 L. Grant Bolton
L. Grant Bolton Jaime C. Piñero
Jaime C. Piñero Bruce A. Barrett
Bruce A. Barrett- 1Division of Plant Science and Technology, University of Missouri, Columbia, MO, United States
- 2Stockbridge School of Agriculture, University of Massachusetts, Amherst, MA, United States
Baits and lures for trapping and monitoring the invasive vinegar fly Drosophila suzukii Matsumura (Diptera: Drosophilidae) are currently derived from fermentation volatiles. Volatile organic compounds from alternative sources could improve monitoring efforts and optimize capture of gravid females. Through electroantennography and behavioral assays, we evaluated male and female D. suzukii responses to blends of selected synthetic fruit volatiles in combination with the fruit compound isoamyl acetate and the strawberry leaf terpenoid β-cyclocitral. Blends that were attractive to both male and female D. suzukii were then evaluated for attractiveness to a non-target drosophilid, Drosophila melanogaster. A simple 3-component blend of isoamyl acetate, β-cyclocitral and methyl butyrate was attractive to D. suzukii, particularly females, relative to fresh blueberry volatiles. The 3-component blend was not attractive to D. melanogaster. Additional research is needed to determine the effectiveness of this blend to attract D. suzukii under field conditions.
Introduction
Phytophagous insects utilize volatile organic compounds (VOCs) emitted from plants for host location (Metcalf and Metcalf, 1992; Bruce et al., 2005), and varying blends of the abundant VOCs can have differing effects on an insect’s searching behavior (Bruce and Pickett, 2011). For example, certain mixtures of VOCs have been shown to be more attractive than their individual components for Manduca sexta (L.) (Lepidoptera: Sphingidae) (Fraser et al., 2003), Aphis fabae Scopoli (Hemiptera: Aphididae) (Webster et al., 2008), Anastrepha ludens (Loew) (Diptera: Tephritidae) (Robacker et al., 1992) and the drosophilids Drosophila melanogaster (Meigen) (Zhu et al., 2003), and Drosophila suzukii Matsumura (Piñero et al., 2019), among other species. Such blends of volatiles can play a vital role in integrated pest management (IPM) systems (Metcalf and Metcalf, 1992; Szendrei and Rodriguez-Saona, 2010), especially for monitoring purposes (Rodriguez-Saona and Stelinski, 2009), mass trapping (El-Sayed et al., 2006) and push-pull applications (Cook et al., 2007).
Drosophila suzukii, also known as the spotted-wing drosophila, is an invasive vinegar fly native to Southeast Asia, but whose distribution now includes many of the small and stone fruit growing regions of the world (Lee et al., 2011a,b; Calabria et al., 2012; Cini et al., 2012; Deprá et al., 2014; Ørsted and Ørsted, 2018). Damage done by D. suzukii comes from its ability to oviposit into ripe or ripening soft fruit with a serrated ovipositor (Walsh et al., 2011; Asplen et al., 2015; Karageorgi et al., 2017). Such damaged fruit then becomes unmarketable through larval feeding (Walsh et al., 2011; Ioriatti et al., 2015) which can reach devastating levels if fly populations are left unmanaged (Bolda et al., 2010; Goodhue et al., 2011).
Current methods for fruit damage prevention begin with an effective monitoring program. Commercially available baits for monitoring D. suzukii include fermentation products, such as apple cider vinegar, grape wine, baker’s yeast, sugar, and water mixture, acetic acid, and ethanol (Cha et al., 2012; Landolt et al., 2012a,b; Iglesias et al., 2014; Kleiber et al., 2014). Generally, fermentation volatiles have been associated with D. suzukii feeding sites and fruit volatiles are used to locate oviposition sites (Karageorgi et al., 2017; Mori et al., 2017). A more D. suzukii-specific lure derived from the headspace components of rice vinegar and wine has been developed and incorporated into commercial lures (Cha et al., 2012, 2014, 2017).
Drosophila suzukii has been reported to oviposit on 130 different hosts (Diepenbrock and McPhie, 2018). The overlapping volatile profiles of suitable host fruits may be the cause for such a broad range of potential fruit hosts (Lance, 1983), and identifying species-specific volatiles has been a challenge. Electroantennogram (EAG) tests have revealed that D. suzukii can detect blends of many VOC compounds emanating from ripening fruit (Abraham et al., 2015; Revadi et al., 2015; Karageorgi et al., 2017; Liu et al., 2018). However, for many phytophagous insects competing background odors from ripening fruit often outcompete synthetic attractants (Schröder and Hilker, 2008), and such a phenomenon may be true for D. suzukii as well (Hamby et al., 2014; Hampton et al., 2014; Abraham et al., 2015; Alnajjar et al., 2017).
The strawberry leaf terpenoid, β-cyclocitral, has been previously reported to be specifically attractive to D. suzukii, but not detected by D. melanogaster and some other commonly occurring drosophilid species (Keesey et al., 2015). Using EAG and behavioral bioassays, Bolton et al. (2019) quantified the attractiveness of nine fruit plant volatiles, including β-cyclocitral, to D. suzukii. These authors reported that β-cyclocitral had a synergistic effect on the attractiveness of ethyl hexanoate and an additive effect when combined with methyl butyrate, ethyl acetate, and isoamyl acetate (Bolton et al., 2019). Piñero et al. (2019) demonstrated that the addition of β-cyclocitral to tart cherry juice synergistically increased attraction in female D. suzukii.
Many drosophilids are attracted to the fruit VOC isoamyl acetate, a ubiquitous odor found in the ripening to early fermenting stages of many fruits (Stökl et al., 2010; Keesey et al., 2015; Revadi et al., 2015). Isoamyl acetate has been shown to reduce attraction in D. suzukii when it is combined with ethanol and acetic acid (Cha et al., 2012). However, D. suzukii attraction to isoamyl acetate may depend upon the concentration used and additional components of a mixture (Revadi et al., 2015). It is known that Hanseniaspora uvarum, a yeast microbe closely associated to D. suzukii (Hamby et al., 2012), emits isobutyl and isoamyl acetate as volatiles and it is reported that D. suzukii had higher antennal sensitivity to isoamyl acetate than D. melanogaster did (Scheidler et al., 2015).
The overall goal of this study was to explore the sensitivity and responsiveness of D. suzukii toward β-cyclocitral and isoamyl acetate when combined with several other selected fruit volatiles. The specific objectives of this study were: (1) To quantify electroantennographic responses of male and female D. suzukii to increasingly complex fruit volatile blends, (2) To characterize the level of male and female attraction to selected volatiles in 3, 4, 5, 6, and 7-component blends, (3) To determine the attraction level of selected blends to a non-target drosophilid, and (4) To compare the level of attraction of male and female D. suzukii to selected mixtures of VOCs to a common host fruit.
Materials and Methods
Fly Colony
Drosophila suzukii flies were reared in 177 mL polypropylene square bottom bottles and fed Nutri-fly™ instant formulation diet (Genesee Scientific, San Diego, CA, United States). Stock bottles were held in an environmental growth chamber set at 24°C and a photoperiod of 16L:8D. The initial colony of flies were obtained in 2014 from a laboratory colony maintained at the Trevor Nichols Research Center (Michigan State University), Fennville, MI.
Volatile Organic Compound Treatments
The following VOCs, associated mostly with fruit ripening, were combined into mixtures with the ester isoamyl acetate (98%, CAS No. 123-92-2) and the strawberry foliar terpenoid, β-cyclocitral (≥95%, CAS No. 432-25-7), and examined for their level of attractiveness to D. suzukii: hexyl acetate (≥98%, CAS No. 142-92-7), methyl butyrate (99%, CAS No. 623-42-7), methyl isovalerate (≥98%, CAS No. 556-24-1), 2-heptanone (99%, CAS No. 110-43-0), ethyl hexanoate (≥98%, CAS No. 123-66-0), ethyl acetate (99.8%, CAS No. 141-78-6), and butyl acetate (≥99%, CAS No. 123-86-4). Isoamyl acetate and β-cyclocitral were selected to form the base of all tested blends based on previous laboratory results (Bolton et al., 2019). All VOC treatments were prepared at diluted concentrations of 10–2–10–8 from the neat solution using laboratory-grade mineral oil as the solvent. Blends were mixed in equal proportions (vol:vol) of each component. Mineral oil alone was tested as a control stimulus. All compounds were purchased from Sigma-Aldrich (St. Louis, MO, United States).
Electrophysiological Bioassays
The degree of antennal sensitivity of D. suzukii to the various blends of volatiles were determined by measuring the response amplitude in electroantennogram (EAG) tests. The examined mixtures consisted of 2- to 7-component blends from the VOCs listed above. The 2–3 component set consisted of eight different blends, the 4-component and the 5-component sets consisted of 10 different blends, and the 6- to 7-component sets consisted of six different blends (Tables 1–8), totaling 24 blends tested in this study. All individual mixtures in a particular component set were exposed randomly to a single fly’s antenna (for three VOC puffs per mixture), and there were 10 flies (per sex) tested per component set.
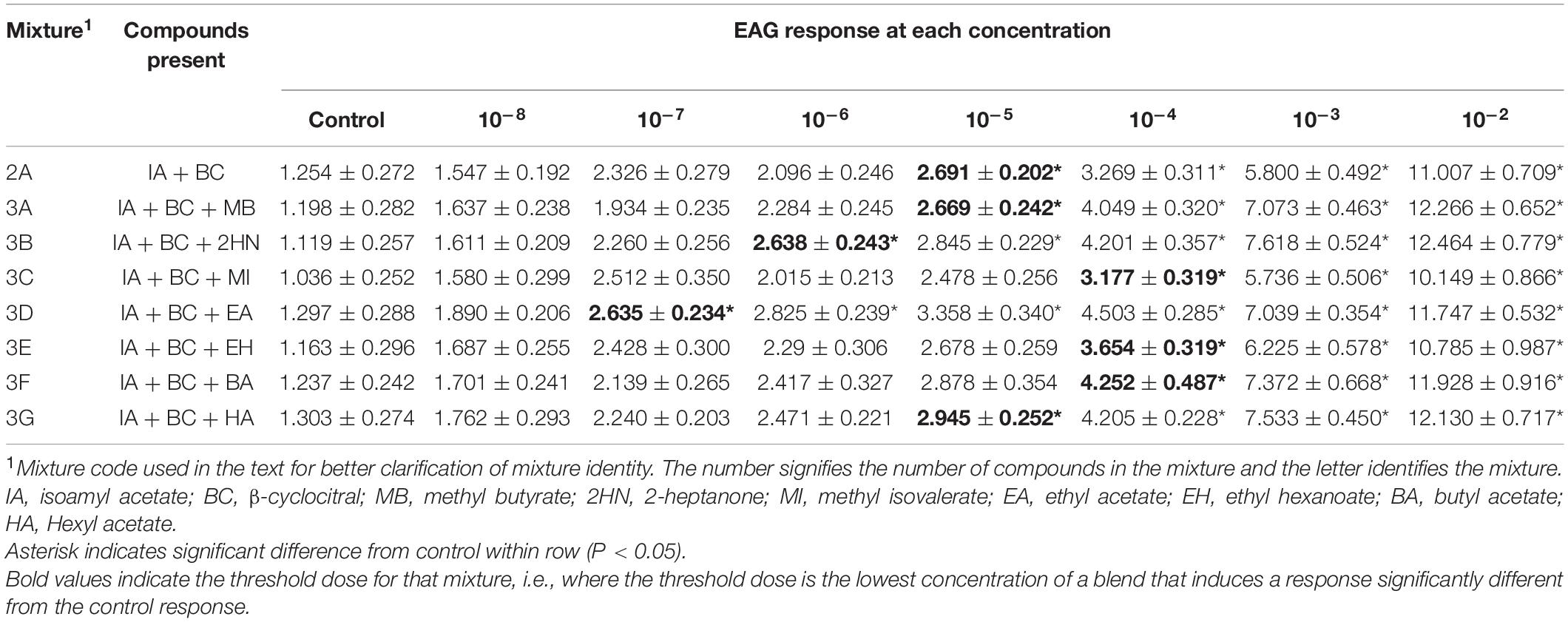
Table 1. Electroantennogram (EAG) responses (mean ± SEM) of female D. suzukii to different concentrations of mixtures with 2–3 compounds.
