 Julio V. Schneider1,2
Julio V. Schneider1,2 Tanja Jungcurt1,3
Tanja Jungcurt1,3 Domingos Cardoso4
Domingos Cardoso4 André M. Amorim5,6
André M. Amorim5,6 Juraj Paule1,3
Juraj Paule1,3 Georg Zizka1,3*
Georg Zizka1,3*- 1Department of Botany and Molecular Evolution, Senckenberg Research Institute and Natural History Museum Frankfurt, Frankfurt am Main, Germany
- 2Entomology III, Department of Terrestrial Zoology, Senckenberg Research Institute and Natural History Museum Frankfurt, Frankfurt am Main, Germany
- 3Institute of Ecology, Evolution and Diversity, Goethe University Frankfurt, Frankfurt am Main, Germany
- 4National Institute of Science and Technology in Interdisciplinary and Transdisciplinary Studies in Ecology and Evolution (INCT IN-TREE), Instituto de Biologia, Universidade Federal da Bahia (UFBA), Salvador, Brazil
- 5Departamento de Ciências Biológicas, Universidade Estadual de Santa Cruz (UESC), Ilhéus, Brazil
- 6Herbário André Maurício Vieira de Carvalho, Centro de Pesquisas do Cacau (CEPEC), Comissão Executiva do Plano da Lavoura Cacaueira (CEPLAC), Itabuna, Brazil
Ochnaceae is a pantropical family with multiple transoceanic disjunctions at deep and shallow levels. Earlier attempts to unravel the processes that led to such biogeographic patterns suffered from insufficient phylogenetic resolution and unclear delimitation of some of the genera. In the present study, we estimated divergence time and ancestral ranges based on a phylogenomic framework with a well-resolved phylogenetic backbone to tackle issues of the timing and direction of dispersal that may explain the modern global distribution of Ochnaceae. The nuclear data provided the more robust framework for divergence time estimation compared to the plastome-scale data, although differences in the inferred clade ages were mostly small. While Ochnaceae most likely originated in West Gondwana during the Late Cretaceous, all crown-group disjunctions are inferred as dispersal-based, most of them as transoceanic long-distance dispersal (LDD) during the Cenozoic. All LDDs occurred in an eastward direction except for the SE Asian clade of Sauvagesieae, which was founded by trans-Pacific dispersal from South America. The most species-rich clade by far, Ochninae, originated from either a widespread neotropical-African ancestor or a solely neotropical ancestor which then dispersed to Africa. The ancestors of this clade then diversified in Africa, followed by subsequent dispersal to the Malagasy region and tropical Asia on multiple instances in three genera during the Miocene-Pliocene. In particular, Ochna might have used the South Arabian land corridor to reach South Asia. Thus, the pantropical distribution of Ochnaceae is the result of LDD either transoceanic or via land bridges/corridors, whereas vicariance might have played a role only along the stem of the family.
Introduction
Dispersal is amongst the most important biogeographical mechanisms shaping the distribution of plant species. In transcontinental range disjunctions, long-distance dispersal (LDD) has even become the more plausible a priori assumption compared to vicariance (de Queiroz, 2005; Renner et al., 2010; Christenhusz and Chase, 2013). Over the years, molecular divergence time estimations revealed that many disjunctions in angiosperms are simply too young to be explained by vicariance, including some of the iconic examples of Gondwanan vicariance such as the southern temperate Nothofagaceae and Proteaceae (Weston and Crisp, 1994; Knapp et al., 2005; Barker et al., 2007). Thus, vicariance is often confined to family level disjunctions, while at genus-level they are mostly inferred as dispersal-based (Renner, 2004).
If vicariance fails to explain transoceanic disjunctions, such distributions can be attributed either to dispersal via land connections between continents or direct dispersal across oceans. The latter may have been facilitated by island chains between continents (stepping-stone dispersal; e.g., Morley, 2003; Harbaugh and Baldwin, 2007; Kainulainen et al., 2017; Ahlstrand et al., 2019) and can be achieved by transport of diaspores in water (e.g., Gallaher et al., 2015), by wind (Muñoz et al., 2004) or animals (e.g., Nogales et al., 2012; Viana et al., 2016) or by rafting of seeds or plants on floating islands (Renner, 2004; van Duzer, 2004). There are many examples of pantropical distributions with trans-Pacific (e.g., Hernandiaceae, Michalak et al., 2010; Monimiaceae, Renner et al., 2010), trans-Atlantic (e.g., Leguminosae, Lavin et al., 2004; Sapotaceae, Bartish et al., 2011; Clusiaceae, Ruhfel et al., 2016; Melastomataceae, Veranso-Libalah et al., 2018) or dispersal across the Indian Ocean (e.g., Arecaceae, Baker and Couvreur, 2013; Rubiaceae, Kainulainen et al., 2017). We might expect some bias in the direction of transoceanic dispersal due to the predominant wind and sea currents (Renner, 2004; Ali and Huber, 2010), albeit over geological times such dispersal seems to have occurred in all directions across angiosperms (e.g., Renner, 2004; de Queiroz, 2005; Crisp et al., 2009; Christenhusz and Chase, 2013). However, more data are needed to reveal potential patterns across time strata and regions.
Besides direct transoceanic LDD, floristic interchange was also available via land connections, including routes that involved migration via the North Atlantic land bridge (NALB; e.g., Tiffney, 1985) and the Beringian land bridge (BLB, Tiffney and Manchester, 2001). The first connected Europe with North America at 45–50° northern latitude and existed during a short period around the Paleocene-Eocene boundary some 55 million years ago (Ma) (Tiffney, 1985; Morley, 2003). This was also the time of the Late Paleocene Thermal Maximum and Early Eocene Climatic Optimum when tropical elements existed at high-latitudes in the so-called boreotropical region. Thus, migration of (sub) tropical and warm-temperate plant species across the NALB was feasible during this period (>65 Ma to ca. 47 Ma; Dick and Pennington, 2019) and used to explain distributions such as in Leguminosae (Lavin and Luckow, 1993), Malpighiaceae (Davis et al., 2002), Meliaceae (Muellner et al., 2006), Rubiaceae (Antonelli et al., 2009; Smedmark et al., 2010), and Sapotaceae (Bartish et al., 2011). The BLB, in turn, was available for the dispersal of terrestrial organisms for a much more extended time from the Early Paleogene to the Pliocene. However, it remains doubtful whether this route was used by tropical elements due to its position at much higher latitudes with climatic conditions having been most likely unfavorable for megathermal species (Tiffney and Manchester, 2001).
The pantropical Ochnaceae (Malpighiales), which is the focus of this study, exhibit interesting geographical patterns that are largely expected to originate from LDD given the previous age estimates for the family (e.g., Xi et al., 2012; Schneider et al., 2017). Disjunct distributions in the family occur across all taxonomic ranks from the subfamily to species level, as well as remarkable differences in species diversity within clades and across continents (Amaral and Bittrich, 2014; Schneider et al., 2014). For example, the monotypic subfamily Medusagynoideae is endemic to the Seychelles, whereas Quiinoideae (4 genera, 46 spp.; Schneider and Zizka, 2016, 2017) are neotropical and Ochnoideae pantropical (29 genera, without accounting for the necessary nomenclatural changes in the poly-/paraphyletic Campylospermum Tiegh. and neotropical Sauvagesia L.; ca. 500 spp.; Amaral and Bittrich, 2014; Schneider et al., 2020, 2021). The latter is subdivided into four tribes (Schneider et al., 2014), with the monotypic Testuleeae confined to tropical Africa, Luxemburgieae being endemic to the Neotropics, and Sauvagesieae and Ochneae being pantropical. Sauvagesieae is the largest tribe in terms of number of genera and is predominantly neotropical, but also contains a SE Asian clade and one African endemic, the monotypic Fleurydora A.Chev. At the genus level, we observe four transoceanic disjunctions, three with an African-Malagasy-Asian distribution (Brackenridgea A.Gray, Campylospermum s.l., Ochna L.; all in subtribe Ochninae) and one with an American-African distribution [Sauvagesia s.l.; only one African species plus the weedy S. erecta L. occurring in the Neo- and Paleotropics, most likely introduced to the Old World by human transport (Amaral and Bittrich, 2014)]. Ochninae is by far the most species-rich clade, uniting about two thirds of the family’s diversity, and among Ochnoideae it is the only one with fleshy, conspicuous diaspores (in addition to the species-poor Malesian Euthemis Jack which has also fleshy but white fruits). All other genera of this subfamily have dry, partially also winged diaspores. Thus, dispersal capacity varies with fruit and seed morphology in Ochnaceae and most likely had some impact on the biogeography.
An earlier attempt to unravel the historical biogeography of Ochnaceae (Bissiengou, 2014) provided first insights into the timing of divergence events and the roles of vicariance vs. LDD. According to that study, Ochnaceae originated in the Neotropics and dispersed to the Old World on multiple instances using the NALB as well as transoceanic LDD. However, these inferences were based on a molecular dataset with insufficient resolution along the phylogenetic backbone of the family, particularly regarding some of the earlier divergence events in Sauvagesieae and among Ochninae, thus leaving unanswered biogeographically important questions such as how and in which direction LDD influenced the radiation of Ochninae. Moreover, the infrageneric sampling was insufficient to infer the ancestral range for several genera with disjunct distributions, including those for which delimitation and monophyly were unclear. Recently, phylogenomic studies based on nuclear loci and plastome data (Schneider et al., 2020, 2021) and by far with the most complete taxon sampling to date provided the required framework for tackling questions about the spatiotemporal diversification of Ochnaceae. Thus, the aim of this study is to obtain estimates of divergence time for Ochnaceae to uncover the ancestral range, the mechanisms that have guided its extant pantropical distribution, and the direction of its dispersal at deep and shallow times.
Materials and Methods
Taxon and Molecular Sampling
We used our previously published datasets obtained from target enrichment of nuclear loci and indirectly captured near-complete plastid genomes based on an Ochnaceae-specific bait kit (Schneider et al., 2020, 2021; Shah et al., 2021) but used a reduced taxon set to include only representative taxa that were needed for addressing our biogeographical questions. Reducing the number of taxa also facilitated the divergence time estimation, with far less computational time required for reaching convergence during the analyses (see below). We selected representative species of all genera of Ochnaceae following the most recent classification of Ochnaceae (Schneider et al., 2014). All non-monotypic tribes within Ochnoideae as well as the subfamilies are represented by a similar proportion of species (ca. 10%; Sauvagesieae with 25% but also with the highest number of genera). For species-rich genera or genera occurring in more than one biogeographical area as defined below, we included representatives of each major clade or area (Supplementary Table 1). This also refers to Campylospermum s.l. and Sauvagesia s.l. which were shown to be poly-/paraphyletic in recent studies (Schneider et al., 2020, 2021), both still with the necessary nomenclatural changes pending. In Campylospermum, only Clade B sensu Schneider et al. (2020) has a disjunct distribution in the present study. We created two datasets, one based on 46 nuclear loci from the family wide “FAM” dataset of Schneider et al. (2020; Philacra Dwyer, Quiina Aubl., and Rhytidanthera Tiegh. lacking), the other one for the plastome data (Rhabdophyllum Tiegh. lacking). Both comprised 59 species each including one outgroup but with slightly different species composition. Some species were represented by different individuals because of excessive missing data in one of the datasets (as outlined in Schneider et al., 2020, 2021; Supplementary Table 1). The nuclear alignment comprised 77,940 bp and the plastid alignment 117,923 bp. The latter was additionally used with sites removed at a 10% gap threshold, which had an alignment length of 33,636 bp. The above mentioned genera missing in the datasets are not expected to influence the outcome of the biogeographic analysis because they are nested in clades without variation in the here defined major geographical areas (see below). Molecular methods from DNA extraction to sequencing and assembly followed Schneider et al. (2020, 2021).
