 Gregorio de Chevalier1,2,3*
Gregorio de Chevalier1,2,3* Sébastien Bouret3
Sébastien Bouret3 Ameline Bardo1,4
Ameline Bardo1,4 Bruno Simmen2
Bruno Simmen2 Cécile Garcia2†
Cécile Garcia2† Sandrine Prat1†
Sandrine Prat1†
- 1UMR 7194, CNRS-Muséum National d’Histoire Naturelle-UPVD, Musée de l’Homme, Paris, France
- 2UMR 7206, CNRS-Museum National d’Histoire Naturelle-Université de Paris, Paris, France
- 3Motivation, Brain and Behavior Team, Institut du Cerveau et de la Moelle Épinière (ICM), INSERM UMRS 1127, CNRS UMR 7225, Pitié-Salpêtrière Hospital, Paris, France
- 4Skeletal Biology Research Centre, School of Anthropology and Conservation, University of Kent, Canterbury, United Kingdom
While the exploitation of aquatic fauna and flora has been documented in several primate species to date, the evolutionary contexts and mechanisms behind the emergence of this behavior in both human and non-human primates remain largely overlooked. Yet, this issue is particularly important for our understanding of human evolution, as hominins represent not only the primate group with the highest degree of adaptedness to aquatic environments, but also the only group in which true coastal and maritime adaptations have evolved. As such, in the present study we review the available literature on primate foraging strategies related to the exploitation of aquatic resources and their putative associated cognitive operations. We propose that aquatic resource consumption in extant primates can be interpreted as a highly site-specific behavioral expression of a generic adaptive foraging decision-making process, emerging in sites at which the local cost-benefit trade-offs contextually favor aquatic over terrestrial foods. Within this framework, we discuss the potential impacts that the unique intensification of this behavior in hominins may have had on the evolution of the human brain and spatial ecology.
Introduction
Under the framework of optimal foraging theory (Stephens and Krebs, 1986), animals are thought to have evolved foraging strategies that increase individual fitness by maximizing the benefits while minimizing the costs and risks related to the exploitation of resources in their natural habitats (Charnov, 1976). In turn, the outcomes of this optimization are suggested to be reflected in a variety of observable behaviors and domains, such as spatial ecology (Pyke, 2019b; Tórrez-Herrera et al., 2020), patch use (Bedoya-Perez et al., 2013) and dietary preferences (Harris et al., 2019). Accordingly, the evolution of foraging strategies must be accompanied by the evolution of (i) cognitive abilities that allow for a contextual computation of costs and benefits resulting in the decision-making processes expressed as behavior (Rosati, 2017) and (ii) their specific neuroanatomical correlates, such as overall brain size (e.g., DeCasien et al., 2017) or the size of specific brain regions involved in those processes (e.g., Louail et al., 2019). As such, considering this framework for investigating the underpinnings of differences in foraging niches and territorial occupation patterns between primate species, and its application in the context of human evolution may provide essential insights on what mechanisms have allowed hominins to inhabit an unmatched variety of ecosystems when compared to non-human primate species.
In this context, two main hypotheses have been formulated to link the decision-making challenges primates face in the wild to the evolution of their complex cognition and brain size: the social and ecological brain hypotheses. According to the former (Dunbar, 1998; Dunbar and Shultz, 2007), cognitive challenges associated with living in large, complex social groups require individuals to keep track of and act on a broader record of competitive and cooperative interactions, which in turn generates selective pressures for adaptive cognitive traits (Byrne, 1996; Brosnan et al., 2010; Massen et al., 2014). As such, the increase in social complexity would comprise the most expressive selective force acting on the evolution of primate cognition and, in turn, of their brain size (Jolly, 1966; Humphrey, 1976; Dunbar, 1998). The second hypothesis, i.e., the ecological brain hypothesis, seeks to explain the evolution of brain size and complex cognition primarily using ecological proxies (Milton, 1981, 1988; Rosati, 2017). In this context, the evolution of cognitive traits related to decision-making, spatial memory and executive control, for example, would be adaptive toward optimizing the trade-offs between costs and benefits of foraging in complex and fluctuating environments (Rosati, 2017; Garcia et al., 2021). In support of this hypothesis, a recent study by DeCasien et al. (2017) using data from a wide diversity of primate species provided evidence that diet is a better predictor of total relative brain size in primates—an measure for global cognitive skills (Deaner et al., 2007; Shultz and Dunbar, 2010)—when compared to social variables, such as group size. Nonetheless, recent research has demonstrated that overall brain size can be a poor proxy for studying the development of specific cognitive skills related to foraging ecology and sociality (Louail et al., 2019). Moreover, it is important to note that these two hypotheses are not mutually exclusive, and each of them can be better at explaining different aspects of primate cognition (Cunningham and Janson, 2007; Rosati, 2017). In this sense, social proxies may be better suited to explain the evolution of cognitive skills related to social learning, for example, and ecological factors, in contrast, may offer a higher explanatory power for the evolution of cognitive skills related to foraging and dietary preferences (Ban et al., 2016; Janmaat et al., 2016; Trapanese et al., 2019). This observation is consistent with evidence presented by DeCasien and Higham (2019), suggesting that the mosaic brain evolution observed in primates is linked to niche specialization, with the evolution of the size of different brain regions being selected according to specific ecological characteristics of distinct primate species (Louail et al., 2019).
Several recent studies aimed at assessing the evolution of cognitive skills involved in finding and exploiting high-quality ephemeral resources in complex terrestrial environments (e.g., Janmaat et al., 2014; Ban et al., 2016; Salmi et al., 2020). In densely forested tropical ecosystems, for example, high-energy foods, such as fruits, may be sparsely distributed and difficult to find, making food search a highly costly activity (Janmaat et al., 2016). As such, the goal of optimal foragers inhabiting these areas is to increase their intake yield while minimizing the energetic expenses derived from food search (Schoener, 1971). One way chimpanzees (Pan troglodytes) have been demonstrated to achieve this is via the modulation of their navigation strategies through the use of less energetically demanding paths when moving between sparsely distributed food sources (Green et al., 2020). In addition, chimpanzees are also known to plan their nest departure time according to the availability, nutritional quality and location of seasonal food resources (Janmaat et al., 2014). However, in the context of energetic optimization, some high-risk, high-yield items such as high-trophic level aquatic resources (both marine and riverine/lacustrine) could also hold major importance in the regulation of energetic balance of primates (Cunnane and Stewart, 2010; Koops et al., 2019), but have been rather overlooked compared to terrestrial resources. This focus on terrestrial resources, albeit partly rooted in the fact that most non-human primate species inhabiting coastal or flooded areas do not forage on aquatic resources (Nowak et al., 2019), may have caused researchers to overlook the relative importance of this behavior in the evolution of some primate species, and particularly in hominins. As pointed out by Archer et al. (2014), for example, the components of archeological assemblages related to aquatic fauna have been far less studied as evidence of early hominin exploitation when compared to terrestrial components, even when the two are found at the same site. As such, this type of issue creates a bias in our reconstruction of the niches occupied by extinct hominins and extant primates, leading to gaps in our understanding of how the consumption of aquatic resources may be linked to the evolution of brain size and cognition in these groups.
Thus, the purpose of this literature review is to discuss primate foraging strategies and their putative associated cognitive operations related to the exploitation of aquatic resources, as well as their implications in the evolution of the primate brain. As such, firstly we present some of the main patterns observed in the aquatic foraging behavior of non-human primates. Then, we review the costs and benefits related to the consumption of aquatic resources, with a particular focus on how marine and freshwater resources contrast with terrestrial resources in their contribution to the energetic optimization in extant non-human primates. We also consider the evidence for intentionality in the exploitation of aquatic resources by hominins and non-human primates and, as such, the extent to which their associated behaviors take place in a planned or opportunistic fashion. In addition, we also aim to specify the potential cognitive operations involved in foraging for aquatic resources, and evaluate the correlation between the evolution of such skills and neuroanatomical changes during the course of human evolution. Finally, by examining the evidence for the emergence of true coastal adaptation in hominins, we aim to assess the implications of the exploitation of coastal resources in the context of territorial occupation, movement ecology, technological complexity and social behavior, as well as the relation of such novelties with the evolution of complex cognitive skills. Overall, the present review seeks to set the foundation for future research attempting to ratify evolutionary interrelationships between aquatic resource exploitation, cognitive ecology and neuroanatomy in extant primates and extinct hominins.
Patterns in the Exploitation of Aquatic Resources by Non-Human Primates
While several primate species that inhabit flooded areas exhibit aquatic behaviors that are relevant to aspects of their ecology, only a fraction of them seems to forage on aquatic foods (Nowak et al., 2019). Together, the five main reviews on the use of aquatic resources by non-human primates show that a total of 26 species (i.e., about 4% of the existing non-human primate species) feed on aquatic fauna (Stewart et al., 2008; Kempf, 2009; Stewart, 2010; Russon et al., 2014; Nowak et al., 2019), out of which ten are found to feed on fish (Russon et al., 2014; Mallick, 2019). Since their publication, developments in this field have caused this number to increase, for example with the first evidence of regular consumption of aquatic fauna (i.e., crabs) by a population of chimpanzees in the Nimba mountains, Guinea (Koops et al., 2019). Still, the total number of species that exploit aquatic resources in general—including fauna and flora—remains disputed, as no systematic reviews are available on the consumption of aquatic flora by primates (Russon et al., 2014). Nevertheless, aquatic foraging has been well documented in several species—e.g., chimpanzees (Nishida, 1980; Sakamaki, 1998; Devos et al., 2002), bonobos (Hohmann et al., 2019), chacma baboons (Lewis et al., 2018) and long-tailed macaques (Tan et al., 2015; Tan, 2017)—leading to speculation on its relative importance for the ecology of a variety of primate groups, and ultimately in the context of hominin evolution (e.g., Boesch et al., 2017; Hohmann et al., 2019; Koops et al., 2019).
Although the exploitation of aquatic fauna and flora may involve distinct acquisition and extraction processes, a few general patterns emerge from the available records of aquatic resource consumption by extant non-human primates. Firstly, behaviors associated with aquatic resource exploitation tend to be highly site-specific, independently of the type of food consumed (Table 1). In chimpanzees, for example, the consumption of aquatic fauna has only been observed in the Nimba mountains (Koops et al., 2019), in spite of the existence of several long-term field projects dedicated to the study of this species in situ (e.g., Pusey et al., 2007; Boesch et al., 2019; Thompson et al., 2020)—also including nearby locations, such as Bossou (Humle et al., 2011; Koops et al., 2019; Matsuzawa, 2019). Notably, the chimpanzee populations in these two locations also differ in their consumption of aquatic flora: while individuals at Bossou have been observed engaging in algae scooping—a behavior characterized by using sticks to scoop algae from water bodies (Matsuzawa, 1996; Humle et al., 2011)—the chimpanzees at Nimba have not (Matsuzawa, 2019). As such, the disparity in observed aquatic foraging behaviors in these two locations has been hypothesized to be a product of differences in resource availability within the ranges of both chimpanzee groups, given the absence of shallow freshwater streams with crabs at Bossou and the scarcity of ponds with algae at Nimba (Matsuzawa, 2019). Nevertheless, chimpanzees represent only one example of the site-specificity of the consumption of aquatic fauna and flora in primates. Differences in aquatic fauna exploitation have been described between chacma baboons troops (Papio ursinus) living at Cape Reserve—which fed on marine invertebrates (Hall, 1962; Lewis and O’Riain, 2019)—and groups inhabiting the Namib desert, which fed on fish from drying desert pools (Hamilton and Tilson, 1985). In addition, this pattern can also hold true for certain species of Pan-American monkeys, such as bearded capuchins (Sapajus libidinosus), which have also been recorded to forage on either marine invertebrates (Santos et al., 2019b) or fishes (Mendes et al., 2000), depending on the studied site.