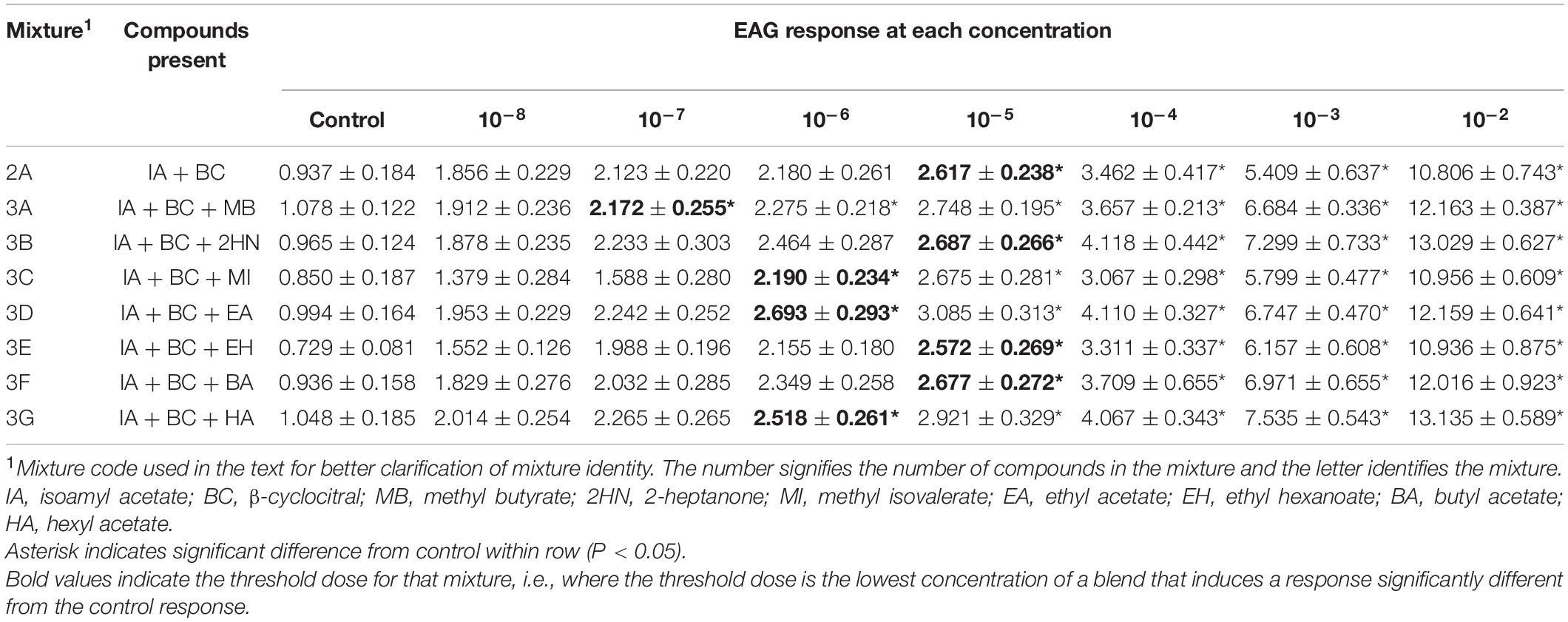
Table 2. Electroantennogram (EAG) responses (mean ± SEM) of male D. suzukii to different concentrations of mixtures with 2-3 compounds.
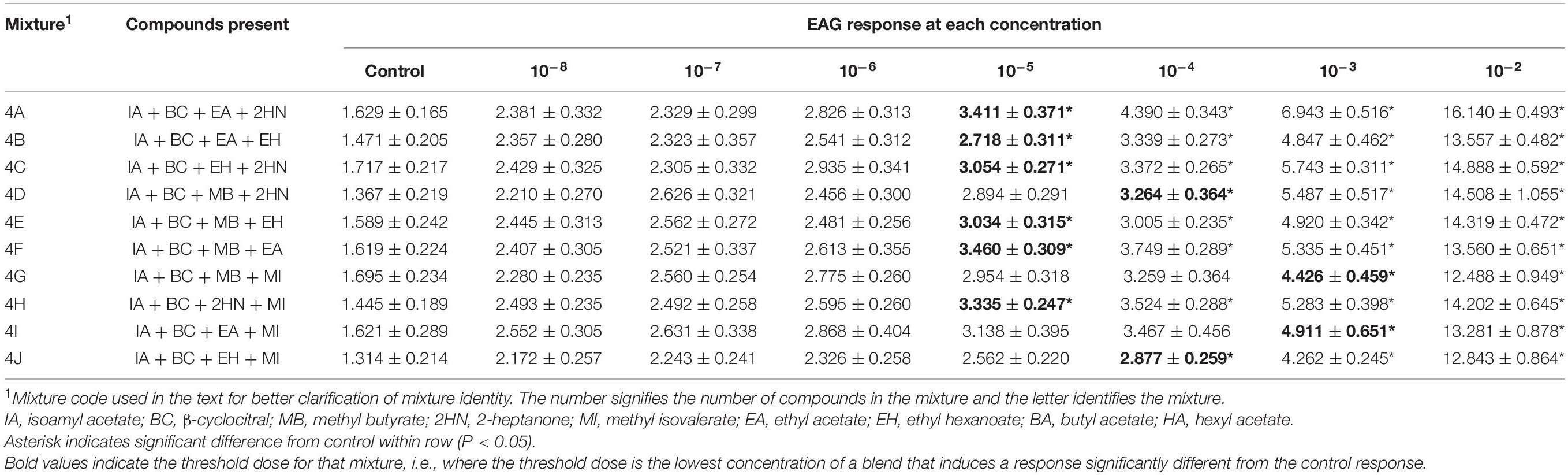
Table 3. Electroantennogram (EAG) responses (mean ± SEM) of female D. suzukii to different concentrations of mixtures with 4 compounds.
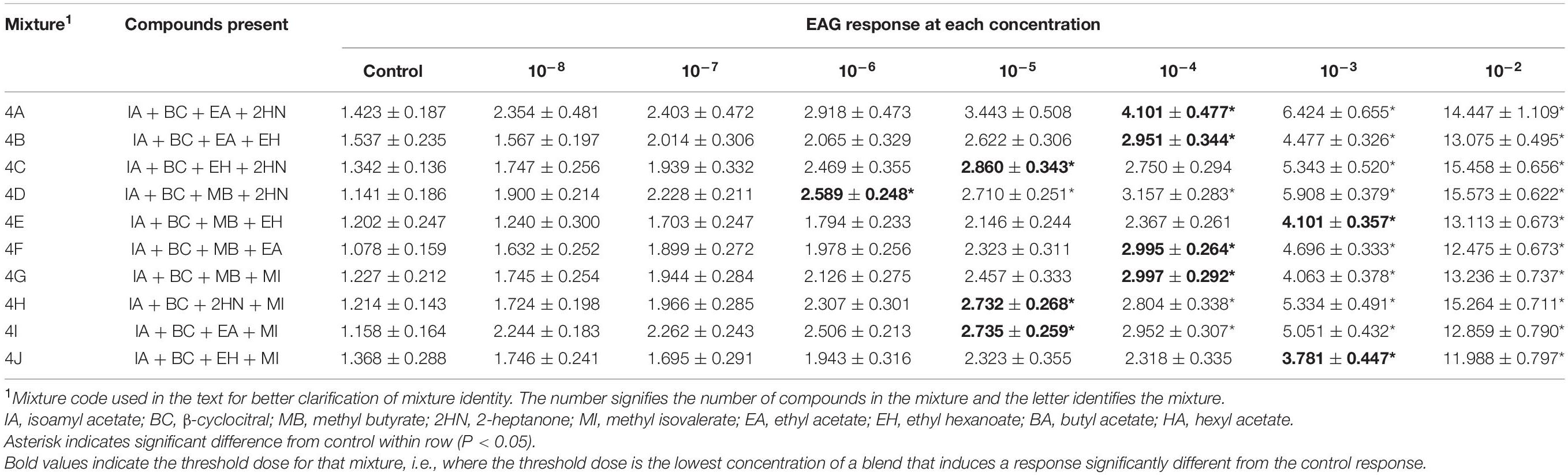
Table 4. Electroantennogram (EAG) responses (mean ± SEM) of male D. suzukii to different concentrations of mixtures with 4 compounds.
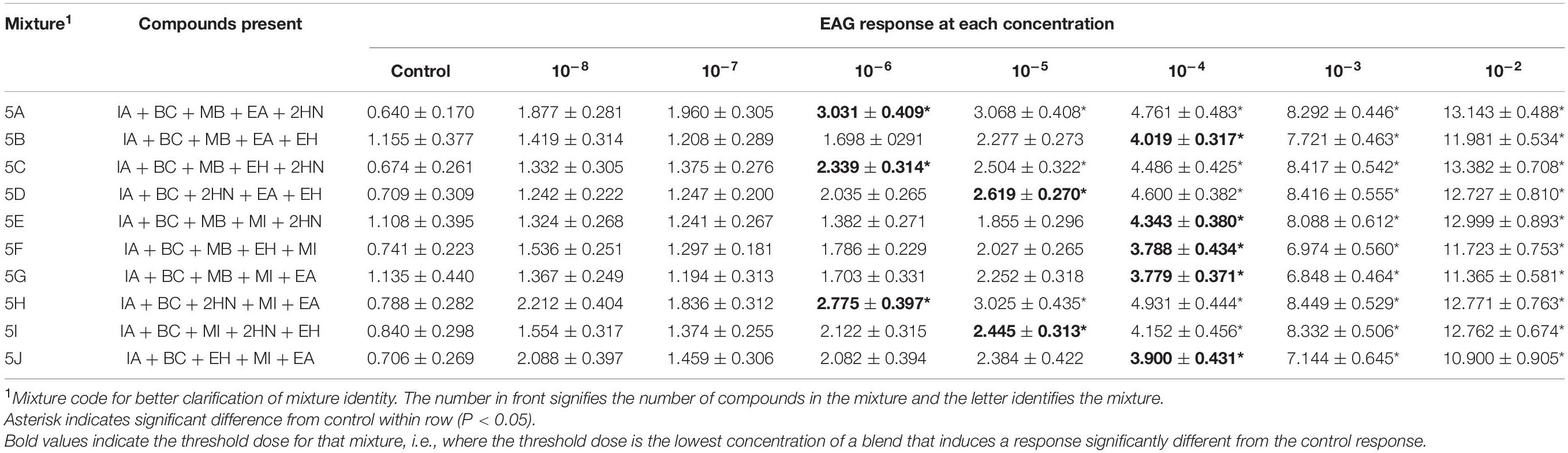
Table 5. Electroantennogram (EAG) responses (mean ± SEM) of female D. suzukii to different concentrations of mixtures with 5 compounds.
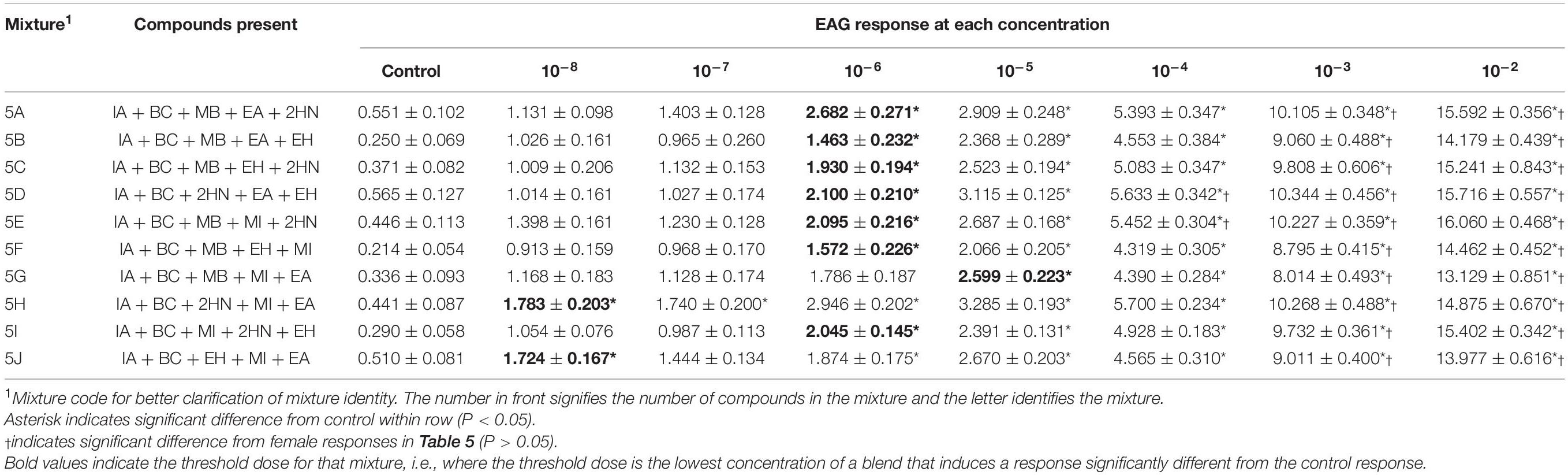
Table 6. Electroantennogram (EAG) responses (mean ± SEM) of male D. suzukii to different concentrations of mixtures with 5 compounds.