Divergence Time Estimation
We simultaneously inferred phylogenetic relationships and divergence times using a Bayesian Markov chain Monte Carlo approach as implemented in BEAST v2.48 (Bouckaert et al., 2014). The analyses were run separately for each dataset. The DNA sequence alignments were loaded into BEAUti with the site models linked, thereby allowing for the convergence of the analyses. Schneider et al. (2020) also demonstrated that both a supermatrix and multispecies coalescent analysis, respectively, converged on similar species trees. We used a GTR + gamma substitution model with four gamma categories and the rates estimated. We selected an uncorrelated relaxed clock with a lognormal distribution. A birth-death speciation prior was specified with the speciation rate parameter set to a range of 0–10,000 and the relative death rate to 0–1, both with a uniform distribution (e.g., Jin and Brown, 2018). The ucld.mean was set as a uniform prior allowing for a range of reasonable substitution rates in plants (0.000001–10) as recommended by S. Ho in Schuster et al. (2013). BEAST analyses were run on the CIPRES Science Gateway v3.3 (Miller et al., 2010) and XSEDE (Towns et al., 2014). Convergence of the runs and the effective sample sizes (ESS) were examined in Tracer v1.7 (Rambaut et al., 2018) after discarding the initial 20% of the generations as burn-in. The analyses were run for up to 400 million generations until the ESS were > 200 (for the nuclear dataset achieved after combining two independent runs). Mean node ages with their 95% highest posterior density (HPD) intervals were summarized and a maximum clade credibility (MCC) tree generated with TreeAnnotator.
In the absence of suitable fossils of Ochnaceae (for the reasoning why the published fossils assigned to Ochnaceae were not used, see Supplementary Material 1), the calibration of the molecular clock was achieved by setting three secondary calibrations for the crown group nodes of Ochnaceae (77.8 Ma, CI = 90.1–60.5 Ma), Ochnoideae (53.0 Ma, CI = 72.4–38.9 Ma) and Quiinoideae (18.1 Ma, CI = 38.7–4.3 Ma) based on the study of Xi et al. (2012) which included 11 species of Ochnaceae and 16 fossil calibrations across Malpighiales. Multiple simultaneous secondary calibrations accommodate better the uncertainty in age estimation (Sauquet et al., 2011; Sauquet, 2013; Schenk, 2016). The standard deviations of the secondary calibrations were chosen to mirror the CIs (i.e., 95% HPD) of the corresponding nodes in Xi et al. (2012) as recommended by Hipsley and Müller (2014). We chose a normal distribution for the secondary calibrations to reflect an equal distribution of the uncertainty across the mean age values.
We assessed the uncertainties in age estimates in the MCC trees of each dataset using infinite-sites plots (Yang and Rannala, 2006; Inoue et al., 2010). Infinite-sites plots compare node ages with the width of their associated 95% confidence interval and are a means to evaluate if the uncertainty in age estimates is due to insufficient DNA sequence data or due to calibration of the molecular clock (dos Reis et al., 2016). Given an infinite amount of sequence data, the points are expected to fall onto a straight line, thus showing a strong correlation. In such cases, the uncertainties in the posterior time estimates are largely due to uncertainties in the calibration. We used BEASTmasteR (Matzke, 2015) for extracting and plotting the mean ages and the width of the 95% HPD from each MCC tree and linear regression for statistics and fitting a regression line.
Ancestral Range Estimations
We estimated the ancestral geographical ranges of Ochnaceae using the maximum-likelihood approach implemented in BioGeoBEARS (Matzke, 2013a,b). An unconstrained dispersal-extinction-cladogenesis (DEC; Ree and Smith, 2008) model was used as well as a likelihood variant of the dispersal-vicariance-analysis (DIVA-like; Ronquist, 1997) with the maximum number of areas set to two. We initially also ran the BAYAREA-LIKE model, which is a simplified likelihood interpretation of the original full Bayesian model implemented in the BayArea program (Landis et al., 2013). However, model fit of BAYAREA-LIKE was poor, so we did not include it for further inferences. We refrained from using the implemented DEC+J model, which emphasizes “jump dispersal,” and statistical comparisons because of fundamental flaws as outlined in Ree and Sanmartín (2018). The estimations were performed with the almost fully supported MCC tree obtained from the divergence time analysis with the nuclear dataset only (with the outgroup pruned) because both the nuclear and plastid trees, respectively, inferred similar relationships and divergence times. Species were assigned to four geographical regions based on the global patterns of endemism in Ochnaceae: Neotropics (A), Africa (B), the Malagasy region (incl. Madagascar, the Seychelles, Comoros, Mascarenes; C), and Asia (incl. Malesia and tropical NE Australia; D). We inferred LDD events by visualizing changes in the most likely ancestral range between nodes on the MCC tree.
Results
Divergence Times
Differences in age estimates between the three datasets (nuclear, plastid, trimmed plastid) are moderate (Table 1 and Supplementary Figures 1–3). The plastid dataset with gap-based trimming generally inferred slightly younger age estimates for the selected 14 clades compared to the un-trimmed plastid dataset (mean difference = 1.8 ± 1.3 Ma; Supplementary Tables 2, 3). Comparing all three datasets, the mean differences between the minimum and maximum age estimates for the crown groups across the selected clades was 3.6 ± 2.7 Ma, for the stem groups 3.4 ± 1.7 Ma. The strongest difference was observed for Luxemburgia A.St.-Hil. (10.9 Ma; Table 1). The infinite-sites plots (Supplementary Figure 4) reveal a stronger correlation between the posterior mean ages and the width of their 95% HPD for the nuclear dataset (R2 = 0.85) than for the plastid datasets (R2 = 0.76 for the untrimmed and R2 = 0.74 for the trimmed), thus indicating that adding more DNA sequence data is less likely to increase the precision of posterior age estimates in the nuclear relative to the plastid datasets, although all three would still benefit from more sequence data. Conversely, the slope (w) is slightly lower for the nuclear dataset (w = 0.44) than for the plastid datasets (w = 0.45 and 0.49, respectively), which means that the uncertainty in age estimates increases more slowly with increasing divergence time in the nuclear dataset. Given the indicators from the infinite-sites plots and the overall moderate differences between the datasets, we refer below to the divergence times obtained with the nuclear dataset.
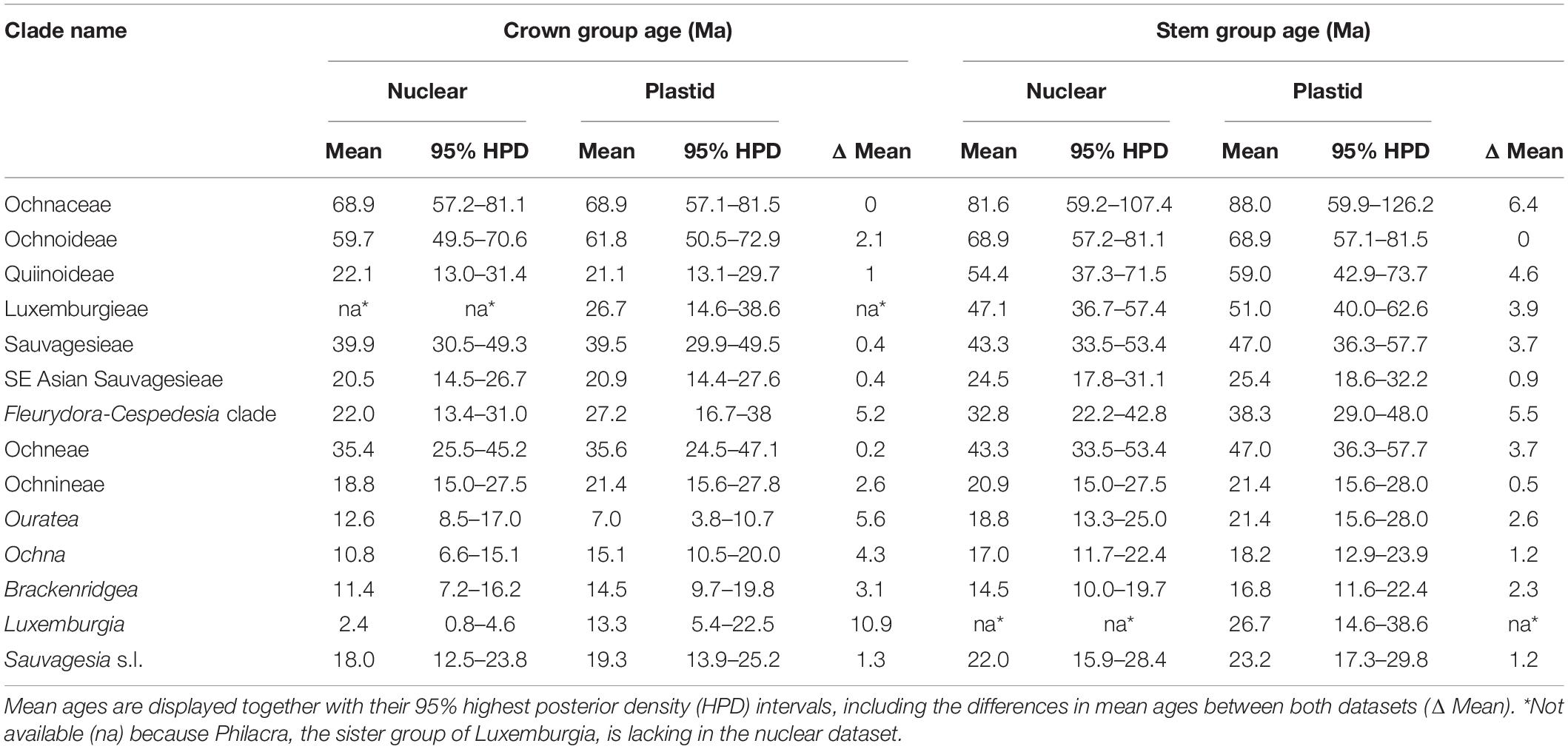
Table 1. Crown and stem group ages of major clades of Ochnaceae as inferred from divergence time estimation (i.e., the maximum clade credibility trees with mean heights obtained from BEAST analysis) based on nuclear and plastid genomic data.
The crown Ochnaceae was dated between 81.1 Ma and 57.2 (HPD; mean: 68.9 Ma), which is the temporal origin of Ochnoideae and the lineage comprised by the most recent common ancestor (MRCA) of Medusagynoideae and Quiinoideae. The stem of Luxemburgieae and the MRCA of Ochneae and Sauvagesieae were dated to 57.4–36.7 Ma (mean: 47.1 Ma), whereas the crown ages of the two major tribes of Ochnoideae, i.e., Ochneae and Sauvagesieae, were estimated 45.2–25.5 Ma (mean: 35.4 Ma) and 49.3–30.5 Ma old (mean: 39.9 Ma), respectively. Among Sauvagesieae, the biogeographically important clade that unites Fleurydora, Cespedesia Goudot, Godoya Ruiz & Pav., and Krukoviella A.C.Sm. (Rhytidanthera not included in the nuclear dataset) has a crown age of 31.0–13.4 Ma (mean: 22.0 Ma), and the clade of all SE Asian representatives of this tribe have a stem age of 31.1–17.8 Ma (mean: 24.5 Ma). The by far most species-rich clade Ochninae originated at 27.5–15.0 Ma (mean: 20.9 Ma). Brackenridgea and Ochna, two genera of Ochninae with disjunct distributions between Africa, Madagascar and Asia, have crown ages of 16.2–7.2 Ma (mean: 11.4 Ma) and 15.1–6.6 Ma (mean: 10.8 Ma), respectively.
