 Shivam Shrotriya
Shivam Shrotriya Hussain S. Reshamwala
Hussain S. Reshamwala Salvador Lyngdoh
Salvador Lyngdoh Yadvendradev V. Jhala
Yadvendradev V. Jhala Bilal Habib
Bilal Habib
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 06 July 2022
Sec. Conservation and Restoration Ecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.815996
This article is part of the Research Topic Conservation of Woolly Wolf: Adaptation, Ecology, Evolution, and Genetics View all 6 articles
The Trans-Himalayan landscape is an extreme cold desert with limited diversity and a low density of wild prey. The landscape has three widespread carnivores—the wolf Canis lupus chanco, snow leopard Panthera uncia, and red fox Vulpes vulpes—competing for similar prey. We studied predation patterns, dietary competition, and niche segregation of three sympatric carnivores in the Indian Trans-Himalayas. The Himalayan or wooly wolf is one of the oldest lineages and has adapted to extreme cold. We further studied the prey selection of the wolf in the high-altitude environment. We collected 1,600 carnivore scats (wolf = 542, snow leopard = 31, and red fox = 1,027), and additional data on 573 snow leopard scats from literature was used in the analyses. We found that livestock was a major contributor to the wolf (56.46%) and snow leopard diet (30.01%). The wolf consumed blue sheep (8.26%) and ibex (3.13%), whereas the snow leopard subsisted on blue sheep (30.79%) and ibex (17.15%), relatively more frequently. The red fox preyed upon small species like pika and marmots (18.85 %); however, livestock carrions (16.49%), fruits and seeds (15.05%), and human-derived material (11.89%) were also consumed frequently. The dietary niche of three carnivore species highly overlapped (Pianka's index = 0.503, simulated mean = 0.419, p = 0.15) due to livestock. The carnivores segregated their diet in prey items originating from the wild. We tested a relationship between carnivore diet variations and prey/predator functional traits using RLQ ordination. Predator morphology traits like body size, weight, and habitat preference significantly affected the prey selection (p < 0.05). The wolf and snow leopard avoided the competition through habitat selection, while the red fox coexisted by exploiting a broader niche. The wolf showed site-specific variation in prey preferences albeit no prey selection at the landscape level (G2 = 6.79, df = 12, p = 0.87). The wolf preferred wild prey over domestic species at the wild prey rich site. The carnivores in this pastoralist landscape have adapted to exploit livestock resources despite facing persecution. Therefore, managing livestock and simultaneously restoring wild prey is crucial for the conservation of the carnivore guild in the Trans-Himalayan ecosystem.
Predation and competition are key interactions that contribute to shaping the carnivore community, their distribution, and in turn the ecology of the region (Chase et al., 2002; Caro and Stoner, 2003; Andheria et al., 2007). The species with similar resource requirements have a high niche overlap (Chase and Leibold, 2003), and negative interactions could lead to competitive exclusion (Hardin, 1960). Therefore, co-occurring carnivore species are expected to have evolved morphological and behavioral adaptations to reduce risks of intraguild competition (Schoener, 1982; Hunter and Caro, 2008). Competitive interactions among the carnivores can result in top carnivores limiting the populations of subordinate carnivores (Creel and Creel, 1996; Donadio and Buskirk, 2006; Bhandari et al., 2021), feeding interference (Linnell and Strand, 2000), or even intraguild predation (Creel and Creel, 1996; Holt and Polis, 1997). Specializing on different food resources is an effective strategy to reduce the predation competition in coexisting carnivores over a long evolutionary time (Connell, 1980; Jones and Barmuta, 1998).
The Trans-Himalayan landscape is roughly demarcated as the northern watershed of the Brahmaputra and the upper Indus rivers spread between the Himalayan mountain range in south and the Bangong-Nujiang Suture zone in north (Sven Hedin's “Trans-Himalaya”, 1910). It is an extreme cold desert with limited diversity and a low density of wild prey species (Namgail, 2009; Mishra et al., 2016). Two large carnivores- the gray wolf Canis lupus and snow leopard Panthera uncia- coexist in the Trans-Himalayan landscape of India, Pakistan, Nepal, and Tibet (Jackson and Wangchuk, 2000; Namgail et al., 2007; Bocci et al., 2017; Chetri et al., 2020). The two predators compete for the same prey base across their distribution from Central Asia to the Himalayas (Jumabay-Uulu et al., 2014; Lyngdoh et al., 2014, 2020; Chetri et al., 2020). However, the mechanism that allows their coexistence despite the consumption of similar prey species remains unclear. The Trans-Himalayan landscape in India also harbors small populations of large and meso-carnivore species such as brown bear Ursus arctos, dhole Cuon alpinus, Eurasian lynx Lynx lynx, beech marten Martes foina, and Tibetan fox Vulpes ferrilata in limited areas (Chundawat and Qureshi, 1999; Pfister, 2004). However, the red fox Vulpes vulpes is another generalist small carnivore species found throughout the landscape and lives as a subordinate predator (Pfister, 2004; Reshamwala et al., 2018). Hence, the wolf, snow leopard, and red fox form the most widespread carnivore guild of the Trans-Himalayan landscape in India.
The body size of predators influences the community structure and resource sharing between species (Simberloff and Dayan, 1991; Hayward et al., 2006; Gómez-Ortiz et al., 2015; Monterroso et al., 2020). While large predators could push away the small predators from the best resources (Linnell and Strand, 2000), they could also provide an additional resource through carrion provisioning (Sivy et al., 2018; Ferretti et al., 2021). Therefore, we hypothesized that the red fox shifts its diet as a subordinate predator to avoid competition with the wolf and snow leopard while exploiting the carrion resource. We further postulated that the two large-bodied carnivores—snow leopard and the wolf—have evolved a behavioral or physical niche separation mechanism to allow coexistence. The primary research objective of this study was to investigate the niche breadth, dietary overlap, and strategies of food resource partitioning among three carnivore species of the Trans-Himalayan landscape in India.
The wolves in the high-altitude ecosystem of the Himalayas and the Tibetan Plateau are an ancient lineage that evolved in isolation from other wolves (Sharma et al., 2004). This sub-species of wolves, C. l. chanco—often termed the Himalayan wolf, Tibetan wolf, or Woolly wolf—has evolved a genetic adaptation to cope with the cold and hypoxic environment (Werhahn et al., 2018; Wang et al., 2020). The wolves of the Trans-Himalayan landscape generate special interest because of their unique evolutionary history. The gray wolf occupies a wide range of habitats, including open scrublands, grasslands, deserts, boreal forests, high arctic, and mountains across the northern hemisphere (Mech and Boitani, 2003; Busch, 2018). As a top predator, their prey selection in different habitats has had a transformative effect on local ecosystems through a trophic cascade (Ripple and Beschta, 2005; Wikenros, 2011; Ripple et al., 2015, but see also MacNulty et al., 2016). Wolves in different habitats have varying prey species, and they adapt their dietary spectrum accordingly (Mech and Boitani, 2003; Newsome et al., 2016). Studying the prey selection of the unique wolf lineage, the Himalayan wolf, is important to understand its dietary adaptations in the Trans-Himalayas. Even though the Trans-Himalaya is often described as a homogenous landscape (Singh et al., 2010), topographical and climatic variations within the landscape change the composition of prey assemblages (Namgail, 2009). Further, interferences from human societies through livestock availability and retaliatory persecution could also impact the feeding behavior of the wolves living in human landscapes (Meriggi and Lovari, 1996; Newsome et al., 2016). Therefore, we expected geographic variations in the prey choices of the wolf within the Trans-Himalayan landscape of India. Understanding the geographic pattern in prey choices of these high-altitude wolves was the second objective of this study.
The Trans-Himalayan landscape forms the northernmost part of India, spanning over 186,000 km2 area along the International border with China and Nepal. The largest area of the Trans-Himalayas falls in the Ladakh Union Territory of India. The landscape also expands to south of Ladakh into the Spiti valley, Himachal Pradesh. This study covered Ladakh and Spiti regions (31.5–35.2° N, 76–79° E) (Figure 1). The Trans-Himalayas have dry land of rugged mountains and open plains where the elevation ranges from 2,800 m in the valleys to 7,000 m high peaks. The mean annual precipitation rarely crosses 100 mm, and mainly occurs in the form of snow during the winter season. The temperature ranges from −30°C in winter to 35°C in summer (Namgail et al., 2009). Although sparse, the vegetation in Ladakh is a combination of steppe vegetation and shrubland (Rawat and Adhikari, 2005). The alpine meadows are dominated by Kobresia spp., Carex spp., Potentilla spp. and Nepeta spp., and shrublands are dominated by Hippophae spp., Salix spp. and Myricaria spp. (Kachroo et al., 1977). Eight species of ungulates provide the main source of wild prey- blue sheep or bharal Pseudois nayaur, Asiatic ibex Capra ibex sibirica, Ladakh urial Ovis vignei vignei, Tibetan argali Ovis ammon hodgsoni, Kiang Equus kiang, Tibetan gazelle Procapra picticaudata, Tibetan antelope Pantholops hodgsoni, and wild yak Bos mutus (Fox et al., 1991; Chundawat and Qureshi, 1999). The Ladakh region is one of the least populated areas with a human density of 4.9 individuals/km2 (Chandramouli, 2013). Most people are agro-pastoralists and can be classified into nomadic, semi-nomadic, and settled villagers. Livestock grazing and associated use of rangelands is widespread across the Trans-Himalayan landscape and has played an important role in the ecological history of the region (Mishra et al., 2001; Bagchi et al., 2004).