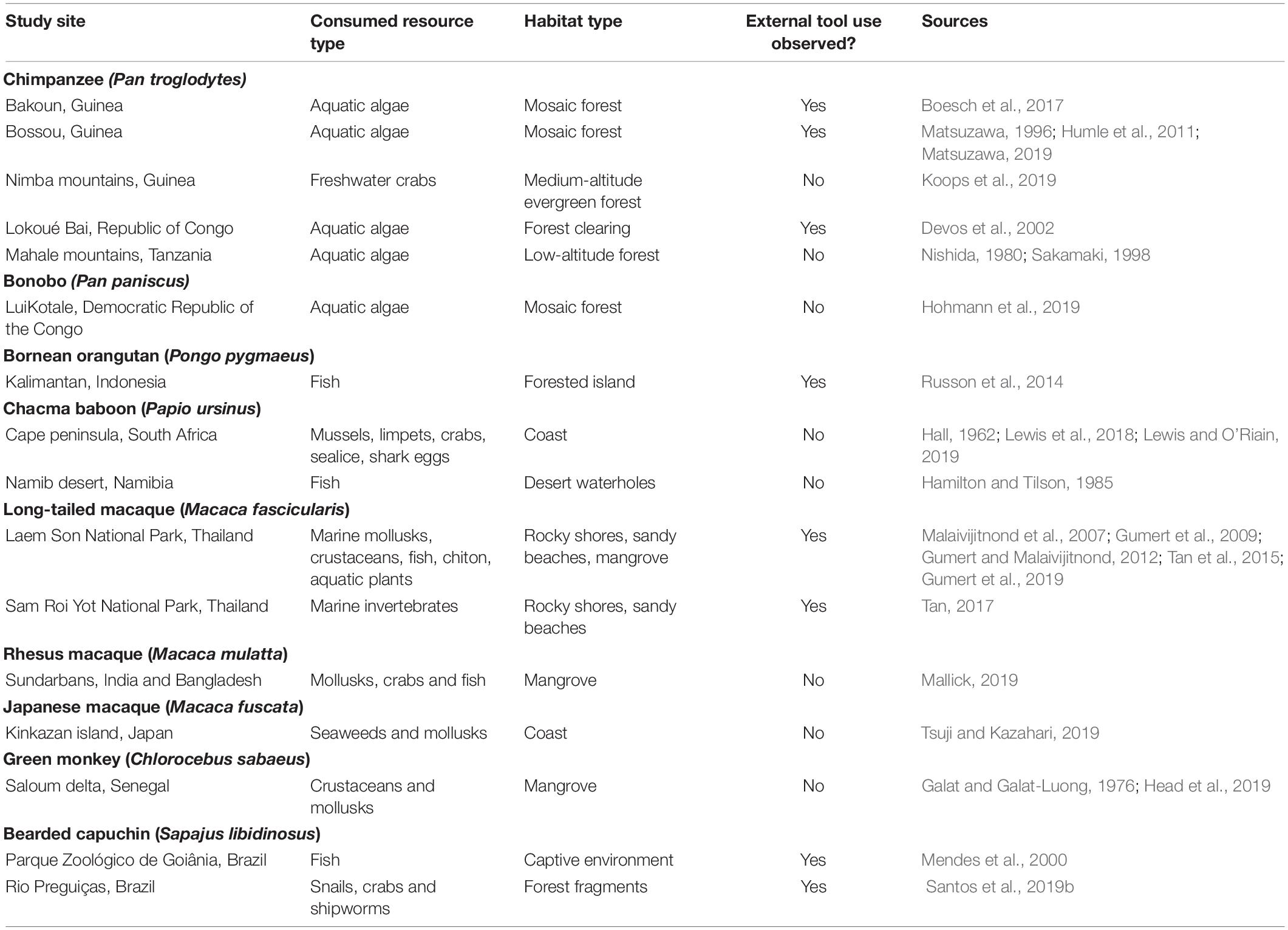
Table 1. Cited examples of aquatic resource consumption by non-human primates.
Secondly, the exploitation of aquatic resources in a variety of sites can also be dependent on demographic and individual factors. This is the case for Orangutans living in a fresh-water island habitat in Borneo, Indonesia, for example, where aquatic foraging behavior depends on individual age and water skills (Russon et al., 2014). At this site, all the individuals who were observed to engage in fish catching and eating were juveniles or adolescents. In this context, Russon et al. (2014) suggested that this pattern may be related to the innovative character of juvenile and adolescent primates, which has been well documented in previous research (e.g., Reader and Laland, 2001; Russon et al., 2010). In addition, orangutan individuals who consumed fish were also more skilled in the water compared to others at the same site (Russon et al., 2014), providing evidence that previously acquired skills of individuals can also affect the development of behaviors involved in aquatic resource exploitation. Furthermore, demographic factors have also been demonstrated to affect the consumption of aquatic crabs by chimpanzees at the Nimba mountains (Koops et al., 2019), where females and infants were found to forage for crabs more frequently and for longer periods when compared to adult males, which was hypothesized to be linked to their reduced access to game meat (Koops et al., 2019).
Thirdly, the exploitation of aquatic resources in primates may or may not rely on the use of tools (Table 1), depending on the species and individual skills. Several species of extant non-human primates have been recorded to exploit aquatic foods using external tools (Russon et al., 2014). A notable example outside of the great apes are Burmese long-tailed macaques (Macaca fascicularis), which employ a variety of stone-hammering techniques for obtaining aquatic foods in coastal environments (Tan et al., 2015; Gumert et al., 2019). For each of these techniques, individuals may employ distinct manual skills, in addition to selecting tool types according to their suitability for the intended task (Gumert et al., 2009; Tan et al., 2015). Besides its specificity regarding the target food item, tool-assisted aquatic resource foraging techniques can also vary within species across different locations. For example, bearded capuchins have been documented to employ different tool-assisted foraging techniques to obtain aquatic foods in distinct study sites, such as the use of baits to prey on fish (Mendes et al., 2000) and shell cracking through percussion (Santos et al., 2019b). In the first case, fishing capuchins would either place food baits under water or hold them partially submerged to attract pond fish, subsequently assuming a fishing body posture that allowed the individual to observe and attempt to capture approaching fishes that were attracted to the bait (Mendes et al., 2000). Invertebrate foraging individuals, on the other hand, used branches as hammers to crack the shells of marine invertebrates in order to forage on them (Santos et al., 2019b). Nevertheless, similarities in techniques used for foraging on aquatic resources have also been observed across sites located at great distances from each other, showing that some of these behaviors can also be geographically widespread. This is the case in some chimpanzee populations which have been observed to employ comparable scooping on foraging for algae in a variety of sites throughout Western (Matsuzawa, 1996; Boesch et al., 2017) and Central Africa (Devos et al., 2002), all of which included some form of tool selection or modification.
Lastly, alike other innovations, behaviors related to the exploitation of aquatic foods can be invented, lost, independently reinvented, acquired through social learning and adapted to and from other tasks unrelated to aquatic foraging (Nishida et al., 2009; Shumaker et al., 2011; Russon et al., 2014; Bandini and Tennie, 2017; Luncz et al., 2017; Mallick, 2019; Santos et al., 2019b). Therefore, in some cases, such behaviors can potentially develop into local traditions with respect to (i) the social position of individuals partaking in it (e.g., Koops et al., 2019), (ii) the type of resource consumed (Matsuzawa, 1996, 2019; Koops et al., 2019), and possibly (iii) the technique used to obtain it (e.g., algae feeding behavior as described in Sakamaki, 1998; Devos et al., 2002; Boesch et al., 2017, albeit the evidence described in Sakamaki, 1998 stems from only one female chimpanzee who was hypothesized to have acquired her distinct algae feeding behavior from her natal group). Furthermore, aspects of exploiting aquatic foods in non-human primates may not only be socially learned from conspecifics, such as shellfish cracking techniques in long-tailed macaques (Tan, 2017), but also be facilitated or primed by the behavior of individuals from other species, such as Homo sapiens (reviewed in Russon et al., 2014). Finally, in some cases, such as in long-tailed macaques, the exploitation of aquatic resources can encompass a variety of local traditions (Tan et al., 2015). Thus, alike other behaviors associated with foraging for terrestrial resources (e.g., tool use; Whiten et al., 1999; Whiten and van de Waal, 2017), it might be hypothesized that, in some cases, the exploitation of aquatic resources by extant non-human primates can be interpreted as a cultural phenomenon, adding to the portfolio of socially transmissible behavioral strategies that different species and populations have developed to forage more optimally on locally available resources.
Trade-Offs Between Aquatic and Terrestrial Foraging
The nature and the importance of aquatic resource exploitation in the context of primate feeding ecology remains a contested topic, as many species of primates that live nearby water bodies or flooded areas often do not feed on aquatic foods (Nowak et al., 2019). Thus, the precise mechanisms through which the consumption of aquatic resources can be beneficial for and influence selection in primate populations remains largely unknown (Hohmann et al., 2019). In addition, the site-specificity of the types of resources consumed, of the demographics of who consume them and of the employed techniques hinders general interpretations of the value of such behavior. Nevertheless, within the framework of optimal foraging theory, there may be several potential ways in which foraging for aquatic resources can be a useful behavioral strategy toward energetic optimization in non-human primates.
Firstly, the nutritional benefits of aquatic fauna consumption would include increased intake of long-chain polyunsaturated fatty acids (LC-PUFAs), which have been regarded as important for the development and function of large primate brains (Joordens et al., 2014). In this sense, the intake of eicosapentaenoic acid (EPA) and arachidonic acid (AA) would be specially increased with aquatic fauna consumption, as these nutrients are rare in terrestrial foods, with the exception of the fatty parts of meat and the brain tissue of some animals (Li et al., 1998; Cordain et al., 2002; Carlson and Kingston, 2007; Stewart, 2010; Joordens et al., 2014). However, brain tissue can be costly to extract, and given the evidence for the health risks associated with its consumption in humans, such as prion diseases (Berger et al., 1997; Domachowske and Suryadevara, 2020), and the susceptibility of non-human primates to such diseases (Dalsgaard, 2002; Race et al., 2009), acquiring EPA and AA through the consumption of brain tissue may not represent an optimal strategy when compared to aquatic fauna. In addition, obtaining fresh terrestrial animal tissue through hunting can be highly energetically costly, dangerous (Boesch, 1994; Tennie et al., 2014) and carry the associated risk of failure (Boesch and Boesch-Achermann, 2000; Gilby and Wrangham, 2007). Comparatively, foraging on some aquatic resources such as mollusks and arthropods can represent a less risky and more energetically efficient alternative to obtain LC-PUFAs for some non-human primates, especially considering the relative abundance and ease of access to these resources in some sites (e.g., Malaivijitnond et al., 2007; Koops et al., 2019). In this sense, this behavior could be particularly advantageous for non-human primate populations that exhibit relatively low hunting opportunities (Koops et al., 2019), and for individuals who have less access to hunting meat, such as females and infants (Fahy et al., 2013; Gilby et al., 2017). Nevertheless, other brain-essential LC-PUFAs, such as docosahexaenoic acid (DHA), can also be metabolically synthesized from precursors, but the relative importance of their dietary uptake vs. their synthesis in the context of brain development remains unclear, and should be experimentally addressed in future studies (Joordens et al., 2014; Koops et al., 2019). Moreover, the quantities of EPA, DHA, and AA found in aquatic fauna can be tied to local conditions, and as such it may vary over time, space and according to the species consumed (Joordens et al., 2014).
Furthermore, it has been hypothesized that the consumption of aquatic flora is important for ensuring sufficient uptake of certain minerals like iodine (Hohmann et al., 2019). Iodine is essential for numerous physiological and developmental functions, including brain development, due to its influence on thyroid function (Venturi and Bégin, 2010). It can be abundant in a diversity of ecosystems, such as coastal and volcanic areas, as well as wetlands (Hohmann et al., 2019). In other terrestrial habitats such as rainforests, on the other hand, access to iodine can be limited (World Health Organization, 2007), with health issues tied to iodine deficiency occurring at relatively high rates in some human populations inhabiting these areas (Phillips et al., 1988; Vanderpas and Moreno-Reyes, 2017). Given the similar detrimental effects that iodine deficiency can have in humans and non-human primates (Mano et al., 1987), it is reasonable to assume that developing a way to access iodine in these iodine-poor environments would be advantageous, and possibly confer a fitness advantage. In this sense, Hohmann et al. (2019) have demonstrated that aquatic algae can provide a rich source of iodine for primates living in rainforest environments, and that a population of bonobos (Pan paniscus) at the Congo basin regularly consumed aquatic herbs as part of their diet. In addition, aquatic algae were found to contain higher levels of several essential minerals (i.e., Mn, Ca, Mg, Fe, K, and I) when compared to ripe fruits and terrestrial herbs (Hohmann et al., 2019). Other species of apes also feed on aquatic herbs (Kempf, 2009), and the potential importance of aquatic plants as sources of minerals in primate diets has also been proposed by other authors (e.g., Boesch et al., 2017).