Ancestral Geographic Ranges
Both the DEC (AIC: 119.2) and DIVA (AIC: 119.1) analyses showed a similar model fit to the data and inferred largely similar ancestral ranges for Ochnaceae. Therefore, we refer below primarily to the DEC analysis and report from the DIVA analysis only in case of substantial conflict. According to the unconstrained estimation of the historical biogeography under the DEC model (Figure 1), the per-area probabilities indicate ambiguous or weakly supported ancestral ranges for the deepest nodes, e.g., the MRCA of Ochnaceae and to some degree also of Ochnoideae and the clade of Quiinoideae and Medusagynoideae. Yet there is some support for a neotropical-African (44%), a neotropical-Malagasy (29%) or a neotropical origin (20% probability) of Ochnaceae (Figure 1, node 59). For Ochnoideae and the clade of Quiinoideae and Medusagynoideae, a widespread neotropical-African or neotropical-Malagasy distribution is the most probable ancestral range with 59% and 58%, respectively (Figure 1, nodes 63 and 60). For the shallower nodes, the estimated ancestral ranges received stronger support with probabilities mostly > 75% (Supplementary Table 4 for a full list of area probabilities and Supplementary Figure 5 for the corresponding node numbers).

Figure 1. Ancestral range estimation of Ochnaceae. The biogeographic reconstruction under the unconstrained DEC model (max. number of areas = 2; dispersal = 0.0045, extinction = 0.0023) visualizes multiple LDD events which in part can be linked with putative past land bridges (North Atlantic land bridge, Afro-Asian land bridge) or stepping-stone dispersal between South America and Africa (vertical bars show the periods when they were supposed to be extant). The pie charts indicate alternative ancestral geographical ranges and their probabilities. The pie charts on the descendant branches show the ranges immediately after the speciation event, whereas the pie charts on the nodes display the range changes along branches before the speciation event. Numbers below branches are for the ancestral range that received that highest probability (only given for selected nodes), numbers above branches correspond to the node numbers in Supplementary Figure 5 and Supplementary Table 4 containing the complete list of probabilities. Species were assigned to the four distribution areas A to D as illustrated on the inset map and the respective tip ranges (colored squares with letter codes at tips). The legend below the inset map displays the color codes for each area, including the area combinations as retrieved in the analysis. The classification system for Ochnaceae is based on Schneider et al. (2014); Lux., Luxemburgieae; MED., Medusagynoideae; QUII, Quiinoideae; T., Testuleeae.
Within Ochnoideae the two major tribes, Sauvagesieae and Ochneae, have a neotropical (93%, node 67) and neotropical-African (77%, node 85) origin, respectively. The deeper nodes in Ochneae, including the MRCA of Ochninae (99%, node 89), share a widespread neotropical-African ancestral range. The cladogenetic event following the origin of Ochninae led to a neotropical (Ouratea Aubl.) and an African clade, i.e., the MRCA of the remaining genera of Ochninae. This African clade showed several range expansions during the Late Miocene to Pliocene reaching the Malagasy region up to five times independently, once in Campylospermum, once in Brackenridgea, and two to three times in Ochna. In each of these genera, there was also one LDD event from Africa to Asia during the Miocene/Pliocene. All inferred LDD events were in eastward direction. The only exception is the clade uniting the SE Asian Sauvagesieae, which is supposed to have taken a westward route from the Neotropics. LDD from the Neotropics to Africa also occurred once in Sauvagesieae during the Miocene (node 69, Figure 1) and once in the Eocene/Oligocene in Ochneae leading to Lophira Banks ex C.F.Gaertn (Figure 2).
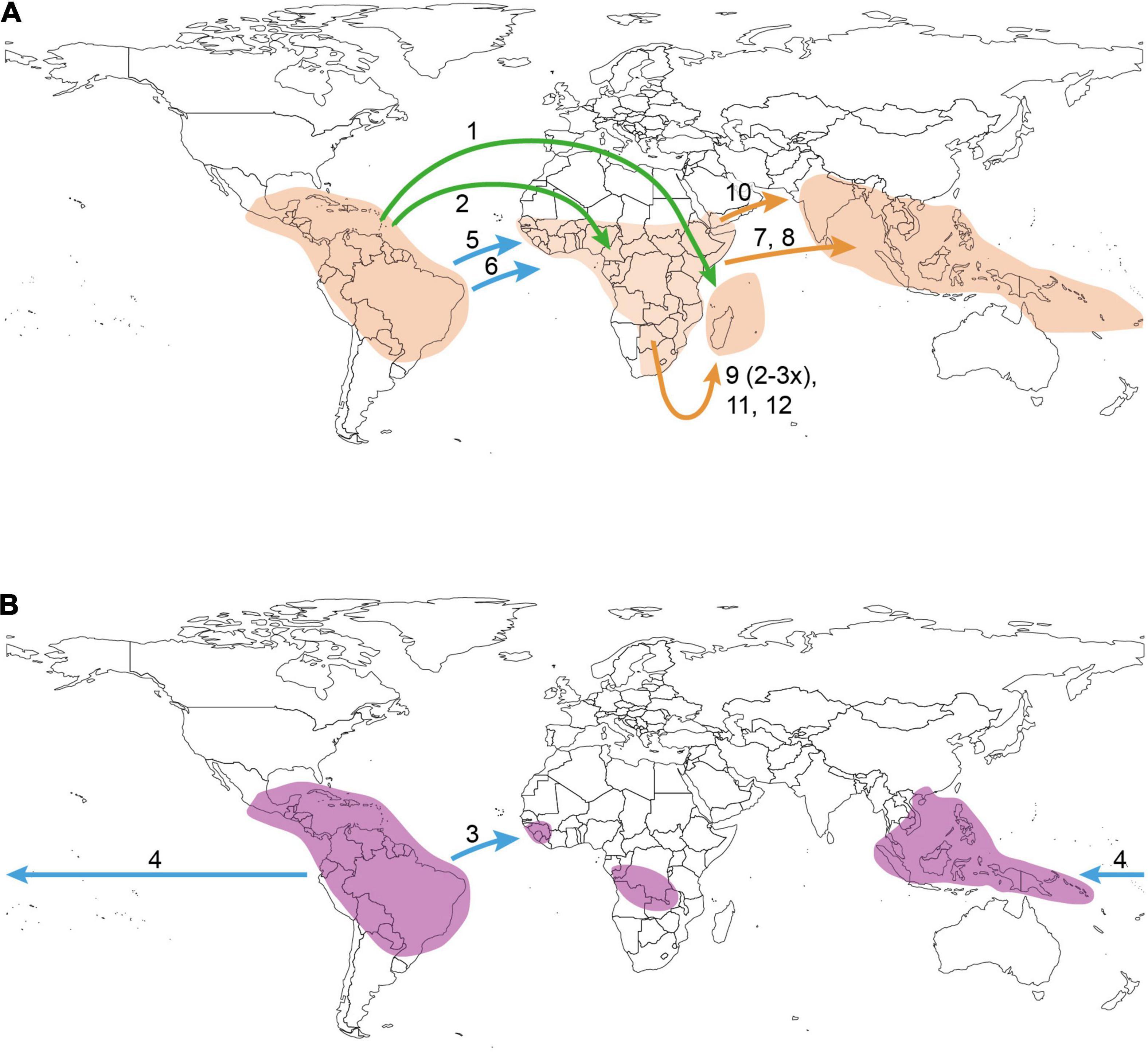
Figure 2. Distribution areas together with the inferred most probable long-distance dispersal (LDD) events of Ochnaceae in Medusagynoideae, Testuleeae, and Ochneae (A) and Sauvagesieae (B). LDD events and their direction are indicated by arrows; the arrow color indicates the time periods of LDD from old (green, Paleogene) to mid-term (blue) and young (orange = Miocene/Pliocene) events. The numbers at the arrows correspond to the LDD events leading to (1) Medusagyne; (2) Testulea; (3) Fleurydora; (4) SE Asian Sauvagesieae; (5) Lophira; (6) African Ochninae; (7) Asian Brackenridgea; (8) Campylospermum serratum; (9) Malagasy Ochna (multiple LDD); (10) Asian Ochna; (11) Malagasy Brackenridgea and (12) Campylospermum. Sauvagesieae is species-poor in Africa with one LDD event explaining the distribution of the monotypic Fleurydora (3) in West Africa (the approximate distribution area of Sauvagesia africana is also given on the map, but this species was not included in the present study; the African occurrences of S. erecta are not indicated). The LDD of the African Ochninae (6) was only inferred by the DIVA-like model.
Discussion
Divergence Time Estimation, Uncertainty, and Impact on Biogeographic Inference
Our divergence time analyses show that the nuclear and plastid genomic sequence data converged on similar age estimates for deep and shallow nodes and that the bulk of the generic diversity of extant Ochnaceae emerged during the Oligocene-Miocene. The latter is consistent with the temporal framework provided in earlier studies that were based on a much more reduced molecular sampling (Schneider and Zizka, 2017; Schneider et al., 2017). However, compared to previous studies, there are some differences in the stem age of Ochnaceae. We inferred mean ages of 81.6 (95% HPD: 107.4–59.2 Ma) for the nuclear and 88.0 Ma (95% HPD: 126.2–59.9 Ma) for the plastid data, respectively, whereas Xi et al. (2012) estimated an age of 101.5 Ma (106.7–96.4 Ma) for the MRCA of Ochnaceae and the clusioids, and Magallón et al. (2015) inferred mean stem group ages of 101.7–92.2 Ma (95% CI: 103.2–83.4 Ma; summarized over different analyses) for Ochnaceae. In a taxonomically narrower study, Ruhfel et al. (2016) obtained mean stem group ages of 115.4 and 120.6 Ma (95% HPD: 124.9–104.2 Ma) for the clusioids, which is the same as for Ochnaceae, depending on where the fossil Paleoclusia Crepet and Nixon (Crepet and Nixon, 1998) was placed for calibration. Thus, our study covered the 95% HPD ranges of these studies but differed in the lower range limit and the mean age. This reflects some of the uncertainty in our divergence time estimation, an issue that was tackled for the biogeographic inference as outlined below.
The first comprehensive molecular phylogenetic analysis of Ochnaceae was based on five DNA regions with a concatenated alignment length of about 4 kb (Bissiengou, 2014; Schneider et al., 2014). In contrast, the nuclear and plastid datasets used in the present study have alignment lengths between 33 and 117 kb. Benefits from this increased molecular sampling are an improved phylogenetic resolution (Schneider et al., 2020, 2021) and a reduced uncertainty in the determination of branch lengths and relative node ages (Inoue et al., 2010; dos Reis et al., 2016). According to the infinite-sites plots, the nuclear dataset produced posterior time estimates with the lowest uncertainty. Despite the robust phylogenetic framework, uncertainty in absolute ages predominantly arises from uncertainty in the calibrations. Generally, fossils provide the most reliable data for the calibration of a molecular clock, when carefully chosen (Parham et al., 2012), while secondary calibrations are considered problematic (e.g., Graur and Martin, 2004; Sauquet, 2013; Schenk, 2016) and only acceptable as a last resort when fossils are lacking, such as often happens with tropical taxa (Sauquet, 2013; Hipsley and Müller, 2014). The major concern with secondary calibrations is that error associated with the primary calibration is compounded into them, thus producing age estimates of dubious reliability. It has been argued that the error is even exacerbated when secondary calibrations are taken from studies that were based on single fossil calibration points, as in earlier studies (Graur and Martin, 2004), or from studies targeting higher taxonomic levels and more distant taxa, i.e., when rates differ between the clade of the primary calibration and the study group (Hipsley and Müller, 2014; Powell et al., 2020). These last points are, however, considered less relevant sources of error in our study because we chose the secondary calibration points from an analysis targeting inter-familial relationships in Malpighiales based on plastome-scale data and 16 fossils (Xi et al., 2012). Also, the primary calibration in the clusioid sister group represents a clade of woody species which are expected to have similar substitution rates as in Ochnaceae. Ochnaceae are predominantly woody shrubs or small trees (some taller trees in Brackenridgea, Lophira, and Testulea Pellegr.) and only a few species of Sauvagesia are herbaceous. So, we expect little impact on evolutionary rates arising from differences in generation time (see Smith and Donoghue, 2008).