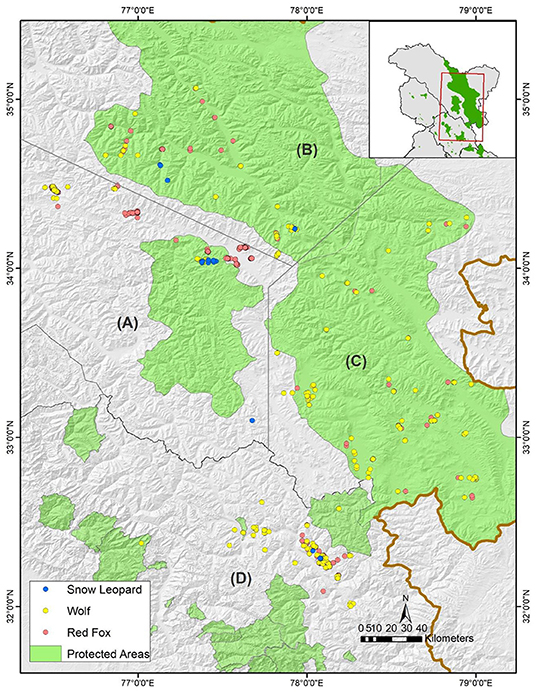
Figure 1. Geographic distribution of the scats of three carnivore species collected from the Trans-Himalayan landscape of Ladakh and Spiti valley, India. The study area was divided into four sub-regions- (A) Western Ladakh, which includes Hemis National Park, Zanskar and Kargil, (B) Nubra-Shyok Wildlife Sanctuary, (C) Changthang Wildlife Sanctuary, and (D) Spiti valley, Himachal Pradesh.
There are three major high-altitude protected areas in Ladakh- Hemis National Park, Changthang Wildlife Sanctuary (WLS), and Karakoram (Nubra-Shyok) WLS. The protected areas in this landscape are typically large and vary in their topography, vegetation, and species assemblages (Chundawat and Qureshi, 1999; Namgail, 2009). We divided the study area into four sub-regions based on geographical features, wild prey species, and the variations of agro-pastoralist systems to understand geographic variations in prey choices of the wolf. The western Ladakh, including Hemis national park, is comparatively rugged with steep slopes of the Zanskar mountain range where blue sheep and Ladakh urial are sighted commonly (Figure 1A). Nubra-Shyok WLS, the northern Ladakh, is part of the Karakoram mountain range where river valleys form a typical desert system and glaciers cover a significant portion (Figure 1B). Most people in Nubra-Shyok WLS are settled villagers, and wild prey here includes mainly blue sheep and the Asiatic ibex. Changthang WLS forms the easternmost region characterized by rolling plains, higher prey diversity (six out of eight ungulates), and nomadic pastoralism (Figure 1C). The Spiti valley in Himachal Pradesh is a narrow river valley adjoining high elevation peaks in the south and Changthang plains in the north (Figure 1D). The agro-pastoralist community is settled in the villages; however, seasonal pastoralists migrate to the grasslands of the valley from the south during summer. Kibber WLS and Pin valley national park are two main protected areas in the Spiti valley where blue sheep and Asiatic ibex are common prey species.
Various field excursions were made to collect the scat samples of the targeted species from 2014 to 2018 in the Ladakh and Spiti regions of the north-western Trans-Himalayan landscape in India. The scats were collected throughout the year, covering the winter (November to April) and summer (May to October) seasons. We collected a total of 1,600 scats of the three carnivore species, which included 542 scats of the Himalayan wolf (winter- 43.36%, summer- 56.64%), 31 scats of snow leopard (winter- 67.74%, summer- 32.26%) and 1,027 scats of red fox (winter- 58.03%, summer- 41.97%) (Figure 1). Scats were searched and picked on naturally occurring trails and other conspicuous sites such as den and rendezvous sites. The trails were not repeated in order to maximize the effort. If more than one scat was present at the site, a single fresh scat was collected from the site. Scats of the carnivores were identified and distinguished in the field based on shape, size, odor, and quantity typical to that of the relative species, following a standard protocol (Vanak and Mukherjee, 2008; Reshamwala et al., 2018). A conservative, multi-criteria approach to differentiate wolf scats from those of other canids was used by Ciucci et al. (1996). Since the field samples of snow leopard scats were limited, we augmented data from published studies on the snow leopard diet in the study area (Lyngdoh et al., 2014) for comparative analyses. Information on a total of 573 additional scats of snow leopard was obtained from three studies-−95 scats from Spiti valley in winter (Bagchi and Mishra, 2006), 217 scats from Spiti valley in summer and 45 scats in winter, 43 scats from Ladakh in summer (Suryawanshi et al., 2017), and 173 scats from Hemis national park, Ladakh collected throughout the year (Chundawat and Rawat, 1994).
Scat analysis is an indirect and non-invasive technique for recording the frequency of occurrence of prey in the diet of mammalian carnivores; hence it is used widely (Floyd et al., 1978; Shahi, 1982; Jhala, 1993; Mukherjee et al., 1994; Habib, 2007; Reshamwala et al., 2018). Scats were tagged with species, date, GPS location, and condition as recent or old at the time of collection and preserved for lab analysis. The standard method of lab analysis of undigested food content in the carnivore scats was followed to identify the prey species and other food (Mukherjee et al., 1994; Bagchi et al., 2003). Scats were washed in running water through a fine sieve of BSS 120 having a pore aperture width of 125 μm so that the digested material could pass through the sieve. Indigestible items including hair, feathers, bones, claws, teeth, chitin remnants of insects, plant material, and human-derived materials (HDM) such as cloth, paper, plastic, and rubber, including poultry waste, were recorded and collected for further identification if required. Dried mammalian hair samples were treated with xylene and alcohol solution and then examined under a microscope to identify prey consumed by medullary hair patterns (Bahuguna et al., 2010). The relative frequency of occurrence of prey items was used for further statistical analyses (Hayward and Kerley, 2005; Lyngdoh et al., 2014). DNA-based verification using the 148-bp region of the mitochondrial Cytochrome-b gene was performed for 118 scats to estimate the errors in field-based identification of the scats (Details in Supplement-A).
The statistical analyses were performed in the open-source program R version 3.6.3 (R Core Team, 2020). Relative biomass consumption of different prey items by the wolf was calculated using an allometric relationship developed by Chakrabarti et al. (2016):
The dietary niche breadth was calculated using Levins' measure (Levins, 1968). The niche overlap among three carnivores was calculated using Pianka's index (Pianka, 1974). We tested if the observed niche overlap was significantly different from the overlap under the null hypothesis, which expects that the carnivores consumed the prey species independent of one another. We used “ra3” randomization algorithm, a conservative approach retaining the species' niche breadth, with 10,000 repeats to test the diet niche overlap in “EcoSimR” package version 0.1.0 (Winemiller and Pianka, 1990; Gotelli and Ulrich, 2012; Gotelli et al., 2015). A hypothesis for diet variations in carnivore species arising from their functional traits was tested using the trait-based RLQ ordination method (Spitz et al., 2014). This three-table ordination method was originally developed to test the relationship of species traits with environmental features (Ter Braak et al., 2012). The method has been used to study the dietary relationships among marine predators and prey species, multi-scale niche study of waterbirds, and dietary analysis of bats (Ridoux, 1994; Scharf et al., 2000; Henry and Cumming, 2017; Arrizabalaga-Escudero et al., 2019). Three lists of (1) predator traits (R list), (2) matrix of prey-predator dietary interactions (L list), and (3) prey traits (Q list) were prepared. Predator and prey trait lists (R and Q lists) are provided in Table 1. The RLQ analyses were performed in the “ade4” package (Dray and Dufour, 2007), following the methods recommended by Dray and Legendre (2008) and Spitz et al. (2014).
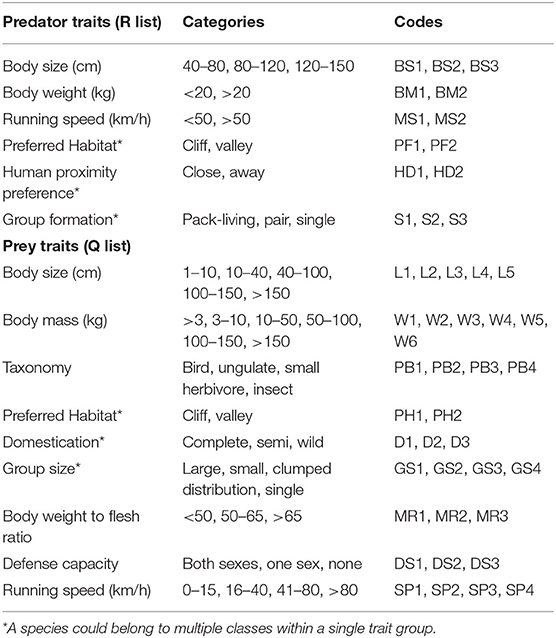
Table 1. Predator and prey traits used for RLQ analysis of the trait-based dietary relationship among carnivores in the Trans-Himalayan landscape, India.
