioeconomics in the United States (Harlan et al., 2006). In the city of Santiago de Chile, areas of higher socioeconomic level present greater abundance and diversity of trees and larger green spaces than areas of a lower socioeconomic level (de la Maza et al., 2002; Reyes and Figueroa Aldunce, 2010; Hernández and Villaseñor, 2018). Our study uncovered that these differences in vegetation cover also led to differences in hummingbird occurrence, implying that people of higher socioeconomic level have greater opportunities to experience nature close to home than people of a lower socioeconomic level. Management actions aiming to increase tree and shrub cover in neighborhoods where people of lower socioeconomic status live will decrease differences in woody cover and hummingbird occurrence, contributing to a more environmentally just city. In this socio-ecological context, woody cover in underprivileged neighborhoods will provide a variety of ecosystem services, such as habitat for wildlife, climate regulation, storm water attenuation, air quality improvement, better human health and wellbeing, in addition to aid achieving sustainable development goals (Roy et al., 2012; Ulmer et al., 2016; Dobbs et al., 2018).
Among all habitat variables explored, those related with woody plants were important for hummingbird occurrence. Several tree species found in urban settlements present specialized adaptations to bird pollination, thus, trees have been found to be more relevant for hummingbirds than herbs (Maruyama et al., 2019). Trees and shrubs provide foraging resources for the Green-backed Firecrown hummingbird that mainly feeds on flowers of woody plants and native mistletoes that parasite trees or shrubs (Aizen et al., 2002; Cuadra-Valdés et al., 2021). In urban land, scattered trees with large floral displays promote territorial behavior, where male hummingbirds actively defend territories from other males (González-Gómez and Vásquez, 2006; Maruyama et al., 2019). Trees and shrubs not only offer foraging resources, but also might improve winter habitat conditions in urban ecosystems by providing refuges to rest, hide from urban predators and protect from rainfall and cold temperatures.
High socioeconomic levels not only promoted a greater occurrence of the Green-backed Firecrown hummingbird in the city through effects mediated by woody and shrub cover, but also through additional unexplored effects. Previous research has found that affluent areas present greater woody plant diversity, greater green space size, greater backyard size and are commonly closer to remnants of native vegetation than poorer areas in the city (Reyes and Figueroa Aldunce, 2010; Hernández and Villaseñor, 2018; Silva, 2021). These variables might promote hummingbird occurrence, as well as the presence of other native bird species in the city (Pauw and Louw, 2012; Villaseñor et al., 2021). Spatial arrangement of vegetation might also be relevant. Hummingbirds might be sensitive to habitat fragmentation and isolation (Gannon et al., 2021), decrease with distance to the natural habitat (e.g., nectar-feeding birds, Pauw and Louw, 2012) or respond to the clumping of feeding resources (Tiebout, 1992). Neighborhood socioeconomics could also relate to the abundance of domestic predators. Preliminary data collected in our study sites shows that free-ranging cats and dogs decrease with neighborhood wealth: on average, 13-fold more cats and 11-fold more dogs were recorded in lower socioeconomic neighborhoods compared with higher socioeconomic neighborhoods (authors, unpublished data). In addition, hummingbird feeders, baths and fountains are more likely to be provided by people of higher socioeconomic level to attract birds, as they have greater purchasing power, their gardens are larger and managed with a greater proportion of woody vegetation cover. Supplementary feeding is common in more developed countries, where up to half of households in the United States, United Kingdom, and Australia offer supplementary food to wild birds (Orros and Fellowes, 2015; and references therein), however, we are unaware of such a widespread practice in our study area. Because some attributes of affluent neighborhoods that are relevant for hummingbird occurrence remain unknown, a more detailed assessment is needed. In addition to the variables previously described, future assessments could include flower availability and abundance (Fontúrbel et al., 2017; Cuadra-Valdés et al., 2021), plant traits (e.g., origin, syndromes, phenology, nectar secretion rates; Maruyama et al., 2019), and spatial variables (Pauw and Louw, 2012).