To account for the increased uncertainty associated with the secondary calibrations, we set the age priors for the calibration points to reflect the confidence intervals of the original age estimates. Hence, we discuss biogeographic scenarios based on rather broad confidence intervals including some extension that mirrors the differences in deep and shallow nodes compared to the ages of nodes in the study of the primary calibrations. For example, the divergence times for the deepest nodes are slightly younger (ca. 8–14 Ma) compared to those from previous studies, whereas more recent ones differ only marginally, but rather tend to be older compared to previous age estimates. For example, crown-group Ochneae has an age of ca. 30 Ma in Xi et al. (2012; therein represented by Elvasia, Lophira and Ochna) but 35.6–33.8 Ma in the present study. The same is observed for Quiinoideae—one of our calibrated nodes—which are 3–4 Ma older than in Xi et al. (2012; therein as Quiinaceae).
Discrepancies in divergence times have also recently been attributed to branch lengths artifacts arising from the acceleration of substitution rates following the K/Pg mass extinction (Berv and Field, 2018). Such mass extinction is supposed to lead to drastically reduced population sizes which entail higher evolutionary rates. Although angiosperms seem to have been less affected by the K/Pg mass extinction (Silvestro et al., 2015), crown-group Ochnaceae began to radiate around the K/Pg boundary according to the inferred divergence times which may well be indicative of some prior depletion of generic diversity. While difficult to reliably assess with the broad confidence intervals of our divergence time estimates, a significant increase in origination rates in the aftermath of the K/Pg mass extinction would be in agreement with the general pattern observed in angiosperms (Silvestro et al., 2015).
The Ancestral Range of Ochnaceae
Our reconstruction of the biogeographic history of the pantropical Ochnaceae revealed a high amount of uncertainty regarding its ancestral range. However, a neotropical or West Gondwanan (comprising South America and Africa) origin is the most likely scenario given the time frame obtained from molecular dating (Xi et al., 2012; Magallón et al., 2015) and the likelihoods of a purely neotropical or neotropical-African ancestral range in the present study. The alternative scenario of a neotropical and Malagasy origin without Africa is not supported by geological evidence because the Malagasy region was part of East Gondwana and separated from West Gondwana between 175 and 140 Ma (McLoughlin, 2001; Morley, 2003; Upchurch, 2008) well before the more likely estimates for the origin of Ochnaceae. A neotropical-African ancestral range was also inferred for the clusioid sister clade (Ruhfel et al., 2016). With the slightly older stem group age inferred for the clusioids in that study and, thus, indirectly for Ochnaceae (which is also covered by our confidence intervals), their origin falls in a time when South America and Africa had begun to separate but were still connected in some parts (the separation of both continents lasted from 137 to 105 Ma; McLoughlin, 2001). Nonetheless, vicariance might have occurred only along the stem of Ochnaceae because the crown-group age interval is too young for any vicariance scenario associated with the separation of South America and Africa. While this is in contrast with other predominantly tropical plant families that have genera with origins rooted in the Gondwanan vicariance (e.g., Mello-Silva et al., 2011; Cai et al., 2016), a LDD explanation for the post-Gondwanan biogeographic history of many other transoceanic clades has been increasingly suggested (e.g., Bartish et al., 2011; Bardon et al., 2016; Dupin et al., 2017). Generally, the time lag between the stem and crown Ochnaceae as well as the potential impact of the K/Pg mass extinction leave a gap that impedes any further inferences about the initial biogeographic history of the family.
Transoceanic Dispersal in Ochnaceae
Both the African and Malagasy components at the crown node of Ochnaceae largely result from the two deep nodes with long branches that lead to the monospecific Medusagynoideae (an endemic of the Seychelles) and Testuleeae (from central-west Africa). Such monospecific lineages branching off in deep time are challenging because they might not only be the result of low diversification rates (i.e., depauperons; Donoghue and Sanderson, 2015) but remnants of formerly more diverse and widespread clades, thus potentially containing a large portion of hidden biogeographic history (e.g., Clayton et al., 2009; Sanmartín and Meseguer, 2016). However, given the time frame for the split of Ochnoideae from the MRCA of Medusagynoideae and Quiinoideae, the most likely scenario involves range expansion by LDD from the Neotropics to Africa along the branch subtending Ochnoideae and leading to Testulea (assuming a neotropical origin as inferred from the probability for the stem of this subfamily) as well as to the Malagasy region along the branch leading to Medusagyne Baker. When Ochnoideae diverged from Medusagynoideae and neotropical Quiinoideae at the end of the Cretaceous or beginning of the Paleogene, East and West Gondwana were long separated and Africa had severed from South America. The Seychelles rafted together with India since the segregation of both from East Gondwana 135 Ma until 66 Ma when they split from the Indian plate (Chatterjee et al., 2013). Therefore, dispersal across the western Indian Ocean to the Malagasy region during the Paleocene-Eocene is the most likely explanation for Medusagyne. This might have occurred directly from Africa or by stepping-stone dispersal via Madagascar or the Indian plate with subsequent survival only on the Seychelles, routes that were inferred frequently for many plant groups (e.g., Strijk et al., 2012; Stride et al., 2014; Kainulainen et al., 2017). There are multiple ways that these taxa (Testuleeae, Medusagynoideae) could have taken to reach Africa. The warm period around the Paleocene-Eocene boundary, i.e., the Late Paleocene Thermal Maximum and Early Eocene Climatic Optimum, allowed the expansion of tropical vegetation toward higher latitudes in the northern hemisphere, thereby opening the boreotropical route with the NALB connecting the New and Old World (Tiffney, 1985; Bissiengou, 2014). Hence, both Testulea and Medusagyne could be the result of dispersal from the Neotropics to Africa through that route. While the boreotropics hypothesis is largely plausible for many extant tropical transoceanic plant clades that were once diverse in the northern hemisphere (Lavin and Luckow, 1993; Herendeen et al., 2022), it is difficult to verify in Ochnaceae because of the lack of a fossil record in support of its former occurrence in the boreotropical flora. An alternative route is via a stepping-stone connection between both continents across the Walvis Ridge and the Rio-Grande Rise available during large parts of the Paleocene-Eocene (Morley, 2003). This would have allowed dispersal via island hopping, as for example suggested for early branching Asteraceae (Katinas et al., 2013) or Meliaceae (Muellner-Riehl and Rojas-Andrés, 2021), although direct dispersal through rafting on floating islands (Renner, 2004; van Duzer, 2004) cannot be ruled out. For the ancestral lineages of Medusagynoideae, we might also assume a western route from across the Americas and the BLB to Asia and further on to the Seychelles. However, it is doubtful whether megathermal taxa had used this route because the BLB was situated at much higher latitudes and provided far less favorable climatic conditions compared to the NALB (Tiffney and Manchester, 2001).
Apart from Testuleeae, the other tribes of Ochnoideae have a neotropical (Luxemburgieae, Sauvagesieae) or neotropical-African ancestral range (Ochneae). While in the first all divergence events until the Oligocene were exclusively within the New World, the deeper nodes of Ochneae until the Early Miocene (i.e., from crown Ochneae to Ochninae) were all inferred as widespread neotropical-African based on the DEC model. However, here is some conflict between the DEC and DIVA analysis because the latter suggests a neotropical ancestral range for the nodes predating crown Ochninae (node 89, Supplementary Figure 6). The DIVA-based inference provides an intuitively more reasonable explanation, assuming independent dispersals from the New World to Africa leading to Lophira and the ancestor of the African Ochninae, in contrast to the assumption of a continuous widespread distribution between the Eocene-Oligocene boundary and the Late to Mid Miocene made by the DEC model. The cladogenetic event that led to the African Lophira happened during the Paleocene-Oligocene, at a time when the boreotropical route for overland dispersal between the Americas and Eurasia/Africa was not available anymore. Thus, direct transoceanic LDD seems to be the most likely route, although island hopping via the Walvis Ridge or Rio-Grande Rise across the Atlantic was still feasible at that time.
Within Sauvagesieae, two dispersals out of the Neotropics occurred, both around the Oligocene-Miocene boundary, and both in taxa with capsular fruits and most likely winged seeds as an ancestral character state (Schneider et al., 2014). Our data suggest transoceanic LDD to Africa for the West-African endemic and monospecific Fleurydora, and there is only a marginal probability for stepping-stone dispersal across the Atlantic from the New World to Africa during the inferred time. The shift to more mesic conditions with forested vegetation in West Africa by the Middle Oligocene (Jacobs, 2004) may have facilitated the establishment of this taxon after LDD, a scenario supported by Melastomateae from closed habitats which colonized Africa via LDD during a similar time frame and which adapted to open habitats during subsequent aridification (Veranso-Libalah et al., 2018). The second transoceanic dispersal founded the SE Asian clade which gave rise to six endemic genera that spread across continental SE Asia and the Indomalayan archipelago. This coincided with the Early Miocene change from seasonally wet to perhumid climates in the Sunda region (Crayn et al., 2015; Morley, 2018) which might have facilitated the successful colonization of SE Asia by the megathermal taxa after the arrival from the Neotropics. Equally important must have been the presence of sandy, nutrient-poor soils because both the SE Asian taxa as well as many of their neotropical allies generally grow on such sites (Amaral, 1991). Successful trans-Pacific dispersal to and colonization of Pacific islands and the Indomalayan archipelago was also inferred for Chiococceae (Rubiaceae) during the same time (Manns et al., 2012), supporting the view that the Early Miocene climate change in this region was crucial for the establishment of elements arriving from the Neotropics. Dispersal by wind or rafting via India is unlikely because dispersal from Africa via Madagascar to India was interrupted well before the appearance of the SE Asian clade. Moreover, Sauvagesieae are poorly represented in Africa (one species each in Sauvagesia and the monotypic Fleurydora) and the MRCAs are all neotropical. With the Early Paleogene land bridges unavailable at the time of dispersal, interoceanic LDD across the Pacific is the only reasonable scenario. The prevailing directionality of the equatorial currents would have allowed rafting over the Pacific Ocean to Malesia. However, with the low velocity of these currents, such a traverse would have taken many months (Gillespie et al., 2011), rendering unlikely the survival under the exposition to seawater, albeit not impossible for a seed. There are also adaptations of seeds to dispersal by water in some Sauvagesieae (Blastemanthus Planch., Wallacea Spruce ex Benth. & Hook.f.) which grow in seasonally inundated rainforests (Amaral, 1991). Nonetheless, it is unlikely that such adaptations would have allowed the crossing of the ocean given the small size of these seeds and the sustained exposition to seawater. Bird dispersal also seems unlikely given the small seeds and the lack of adaptations for epizoochorous dispersal as well as the known flyways (see Gillespie et al., 2011). Thus, rafting across the Pacific still seems the most reasonable explanation.
The genus Sauvagesia s.l. nicely illustrates how taxonomic delimitation can have a profound impact on biogeographic inferences. Earlier concepts of a broadly defined and pantropical Sauvagesia (Sastre, 1971a; Amaral, 1991, 2006) suggested a Malesian origin of the genus with subsequent dispersal to Africa and the Neotropics (Sastre, 1971b). However, molecular phylogenetic studies revealed that the Malesian elements, i.e., the former genera Indovethia Boerl. and Neckia Korth., are independent from Sauvagesia s.l. and are part of the SE Asian clade within Sauvagesieae (Schneider et al., 2014, 2020, 2021). Although we did not include the African species S. africana (Baill.) Bamps in the present study, Sauvagesia s.l. undoubtedly has a neotropical origin. A better sampling here within paraphyletic Sauvagesia s.l. would also help us further explore that its two independent lineages have each radiated as ecologically and biogeographically distinct. Sauvagesia s.str. is mostly comprised of widespread, herbaceous species with a broad ecological amplitude and predilection to disturbance-prone settings. The other Sauvagesia lineage has mostly radiated in the open, grassy and rocky vegetation on the mountaintops of the Brazilian Espinhaço range (e.g., Cardoso and Harley, 2015; Queiroz-Lima et al., 2018), where many other extraordinarily diverse plant radiations are often composed of more geographically confined or narrowly endemic species (e.g., Ribeiro et al., 2014; Rando et al., 2016; Alcantara et al., 2018; Vasconcelos et al., 2020; Rapini et al., 2021).
