
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 06 May 2022
Sec. Behavioral and Evolutionary Ecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.841744
 Mohammed Athamnia1,2†Bourhane-Eddine Belabed3†
Mohammed Athamnia1,2†Bourhane-Eddine Belabed3† Kenz Raouf Samraoui4,5†Abdennour Boucheker1,6†Laïd Touati1,7†Farrah Samraoui1,2†Hamed A. El-Serehy8†
Kenz Raouf Samraoui4,5†Abdennour Boucheker1,6†Laïd Touati1,7†Farrah Samraoui1,2†Hamed A. El-Serehy8† Boudjéma Samraoui1,5*†
Boudjéma Samraoui1,5*†
Global changes are inducing migratory birds to advance their annual cycle. However, changes in the time of arrival at their breeding grounds have significant fitness implications. This study aims to identify factors affecting the variability in arrival time of migratory white storks (Ciconia ciconia L.) and to determine if their arrival at North African breeding grounds is occurring earlier. We monitored the arrival of ringed white storks at a breeding colony in Algeria between 2017 and 2021. The birds arrived at this breeding colony over an extended period spanning mid-December to mid-June each year. We found that stork arrival was negatively correlated with age and year of arrival, with older birds arriving first and stragglers consisting of first- and second-year birds arriving later. Notably, arrivals have been shifting toward earlier dates at this breeding ground. Furthermore, cluster analysis of arrival dates for each age-class revealed two distinct groups comprising early and late arrivals. Advancement of the annual cycle of the North African white stork population is consistent with phenological shifts induced by global changes and that have been recorded globally in a wide range of living organisms.
Arrival time is a key life history trait for long-distance migrants endeavoring to reach their breeding grounds when environmental conditions are optimal for migration and breeding (Tomotani et al., 2021). It is also central to our understanding of how evolution has shaped bird migration (Gienapp et al., 2008), particularly in light of the trade-off between costs and benefits inherent to migratory behavior (Møller, 1994).
Several studies have explored factors influencing arrival time of migratory birds (reviewed by Newton, 2007a). Among these, bird age is known to affect arrival time (Møller, 1994; Lozano et al., 1996; Moore et al., 2003). However, interindividual variation in arrival time has often been viewed simply as differing between juveniles and adults (Rotics et al., 2018), with research consistently showing that first-year juveniles fare far worse than adults (Dittmann and Becker, 2003; Sergio et al., 2014) during their grueling first migratory flight because they expend considerably more energy in costly flapping flight (Rotics et al., 2016). Given the causal link between phenology and individual fitness (Drent et al., 2003), this early handicap for young birds may have long-lasting effects, as arrival time is negatively correlated with age (Belabed et al., 2019).
Other intrinsic factors that mediate variation in arrival time include sex (Ouwehand and Both, 2017), physical condition (Matyjasiak et al., 2013; Cooper et al., 2015), and overwintering location (Rotics et al., 2018). However, additional factors are difficult to disentangle from age. For instance, older birds may have better navigation skills (Perdeck, 1967; Mueller et al., 2013), perform better or rely more on soaring flight (Harel et al., 2016; Rotics et al., 2016), overwinter closer to breeding grounds (Teitelbaum et al., 2016), exhibit more experience in finding optimal flight paths (Mueller et al., 2013; Sergio et al., 2014), or exert social dominance on congeners enroute (Moore et al., 2003).
Importantly, interannual variability in arrival time has been shown to co-vary with environmental factors such as temperature at the breeding grounds and along the migration route (Hüppop and Hüppop, 2003; Marra et al., 2005; Gordo, 2007; Gordo et al., 2013; Vaitkuviene et al., 2015). There is now global consensus that human activity has caused the rapid rise in the average temperature of the Earth’s surface (IPCC, 2014). To date, many studies have uncovered congruence between temperature-related shifts and altered species phenology and distribution, with species advancing their annual cycles (Walther et al., 2002; Root et al., 2003; Menzel et al., 2006) and shifting their altitudinal or latitudinal distributions (Chen et al., 2011; Boisvert-Marsh et al., 2014).
One subtle threat has come from insights into migrant birds and changing selection pressures on avian life history traits (Coppack and Both, 2002; Both and Visser, 2005), leading to possible trophic mismatches between the timing of breeding and food supply (Visser et al., 1998, 2004; Menzel et al., 2006). Consequently, species that are unable to shift their breeding period to match advances in seasonal phenology are in decline (Both et al., 2006; Møller et al., 2008; Saino et al., 2011). Thus, given the strong associations between global warming and other anthropogenic activities such as food subsidies provided by landfills (Gilbert et al., 2016), breeding and population dynamics, it is important to monitor changes that may provide important insights into the plasticity displayed by long-distance migratory birds and their possible responses to climate change.
As a long-distance migrant now known to be arriving earlier at its breeding grounds (Ptaszyk et al., 2003), the white stork is an excellent biological model for exploring the evolution of bird migration and the impact of global changes. Timings of both arrival and reproductive performance are closely linked, implying that strong selective pressures act on these two life history traits (Tryjanowski et al., 2004; Belabed et al., 2019). However, attempts to define a relationship between arrival time and reproductive success in birds (e.g., number of chicks raised) have generated apparently contradictory results, with some studies uncovering a significant link (Tryjanowski et al., 2004; Belabed et al., 2019), whereas others did not (Vergara et al., 2007; Fulin et al., 2009). Thus, more research is needed to disentangle all of the factors (age, body condition, climate, etc.) affecting the timing of arrival and their relationships with other life history traits. Considering the inherent plasticity of bird migration (Newton, 2007a) and the intricate link between phenology and fitness, predicting the effects of global warming on bird migration patterns is complex yet essential to inform conservation efforts. The study colony may provide insights on how the breeding phenology of long-lived migrants varies when there is no strong selective pressure for matching with local spring timing as the urban food subsidies may dilute such selective pressure. Thus, modeling the time of arrival of migrant species at their breeding grounds enables key factors influencing bird migration and reproductive success to be identified. Such modeling approaches may also help predict changes in population dynamics resulting from variations in these driving factors. In this study, we aim to identify factors affecting variability in the arrival time of white storks at a North African breeding ground and we particularly wanted to answer the following questions:
1. Is age a driver of arrival time in the white stork?
2. Is the study population advancing its arrival at its breeding ground?
3. Within each age-class, are there individuals that arrive consistently earlier than others?
Algeria supports a large population of white storks located at the southern edge of the species’ breeding range, where they are clustered in loose colonies. One such colony in an olive grove near Dréan, Algeria (36° 41.170′ N, 7° 41.520′ E), is situated 300 m away from the largest landfill site in the region (Bouriach et al., 2015). The breeding season at Dréan extends from the end of February to mid-July (Belabed et al., 2019). From 2011 to the current time, the breeding ecology of the white stork population at this site has been monitored and, every year before fledging, surviving chicks are fitted with a Darvic PVC ring bearing a unique code. Between 2017 and 2021, the size of the colony fluctuated between 210 and 265 breeding pairs and starting from 2017, every week we recorded the first arrival date of each ringed white stork at the colony using telescopes (Belabed et al., 2019). The weekly transect includes the olive grove where breeding occurs, the adjacent landfill, and the bordering fields. In addition, during the breeding season (March to July), an additional weekly visit is carried out to monitor active nests but any new arrival in the colony and environs is also recorded. Arrival dates were determined as day-of-the-year (DOY) starting from January 1.