Both exotic and native species provide relevant foraging resources for hummingbirds, however, alien plants can interfere in ecological networks. For instance, native and exotic plants can compete for pollinators (Feinsinger, 1987), where alien plants can attract more native pollinators (Morales and Aizen, 2002) and present more hummingbird interactions than native plants (Maruyama et al., 2019). This phenomenon will likely affect plant recruitment and long-term viability of native vegetation that is already deteriorated and under high threat. In Chile, even though highly attractive native resources exist, such as Tristerix spp., exotic species such as Eucalyptus spp. act as magnet species, changing hummingbird abundance, visitation rates and activity patterns, altering pollination interactions not only in the site but also in the surrounding landscape (Cuadra-Valdés et al., 2021). Given that vegetation in Santiago is strongly dominated by exotic species (86% of trees are exotic; Hernández and Villaseñor, 2018), their ecological impacts need to be better understood.
With climate change and growing urbanization, monitoring programs are key to determine the status, detect changes and, thus, implement early conservation action. The mega drought affecting Central Chile since 2010 (Garreaud et al., 2020) has created drier conditions in natural areas, thus greener plants in the city might be attracting more birds. However, due to the lack of long-term monitoring programs for terrestrial birds in our study area, we only can hypothesize on the effects of this mega drought on birds. In addition, several changes could be occurring in bird species in our study area because urbanization alters environmental conditions that might lead to changes in phenology, behavior and/or distribution patterns. For instance, reduced migration and range expansion have been reported for birds in urban areas offering milder conditions than surrounding environments and providing abundant supplementary food through the year - by the widespread use of feeding stations, food scraps, or non-native plants (Greig et al., 2017). Urban areas can alter the phenology of bird species to different degrees, from earlier arrival times to some migratory species becoming sedentary in cities (Cubie, 2014; Greig et al., 2017). These changes in bird ecology influence ecological networks, where changes in a keystone species may threaten the viability of ecosystems.
The Green-backed Firecrown hummingbird is the most abundant and most important vertebrate pollinator in south temperate forests of South America (Aizen et al., 2002; Vergara et al., 2014). It is a dominant species in natural areas such as old-growth forests (Díaz et al., 2005), but can be abundant in modified environments, such as eucalypt plantations (Fontúrbel et al., 2017) and green spaces within cities (Villaseñor and Escobar, 2019). Despite the relevance of the Green-backed Firecrown hummingbird for ecosystem functions and its susceptibility to environmental modification, investigations focused on the ecology of the species in modified environments are scarce [but see growing work by Fontúrbel et al. (2017), Cuadra-Valdés et al. (2021)]. Implementing monitoring programs along its range will contribute to understand the effect of anthropogenic change on the species and its ecosystem network.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethical review and approval was not required for the animal study because this is an observational study (bird counts). No animals were captured or harmed to perform the research.
NV conceived the idea, obtained funding, designed the study, performed statistical analyses, and wrote the manuscript. ME coordinated fieldwork, collected data, and contributed with comments and editions on the manuscript. Both authors contributed to the article and approved the submitted version.
Government of Chile, ANID-FONDECYT 11201045 granted to NV.
ME was employed by Manque Bioexploraciones SpA.
The remaining author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank funding from ANID-FONDECYT 11201045 (granted to NV). We also thank Macarena Silva and Catalina Muñoz-Pacheco who contributed to collect data, Trevor Walter for proofreading and providing suggestions, and two reviewers for their comments on a previous version of the manuscript.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.850065/full#supplementary-material
Aizen, M. A., Vázquez, D. P., and Smith-Ramírez, C. (2002). Historia natural y conservación de los mutualismos planta-animal del bosque templado de Sudamérica austral. Rev. Chil. Hist. Nat. 75, 79–97.
Amaya-Espinel, J. D., and Hostetler, M. E. (2019). The value of small forest fragments and urban tree canopy for Neotropical migrant birds during winter and migration seasons in Latin American countries: a systematic review. Landsc. Urban Plan. 190:103592. doi: 10.1016/j.landurbplan.2019.103592
Aronson, M. F. J., La Sorte, F. A., Nilon, C. H., Katti, M., Goddard, M. A., Lepczyk, C. A., et al. (2014). A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. B Biol. Sci. 281:20133330. doi: 10.1098/rspb.2013.3330
Benito, J. F., Escobar, M. A. H., and Villaseñor, N. R. (2019). Conservation in the city: how does habitat structure influence the abundance of individual bird species in a Latin American metropolis? Gayana 83, 114–125. doi: 10.4067/S0717-65382019000200114
Bibby, C. J., Burgess, N. D., Hill, D. A., and Mustoe, S. (2000). Bird Census Techniques, Second ed. Great Britain: Academic Press.