Transoceanic dispersal is also suggested for some of the disjunct distributions in Ochninae. Following the DIVA analysis, the African Ochninae originated from dispersal from the Neotropics to Africa during the Miocene at a time when land bridges or island connections were unavailable, contrary to earlier assumptions of a Gondwanan vicariance (Amaral, 1991). Africa must have been reached by transoceanic LDD followed by an extensive radiation, including the colonization of savannas. This radiation occurred at a time when savannas spread and diversified (Cerling et al., 1997; Jacobs, 2004; Simon et al., 2009; Maurin et al., 2014), which might have opened up new opportunities for LDD, for example, via drier corridors. Alternatively, as inferred for Melastomateae (Veranso-Libalah et al., 2018), the adaptation to drier open habitats post-dated the establishment in Africa. Three of the five African genera of Ochninae (thus, only those that also occur in East Africa) further dispersed to the Malagasy region and Asia. All these dispersal events occurred in a time period between about 10 and 5 Ma, when floristic exchange overland of megathermal angiosperms between Africa and Asia was feasible via the Arabian Peninsula as suggested, for example, for Annonaceae (Onstein et al., 2019). This corridor became available after the closure of the Thetyan seaway at about 20 Ma in the Early Miocene (Patnaik, 2015) and due to more favorable conditions with woodland and grassland vegetation (Kingston and Hill, 1999; Gilbert et al., 2014). Therefore, dispersal via this land connection is a reasonable explanation for the presence of these taxa in Asia. A further argument in favor of the overland dispersal hypothesis is the existence of one Ochna species on the southern Arabian Peninsula. Ochna is also the genus with the westernmost distribution in Asia, occurring in India and Sri Lanka. So, it is plausible to assume a more contiguous African-Asian distribution area for this genus during that period. The disruption of this distribution area across western South Asia may have resulted from the increasing drought during the Late Miocene to Pliocene (Morley, 2018). Although it seems reasonable that Ochna had used this way to reach Asia, our analysis does not preclude an alternative scenario which involves LDD directly across the Indian Ocean as has been inferred for other fleshy-fruited taxa (e.g., Bartish et al., 2011; de Wilde et al., 2011).
The other two disjunct genera, Brackenridgea and Campylospermum s.l., are largely confined to areas far more east, i.e., to mainland SE Asia and the Indomalayan archipelago (Kanis, 1968), thus having much larger distribution gaps. Therefore, we might also assume LDD across the Indian Ocean, either directly or via the “Lemurian” stepping-stones (Schatz, 1996; Meimberg et al., 2001; Warren et al., 2010). The last must have occurred via island hopping because there is no geological evidence for a continuous land connection between these regions (Schatz, 1996). The stepping stones included Madagascar, the Seychelles and perhaps also some other islands of the Indian Ocean that emerged during periods of low sea-level or due to volcanic activity since the Late Miocene (Emerick and Duncan, 1982; Warren et al., 2010; Weigelt et al., 2016). The presence of Ochninae on several of these islands further supports this hypothesis, which has also been invoked to explain similar disjunctions in other plant genera (e.g., Yuan et al., 2005; de Wilde et al., 2011). However, our ancestral range analysis indicates that the Asian taxa do not descend from the taxa of the Malagasy region in Brackenridgea and Campylospermum.
The strongest biogeographic link of the Malagasy biodiversity is with Africa (Yoder and Nowak, 2006) as is true for many plant families (Buerki et al., 2013), including Ochnaceae. Earlier dispersal to Madagascar via rafting from Africa (e.g., on floating islands) was facilitated by the eastward sea currents that prevailed from the Eocene to the Oligocene. Later, with the beginning of the Miocene, these currents shifted and were perennially directed toward Africa, thereby impeding the seaway to the Malagasy region (Ali and Huber, 2010). Given the Miocene-Pliocene ages of the dispersal to the Malagasy region, it is unlikely that Ochninae have used this way. Alternatively, they have been transported by birds, which are supposedly the main dispersers of these fleshy-fruited taxa (Amaral, 1991). This is a commonly invoked explanation (e.g., Renner, 2004; Kainulainen et al., 2017). However, Africa is comparatively poor in frugivorous bird species (Kissling et al., 2009), and few Malagasy species are actually frugivorous and none of them is migratory (Hawkins and Goodman, 2003; Albert-Daviaud et al., 2018). This paucity of frugivorous avian dispersal suggests that such dispersal events have occurred only sporadically, which is also reflected in the high degree of endemism (Kainulainen et al., 2017).
Interestingly, transoceanic LDD in Ochnaceae is not confined to fleshy-fruited taxa. Both major dispersal units of Ochnoideae—fleshy drupelets in Ochninae and capsular or samaroid to nut-like fruits, in part with winged seeds, in Lophira and Sauvagesieae (see Schneider et al., 2014)—were able to disperse via transoceanic LDD. The multiple colonization events of the Malagasy region from Africa by Ochninae (at least three in Ochna and one each in Brackenridgea and Campylospermum) and potentially further on to Asia confirm that such events occurred sufficiently frequent as is also documented by the hundreds of disjunct plant genera for which transoceanic exchange of diaspores between Africa, the Malagasy region and Asia is likely (e.g., Thorne, 1973; Schatz, 1996; Renner, 2004; Yuan et al., 2005; Yoder and Nowak, 2006; de Wilde et al., 2011; Strijk et al., 2012; Kainulainen et al., 2017; Razafimandimbison et al., 2020). Whether dispersal occurred the way back to Africa in Ochna, as suggested in one instance by our data, remains to be corroborated with a more complete taxon sampling.
Conclusion
Ochnaceae is another example of a pantropically distributed plant clade for which continental disjunctions are too young to be explained by vicariance. The here inferred LDD events involved either direct dispersal across oceans or via land connections or stepping stones, all but one in eastward direction. Two taxa might have used the NALB and the boreotropical route to reach Africa and the Malagasy region from the Neotropics, and some Ochninae could have used the Afro-Asian corridor via the Arabian Peninsula on their way to Asia. At least two of the Asian lineages of the Ochninae genera resulted from independent dispersal events from Africa, not from the Malagasy region. The latter was colonized at least three times independently from Africa in Ochna, confirming the importance of Africa as a source of dispersal to the Malagasy region. However, due to the limited intrageneric taxon sampling in Ochninae in the present study, especially in Ochna, the number and direction of dispersal events associated with the colonization of the Malagasy region and Asia as well as the radiation and dispersal within the Malagasy region remain to be uncovered. Thus, future research should focus on genus-level reconstructions of the historical biogeography with a comprehensive taxon sampling of these target taxa.
Here we have revealed new insights into the understanding of Ochnaceae interchange across biogeographic regions. Beyond further exploring the impact of geography (e.g., geographic barriers) on the historical biogeography within Ochnaceae, a denser, more complete taxon sampling would also enable us to understand how ecology has shaped the evolutionary history of the family. Lessons from globally distributed plant clades that are ecologically confined within biomes suggest the ease with which lineages can transcend geographic barriers (e.g., Lavin et al., 2004; Crisp et al., 2009; Thiv et al., 2011; Gagnon et al., 2019; Ringelberg et al., 2020), i.e., for such groups it seems to be “easier to move than to evolve” (Donoghue, 2008). More biogeographically realistic models have been developed (Landis et al., 2021a,b) that explicitly model the evolutionary accessibility of distinct environments (biomes) that create geographical opportunity (Edwards and Donoghue, 2013), i.e., permitting to reveal when lineages switch between biomes depending on the temporal availability and geographical connectivity of biomes (Landis et al., 2021a,b). Thus, such models are hoped to better describe how phylogenetic biome conservatism, evolutionary biome switches, and geographic (vicariance) processes shaped the Ochnaceae diversification history across the global biogeographic regions.
Data Availability Statement
Publicly available datasets were analyzed in this study. This data can be found here: http://www.ncbi.nlm.nih.gov/bioproject/602196. The DNA sequence alignments, tree files, and area coding generated and used for the ancestral area analysis are available from: doi: 10.5281/zenodo.5654525.
Author Contributions
JS and GZ designed the research. JP, JS, and TJ carried out the data analyses. JS, DC, and AA contributed to the collection of plant material. JS performed the molecular experiments and drafted the manuscript with contributions from all authors.
Funding
This study was supported by grants from the Deutsche Forschungsgemeinschaft (DFG; ZI 557/14-1), the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq; Research Productivity Fellowship, grants nos. 308244/2018-4 and 312404/2018-2) and the Fundação de Amparo à Pesquisa do Estado da Bahia (FAPESB; grant no. APP0037/2016). Open access publication was funded by the Goethe University Frankfurt (institutional membership).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We are grateful to the herbaria BR, CEPEC, FR, GB, L, LZ, MO, NY, P, U, and WAG, as well as the Botanischer Garten und Botanisches Museum Berlin-Dahlem and Royal Botanic Garden Edinburgh for the permission to use their specimens; to GenXPro for the permission to use their lab facilities; to Kristen Porter-Utley for providing leaf material; and to Pedro Lage Vianna, Roberto Salas, Adriana Queiroz-Lima, Cássia Bitencourt, and Quézia Cavalcante for assistance during field work. Brazilian accessions of Ochnaceae used in this study are all registered into the Sistema Nacional de Gestão do Patrimônio Genético e do Conhecimento Tradicional Associado (SisGen) under Cadastro de Remessa N° R391A8F.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.813336/full#supplementary-material
Supplementary Figure 1 | Maximum clade credibility tree obtained from BEAST analysis with divergence times of Ochnaceae based on the nuclear loci. Numbers on branches are posterior probabilities (PP) < 1 (all other nodes had PP = 1). The bars at nodes are the 95% HPD ranges (mean heights). The circles on nodes (A–C) indicate the secondary calibration points as obtained from Xi et al. (2012). The geological time scale includes the epochs for the periods of Paleogene, Neogene and Quaternary (Paleo., Paleocene; Oligoc., Oligocene). The classification system for Ochnaceae is based on Schneider et al. (2014); Elvas., Elvasiinae; Lux., Luxemburgieae; MED., Medusagynideae; QUII, Quiinoideae; T., Testuleeae.
Supplementary Figure 2 | Maximum clade credibility tree obtained from BEAST analysis with divergence times of Ochnaceae based on the plastome data (alignment length: 117,923 bp). Numbers on branches are posterior probabilities (PP) < 1 (all other nodes had PP = 1). The bars at nodes are the 95% HPD ranges (mean heights). The circles on nodes (A–C) indicate the secondary calibration points as obtained from Xi et al. (2012). The classification system for Ochnaceae is based on Schneider et al. (2014); Elvas., Elvasiinae; Lux., Luxemburgieae; MED., Medusagynideae; QUII, Quiinoideae; T., Testuleeae.
Supplementary Figure 3 | Maximum clade credibility tree obtained from BEAST analysis with divergence times of Ochnaceae based on the plastome data with sites removed at a threshold of 10% gaps (alignment length: 33,636 bp). Numbers on branches are posterior probabilities (PP) < 1 (all other nodes had PP = 1). The bars at nodes are the 95% HPD ranges (mean heights). The circles on nodes (A–C) indicate the secondary calibration points as obtained from Xi et al. (2012). The classification system for Ochnaceae is based on Schneider et al. (2014); Elvas., Elvasiinae; Lux., Luxemburgieae; MED., Medusagynideae; QUII, Quiinoideae; T., Testuleeae.
Supplementary Figure 4 | Infinite-sites plot for the nuclear and plastid data of Ochnaceae. Each point corresponds to an internal node of the maximum clade credibility tree. The 95% HPD width is the difference of the 97.5 and 2.5% limits. The slope indicates the increase in uncertainty in the posterior HPD for every million years of species divergence.