Initial data exploration was carried out by seeking outliers and assessment for zero inflation. To test the influence of age and year on arrival date and to avoid pseudoreplication, we conducted a generalized additive mixed model (GAMM) using a log-link function and a Poisson error distribution. The GAMM was performed using the package “gamm4” with the default penalized thin plate regressions splines and the default number of basis functions (Wood et al., 2012). Bird age and year of arrival were used as fixed effects, whereas individuals were included as random variables to account for repeat records and intra-individual variance. Sex could not be incorporated as an independent variable because not all birds had been sexed. Model validation included tests of homogeneity, normality, influential observations, and independence, which were carried out to check model robustness. To account for the residual variance within each age class, we ran a hierarchical cluster analysis (HCA) (Jain et al., 1999) on arrival time of a cohort of individuals aged 2–10 years between 2017 and 2021. HCA was conducted using the package “MASS” (Venables and Ripley, 2002). Furthermore, a linear regression between arrival date (response variable) and age (explanatory variable) was performed for each cluster resulting from the HCA using the package “stats.” All statistical analyses were performed using R (R Development Core Team, 2021).
Between 2017 and 2021, we collected 410 records from 153 individual birds (aged 1–10 years) ringed as fledglings between 2011 and 2020 (Table 1) and corresponding to 1–5 records per individual. Over the past 5 years, first arrival dates varied, with the earliest being 19 December (9- and 10-year-old individuals) and the latest being 7 June (1-year old individuals) (Figure 1A). Density plots of numbers of arrivals against arrival dates for different years clearly indicate a consistent trend for earlier arrival dates in more recent years (Figure 1B), a trend that is less apparent for the second quartiles, and that dissipates for the final two quartiles.
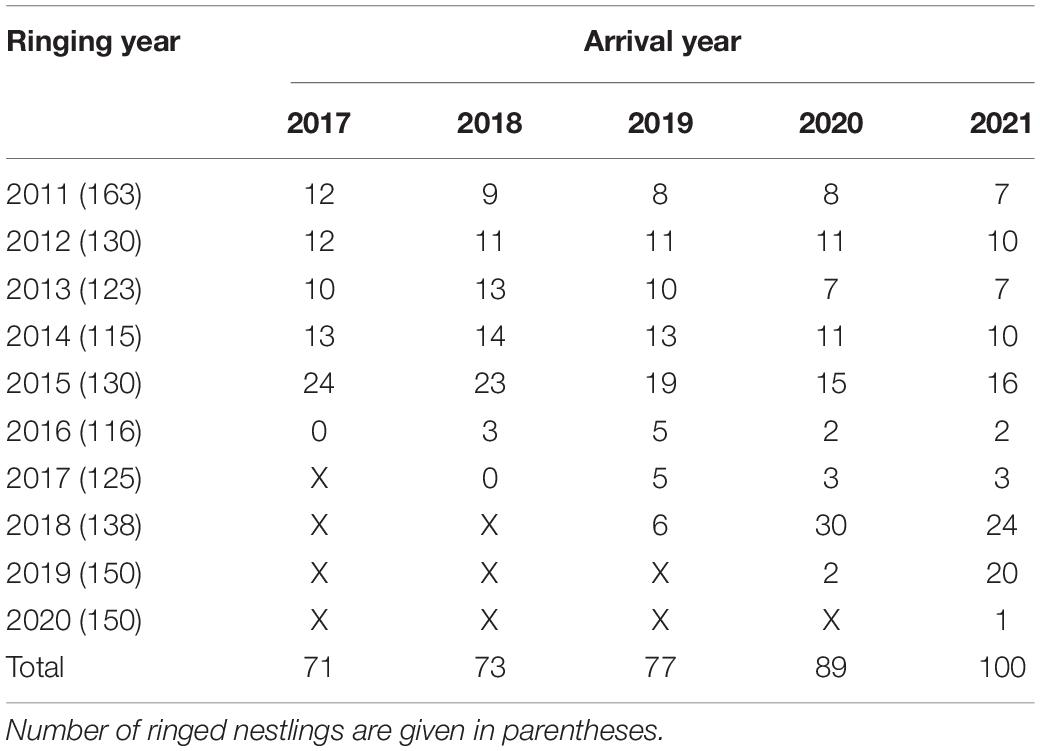
Table 1. Numbers of returning white storks recorded between 2017 and 2021 (columns) that had been ringed between 2011 and 2020 (rows).
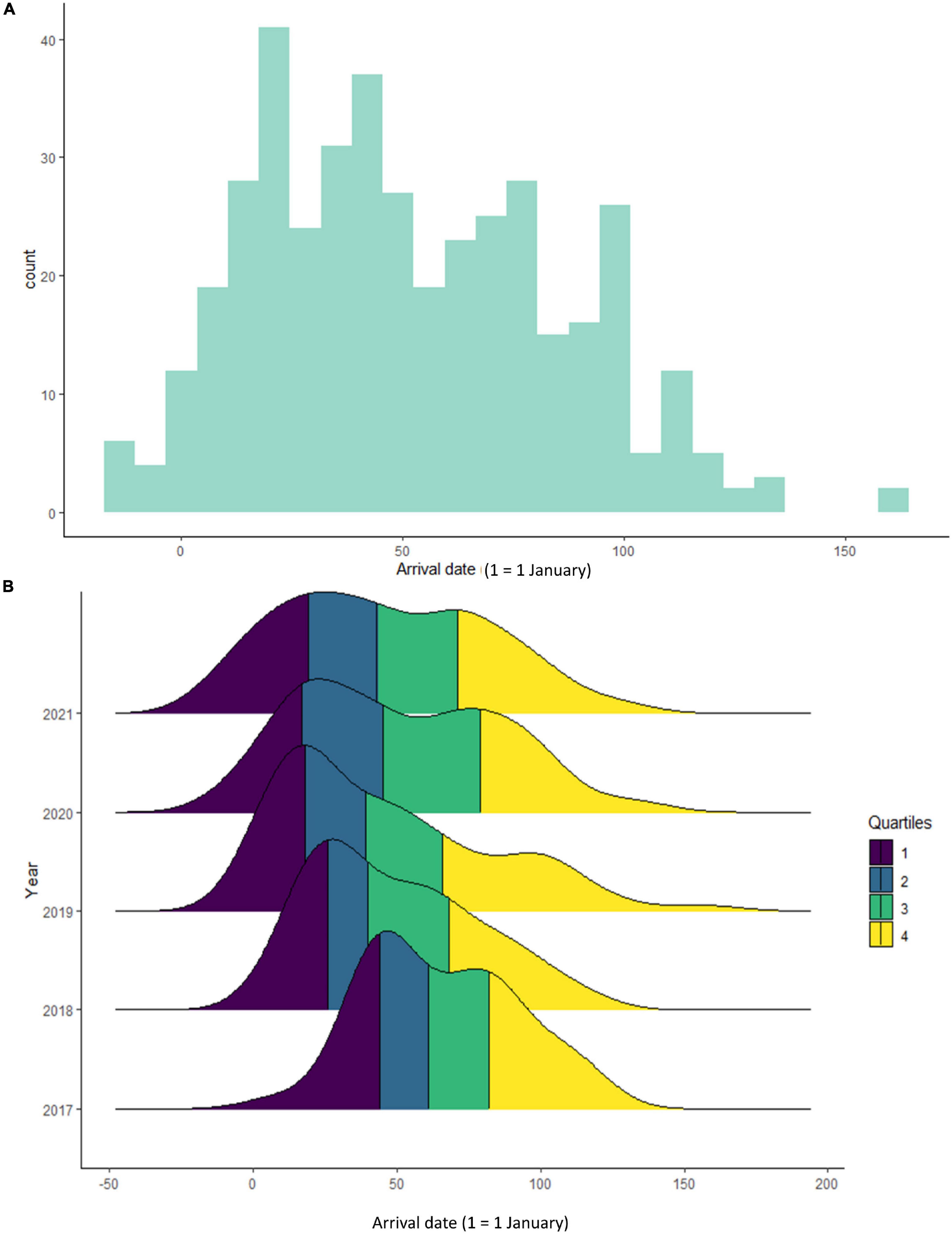
Figure 1. Arrival dates of white storks at the Dréan colony (Algeria) between 2017 and 2021: (A) histogram, (B) density plots displaying quartiles.