Jacobs' selectivity index was used to determine the prey selectivity of the wolf using the formula:
where, ri is the proportion of species i among all the prey species consumed at a site and pi is the proportion of species i in the available prey community (Jacobs, 1974). We calculated prey selectivity separately at four geographic sub-regions of the landscape to understand site-specific differences in the diet of the wolf. The landscape was gridded into 10 × 10 km2 cells to remove the biases arising from unequal sampling among the regions. Only ten wolf scats per grid were selected randomly for the preference analysis. Literature references were used for the availability of the wild prey species found in the landscape (Namgail, 2009; Lyngdoh et al., 2014; Shrotriya et al., 2015; Suryawanshi et al., 2017; Habib et al., 2021). Livestock availability data were obtained from the Department of Animal Husbandry, Leh (Supplement-B). The log-likelihood test ratio was computed to test the differences in prey preference of the wolf across four sub-regions of the landscape (Karanth and Sunquist, 2000).
Livestock (cattle, yak, goat, sheep, horse, and donkey) was a major contributor to the wolf diet in the landscape (56.46%) (Table 2). The wolf consumed blue sheep (8.26%) and ibex (3.13%) among the wild ungulates. The small prey items such as pika (13.54%) and marmot (6.99%) were quite frequently represented in the wolf diet. Compared to the wolf, livestock contribution in the snow leopard diet was low (30.01%) (Table 2). Blue sheep (30.79%) was the major contributor to the snow leopard diet, followed by ibex (17.15%). Plant items were reported quite frequently in the data obtained from Chundawat and Rawat (1994). Small prey items such as pika (15.66%) and marmots (3.19%) were often present in the red fox diet (Table 2). However, the fox also frequently consumed livestock (16.49%), fruits and seeds (15.05%), and other human-derived materials along with poultry (11.89%).
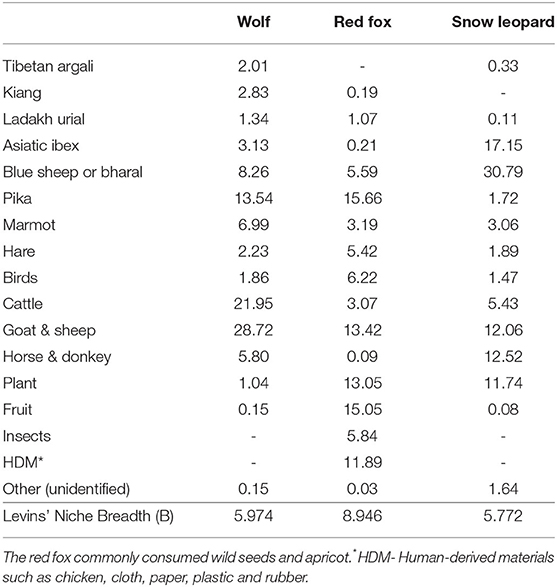
Table 2. Relative frequency of occurrence (%) of different prey and food categories in the diet of three carnivores (Himalayan wolf- 542 scats, red fox- 970, snow leopard- 606) and their niche breadth.
Only a portion of the scats collected (n = 118) could be verified by DNA-based identification. The misclassification error for the wolf scats was 5.15%, while none of the sampled scats of snow leopard could be verified (Supplement-A).
Although the dietary range of all the three carnivores varied in contribution from each food item, we did not find a significant diet niche separation among the three carnivores (Pianka's mean observed index = 0.503, randomized simulated mean index = 0.419, p = 0.15). Most food items were consumed by all three carnivore species, varying only in the contribution of each item (Figure 2, Table 2). Livestock consumption, either by hunting directly or utilizing it through scavenging, was the major source of the niche overlap. Ordination based on RLQ analysis revealed that the consumption of large body-sized prey species such as ibex, argali, kiang, yak, and the horse was associated with the wolf and snow leopard, while small prey such as pika, marmot, hare, birds, and insects grouped with the red fox (Figure 3). However, the body size of the prey species could not clearly differentiate between the diet of the wolf and snow leopard. No evidence was found for the effect of prey traits on dietary relationships. Predator morphology traits such as body size and body weight, and habitat preference in relation to human presence significantly affected the variation in prey selection (p < 0.05) (Figure 3).
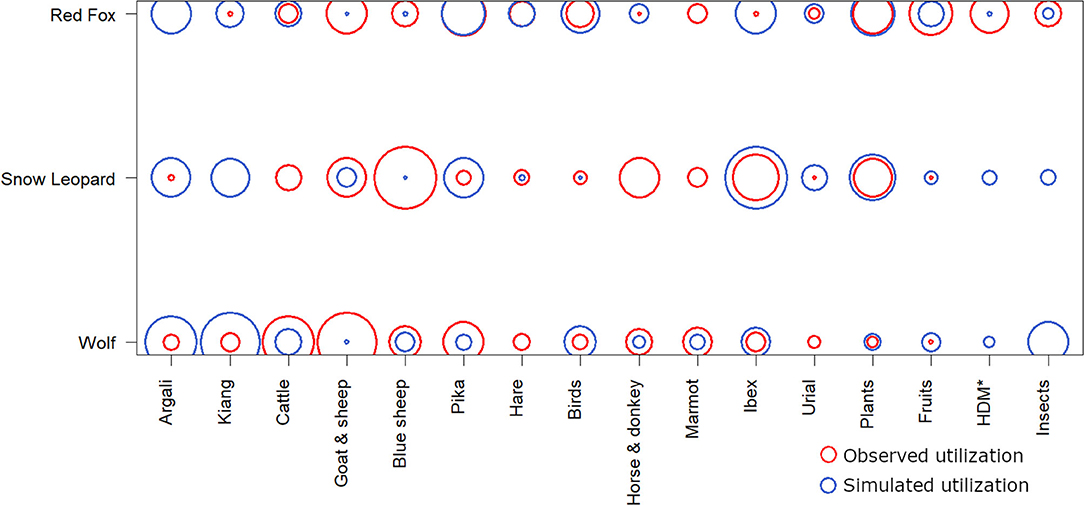
Figure 2. Comparative dietary niche of the red fox, snow leopard and the wolf (Pianka's mean observed index = 0.503 simulated mean = 0.419; p = 0.15). Larger circles represent relatively higher utilization of the food item. For example, the snow leopard consumed blue sheep more than the red fox and wolf. Further, goat and sheep were consumed more, while ibex was consumed less than the simulated utilization.
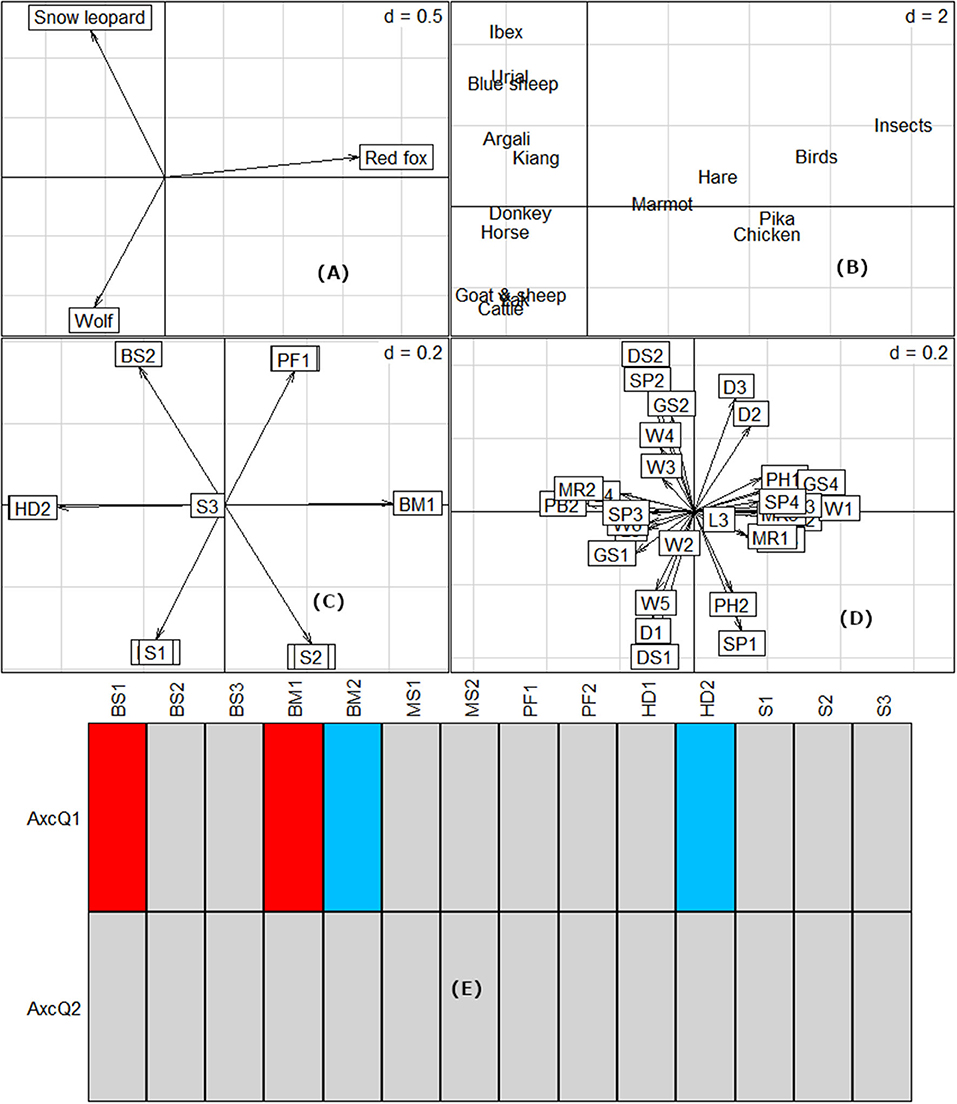
Figure 3. RLQ ordination of the predator (A) and prey raw scores (B) along the first two axes. The ordination is based on the canonical weights of predator (C) and prey (D) traits and their dietary relationships. None of the prey traits influenced the dietary choices. However, predator traits of body size, body mass and habitat preference significantly influenced the dietary relationship of the three carnivores. The traits marked in red are significant at p < 0.05, while the traits marked in blue are significant at p < 0.1 (Refer to the trait codes in Table 1).