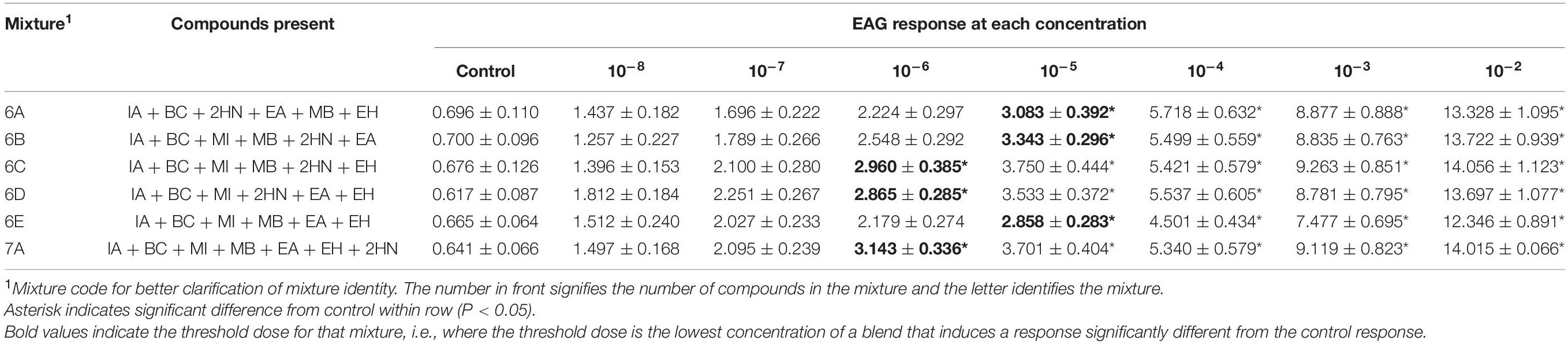
Table 7. Electroantennogram (EAG) responses (mean ± SEM) of female D. suzukii to different concentrations of mixtures with 6-7 compounds.
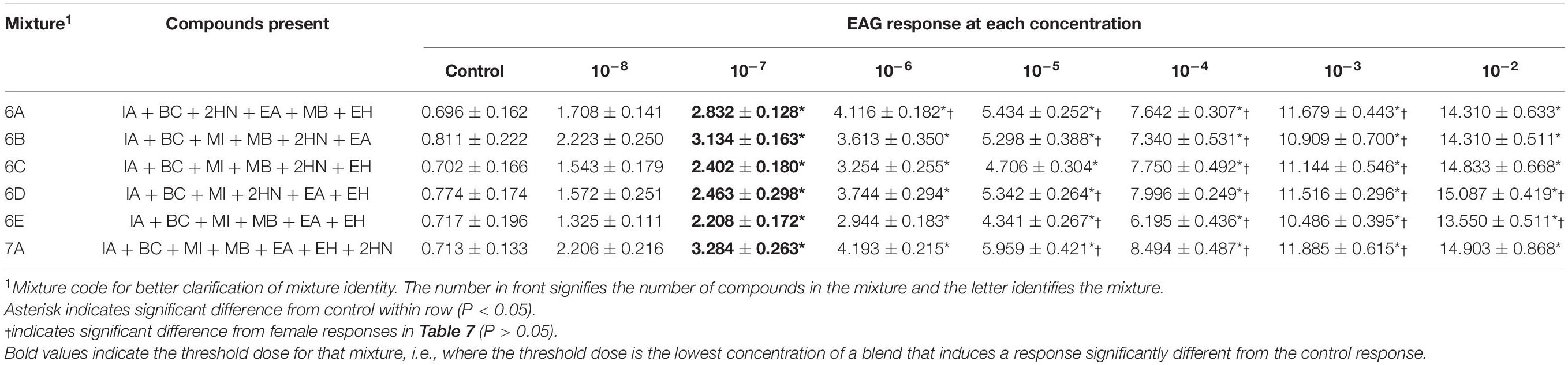
Table 8. Electroantennogram (EAG) responses (mean ± SEM) of male D. suzukii to different concentrations of mixtures with 6-7 compounds.
The EAG preparations consisted of an individual D. suzukii (7–10 days old) first secured in a micropipette tip (the end of the pipette tip was cut to expose the fly’s head). Glass capillary tubes were pulled to form glass probes, which were then filled with Drosophila Ringer’s solution. The probes were inserted over Ag-AgCl wires into micromanipulators (MP-15; Syntech, Hilversum, Netherlands). The first (or grounding) probe was inserted into the eye of the fly and the second (or recording) probe was then positioned on the surface of the anterior third segment of the fly’s antenna.
A stream of humidified and charcoal-purified air was constantly passed over the antennal preparation (0.4 liters/min) through a 15 mm diameter glass tube (constant air tube) that was proximal to the antennae. From each of the tested VOC mixtures, a single 10 μl drop of the diluted VOC sample stock was placed onto 1 cm2 filter paper disks, then placed into a borosilicate glass Pasteur pipette (puff cartridge) (15 cm length × 6 mm dia), with the tip of the cartridge inserted into the constant air tube. The insertion point of the puff cartridge was positioned 13 cm from the end of the constant air tube where the fly’s antennal preparation was stationed. These VOC puff cartridge preparations were freshly created for each new set of tested insect antennae. Control puffs consisted of air passed over a filter paper disk treated with a 10 μl droplet of mineral oil.
The EAG signals from the antennae were passed through a 2-channel signal acquisition interface controller (IDAC-2; Syntech, Hilversum, Netherlands) then recorded and processed using Syntech software (GC-EAD 2010). Both constant airflow and air puffs were generated with a stimulus flow controller (CS-55; Syntech, Hilversum, Netherlands). Three puffs of the same VOC treatment (one second in duration at 100 ml/min) were delivered 30 s apart for each stimulus and averaged, with one minute allowed between every new dilution and treatment compound. Seven dilutions of each treatment were tested in ascending order, ranging from the lowest (10–8) to the highest (10–2) concentrations. Before and after recording a mixture series, a control stimulus (mineral oil) and a standard stimulus of Z-3-hexen-1-ol were applied (Bolton et al., 2019). All amplitudes of the responses were measured as an absolute value.
Behavioral Bioassays With Drosophila suzukii
We sought to conduct a comprehensive assessment of male and female D. suzukii responses to simpler and more complex mixtures, and this necessitated the inclusion of a large number of treatments. The level of behavioral attraction that D. suzukii exhibited toward the various component VOC mixtures was evaluated in screened cages (33 cm3) (BugDorm-43030F, MegaView Science Co., Ltd., Taichung, Taiwan).
In each cage, a 5 ml polystyrene cup (33 mm diameter) (Globe Scientific Inc., Mahwah, NJ, United States, Item No. 3601) was placed in two diagonally opposing corners. One of the cups contained 200 μl of mineral oil (control) and the other cup contained 200 μl of one of the mixture treatments diluted in mineral oil to a 10–2 dose. The mineral oil facilitated capture and drowning of flies as they entered and explored the interior of each cup (Bolton et al., 2019). Twenty D. suzukii (same sex), 7–10 days old and anesthetized with CO2, were placed in a small petri dish and released into a cage after an hour of recovery. A moist paper towel was placed in the center of the cage as a water source for the flies. The cages were then placed in a desktop environmental chamber (interior dimensions: 67 × 76 × 43 cm) programmed for a 16L:8D photoperiod and constant temperature of 23°C (Percival, Model 130BLL, Perry, IA, United States). After 24 h, the cages were removed, and the number of flies captured in the control and treatment cups were counted and recorded. Both male and female flies were tested, and each VOC component blend mixture had six replicate cages per fly sex per dose.
Behavioral Bioassays With Drosophila melanogaster
Nineteen synthetic VOC component mixtures that were electrophysiologically sensitive to both male and female D. suzukii and that also induced a high fly recapture rate in the cage bioassays were used in cage studies with D. melanogaster. This experiment determined the relative selectivity of each selected VOC mixture blend [isoamyl acetate + β-cyclocitral (known as “base”), base + methyl butyrate, base + 2-heptanone, base + methyl isovalerate, base + ethyl acetate, base + ethyl hexanoate, base + ethyl acetate + methyl butyrate, base + methyl isovalerate + ethyl hexanoate, base + ethyl acetate + 2-heptanone, base + ethyl hexanoate + ethyl acetate + methyl butyrate, base + ethyl hexanoate + 2-heptanone + methyl butyrate, base + methyl isovalerate + ethyl acetate + methyl butyrate, base + methyl isovalerate + ethyl acetate + 2-heptanone, base + ethyl hexanoate + ethyl acetate + 2-heptanone, base + methyl isovalerate + ethyl hexanoate + ethyl acetate + 2-heptanone, base + methyl isovalerate + ethyl acetate + 2-heptanone + methyl butyrate, base + methyl isovalerate + ethyl hexanoate + 2-heptanone + methyl butyrate, base + methyl isovalerate + ethyl hexanoate + ethyl acetate + methyl butyrate, base + methyl isovalerate + ethyl hexanoate + ethyl acetate + 2-heptanone + methyl butyrate], with D. melanogaster acting as a model non-target species. The experimental design for the cage bioassays was the same as described above for D. suzukii (except that only 19 selected VOC mixtures were used).
Comparison of Selected Mixtures Against Blueberry Volatiles
Lures used for trapping often have to compete against ripening fruit volatiles in fields and orchards, and less attractive lures and traps that don’t effectively capture or kill D. suzukii can result in a spillover effect, leading to an increase in fruit infestation (Hampton et al., 2014; Alnajjar et al., 2017). To determine whether the selected VOC component blends were more attractive than a common fruit host, the four most promising blends from the cage bioassays that were attractive to D. suzukii but not to D. melanogaster, were compared against blueberry fruit with a moderate level of attractiveness to D. suzukii (Abraham et al., 2015; Urbaneja-Bernat et al., 2021). Blueberry was used due to its availability, the space limitations of the trap cups, and the need to avoid cutting fruit due to the emission of induced volatiles. Thirty to 40 D. suzukii flies (sex ratio 1:1) were prepared in the same way as previously described. Trap cups were also prepared in a similar manner, with the VOC mixture being placed in one trap and one uncut blueberry (1.5–2.0 g) placed in another cup with 200 μL of mineral oil underneath the blueberry. Other elements of the experimental design were identical to the previously described behavioral assays.
Data Analysis
The EAG treatments were arranged as a 2 × 8 factorial (2 sexes and 7 doses + control) in which the main plot effect was “sex” and the sub-plot effect was “dose” and “sex” by “dose.” EAG responses (amplitudes) were analyzed with a split plot analysis of variance (ANOVA) using SAS PROC GLIMMIX (SAS 9.4, SAS Institute Inc., 2017 Cary, NC, United States) as described in Littell et al. (1998). A Dunnett’s post hoc test was used to assess mean differences from controls. The EAG responses that are statistically distinguishable from the control indicated a threshold dose. Threshold dose is defined as the lowest concentration of a blend that induces a response significantly different from control responses (Raguso et al., 1996). Behavioral assay data (x + 0.01 to account for zero values) were prepared using Logit Link, a binomial distribution and a completely randomized design. Data were analyzed with ANOVA using SAS PROC GLIMMIX and the model using a ratio of T/T + C, where T is the number of flies that chose the treatment and C is the number of flies that chose the control.
Results
Electrophysiological Responses
2-3 Compound Mixtures
Electroantennogram (EAG) responses of all 2- and 3-component blends increased as concentration increased. However, there was no interaction between sex and concentration for any of the mixtures [ANOVA, Mixture 2A, F(1,7) = 0.20, P = 0.9856; Mixture 3A, F(1,7) = 0.32, P = 0.9427; Mixture 3B, F(1,7) = 0.24, P = 0.9742; Mixture 3C, F(1,7) = 0.76, P = 0.6245; Mixture 3D, F(1,7) = 0.32, P = 0.9432; Mixture 3E, F(1,7) = 0.10, P = 0.9981; Mixture 3F, F(1,7) = 0.12, P = 0.9970; Mixture 3G, F(1,7) = 0.55, P = 0.7918]. Female EAG responses were detectable at a minimum threshold at the 10–4 dose (Table 1), except for mixtures 2A, 3A, and 3G which were detected at 10–5, and 3B and 3D which were detected at10–6 and 10–7 concentrations, respectively. Overall, males were more sensitive to the blends (i.e., responded to lower threshold doses) than the females. All male EAG responses at the 10–5 dose were significantly different from the control (Table 2). Three mixtures had a male sensitivity threshold at the 10–6 dose (Mixtures 3C, 3D, and 3G) and one mixture was first detected by males at the 10–7 dose (Mixtures 3A).