A preliminary analysis by applying loess smoothing curves seemed to confirm a negative relationship between age and arrival date (Figure 2A). The GAMM model indicated a non-linear relationship between date of arrival and age, with older birds arriving earlier at the Algerian breeding ground (Table 2). This negative relationship is somewhat linear for birds aged between 1 and 5 years, with the slope gradually leveling off for older birds (Figure 2B). Our GAMM results also revealed that, after accounting for age, there is a significant trend for birds to arrive earlier at Dréan over the last 5 years (Table 2 and Figure 2C). In fact, the distribution of arrival dates indicates that the Dréan colony is arriving earlier at this breeding ground by ca. 9 days over the last 4 years.
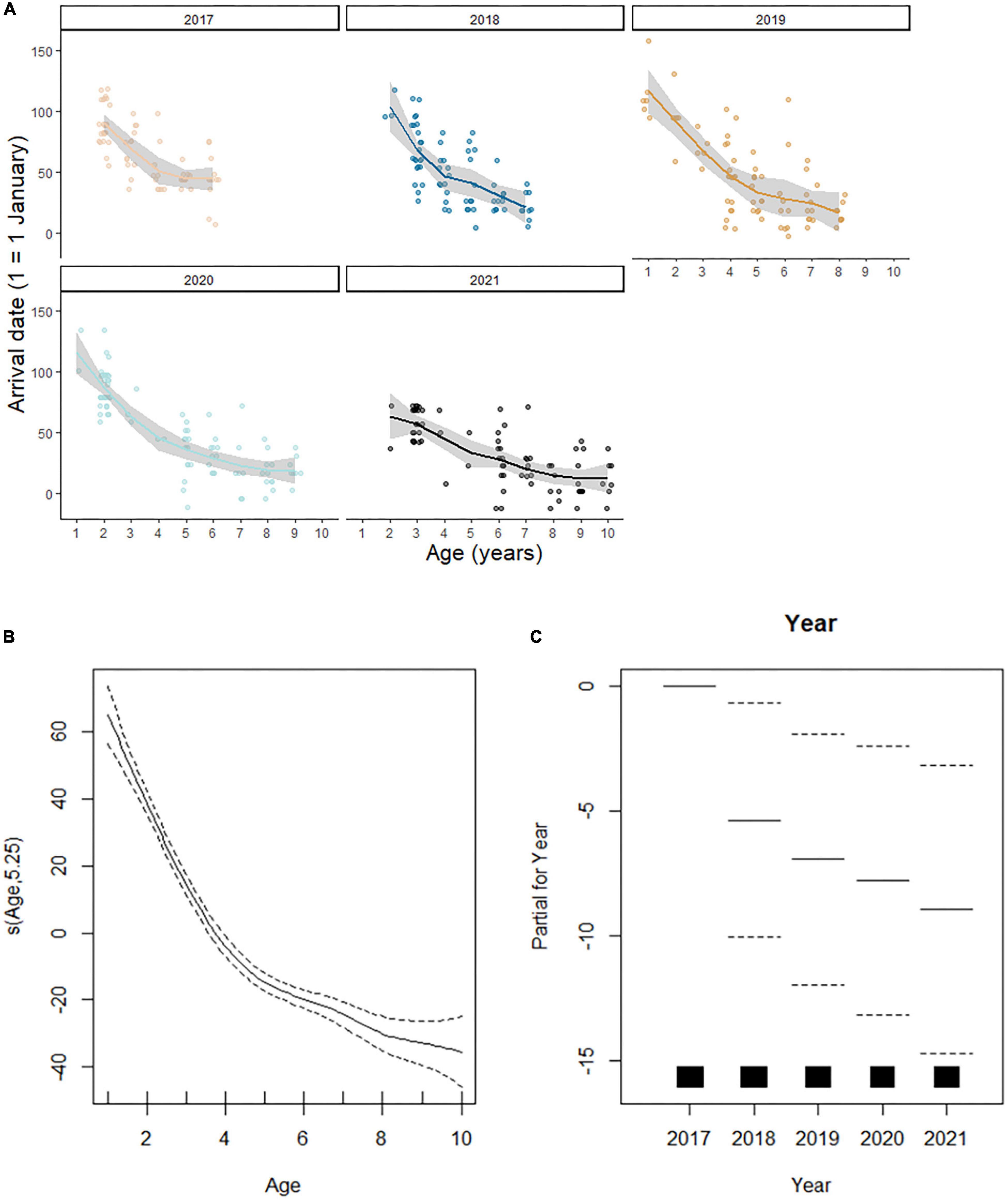
Figure 2. Scatter plots of yearly arrival dates for white storks against bird age between 2017 and 2021 (A), GAMM model fit displaying arrival dates against age (B), and GAMM model displaying arrival dates against year (C).
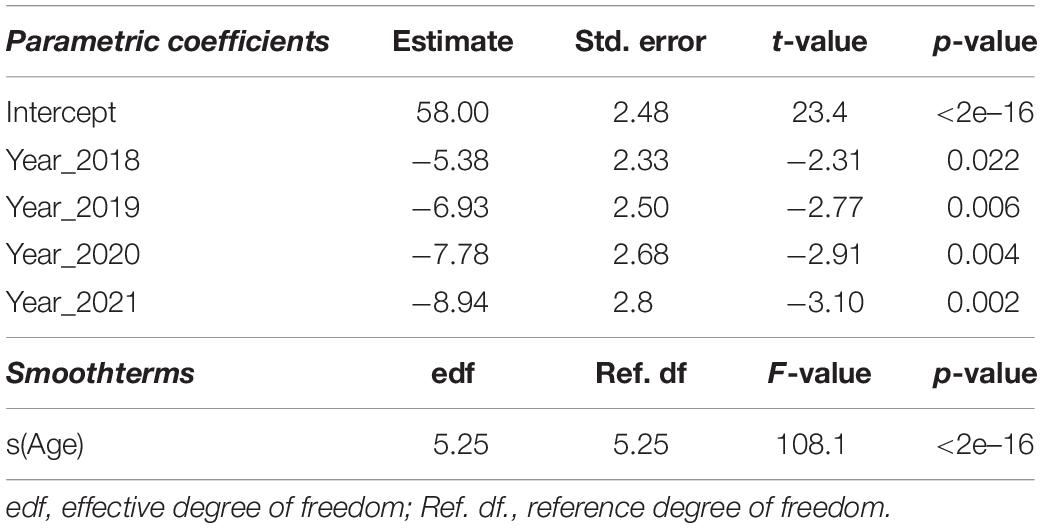
Table 2. Estimated regression parameters, standard errors, t-values, and p-values for the GAMM.
In addition, an HCA indicated that 2–10-year-old birds could be separated into two distinct clusters, which we have labeled “early” and “late” arrivers. Although these two clusters display slight overlap in their distributions of arrival dates (Figure 3), perhaps due to sex effects, arrival dates within each cluster is consistently distinct across all five age-classes we considered, as confirmed by linear regressions (Figure 4). Between 2017 and 2021, and within each age class, “early” birds, as defined by HCA, arrived regularly ahead of time of “late” birds. The disappearance of the slopes of the linear regressions after 2018 simply reflects the absence of 2–3-year-old birds after that year.