The biomass correction in the diet of the wolf followed the pattern obtained from the frequency of occurrence for most of the food items. However, over-representation of the small prey items such as pika and marmot was corrected when accounted for biomass consumption (Figure 4). While pika (4.37%) and marmot (4.72%) provided less biomass to the wolf, large prey items contributed more biomass than represented by the frequency of occurrence; for example, blue sheep contributed 10.01% biomass. A log-likelihood test ratio for prey preferences of the Himalayan wolf showed that statistically significant preferences for prey items did not exist at the landscape level (G2 = 6.79, df = 12, p = 0.871). However, the dietary preferences of the wolf varied across all four sub-regions of the landscape (Figure 5). Wild ungulates such as argali, blue sheep, ibex, and urial were often consumed more than the availability, and small prey items such as pika, marmot, and hare frequently showed a negative selection. Preferences for the livestock varied across the landscape. The livestock species were preferred in the Nubra-Shyok WLS, Spiti valley, and the western Ladakh region. However, the preference for livestock decreased in eastern Ladakh, where Changthang WLS exists (Figure 5). It is to be noted that Changthang also holds the highest overall wild prey diversity and density.
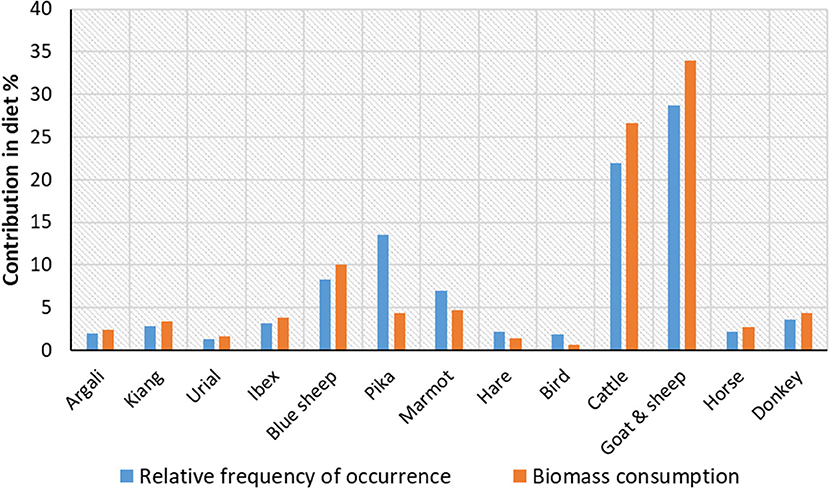
Figure 4. Contribution of different diet categories in the diet of wolf in the Trans-Himalayan landscape, India. Biomass provides a better representation than the frequency of occurrences for small prey items such as pika and marmot.
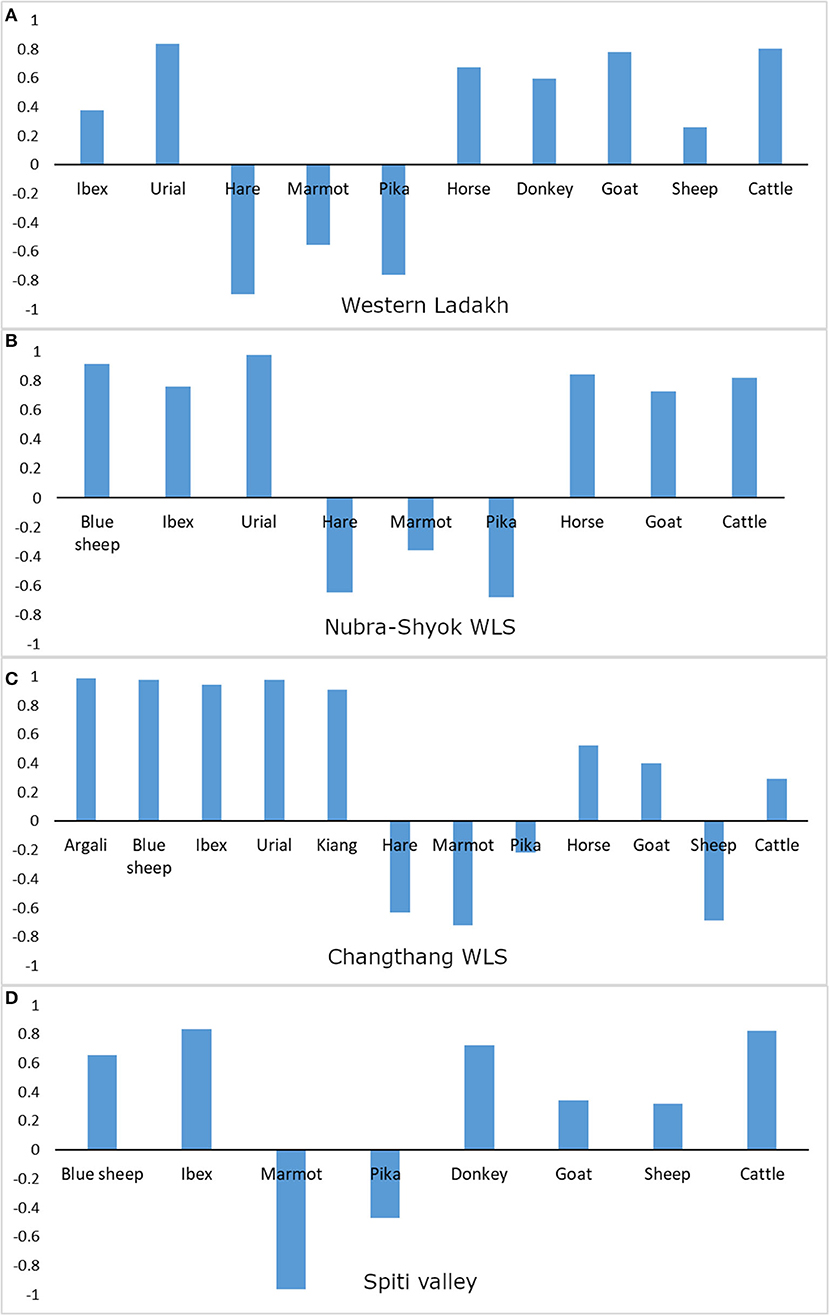
Figure 5. Jacob's index values for prey items of the wolf in different sub-regions of the Trans-Himalayan landscape in India. (A) Western Ladakh, including Hemis National Park, Zanskar and Kargil, (B) Nubra-Shyok Wildlife Sanctuary, (C) Changthang Wildlife Sanctuary, and (D) Spiti valley, Himachal Pradesh. Positive values indicate relatively higher consumption than the availability, and negative values indicate vice versa.
Intraguild diet competition among carnivores depends on their evolutionary history together and the availability of the resources (Connell, 1980; Chase and Leibold, 2003; Hunter and Caro, 2008). Three carnivore species of the Trans-Himalayan landscape in India—the wolf, snow leopard, and red fox—exhibited a very high overlap of the food items in this study. Out of 17 food categories, only insects and human-derived material were absent from the diet of snow leopard and the wolf (Figure 2 and Table 2). Kiang was absent from the snow leopard diet, and Tibetan argali was absent from the red fox diet. Kiang is distributed throughout eastern Ladakh, while argali is found in a few small pockets in Changthang WLS (Namgail, 2009). We found the body size and body mass to be the most influential predator traits determining the intraguild feeding relationship of the carnivores in this landscape (Figure 3). Body size differences of predators are key drivers of their dietary choices, as it influences the ability of carnivores to hunt different prey species and restrict the mesopredators to small prey (Simberloff and Dayan, 1991; Monterroso et al., 2020). In this study, the red fox was a small body-sized mesopredator species, resulting in its diet of small animals. The generalist and opportunistic habits of small-sized carnivores allow exploiting a wide range of prey and favor their coexistence with top carnivores (Jaksic et al., 1996; Gómez-Ortiz et al., 2015). Red fox also had the widest niche breadth (B = 8.946), including insects, various fruits and seeds, and human-derived materials. Red foxes in this landscape have adapted to utilize garbage from human sources, increasing their densities around the human settlements (Reshamwala et al., 2018). Apricots formed a large chunk of their fruit consumption in western Ladakh, while wild berries were frequently consumed in Nubra-Shyok WLS. The presence of livestock and large wild prey species in the red fox diet resulted in its dietary overlap with the wolf and snow leopard (Figure 2). However, the large prey could be originated from the secondary consumption of the carrions originally hunted by the top predators or died naturally (Ghoshal et al., 2016; Reshamwala et al., 2018; Ferretti et al., 2021).