Chamberlain, D. E., Henry, D. A. W., Reynolds, C., Caprio, E., and Amar, A. (2019). The relationship between wealth and biodiversity: a test of the Luxury Effect on bird species richness in the developing world. Glob. Chang. Biol. 25, 3045–3055. doi: 10.1111/gcb.14682
Cuadra-Valdés, J., Vizentin-Bugoni, J., and Fontúrbel, F. E. (2021). An exotic magnet plant alters pollinator abundance and behavior: a field test with a native mistletoe. Biol. Invasions 23, 2515–2525. doi: 10.1007/s10530-021-02519-2
Cubie, D. (2014). Site fidelity, residency, and sex ratios of wintering ruby-throated hummingbirds (Archilochus colubris) on the southeastern U.S. Atlantic Coast. Wilson J. Ornithol. 126, 775–778. doi: 10.1676/14-005.1
de la Maza, C. L., Rodríguez, R., Bown, H., Hernández, J., and Escobedo, F. (2002). Vegetation diversity in the Santiago de Chile urban ecosystem. Arboric. J. 26, 347–357.
de Zoeten, T., and Pulido, F. (2020). How migratory populations become resident. Proc. R. Soc. B Biol. Sci. 287:20193011. doi: 10.1098/rspb.2019.3011
Díaz, I. A., Armesto, J. J., Reid, S., Sieving, K. E., and Willson, M. F. (2005). Linking forest structure and composition: avian diversity in successional forests of Chiloé Island, Chile. Biol. Conserv. 123, 91–101. doi: 10.1016/j.biocon.2004.10.011
Dingle, H., and Drake, V. A. (2007). What is migration? Bioscience 57, 113–121. doi: 10.1641/B570206
Dobbs, C., Escobedo, F. J., Clerici, N., de la Barrera, F., Eleuterio, A. A., MacGregor-Fors, I., et al. (2018). Urban ecosystem Services in Latin America: mismatch between global concepts and regional realities? Urban Ecosyst. 22, 173–187. doi: 10.1007/S11252-018-0805-3
Dokter, A. M., Farnsworth, A., Fink, D., Ruiz-Gutierrez, V., Hochachka, W. M., La Sorte, F. A., et al. (2018). Seasonal abundance and survival of North America’s migratory avifauna determined by weather radar. Nat. Ecol. Evol. 2, 1603–1609. doi: 10.1038/s41559-018-0666-4
Escobedo, F. J., Clerici, N., Staudhammer, C. L., and Corzo, G. T. (2015). Socio-ecological dynamics and inequality in Bogotá, Colombia’s public urban forests and their ecosystem services. Urban For. Urban Green. 14, 1040–1053.
Escobedo, F. J., Palmas-Perez, S., Dobbs, C., Gezan, S., and Hernandez, J. (2016). Spatio-temporal changes in structure for a Mediterranean urban forest: Santiago, Chile 2002 to 2014. Forests 7:121.
Estades, C. F., and Temple, S. A. (1999). Deciduous-forest bird communities in a fragmented landscape dominated by exotic pine plantations. Ecol. Appl. 9, 573–585.