In this context, given the diverse nature of behaviors linked to aquatic resource exploitation in non-human primates, the costs and benefits of consuming these items have also been hypothesized to vary temporally, spatially and across sex and age. Long-tailed macaques, for example, show higher rates of aquatic foraging during periods when the abundance of ripe fruit is reduced (Yeager, 1996; Malaivijitnond et al., 2007). In this case, the consumption of aquatic resources could potentially serve as a fallback source of nutrients in periods when other food sources are scarcer (Stewart, 2010). In Bakoun, Guinea, fishing for algae among chimpanzees is frequent during the dry season and absent during the wet season (Boesch et al., 2017). Aquatic algae is a highly preferred food item at this site, which has been suggested to play an important role in satisfying chimpanzee dietary requirements by providing a source of protein, carbohydrates, lipids and minerals (Becker, 2007; Tipnee et al., 2015; Boesch et al., 2017). Given the limited availability of aquatic algae in Bakoun, which is restricted to the dry season, its frequent consumption during such periods likely represents a more optimal behavioral strategy considering the otherwise limited access to abundant terrestrial foods and water at this location (Boesch et al., 2017). Contrastingly, in sites where ponds with aquatic algae are scarcely found, algae foraging behavior would likely incur higher search costs and lower yields, which in turn would not be as energetically profitable. This is the case for chimpanzees inhabiting the Nimba mountains, where algae scooping behavior has not yet been observed (Matsuzawa, 2019), and instead chimpanzees feed on aquatic crabs, which are more widely available and likely represent a more optimal local foraging strategy (Koops et al., 2019; Matsuzawa, 2019). Unlike the aquatic algae foraging in Bakoun, crab consumption on Nimba mountains chimpanzees occurs independently of seasonality, and is instead dependent on demographic factors, with females and infants engaging in this behavior significantly more often than adult males (Koops et al., 2019). In this context, Koops et al. (2019) proposed that the associated benefits of crab foraging for females may be linked to an increase in access to salts. In addition, given the small size of the majority of crabs found at this site (Koops et al., 2019), crab fishing also poses less injury risks when compared to hunting (e.g., getting wounded; Busse, 1977) and ant-dipping (e.g., suffering painful bites; Humle et al., 2009). As such, this activity could be suitable for immature individuals practicing their foraging skills (Koops et al., 2019).
Nevertheless, foraging on aquatic resources is not devoid of associated risks. Freshwater crabs, for example, may be a source of parasites that cause human lung fluke disease, which is known to also affect non-human primates (Sachs and Voelker, 1975; Voelker and Sachs, 1977; Friant et al., 2015). Moreover, primates in close proximity to water bodies may be susceptible to predators such as crocodilians in some environments (Cowlishaw, 1994; Hill and Dunbar, 1998; Cheney et al., 2004). At coastal sites, primates engaging on aquatic foraging may also face challenges related to accessing frequently submerged food items (e.g., mollusks and invertebrates). The predation of marine organisms by chacma baboons in the intertidal zone at the Cape Peninsula, for example, has been negatively correlated to increases in risks associated to coastal foraging (e.g., wave height, offshore wind speed and increasing tides; Lewis and O’Riain, 2019). In addition, the use of flooded habitats may also involve increases in energetic costs associated with thermoregulation (Head et al., 2019) and with the extension of ranging areas and travel distances (Santos et al., 2019a). In this context, such cost-benefit trade-offs may be more likely to affect aquatic foraging when the relative payoff of such behavior is higher. In line with this premise, a recent study on Japanese macaques (Macaca fuscata) has found that their seafood feeding behavior is only linked to tidal cycles in months when forests are poorer in resources, favoring behaviors that increase the efficiency of aquatic resource exploitation (Tsuji and Kazahari, 2019). Finally, in some cases, the exploitation of aquatic foods can also involve high extraction costs, especially when it concerns the consumption of encapsulated foods (e.g., shelled mollusks and crustaceans). To counter this issue, some non-human primate populations resort to the use of percussive tools, presumably to increase the efficiency of the acquisition of aquatic resources (Gumert et al., 2019; Santos et al., 2019b). Such use of percussive tools in the context of aquatic foraging can be cognitively demanding (Santos et al., 2019b), which may help to explain why this behavior has only been observed in species for which the use of percussive techniques for the extraction of encapsulated forest foods has also been recorded: long-tailed macaques (Gumert et al., 2019) and bearded capuchins (Santos et al., 2019b). Nevertheless, it is important to note that there are also examples of other non-human primate species that are able to intensively exploit shelled aquatic resources without using external tools, such as green monkeys (Galat and Galat-Luong, 1976; Head et al., 2019) and rhesus macaques (Mallick, 2019).
Intentionality in the Consumption of Aquatic Resources
Within the framework of the optimal foraging theory, individuals would evolve cognitive skills that allow them to exhibit more efficient foraging decision-making strategies in their local environment (Charnov, 1976; Stephens and Krebs, 1986; Pyke, 2019a). Thus, when the benefits of exploiting aquatic resources outweigh its risks and costs, it can be expected that this behavior will take place intentionally (i.e., as the expected outcome of a decision-making process) rather than opportunistically (i.e., via a set of simple reflexes based on direct perceptions that enable animals to maximize intake and minimize costs without any cognitive operation). In this sense, while some non-human primate species have been shown to make use of aquatic habitats intentionally, for example, for predation avoidance choice (see Otani et al., 2020), establishing the intentionality of aquatic resource exploitation would require evidence for planning and anticipation of a specific feeding event by an individual. In this context, although it has been suggested that deliberate aquatic resource consumption in primates can be detected from search behavior (Russon et al., 2014), the most reliable available proxy to establish intentionality is likely tool selection followed by tool use.
In order to select, prepare and flexibly use tools, animals need both to understand causality and to be able to plan sequences of actions (Musgrave and Sanz, 2019). In other words, when an individual selects and uses a tool for foraging, it must first be able to anticipate the required task to obtain the intended resource, subsequently engaging with the chosen tool in a way that is suitable for accomplishing the envisioned goal. Algae-scooping chimpanzees in Bossou, for example, are known to preferably use fern stalks when manufacturing the sticks that are used to retrieve algae from ponds, which in turn has been hypothesized to be due to its suitability for the task, as the small hooks on fern stalks may increase algae gathering efficiency (Matsuzawa, 2019). Likewise, long-tailed macaques are known to select aquatic foraging tools according to their intended use, employing different techniques depending on prey species (Gumert et al., 2009). Additionally, tool-use has been suggested to depend on the prior computation of the relative costs and benefits of tool-assisted foraging when compared to other foraging modes (Musgrave and Sanz, 2019). In Bakoun, where algae fishing becomes a highly profitable foraging strategy during the dry season, chimpanzees have never been observed to collect algae with their hands, and instead used stick tools to retrieve the algae in all instances (Boesch et al., 2017). This preference for tool use has been proposed to increase the yield of algae consumption—as deeper sections of water bodies contain more algae—while at the same time reducing the associated costs (e.g., thermoregulation) by eliminating the need for immersion in the water (Boesch et al., 2017).
Furthermore, intentional aquatic resource exploitation can involve complex decision-making strategies such as evaluating what, where and when to eat in fluctuating terrestrial environments (Trapanese et al., 2019). As such, the emergence of more sophisticated aquatic foraging behaviors in primates, such as fish-eating, was suggested to involve a gradual increase in behavioral complexity over time (Russon et al., 2014). According to this hypothesis, this behavior would start with the inadvertent acquisition of fish by naïve individuals, successively progressing into opportunistic hand-catching, followed by intentional hand-catching and finally developing into tool-assisted fish acquisition (Stewart, 2010). This hypothesis is supported by a study on fish-eating by orangutan populations in Borneo, where fish-catching that involved tool use was preceded by all hypothesized precursors (Russon et al., 2014). Comparatively, for early hominins inhabiting the coast, intertidal zones would provide a richness of edible seaweed, shellfish and fish, which when combined with other resources that wash up on the shore, such as carcasses, would provide an ideal scenario for opportunistic scavenging without the need for advanced technology (Erlandson, 2017). Over time, such practice would open up possibilities for the development of more complex and efficient strategies for foraging on marine resources. This scenario would not only be consistent with Stewart’s (2010) hypothesis, but also resemble previously proposed ways in which terrestrial hunting could have developed in early hominins, beginning with confrontational scavenging, progressing toward cooperative exhaustion pursuits and ultimately leading to more complex hunting strategies involving long-distance weapons and other tools (Garcia et al., 2021). However, the scarcity of available research on the evolution of the techniques that primates use to exploit aquatic resources indicates that the existence of such a generalized pathway toward the development of this behavior remains speculative (Russon et al., 2014). In addition, different populations and species may also acquire aquatic fauna exploitation behavior through distinct pathways according to variations in local ecological conditions and required techniques (Russon et al., 2014). In this sense, it is also likely that human populations inhabiting different ecosystems started to exploit aquatic resources through distinct behavioral pathways, whether that included the consumption of fish or not.
Within this framework, when trying to infer intentionality in early hominin aquatic resource use, scientists cannot rely on direct observation, and instead must use indirect proxies. The difficulties associated with this methodology are manifold. Firstly, not all types of aquatic resource exploitation produces remains that may become material evidence, such as the consumption of aquatic flora without the use of tools. Secondly, even when tools are used, they may be made from soft material, such as those used by some non-human primates to catch algae (e.g., Matsuzawa, 1996; Boesch et al., 2017). As such, their remains are unlikely to be preserved in the archeological record, which in turn may create a bias in the material evidence preventing the accurate reconstruction of the emergence of aquatic resource exploitation in primates and extinct hominins (McGrew, 2010). In this context, most of the available evidence for aquatic foraging in early hominins comes from taphonomic analyses, frequently from sites where paleoanthropological remains are associated with aquatic fauna remains (Will et al., 2019). The use of this methodology has allowed scientists to suggest that Plio-Pleistocene hominins were exploiting aquatic resources as early as 1.95 Ma at the Turkana Basin (Archer et al., 2014). In addition, it also provided evidence for shellfish foraging and for shell tool manufacturing by Homo erectus in Java around 0.5 Ma (Joordens et al., 2015). Nevertheless, reliably determining whether this type of exploitation was opportunistic or not solely from faunal assemblages can be challenging, and scientists must also rely on complementary research methods that allow for dietary reconstruction, such as isotopic analyses (e.g., Lewis and Sealy, 2018). In the Neanderthal site of Vanguard cave, in Gibraltar, for example, while the taphonomic evidence points to the collection, processing and consumption of aquatic resources during the Middle Paleolithic, the absence of isotopic evidence for this behavior—as well as the absence of associated tools—suggests that this practice was opportunistic (Stringer et al., 2008; Richards and Trinkaus, 2009; McLeod, 2018). A variety of other sites provide evidence for the consumption of aquatic animals by Neanderthals in France (Hardy and Moncel, 2011), Greece, Italy, Portugal, Spain (Cortés-Sánchez et al., 2011) and Belgium (Guillaud et al., 2021), albeit without isotopic or tool-related evidence for intentional exploitation. The majority of the evidence for intentional and intensive exploitation of aquatic resources within hominins comes from Anatomically Modern Humans (AMHs), with the first record of tool-associated mollusk exploitation dating back to around 164 ka from Pinnacle Point, South Africa (Marean et al., 2007). Despite the existence of some described fishing tools from the upper Paleolithic (e.g., Gramsch et al., 2013), most lithic evidence for tool-associated fishing comes from much later during the Mesolithic (Cleyet-Merle, 1990). In this context, isotopic analyses of AMH remains also point toward a greater reliance on aquatic resources when compared with Neanderthals during the Paleolithic, albeit with significant regional variation (Richards and Trinkaus, 2009).