Despite the similar niche breadth, the wolf (B = 5.974) and snow leopard (B = 5.772) showed prey-specific variations in consumption rates. For example, Asiatic ibex and blue sheep were consumed by snow leopard more frequently compared to the wolf. The wolf in the mountain landscape is often associated with the valleys and flat areas (Paquet et al., 1996; Habib et al., 2021), while the snow leopard selects high ruggedness areas with cliffs and steep slopes (Watts et al., 2019). Their habitat preferences also relate to their hunting strategies. The snow leopard often hunts individually by ambush method, while wolves are pack hunters using chase down method. Valley flats in the mountain regions are also the areas preferred by humans for settling; hence closeness to human settlement significantly affected the predation choices in our analysis (Figure 3). The human settlements are also associated with the presence of livestock, which was a common prey resource for all three carnivores. Habitat characteristics played a role in the prey preferences of snow leopard and the wolf in other parts of the Himalayas as well (Sharma et al., 2007; Chetri et al., 2017).
Except for a few recent and site-specific studies (e.g., Bocci et al., 2017; Chetri et al., 2017; Ud Din et al., 2017; Jamtsho and Katel, 2019; Werhahn et al., 2019; Lyngdoh et al., 2020), the diets and feeding behavior of the wolves of the Himalayan landscape are understudied. Newsome et al. (2016) reviewed the diet of all the gray wolves of the world using 177 studies. The representation from Asia was limited to only 15 studies, and no study from the Himalayan region was included in the review. Later, Lyngdoh et al. (2020) specifically reviewed 22 studies on the dietary pattern of the Himalayan wolf in central and southeast Asia. Lyngdoh et al. (2020) found that livestock consumption (54.92%) was widespread in the Himalayan wolf diet across the mountain areas of Asia. A total of 39 prey species were recorded in the wolf diet with highly localized selection for rare species such as Przewalski's horse Equus ferus przewalskii. Our data suggest a similar pattern with a high livestock consumption (56.46%) in the Trans-Himalayan landscape of India. Tibetan argali is a rare and localized wild ungulate species in Changthang WLS, which had the highest predation selection ratio by the wolf (Jackob's index = 0.98). This information is potentially valuable for the conservation of argali, which is one of the most threatened ungulate species in Ladakh with an estimated population of 300–360 individuals only (Namgail et al., 2009). Wolf predation was considered one of the major threats to the conservation of endangered Przewalski's gazelle Procapra przewalskii in Qinghai, China (Liu and Jiang, 2003).
Predation by the wolf is a hierarchical selection process (McPhee et al., 2012). They usually consume medium-size prey (Kunkel and Pletscher, 2001), but specialization for hunting the large prey is also reported commonly (Ripple et al., 2001; Feldhamer et al., 2003; Ripple and Beschta, 2012). Several studies from the European region found that wolves hunted on medium-sized prey comparatively more than the large prey such as European bison Bison bonasus, and moose Alces alces (Nowak et al., 2011; Wagner et al., 2012). Our results align with these studies as the wolves in the Trans-Himalayas consumed medium-sized prey such as argali, ibex, blue sheep, goat, and sheep more than large-sized prey such as kiang, yak, horse, and donkey (Table 2). However, we also found a considerable presence of small prey items in the wolf diet. During the field surveys, we observed a few instances where individual wolves showed behavioral specialization to hunt small prey such as marmot. The Trans-Himalayan landscape shows extreme weather where prey availabilities might fluctuate with the seasons. In addition, nomadic pastoralism could bring strong seasonal changes in the availability of livestock. Small species in such cases could allow the wolves to switch to locally abundant prey. Newsome et al. (2016) also observed that the Asian studies reported a relatively higher amount of small prey such as rodents than the European studies on the wolf diet.
While the wolf can be considered a diet generalist throughout the landscape (Figure 3), it showed a variation in site-specific preferences (Figure 5). Livestock was highly preferred by the wolf in the western Ladakh and Spiti valley, where wild prey diversity and availability are low compared to other parts of the landscape (Figure 5). In Nubra-Shyok WLS, the wolf showed almost equal preferences for wild and domestic prey. Pastoralist practices are the least common in Nubra-Shyok WLS, and domestic prey availability is the lowest (Supplement-A). Despite prevailing nomadic pastoralism and higher availability of domestic prey in Changthang WLS, prey selectivity was high for wild prey species. Wolves are known to subsist on livestock throughout their distribution range (Mech and Boitani, 2003; Newsome et al., 2016). However, evidence exists that wolves prefer wild prey over domestic prey if the wild prey populations occur at relatively higher densities (Meriggi and Lovari, 1996; Sidorovich et al., 2003; Newsome et al., 2016). The patterns of site-specific preferences in this study reflect the current wolf predation dynamics of the landscape, which could change with the availability of domestic and wild prey in future. Recovery of wild ungulate prey is considered an important measure to reduce the extent of livestock depredation by the wolf (Werhahn et al., 2019). In contrast, Treves et al. (2004) showed that an increase in wild prey availability resulted in an increased presence of wolves, which could intensify the livestock depredation (Newsome et al., 2016). The gray wolf showed both type II and type III responses to wild ungulate prey (Garrott et al., 2007). However, our study suggested that higher livestock presence in Changthang WLS did not increase livestock predation, possibly due to better wild prey availability. The Indian wolf C. l. pallipes, the closest sub-species to the Himalayan or woolly wolf (Sharma et al., 2004; Hennelly et al., 2021), also highly depends on livestock (Shahi, 1982; Kumar and Rahmani, 2000; Jethva and Jhala, 2004; Habib, 2007). The Indian wolf also showed a low livestock consumption when blackbuck Antilope cervicapra was abundantly available (Jhala, 1993).
A high niche overlap in the diets of three widespread carnivore species in the Trans-Himalayan landscape indicates that the sympatric carnivores face intense competition (Hardin, 1960; Schoener, 1982; Chase and Leibold, 2003). The Trans-Himalayan landscape is among the least productive graminoid-dominated ecosystems (Mishra, 2001). As a result, the low diversity and density of the wild prey species in the landscape (Namgail, 2009) could raise predation competition. Our results suggest that the wolf and snow leopard avoided the competition through habitat preference, while the red fox coexisted by expanding its niche. Livestock was a common and abundant resource available in this agro-pastoralist landscape (Mishra et al., 2001), which allowed the diet overlap, facilitating the coexistence of the carnivores. The landscape has a history of pastoralism for at least three millennia (Handa, 1994; Schaller, 1998). Hence the carnivores of the region might have adapted to the presence and availability of livestock and developed strategies to exploit livestock resources despite facing persecution.
A major limitation of this study was the insufficiency of snow leopard scat samples from the field. We augmented the data from literature scattered from 1994 to 2017, which may lack comparative representativeness. Therefore, the snow leopard diet analyses assumed that its diet niche did not change significantly within this time frame. The prey availability estimates of small species such as pika and marmot were older and based on extrapolation of total counts, unlike the estimates of wild prey based on distance sampling. We carried out DNA-based verification of the scats only for a subset of the samples. The field identification of the wolf scats was reliably confirmed, and red fox scats were distinguishable from large carnivore scats based on physical characteristics of shape, size and smell (Reshamwala et al., 2018). The majority of the snow leopard data in this study were from the literature. Based on the field knowledge and the molecular verification, we believe that the misclassification errors were low, randomly distributed and unlikely to alter the observed predation patterns. Although our results carry associated errors, this study helps understand the dietary competition of three widespread carnivores in the Indian Trans Himalayas. Feral dogs have a considerable presence in the landscape and are known to compete with carnivorous wildlife for resources (Ghoshal et al., 2016; Home et al., 2017; Reshamwala et al., 2018). We did not account for the effect of feral dogs on dietary ecology of other carnivores. A possible interference competition of feral dogs with wolves and foxes might be limiting their access to food resources close to human settlements. However, the presence of dogs is clumped, and a fine-scale study on this aspect should be carried out in the future.
Livestock predation by the carnivores and their retaliatory killing is a primary concern for the local economy and conservation in the Trans-Himalayan landscape of India (Bagchi et al., 2004; Suryawanshi et al., 2017). Therefore, managing livestock in relation to wild prey is crucial for the Trans-Himalayan ecosystem. Severe reduction of the livestock could result in population decrease for all the carnivores as well as heighten competition, especially due to depressed wild ungulate populations. Changing practices and modernization is leading to decreased livestock availability in the past few decades (Dollfus, 2012). Therefore, the management should simultaneously focus on wild prey restoration, which is likely to change the preferences of the wolf and other carnivores. In addition, reducing persecution is suggested as a more effective method for restoring large carnivores than restoring their prey (Bleyhl et al., 2021). The snow leopard is a flagship conservation species in the Trans-Himalayan landscape of India. The Himalayan or woolly wolf also needs to be elevated in conservation programs, promoting public acceptance and reducing persecution.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was reviewed and approved by Wildlife Institute of India Ethics Committee.
BH and SS conceptualized and planned the study. SS, HR, and SL conducted fieldwork and collected the data. SS carried out the formal analysis and wrote the manuscript with inputs from YJ and BH. All the authors edited and reviewed the final manuscript.