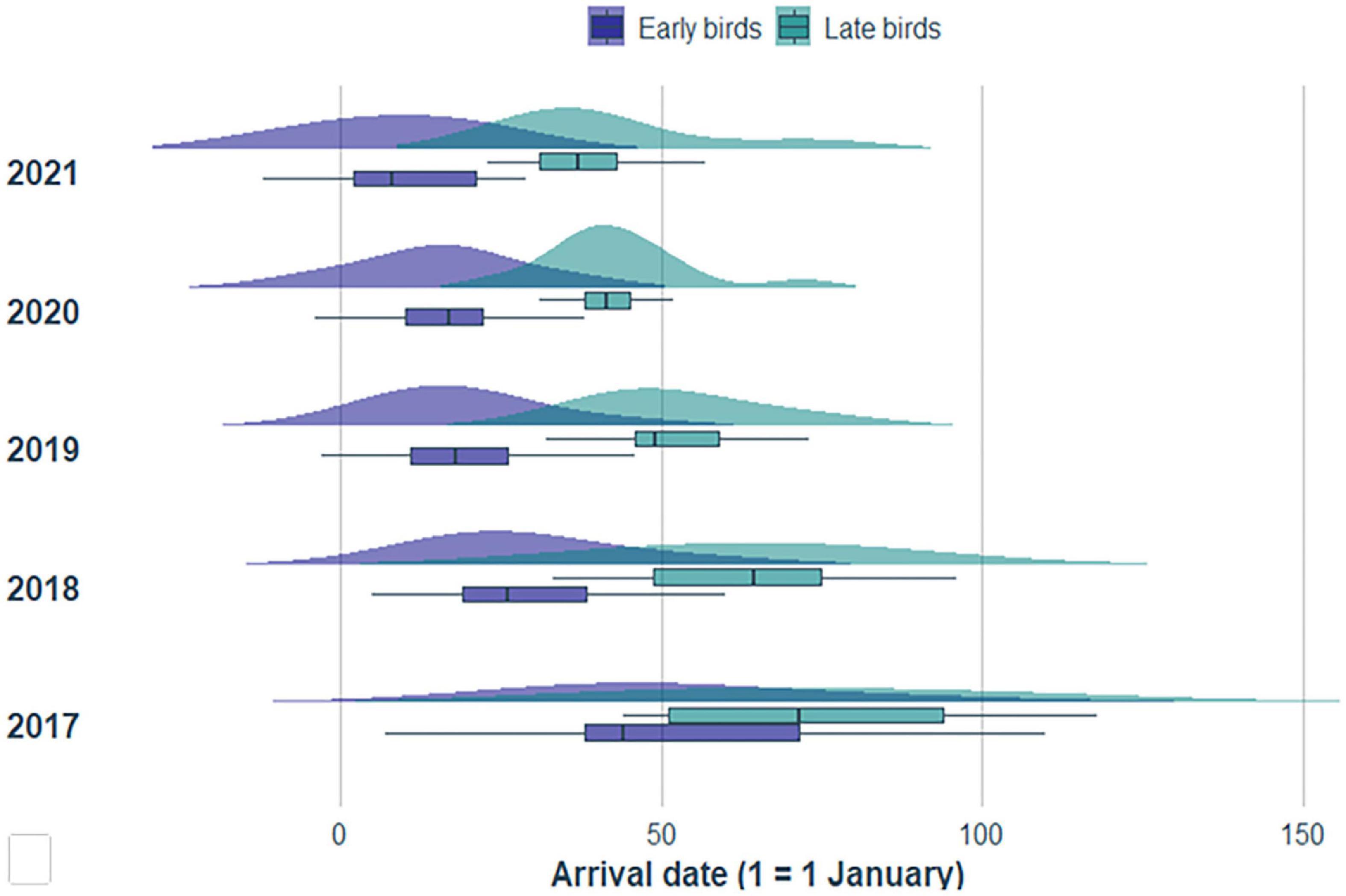
Figure 3. Distributions and boxplots of arrival dates for “early-” and “late-arriving” white storks between 2017 and 2021.
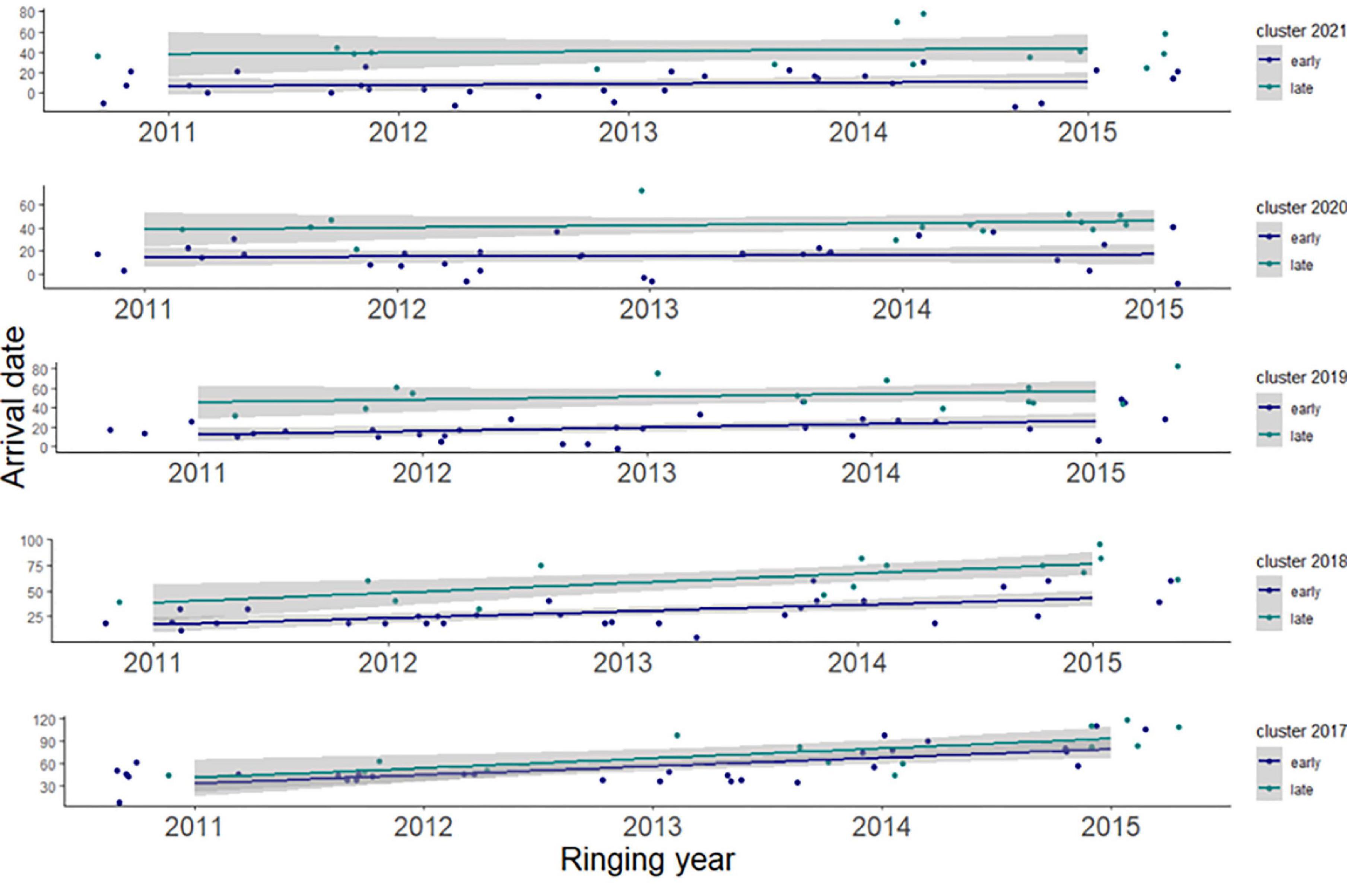
Figure 4. Linear regressions for the “early” and “late” arrival dates of white storks, sorted by age (represented by ringing year, x-axis). Data points have been jittered across the x-axis.
Our study presents evidence that age is the main determinant of the onset of arrival of white storks at their North African breeding grounds. Arrival at these breeding grounds is negatively correlated with age, with older birds arriving before younger individuals, allowing them to settle predominantly on their previous nest sites and thereby avoid the energetic costs of ousting other birds or building new nests. Accordingly, age is also a key factor in egg-laying time and breeding success given their known links to time of arrival (Belabed et al., 2019).