However, the period in which extinct hominins and modern humans may have started to intentionally forage for aquatic resources is likely underestimated for two reasons. Firstly, the post-glacial sea level rise during the Holocene has likely limited the archeological record from coastal hominin occupations from the Pleistocene (Bailey et al., 2007; Erlandson, 2017), potentially resulting in a significant loss in evidence of early coastal—and consequently aquatic—resource exploitation. Secondly, not all behaviors associated with intentional aquatic resource exploitation require technological assistance, as it is evidenced by data collected from non-human primates (Russon et al., 2014) and from ethnographical studies in modern indigenous populations consuming similar resources (Stewart, 1994). In this sense, earlier, less complex aquatic foraging behaviors could be absent from the archeological record due to lack of material remains (e.g., tools). As such, while the usage of taphonomic, lithic and observational data has promoted significant advances in our knowledge about the intentional use of aquatic resource in non-human primates and hominins, further empirical evidence is needed (1) to better detect intentionality in the use of aquatic resources in non-human primates, (2) to reconstruct the technological patterns of the emergence of this behavior, (3) to determine which early Pleistocene hominins intentionally exploited aquatic resources and finally (4) to allow for comparisons between the aquatic resource consumption patterns of non-human primates and hominins.
Aquatic Foods and Hominin Brain Size
Coastal, riverine and lacustrine ecosystems are complex and fluctuating environments, in which the availability of resources may be affected by tidal regimes, weather and seasonality. Thus, as is the case in other environments with rapidly changing conditions, primates inhabiting such areas are likely to mobilize cognitive skills that reflect a high level of behavioral flexibility in their foraging decisions (Trapanese et al., 2019; Garcia et al., 2021). In this sense, foraging in aquatic environments could imply a diversity of behavioral responses ranging from simple reflexes to more complex mental representations of goals, their values and the potential courses of action required to make decisions and optimize energetic balance in a social context. At the cognitive level, these decision-making processes are based on a set of skills—for example, episodic memory, planning and value-based decision making—which allow individuals to optimize the outcomes of their actions both at the individual and collective levels (Garcia et al., 2021). Considering the putative benefits of the consumption of aquatic foods for brain development and function (Joordens et al., 2014; Hohmann et al., 2019), many authors have proposed a potential link between the exploitation of these food sources and the evolution of brain size and cognition in hominins (e.g., Marean et al., 2007; Archer et al., 2014; Russon et al., 2014; Will et al., 2019).
In this context, while there is an overlap between the onset of aquatic resource exploitation and an increase in hominin encephalization (Braun et al., 2010; Joordens et al., 2014), it seems unlikely that the former would have triggered the latter for a variety of reasons. Firstly, intensive use of aquatic resources does not necessarily depend on high technological prowess (Carlson and Kingston, 2007; Cunnane et al., 2007; Archer et al., 2014; Russon et al., 2014). As such, the extent to which an increase in reliance on aquatic foraging behavior would generate selective advantages for larger brains remains unclear. Early hominins such as Ardipithecus and some species of Australopithecus. would have likely lived in similar habitats as chimpanzees, the latter being known to exploit aquatic resources in forested environments (Koops et al., 2019). Considering the similarities in brain size between early hominins and chimpanzees (Klein, 2009; Carlson et al., 2011), as well as similarities between the dietary needs of chimpanzees and humans for some brain-selective nutrients (e.g., iodine; Mano et al., 1987), it is possible to hypothesize that these early hominins may have also exploited aquatic resources (Koops et al., 2019), albeit without the onset of the brain expansion process observed only in later hominins. However, this hypothesis strongly relies on nutritional data, and the knowledge about which ecological conditions can contribute to the onset of aquatic resource exploitation remains scarce (Koops et al., 2019). Secondly, the pattern observed in aquatic foraging non-human primates is that of the adaptation of previous terrestrial foraging tool-use behavior and skills into aquatic foraging (Shumaker et al., 2011; Russon et al., 2014). Indeed, by the time when the first evidence for intensive aquatic resource use in hominins is recognized (ca. 1.95 Ma; Braun et al., 2010; Archer et al., 2014), hominins had already been making and using stone-tools for over a million years (McPherron et al., 2010; Harmand et al., 2015). Comparatively, by the time that the first evidence for tool-assisted aquatic resource exploitation appears in the archeological record of AMHs (ca. 164 ka in South Africa; Marean et al., 2007), Neanderthals and AMHs had already been engaging in complex forms of social hunting for over 100 ka (ca. 300 ka in Europe, Conard et al., 2020; and between 259 and 125 ka in South Africa, Bamford and Henderson, 2003). Carrying out these hunting practices, in turn, would depend on complex planning skills associated with the manufacturing of tools (e.g., wooden spears), and involving a variety of cognitive mechanisms such as working memory, context-specific goal value representations and the computation of trade-offs associated with costs and benefits (Garcia et al., 2021). In this sense, the onset of aquatic resource exploitation would not necessarily trigger an increase in cognitive complexity and brain size, as it may instead derive from the behavioral plasticity associated with already existing cognitive skills, expressed in a process of adaptive decision-making. Accordingly, we suggest that the emergence of aquatic resource use per se, as well as the onset of tool-assisted aquatic resource exploitation can be more parsimoniously explained as the product of a local shift in foraging cost/benefit trade-offs favoring aquatic over terrestrial resources, followed by a behavioral adaptation that optimized energetic and nutritional balance in this novel ecological context. This shift in cost-benefit trade-offs, in turn, could have been induced by a variety of locally specific factors, such as (i) seasonality (e.g., aquatic foods as fallback resources; Stewart, 2010; Archer et al., 2014), (ii) technological advancement (e.g., the repurposing of previously used skills and tools; Shumaker et al., 2011; Russon et al., 2014), (iii) physiological changes (e.g., bigger brains requiring more efficient energetic intake and more brain-selective nutrients; Navarrete et al., 2011; Potts, 2011; Joordens et al., 2014; Hohmann et al., 2019), and (iv) demographic conditions (e.g., higher benefits of aquatic resource exploitation for females and immature individuals; Koops et al., 2019).
Moreover, when considering the spatial, technological and demographic specificity of the emergence of aquatic foraging behavior in some of our closest non-human relatives (Russon et al., 2014), any overarching explanation seeking to causally link the local emergence of such behavior with a subsequent sharp development of the global cognitive abilities and brain expansion of a species seems implausible. Likewise, Joordens et al. (2014) rule out the possibility of a driving role of aquatic resource consumption on the expansion of the human brain. According to these authors, if the early evolution of larger brains in Homo was driven by aquatic resource exploitation, we would expect to see a similar pattern in the context of other species that shifted toward foraging for these resources. In this context, while there is evidence that, in some animal groups, aquatic foraging species have the largest absolute brain size (e.g., herpestids, mustelids, and procyonids; Shabel, 2010), and that a shift toward aquatic foraging in cetaceans is linked to brain expansion (Marino, 2007; Xu et al., 2012), the magnitude of these associations is not comparable to the increase in brain size and cognition complexity as seen in hominins, and as such more factors are needed to explain this evolutionary transition (Joordens et al., 2014). In addition, the association between aquatic foraging and brain size does not seem hold true within non-human primate groups either, where the brains of some species that forage intensively for aquatic resources (e.g., long-tailed macaques; Tan et al., 2015) are not enlarged when compared with those of other closely related terrestrial foraging species (Navarrete et al., 2018).
Nevertheless, the exploitation of aquatic resources could have had an important role as a facilitator in the evolution of brain size and complex cognition in hominins, potentially providing a crucial fuel for our encephalization when combined with the consumption of meat (Kyriacou, 2017; Hohmann et al., 2019). Indeed, studies on human nutritional requirements provide evidence that even the consumption of a small amount of aquatic fauna could provide enough LC-PUFAs and other brain-essential micronutrients to meet daily nutritional needs of hominins (Kyriacou et al., 2014, 2016). In this sense, some researchers have hypothesized that the evolution of enlarged fat deposits in humans, for example, could be linked with an increase in aquatic resource exploitation, with adipose tissue providing an efficient way to store a surplus of consumed LC-PUFAs, and as such serving as a buffer ensuring a steady supply of brain-essential nutrients in fluctuating environments (Joordens et al., 2014). This buffering effect, in turn, could have been essential to guarantee proper cognitive function in a large part of the population during food shortage events, and particularly for individuals with decreased access to game meat, such as females and developing immature individuals (Koops et al., 2019; Will et al., 2019; Lombard and Kyriacou, 2020). As such, aquatic resource exploitation could bring about both proximate (e.g., nutritional and energetic) and ultimate (e.g., increased survival and fecundity) adaptive benefits, potentially conferring individuals or groups of individuals that engage in this behavior a selective advantage (Will et al., 2019).
In non-human primates, however, such innovative behaviors are more likely to be lost over time, either through a cessation in the display of an innovative behavior by an innovator or through the direct loss of innovator individuals (Perry et al., 2003; Nishida et al., 2009; Russon et al., 2014). Indeed, data from recent studies in captive primates suggested that besides social learning, individual learning may also influence the appearance of some behaviors such as tool-use (Bandini and Tennie, 2017, 2018, 2019; Bandini et al., 2021). Notably, in the context of aquatic foraging, Bandini and Tennie (2017) demonstrated that captive naïve chimpanzees can independently express algae-scooping behavior if provided with the necessary materials without any social learning involved. These results also suggested that while tested naïve individuals had the inherent cognitive skills to engage in algae-scooping, the emergence of such behavior was dependent on a transition to a new environmental context in which the motivation for them to do so was present (Bandini and Tennie, 2017). Thus, while there is evidence that aquatic exploitation behaviors can be socially learned in the wild (Russon et al., 2014; Tan, 2017), this does not necessarily mean that they will be socially learned, and neither that innovative behaviors will be successfully transmitted across generations and become local traditions.
As such, we hypothesize that observed disparities in the complexity and intensity with which humans and non-human primates exploit aquatic resources may have largely emerged from our increased capacity for retention and improvement of innovations over time through cultural transmission. In this sense, by increasing our dietary breadth and by fueling population growth through increased survival and fecundity (Will et al., 2019), the emergence and intensification of aquatic foraging behavior in hominins could have broadened the diversity of social and environmental contexts to which hominins would have to apply their decision-making skills. In turn, this process would have created novel cognitive challenges requiring more effective decision-making in these more complex ecological contexts. Therefore, while the exploitation of aquatic resources may not have triggered the onset of hominin brain expansion, it could have contributed to an intensification of the selection for more advanced global cognitive skills translated into larger brains. These larger brains, in turn, would require an increase in energetic intake (Navarrete et al., 2011; Potts, 2011), and especially of brain-selective nutrients, which tend to be abundant in aquatic foods (Joordens et al., 2014; Hohmann et al., 2019). As such, this process could then lead to a shift in the cost/benefit ratio of foraging on aquatic foods by increasing the payoff of this activity. In this context, given the strategic and non-random nature of social learning, this increase in payoff would selectively favor the social spread of innovations that optimize aquatic resource exploitation (Laland, 2004; Rendell et al., 2011; Whitehead et al., 2019). Additionally, this could have also influenced the selection for physiological traits that allow for better storage of important brain nutrients such as LC-PUFAs (Joordens et al., 2014). Finally, in areas where aquatic resources are abundant and diverse, these adaptations could fuel additional stable population growth and promote further expansion in the cultural repertoire of hominins (Kolodny et al., 2016; Will et al., 2019). Markedly, this adaptive scenario would only be possible if behavioral innovations associated to the optimization of aquatic resource exploitation could be successfully transmitted between individuals and across generations, which would depend on positive selection for traits that increase reliance on social learning and culture in such populations. This interpretation is consistent with studies suggesting a link between coastal adaptation and selective pressures for prosociality in hominins (Marean, 2014, 2016) and also with evidence from modeling research demonstrating that even small-scale population growth can lead to disproportional effects on cultural accumulation through a positive feedback loop process (Creanza et al., 2017). Still, in order to verify whether the relationship between aquatic resource exploitation, cognition and brain expansion in hominins constitutes an example of gene-culture coevolution, further empirical evidence is needed.