MBZ species conservation fund provided initial funding at the beginning of the study. Grant-In Aid, Wildlife Institute of India provided major funds for the study.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Dean and Director, Wildlife Institute of India are acknowledged for institutional support and funding. We thank the Department of Wildlife Protection, Leh-Ladakh and Forest Department, Himachal Pradesh for necessary permissions. In particular, Mr. Jigmet Takpa, Mr. Intesar Suhail, Mr. Tsering Angchok and Mr. Pankaj Raina of the Department of Wildlife Protection, Leh, helped thoughout the fieldwork. The field staff of the concerned departments and a number of volunteers participated in field surveys and data collection. Dawa Tashi, Rigzin Tamchos, Neeraj Mahar, Anuj Patil, Bhushan Sayanke and Vinod Thakur are thanked for assisting during laboratory analyses. Dr. SP Goyal reviewed and improved an initial draft of the manuscript.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.815996/full#supplementary-material
Andheria, A. P., Karanth, K. U., and Kumar, N. S. (2007). Diet and prey profiles of three sympatric large carnivores in Bandipur Tiger Reserve, India. J. Zool. 273, 169–175. doi: 10.1111/j.1469-7998.2007.00310.x
Arrizabalaga-Escudero, A., Merckx, T., García-Baquero, G., Wahlberg, N., Aizpurua, O., Garin, I., et al. (2019). Trait-based functional dietary analysis provides a better insight into the foraging ecology of bats. J. Anim. Ecol. 88, 1587–1600. doi: 10.1111/1365-2656.13055
Bagchi, S., Goyal, S. P., and Sankar, K. (2003). Prey abundance and prey selection by tigers (Panthera tigris) in a semi-arid, dry deciduous forest in western India. J. Zool. 260, 285–290. doi: 10.1017/S0952836903003765
Bagchi, S., and Mishra, C. (2006). Living with large carnivores: predation on livestock by the snow leopard (Uncia uncia). J. Zool. 268, 217–224. doi: 10.1111/j.1469-7998.2005.00030.x
Bagchi, S., Mishra, C., and Bhatnagar, Y. V. (2004). Conflicts between traditional pastoralism and conservation of Himalayan ibex (Capra sibirica) in the Trans-Himalayan mountains. Anim. Conserv. 7, 121–128. doi: 10.1017/S1367943003001148
Bahuguna, A., Sahajpal, V., Goyal, S. P., Mukherjee, S. K., and Thakur, V. (2010). Species Identification From Guard Hair of Selected Indian Mammals: A Reference Guide. Dehradun: Wildlife Institute of India.
Bhandari, A., Ghaskadbi, P., Nigam, P., and Habib, B. (2021). Dhole pack size variation: assessing the effect of Prey availability and Apex predator. Ecol. Evol. 11, 4774–4785. doi: 10.1002/ece3.7380
Bleyhl, B., Ghoddousi, A., Askerov, E., Bocedi, G., Breitenmoser, U., Manvelyan, K., et al. (2021). Reducing persecution is more effective for restoring large carnivores than restoring their prey. Ecol. Appl. 31:e02338. doi: 10.1002/eap.2338
Bocci, A., Lovari, S., Khan, M. Z., and Mori, E. (2017). Sympatric snow leopards and Tibetan wolves: coexistence of large carnivores with human-driven potential competition. Eur. J. Wildl. Res. 63, 92–100. doi: 10.1007/s10344-017-1151-0
Caro, T. M., and Stoner, C. J. (2003). The potential for interspecific competition among African carnivores. Biol. Conserv. 110, 67–75. doi: 10.1016/S0006-3207(02)00177-5
Chakrabarti, S., Jhala, Y. V., Dutta, S., Qureshi, Q., Kadivar, R. F., and Rana, V. J. (2016). Adding constraints to predation through allometric relation of scats to consumption. J. Anim. Ecol. 85, 660–670. doi: 10.1111/1365-2656.12508
Chandramouli, C. (2013). Census of India 2011, Primary Census Abstract. New Delhi: Office of the Registrar General and Census Commissioner, India.
Chase, J. M., Abrams, P. A., Grover, J. P., Diehl, S., Chesson, P., Holt, R. D., et al. (2002). The interaction between predation and competition: a review and synthesis. Ecol. Lett. 5, 302–315. doi: 10.1046/j.1461-0248.2002.00315.x
Chase, J. M., and Leibold, M. A. (2003). Ecological Niches: Linking Classical and Contemporary Approaches. Chicago: University of Chicago Press. doi: 10.7208/chicago/9780226101811.001.0001
Chetri, M., Odden, M., Devineau, O., McCarthy, T., and Wegge, P. (2020). Multiple factors influence local perceptions of snow leopards and Himalayan wolves in the central Himalayas, Nepal. PeerJ 8:e10108. doi: 10.7717/peerj.10108
Chetri, M., Odden, M., and Wegge, P. (2017). Snow leopard and himalayan wolf: food habits and prey selection in the Central Himalayas, Nepal. PLoS ONE 12:e0170549. doi: 10.1371/journal.pone.0170549
Chundawat, R. S., and Qureshi, Q. (1999). Planning Wildlife Conservation in Leh and Kargil Districts of Ladakh, Jammu and Kashmir. Final Report. Dehradun: Wildlife Institute of India.
Chundawat, R. S., and Rawat, G. S. (1994). “Food habits of snow leopard in Ladakh, India,” in Proceedings of the International Snow Leopard Symposium, 1989, eds J. L. Fox and D. Juzeng (Seattle: International Snow Leopard Trust), 127–132.
Ciucci, P., Boitani, L., Pelliccioni, E. R., Rocco, M., and Guy, I. (1996). A comparison of scat-analysis methods to assess the diet of the wolf Canis lupus. Wildl. Biol. 2, 37–48. doi: 10.2981/wlb.1996.006
Connell, J. H. (1980). Diversity and the coevolution of competitors, or the ghost of competition past. Oikos 35, 131–138. doi: 10.2307/3544421
Creel, S., and Creel, N. M. (1996). Limitation of African wild dogs by competition with larger carnivores. Conserv. Biol. 10, 526–538. doi: 10.1046/j.1523-1739.1996.10020526.x
Dollfus, and Dollfus, P. (2012). Transformation processes in nomadic pastoralism in Ladakh. J. Assoc. Nepal Himal. Stud. 32, 61–72. Available online at: https://digitalcommons.macalester.edu/himalaya/vol32/iss1/15/
Donadio, E., and Buskirk, S. W. (2006). Diet, morphology, and interspecific killing in carnivora. Am. Nat. 167, 524–536. doi: 10.1086/501033
Dray, S., and Dufour, A.-B. (2007). The ade4 package: implementing the duality diagram for ecologists. J. Stat. Softw. 22, 1–20. doi: 10.18637/jss.v022.i04
Dray, S., and Legendre, P. (2008). Testing the species traits-environment relationships: the fourth-corner problem revisited. Ecology 89, 3400–3412. doi: 10.1890/08-0349.1
Feldhamer, G. A., Thompson, B. C., and Chapman, J. A. (2003). Wild Mammals of North America: Biology, Management, and Conservation. Baltimore, MD: JHU Press.
Ferretti, F., Pacini, G., Belardi, I., ten Cate, B., Sensi, M., Oliveira, R., et al. (2021). Recolonizing wolves and opportunistic foxes: interference or facilitation? Biol. J. Linn. Soc. 132, 196–210. doi: 10.1093/biolinnean/blaa139
Floyd, T. J., Mech, L. D., and Jordan, P. A. (1978). Relating wolf scat content to prey consumed. J. Wildl. Manage. 42, 528–532. doi: 10.2307/3800814
Fox, J. L., Nurbu, C., and Chundawat, R. S. (1991). The mountain ungulates of Ladakh, India. Biol. Conserv. 58, 167–190. doi: 10.1016/0006-3207(91)90118-S
Garrott, R. A., Bruggeman, J. E., Becker, M. S., Kalinowski, S. T., and White, P. J. (2007). Evaluating prey switching in wolf–ungulate systems. Ecol. Appl. 17, 1588–1597. doi: 10.1890/06-1439.1
Ghoshal, A., Bhatnagar, Y. V., Mishra, C., and Suryawanshi, K. R. (2016). Response of the red fox to expansion of human habitation in the Trans-Himalayan mountains. Eur. J. Wildl. Res. 62, 131–136. doi: 10.1007/s10344-015-0967-8
Gómez-Ortiz, Y., Monroy-Vilchis, O., and Mendoza-Martínez, G. D. (2015). Feeding interactions in an assemblage of terrestrial carnivores in central Mexico. Zool. Stud. 54:e16. doi: 10.1186/s40555-014-0102-7
Gotelli, N., Hart, E., and Ellison, A. (2015). EcoSimR: Null Model Analysis for Ecological Data. R package version 0.1.0. Available online at: https://github.com/GotelliLab/EcoSimR/
Gotelli, N. J., and Ulrich, W. (2012). Statistical challenges in null model analysis. Oikos 121, 171–180. doi: 10.1111/j.1600-0706.2011.20301.x
Habib, B. (2007). Ecology of Indian Wolf (Canis lupus pallipes Sykes, 1831), and Modeling Its Potential Habitat in the Great Indian Bustard Sanctuary (Ph.D. Thesis), Maharashtra, India.
Habib, B., Jhala, Y. V., Lyngdoh, S., Shrotriya, S., and Reshamwala, S. R. (2021). Ecology and Conservation of Himalayan Wolf. Final Project Report. Dehradun, India: Wildlife Institute of India.
Handa, O. P. (1994). Tabo Monastery and Buddhism in Trans-Himalaya: A Thousand Years of Existence of the Tabo Chos-Khor. Shimla: Indus Publishing Company.