Supplementary Figure 5 | Maximum clade credibility tree obtained from BEAST analysis with node numbers. The node numbers refer to Supplementary Table 4.
Supplementary Figure 6 | Ancestral range estimation of Ochnaceae based on the Dispersal-Vicariance Analysis (DIVA-like). The pie charts indicate alternative ancestral geographical ranges and their probabilities. Numbers above branches correspond to the node numbers in Supplementary Figure 5. Species were assigned to the four distribution areas A–D as illustrated on the inset map and the respective tip ranges (colored squares with letter codes at tips). The classification system for Ochnaceae is based on Schneider et al. (2014); Lux., Luxemburgieae, MED., Medusagynoideae, QUII, Quiinoideae, T., Testuleeae.
References
Ahlstrand, N. I., Verstraete, B., Hassemer, G., Dunbar-Co, S., Hoggard, R., Meudt, H. M., et al. (2019). Ancestral range reconstruction of remote oceanic island species of Plantago (Plantaginaceae) reveals differing scales and modes of dispersal. J. Biogeogr. 46, 706–722. doi: 10.1111/jbi.13525
Albert-Daviaud, A., Perillo, S., and Stuppy, W. (2018). Seed dispersal syndromes in the Madagascan flora: the unusual importance of primates. Oryx 52, 418–426. doi: 10.1017/S0030605317001600
Alcantara, S., Ree, R. H., and Mello-Silva, R. (2018). Accelerated diversification and functional trait evolution in Velloziaceae reveal new insights into the origins of the campos rupestres exceptional floristic richness. Ann. Bot. 122, 165–180. doi: 10.1093/aob/mcy063
Ali, J. R., and Huber, M. (2010). Mammalian biodiversity on Madagascar controlled by ocean currents. Nature 463, 653–656. doi: 10.1038/nature08706
Amaral, M. C. E. (2006). Inclusion of Sinia in Sauvagesia (Ochnaceae). Novon 16, 1–2. doi: 10.3417/1055-3177(2006)16[1:IOSISO]2.0.CO;2
Amaral, M. C. E., and Bittrich, V. (2014). “Ochnaceae,” in The Families and Genera of Vascular Plants, Vol. 11, ed. K. Kubitzki (Heidelberg: Springer), 253–268. doi: 10.1007/978-3-642-39417-1_19
Antonelli, A., Nylander, J. A. A., Persson, C., and Sanmartín, I. (2009). Tracing the impact of the Andean uplift on Neotropical plant evolution. Proc. Natl. Acad. Sci. U.S.A. 106, 9749–9754. doi: 10.1073/pnas.0811421106
Baker, W. J., and Couvreur, T. L. P. (2013). Global biogeography and diversification of palms sheds light on the evolution of tropical lineages. I. Historical biogeography. J. Biogeogr. 40, 274–285. doi: 10.1111/j.1365-2699.2012.02795.x
Bardon, L., Sothers, C., Prance, G. T., Malé, P. J. G., Xi, Z., Davis, C. C., et al. (2016). Unraveling the biogeographical history of Chrysobalanaceae from plastid genomes. Amer. J. Bot. 103, 1089–1102. doi: 10.3732/ajb.1500463
Barker, N. P., Weston, P. H., Rutschmann, F., and Sauquet, H. (2007). Molecular dating of the “Gondwanan” plant family Proteaceae is only partially congruent with the timing of the break-up of Gondwana. J. Biogeogr. 34, 2012–2027. doi: 10.1111/j.1365-2699.2007.01749.x
Bartish, I. V., Antonelli, A., Richardson, J. E., and Swenson, U. (2011). Vicariance or long-distance dispersal: historical biogeography of the pantropical subfamily Chrysophylloideae (Sapotaceae). J. Biogeogr. 38, 177–190. doi: 10.1111/j.1365-2699.2010.02389.x
Berv, J. S., and Field, D. J. (2018). Genomic signature of an avian Lilliput Effect across the K-Pg extinction. Syst. Biol. 67, 1–13. doi: 10.1093/sysbio/syx064
Bissiengou, P. (2014). Systematics, Evolution and Historical Biogeography of the Family Ochnaceae with Emphasis on the Genus Campylospermum. Dissertation thesis. Wageningen: Wageningen University.
Bouckaert, R., Heled, J., Kühnert, D., Vaughan, T., Wu, C.-H., Xie, D., et al. (2014). BEAST 2: a software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 10:e1003537. doi: 10.1371/journal.pcbi.1003537
Buerki, S., Devey, D. S., Callmander, M. W., Phillipson, P. B., and Forest, F. (2013). Spatio-temporal history of the endemic genera of Madagascar. Bot. J. Linn. Soc. 171, 304–329. doi: 10.1111/boj.12008
Cai, L., Xi, Z., Peterson, K., Rushworth, C., Beaulieu, J., and Davis, C. C. (2016). Phylogeny of Elatinaceae and the tropical Gondwanan origin of the Centroplacaceae (Malpighiaceae. Elatinaceae) clade. PLoS One 11:e0161881. doi: 10.1371/journal.pone.0161881
Cardoso, D. B. O. S., and Harley, R. M. (2015). Sauvagesia paganuccii (Ochnaceae), a new species endemic to campo rupestre vegetation of Bahia. Brazil. Syst. Bot. 40, 776–781. doi: 10.1600/036364415X689230
Cerling, T. E., Harris, J. M., MacFadden, B. J., Leakey, M. G., Quade, J., Eisenmann, V., et al. (1997). Global vegetation change through the Miocene/Pliocene boundary. Nature 389, 153–158. doi: 10.1038/38229
Chatterjee, S., Goswami, A., and Scotese, C. R. (2013). The longest voyage: tectonic, magmatic, and paleoclimatic evolution of the Indian plate during its northward flight from Gondwana to Asia. Gondwana Res. 23, 238–267. doi: 10.1016/j.gr.2012.07.001
Christenhusz, M. J., and Chase, M. W. (2013). Biogeographical patterns of plants in the Neotropics – dispersal rather than plate tectonics is most explanatory. Bot. J. Linn. Soc. 171, 277–286. doi: 10.1111/j.1095-8339.2012.01301.x
Clayton, J. W., Soltis, P. S., and Soltis, D. E. (2009). Recent longdistance dispersal overshadows ancient biogeographical patterns in a pantropical Angiosperm family (Simaroubaceae. Sapindales). Syst. Biol. 58, 395–410. doi: 10.1093/sysbio/syp041
Crayn, D. M., Costion, C., and Harrington, M. G. (2015). The Sahul–Sunda floristic exchange: dated molecular phylogenies document Cenozoic intercontinental dispersal dynamics. J. Biogeogr. 42, 11–24. doi: 10.1111/jbi.12405
Crepet, W. L., and Nixon, K. C. (1998). Fossil Clusiaceae from the Late Cretaceous (Turonian) of New Jersey and implications regarding the history of bee pollination. Amer. J. Bot. 85, 1122–1133. doi: 10.2307/2446345
Crisp, M. D., Arroyo, M. T. K., Cook, L. G., Gandolfo, M. A., Jordan, G. J., McGlone, M. S., et al. (2009). Phylogenetic biome conservatism on a global scale. Nature 458, 754–756. doi: 10.1038/nature07764
Davis, C. C., Bell, C. D., Mathews, S., and Donoghue, M. J. (2002). Laurasian migration explains Gondwanan disjunctions: evidence from Malpighiaceae. Proc. Natl. Acad. Sci. U.S.A. 99, 6833–6837. doi: 10.1073/pnas.102175899
de Queiroz, A. (2005). The resurrection of oceanic dispersal in historical biogeography. Trends Ecol. Evol. 20, 68–73. doi: 10.1016/j.tree.2004.11.006
de Wilde, J. J. F. E., Hughes, M., Rodda, M., and Thomas, D. C. (2011). Pliocene intercontinental dispersal from Africa to Southeast Asia highlighted by the new species Begonia afromigrata (Begoniaceae). Taxon 60, 1685–1692. doi: 10.1002/tax.606013
Dick, C. W., and Pennington, R. T. (2019). History and geography of neotropical tree diversity. Ann. Rev. Ecol. Evol. 50, 279–301. doi: 10.1146/annurev-ecolsys-110617-062314
Donoghue, M. J. (2008). A phylogenetic perspective on the distribution of plant diversity. Proc. Natl. Acad. Sci. U.S.A. 105, 11549–11555. doi: 10.1073/pnas.0801962105
Donoghue, M. J., and Sanderson, M. J. (2015). Confluence, synnovation, and depauperons in plant diversification. New Phytol. 207, 260–274. doi: 10.1111/nph.13367
dos Reis, M., Donoghue, P. C. J., and Yang, Z. (2016). Bayesian molecular clock dating of species divergences in the genomics era. Nat. Rev. Genet. 17, 71–80. doi: 10.1038/nrg.2015.8
Dupin, J., Matzke, N. J., Särkinen, T., Knapp, S., Olmstead, R., Bohs, L., et al. (2017). Bayesian estimation of the global biogeographic history of the Solanaceae. J. Biogeogr. 44, 887–899. doi: 10.1111/jbi.12898
Edwards, E. J., and Donoghue, M. J. (2013). Is it easy to move and easy to evolve? Evolutionary accessibility and adaptation. J. Exp. Bot. 64, 4047–4052. doi: 10.1093/jxb/ert220
Emerick, C. M., and Duncan, R. A. (1982). Age progressive volcanism in the Comoros Archipelago, western Indian Ocean and implications for Somali plate tectonics. Earth Planet. Lett. 60, 415–428. doi: 10.1016/0012-821X(82)90077-2
Gagnon, E., Ringelberg, J. J., Bruneau, A., Lewis, G. P., and Hughes, C. E. (2019). Global succulent biome phylogenetic conservatism across the pantropical Caesalpinia group (Leguminosae). New Phytol. 222, 1994–2008. doi: 10.1111/nph.15633
Gallaher, T., Callmander, M. W., Buerki, S., and Keeley, S. C. (2015). A long distance dispersal hypothesis for the Pandanaceae and the origins of the Pandanus tectorius complex. Mol. Phylogenet. Evol. 83, 20–32. doi: 10.1016/j.ympev.2014.11.002
Gilbert, C. C., Bibi, F., Hill, A., and Beech, M. J. (2014). Early guenon from the late Miocene Baynunah Formation, Abu Dhabi, with implications for cercopithecoid biogeography and evolution. Proc. Natl. Acad. Sci. U.S.A. 111, 10119–10124. doi: 10.1073/pnas.1323888111
Gillespie, R. G., Baldwin, B. G., Waters, J. M., Fraser, C. I., Nikula, R., and Roderick, G. K. (2011). Long-distance dispersal: a framework for hypothesis testing. Trends Ecol. Evol. 27, 47–56. doi: 10.1016/j.tree.2011.08.009
Graur, D., and Martin, W. (2004). Reading the entrails of chickens: molecular timescales of evolution and the illusion of precision. Trends Genet. 20, 80–86. doi: 10.1016/j.tig.2003.12.003
Harbaugh, D. T., and Baldwin, B. G. (2007). Phylogeny and biogeography of the sandalwoods (Santalum, Santalaceae): repeated dispersals throughout the Pacific. Amer. J. Bot. 94, 1028–1040. doi: 10.3732/ajb.94.6.1028
Hawkins, A. F. A., and Goodman, S. M. (2003). “Introduction to the birds,” in The Natural History of Madagascar, eds S. M. Goodman and J. P. Benstead (Chicago, IL: University of Chicago Press), 1019–1044.