Implications for Hominin Spatial Ecology
While a variety of cultural behaviors has been documented in non-human primate species (Whiten, 2011; Whiten and van de Waal, 2017), the extent to which humans are able to accumulate and disseminate cultural changes over generations is unmatched (Mesoudi and Thornton, 2018). This increased capacity for cumulative cultural evolution is thought to have allowed hominins to inhabit a wide range of ecosystems, making use of a remarkable diversity of survival tools and techniques developed over time (Boyd and Richerson, 1996). As such, the extent to which hominins have been able to adapt to inhabiting coastal landscapes is unparalleled among primates, albeit there are examples of secondary adaptations to aquatic lifestyles in a variety of vertebrate species (Mazin and de Buffrénil, 2001; Houssaye and Fish, 2016; Davis, 2019). In this sense, coastally adapted populations exhibit a set of behavioral traits which allowed them to strategically occupy coastal and near-coastal zones in settlement systems with reduced mobility, and where the systematic consumption of marine resources would take place in accordance with tidal regimes and fluctuating food availability (Marean, 2014; Will et al., 2019). Thus, true coastal adaptation would not only involve the habitual consumption of aquatic resources, but also the incorporation of marine resources into the local material culture and the display of specialized technologies that allow for the more efficient exploitation of such resources (Will et al., 2019). Similarly, some non-human primate species that intensively exploit aquatic resources also exhibit other adaptations that are accessory to aquatic foraging, such as agile swimming in rhesus macaques (Mallick, 2019) and the intensification of percussive tool use in bearded capuchins (Santos et al., 2019b). As such, the success of primates—hominins included—in colonizing flooded habitats seems to depend on their capacity to efficiently exploit aquatic resources, which likely involves a variety of complementary adaptations.
In this context, a growth in the effectiveness of coastal resource extraction combined with an increase in sedentism can cause the depletion rate of some high-quality marine food stocks to accelerate. At coastal archeological sites in South Africa, for example, there is evidence for a decline in shellfish size from middle (MSA; ∼120–60 ka) to the late stone age (LSA; ∼12–1 ka) deposits, presumably as a result of hominin impact (Klein and Steele, 2013). A decrease in limpet size associated to late Neanderthal exploitation has also been documented during the Upper Paleolithic (UP) in Europe, which may potentially be associated to increases in population growth and density during that period (Stiner et al., 1999). In addition, modeling research on cultural evolution shows that, through changing resource availability, technological development may result in further population growth (Kolodny et al., 2016), which in turn could expedite the reduction in the local availability of high-quality marine foods (Klein et al., 2004; Niespolo et al., 2021). In this context, a decrease in size and quality of marine stocks could cause a reduction in the yield, and consequently also in the benefit of foraging on local aquatic resources. Moreover, higher population sizes and densities would also generate an increase in intraspecific competition for such resources, raising the costs and risks associated to acquiring them. In response to a shift in the cost-benefit relation of foraging in marine foods, populations would have two ways in which to flexibly adapt: (i) through further technological development that would increase food access (e.g., shifting toward the consumption of more agile, harder-to-catch prey; Stiner et al., 1999) or (ii) through territorial expansion or migration to other inland or coastal areas (e.g., Walter et al., 2000).
Indeed, it has been proposed that some of the earlier terrestrial hominin dispersals out of Africa and into Eurasia have been influenced by food availability and facilitated by similarities in access to resources across sites without predetermination (Prat, 2018). Comparatively, a similar process could have pushed for hominin dispersal along the coast and through the sea. This is specially the case considering similarities in resource availability between some coastal zones, and the presence of intricate oceanic current systems in areas with higher density of islands which may further facilitate dispersal (Erlandson, 2017). In this sense, this facilitation may have been crucial to the success of such hominin dispersals, especially considering that water bodies often also represent substantial biogeographic barriers in the context of non-human primate spatial ecology (e.g., Harcourt and Wood, 2012; Boubli et al., 2015; Shekelle et al., 2019). Notably, a variety of authors has proposed the existence of migration corridors along coastal zones which would likely not constitute substantial ecological barriers for oceanic movement, e.g., the kelp highway along the Pacific Rim (Erlandson et al., 2007, 2015) and the mangrove rim along the Indian Ocean (Kathiresan and Rajendran, 2005). Therefore, dispersals along such coastal migration corridors could have been associated with low technological costs and reduced failure risks, particularly for populations with a high degree of adaptedness to a coastal lifestyle.
Accordingly, a scenario of progressive colonization of different coastal ecosystems could have pushed hominins to display adaptive behaviors in response to novel subsistence challenges and opportunities, likely involving the cultural accumulation and diversification of novel technologies used to exploit aquatic resources and of technologies necessary for water crossings (Gaffney, 2021). Incrementally, this could culminate in the development of maritime adaptations by some human populations, involving intensive use of boats and other sea-going vessels, off-shore exploitation of marine resources and more frequent long-distance sea traveling (Will et al., 2019). Within this framework, an increase in complexity of coastal occupations and water crossings would also impose novel relevant cognitive challenges to hominins (Leppard, 2015b; Leppard and Runnels, 2017). Unlike passive dispersal, strategic longer distance water crossings would require the use of composite technologies, further capacity for projecting future actions to achieve to out-of-sight goals and an ability to engage in more complex cooperative planning interactions (Leppard, 2015a). In this sense, this process could, at least partially, account for observed differences in frequency and intensity of sea dispersals between early and late Pleistocene hominins, with the latter exhibiting higher adaptedness to coastal ecosystems and being more cognitively ready to flexibly adapt to novel environments. Together, such adaptations would also have putatively allowed populations to increase the yield of their foraging behavior and to diversify the array of readily available aquatic resources. In turn, this could confer such populations with a further adaptive advantage, allowing for the progressive increases in efficiency and spread of demographic expansions via water crossings observed in the paleoanthropological record from the Early to the Late Pleistocene (Gaffney, 2021).
General Discussion and Perspectives
Although the consumption of aquatic resources has been reported in populations of several non-human primate species (Stewart et al., 2008; Kempf, 2009; Stewart, 2010; Russon et al., 2014; Koops et al., 2019), little is currently known about the ecological conditions that favor the emergence and permanence of this behavior in some—but not all—populations that have access to aquatic resources in the wild. In part, this has to do with the lack of studies tracking the emergence of aquatic foraging behavior in a wild setting, of which Russon et al. (2014) is an exception, having documented the appearance of fish-eating behavior among ex-captive, rehabilitated orangutans in Borneo. Evidence from this study, in turn, seems to be in line with Stewart’s (2010) hypothesis that the emergence of complex aquatic resource exploitation would be preceded by simpler behavioral precursors, with the transition from simpler to more complex behaviors gradually developing over time. Still, when considering the myriad of local specificities that may affect aquatic foraging behavior in non-human primates—such as local food availability (e.g., Matsuzawa, 2019), demography (e.g., Koops et al., 2019) and employed techniques (e.g., Sakamaki, 1998; Boesch et al., 2017)—any individual explanation seeking to globally explain the emergence of such behaviors is unlikely to offer a high explanatory power in a local context. Instead, future research may benefit from adopting a comparative approach for studying why non-human primates may favor aquatic over terrestrial resources in some ecological contexts, but not in others. By doing this, researchers would be able to get a better understanding of what cognitive processes may cause some non-human primate populations to transition from an entirely terrestrial diet to one that also includes aquatic foods. Besides, this approach could also aid in assessing the extent to which this transition may be favored by local cost-benefit trade-offs and how such behaviors perish or persist over time across different populations and species.
Regardless of the mechanisms behind their emergence, the presence of widespread and intensive aquatic foraging behaviors in some non-human primate populations (e.g., long-tailed macaques; Gumert et al., 2009; Gumert and Malaivijitnond, 2012; Tan et al., 2015; Tan, 2017) hints at an important role played by these items in their local foraging ecology. According to the optimal foraging theory, this should be specially the case for species that forage for aquatic resources intentionally, as their observed foraging strategies are expected to have resulted from cognitive processes that optimize the energetic balance in dynamic ecological contexts (Charnov, 1976; Stephens and Krebs, 1986). However, considering that the available evidence for intentionality in aquatic resource exploitation by non-human primates mainly comes from planning behavior related to tool selection and use, our current record of how many species consume aquatic resources intentionally likely represents an underestimation. As such, the use of other proxies (see below) to investigate intentionality in non-human primate aquatic foraging is central to future research, and particularly in the case of species that do not use tools when foraging in coastal, riverine or lacustrine environments. In the context of terrestrial foraging, for example, one method that has been proven useful for determining intentionality has been the use of a combination of movement, environmental and behavioral data to study the decision-making processes of primates in the wild on a contextual ecological basis, which in turn helps shed light on what information individuals are acting on and what cognitive skills are associated with their observed behaviors (e.g., Janmaat et al., 2014; Ban et al., 2016; Salmi et al., 2020; but see Janmaat et al., 2021 for a review of this method). By applying such novel methods, researchers could be able to determine, for example, whether the aquatic foraging behavior of coastal-dwelling primates are part of a plan (e.g., if they schedule their foraging to tides) or how the seasonal availability of different aquatic resources spatiotemporally affects their behavior. In addition, this could also allow for determining whether aquatic foods may truly be preferred to terrestrial ones on some environmental contexts, or whether they may simply represent fallback resources in times of terrestrial food shortages.
Moreover, with hominins being the only primate group in which true coastal adaptation has evolved (Will et al., 2019)—and consequently also the one in which the most intensive and complex exploitation of aquatic resources can be observed—such diversification in methodologies could be crucial for a better appraisal of currently existing hypotheses on how the consumption of aquatic resources may be linked to the evolution of the hominin brain and cognitive skills. Additionally, in light of the putative cultural nature of coastal adaptation in hominins (Marean, 2014; Will et al., 2019), future studies could also benefit from assessing foraging decision-making processes at the collective level and the social transmission of behaviors between individuals as an important factor influencing the development of aquatic foraging in extinct hominins. This approach would be of particular value when considering the patterns of aquatic resource exploitation in AMHs, as our species is not only the one for which the largest body of evidence for coastal adaptedness is available, but also the only one in which advanced maritime adaptations have been detected (Erlandson, 2001, 2017; O’Connor et al., 2011; Fujita et al., 2016)—the latter likely being unique in the complexity of cognitive skills and collective decision-making efforts required to achieve it (Leppard, 2015a,b; Leppard and Runnels, 2017). That said, the relatively lower amount of evidence for a high degree of coastal and maritime adaptedness in other hominin species (e.g., Neanderthals and Homo erectus) may also be a product of the loss of coastal archeological remains of such hominins due to the post-glacial sea level rises during the Holocene (Bailey et al., 2007; Erlandson, 2017). Finally, an important knowledge gap remains with respect to how the exploitation of aquatic resources in hominins may have been linked to the regular use of fire and cooking. Given the centrality of cooking in the evolutionary context of hominin foraging ecology (Carmody and Wrangham, 2009), the association between this activity and aquatic foraging becomes specially important when considering that the detection of widespread regular use of fire in the paleoanthropological record (ca. 400–350 ka) predates the earliest available evidence for the appearance of intensive aquatic resource exploitation in AMHs—ca. 164 ka—by over 150,000 years (Marean et al., 2007; MacDonald et al., 2021). Nevertheless, accurate detection of burning traces on aquatic fauna remains can be a challenging and inconclusive process (Guillaud et al., 2021), and more evidence is still needed in order to better assess when hominins first started cooking aquatic foods.