4-Compound Mixtures
The EAG responses to the ten 4-compound mixtures that were evaluated increased significantly with increased concentration for both sexes, although there was no interaction between sex and concentration for any of the mixtures [Mixture 4A, F(1,7) = 0.72, P = 0.6589; Mixture 4B, F(1,7) = 0.29, P = 0.9583; Mixture 4C, F(1,7) = 0.55, P = 0.7928; Mixture 4D, F(1,7) = 0.71, P = 0.6628; Mixture 4E, F(1,7) = 0.35, P = 0.9292; Mixture 4F, F(1,7) = 0.18, P = 0.9896; Mixture 4G, F(1,7) = 0.60, P = 0.7571; Mixture 4H, F(1,7) = 1.36, P = 0.2276; Mixture 4I, F(1,7) = 0.10, P = 0.9980; Mixture 4J, F(1,7) = 0.22, P = 0.9809].
The threshold dose for female EAG responses was the 10–4 dose for mixtures 4D, 4G, 4I, 4J, and the 10–5 dose for mixtures 4A, 4B, 4C, 4E, 4F, and 4H (Table 3). Male responses (Table 4) showed similar results at a 10–4 threshold (Mixtures 4A, 4B, 4F, and 4G) or 10–5 (4C, 4H, and 4I). Mixture 4D was different from the control at the 10–6 dose and mixture 4J was first detected at 10–3.
5-Compound Mixtures
For all ten mixtures comprising five compounds, there were interactions between sex and the concentration of the mixture [Mixture 5A, F(1,7) = 5.97, P < 0.001; Mixture 5B, F(1,7) = 4.40, P = 0.0002; Mixture 5C, F(1,7) = 2.21, P = 0.0369; Mixture 5D, F(1,7) = 5.38, P < 0.001; Mixture 5E, F(1,7) = 4.99, P < 0.001; Mixture 5F, F(1,7) = 6.10, P < 0.001; Mixture 5G, F(1,7) = 2.15, P < 0.001; Mixture 5H, F(1,7) = 2.79, P = 0.0095; Mixture 4I, F(1,7) = 5.96, P < 0.001; Mixture 5J, F(1,7) = 4.20, P = 0.0003]. Males and females responded differently depending on the concentration level of the mixture. For example, male responses at the two highest doses (10–3 and 10–2) for all 5-component mixtures were significantly higher than female responses at those doses (Table 6). Additionally, male responses at 10–4 doses for mixtures 5D and 5E were higher than female responses. In general, males detected mixtures at lower concentrations (Table 6) than females (Table 5). Males had a threshold dose between 10–6 and 10–5 (Table 6), with two mixtures being detected at the lowest concentration (Mixture 5H and 5J). Females were less sensitive than the males to these mixtures, with five mixtures being first detected at the 10–4 concentration (5B, 5E, 5F, 5G, and 5J) two mixtures detected at 10–5 (5D and 5I) and three mixtures detected at 10–6 (5A, 5C, and 5H).
6-7 Compound Mixtures
Only three mixtures with six compounds present and the compound with seven compounds present had interactions between sex and concentration of the mixtures [Mixture 6A, F(1,7) = 2.16, P = 0.0411; Mixture 6D, F(1,7) = 3.11, P = 0.0043; Mixture 6E, F(1,7) = 3.53, P = 0.0016; Mixture 7A, F(1,7) = 2.19, P = 0.0387]. The remaining mixtures had no interaction [Mixture 6B, F(1,7) = 1.10, P = 0.3681; Mixture 6C, F(1,7) = 1.44, P = 0.1956]. Female responses to Mixture 6A (Table 7) were significantly lower than male responses at the 10–3, 10–4, 10–5, and 10–6 doses (Table 8). Male responses to mixtures 6D and 6E were different from females at 10–2, 10–3, 10–4, and 10–5 doses. Male responses to mixture 7A were only significantly different from females at 10–3, 10–4, and 10–5 doses. Males detected mixtures at the 10–7 threshold dose (Table 8) and were more sensitive to these mixtures than females (Table 7).
Behavioral Responses of Drosophila suzukii
2-3 Compound Mixtures
While the overall main effects for treatment and sex were insignificant [F(8,90) = 0.46 P = 0.8810; F(1,90) = 0.32 P = 0.5715], there was an interaction between the two [F(8,90) = 2.98 P = 0.0052]. The 2-component mixture of isoamyl acetate + β-cyclocitral (the base combination for all 3-component VOC treatments) captured significantly more male and female D. suzukii than the competing control treatment (base, t = 3.02, d.f. = 90, P = 0.0033; t = 2.46, d.f. = 90, P = 0.0160). In fact, all of the 3-component mixtures captured significantly more male flies than the control treatments (base + methyl butyrate, t = 2.85, d.f. = 90, P = 0.0055; base + 2-heptanone, t = 3.23, d.f. = 90, P = 0.0017; base + methyl isovalerate, t = 3.78, d.f. = 90, P = 0.0003; base + ethyl acetate, t = 2.02, d.f. = 90, P = 0.0461; base + ethyl hexanoate, t = 3.25, d.f. = 90, P = 0.0016; base + butyl acetate, t = 4.52, d.f. = 90, P < 0.0001; base + hexyl acetate, t = 3.83, d.f. = 90, P = 0.0002). The component mixture containing butyl acetate attracted the greatest mean number of male D. suzukii (Figure 1). For female D. suzukii, all of the 3-component mixtures (base + methyl butyrate, t = 4.62, d.f. = 90, P < 0.0001; base + 2-heptanone, t = 3.91, d.f. = 90, P = 0.0002; base + methyl isovalerate, t = 3.85, d.f. = 90, P = 0.0002; base + ethyl acetate, t = 4.80, d.f. = 90, P < 0.0001; base + ethyl hexanoate, t = 4.63, d.f. = 90, P < 0.0001), except those containing butyl acetate and hexyl acetate (base + butyl acetate, t = −1.26, d.f. = 90, P = 0.2125; base + hexyl acetate, t = 1.16, d.f. = 90, P = 0.24803), attracted significantly more females that the competing control treatments.
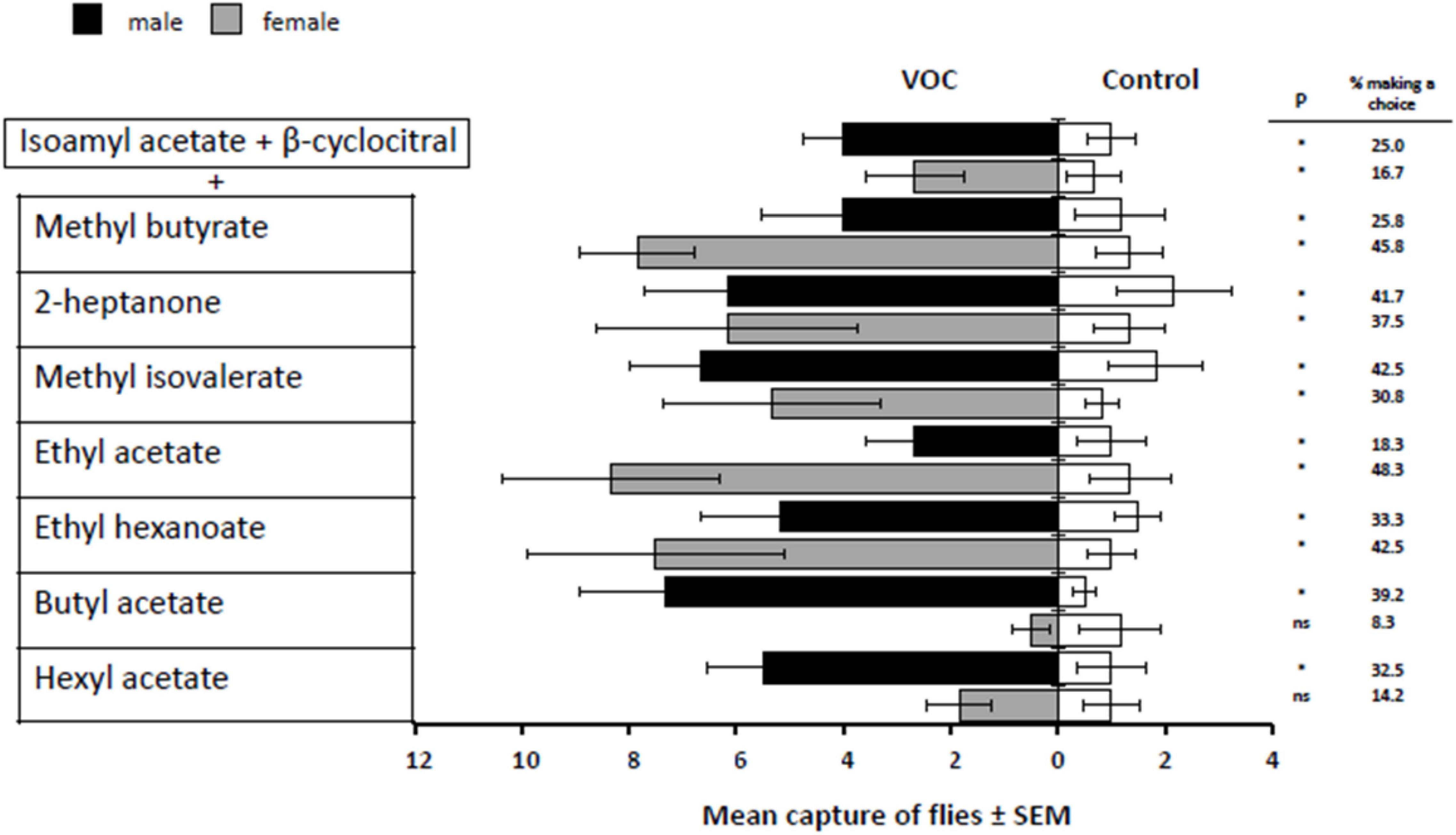
Figure 1. Response of male and female D. suzukii to mixtures of one fruit volatile added to isoamyl acetate and β-cyclocitral (VOC) against the mineral oil solvent (Control). P-values with * indicate P < 0.05 and “ns” indicates a non-significant difference where P > 0.05.
The 3-component mixture containing the base + ethyl acetate captured the greatest mean number of females (Figure 1). Interestingly, the mean female responses to the methyl butyrate-containing blend were almost double to that of the male responses, and the female responses to base + ethyl acetate was nearly three times greater than the male responses. Conversely, the responses to the base + 2-heptanone, methyl isovalerate and ethyl hexanoate treatments were similar for both sexes. Overall, the greatest increase in mean female captures involving the base 2-component mixture (isoamyl acetate + β-cyclocitral) occurred with the addition of either methyl butyrate, ethyl acetate or ethyl hexanoate. The addition of one of those components resulted in nearly a threefold increase in female captures (Figure 1).
4-Compound Mixtures
Adding two additional volatiles to the base mixture (isoamyl acetate + β-cyclocitral) captured significantly more male and female D. suzukii than the competing control treatments [F(9,100) = 2.22, P = 0.0263]. The pair of compounds added to the 2-component base mixture that increased captures of both males and females in the capturing devices were the (1) ethyl acetate + 2-heptanone (male, t = 3.30, d.f. = 100, P = 0.0013; female, t = 3.55, d.f. = 100, P = 0.0006), (2) ethyl acetate + methyl butyrate (male, t = 2.67, d.f. = 100, P = 0.0089; female, t = 3.12, d.f. = 100, P = 0.0023), and (3) methyl isovalerate + ethyl hexanoate treatments (male, t = 4.28, d.f. = 100, P < 0.0001; female, t = 4.05, d.f. = 100, P = 0.0001) (Figure 2). The treatments that attracted significantly more males, but not females, over the control treatments were (1) ethyl hexanoate + 2-heptanone (male, t = 2.10, d.f. = 100, P = 0.0382; female, t = 1.16, d.f. = 100, P = 0.2477), (2) 2-heptanone + methyl butyrate (male, t = 2.85, d.f. = 100, P = 0.0053; female, t = 0.49, d.f. = 100, P = 0.6243), and (3) methyl isovalerate + ethyl acetate treatments (male, t = 3.58, d.f. = 100, P = 0.0005; female, t = 1.44, d.f. = 100, P = 0.1519). The treatments that attracted significantly more females, but not males, over the control treatments were (1) ethyl acetate + ethyl hexanoate (male, t = 1.80, d.f. = 100, P = 0.0755; female, t = 2.08, d.f. = 100, P = 0.0401) and (2) methyl isovalerate + 2-heptanone (male, t = 1.05, d.f. = 100, P = 0.2953; female, t = 2.12, d.f. = 100, P = 0.0365). The two treatments that did not reveal a significant difference between male and female captures from the controls were (1) ethyl hexanoate + methyl butyrate (male, t = 1.79, d.f. = 100, P = 0.0772; female, t = 0.95, d.f. = 100, P = 0.3461) and (2) methyl isovalerate + methyl butyrate (male, t = 0.89, d.f. = 100, P = 0.3757; female, t = 1.89, d.f. = 100, P = 0.0611).