Fan, Y., Chen, J., Shirkey, G., John, R., Wu, S. R., Park, H., et al. (2016). Applications of structural equation modeling (SEM) in ecological studies: an updated review. Ecol. Process. 5:19. doi: 10.1186/s13717-016-0063-3
Feinsinger, P. (1987). Effects of plant species on each other’s pollination: is community structure influenced? Trends Ecol. Evol. 2, 123–126. doi: 10.1016/0169-5347(87)90052-8
Fontúrbel, F. E., Salazar, D. A., and Medel, R. (2017). Increased resource availability prevents the disruption of key ecological interactions in disturbed habitats. Ecosphere 8:e01768. doi: 10.1002/ecs2.1768
Gannon, D. G., Hadley, A. S., and Frey, S. J. K. (2021). Pairing automated mark–recapture and social network models to explore the effects of landscape configuration on hummingbird foraging patterns. Biol. Lett. 17:20210188. doi: 10.1098/RSBL.2021.0188
Garreaud, R. D., Boisier, J. P., Rondanelli, R., Montecinos, A., Sepúlveda, H. H., and Veloso-Aguila, D. (2020). The Central Chile Mega Drought (2010–2018): a climate dynamics perspective. Int. J. Climatol. 40, 421–439. doi: 10.1002/joc.6219
GfK Chile (2019). Estilo de Vida de los Nuevos Grupos Socioeconómicos de Chile. Available online at: https://www.gfk.com/fileadmin/user_upload/country_one_pager/CL/GfK_GSE_190502_FINAL.pdf [(accessed February 3, 2022]).
González-Gómez, P. L., and Vásquez, R. A. (2006). A field study of spatial memory in Green-backed Firecrown hummingbirds (Sephanoides sephaniodes). Ethology 112, 790–795. doi: 10.1111/j.1439-0310.2006.01223.x
Greig, E. I., Wood, E. M., and Bonter, D. N. (2017). Winter range expansion of a hummingbird is associated with urbanization and supplementary feeding. Proc. R. Soc. B Biol. Sci. 284:20170256. doi: 10.1098/rspb.2017.0256
Harlan, S. L., Brazel, A. J., Prashad, L., Stefanov, W. L., and Larsen, L. (2006). Neighborhood microclimates and vulnerability to heat stress. Soc. Sci. Med. 63, 2847–2863. doi: 10.1016/j.socscimed.2006.07.030
Hernández, H. J., and Villaseñor, N. R. (2018). Twelve-year change in tree diversity and spatial segregation in the Mediterranean city of Santiago, Chile. Urban For. Urban Green. 29, 10–18. doi: 10.1016/j.ufug.2017.10.017
INE - MINVU (2018). Metodología para medir el crecimiento urbano de las ciudades de Chile. IX Ciudad y Territorio. Santiago: Ministerio de Vivienda y Urbanismo.
INE (2015). Medio Ambiente, Informe Anual. Período de información: 2009 – 2013. Santiago: Instituto Nacional de Estadísticas.
INE (2019). Estimaciones y proyecciones de la población de Chile 2002-2035, totales regionales, población urbana y rural. Santiago: Instituto Nacional de Estadísticas.
IPBES (2019). Global Assessment Report on Biodiversity and Ecosystem Services. Bonn: IPBES Secretariat.
Kamp, J., Oppel, S., Ananin, A. A., Durnev, Y. A., Gashev, S. N., Hölzel, N., et al. (2015). Global population collapse in a superabundant migratory bird and illegal trapping in China. Conserv. Biol. 29, 1684–1694. doi: 10.1111/cobi.12537
Kauffman, M. J., Cagnacci, F., Chamaillé-Jammes, S., Hebblewhite, M., Hopcraft, J. G., Merkle, J. A., et al. (2021). Mapping out a future for ungulate migrations. Science 372, 566–569. doi: 10.1126/science.abf0998
Kinzig, A. P., Warren, P., Martin, C., Hope, D., and Katti, M. (2005). The effects of human socioeconomic status and cultural characteristics on urban patterns of biodiversity. Ecol. Soc. 10:23.
Koleček, J., Reif, J., Šálek, M., Hanzelka, J., Sottas, C., and Kubelka, V. (2021). Global population trends in shorebirds: migratory behaviour makes species at risk. Sci. Nat. 108:9. doi: 10.1007/s00114-021-01717-1
Leong, M., Dunn, R. R., and Trautwein, M. D. (2018). Biodiversity and socioeconomics in the city: a review of the luxury effect. Biol. Lett. 14:20180082. doi: 10.1098/rsbl.2018.0082
Leveau, L. M. (2021). Big cities with small green areas hold a lower proportion of migrant birds: a global analysis. Urban For. Urban Green. 57:126953. doi: 10.1016/j.ufug.2020.126953
Loss, S. R., Ruiz, M. O., and Brawn, J. D. (2009). Relationships between avian diversity, neighborhood age, income, and environmental characteristics of an urban landscape. Biol. Conserv. 142, 2578–2585. doi: 10.1016/j.biocon.2009.06.004
Luck, G. W., Smallbone, L. T., O’Brien, R., and O’Brien, R. (2009). Socio-economics and vegetation change in urban ecosystems: patterns in space and time. Ecosystems 12:604. doi: 10.1007/s10021-009-9244-6
MacGregor-Fors, I., and Schondube, J. E. (2011). Gray vs. green urbanization: relative importance of urban features for urban bird communities. Basic Appl. Ecol. 12, 372–381. doi: 10.1016/j.baae.2011.04.003
Martínez-Piña, D., and González-Cifuentes, G. (2017). Aves de Chile: guía de campo y Breve Historia Natural. Santiago: Ediciones del Naturalista.