Age-dependent effects may affect fitness profoundly and perpetuate long into a bird’s lifespan (Martin, 1995). There are several possible mechanisms by which age may influence the arrival time of migrant birds. Older birds may select overwintering sites close to their breeding grounds (Teitelbaum et al., 2016), whereas younger birds might disperse further to avoid competition. Older birds may also be better flyers (Rotics et al., 2016) or have amassed greater experience in orienting themselves during travel (Thorup et al., 2003). They may also possess better knowledge of foraging sites (Mueller et al., 2013; Sergio et al., 2014), thereby allowing them to achieve better body condition sooner and facilitating earlier initiation of migration. The bimodal distribution of the arrival dates and the varying shape of the fourth quartile recorded at Dréan might be explained by the differential arrival of resident birds and recruits, but other hypotheses cannot be ruled out. Further studies are necessary to unravel all the mechanisms by which older migratory birds tend to precede younger ones at breeding grounds.
Even after accounting for age, there still remains a considerable amount of variation in arrival dates, with one cohort of birds consistently arriving early and another arriving later (Nolan, 1978; this study). Many explanations have been proposed to account for this variability observed in many migratory bird species (Drent et al., 2003), some of which may not be mutually exclusive (wintering site location, sex, body condition, etc.).
While the specific wintering locations are not known for the Dréan population as only three records/recoveries have been obtained so far: Niger, coastal Libya and Israel (unpublished data), Algerian and Tunisian white storks are known to winter in and around the Lake Chad basin with some individuals dispersing widely in West, Central, and Eastern Africa (Heim de Balsac and Mayaud, 1962; Schulz, 1988; Lauthe, 1989).
To date, few studies have investigated the wintering distribution of North African white storks in any systematic way (Mullié et al., 1995). However, a recent study of juveniles, fitted with high-resolution, solar GSM-GPS-ACC loggers, from neighboring Tunisia indicated that they winter during the first 5 months in a relatively narrow latitudinal band south of the Sahel zone (Flack et al., 2016). If confirmed and extended to all age and sex groups, this lack of strong latitudinal segregation in wintering grounds among North African white storks would imply limited effect from wintering location on arrival date at the breeding grounds.
Furthermore, sex and genetic components cannot be excluded as factors contributing to this age-independent variability. For instance, protandry (males systematically arriving before females) has not been demonstrated unequivocally in white storks (Barbraud and Barbraud, 1999; Vergara et al., 2007; see Gordo et al., 2013), though discrepancies among datasets may be attributable to sexing errors or small sampling sizes (Rotics et al., 2018). Accordingly, further detailed studies are needed to investigate this phenomenon and test the protandry hypothesis.
In addition, our study indicates that white storks are arriving at the breeding grounds earlier in more recent years. To circumvent shortcomings associated with previous analytical approaches (discussed further below), we assessed the entire distribution of arrival dates for the colony at Dréan. In doing so, we have shed light on the importance of the colony age structure, as revealed by the different trend displayed by the third and fourth quartiles of our density distributions (Figure 1B) relative to the first and second quartiles, with the pattern of the third and fourth quartiles very much dependent on survival of younger birds. This latter scenario is supported by the notable lateral extension of the fourth quartiles in years when colony recruitment is high, i.e., late arrivals are more abundant. German and Austrian young storks have been shown to overwinter “closer-to-home” (Cheng et al., 2019). This strategy implying shorter distance migrantions increases their survival probability and allow them to arrive earlier at their breeding grounds (Cheng et al., 2019). Such flexible strategy could explain the rapid shift in arrival time but this hypothesis needs to be tested for North African white storks.
Previous studies of the arrival time of migrant birds, especially long-distance migrants, have suffered from methodological limitations (Gordo et al., 2013) or lack strong theoretical frameworks. Thus, much of the published research to date has been descriptive in nature and is focused on a limited set of species. Some previous analyses of migrant arrival dates assessed the first field record of a given species in each year (Lehikoinen et al., 2004; Knudsen et al., 2011), but such analyses are sensitive to population size and sampling effort biases and only pertain to the left-hand portion of the arrival date distribution (Miller-Rushing et al., 2008; Wilson, 2013). Other studies focused on the mean or median date of arrival (Tryjanowski and Sparks, 2001; Kullberg et al., 2015), whereas others employed quartiles to distinguish early and late breeders (Wilson, 2007; Lourenço et al., 2011; Janiszewski et al., 2013).
Despite our limited time-series (5 years), we argue that our results unambiguously demonstrate age as the predominant driver of arrival timing patterns in this white stork colony. Furthermore, the fact that we detected a trend in earlier migration (∼-2.3 days year–1) within such a small time frame suggests that climate change might be having a stronger impact now than reported previously (Knudsen et al., 2011). Alternatively, since climate change patterns differ spatially, local bird populations may respond accordingly (Martin et al., 2016). Although direct comparisons are fraught with methodological difficulties, the change in arrival date for North African white storks is far greater than the mean −0.4 days year–1 earlier migration recorded for European migrating birds (Lehikoinen et al., 2004). Similarly, white storks in western Poland have been recorded as arriving in wintering grounds ca. 10 days earlier than they did a century ago (Ptaszyk et al., 2003). Moreover, local environmental changes may also influence arrival dates, with earlier arrival dates of −0.31 and −0.62 days year–1 for rural and urban Polish birds, respectively (Tryjanowski et al., 2013). Given that certain environmental changes are likely to vary in a non-linear fashion, even faster responses might be expected. Thus, further studies are warranted to explore if these observed changes in bird migratory patterns are part of a long-term trend.
There are significant fitness benefits associated with arriving early at breeding sites, but there are also notable costs. For instance, early arrival time has been linked to enhanced breeding success (Smith and Moore, 2005), with early-arriving birds acquiring better quality nesting locations and territories (Bensch and Hasselquist, 1991; Aebischer et al., 1996), having greater success at finding mates (Møller, 1994; Lozano et al., 1996; Smith and Moore, 2005; Gunnarsson and Tómasson, 2011; Janiszewski et al., 2013), and producing early-hatching offspring that may exhibit greater survival (Verboven and Visser, 1998; Monros et al., 2002; Lok et al., 2017).
However, the arrival time at breeding sites of migrant birds is usually constrained by stabilizing selection (Kissner et al., 2003), with costs associated with low food supply and adverse weather conditions during migration and upon arrival at the breeding grounds (Whitmore et al., 1977; Brown and Brown, 2000; Newton, 2007b). This trade-off may vary according to geographical regions. For instance, early-arriving birds in North Africa may face less onerous costs than birds in central Europe, even though infrequent weather extremes do occur in the former so that sharp temperature declines have led to mass breeding failures in mixed colonies of herons (B.S. unpublished).
Another cost associated with early arrival is that food may be in short supply. Indeed, a common concern is that the impact of global warming on communities may lead to a decoupling of phenological relationships between species and their prey, resulting in mismatches and population decline (Durant et al., 2007; Møller et al., 2008). However, landfill sites (such as occur near Dréan) and availability of other anthropogenic food sources may offset such a limitation. Indeed, many birds, including white storks, regularly rely upon urban food subsidies during the breeding season (Gilbert et al., 2016). Over the last two decades, a multiplication of landfill sites has considerably altered the Algerian landscape, leading to increased population sizes of commensal species such as cattle egrets Ardea ibis, yellow-legged gulls Larus michahellis, and white storks. Consequently, selective pressures on white storks to synchronize their breeding cycle with food supplies may slacken (Lack, 1968).
Climate change is widely accepted as causing phenological shifts worldwide, including changes in migration schedules that represent compelling evidence of adaptive responses by living organisms. For instance, previous analyses of environmental factors affecting the spring arrival of migrant birds revealed an association with mean spring temperatures and fluctuations of the North Atlantic Oscillation (NAO), which drives weather patterns in western Europe, North Africa, and North America (Hüppop and Hüppop, 2003; Vähätalo et al., 2004; Wilson, 2007, 2009). Moreover, climatic conditions at overwintering sites have been shown to influence the migration schedules of long-distance migrants (Gordo et al., 2005).