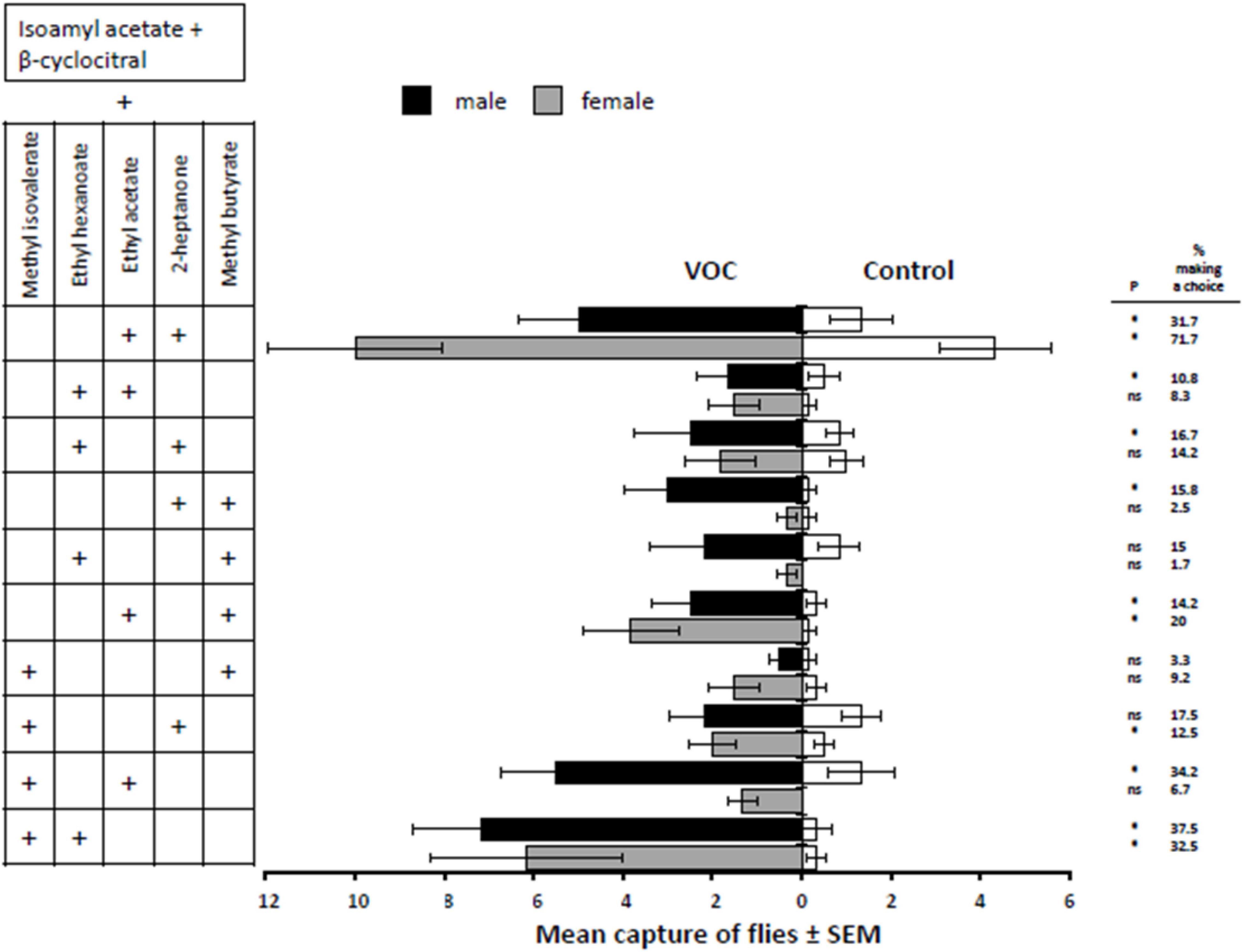
Figure 2. Response of male and female D. suzukii to mixtures of two fruit volatiles added to isoamyl acetate and β-cyclocitral (VOC) against the mineral oil solvent (Control). P-values with * indicate P < 0.05 and “ns” indicates a non-significant difference where P > 0.05.
The treatment that captured the greatest mean number of female D. suzukii was the base + ethyl acetate + 2-heptanone (with nearly 72% making a choice), and the most attractive treatment for the males was the base + methyl isovalerate + ethyl hexanoate (with nearly 38% of the males making a choice) (Figure 2).
5-Compound Mixtures
The majority of the 5-component mixtures (all having the 2-component base mixture of isoamyl acetate + β-cyclocitral) were more attractive to both male and female flies than the respective controls [F(9,100) = 2.48, P = 0.0135]. Of the ten treatments, only two were not attractive to males: (1) base + methyl butyrate + ethyl acetate + 2-heptanone (male, t = 0.75, d.f. = 100, P = 0.4566; female, t = 5.40, d.f. = 100, P < 0.001), and (2) base + methyl isovalerate + methyl butyrate + 2-heptanone (male, t = 1.34, d.f. = 100, P = 0.1844; female, t = 4.17, d.f. = 100, P < 0.0001) (Figure 3). The two most female-attractive treatments were (1) base + ethyl acetate + 2-heptanone + methyl butyrate (Figure 3) and (2) base + methyl isovalerate + ethyl acetate + methyl butyrate (male, t = 3.62, d.f. = 100, P = 0.0005; female, t = 4.60, d.f. = 100, P < 0.0001). Two treatments, (1) base + ethyl hexanoate + ethyl acetate + methyl butyrate (male, t = 4.39, d.f. = 100, P < 0.0001; female, t = 3.95, d.f. = 100, P = 0.0001) and (2) base + methyl isovalerate + ethyl hexanoate + methyl butyrate (male, t = 4.55, d.f. = 100, P < 0.0001; female, t = 3.23, d.f. = 100, P = 0.0017), were the most attractive to males, both having about 40% of the males responding and making a choice (Figure 3). The remaining mixtures that were attractive to males and females were (1) base + 2-heptanone + methyl butyrate + ethyl hexanoate (male, t = 3.14, d.f. = 100, P = 0.0022; female, t = 4.33, d.f. = 100, P < 0.0001), (2) base + 2-heptanone + ethyl acetate + ethyl hexanoate (male, t = 3.96, d.f. = 100, P = 0.0001; female, t = 4.48, d.f. = 100, P < 0.0001), (3) base + methyl isovalerate + 2-heptanone + ethyl acetate (male, t = 3.83, d.f. = 100, P = 0.0002; female, t = 4.26, d.f. = 100, P < 0.0001), (4) base + methyl isovalerate + 2-heptanone + ethyl hexanoate (male, t = 3.31, d.f. = 100, P = 0.0013; female, t = 2.12, d.f. = 100, P = 0.0365), and (5) base + methyl isovalerate + ethyl acetate + ethyl hexanoate treatments (male, t = 4.01, d.f. = 100, P = 0.0001; female, t = 3.68, d.f. = 100, P = 0.0004).
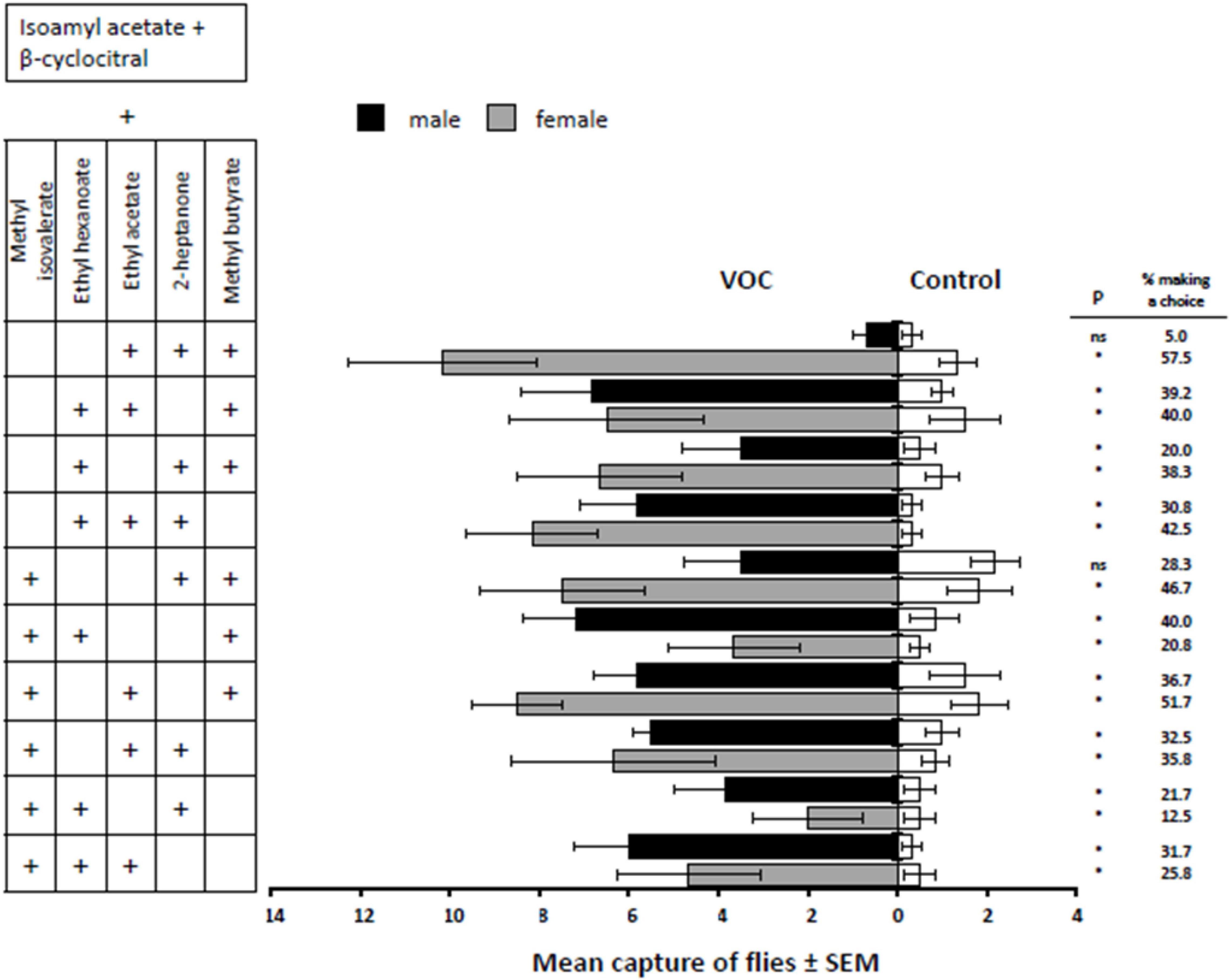
Figure 3. Response of male and female D. suzukii to mixtures of three fruit volatiles added to isoamyl acetate and β-cyclocitral (VOC) against the mineral oil solvent (Control). P-values with * indicate P < 0.05 and “ns” indicates a non-significant difference where P > 0.05.
6-7 Compounds
While the overall main effects for treatment and sex for the five 6-compound mixtures and single 7-compound mixture were insignificant [F(5,60) = 1.24, P = 0.3016 F(1,60) = 0.01 P = 0.9043], there was an interaction between the two [F(5,60) = 3.29, P = 0.0163]. The two treatments that were the most attractive to female D. suzukii were (1) methyl isovalerate + ethyl hexanoate + ethyl acetate + methyl butyrate (t = 4.60, d.f. = 50, P < 0.0001) and (2) methyl isovalerate + ethyl hexanoate + 2-heptanone + methyl butyrate (t = 4.53, d.f. = 50, P < 0.0001), both treatments having close to a 71 and 51% response, respectively, of the released females (Figure 4). The treatment most attractive to males was also methyl isovalerate + ethyl hexanoate + ethyl acetate + methyl butyrate (t = 4.39, d.f. = 50, P < 0.0001). It is interesting to note that when methyl isovalerate was removed from the blend of volatiles, the percentage of female response dropped from 71 to 10% capture. No such drastic a reduction in male captures was evident with the removal of any compound (Figure 4). Additionally, when 2-heptanone was removed from the 7-component blend, the percentage of male and female flies making a choice nearly doubled.