Hardin, G. (1960). The competitive exclusion principle. Science 131, 1292–1297. doi: 10.1126/science.131.3409.1292
Hayward, M. W., Henschel, P., O'Brien, J., Hofmeyr, M., Balme, G., and Kerley, G. I. H. (2006). Prey preferences of the leopard (Panthera pardus). J. Zool. 270, 298–313. doi: 10.1111/j.1469-7998.2006.00139.x
Hayward, M. W., and Kerley, G. I. H. (2005). Prey preferences of the lion (Panthera leo). J. Zool. 267, 309–322. doi: 10.1017/S0952836905007508
Hennelly, L. M., Habib, B., Modi, S., Rueness, E. K., Gaubert, P., and Sacks, B. N. (2021). Ancient divergence of Indian and Tibetan wolves revealed by recombination-aware phylogenomics. Mol. Ecol. 30, 6687–6700. doi: 10.1111/mec.16127
Henry, D. A. W., and Cumming, G. S. (2017). Can waterbirds with different movement, dietary and foraging functional traits occupy similar ecological niches? Landsc. Ecol. 32, 265–278. doi: 10.1007/s10980-016-0449-8
Holt, R. D., and Polis, G. A. (1997). A theoretical framework for intraguild predation. Am. Nat. 149, 745–764. doi: 10.1086/286018
Home, C., Pal, R., Sharma, R. K., Suryawanshi, K. R., Bhatnagar, Y. V., and Vanak, A. T. (2017). Commensal in conflict: Livestock depredation patterns by free-ranging domestic dogs in the Upper Spiti Landscape, Himachal Pradesh, India. Ambio 46, 655–666. doi: 10.1007/s13280-016-0858-6
Hunter, J., and Caro, T. (2008). Interspecific competition and predation in American carnivore families. Ethol. Ecol. Evol. 20, 295–324. doi: 10.1080/08927014.2008.9522514
Jackson, R., and Wangchuk, R. (2000). “People-wildlife conflicts in the trans-Himalaya,” in Management Planning Workshop for the Trans-Himalayan Protected Areas (Leh: Wildlife Institute of India), 1–10.
Jacobs, J. (1974). Quantitative measurement of food selection - a modification of the forage ratio and Ivlev's electivity index. Oecologia 14, 413–417. doi: 10.1007/BF00384581
Jaksic, F. M., Feinsinger, P., and Jimenez, J. E. (1996). Ecological redundancy and long-term dynamics of vertebrate predators in Semiarid Chile. Conserv. Biol. 10, 252–262. doi: 10.1046/j.1523-1739.1996.10010252.x
Jamtsho, Y., and Katel, O. (2019). Livestock depredation by snow leopard and Tibetan wolf: implications for herders' livelihoods in Wangchuck Centennial National Park, Bhutan. Pastoralism 9:1. doi: 10.1186/s13570-018-0136-2
Jethva, B. D., and Jhala, Y. V. (2004). Foraging ecology, economics and conservation of Indian wolves in the Bhal region of Gujarat, Western India. Biol. Conserv. 116, 351–357. doi: 10.1016/S0006-3207(03)00218-0
Jhala, Y. V. (1993). Predation on Blackbuck by Wolves in Velavadar National Park, Gujarat, India. Conserv. Biol. 7, 874–881. doi: 10.1046/j.1523-1739.1993.740874.x
Jones, M. E., and Barmuta, L. A. (1998). Diet overlap and relative abundance of sympatric dasyurid carnivores: a hypothesis of competition. J. Anim. Ecol. 67, 410–421. doi: 10.1046/j.1365-2656.1998.00203.x
Jumabay-Uulu, K., Wegge, P., Mishra, C., and Sharma, K. (2014). Large carnivores and low diversity of optimal prey: a comparison of the diets of snow leopards Panthera uncia and wolves Canis lupus in Sarychat-Ertash Reserve in Kyrgyzstan. Oryx 48, 529–535. doi: 10.1017/S0030605313000306
Kachroo, P., Sapru, B. L., and Dhar, U. (1977). Flora of Ladakh: an Ecological and Taxonomical Appraisal. Dehradun, India: Bishen Singh Mahendra Pal Singh publishers, Dehradun, India.
Karanth, K. U., and Sunquist, M. E. (2000). Behavioural correlates of predation by tiger (Panthera tigris), leopard (Panthera pardus) and dhole (Cuon alpinus) in Nagarahole, India. J. Zool. 250, 255–265. doi: 10.1111/j.1469-7998.2000.tb01076.x
Kumar, S., and Rahmani, A. (2000). Livestock depredation by wolves in the Great Indian Bustard Sanctuary, Nannaj (Maharashtra), India. J. Bombay Nat. Hist. Soc. 97, 340–348.
Kunkel, K., and Pletscher, D. H. (2001). Winter hunting patterns of wolves in and Near Glacier National Park, Montana. J. Wildl. Manage. 65, 520–530. doi: 10.2307/3803105
Linnell, J. D. C., and Strand, O. (2000). Interference interactions, coexistence and conservation of mammalian carnivores. Divers. Distrib. 6, 169–176. doi: 10.1046/j.1472-4642.2000.00069.x
Liu, B., and Jiang, Z. (2003). Diet composition of wolves Canis lupus in the northeastern Qinghai-Tibet Plateau, China. Acta Theriol. 48, 255–263. doi: 10.1007/BF03194165
Lyngdoh, S., Shrotriya, S., Goyal, S. P., Clements, H., Hayward, M. W., and Habib, B. (2014). Prey preferences of the snow leopard (Panthera uncia): regional diet specificity holds global significance for conservation. PLoS ONE 9:e88349. doi: 10.1371/journal.pone.0088349
Lyngdoh, S. B., Habib, B., and Shrotriya, S. (2020). Dietary spectrum in Himalayan wolves: comparative analysis of prey choice in conspecifics across high-elevation rangelands of Asia. J. Zool. 310, 24–33. doi: 10.1111/jzo.12724
MacNulty, D. R., Stahler, D. R., Wyman, C. T., Ruprecht, J., and Smith, D. W. (2016). The challenge of understanding Northern Yellowstone elk dynamics after wolf reintroduction. Yellowstone Sci. 24, 25–33. Available online at: https://qcnr.usu.edu/labs/macnulty_lab/files/MacNulty%20et%20al%202016a.pdf
McPhee, H. M., Webb, N. F., and Merrill, E. H. (2012). Hierarchical predation: wolf (Canis lupus) selection along hunt paths and at kill sites. Can. J. Zool. 90, 555–563. doi: 10.1139/z2012-021
Mech, L. D., and Boitani, L. (2003). Wolves: Behavior, Ecology, and Conservation. Chicago: University of Chicago Press. doi: 10.7208/chicago/9780226516981.001.0001
Meriggi, A., and Lovari, S. (1996). A review of wolf predation in Southern Europe: does the wolf prefer wild prey to livestock? J. Appl. Ecol. 33, 1561–1571. doi: 10.2307/2404794
Mishra, C. (2001). High Altitude Survival Conflicts Between Pastoralism and Wildlife in the Trans-Himalaya (Ph.D. Thesis).
Mishra, C., Bhatnagar, Y. V., and Suryawanshi, K. R. (2016). “Species Richness and Size Distribution of Large Herbivores in the Himalaya,” in The Ecology of Large Herbivores in South and Southeast Asia, eds. F. S. Ahrestani and M. Sankaran (Springer Science+Business Media Dordrecht), 89–97. doi: 10.1007/978-94-017-7570-0_3
Mishra, C., Prins, H. H. T., and Van Wieren, S. E. (2001). Overstocking in the Trans-Himalayan rangelands of India. Environ. Conserv. 28, 279–283. doi: 10.1017/S0376892901000297
Monterroso, P., Díaz-Ruiz, F., Lukacs, P. M., Alves, P. C., and Ferreras, P. (2020). Ecological traits and the spatial structure of competitive coexistence among carnivores. Ecology 101:e03059. doi: 10.1002/ecy.3059
Mukherjee, S., Goyal, S., and Chellam, R. (1994). Refined techniques for the analysis of Asiatic lion Panthera leo persica scats. Acta Theriol. 39, 425–430. doi: 10.4098/AT.arch.94-50
Namgail, T. (2009). Geography of Mammalian Herbivores in the Indian Trans-Himalaya: patterns and processes.
Namgail, T., Fox, J. L., and Bhatnagar, Y. V. (2007). Carnivore-caused livestock mortality in Trans-Himalaya. Environ. Manage. 39, 490–496. doi: 10.1007/s00267-005-0178-2
Namgail, T., Fox, J. L., and Bhatnagar, Y. V. (2009). Status and distribution of the Near Threatened Tibetan argali Ovis ammon hodgsoni in Ladakh, India: effect of a hunting ban. Oryx 43, 288–291. doi: 10.1017/S0030605308000264
Newsome, T. M., Boitani, L., Chapron, G., Ciucci, P., Dickman, C. R., Dellinger, J. A., et al. (2016). Food habits of the world's grey wolves. Mamm. Rev. 46, 255–269. doi: 10.1111/mam.12067
Nowak, S., Mysłajek, R. W., Kłosińska, A., and Gabryś, G. (2011). Diet and prey selection of wolves (Canis lupus) recolonising Western and Central Poland. Mamm. Biol. 76, 709–715. doi: 10.1016/j.mambio.2011.06.007
Paquet, P. C., Wierzchowski, J., and Callagan, C. (1996). “Effects of human activity on gray wolves in the Bow River Valley, Banff National Park, Alberta,” in A cumulative effects assesment and futures outlook for the Banff Bow Valley, eds J. Green, C. Pacas, S. Bayley, and L. Cornwell (Ottawa: Banff Bow Valley Study, Department of Canadian Heritage) p. 74–120.