In summary, by adaptively responding to novel ecological challenges linked to local changes in foraging cost-benefit trade-offs, hominin populations have been able to strategically expand their niches through diversifying their diets to include a wide variety of aquatic resources. Over time, the intensification of aquatic resource exploitation combined with higher sedentism and population growth may have led to the development of long-lasting coastal adaptations in AMHs, later followed by maritime adaptations involving advanced sea-faring technologies and deliberate water crossings toward new geographical regions (Erlandson, 2017; Will et al., 2019). Markedly, these ecological shifts were likely linked to key cognitive skills in hominins allowing for a higher level of adaptive flexibility (Leppard, 2015b), which when combined with an increased capacity for accumulating cultural changes over time (Mesoudi and Thornton, 2018), may have translated into an unmatched readiness to adapt to novel ecological niches (Boyd and Richerson, 1996; Wright et al., 2010; Gaffney, 2021). Thus, it is clear that the unique ways in which some coastal populations have been able to exploit aquatic resources most likely had a deep impact on the ecological trajectory of hominins. In this sense, developing a better comparative understanding of how such behavior can emerge in hominins and non-human primates may have profound implications for how we interpret hominin evolution.
Author Contributions
CG and SP provided the initial ideas for this review. GC wrote the body of the manuscript. All authors have participated in the writing, conception and discussion of ideas presented in the final version of this review.
Funding
The publication of this review was funded by the ANR HOMTECH (ANR-17-CE27-0005). This work was supported by the Centre National de la Recherche Scientifique (MITI CNRS 80 PRIME, Flexidiet Project).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank the editor and the two reviewers for their constructive comments.
References
Archer, W., Braun, D. R., Harris, J. W. K., McCoy, J. T., and Richmond, B. G. (2014). Early pleistocene aquatic resource use in the Turkana Basin. J. Hum. Evol. 77, 74–87. doi: 10.1016/j.jhevol.2014.02.012
Bailey, G. N., Flemming, N. C., King, G. C. P., Lambeck, K., Momber, G., Moran, L. J., et al. (2007). Coastlines, submerged landscapes, and human evolution: the Red Sea Basin and the Farasan Islands. J. Isl. Coast. Archaeol. 2, 127–160. doi: 10.1080/15564890701623449
Bamford, M. K., and Henderson, Z. L. (2003). A reassessment of the wooden fragment from Florisbad, South Africa. J. Archaeol. Sci. 30, 637–650. doi: 10.1016/S0305-4403(02)00245-5
Ban, S. D., Boesch, C., N’Guessan, A., N’Goran, E. K., Tako, A., and Janmaat, K. R. L. (2016). Taï chimpanzees change their travel direction for rare feeding trees providing fatty fruits. Anim. Behav. 118, 135–147. doi: 10.1016/j.anbehav.2016.05.014
Bandini, E., and Tennie, C. (2017). Spontaneous reoccurrence of “scooping”, a wild tool-use behaviour, in naïve chimpanzees. PeerJ 5:e3814. doi: 10.7717/peerj.3814
Bandini, E., and Tennie, C. (2018). Naive, captive long-tailed macaques (Macaca fascicularis fascicularis) fail to individually and socially learn pound-hammering, a tool-use behaviour. R. Soc. Open Sci. 5:171826. doi: 10.1098/rsos.171826
Bandini, E., and Tennie, C. (2019). Individual acquisition of “stick pounding” behavior by naïve chimpanzees. Am. J. Primatol. 81:e22987.
Bandini, E., Grossmann, J., Funk, M., Albiach-Serrano, A., and Tennie, C. (2021). Naïve orangutans (Pongo abelii and Pongo pygmaeus) individually acquire nut-cracking using hammer tools. Am. J. Primatol. 83, 1–15. doi: 10.1002/ajp.23304
Becker, E. W. (2007). Micro-algae as a source of protein. Biotechnol. Adv. 25, 207–210. doi: 10.1016/j.biotechadv.2006.11.002
Bedoya-Perez, M. A., Carthey, A. J. R., Mella, V. S. A., McArthur, C., and Banks, P. B. (2013). A practical guide to avoid giving up on giving-up densities. Behav. Ecol. Sociobiol. 67, 1541–1553. doi: 10.1007/s00265-013-1609-3
Berger, J. R., Waisman, E., and Weisman, B. (1997). Creutzfeldt-jakob disease and eating squirrel brains. Lancet 350:642. doi: 10.1016/s0140-6736(05)63333-8
Boesch, C. (1994). Cooperative hunting in wild chimpanzees. Anim. Behav. 48, 653–667. doi: 10.1006/anbe.1994.1285
Boesch, C., and Boesch-Achermann, H. (2000). The Chimpanzees of the Taï Forest: Behavioural Ecology and Evolution. Oxford: Oxford University Press.
Boesch, C., Kalan, A. K., Agbor, A., Arandjelovic, M., Dieguez, P., Lapeyre, V., et al. (2017). Chimpanzees routinely fish for algae with tools during the dry season in Bakoun, Guinea. Am. J. Primatol. 79, 1–7. doi: 10.1002/ajp.22613
Boesch, C., Wittig, R., Crockford, C., Vigilant, L., Deschner, T., and Leendertz, F. (2019). The Chimpanzees of the Taï Forest: 40 Years of Research. Cambridge: Cambridge University Press.
Boubli, J. P., Ribas, C., Lynch Alfaro, J. W., Alfaro, M. E., da Silva, M. N. F., Pinho, G. M., et al. (2015). Spatial and temporal patterns of diversification on the amazon: a test of the riverine hypothesis for all diurnal primates of Rio Negro and Rio Branco in Brazil. Mol. Phylogenet. Evol. 82, 400–412. doi: 10.1016/j.ympev.2014.09.005
Boyd, R., and Richerson, J. (1996). Why culture is common, but cultural evolution is rare. Proc. Br. Acad. 88, 77–93.
Braun, D. R., Harris, J. W. K., Levin, N. E., McCoy, J. T., Herries, A. I. R., Bamford, M. K., et al. (2010). Early hominin diet included diverse terrestrial and aquatic animals 1.95 Ma in East Turkana, Kenya. Proc. Natl. Acad. Sci. U.S.A. 107, 10002–10007. doi: 10.1073/pnas.1002181107
Brosnan, S. F., Salwiczek, L., and Bshary, R. (2010). The interplay of cognition and cooperation. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 365, 2699–2710. doi: 10.1098/rstb.2010.0154
Busse, C. D. (1977). Chimpanzee predation as a possible factor in the evolution of red colobus monkey social organization. Evolution 31, 907–911. doi: 10.1111/j.1558-5646.1977.tb01087.x
Byrne, R. W. (1996). Machiavellian intelligence. Evol. Anthropol. 5, 172–180. doi: 10.1002/(SICI)1520-650519965:5<172::AID-EVAN6<3.0.CO;2-H
Carlson, B. A., and Kingston, J. D. (2007). Docosahexaenoic acid, the aquatic diet, and hominin encephalization: difficulties in establishing evolutionary links. Am. J. Hum. Biol. 19, 132–141. doi: 10.1002/ajhb.20579
Carlson, K. J., Stout, D., Jashashvili, T., de Ruiter, D. J., Tafforeau, P., Carlson, K., et al. (2011). The endocast of MH1, Australopithecus sediba. Science 333, 1402–1407. doi: 10.1126/science.1203922
Carmody, R. N., and Wrangham, R. W. (2009). The energetic significance of cooking. J. Hum. Evol. 57, 379–391. doi: 10.1016/j.jhevol.2009.02.011
Charnov, E. L. (1976). Optimal foraging, the marginal value theorem. Theor. Popul. Biol. 9, 129–136. doi: 10.1016/0040-5809(76)90040-x
Cheney, D. L., Seyfarth, R. M., Fischer, J., Beehner, J., Bergman, T., Johnson, S. E., et al. (2004). Factors affecting reproduction and mortality among baboons in the Okavango Delta, Botswana. Int. J. Primatol. 25, 401–428. doi: 10.1023/B:IJOP.0000019159.75573.13
Conard, N. J., Serangeli, J., Bigga, G., and Rots, V. (2020). A 300,000-year-old throwing stick from Schöningen, northern Germany, documents the evolution of human hunting. Nat. Ecol. Evol. 4, 690–693. doi: 10.1038/s41559-020-1139-0
Cordain, L., Watkins, B. A., Florant, G. L., Kelher, M., Rogers, L., and Li, Y. (2002). Fatty acid analysis of wild ruminant tissues: evolutionary implications for reducing diet-related chronic disease. Eur. J. Clin. Nutr. 56, 181–191. doi: 10.1038/sj.ejcn.1601307
Cortés-Sánchez, M., Morales-Muñiz, A., Simón-Vallejo, M. D., Lozano-Francisco, M. C., Vera-Peláez, J. L., Finlayson, C., et al. (2011). Earliest known use of marine resources by neanderthals. PLoS One 6:e24026. doi: 10.1371/journal.pone.0024026
Cowlishaw, G. (1994). Vulnerability to predation in baboon populations. Behaviour 131, 293–304. doi: 10.1163/156853994X00488
Creanza, N., Kolodny, O., and Feldman, M. W. (2017). Greater than the sum of its parts? Modelling population contact and interaction of cultural repertoires. J. R. Soc. Interface 14:171. doi: 10.1098/rsif.2017.0171
Cunnane, S. C., Plourde, M., Stewart, K., and Crawford, M. A. (2007). Docosahexaenoic acid and shore-based diets in hominin encephalization: a rebuttal. Am. J. Hum. Biol. Off. J. Hum. Biol. Counc. 19, 578–581. doi: 10.1002/ajhb.20673
Cunnane, S., and Stewart, K. (2010). Human Brain Evolution: The Influence of Freshwater and Marine Food Resources. Hoboken, NJ: John Wiley & Sons.
Cunningham, E., and Janson, C. (2007). A socioecological perspective on primate cognition, past and present. Anim. Cogn. 10, 273–281. doi: 10.1007/s10071-007-0078-3
Dalsgaard, N. J. (2002). Prion diseases. An overview. APMIS 110, 3–13. doi: 10.1034/j.1600-0463.2002.100102.x
Davis, R. W. (2019). Marine Mammals: Adaptations for an Aquatic Life. Berlin: Springer, doi: 10.1007/978-3-319-98280-9
Deaner, R. O., Isler, K., Burkart, J., and van Schaik, C. (2007). Overall brain size, and not encephalization quotient, best predicts cognitive ability across non-human primates. Brain. Behav. Evol. 70, 115–124. doi: 10.1159/000102973
DeCasien, A. R., and Higham, J. P. (2019). Primate mosaic brain evolution reflects selection on sensory and cognitive specialization. Nat. Ecol. Evol. 3, 1483–1493. doi: 10.1038/s41559-019-0969-0
DeCasien, A. R., Williams, S. A., and Higham, J. P. (2017). Primate brain size is predicted by diet but not sociality. Nat. Ecol. Evol. 1:112. doi: 10.1038/s41559-017-0112
Devos, C., Gatti, S., and Levréo, F. (2002). New record of algae feeding and scooping by Pan t. troglodytes at Lokoué Bai in Odzala National Park, Republic of Congo. Pan Africa News 9, 19–21.
Domachowske, J., and Suryadevara, M. (2020). Prion Disease in Clinical Infectious Diseases Study Guide: A Problem Based Approach. Cham: Springer International Publishing, 201–203. doi: 10.1007/978-3-030-50873-9_32
Santos, R. R., Sousa, A. A., de, Fragaszy, D. M., and Ferreira, R. G. (2019b). “The role of tools in the feeding ecology of bearded capuchins living in mangroves,” in Primates in Flooded Habitats: Ecology and Conservation, eds A. A. Barnett, I. Matsuda, and K. Nowak (Cambridge: Cambridge University Press), 59–63.
Santos, R. R., Bridgeman, L. L., Supriatna, J., Siregar, R., Winarni, N., and Salmi, R. (2019a). “Behavioural ecology of mangrove primates and their neighbours,” in Primates in Flooded Habitats: Ecology and Conservation, eds A. A. Barnett, I. Matsuda, and K. Nowak (Cambridge: Cambridge University Press), 124–133.