Herendeen, P. S., Cardoso, D. B. O. S., Herrera, F., and Wing, S. L. (2022). Fossil papilionoids of the Bowdichia clade (Leguminosae) from the Paleogene of North America. Amer. J. Bot. 109, 130–150. doi: 10.1002/ajb2.1808
Hipsley, C. A., and Müller, J. (2014). Beyond fossil calibrations: realities of molecular clock practices in evolutionary biology. Front. Genet. 5:138. doi: 10.3389/fgene.2014.00138
Inoue, J., Donoghue, P. C. J., and Yang, Z. (2010). The impact of the representation of fossil calibrations on Bayesian estimation of species divergence times. Syst. Biol. 59, 74–89. doi: 10.1093/sysbio/syp078
Jacobs, B. F. (2004). Palaeobotanical studies from tropical Africa: relevance to the evolution of forest, woodland, and savanna biomes. Phil. Transact. Roy. Soc. London B 359, 1573–1583. doi: 10.1098/rstb.2004.1533
Jin, Y., and Brown, R. P. (2018). Partition number, rate priors, and unreliable divergence times in Bayesian phylogenetic dating. Cladistics 34, 568–573. doi: 10.1111/cla.12223
Kainulainen, K., Razafimandimbison, S. G., Wikström, N., and Bremer, B. (2017). Island hopping, long-distance dispersal and species radiation in the western Indian Ocean: historical biogeography of the Coffeeae alliance (Rubiaceae). J. Biogeogr. 44, 1966–1979. doi: 10.1111/jbi.12981
Katinas, L., Crisci, J. V., Hoch, P., Tellería, M. C., and Apodaca, M. J. (2013). Trans-oceanic dispersal and evolution of early composites (Asteraceae). Perspect. Pl. Ecol Evol. Syst. 15, 269–280. doi: 10.1016/j.ppees.2013.07.003
Kingston, J. D., and Hill, A. (1999). “Late Miocene palaeoenvironments in Arabia: a synthesis,” in Fossil Vertebrates of Arabia: With Emphasis on the Late Miocene Faunas, Geology, and Palaeoenvironments of the Emirate of Abu Dhabi, eds P. J. Whybrow and A. Hill (New Haven, CT: Yale University Press), 389–407.
Kissling, W. D., Böhning-Gaese, K., and Jetz, W. (2009). The global distribution of frugivory in birds. Global Ecol. Biogeogr. 18, 150–162. doi: 10.1111/j.1466-8238.2008.00431.x
Knapp, M., Stockier, K., Havell, D., Delsuc, F., Sebastiani, F., and Lockhart, P. J. (2005). Relaxed molecular clock provides evidence for long-distance dispersal of Nothofagus (southern beech). PLoS Biol. 3:e14. doi: 10.1371/journal.pbio.0030014
Landis, M. J., Edwards, E. J., and Donoghue, M. J. (2021a). Modeling phylogenetic biome shifts on a planet with a past. Syst. Biol. 70, 86–107. doi: 10.1093/sysbio/syaa045
Landis, M. J., Eaton, D. A. R., Clement, W. L., Park, B., Spriggs, E. L., Sweeney, P., et al. (2021b). Joint phylogenetic estimation of geographic movements and biome shifts during the global diversification of Viburnum. Syst. Biol. 70, 67–85. doi: 10.1093/sysbio/syaa027
Landis, M. J., Matzke, N. J., Moore, B. R., and Huelsenbeck, J. P. (2013). Bayesian analysis of biogeography when the number of areas is large. Syst. Biol. 62, 789–804. doi: 10.1093/sysbio/syt040
Lavin, M. J., and Luckow, M. (1993). Origins and relationships of tropical North America in the context of the boreotropics hypothesis. Amer. J. Bot. 80, 1–14. doi: 10.1002/j.1537-2197.1993.tb13761.x
Lavin, M., Schrire, B., Lewis, G., Pennington, T., Delgado-Salinas, A., Thulin, M., et al. (2004). Metacommunity process rather than continental tectonic history better explains geographically structured phylogenies in legumes. Phil. Trans. R. Soc. Lond. B 359, 1509–1522. doi: 10.1098/rstb.2004.1536
Magallón, S., Gómez-Acevedo, S., Sánchez-Reyes, L. L., and Hernández-Hernández, T. (2015). A metacalibrated time-tree documents the early rise of flowering plant phylogenetic diversity. New Phytol. 207, 437–453. doi: 10.1111/nph.13264
Manns, U., Wikström, N., Taylor, C. M., and Bremer, B. (2012). Historical biogeography of the predominantly neotropical subfamily Cinchonoideae (Rubiaceae): into or out of America? Int. J. Pl. Sci. 173, 261–286. doi: 10.1086/663971
Matzke, N. J. (2013a). BioGeoBEARS: Biogeography With Bayesian (and Likelihood) Evolutionary Analysis in R Scripts. R package, version 1.2.2. Available online at: https://github.com/nmatzke/BioGeoBEARS (accessed February 10, 2021).
Matzke, N. J. (2013b). Probabilistic historical biogeography: new models for founder-event speciation, imperfect detection, and fossils allow improved accuracy and model-testing. Front. Biogeogr. 5:242–248. doi: 10.21425/F5FBG19694
Matzke, N. J. (2015). BEASTmasteR: R Tools for Automated Conversion of NEXUS Data to BEAST2 XML Format, for Fossil tip-Dating and Other Uses. Instructions at PhyloWiki. Available online at: http://phylo.wikidot.com/beastmaster (accessed March 09, 2021).
Maurin, O., Davies, T. J., Burrows, J. E., Daru, B. H., Yessoufou, K., Muasya, A. M., et al. (2014). Savanna fire and the origins of the ‘underground forests’ of Africa. New Phytol. 204, 201–214. doi: 10.1111/nph.12936
McLoughlin, S. (2001). The breakup history of Gondwana and its impact on pre-Cenozoic floristic provincialism. Aust. J. Bot. 49, 271–300. doi: 10.1071/BT00023
Meimberg, H., Wistuba, A., Dittrich, P., and Heubl, G. (2001). Molecular phylogeny of Nepenthaceae based on cladistic analysis of plastid trnK intron sequence data. Plant Biol. 3, 164–175. doi: 10.1055/s-2001-12897
Mello-Silva, R., Santos, D. Y. A. C., Salatino, M. L. F., Motta, L. B., Cattai, M. B., Sasaki, D., et al. (2011). Five vicarious genera from Gondwana: the Velloziaceae as shown by molecules and morphology. Ann. Bot. 108, 87–102. doi: 10.1093/aob/mcr107
Michalak, I., Zhang, L.-B., and Renner, S. S. (2010). Trans-Atlantic, trans-Pacific and trans-Indian Ocean dispersal in the small Gondwanan Laurales family Hernandiaceae. J. Biogeogr. 37, 1214–1226. doi: 10.1111/j.1365-2699.2010.02306.x
Miller, M. A., Pfeiffer, W., and Schwartz, T. (2010). “Creating the CIPRES Science Gateway for inference of large phylogenetic trees,” in Proceedings of the Gateway Computing Environments Workshop (GCE), 14 November 2010, (New Orleans, LA), 1–8. doi: 10.1109/GCE.2010.5676129
Morley, R. J. (2003). Interplate dispersal paths for megathermal angiosperms. Perspect. Pl. Ecol. Evol. Syst. 6, 5–20. doi: 10.1078/1433-8319-00039
Morley, R. J. (2018). Assembly and division of the South and South-East Asian flora in relation to tectonics and climate change. J. Trop. Ecol. 34, 209–234. doi: 10.1017/S0266467418000202
Muellner, A. N., Savolainen, V., Samuel, R., and Chase, M. W. (2006). The mahogany family “out-of-Africa”: divergence time estimation, global biogeographic patterns inferred from plastid rbcL DNA sequences, extant, and fossil distribution of diversity. Mol. Phylogenet. Evol. 40, 236–250. doi: 10.1016/j.ympev.2006.03.001
Muellner-Riehl, A. N., and Rojas-Andrés, B. M. (2021). Biogeography of Neotropical Meliaceae: geological connections, fossil and molecular evidence revisited. Braz. J. Bot. doi: 10.1007/s40415-021-00770-4
Muñoz, J., Felicísimo, A. M., Cabezas, F., Burgaz, A. R., and Martínez, I. (2004). Wind as a long-distance dispersal vehicle in the Southern Hemisphere. Science 304, 1144–1147. doi: 10.1126/science.1095210
Nogales, M., Heleno, R., Traveset, A., and Vargas, P. (2012). Evidence for overlooked mechanisms of long-distance seed dispersal to and between oceanic islands. New Phytol. 194, 313–317. doi: 10.1111/j.1469-8137.2011.04051.x
Onstein, R. E., Kissling, W. D., Chatrou, L. W., Couvreur, T. L. P., Morlon, H., and Sauquet, H. (2019). Which frugivory-related traits facilitated historical long-distance dispersal in the custard apple family (Annonaceae)? J. Biogeogr. 46, 1874–1888. doi: 10.1111/jbi.13552
Parham, J. F., Donoghue, P. C. J., Bell, C. J., Calway, T. D., Head, J. J., Holroyd, P. A., et al. (2012). Best practices for justifying fossil calibrations. Syst. Biol. 61, 346–359. doi: 10.1093/sysbio/syr107
Patnaik, R. (2015). Diet and habitat changes among Siwalik herbivorous mammals in response to Neogene and Quaternary climate changes: an appraisal in the light of new data. Quat. Int. 371, 232–243. doi: 10.1016/j.quaint.2014.11.025
Powell, C. L. E., Waskin, S., and Battistuzzi, F. U. (2020). Quantifying the error of secondary vs. distant primary calibrations in a simulated environment. Front. Genet. 11:252. doi: 10.3389/fgene.2020.00252
Queiroz-Lima, A., Amorim, A. M. A., and Cardoso, D. B. O. S. (2018). Two more elegant species of the neglected Sauvagesia elegantissima complex (Ochnaceae). Syst. Bot. 43, 221–230. doi: 10.1600/036364418X697076
Rambaut, A., Drummond, A. J., Xie, D., Baele, G., and Suchard, M. A. (2018). Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 67, 901–904. doi: 10.1093/sysbio/syy032
Rando, J. G., Zuntini, A. R., Conceição, A. S., van den Berg, C., Pirani, J. R., and Queiroz, L. P. (2016). Phylogeny of Chamaecrista ser. Coriaceae (Leguminosae) unveils a lineage recently diversified in Brazilian campo rupestre vegetation. Int. J. Plant Sci. 177, 3–17. doi: 10.1086/683846
Rapini, A., Bitencourt, C., Luebert, F., and Cardoso, D. (2021). An escape-to-radiate model for explaining the high plant diversity and endemism in campos rupestres. Biol. J. Linn. Soc. 133, 481–498. doi: 10.1093/biolinnean/blaa179
Razafimandimbison, S., Kainulainen, K., Senterre, B., Morel, C., and Rydin, C. (2020). Phylogenetic affinity of an enigmatic Rubiaceae from the Seychelles revealing a recent biogeographic link with Central Africa: gen. nov. Seychellea and trib. nov. Seychelleeae. Mol. Phylogenet. Evol. 143, 106685. doi: 10.1016/j.ympev.2019.106685
Ree, R. H., and Sanmartín, I. (2018). Conceptual and statistical problems with the DEC+J model of founder-event speciation and its comparison with DEC via model selection. J. Biogeogr. 45, 741–749. doi: 10.1111/jbi.13173
Ree, R. H., and Smith, S. A. (2008). Maximum likelihood inference of geographic range evolution by dispersal, local extinction, and cladogenesis. Syst. Biol. 57, 4–14. doi: 10.1080/10635150701883881
Renner, S. (2004). Plant dispersal across the tropical Atlantic by wind and sea currents. Int. J. Plant Sci. 165, S23–S33. doi: 10.1086/383334
Renner, S. S., Strijk, J. S., Strasberg, D., and Thébaud, C. (2010). Biogeography of the Monimiaceae (Laurales): a role for East Gondwana and long distance dispersal, but not West Gondwana. J. Biogeogr. 37, 1227–1238. doi: 10.1111/j.1365-2699.2010.02319.x
Ribeiro, P. L., Rapini, A., Damascena, L. S., and van den Berg, C. (2014). Plant diversification in the Espinhaço Range: insights from the biogeography of Minaria (Apocynaceae). Taxon 63, 1253–1264. doi: 10.12705/636.16
Ringelberg, J. J., Zimmermann, N. E., Weeks, A., Lavin, M., and Hughes, C. E. (2020). Biomes as evolutionary arenas: convergence and conservatism in the trans-continental succulent biome. Global Ecol. Biogeogr. 29, 1100–1113. doi: 10.1111/geb.13089
Ronquist, F. (1997). Dispersal-vicariance analysis: a new approach to the quantification of historical biogeography. Syst. Biol. 46, 195–203. doi: 10.1093/sysbio/46.1.195
Ruhfel, B. R., Bove, C. P., Philbrick, C. T., and Davis, C. C. (2016). Dispersal largely explains the Gondwanan distribution of the ancient tropical clusioid plant clade. Amer. J. Bot. 103, 1–12. doi: 10.3732/ajb.1500537
Sanmartín, I., and Meseguer, A. S. (2016). Extinction in phylogenetics and biogeography: from timetrees to patterns of biotic assemblage. Front. Genet. 7:35. doi: 10.3389/fgene.2016.00035
Sastre, C. (1971a). Essai de taxonomie numérique et schéma évolutif du genre Sauvagesia L. Sellowia 23, 9–44.