Pianka, E. R. (1974). Niche overlap and diffuse competition. Proc. Natl. Acad. Sci. U.S.A. 71, 2141–2145. doi: 10.1073/pnas.71.5.2141
R Core Team. (2020). R: A language and environment for statistical computing. Version 3.6.3. R Foundation for Statistical Computing, Vienna, Austria. Available online at: https://www.R-project.org/.
Rawat, G. S., and Adhikari, B. S. (2005). Floristics and distribution of plant communities across moisture and topographic gradients in Tso Kar basin, Changtnang plateau, eastern Ladakh. Arctic, Antarct. Alp. Res. 37, 539–544. doi: 10.1657/1523-0430(2005)037[0539:FADOPC]2.0.CO;2
Reshamwala, H. S., Shrotriya, S., Bora, B., Lyngdoh, S., Dirzo, R., and Habib, B. (2018). Anthropogenic food subsidies change the pattern of red fox diet and occurrence across Trans-Himalayas, India. J. Arid Environ. 150, 15–20. doi: 10.1016/j.jaridenv.2017.12.011
Ridoux, V. (1994). The diets and dietary segregation of seabirds at the subantarctic Crozet Islands. Mar. Ornithol. 22, 1–192.
Ripple, W. J., and Beschta, R. L. (2005). Linking wolves and plants: aldo leopold on trophic cascades. Bioscience 55, 613–621. doi: 10.1641/0006-3568(2005)055[0613:LWAPAL]2.0.CO;2
Ripple, W. J., and Beschta, R. L. (2012). Trophic cascades in Yellowstone: the first 15 years after wolf reintroduction. Biol. Conserv. 145, 205–213. doi: 10.1016/j.biocon.2011.11.005
Ripple, W. J., Beschta, R. L., and Painter, L. E. (2015). Trophic cascades from wolves to alders in Yellowstone. For. Ecol. Manage. 354, 254–260. doi: 10.1016/j.foreco.2015.06.007
Ripple, W. J., Larsen, E. J., Renkin, R. A., and Smith, D. W. (2001). Trophic cascades among wolves, elk and aspen on Yellowstone National Park's northern range. Biol. Conserv. 102, 227–234. doi: 10.1016/S0006-3207(01)00107-0
Scharf, F. S., Juanes, F., and Rountree, R. A. (2000). Predator size-prey size relationships of marine fish predators: interspecific variation and effects of ontogeny and body size on trophic-niche breadth. Mar. Ecol. Prog. Ser. 208, 229–248. doi: 10.3354/meps208229
Schoener, T. W. (1982). The controversy over interspecific competition: despite spirited criticism, competition continues to occupy a major domain in ecological thought. Am. Sci. 70, 586–595.
Shahi, P. S. (1982). Status of the gray wolf (Canis lupus pallipes) in India: a preliminary survey. J. Bombay Nat. Hist. Soc. 79, 493–502.
Sharma, D. K., Maldonado, J. E., Jhala, Y. V., and Fleischer, R. C. (2004). Ancient wolf lineages in India. Proc. R. Soc. London. B 271, S1–S4. doi: 10.1098/rsbl.2003.0071
Sharma, S., Dutta, T., and Bhatnager, Y. V. (2007). Snow leopard in Himalaya: Resource Partitioning and Coexistence with Tibetan Wolf. in Felid Biology and Conservation, eds J. Hughes and R. Mercer (The Wildlife Conservation Research Unit, Oxford University), 87.
Shrotriya, S., Reshamwala, H. S., Mahar, N., Habib, B., Suhail, I., and Takpa, J. (2015). Distribution and Population Estimation of Ungulates in Changthang Region, Ladakh, Jammu and Kashmir, India. Dehradun: Wildlife Institute of India.
Sidorovich, V. E., Tikhomirova, L. L., and Jedrzejewska, B. (2003). Wolf Canis lupus numbers, diet and damage to livestock in relation to hunting and ungulate abundance in northeastern Belarus during 1990–2000. Wildl. Biol. 9, 103–111. doi: 10.2981/wlb.2003.032
Simberloff, D., and Dayan, T. (1991). The guild concept and the structure of ecological communities. Annu. Rev. Ecol. Syst. 22, 115–143. doi: 10.1146/annurev.es.22.110191.000555
Singh, N. J., Yoccoz, N. G., Lecomte, N., Côté, S. D., and Fox, J. L. (2010). Scale and selection of habitat and resources: Tibetan argali (Ovis ammon hodgsoni) in high-altitude rangelands. Can. J. Zool. 88, 436–447. doi: 10.1139/Z10-015
Sivy, K. J., Pozzanghera, C. B., Colson, K. E., Mumma, M. A., and Prugh, L. R. (2018). Apex predators and the facilitation of resource partitioning among mesopredators. Oikos 127, 607–621. doi: 10.1111/oik.04647
Spitz, J., Ridoux, V., and Brind'Amour, A. (2014). Let's go beyond taxonomy in diet description: testing a trait-based approach to prey-predator relationships. J. Anim. Ecol. 83, 1137–1148. doi: 10.1111/1365-2656.12218
Suryawanshi, K. R., Redpath, S. M., Bhatnagar, Y. V., Ramakrishnan, U., Chaturvedi, V., Smout, S. C., et al. (2017). Impact of wild prey availability on livestock predation by snow leopards. R. Soc. Open Sci. 4:170026. doi: 10.1098/rsos.170026
Ter Braak, C. J. F., Cormont, A., and Dray, S. (2012). Improved testing of species traits–environment relationships in the fourth-corner problem. Ecology 93, 1525–1526. doi: 10.1890/12-0126.1
Treves, A., Naughton-Treves, L., Harper, E. K., Mladenoff, D. J., Rose, R. A., Sickley, T. A., et al. (2004). Predicting human-carnivore conflict: a spatial model derived from 25 years of data on wolf predation on livestock. Conserv. Biol. 18, 114–125. doi: 10.1111/j.1523-1739.2004.00189.x
Ud Din, J., Ali, H., Ali, A., Younus, M., Mehmood, T., Norma-Rashid, Y., et al. (2017). Pastoralist-predator interaction at the roof of the world: Conflict dynamics and implications for conservation. Ecol. Soc. 22:32. doi: 10.5751/ES-09348-220232
Vanak, A. T., and Mukherjee, S. (2008). Identification of scat of Indian fox, jungle cat and golden jackal based on morphometrics. J. Bombay Nat. Hist. Soc. 105:212.
Wagner, C., Holzapfel, M., Kluth, G., Reinhardt, I., and Ansorge, H. (2012). Wolf (Canis lupus) feeding habits during the first eight years of its occurrence in Germany. Mamm. Biol. 77, 196–203. doi: 10.1016/j.mambio.2011.12.004
Wang, M.-S., Wang, S., Li, Y., Jhala, Y., Thakur, M., Otecko, N. O., et al. (2020). Ancient hybridization with an unknown population facilitated high-altitude adaptation of canids. Mol. Biol. Evol. 37, 2616–2629. doi: 10.1093/molbev/msaa113
Watts, S. M., McCarthy, T. M., and Namgail, T. (2019). Modelling potential habitat for snow leopards (Panthera uncia) in Ladakh, India. PLoS ONE 14:e0211509. doi: 10.1371/journal.pone.0211509
Werhahn, G., Kusi, N., Li, X., Chen, C., Zhi, L., Lázaro Martín, R., et al. (2019). Himalayan wolf foraging ecology and the importance of wild prey. Glob. Ecol. Conserv. 20:e00780. doi: 10.1016/j.gecco.2019.e00780
Werhahn, G., Senn, H., Ghazali, M., Karmacharya, D., Sherchan, A. M., Joshi, J., et al. (2018). The unique genetic adaptation of the Himalayan wolf to high-altitudes and consequences for conservation. Glob. Ecol. Conserv. 16:e00455. doi: 10.1016/j.gecco.2018.e00455
Wikenros, C. (2011). The Return of the Wolf: Effects on Prey, Competitors and Scavengers (Ph. D. thesis). Swedish University of Agricultural Sciences, Uppsala, Sweden.
Keywords: diet competition, himalayan wolf, Ladakh, prey selection, RLQ ordination
Citation: Shrotriya S, Reshamwala HS, Lyngdoh S, Jhala YV and Habib B (2022) Feeding Patterns of Three Widespread Carnivores—The Wolf, Snow Leopard, and Red Fox—in the Trans-Himalayan Landscape of India. Front. Ecol. Evol. 10:815996. doi: 10.3389/fevo.2022.815996
Received: 16 November 2021; Accepted: 14 June 2022;
Published: 06 July 2022.
Edited by:
Enrique Martínez-Meyer, National Autonomous University of Mexico, MexicoReviewed by:
Tariq Mahmood, Pir Mehr Ali Shah Arid Agriculture University, PakistanCopyright © 2022 Shrotriya, Reshamwala, Lyngdoh, Jhala and Habib. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bilal Habib, YmhAd2lpLmdvdi5pbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.