Dunbar, R. I. M. (1998). The social brain hypothesis. Evol. Anthropol. 6, 178–190. doi: 10.1002/(SICI)1520-650519986:5<178::AID-EVAN5<3.0.CO;2-8
Dunbar, R. I. M., and Shultz, S. (2007). Evolution in the social brain. Science 317, 1344–1347. doi: 10.1126/science.1145463
Erlandson, J. M. (2001). The archaeology of aquatic adaptations: paradigms for a new millennium. J. Archaeol. Res. 9, 287–350. doi: 10.1023/A:1013062712695
Erlandson, J. M. (2017). “Coastlines, marine ecology, and maritime dispersals in human history,” in Human Dispersal and Species Movement: From Prehistory to the Present, eds N. Boivin, R. Crassard, and M. Petraglia (Cambridge University Press), 147–163. doi: 10.1017/9781316686942.007
Erlandson, J. M., Braje, T. J., Gill, K. M., and Graham, M. H. (2015). Ecology of the kelp highway: did marine resources facilitate human dispersal from Northeast Asia to the Americas? J. Isl. Coast. Archaeol. 10, 392–411. doi: 10.1080/15564894.2014.1001923
Erlandson, J. M., Graham, M. H., Bourque, B. J., Corbett, D., Estes, J. A., and Steneck, R. S. (2007). The kelp highway hypothesis: marine ecology, the coastal migration theory, and the peopling of the Americas. J. Isl. Coast. Archaeol. 2, 161–174. doi: 10.1080/15564890701628612
Fahy, G. E., Richards, M., Riedel, J., Hublin, J.-J., and Boesch, C. (2013). Stable isotope evidence of meat eating and hunting specialization in adult male chimpanzees. Proc. Natl. Acad. Sci. U.S.A. 110, 5829–5833. doi: 10.1073/pnas.1221991110
Friant, S., Brown, K., Saari, M. T., Segel, N. H., Slezak, J., and Goldberg, T. L. (2015). Lung fluke (Paragonimus africanus) infects Nigerian red-capped mangabeys and causes respiratory disease. Int. J. Parasitol. Parasites Wildl. 4, 329–332. doi: 10.1016/j.ijppaw.2015.08.003
Fujita, M., Yamasaki, S., Katagiri, C., Oshiro, I., Sano, K., Kurozumi, T., et al. (2016). Advanced maritime adaptation in the western Pacific coastal region extends back to 35,000-30,000 years before present. Proc. Natl. Acad. Sci. U.S.A. 113, 11184–11189. doi: 10.1073/pnas.1607857113
Gaffney, D. (2021). Pleistocene water crossings and adaptive flexibility within the homo genus. J. Archaeol. Res. 29, 255–326. doi: 10.1007/s10814-020-09149-7
Galat, G., and Galat-Luong, A. (1976). La colonisation de la mangrove par Cercopithecus aethiops sabaeus au Sénégal. La Terre La Vie Rev. d’Ecol. Appl. 30, 3–30.
Garcia, C., Bouret, S., Druelle, F., and Prat, S. (2021). Balancing costs and benefits in primates: ecological and palaeoanthropological views. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 376:20190667. doi: 10.1098/rstb.2019.0667
Gilby, I. C., and Wrangham, R. W. (2007). Risk-prone hunting by chimpanzees (Pan troglodytes schweinfurthii) increases during periods of high diet quality. Behav. Ecol. Sociobiol. 61, 1771–1779. doi: 10.1007/s00265-007-0410-6
Gilby, I. C., Machanda, Z. P., O’Malley, R. C., Murray, C. M., Lonsdorf, E. V., Walker, K., et al. (2017). Predation by female chimpanzees: toward an understanding of sex differences in meat acquisition in the last common ancestor of Pan and Homo. J. Hum. Evol. 110, 82–94. doi: 10.1016/j.jhevol.2017.06.015
Gramsch, B., Beran, J., Hanik, S., and Sommer, R. S. (2013). A Palaeolithic fishhook made of ivory and the earliest fishhook tradition in europe. J. Archaeol. Sci. 40, 2458–2463. doi: 10.1016/j.jas.2013.01.010
Green, S. J., Boruff, B. J., Bonnell, T. R., and Grueter, C. C. (2020). Chimpanzees use least-cost routes to out-of-sight goals. Curr. Biol. 30, 4528–4533.e5. doi: 10.1016/j.cub.2020.08.076
Guillaud, E., Béarez, P., Daujeard, C., Defleur, A. R., Desclaux, E., Roselló-Izquierdo, E., et al. (2021). Neanderthal foraging in freshwater ecosystems: a reappraisal of the Middle Paleolithic archaeological fish record from continental Western Europe. Quat. Sci. Rev. 252:106731. doi: 10.1016/j.quascirev.2020.106731
Gumert, M. D., and Malaivijitnond, S. (2012). Marine prey processed with stone tools by burmese long-tailed macaques (Macaca fascicularis aurea) in intertidal habitats. Am. J. Phys. Anthropol. 149, 447–457. doi: 10.1002/ajpa.22143
Gumert, M. D., Kluck, M., and Malaivijitnond, S. (2009). The physical characteristics and usage patterns of stone axe and pounding hammers used by long-tailed macaques in the Andaman Sea region of Thailand. Am. J. Primatol. 71, 594–608. doi: 10.1002/ajp.20694
Gumert, M. D., Tan, A., and Malaivijitnond, S. (2019). “Long-tailed macaque stone tool use in intertidal habitats,” in Primates in Flooded Habitats: Ecology and Conservation, eds A. A. Barnett, I. Matsuda, and K. Nowak (Cambridge: Cambridge University Press), 144–147.
Hall, K. R. L. (1962). Numerical data, maintenance activities and locomotion in the wild chacma baboon, Papio ursinus. Proc. Zool. Soc. London 139, 181–220. doi: 10.1111/j.1469-7998.1962.tb01827.x
Hamilton, W. J. III, and Tilson, R. L. (1985). Fishing baboons at desert waterholes. Am. J. Primatol. 8, 255–257. doi: 10.1002/ajp.1350080308
Harcourt, A. H., and Wood, M. A. (2012). Rivers as barriers to primate distributions in Africa. Int. J. Primatol. 33, 168–183. doi: 10.1007/s10764-011-9558-z
Hardy, B. L., and Moncel, M. H. (2011). Neanderthal use of fish, mammals, birds, starchy plants and wood 125-250,000 years ago. PLoS One 6:e23768. doi: 10.1371/journal.pone.0023768
Harmand, S., Lewis, J. E., Feibel, C. S., Lepre, C. J., Prat, S., Lenoble, A., et al. (2015). 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya. Nature 521, 310–315. doi: 10.1038/nature14464
Harris, L. R., Harris, K. A., and Nel, R. (2019). Food preferences of two sandy beach scavengers with different foraging strategies. Estuar. Coast. Shelf Sci. 219, 120–127. doi: 10.1016/j.ecss.2019.02.001
Head, J., Healy, A., and Nowak, K. (2019). “Primates of african mangroves,” in Primates in Flooded Habitats: Ecology and Conservation, eds A. A. Barnett, I. Matsuda, and K. Nowak (Cambridge: Cambridge University Press), 77–88.
Hill, R. A., and Dunbar, R. I. M. (1998). An evaluation of the roles of predation rate and predation risk as selective pressures on primate grouping behaviour. Behaviour 135, 411–430.
Hohmann, G., Ortmann, S., Remer, T., and Fruth, B. (2019). Fishing for iodine: what aquatic foraging by bonobos tells us about human evolution. BMC Zool. 4:5. doi: 10.1186/s40850-019-0043-z
Houssaye, A., and Fish, F. E. (2016). Functional (secondary) adaptation to an aquatic life in vertebrates: an introduction to the symposium. Integr. Comput. Biol. 56, 1266–1270. doi: 10.1093/icb/icw129
Humle, T., Snowdon, C. T., and Matsuzawa, T. (2009). Social influences on ant-dipping acquisition in the wild chimpanzees (Pan troglodytes verus) of Bossou, Guinea, West Africa. Anim. Cogn. 12(Suppl. 1), S37–S48. doi: 10.1007/s10071-009-0272-6
Humle, T., Yamakoshi, G., and Matsuzawa, T. (2011). “Algae scooping remains a puzzle BT,” in The Chimpanzees of Bossou and Nimba, eds T. Matsuzawa, T. Humle, and Y. Sugiyama (Tokyo: Springer Japan), 117–122. doi: 10.1007/978-4-431-53921-6_12
Humphrey, N. (1976). “The social function of intellect,” in Growing Points in Ethology, eds P. P. G. Bateson and R. A. Hinde (Cambridge: Cambridge univ press), 303–317.
Janmaat, K. R. L., Boesch, C., Byrne, R., Chapman, C. A., Goné Bi, Z. B., Head, J. S., et al. (2016). Spatio-temporal complexity of chimpanzee food: how cognitive adaptations can counteract the ephemeral nature of ripe fruit. Am. J. Primatol. 78, 626–645. doi: 10.1002/ajp.22527
Janmaat, K. R. L., de Guinea, M., Collet, J., Byrne, R. W., Robira, B., van Loon, E., et al. (2021). Using natural travel paths to infer and compare primate cognition in the wild. iScience 24:102343. doi: 10.1016/j.isci.2021.102343
Janmaat, K. R. L., Polansky, L., Ban, S. D., and Boesch, C. (2014). Wild chimpanzees plan their breakfast time, type, and location. Proc. Natl. Acad. Sci. U.S.A. 111, 16343–16348. doi: 10.1073/pnas.1407524111
Jolly, A. (1966). Lemur social behavior and primate intelligence. Science 153, 501–506. doi: 10.1126/science.153.3735.501
Joordens, J. C. A., d’Errico, F., Wesselingh, F. P., Munro, S., de Vos, J., Wallinga, J., et al. (2015). Homo erectus at Trinil on Java used shells for tool production and engraving. Nature 518, 228–231. doi: 10.1038/nature13962
Joordens, J. C. A., Kuipers, R. S., Wanink, J. H., and Muskiet, F. A. J. (2014). A fish is not a fish: patterns in fatty acid composition of aquatic food may have had implications for hominin evolution. J. Hum. Evol. 77, 107–116. doi: 10.1016/j.jhevol.2014.04.004
Kathiresan, K., and Rajendran, N. (2005). Mangrove ecosystems of the Indian Ocean region. Indian J. Mar. Sci. 34, 104–113.
Kempf, E. (2009). Patterns of water use in primates. Folia Primatol. 80, 275–294. doi: 10.1159/000252586
Klein, R. G. (2009). The Human Career: Human Biological and Cultural Origins. Chicago, CA: University of Chicago Press.