Maruyama, P. K., Bonizário, C., Marcon, A. P., D’Angelo, G., da Silva, M. M., da Silva Neto, E. N., et al. (2019). Plant-hummingbird interaction networks in urban areas: generalization and the importance of trees with specialized flowers as a nectar resource for pollinator conservation. Biol. Conserv. 230, 187–194. doi: 10.1016/j.biocon.2018.12.012
McKinney, M. L. (2002). Urbanization, biodiversity, and conservation. Bioscience 52, 883–890. doi: 10.1641/0006-3568(2002)052[0883:ubac]2.0.co;2
Morales, C. L., and Aizen, M. A. (2002). Does invasion of exotic plants promote invasion of exotic flower visitors? A case study from the temperate forests of the Southern Andes. Biol. Invasions 4, 87–100. doi: 10.1023/A:1020513012689
Muñoz-Pacheco, C. B., and Villaseñor, N. R. (2021). Avian species richness in cities: a review of the Spanish-language literature from the Southern Cone of South America. Urban Ecosyst. doi: 10.1007/s11252-021-01180-w
Myers, N., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A. B., and Kent, J. (2000). Biodiversity hotspots for conservation priorities. Nature 403, 853–858. doi: 10.1038/35002501
Orros, M. E., and Fellowes, M. D. E. (2015). Wild bird feeding in an urban area: intensity, economics and numbers of individuals supported. Acta Ornithol. 50, 43–58. doi: 10.3161/00016454AO2015.50.1.006
Pauw, A., and Louw, K. (2012). Urbanization drives a reduction in functional diversity in a guild of nectar-feeding birds. Ecol. Soc. 17:27. doi: 10.5751/ES-04758-170227
R Core Team (2021). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Reyes, S., and Figueroa Aldunce, I. (2010). Distribución, superficie y accesibilidad de las áreas verdes en Santiago de Chile. Rev. Estud. Urbano Reg. 36, 89–110.
Robinson, R. A., Crick, H. Q. P., Learmonth, J. A., Maclean, I. M. D., Thomas, C. D., Bairlein, F., et al. (2009). Travelling through a warming world: climate change and migratory species. Endanger. Species Res. 7, 87–99. doi: 10.3354/esr00095
Rodríguez, P., Valenzuela, J., Truffello, R., Ulloa, J., Matas, M., Quintana, D., et al. (2019). Un modelo de identificación de requerimientos de nueva infraestructura pública en educación básica. Santiago: Ministerio de Educación.
Rosseel, Y. (2012). lavaan: an R package for structural equation modeling. J. Stat. Softw. 2, 1–36. doi: 10.3389/fpsyg.2014.01521
Roy, S., Byrne, J., and Pickering, C. (2012). A systematic quantitative review of urban tree benefits, costs, and assessment methods across cities in different climatic zones. Urban For. Urban Green. 11, 351–363. doi: 10.1016/J.UFUG.2012.06.006
Sanderson, E. W., and Huron, A. (2011). Conservation in the city. Conserv. Biol. 25, 421–423. doi: 10.1111/j.1523-1739.2011.01683.x
Silva, C. P., García, C. E., Estay, S. A., Barbosa, O., and Chapman, M. G. (2015). Bird richness and abundance in response to urban form in a Latin American City: Valdivia, Chile as a Case Study. PLoS One 10:e0138120. doi: 10.1371/journal.pone.0138120
Silva, M. (2021). Segregación socioambiental y riqueza de aves en la ciudad de Santiago, Chile. Ph.D.thesis. Santiago: Universidad de Chile.