Sastre, C. (1971b). Distribution géographique des espèces de Sauvagesia L. Compt. Rend. Sommaires Scéances Soc. Biogéogr. 421, 47–59.
Sauquet, H. (2013). A practical guide to molecular dating. Comptes Rendus Palevol. 12, 355–367. doi: 10.1016/j.crpv.2013.07.003
Sauquet, H., Ho, S. Y. W., Gandolfo, M. A., Jordan, G. J., Wilf, P., Cantrill, D. J., et al. (2011). Testing the impact of calibration on molecular divergence times using a fossil-rich group: the case of Nothofagus (Fagales). Syst. Biol. 61, 289–313. doi: 10.1093/sysbio/syr116
Schatz, G. E. (1996). “Malagasy/Indo-Australo-Malesian phytogeographic connections,” in Biogéographie de Madagascar, ed. W. R. Lourenço (Paris: ORSTOM), 73–84.
Schenk, J. J. (2016). Consequences of secondary calibrations on divergence time estimates. PLoS One 11:e0148228. doi: 10.1371/journal.pone.0148228
Schneider, J. V., and Zizka, G. (2017). Phylogeny, taxonomy, and biogeography of Neotropical Quiinoideae (Ochnaceae s.l.). Taxon 66, 855–867. doi: 10.12705/664.4
Schneider, J. V., Bissiengou, P., Amaral, M. C. E., Fay, M. F., Thines, M., Sosef, M. S. M., et al. (2014). Phylogenetics, ancestral state reconstruction, and a new infrafamilial classification of the pantropical Ochnaceae s.l. (Medusagynaceae, Ochnaceae s.str., Quiinaceae) based on five DNA regions. Mol. Phylogenet. Evol. 78, 199–214. doi: 10.1016/j.ympev.2014.05.018
Schneider, J. V., Habersetzer, J., Rabenstein, R., Wesenberg, J., Wesche, K., and Zizka, G. (2017). Water supply and demand remain coordinated during breakdown of the global scaling relationship between leaf size and major vein density. New Phytol. 214, 473–486. doi: 10.1111/nph.14382
Schneider, J. V., Jungcurt, T., Cardoso, D., Amorim, A. M., Töpel, M., Andermann, T., et al. (2020). Phylogenomics of the tropical plant family Ochnaceae using targeted enrichment of nuclear genes and 250+ taxa. Taxon 70, 48–71. doi: 10.1002/tax.12421
Schneider, J. V., Paule, J., Jungcurt, T., Cardoso, D., Amorim, A. M., Berberich, T., et al. (2021). Resolving recalcitrant clades in the pantropical Ochnaceae: insights from comparative phylogenomics of plastome and nuclear genomic data derived from targeted sequencing. Front. Plant Sci. 12:638650. doi: 10.3389/fpls.2021.638650
Schuster, T. M., Setaro, S. D., and Kron, K. (2013). Age estimates for the buckwheat family Polygonaceae based on sequence data calibrated by fossils and with a focus on the amphi-pacific Muehlenbeckia. PLoS One 8:e61261. doi: 10.1371/journal.pone.0061261
Shah, T., Schneider, J. V., Zizka, G., Maurin, O., Baker, W., Forest, F., et al. (2021). Joining forces in Ochnaceae phylogenomics: a tale of two targeted sequencing probe kits. Amer. J. Bot. 108, 1201–1216. doi: 10.1002/ajb2.1682
Silvestro, D., Cascales-Miñana, B., Bacon, C. D., and Antonelli, A. (2015). Revisiting the origin and diversification of vascular plants through a comprehensive Bayesian analysis of the fossil record. New Phytol. 207, 425–436. doi: 10.1111/nph.13247
Simon, M. F., Grether, R., de Queiroz, L. P., Skema, C., Pennington, R. T., and Hughes, C. E. (2009). Recent assembly of the Cerrado, a Neotropical plant diversity hotspot, by in situ evolution of adaptations to fire. Proc. Natl. Acad. Sci. U.S.A. 106, 20359–20364. doi: 10.1073/pnas.0903410106
Smedmark, J. E. E., Eriksson, T., and Bremer, B. (2010). Divergence time uncertainty and historical biogeography reconstruction: an example from Urophylleae (Rubiaceae). J. Biogeogr. 37, 2260–2274. doi: 10.1111/j.1365-2699.2010.02366.x
Smith, S. A., and Donoghue, M. J. (2008). Rates of molecular evolution are linked to life history in flowering plants. Science 322, 86–89. doi: 10.1126/science.1163197
Stride, G., Nylinder, S., and Swenson, U. (2014). Revisiting the biogeography of Sideroxylon (Sapotaceae) and an evaluation of the taxonomic status of Argania and Spiniluma. Austr. Syst. Bot. 27, 104–118. doi: 10.1071/SB14010
Strijk, J. S., Noyes, R. D., Strasberg, D., Cruaud, C., Gavory, F., Chase, M. W., et al. (2012). In and out of Madagascar: dispersal to peripheral islands, insular speciation and diversification of Indian Ocean daisy trees (Psiadia, Asteraceae). PLoS One 7:e42932. doi: 10.1371/journal.pone.0042932
Thiv, M., van der Niet, T., Rutschmann, F., Thulin, M., Brune, T., and Linder, H. P. (2011). Old-New World and trans-African disjunctions of Thamnosma (Rutaceae): intercontinental long-distance dispersal and local differentiation in the succulent biome. Amer. J. Bot. 98, 76–87. doi: 10.3732/ajb.1000339
Thorne, R. F. (1973). “Floristic relationships between tropical Africa and tropical America,” in Tropical Forest Ecosystems in Africa and South America: A Comparative Review, eds B. J. Meggers and W. D. Duckworth (Washington, DC: Smithsonian Institution), 27–47.
Tiffney, B. H. (1985). The Eocene North Atlantic land bridge: its importance in Tertiary and modern phytogeography of the Northern Hemisphere. J. Arnold Arboretum 66, 243–273. doi: 10.5962/bhl.part.13183
Tiffney, B. H., and Manchester, S. R. (2001). The use of geological and paleontological evidence in evaluating plant phylogeographic hypotheses in the Northern Hemisphere Tertiary. Int. J. Plant Sci. 162, S3–S17. doi: 10.1086/323880
Towns, J., Cockerill, T., Dahan, M., Foster, I., Gaither, K., Grimshaw, A., et al. (2014). XSEDE: accelerating scientific discovery. Comput. Sci. Engineer. 16, 62–74. doi: 10.1109/MCSE.2014.80
Upchurch, P. (2008). Gondwanan break-up: legacies of a lost world? Trends Ecol. Evol. 23, 229–236. doi: 10.1016/j.tree.2007.11.006
Vasconcelos, T. N. C., Alcantara, S., Andrino, C. O., Forest, F., Reginato, M., Simon, M. F., et al. (2020). Fast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancient Neotropical mountains. Proc. R. Soc. B 287:20192933. doi: 10.1098/rspb.2019.2933
Veranso-Libalah, M. C., Kadereit, G., Stone, R. D., and Couvreur, T. L. P. (2018). Multiple shifts to open habitats in Melastomateae (Melastomataceae) congruent with the increase of African Neogene climatic aridity. J. Biogeogr. 45, 1420–1431. doi: 10.1111/jbi.13210
Viana, D. S., Gangoso, L., Bouten, W., and Figuerola, J. (2016). Overseas seed dispersal by migratory birds. Proc. R. Soc. B 283:20152406. doi: 10.1098/rspb.2015.2406
Warren, B. H., Strasberg, D., Bruggemann, J. H., Prys-Jones, R. P., and Thébaud, C. (2010). Why does the biota of the Madagascar region have such a strong Asiatic flavour? Cladistics 26, 526–538. doi: 10.1111/j.1096-0031.2009.00300.x
Weigelt, P., Steinbauer, M. J., Cabral, J. S., and Kreft, H. (2016). Late Quaternary climate change shapes island biodiversity. Nature 532, 99–102. doi: 10.1038/nature17443
Weston, P., and Crisp, M. D. (1994). Cladistic biogeography of waratahs (Proteaceae: Embothrieae) and their allies across the Pacific. Austr. Syst. Bot. 7, 225–249. doi: 10.1071/SB9940225
Xi, Z., Ruhfel, B. R., Schaefer, H., Amorim, A. M., Sugumaran, M., Wurdack, K. J., et al. (2012). Phylogenomics and a posteriori data partitioning resolve the Cretaceous angiosperm radiation Malpighiales. Proc. Natl. Acad. Sci. U.S.A. 109, 17519–17524. doi: 10.1073/pnas.1205818109
Yang, Z., and Rannala, B. (2006). Bayesian estimation of species divergence times under a molecular clock using multiple fossil calibrations with soft bounds. Mol. Biol. Evol. 23, 212–226. doi: 10.1093/molbev/msj024
Yoder, A. D., and Nowak, M. D. (2006). Has vicariance or dispersal been the predominant biogeographic force in Madagascar? Only time will tell. Annu. Rev. Ecol. Evol. Syst. 37, 405–431. doi: 10.1146/annurev.ecolsys.37.091305.110239
Yuan, Y.-M., Wohlhauser, S., Möller, M., Klackenberg, J., Callmander, M. W., and Küpfer, P. (2005). Phylogeny and biogeography of Exacum (Gentianaceae): a disjunctive distribution in the Indian Ocean Basin resulted from long distance dispersal and extensive radiation. Syst. Biol. 54, 21–34. doi: 10.1080/10635150590905867
Keywords: boreotropics, historical biogeography, Malpighiales, land bridges, molecular clock, phylogenomics, spatio-temporal evolution
Citation: Schneider JV, Jungcurt T, Cardoso D, Amorim AM, Paule J and Zizka G (2022) Predominantly Eastward Long-Distance Dispersal in Pantropical Ochnaceae Inferred From Ancestral Range Estimation and Phylogenomics. Front. Ecol. Evol. 10:813336. doi: 10.3389/fevo.2022.813336
Received: 11 November 2021; Accepted: 03 January 2022;
Published: 10 February 2022.
Edited by:
Hang Sun, Kunming Institute of Botany, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Yaowu Xing, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences (CAS), ChinaRowan J. Schley, University of Exeter, United Kingdom
Copyright © 2022 Schneider, Jungcurt, Cardoso, Amorim, Paule and Zizka. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Georg Zizka, R2VvcmcuWml6a2FAc2VuY2tlbmJlcmcuZGU=