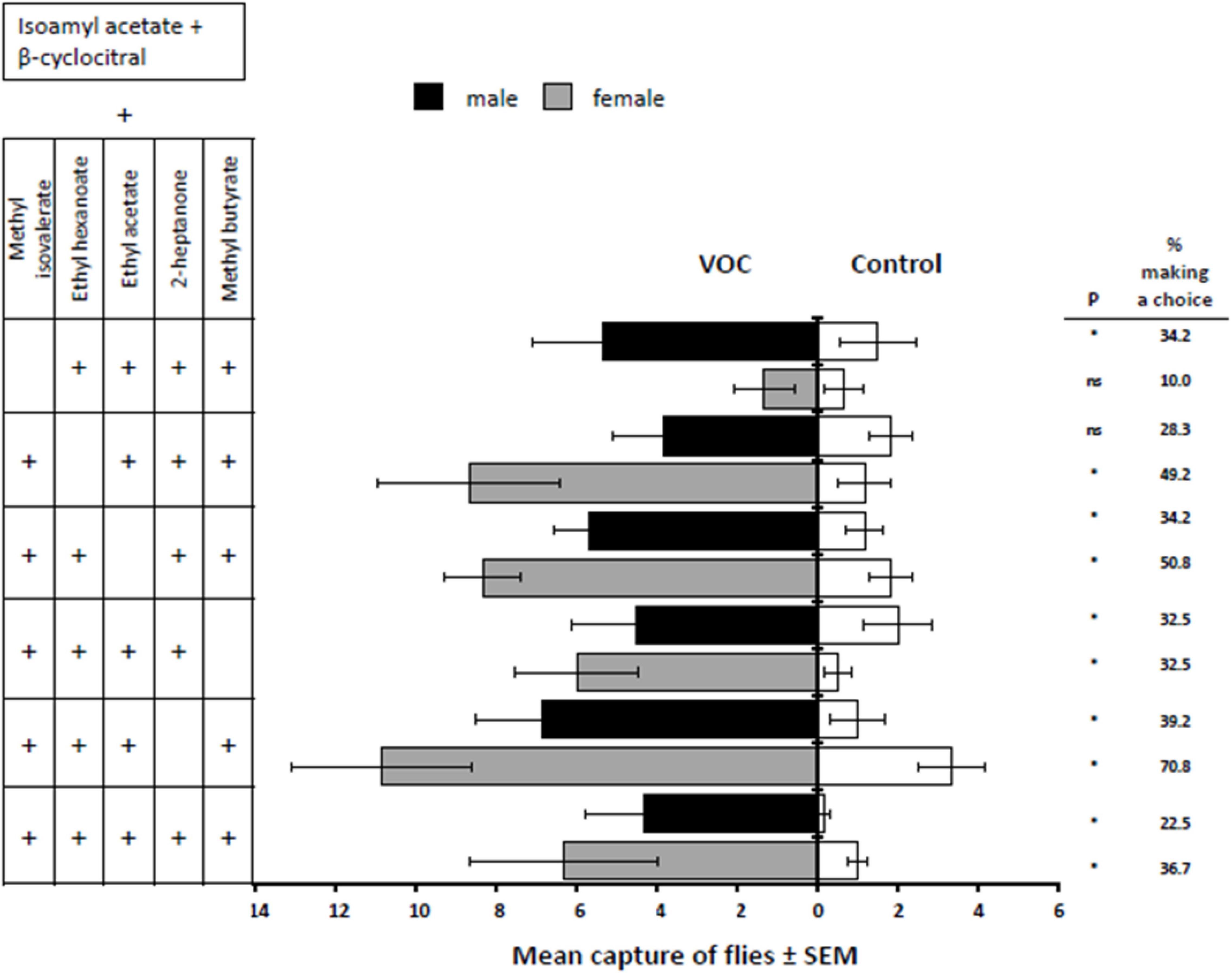
Figure 4. Response of male and female D. suzukii to all selected fruit volatiles added to isoamyl acetate and β-cyclocitral (VOC) against the mineral oil solvent (Control) and a subtraction strategy where one volatile from the entire blend was removed. P-values with * indicate P < 0.05 and “ns” indicates a non-significant difference where P > 0.05.
Behavioral Responses of Drosophila melanogaster
Of the nineteen mixtures that were evaluated with D. melanogaster six mixtures were found to not be attractive to either sex (Figure 5). For example, isoamyl acetate + β-cyclocitral alone (the base combination for all 3–7 component VOC treatments) was not significantly attractive to either sex (female, t = 0.69, P = 0.4941; male, t = 1.73, P = 0.0851). Such a lack of significant attraction still occurred despite the addition of methyl butyrate (female, t = 1.35, P = 0.1804; male, t = 1.83 P = 0.0693), 2-heptanone (female, t = 1.10, P = 0.2751; male, t = 1.47, P = 0.1438) and methyl isovalerate (female, t = 0.80, P = 0.4247; male, t = 1.96, P = 0.0517). These three mixtures were attractive to both male and female D. suzukii (Figure 1) and had higher responses to these volatiles than D. melanogaster (Figure 5). The 5-compound mixture consisting of base + methyl isovalerate + ethyl acetate + methyl butyrate was not significantly more attractive to male and female D. melanogaster than the solvent control (female, t = 1.19, P = 0.2352; male, t = 1.52, P = 0.1292); however, this 5-component mixture was highly attractive to both sexes of D. suzukii (Figure 3). Finally, a 6-compound mixture that lacked 2-heptanone was not attractive to either sex of D. melanogaster (female, t = 1.20, P = 0.2328; male, t = 1.19, P = 0.2352), but was attractive to male and female D. suzukii, especially with females, with 70.8% making a choice (Figure 4).
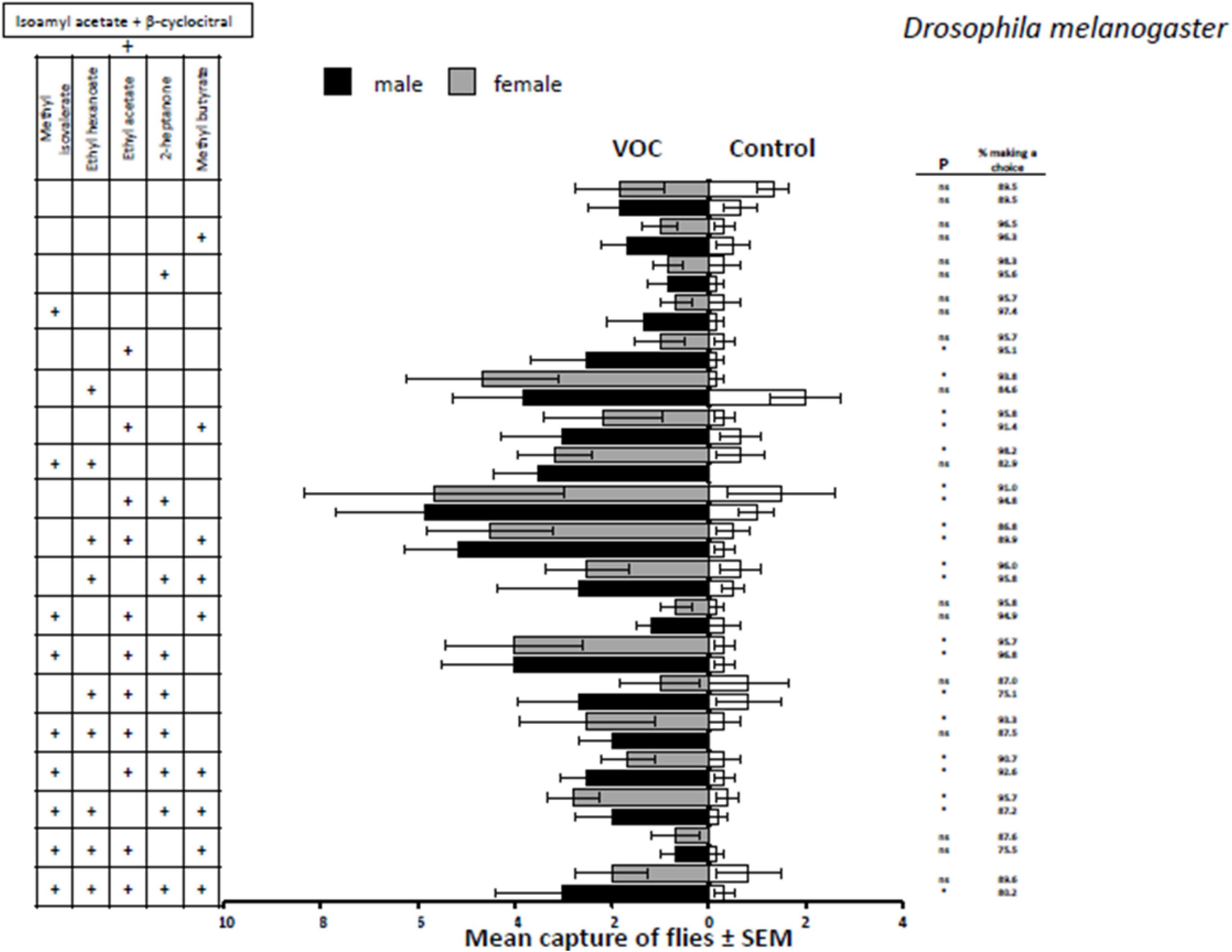
Figure 5. Response of male and female D. melanogaster to selected mixtures of fruit volatiles added to isoamyl acetate and β-cyclocitral (VOC) against the mineral oil solvent (Control). P-values with * indicate P < 0.05.
Comparison of Selected Mixtures Against Blueberry Volatiles
Four mixtures (two 3-component blends, one 5- and one 6-component blend) that were attractive to D. suzukii but unattractive to D. melanogaster were compared against volatiles emitted by a single blueberry (Figure 6). Of those four mixtures, only the 3-component mixture with methyl butyrate (added to the 2-component base mixture of isoamyl acetate + β-cyclocitral) was significantly more attractive to females than the blueberry fruit (t = −2.42, P = 0.0173); however, the males were not. The levels of attractiveness of the remaining VOC mixtures toward D. suzukii were not significantly different from that of the blueberry fruit treatments, indicating an adequate level of D. suzukii response to the synthetic mixtures.