Given that early arrival is an important determinant of breeding success (Belabed et al., 2019), we anticipate that the Algerian colony of white storks will continue to arrive early at Dréan in the absence of stabilizing selection (Brinkhof et al., 2002). Climate-driven evolutionary changes may not only affect the timing of spring migration, but also influence migratory behavior. Where costs associated with early arrival are partially offset by the presence of landfills, it may become advantageous for white storks to reside year-round in breeding grounds if the time frame to be devoted to migration can be shortened considerably and food subsidies are available. Indeed, over the last three decades, white storks have taken advantage of the large-scale habitat transformation engendered by the multiplication of landfills, overwintering at certain breeding grounds in southern Europe and North Africa rather than migrating at all (Tortosa et al., 1995; Samraoui, 1998; Gilbert et al., 2016). Nevertheless, the strong dependence of white storks on food wastes entails other perils, particularly biocontamination (Plaza and Lambertucci, 2020).
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was reviewed and approved by the Algerian Ministère de l’Enseignement Supérieur et de la Recherche Scientifique.
BS and HE-S conceived the ideas and designed methodology. MA, B-EB, KS, AB, and LT collected the data. BS and KS analyzed the data. BS and FS led the writing of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
This study was financially supported by King Saud University, Riyadh, Saudi Arabia, through the Researchers Supporting Project Number (RSP-2021/19).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
This work is dedicated to the memory of “Kouka” Athamnia. We are most grateful to the two reviewers for their helpful comments and suggestions. Last but not least, we thank King Saud University, Riyadh, Saudi Arabia, and its Researchers Supporting Project Number (RSP-2021/19) for material support.
Aebischer, A., Perrin, N., Krieg, M., Studer, J., and Meyer, D. R. (1996). The role of territory choice, mate choice and arrival date on breeding success in the Savi’s warbler Locustella luscinioides. J. Avian Biol. 27, 134–152. doi: 10.2307/3677143
Barbraud, C., and Barbraud, J. C. (1999). Is there age assortative mating in the European white stork? Waterbirds 22, 478–481. doi: 10.2307/1522129
Belabed, B.-E., Athamnia, M., Touati, L., Samraoui, F., Boucheker, A., and Samraoui, B. (2019). The early bird catches the worm: age-specific arrival time influences reproductive performance in the white stork Ciconia ciconia. Bird Study 66, 121–129. doi: 10.1080/00063657.2019.1618240
Bensch, S., and Hasselquist, D. (1991). Territory infidelity in the polygynous Great Reed warbler Acrocephalus arundinaceus: the effect of variation in territory attractiveness. J. Anim. Ecol. 60, 857–871. doi: 10.2307/5418
Boisvert-Marsh, L., Périé, C., and de Blois, S. (2014). Shifting with climate? Evidence for recent changes in tree species distribution at high latitudes. Ecosphere 5:83. doi: 10.1890/ES14-00111.1
Both, C., Bouwhuis, S., Lessells, C. M., and Visser, M. E. (2006). Climate change and population declines in a long-distance migratory bird. Nature 441, 81–83. doi: 10.1038/nature04539
Both, C., and Visser, M. E. (2005). The effect of climate change on the correlation between avian life-history traits. Glob. Change Biol. 11, 1606–1613. doi: 10.1111/j.1365-2486.2005.01038.x
Bouriach, M., Samraoui, F., Souilah, R., Houma, I., Razkallah, I., Alfarhan, A. H., et al. (2015). Is a core-periphery gradient a determinant factor of breeding performance in the colonial breeding white storks Ciconia ciconia? Acta Ornithol. 50, 149–156. doi: 10.3161/00016454ao2015.50.2.003
Brinkhof, M. W. G., Cavé, A. J., Daan, S., and Perdeck, A. C. (2002). Timing of current reproduction directly affects future reproductive output in European coots. Evolution 56, 400–411. doi: 10.1111/j.0014-3820.2002.tb01349.x
Brown, C. R., and Brown, M. B. (2000). Weather-mediated natural selection on arrival time in cliff swallows (Petrochelidon pyrrhonota). Behav. Ecol. Sociobiol. 47, 339–345. doi: 10.1007/s002650050674
Chen, I.-C., Hill, J. K., Ohlemüller, R., Roy, D. B., and Thomas, C. D. (2011). Rapid range shifts of species associated with high levels of climate warming. Science 333, 1024–1026. doi: 10.1126/science.1206432
Cheng, Y., Fiedler, W., Wikelski, M., and Flack, A. (2019). “Closer-to-home” strategy benefits juvenile survival in a long-distance migratory bird. Ecol. Evol. 9, 8945–8952. doi: 10.1002/ece3.5395
Cooper, N. W., Sherry, T. W., and Marra, P. P. (2015). Experimental reduction of winter food decreases body condition and delays migration in a long-distance migratory bird. Ecology 96, 1933–1942. doi: 10.1890/14-1365.1
Coppack, T., and Both, C. (2002). Predicting life-cycle adaptation of migratory birds to global climate change. Ardea 90, 369–378. doi: 10.5253/arde.v90i3.p369
Dittmann, T., and Becker, P. H. (2003). Sex, age, experience and condition as factors affecting arrival date in prospecting common terns, Sterna hirundo. Anim. Behav. 65, 981–986. doi: 10.1006/anbe.2003.2128
Drent, R., Both, C., Green, M., Madsen, J., and Piersma, T. (2003). Pay-offs and penalties of competing migratory schedules. Oikos 103, 274–292. doi: 10.1034/j.1600-0706.2003.12274.x
Durant, J. M., Hjermann, D. Ø., Ottersen, G., and Stenseth, N. C. (2007). Climate and the match or mismatch between predator requirements and resource availability. Clim. Res. 33, 271–283. doi: 10.3354/cr033271
Flack, A., Fiedler, W., Blas, J., Pokrovsky, I., Mitropolsky, M., Aghababyan, K., et al. (2016). Costs of migratory decisions: a comparison across eight white stork populations. Sci. Adv. 2:e1500931. doi: 10.1126/sciadv.1500931
Fulin, M., Jerzak, L., Sparks, T. H., and Tryjanowski, P. (2009). Relationship between arrival date, hatching date and breeding success of the white stork (Ciconia ciconia) in Slovakia. Biologia 64, 361–364. doi: 10.2478/s11756-009-0037-3
Gienapp, P., Teplitsky, C., Alho, J. S., Mills, J. A., and Merilä, J. (2008). Climate change and evolution: disentangling environmental and genetic responses. Mol. Ecol. 17, 167–178. doi: 10.1111/j.1365-294X.2007.03413.x
Gilbert, N. I., Correia, R. A., Silva, J. P., Pacheco, C., Catry, I., Atkinson, P. W., et al. (2016). Are white storks addicted to junk food? Impacts of landfill use on the movement and behavior of resident white storks (Ciconia ciconia) from a partially migratory population. Mov. Ecol. 4:7. doi: 10.1186/s40462-016-0070-0
Gordo, O. (2007). Why are bird migration dates shifting? A review of weather and climate effects on avian migratory phenology. Clim. Res. 35, 37–58. doi: 10.3354/cr00713
Gordo, O., Brotons, L., Ferrer, X., and Comas, P. (2005). Do changes in climate patterns in wintering areas affect the timing of the spring arrival of trans-Saharan migrant birds? Glob. Change Biol. 11, 12–21. doi: 10.1111/j.1365-2486.2004.00875.x
Gordo, O., Tryjanowski, P., Kosicki, J. Z., Fulin, M., and Both, C. (2013). Complex phenological changes and their consequences in the breeding success of a migratory bird, the white stork Ciconia ciconia. J. Anim. Ecol. 82, 1072–1086. doi: 10.1111/1365-2656.12084
Gunnarsson, T. G., and Tómasson, G. (2011). Flexibility in spring arrival of migratory birds at northern latitudes under rapid temperature changes. Bird Study 58, 1–12. doi: 10.1080/00063657.2010.526999
Harel, R., Horvitz, N., and Nathan, R. (2016). Adult vultures outperform juveniles in challenging thermal soaring conditions. Sci. Rep. 6:27865. doi: 10.1038/srep27865
Heim de Balsac, H., and Mayaud, N. (1962). Les Oiseaux du Nord-Ouest de l’Afrique: Distribution Géographique, Écologie, Migrations, Reproduction. Paris: P. Lechevalier.