Silva-Rodríguez, E. A., Ortega-Solís, G. R., and Jiménez, J. E. (2008). Descripción del ensamble de aves en un agroecosistema del sur de Chile. Boletín Chil. Ornitol. 14, 81–91.
Sol, D., Trisos, C., Múrria, C., Jeliazkov, A., González-Lagos, C., Pigot, A. L., et al. (2020). The worldwide impact of urbanisation on avian functional diversity. Ecol. Lett. 23, 962–972. doi: 10.1111/ele.13495
Stratford, J. A., and Robinson, W. D. (2005). Distribution of neotropical migratory bird species across an urbanizing landscape. Urban Ecosyst. 81, 59–77. doi: 10.1007/S11252-005-1419-0
Strohbach, M. W., Haase, D., and Kabisch, N. (2009). Birds and the city: urban biodiversity, land use, and socioeconomics. Ecol. Soc. 14:31. doi: 10.5751/ES-03141-140231
Tiebout, H. M. (1992). Comparative energetics of divergent foraging modes: a doubly labelled water experiment on hummingbird competition. Anim. Behav. 44, 895–906. doi: 10.1016/S0003-3472(05)80585-8
Tryjanowski, P., Sparks, T. H., Kuźniak, S., Czechowski, P., and Jerzak, L. (2013). Bird migration advances more strongly in urban environments. PLoS One 8:e63482. doi: 10.1371/journal.pone.0063482
Ulmer, J. M., Wolf, K. L., Backman, D. R., Tretheway, R. L., Blain, C. J., O’Neil-Dunne, J. P., et al. (2016). Multiple health benefits of urban tree canopy: the mounting evidence for a green prescription. Health Place 42, 54–62. doi: 10.1016/J.HEALTHPLACE.2016.08.011
Vergara, P. M., Rivera-Hutinel, A., Farías, A. A., Cofré, H., Samaniego, H., and Ingo, J. H. (2014). “Aves y mamíferos del bosque,” in Ecología Forestal: Bases para el Manejo Sustentable y Conservación de los Bosques Nativos de Chile, eds C. Donoso, M. E. González, and A. Lara (Valdivia: Universidad Austral de Chile).
Villaseñor, N. R., and Escobar, M. A. H. (2019). Cemeteries and biodiversity conservation in cities: how do landscape and patch-level attributes influence bird diversity in urban park cemeteries? Urban Ecosyst. 22, 1037–1046. doi: 10.1007/s11252-019-00877-3
Villaseñor, N. R., Truffello, R., and Reyes-Paecke, S. (2021). Greening at multiple scales promote biodiverse cities: a multi-scale assessment of drivers of Neotropical birds. Urban For. Urban Green. 66:127394. doi: 10.1016/j.ufug.2021.127394
Villegas, M., Garitano-Zavala, Á, Villegas, M., and Garitano-Zavala, Á (2010). Bird community responses to different urban conditions in La Paz, Bolivia. Urban Ecosyst. 13, 375–391. doi: 10.1007/s11252-010-0126-7
Visser, M. E., Perdeck, A. C., Van Balen, J. H., and Both, C. (2009). Climate change leads to decreasing bird migration distances. Glob. Chang. Biol. 15, 1859–1865. doi: 10.1111/j.1365-2486.2009.01865.x
Keywords: environmental justice, hummingbird, luxury effect, neotropical austral migrants, Sephanoides sephaniodes, urbanization, wildlife-friendly cities, nectar-feeding bird
Citation: Villaseñor NR and Escobar MAH (2022) Linking Socioeconomics to Biodiversity in the City: The Case of a Migrant Keystone Bird Species. Front. Ecol. Evol. 10:850065. doi: 10.3389/fevo.2022.850065
Received: 07 January 2022; Accepted: 22 February 2022;
Published: 15 March 2022.
Edited by:
Álvaro Garitano-Zavala, Universidad Mayor de San Andrés, BoliviaReviewed by:
Alex Jahn, Indiana University, United StatesCopyright © 2022 Villaseñor and Escobar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nélida R. Villaseñor, dmlsbGFzZW5vckB1Y2hpbGUuY2w=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.