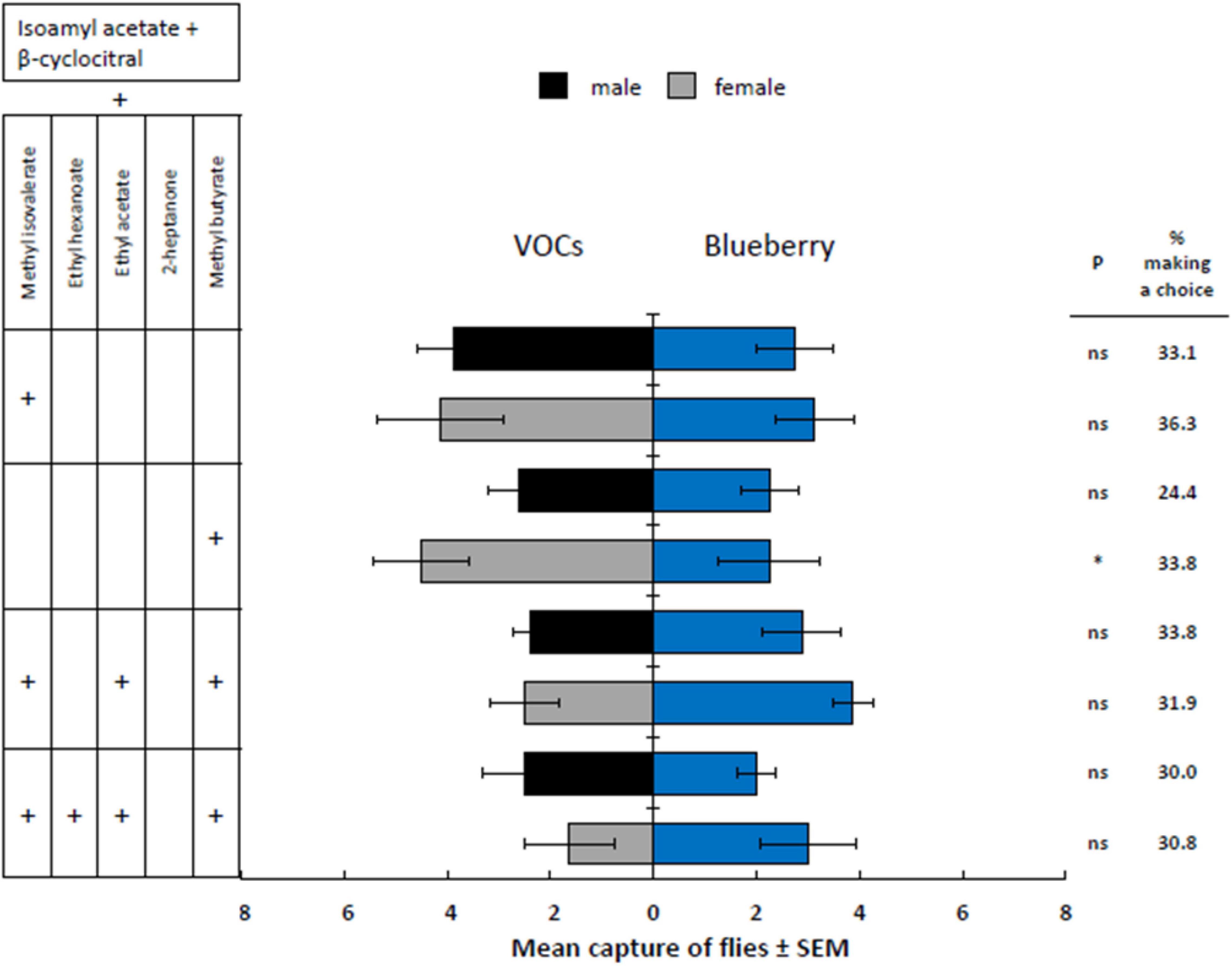
Figure 6. Response of male and female D. suzukii to mixtures of fruit volatiles added to isoamyl acetate and β-cyclocitral (VOCs) against a whole blueberry (Blueberry). P-values with * indicate P < 0.05 and “ns” indicates a non-significant difference where P > 0.05.
Discussion
This study has demonstrated that a simple, high concentration blend of isoamyl acetate + β-cyclocitral + methyl butyrate (in equal proportions) is (1) attractive to both sexes of D. suzukii, (2) not attractive to the non-target species D. melanogaster, and (3) is more attractive to females than blueberry fruit. Moreover, methyl isovalerate, methyl butyrate, and ethyl acetate in various combinations with isoamyl acetate + β-cyclocitral were attractive to D. suzukii but not to D. melanogaster.
Our EAG results indicated that as the complexity of the mixtures increased, D. suzukii ability to detect the volatiles at lower concentrations increased. Overall, males were able to detect mixtures with 5–7 components at lower concentrations than females, and male EAG responses were typically higher than those recorded in females. Generally, antennal sensitivity did not correlate with behavioral activity; however, a 3-component mixture of isoamyl acetate + β-cyclocitral + ethyl acetate was detected by females at the lowest threshold of all 3-component mixtures, and females were more sensitive to this mixture than males, both electrophysiologically and behaviorally. Similar results were obtained for males exposed to a 4-component mixture (isoamyl acetate + β-cyclocitral + 2-heptanone + methyl butyrate). However, this pattern was not true for all mixtures that produced significant electrophysiological results for males or females. What is driving the difference between male and female responses is still unclear. D. suzukii has been shown to be attracted to a synthetic blend of volatiles from raspberry headspace in laboratory assays (Abraham et al., 2015) and in cage bioassays, males responded more to the synthetic blend than females. Males were more electrophysiologically and behaviorally sensitive to the fruit volatile benzyl acetate than females.
The individual addition of methyl butyrate, 2-heptanone, ethyl acetate, and ethyl hexanoate to the base component mixture of isoamyl acetate + β-cyclocitral enhanced the attractiveness to females. Males, on the other hand, were not as attracted to base + ethyl acetate, but were attracted to the addition of butyl acetate and hexyl acetate. This data supports the synergistic effects that β-cyclocitral has on several fruit volatiles, specifically on ethyl acetate and ethyl hexanoate (Bolton et al., 2019). Previously, Piñero et al. (2019) documented synergistic interactions between β-cyclocitral and isoamyl acetate in male but not female D. suzukii. Although 2-heptanone was considered antagonistic to D. suzukii attraction (Bolton et al., 2019), the addition of other fruit volatiles negated those effects in this study. However, when 2-heptanone was removed from the 7-component blend, there was a higher numerical response from females.
As the blend of volatiles increased in complexity, there was an increase in attraction to the 5-, 6-, and 7-compound blends. Despite being more attractive than a mineral oil control, the more complex mixtures were not more attractive than the volatiles emanating from whole blueberry. For example, the simple blend of isoamyl acetate + β-cyclocitral + methyl butyrate was more attractive than blueberry to D. suzukii but not to the non-target D. melanogaster. This combination of volatiles may be promising for field trapping and monitoring. For example, a field study that evaluated the effect of adding methyl butyrate at high concentration over a yeast/sugar drowning solution caught more D. suzukii and less non-targets than just the yeast/sugar drowning solution alone (G. Bolton unpublished data). The same study evaluated a high concentration of isoamyl acetate, which had similar results to methyl butyrate. These two compounds are also present in the blush red phase of strawberry and D. suzukii responds earlier electrophysiologically to these compounds than D. melanogaster or D. biarmipes Malloch (Keesey et al., 2015). Therefore, the combination of these two compounds may play an important role in the insect’s ability to locate ripening fruit.
Drosophila suzukii responds to the volatile profiles of many fruit hosts (Burrack et al., 2013) and a species-specific blend or ratio may not be as important in host location for D. suzukii as it is for other pest species (Bruce et al., 2005; Bruce and Pickett, 2011; Abraham et al., 2015; Revadi et al., 2015). Previous research investigating volatiles from several host fruits has led to a wide range of promising compounds attractive to D. suzukii (Abraham et al., 2015; Feng et al., 2018; Dewitte et al., 2021; Urbaneja-Bernat et al., 2021), although there is little volatile overlap between these suggested blends (Cloonan et al., 2018). For example, isoamyl acetate was an attractive component of the volatile profile of wild blueberries (Urbaneja-Bernat et al., 2021), but wasn’t in blackberry (Dewitte et al., 2021), apple (Feng et al., 2018), or raspberry (Abraham et al., 2015). Our results demonstrate that several mixtures and blends of high concentrations of overlapping fruit volatiles including isoamyl acetate, in addition to β-cyclocitral, are attractive to D. suzukii. Such findings support the notion that phytophagous insects rely on commonly distributed plant volatiles blends, despite a probable redundancy in the mixture (Bruce and Pickett, 2011). It is conceivable that overlapping fruit volatiles may extend the host range of an insect and thus promote polyphagy (Lance, 1983). A “super-blend” of overlapping volatiles would not rely on using VOCs from one particular fruit, but instead, use commonly attractive “host” VOCs and certain combinations that the insect is sensitive to (Del Socorro et al., 2010).
Overall, the simplest blend that was most attractive to females in this study was isoamyl acetate + β-cyclocitral + ethyl hexanoate + 2-heptanone, although this blend was also highly attractive to our non-target model, D. melanogaster. Therefore, a simple blend like isoamyl acetate + β-cyclocitral + methyl butyrate, that was attractive to female D. suzukii, not attractive to D. melanogaster, and more attractive than whole blueberry fruit, needs further investigation. Future work is also needed to determine if different ratios of each compound would optimize attraction both in laboratory assays and in field studies. Field studies using the most attractive mixtures need to be conducted to corroborate our laboratory results. Additionally, testing attractive blends against more attractive fruit other than blueberry, such as strawberry and raspberry (Revadi et al., 2015), would determine whether this blend of volatiles capture adequate numbers of D. suzukii with differing competing background cues. These efforts to find an attractive blend of fruit ripening volatiles could improve early monitoring of D. suzukii when fruit is not yet susceptible.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
BB and LB designed the study. LB carried out the study, did the statistical analysis, and wrote the manuscript with contributions from BB and JP. All authors contributed to the article and approved the submitted version.
Funding
This research was partially funded through two United States Department of Agriculture (USDA), Specialty Crops Block Grant Programs (SCBGP) via the Missouri Department of Agriculture (MDA) (Agreement Numbers 14SCBGPMO0029 and 16SCBGPMO0003).
Author Disclaimer
The contents of this paper are solely the responsibility of the authors and do not necessarily represent the official views of the USDA or MDA.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Susan Kelso, Natalie Diesel, Jordan Hoffman, and Nicole Pruess for their technical support, and Mark Ellersieck (University of Missouri) for assistance with the statistical analyses.
References
Abraham, J., Zhang, A. J., Angeli, S., Abubeker, S., Michel, C., Feng, Y., et al. (2015). Behavioral and antennal responses of Drosophila suzukii (Diptera: Drosophilidae) to volatiles from fruit extracts. Environ. Entomol. 44, 356–367. doi: 10.1093/ee/nvv013
Alnajjar, G., Judith, C., and Drummond, F. (2017). Behavioral and preventative management of Drosophila suzukii Matsumura (Diptera: Drosophilidae) in Maine wild blueberry (Vaccinium angustifolium Aiton) through attract and kill trapping and insect exclusion-netting. Int. J. Entomol. Nematol. 3, 51–56.
Asplen, M. K., Anfora, G., Biondi, A., Choi, D., Chu, D., Daane, K. M., et al. (2015). Invasion biology of spotted-wing Drosophila (Drosophila suzukii): a global perspective and future priorities. J. Pest Sci. 88, 469–494.
Bolda, M. P., Goodhue, R. E., and Zalom, F. G. (2010). Spotted wing Drosophila: potential economic impact of a newly established pest. Agric. Res. Econ. Update 13, 5–8.
Bolton, L. G., Piñero, J. C., and Barrett, B. A. (2019). Electrophysiological and behavioral responses of Drosophila suzukii (Diptera: Drosophilidae) towards the leaf volatile β-cyclocitral and selected fruit-ripening volatiles. Environ. Entomol. 48, 1049–1055. doi: 10.1093/ee/nvz092
Bruce, T. J., and Pickett, J. A. (2011). Perception of plant volatile blends by herbivorous insects–finding the right mix. Phytochemistry 72, 1605–1611. doi: 10.1016/j.phytochem.2011.04.011
Bruce, T. J., Wadhams, L. J., and Woodcock, C. M. (2005). Insect host location: a volatile situation. Trends Plant Sci. 10, 269–274. doi: 10.1016/j.tplants.2005.04.003
Burrack, H. J., Fernandez, G. E., Spivey, T., and Kraus, D. A. (2013). Variation in selection and utilization of host crops in the field and laboratory by Drosophila suzukii Matsumura (Diptera: Drosophilidae), an invasive frugivore. Pest Manag. Sci. 69, 1173–1180. doi: 10.1002/ps.3489
Calabria, G., Máca, J., Bächli, G., Serra, L., and Pascual, M. (2012). First records of the potential pest species Drosophila suzukii (Diptera: Drosophilidae) in Europe. J. Appl. Entomol. 136, 139–147.
Cha, D. H., Adams, T., Rogg, H., and Landolt, P. J. (2012). Identification and field evaluation of fermentation volatiles from wine and vinegar that mediate attraction of spotted wing Drosophila, Drosophila suzukii. J. Chem. Ecol. 38, 1419–1431. doi: 10.1007/s10886-012-0196-5
Cha, D. H., Adams, T., Werle, C. T., Sampson, B. J., Adamczyk, J. J. Jr., Rogg, H., et al. (2014). A four-component synthetic attractant for Drosophila suzukii (Diptera: Drosophilidae) isolated from fermented bait headspace. Pest. Manag. Sci. 70, 324–331. doi: 10.1002/ps.3568
Cha, D. H., Landolt, P. J., and Adams, T. B. (2017). Effect of chemical ratios of a microbial based feeding attractant on trap catch of Drosophila suzukii (Diptera: Drosophilidae). Environ. Entomol. 46, 907–915. doi: 10.1093/ee/nvx079
Cini, A., Ioriatti, C., and Anfora, G. (2012). A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bull. Insectol. 65, 149–160.