Hüppop, O., and Hüppop, K. (2003). North Atlantic Oscillation and timing of spring migration in birds. Proc. R. Soc. Lond. B Biol. Sci. 270, 233–240. doi: 10.1098/rspb.2002.2236
IPCC (2014). Climate Change (2014): Synthesis Report. Available online at: https://www.ipcc.ch/site/assets/uploads/2018/05/SYR_AR5_FINAL_full_wcover.pdf (accessed December 21, 2021).
Jain, A., Murty, M., and Flynn, P. (1999). Data clustering: a review. ACM Comput. Surv. 31, 264–323. doi: 10.1145/331499.331504
Janiszewski, T., Minias, P., and Wojciechowski, Z. (2013). Reproductive consequences of early arrival at breeding grounds in the white stork Ciconia ciconia. Bird Study 60, 280–284. doi: 10.1080/00063657.2013.778227
Kissner, K. J., Weatherhead, P. J., and Francis, C. M. (2003). Sexual size dimorphism and timing of spring migration in birds. J. Evol. Biol. 16, 154–162. doi: 10.1046/j.1420-9101.2003.00479.x
Knudsen, E., Lindén, A., Both, C., Joneén, N., Pulido, F., Saino, N., et al. (2011). Challenging claims in the study of migratory birds and climate change. Biol. Rev. 86, 928–946. doi: 10.1111/j.1469-185X.2011.00179.x
Kullberg, C., Fransson, T., Hedlund, J., Jonzén, N., Langvall, O., Nilsson, J., et al. (2015). Change in spring arrival of migratory birds under an era of climate change, Swedish data from the last 140 years. Ambio 44(Suppl. 1), S69–S77. doi: 10.1007/s13280-014-0600-1
Lack, D. (1968). Ecological Adaptations for Breeding in Birds. London: Methuen. doi: 10.1017/s003060530000750x
Lauthe, P. (1989). “Status, verbreitung und zugdes weißstorchs in Tunesien,” in Proc. 1st Int. Stork Conserv. Symp. Schriftenreihe des DDA 10 White Stork. Status and Conservation, eds G. Rheinwald, J. Ogden, and H. Schulz, 189–194.
Lehikoinen, E., Sparks, T. H., and Žalakevičius, M. (2004). Arrival and departure dates. Adv. Ecol. Res. 35, 1–31. doi: 10.1016/S0065-2504(04)35001-4
Lok, T., Veldhoen, L., Overdijk, O., Tinbergen, J. M., and Piersma, T. (2017). An age-dependent fitness cost of migration? Old trans-Saharan migrating spoonbills breed later than those staying in Europe, and late breeders have lower recruitment. J. Anim. Ecol. 86, 998–1009. doi: 10.1111/1365-2656.12706
Lourenço, P. M., Kentie, R., Schroeder, J., Groen, N. M., Hooijmeijer, J. C. E. W., and Piersma, T. (2011). Repeatable timing of northward departure, arrival and breeding in Black-tailed Godwits Limosa l. limosa, but no domino effects. J. Ornithol. 152, 1023–1032. doi: 10.1007/s10336-011-0692-3
Lozano, G. A., Perreault, S., and Lemon, R. E. (1996). Age, arrival date and reproductive success of male American redstarts Setophaga ruticilla. J. Avian Biol. 27, 164–170. doi: 10.2307/3677146
Marra, P. P., Francis, C. M., Mulvihill, R. S., and Moore, F. R. (2005). The influence of climate on the timing and rate of spring bird migration. Oecologia 142, 307–315. doi: 10.1007/s00442-004-1725-x
Martin, B., Onrubia, A., and Ferrer, M. (2016). Migration timing responses to climate change differ between adult and juvenile white storks across Western Europe. Clim. Res. 69, 9–23. doi: 10.3354/cr01390
Martin, K. (1995). Patterns and mechanisms for age-dependent reproduction and survival in birds. Am. Zool. 35, 340–348. doi: 10.1093/icb/35.4.340
Matyjasiak, P., Olejniczak, I., Boniecki, P., and Møller, A. P. (2013). Wing characteristics and spring arrival date in Barn Swallows Hirundo rustica. Acta Ornithol. 48, 81–92. doi: 10.3161/000164513x670025
Menzel, A., Sparks, T. H., Estrella, N., Koch, E., Aasa, A., Ahas, R., et al. (2006). European phenological response to climate change matches the warming pattern. Glob. Change Biol. 12, 1969–1976. doi: 10.1111/j.1365-2486.2006.01193.x
Miller-Rushing, A. J., Loyd-Evans, T. L., Primack, R. B., and Satzinger, P. (2008). Bird migration times, climate change, and changing population sizes. Glob. Change Biol. 14, 1959–1972. doi: 10.1111/j.1365-2486.2008.01619.xr2008
Møller, A. P. (1994). Phenotype-dependent arrival time and its consequences in a migratory bird. Behav. Ecol. Sociobiol. 35, 115–122. doi: 10.1007/s002650050077
Møller, A. P., Rubolini, D., and Lehikoinen, E. (2008). Populations of migratory bird species that did not show a phenological response to climate change are declining. Proc. Natl. Acad. Sci. U.S.A. 105, 16195–16200. doi: 10.1073/pnas.0803825105
Monros, J. S., Belda, E. J., and Barba, E. (2002). Post-fledging survival of individual great tits: the effect of hatching date and fledging mass. Oikos 99, 481–488. doi: 10.1034/j.1600-0706.2002.11909.x
Moore, F. R., Mabey, S., and Woodrey, M. (2003). “Priority access to food in migratory birds: age, sex and motivational asymmetries,” in Avian Migration, eds P. Berthold, E. Gwinner, and E. Sonnenschein (Berlin: Springer), 281–292. doi: 10.1007/978-3-662-05957-9_19
Mueller, T., O’Hara, R. H., Converse, S. J., Urbanek, R. P., and Fagan, W. F. (2013). Social learning of migratory performance. Science 341, 999–1002. doi: 10.1126/science.1237139
Mullié, W. C., Brouwer, J., and Schott, P. (1995). “Numbers, distribution and habitat of wintering White Storks in the east-central Sahel in relation to rainfall, food and anthropogenic influences,” in Proceedings of the International Symposium on the White Stork (Western Population), eds O. Biber, P. Enggist, C. Marti, and T. Salathé (Basel), 219–240. doi: 10.2307/1521455
Newton, I. (2007b). Weather-related mass-mortality events in migrants. Ibis 149, 453–467. doi: 10.1111/j.1474-919x.2007.00704.x
Nolan, V. Jr. (1978). The ecology and behavior of the prairie warbler Dendroica discolor. Ornithol. Monogr. 26, 1–595. doi: 10.2307/40166732
Ouwehand, J., and Both, C. (2017). African departure rather than migration speed determines variation in spring arrival in pied flycatchers. J. Anim. Ecol. 86, 88–97. doi: 10.1111/1365-2656
Plaza, P. I., and Lambertucci, S. A. (2020). How are garbage dumps impacting vertebrate demography, health, and conservation? Glob. Ecol. Conserv. 12, 9–20. doi: 10.1016/j.gecco.2017.08.002
Ptaszyk, J., Kosicki, J., Sparks, T. H., and Tryjanowski, P. (2003). Changes in the timing and pattern of arrival of the White Stork (Ciconia ciconia) in western Poland. J. Ornithol. 144, 323–329. doi: 10.1046/j.1439-0361.2003.03011.x
R Development Core Team (2021). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Root, T. L., Price, J. T., Hall, K. R., Schneider, S. H., Rosenzweig, C., and Pounds, J. A. (2003). Fingerprints of global warming on wild animals and plants. Nature 421, 57–60. doi: 10.1038/nature01333
Rotics, S., Kaatz, M., Resheff, Y. S., Feldman, S., Zurell, D., Sapir, N., et al. (2016). The challenges of the first migration: movement and behavior of juvenile versus adult white storks with insights regarding juvenile mortality. J. Anim. Ecol. 85, 938–947. doi: 10.1111/1365-2656.12525
Rotics, S., Kaatz, M., Turjeman, S., Zurell, D., Wikelski, M., Sapir, N., et al. (2018). Early arrival at breeding grounds: causes, costs and a trade-off with overwintering latitude. J. Anim. Ecol. 87, 1627–1638. doi: 10.1111/1365-2656.12898
Saino, N., Ambrosini, R., Rubolini, D., von Hardenberg, J., Provenzale, A., Hüppop, K., et al. (2011). Climate warming, ecological mismatch at arrival and population decline in migratory birds. Proc. R. Soc. Lond. B Biol. Sci. 278, 835–842. doi: 10.1098/rspb.2010.1778
Schulz, H. (1988). Weißstorchzug – Ôkologie, Gefâhrdung und Schutz des Weißstorchs in Africa und Nahost. Weikersheim: Verlag Josef Margraf.