Cloonan, K. R., Abraham, J., Angeli, S., Sied, Z., and Rodriguez-Saona, C. (2018). Advances in the chemical ecology of the spotted wing Drosophila (Drosophila suzukii) and its applications. J. Chem. Ecol. 44, 922–939. doi: 10.1007/s10886-018-1000-y
Cook, S. M., Khan, Z. R., and Pickett, J. A. (2007). The use of push-pull strategies in integrated pest management. Annu. Rev. Entomol. 52, 375–400. doi: 10.1146/annurev.ento.52.110405.091407
Del Socorro, A. P., Gregg, P. C., Alter, D., and Moore, C. J. (2010). Development of a synthetic plant volatile-based attracticide for female noctuid moths. I. Potential sources of volatiles attractive to Helicoverpa armigera (Hübner)(Lepidoptera: Noctuidae). Austral. J. Entomol. 49, 10–20. doi: 10.1111/j.1440-6055.2009.00733.x
Deprá, M., Poppe, J. L., Schmitz, H. J., De Toni, D. C., and Valente, V. L. S. (2014). The first records of the invasive pest Drosophila suzukii in the South American continent. J. Pest. Sci. 87, 379–383. doi: 10.1007/s10340-014-0591-5
Dewitte, P., Kerckvoorde, V. V., Beliën, T., Bylemans, D., and Wenseleers, T. (2021). Identification of blackberry (Rubus fruticosus) volatiles as Drosophila suzukii attractants. Insects 12:417. doi: 10.3390/insects12050417
Diepenbrock, L. M., and McPhie, D. R. (2018). Use of a novel host plant family, Basallaceae, by Drosophila suzukii (Diptera: Drosophilidae). Proc. Entomol. Soc. Wash. 120, 255–260. doi: 10.4289/0013-8797.120.2.255
El-Sayed, A. M., Suckling, D. M., Wearing, C. H., and Byers, J. A. (2006). Potential of mass trapping for long-term pest management and eradication of invasive species. J. Econ. Entomol. 99, 1550–1564. doi: 10.1603/0022-0493-99.5.1550
Feng, Y., Bruton, R., Park, A., and Zhang, A. (2018). Identification of attractive blend for spotted wing Drosophila, Drosophila suzukii, from apple juice. J. Pest Sci. 91, 1251–1267. doi: 10.1007/s10340-018-1006-9
Fraser, A. M., Mechaber, W. L., and Hildebrand, J. G. (2003). Electroantennographic and behavioral responses of the sphinx moth Manduca sexta to host plant headspace volatiles. J. Chem. Ecol. 29, 1813–1833. doi: 10.1023/a:1024898127549
Goodhue, R. E., Bolda, M., Farnsworth, D., Williams, J. C., and Zalom, F. G. (2011). Spotted wing Drosophila infestation of California strawberries and raspberries: economic analysis of potential revenue losses and control costs. Pest. Manag. Sci. 67, 1396–1402. doi: 10.1002/ps.2259
Hamby, K. A., Bolda, M. P., Sheehan, M. E., and Zalom, F. G. (2014). Seasonal monitoring for Drosophila suzukii (Diptera: Drosophilidae) in California commercial raspberries. Environ. Entomol. 43, 1008–1018. doi: 10.1603/EN13245
Hamby, K. A., Hernández, A., Boundy-Mills, K., and Zalom, F. G. (2012). Associations of yeasts with spotted-wing Drosophila (Drosophila suzukii; Diptera: Drosophilidae) in cherries and raspberries. Appl. and Environ. Microbiol. 78, 4869–4873. doi: 10.1128/AEM.00841-12
Hampton, E., Koski, C., Barsoian, O., Faubert, H., Cowles, R. S., and Alm, S. R. (2014). Use of early ripening cultivars to avoid infestation and mass trapping to manage Drosophila suzukii (Diptera: Drosophilidae) in Vaccinium corymbosum (Ericales: Ericaceae). J. Econ. Entomol. 107, 849–857. doi: 10.1603/EC14232
Iglesias, L. E., Nyoike, T. W., and Liburd, O. E. (2014). Effect of trap design, bait type, and age on captures of Drosophila suzukii (Diptera: Drosophilidae) in berry crops. J. Econ. Entomol. 107, 1508–1518. doi: 10.1603/ec13538
Ioriatti, C., Walton, V., Dalton, D., Anfora, G., Grassi, A., Maistri, S., et al. (2015). Drosophila suzukii (Diptera: Drosophilidae) and its potential impact to wine grapes during harvest in two cool climate wine grape production regions. J. Econ. Entomol. 108, 1148–1155. doi: 10.1093/jee/tov042
Karageorgi, M., Bräcker, L. B., Lebreton, S., Minervino, C., Cavey, M., Siju, K. P., et al. (2017). Evolution of multiple sensory systems drives novel egg-laying behavior in the fruit pest Drosophila suzukii. Curr. Biol. 27, 847–853. doi: 10.1016/j.cub.2017.01.055
Keesey, I. W., Knaden, M., and Hansson, B. S. (2015). Olfactory specialization in Drosophila suzukii supports and ecological shift in host preferences from rotten to fresh fruit. J. Chem. Ecol. 41, 121–128. doi: 10.1007/s10886-015-0544-3
Kleiber, J. R., Unelius, C. R., Lee, J. C., Suckling, D. M., Qian, M. C., and Bruck, D. J. (2014). Attractiveness of fermentation and related products to spotted wing Drosophila (Diptera: Drosophilidae). Environ. Entomol. 43, 439–447. doi: 10.1603/EN13224
Lance, D. R. (1983). “Host-seeking behavior of the gypsy moth: the influence of polyphagy and highly apparent host plants,” in Herbivorous Insects: Host-seeking Behavior and Mechanisms, ed. S. Ahmad (New York, NY: Academic Press), 210–224.
Landolt, P. J., Adams, T., and Rogg, H. (2012a). Trapping spotted wing Drosophila, Drosophila suzukii (Matsumura) (Diptera: Drosophilidae), with combinations of vinegar and wine, and acetic acid and ethanol. J. Appl. Entomol. 136, 148–154. doi: 10.1111/j.1439-0418.2011.01646.x
Landolt, P. J., Adams, T., Davis, T. S., and Rogg, H. (2012b). Spotted wing Drosophila, Drosophila suzukii (Diptera: Drosophilidae), trapped with combinations of wines and vinegars. Fla Entomol. 95, 326–332. doi: 10.1653/024.095.0213
Lee, J. C., Bruck, D. J., Dreves, A. J., Ioriatti, C., Vogt, H., and Baufeld, P. (2011a). In focus: spotted wing Drosophila, Drosophila suzukii, across perspectives. Pest Manag. Sci. 67, 1349–1351. doi: 10.1002/ps.2271
Lee, J. C., Bruck, D. J., Curry, H., Edwards, D., Haviland, D. R., Van Steenwyk, R. A., et al. (2011b). The susceptibility of small fruits and cherries to the spotted-wing Drosophila, Drosophila suzukii. Pest Manag. Sci. 67, 1358–1367. doi: 10.1002/ps.2225
Littell, R. C., Henry, P. R., and Ammerman, C. B. (1998). Statistical analysis of repeated measures data using SAS procedures. J. Anim. Sci. 76, 1216–1231. doi: 10.2527/1998.7641216x
Liu, Y., Dong, W., Zhang, F., Kenis, M., Griepink, F., Zhang, J., et al. (2018). Identification of active components from volatiles of Chinese bayberry, Myrica rubra attractive to Drosophila suzukii. Arthropod. Plant Interact. 12, 435–442. doi: 10.1007/s11829-018-9595-z
Metcalf, R. L., and Metcalf, E. R. (1992). Plant Kairomones in Insect Ecology and Control. London: Chapman and Hall Ltd.
Mori, B. A., Whitener, A. B., Leinweber, Y., Revadi, S., Beers, E. H., Witzgall, P., et al. (2017). Enhanced yeast feeding following mating facilitates control of the invasive fruit pest Drosophila suzukii. J. Appl. Ecol. 54, 170–177.
Ørsted, I. V., and Ørsted, M. (2018). Species distribution models of the Spotted Wing Drosophila (Drosophila suzukii, Diptera: Drosophilidae) in its native and invasive range reveal an ecological niche shift. J. Appl. Ecol. 56, 423–435. doi: 10.1111/1365-2664.13285
Piñero, J. C., Barrett, B. A., Bolton, L. G., and Follett, P. A. (2019). β-cyclocitral synergizes the response of adult Drosophila suzukii (Diptera: Drosophilidae) to fruit juices and isoamyl acetate in a sex-dependent manner. Sci. Rep. 9:10574. doi: 10.1038/s41598-019-47081-z
Raguso, R. A., Light, D. M., and Pickersky, E. (1996). Electroantennogram responses of Hyles lineata (Sphingidae: Lepidoptera) to volatile compounds from Clarkia breweri (Onagraceae) and other moth-pollinated flowers. J. Chem. Ecol. 22, 1735–1766. doi: 10.1007/BF02028502
Revadi, S., Vitagliano, S., Rossi Stacconi, M. V., Ramasamy, S., Mansourian, S., Carlin, S., et al. (2015). Olfactory responses of Drosophila suzukii females to host plant volatiles. Physiol. Entomol. 40, 54–64.
Robacker, D. C., Warfield, W. C., and Flath, R. A. (1992). A four-component attractant for the Mexican fruit fly, Anastrepha ludens (Diptera: Tephritidae), from host fruit. J. Chem. Ecol. 18, 1239–1254. doi: 10.1007/BF00980077
Rodriguez-Saona, C. R., and Stelinski, L. L. (2009). “Behavior-modifying strategies in IPM: theory and practice,” in Integrated Pest Management: Innovation-Development Process, eds R. Peshin and A. K. Dhawan (Dordrecht: Springer), 263–315. doi: 10.1007/978-1-4020-8992-3_11
Scheidler, N. H., Liu, C., Hamby, K. A., Zalom, F. G., and Syed, Z. (2015). Volatile codes: correlation of olfactory signals and reception in Drosophila-yeast chemical communication. Sci. Rep. 5, 1–13. doi: 10.1038/srep14059
Schröder, R., and Hilker, M. (2008). The relevance of background odor in resource location by insects: a behavioral approach. Bioscience 58, 308–316. doi: 10.3389/fphys.2018.00049
Stökl, J., Strutz, A., Dafni, A., Svatos, A., Doubsky, J., Knaden, M., et al. (2010). A deceptive pollination system targeting drosophilids through olfactory mimicry of yeast. Cur. Biol. 20, 1846–1852. doi: 10.1016/j.cub.2010.09.033
Szendrei, Z., and Rodriguez-Saona, C. (2010). A meta-analysis of insect pest behavioral manipulation with plant volatiles. Ent. Exp. Appl. 134, 201–210. doi: 10.1111/j.1570-7458.2009.00954.x
Urbaneja-Bernat, P., Cloonan, K., Zhang, A. J., Salazar-Mendoza, P., and Rodriguez-Saona, C. (2021). Fruit volatiles mediate differential attraction of Drosophila suzukii to wild and cultivated blueberries. J. Pest Sci. 94, 1249–1263. doi: 10.1007/s10340-021-01332-z
Walsh, D. B., Bolda, M. P., Goodhue, R. E., Dreves, A. J., Lee, J., Bruck, D. J., et al. (2011). Drosophila suzukii (Diptera: Drosophilidae): invasive pest of ripening soft fruit expanding its geographic range and damage potential. J. Integr. Pest Manag. 2, G1–G7.
Webster, B., Bruce, T., Dufour, S., Birkemeyer, C., Birkett, M., Hardie, J., et al. (2008). Identification of volatile compounds used in host location by the black bean aphid, Aphis fabae. J. Chem. Ecol. 34, 1153–1161. doi: 10.1007/s10886-008-9510-7
Keywords: semiochemicals, behavior, monitoring, odorants, olfaction
Citation: Bolton LG, Piñero JC and Barrett BA (2022) Behavioral Responses of Drosophila suzukii (Diptera: Drosophilidae) to Blends of Synthetic Fruit Volatiles Combined With Isoamyl Acetate and β-Cyclocitral. Front. Ecol. Evol. 10:825653. doi: 10.3389/fevo.2022.825653
Received: 30 November 2021; Accepted: 11 February 2022;
Published: 10 March 2022.
Edited by:
John Abraham, University of Cape Coast, GhanaReviewed by:
Ahmed M. Saveer, North Carolina State University, United StatesDanielle Kirkpatrick, Trécé, Inc., United States
Thomas Buxton, University of Ghana, Ghana
Copyright © 2022 Bolton, Piñero and Barrett. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bruce A. Barrett, QmFycmV0dEJAbWlzc291cmkuZWR1