Sergio, F., Tanferna, A., de Stephanis, R., Jimenez, L. L., Blas, J., Tavecchia, G., et al. (2014). Individual improvements and selective mortality shape lifelong migratory performance. Nature 515, 410–413. doi: 10.1038/nature13696
Smith, R. J., and Moore, F. R. (2005). Arrival timing and seasonal reproductive performance in a long-distance migratory landbird. Behav. Ecol. Sociobiol. 57, 231–239. doi: 10.1007/s00265-004-0855-9
Teitelbaum, C. S., Converse, S., Fagan, W. F., Bohning-Gaese, K., O’Hara, R. B., Lacy, A. E., et al. (2016). Experience drives innovation of new migration patterns of whooping cranes in response to global change. Nat. Commun. 7:12793. doi: 10.1038/ncomms12793
Thorup, K., Alerstam, T., Hake, M., and Kjellen, N. (2003). Bird orientation: compensation for wind drift in migrating raptors i sage dependent. Proc. R. Soc. Lond. B Biol. Sci. 270, S8–S11. doi: 10.1098/rsbl.2003.0014
Tomotani, B. M., Gienapp, P., de la Hera, I., Terpstra, M., Pulido, F., and Visser, M. E. (2021). Integrating causal and evolutionary analysis of life-history evolution: arrival date in a long-distance migrant. Front. Ecol. Evol. 9:630823. doi: 10.3389/fevo.2021.630823
Tortosa, F. S., Mañez, M., and Barcell, M. (1995). Wintering white storks (Ciconia ciconia) in South West Spain in the years 1991 and 1992. Vogelwarte 38, 41–45.
Tryjanowski, P., and Sparks, T. H. (2001). Is the detection of the first arrival date of migrating birds influenced by population size? A case study of the red-backed shrike Lanius collurio. Int. J. Biometeorol. 45, 217–219. doi: 10.1007/s00484-001-0112-0
Tryjanowski, P., Sparks, T. H., Kuźniak, S., Czechowski, P., and Jerzak, L. (2013). Bird migration advances more strongly in urban environments. PLoS One 8:e63482. doi: 10.1371/journal.pone.0063482
Tryjanowski, P., Sparks, T. H., Ptaszyk, J., and Kosicki, J. (2004). Do white storks Ciconia ciconia always profit from an early return to their breeding grounds? Bird Study 51, 222–227. doi: 10.1080/00063650409461357
Vähätalo, A. V., Raino, K., Lehikoinen, A., and Lehikoinen, E. (2004). Spring arrival of birds depends on the North Atlantic Oscillation. J. Avian Biol. 35, 210–216. doi: 10.1111/j.0908-8857.2004.03199.x
Vaitkuviene, D., Dagys, M., Bartkeviciene, G., and Romanovskaja, D. (2015). The effect of weather variables on the white stork (Ciconia ciconia) spring migration phenology. Ornis Fenn. 92, 43–52.
Verboven, N., and Visser, M. E. (1998). Seasonal variation in local recruitment of great tits: the importance of being early. Oikos 81, 511–524. doi: 10.2307/3546771
Vergara, P., Aguirre, J. I., and Fernández-Cruz, M. (2007). Arrival date, age and breeding success in white stork Ciconia ciconia. J. Avian Biol. 38, 573–579. doi: 10.1111/j.0908-8857.2007.03983.x
Visser, M. E., Both, C., and Lambrechts, M. M. (2004). Global climate change leads to mistimed avian reproduction. Adv. Ecol. Res. 35, 89–110. doi: 10.1016/s0065-2504(04)35005-1
Visser, M. E., van Noordwijk, A. J., Tinbergen, J. M., and Lessells, C. M. (1998). Warmer springs lead to mistimed reproduction in great tits (Parus major). Proc. R. Soc. Lond. B Biol. Sci. 265, 1867–1870. doi: 10.1098/rspb.1998.0514
Walther, G.-R., Post, E., Convey, P., Menzel, A., Parmesan, C., Beebee, T. J. C., et al. (2002). Ecological responses to recent climate change. Nature 416, 389–395. doi: 10.1038/416389a
Whitmore, R. C., Mosher, J. A., and Frost, H. H. (1977). Spring migrant mortality during unseasonable weather. Auk 94, 778–781. doi: 10.2307/4085279
Wilson, W. H. Jr. (2007). Spring arrival dates of migratory breeding birds in Maine: sensitivity to climate change. Wilson J. Ornithol. 119, 667–679. doi: 10.1676/06-044.1
Wilson, W. H. Jr. (2009). Variability of arrival dates of Maine migratory birds: implications for detecting climate change. Northeast. Nat. 16, 443–454. doi: 10.1656/045.016.n311
Wilson, W. H. Jr. (2013). A deeper statistical examination of arrival dates of migratory breeding birds in relation to global climate change. Biology 2, 742–754. doi: 10.3390/biology2020742
Keywords: arrival time, Ciconiiformes, climate change, interannual variation, interindividual variation, long-distance migration, North Africa
Citation: Athamnia M, Belabed B-E, Samraoui KR, Boucheker A, Touati L, Samraoui F, El-Serehy HA and Samraoui B (2022) Variability in Arrival Time of White Storks (Ciconia ciconia L.): Impact of Age, Interindividual Variation, and Global Change. Front. Ecol. Evol. 10:841744. doi: 10.3389/fevo.2022.841744
Received: 22 December 2021; Accepted: 09 March 2022;
Published: 06 May 2022.
Edited by:
Brett K. Sandercock, Norwegian Institute for Nature Research (NINA), NorwayReviewed by:
Eunbi Kwon, Max Planck Institute for Ornithology, GermanyCopyright © 2022 Athamnia, Belabed, Samraoui, Boucheker, Touati, Samraoui, El-Serehy and Samraoui. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Boudjéma Samraoui, YnNhbXJhb3VpQGdtYWlsLmNvbQ==
†ORCID: Mohammed Athamnia, orcid.org/0000-0002-0383-8058; Farrah Samraoui, orcid.org/0000-0003-2407-3004; Bourhane-Eddine Belabed, orcid.org/0000-0001-9807-7206; Kenz Raouf Samraoui, orcid.org/0000-0002-5132-6276; Boudjéma Samraoui, orcid.org/0000-0002-0608-9021; Abdennour Boucheker, orcid.org/0000-0001-9433-9934; Laïd Touati, orcid.org/0000-0002-6716-9914; Hamed A. El-Serehy, orcid.org/0000-0002-7686-0010
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.