Klein, R. G., and Steele, T. E. (2013). Archaeological shellfish size and later human evolution in Africa. Proc. Natl. Acad. Sci. U.S.A. 110, 10910–10915. doi: 10.1073/pnas.1304750110
Klein, R. G., Avery, G., Cruz-Uribe, K., Halkett, D., Parkington, J. E., Steele, T., et al. (2004). The Ysterfontein 1 Middle Stone Age site, South Africa, and early human exploitation of coastal resources. Proc. Natl. Acad. Sci. U.S.A. 101, 5708–5715. doi: 10.1073/pnas.0400528101
Kolodny, O., Creanza, N., and Feldman, M. W. (2016). Game-changing innovations: how culture can change the parameters of its own evolution and induce abrupt cultural shifts. PLoS Comput. Biol. 12:e1005302. doi: 10.1371/journal.pcbi.1005302
Koops, K., Wrangham, R. W., Cumberlidge, N., Fitzgerald, M. A., van Leeuwen, K. L., Rothman, J. M., et al. (2019). Crab-fishing by chimpanzees in the Nimba Mountains, Guinea. J. Hum. Evol. 133, 230–241. doi: 10.1016/j.jhevol.2019.05.002
Kyriacou, K. (2017). Intertidal shellfish as a source of protein and energy for the Middle Stone Age inhabitants of the southwestern Cape and northern Kwazulu-Natal, South Africa. Quat. Int. 438, 30–39. doi: 10.1016/j.quaint.2017.01.032
Kyriacou, K., Blackhurst, D. M., Parkington, J. E., and Marais, A. D. (2016). Marine and terrestrial foods as a source of brain-selective nutrients for early modern humans in the southwestern Cape, South Africa. J. Hum. Evol. 97, 86–96. doi: 10.1016/j.jhevol.2016.04.009
Kyriacou, K., Parkington, J. E., Marais, A. D., and Braun, D. R. (2014). Nutrition, modernity and the archaeological record: coastal resources and nutrition among Middle Stone Age hunter-gatherers on the western Cape coast of South Africa. J. Hum. Evol. 77, 64–73. doi: 10.1016/j.jhevol.2014.02.024
Leppard, T. P. (2015b). The evolution of modern behaviour and its implications for maritime dispersal during the palaeolithic. Cambridge Archaeol. J. 25, 829–846. doi: 10.1017/S0959774315000098
Leppard, T. P. (2015a). Passive dispersal versus strategic dispersal in island colonization by hominins. Curr. Anthropol. 56, 590–595. doi: 10.1086/682325
Leppard, T. P., and Runnels, C. (2017). Maritime hominin dispersals in the Pleistocene: advancing the debate. Antiquity 91, 510–519. doi: 10.15184/aqy.2017.16
Lewis, M. C., and O’Riain, M. J. (2019). “The ecology of chacma baboon foraging in the marine intertidal zone of the Cape peninsula, South Africa,” in Primates in Flooded Habitats: Ecology and Conservation, eds A. A. Barnett, I. Matsuda, and K. Nowak (Cambridge: Cambridge University Press), 148–151. doi: 10.1017/9781316466780.021
Lewis, M. C., and Sealy, J. C. (2018). Coastal complexity: ancient human diets inferred from bayesian stable isotope mixing models and a primate analogue. PLoS One 13:e0209411. doi: 10.1371/journal.pone.0209411
Lewis, M. C., West, A. G., and O’Riain, M. J. (2018). Isotopic assessment of marine food consumption by natural-foraging chacma baboons on the Cape peninsula, South Africa. Am. J. Phys. Anthropol. 165, 77–93. doi: 10.1002/ajpa.23332
Li, D., Ng, A., Mann, N. J., and Sinclair, A. J. (1998). Contribution of meat fat to dietary arachidonic acid. Lipids 33, 437–440. doi: 10.1007/s11745-998-0225-7
Lombard, M., and Kyriacou, K. (2020). Hunter-Gatherer Women. Oxford: Oxford Research Encyclopedia of Anthropology.
Louail, M., Gilissen, E., Prat, S., Garcia, C., and Bouret, S. (2019). Refining the ecological brain: strong relation between the ventromedial prefrontal cortex and feeding ecology in five primate species. Cortex 118, 262–274. doi: 10.1016/j.cortex.2019.03.019
Luncz, L. V., Svensson, M. S., Haslam, M., Malaivijitnond, S., Proffitt, T., and Gumert, M. (2017). Technological response of wild macaques (Macaca fascicularis) to anthropogenic change. Int. J. Primatol. 38, 872–880. doi: 10.1007/s10764-017-9985-6
MacDonald, K., Scherjon, F., van Veen, E., Vaesen, K., and Roebroeks, W. (2021). Middle Pleistocene fire use: the first signal of widespread cultural diffusion in human evolution. Proc. Natl. Acad. Sci. U.S.A 118:e2101108118. doi: 10.1073/pnas.2101108118
Malaivijitnond, S., Lekprayoon, C., Tandavanittj, N., Panha, S., Cheewatham, C., and Hamada, Y. (2007). Stone-tool usage by thai long-tailed macaques (Macaca fascicularis). Am. J. Primatol. 69, 227–233. doi: 10.1002/ajp.20342
Mallick, J. K. (2019). “Primates in the Sundarbans of India and Bangladesh,” in Primates in Flooded Habitats: Ecology and Conservation, eds A. A. Barnett, I. Matsuda, and K. Nowak (Cambridge: Cambridge University Press), 110–123. doi: 10.1017/9781316466780.017
Mano, M. T., Potter, B. J., Belling, G. B., Chavadej, J., and Hetzel, B. S. (1987). Fetal brain development in response to iodine deficiency in a primate model (Callithrix jacchus jacchus). J. Neurol. Sci. 79, 287–300. doi: 10.1016/0022-510x(87)90236-x
Marean, C. W. (2014). The origins and significance of coastal resource use in Africa and Western Eurasia. J. Hum. Evol. 77, 17–40. doi: 10.1016/j.jhevol.2014.02.025
Marean, C. W. (2016). The transition to foraging for dense and predictable resources and its impact on the evolution of modern humans. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 371:239. doi: 10.1098/rstb.2015.0239
Marean, C. W., Bar-Matthews, M., Bernatchez, J., Fisher, E., Goldberg, P., Herries, A. I. R., et al. (2007). Early human use of marine resources and pigment in South Africa during the Middle Pleistocene. Nature 449, 905–908. doi: 10.1038/nature06204
Marino, L. (2007). Cetacean brains: how aquatic are they? Anat. Rec. 290, 694–700. doi: 10.1002/ar.20530
Massen, J. J. M., Szipl, G., Spreafico, M., and Bugnyar, T. (2014). Ravens intervene in others’ bonding attempts. Curr. Biol. 24, 2733–2736. doi: 10.1016/j.cub.2014.09.073
Matsuzawa, T. (1996). A newly found tool-use by wild chimpanzees: algae scooping. Primate Res 12:283.
Matsuzawa, T. (2019). Chimpanzees foraging on aquatic foods: algae scooping in Bossou. Primates 60, 317–319. doi: 10.1007/s10329-019-00733-0
Mazin, J.-M., and de Buffrénil, V. (2001). Secondary Adaptation of Tetrapods to Life in Water. in Proceedings of the International Meeting, Poitiers, 1996. Verlag Dr. Friedrich Pfeil.
McGrew, W. C. (2010). In search of the last common ancestor: new findings on wild chimpanzees. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 365, 3267–3276. doi: 10.1098/rstb.2010.0067
McLeod, H. (2018). Plenty of fish in the sea? A comparison of marine resource use in early hominins. Ethnogr 4, 36–39.
McPherron, S. P., Alemseged, Z., Marean, C. W., Wynn, J. G., Reed, D., Geraads, D., et al. (2010). Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia. Nature 466, 857–860. doi: 10.1038/nature09248
Mendes, F. D., Martins, L. B., Pereira, J. A., and Marquezan, R. F. (2000). Fishing with a bait: a note on behavioural flexibility in Cebus apella. Folia Primatol. 71, 350–352. doi: 10.1159/000021759
Mesoudi, A., and Thornton, A. (2018). What is cumulative cultural evolution? Proc. Biol. Sci. 285:712. doi: 10.1098/rspb.2018.0712
Milton, K. (1981). Distribution patterns of tropical plant foods as an evolutionary stimulus to primate mental development. Am. Anthropol. 83, 534–548. doi: 10.1525/aa.1981.83.3.02a00020
Milton, K. (1988). “Foraging behaviour and the evolution of primate intelligence,” in Machiavellian Intelligence: Social Expertise and The Evolution of Intellect in Monkeys, Apes, and Humans, eds R. W. Byrne and A. Whiten (New York, NY: Clarendon Press/Oxford University Press), 285–305.
Musgrave, S., and Sanz, C. (2019). “Animal tool use,” in Encyclopedia of Animal Behavior, 2nd Edn, Vol. 1, ed. J. C. Choe (San Diego, CA: Elsevier Academic Press), 310–317. doi: 10.1016/B978-0-12-809633-8.90154-7
Navarrete, A. F., Blezer, E. L. A., Pagnotta, M., de Viet, E. S. M., Todorov, O. S., Lindenfors, P., et al. (2018). Primate brain anatomy: new volumetric MRI measurements for neuroanatomical studies. Brain. Behav. Evol. 91, 109–117. doi: 10.1159/000488136
Navarrete, A., van Schaik, C. P., and Isler, K. (2011). Energetics and the evolution of human brain size. Nature 480, 91–93. doi: 10.1038/nature10629
Niespolo, E. M., Sharp, W. D., Avery, G., and Dawson, T. E. (2021). Early, intensive marine resource exploitation by Middle Stone Age humans at Ysterfontein 1 rockshelter, South Africa. Proc. Natl. Acad. Sci. U.S.A. 118:42118. doi: 10.1073/pnas.2020042118
Nishida, T. (1980). Local differences in responses to water among wild chimpanzees. Folia Primatol. 33, 189–209. doi: 10.1159/000155934
Nishida, T., Matsusaka, T., and McGrew, W. C. (2009). Emergence, propagation or disappearance of novel behavioral patterns in the habituated chimpanzees of Mahale: a review. Primates 50, 23–36. doi: 10.1007/s10329-008-0109-y
Nowak, K., Barnett, A. A., and Matsuda, I. (2019). Primates in Flooded Habitats: Ecology and Conservation. Cambridge: Cambridge University Press, doi: 10.1017/9781316466780
O’Connor, S., Ono, R., and Clarkson, C. (2011). Pelagic fishing at 42,000 years before the present and the maritime skills of modern humans. Science 334, 1117–1121. doi: 10.1126/science.1207703
Otani, Y., Bernard, H., Wong, A., Tangah, J., Tuuga, A., Hanya, G., et al. (2020). Factors influencing riverine utilization patterns in two sympatric macaques. Sci. Rep. 10:15749. doi: 10.1038/s41598-020-72606-2
Perry, S., Baker, M., Fedigan, L., Gros-Louis, J., Jack, K., MacKinnon, K. C., et al. (2003). Social conventions in wild white-faced capuchin monkeys: evidence for traditions in a neotropical primate. Curr. Anthropol. 44, 241–268. doi: 10.1086/345825
Phillips, D. I. W., Lusty, T. I., Osmond, C., and Church, D. (1988). Iodine supplementation: comparison of oral or intramuscular iodized oil with oral potassium iodide. A controlled trial in Zaire. Int. J. Epidemiol. 17, 142–147. doi: 10.1093/ije/17.1.142
Prat, S. (2018). First hominin settlements out of Africa. tempo and dispersal mode: review and perspectives. Comput. Rendus Palevol. 17, 6–16. doi: 10.1016/j.crpv.2016.04.009
Pusey, A. E., Pintea, L., Wilson, M. L., Kamenya, S., and Goodall, J. (2007). The contribution of long-term research at Gombe National Park to chimpanzee conservation. Conserv. Biol. 21, 623–634. doi: 10.1111/j.1523-1739.2007.00704.x
Pyke, G. H. (2019b). “Animal movements – an optimal foraging theory approach,” in Encyclopedia of Animal Behavior, 2nd Edn, Vol. 2, ed. J. C. Choe (San Diego, CA: Elsevier Academic Press), 149–156. doi: 10.1016/B978-0-12-809633-8.90160-2
Pyke, G. H. (2019a). “Optimal foraging theory: an introduction,” in Encyclopedia of Animal Behavior, 2nd Edn, Vol. 2, ed. J. C. Choe (San Diego, CA: Elsevier Academic Press), 111–117. doi: 10.1016/B978-0-12-809633-8.01156-0
Race, B., Meade-White, K. D., Miller, M. W., Barbian, K. D., Rubenstein, R., LaFauci, G., et al. (2009). Susceptibilities of nonhuman primates to chronic wasting disease. Emerg. Infect. Dis. 15, 1366–1376. doi: 10.3201/eid1509.090253
Reader, S. M., and Laland, K. N. (2001). Primate innovation: sex, age and social rank differences. Int. J. Primatol. 22, 787–805. doi: 10.1023/A:1012069500899
Rendell, L., Fogarty, L., Hoppitt, W. J. E., Morgan, T. J. H., Webster, M. M., and Laland, K. N. (2011). Cognitive culture: theoretical and empirical insights into social learning strategies. Trends Cogn. Sci. 15, 68–76. doi: 10.1016/j.tics.2010.12.002
Richards, M. P., and Trinkaus, E. (2009). Isotopic evidence for the diets of European Neanderthals and early modern humans. Proc. Natl. Acad. Sci. U.S.A. 106, 16034–16039. doi: 10.1073/pnas.0903821106
Rosati, A. G. (2017). Foraging cognition: reviving the ecological intelligence hypothesis. Trends Cogn. Sci. 21, 691–702. doi: 10.1016/j.tics.2017.05.011
Russon, A. E., Compost, A., Kuncoro, P., and Ferisa, A. (2014). Orangutan fish eating, primate aquatic fauna eating, and their implications for the origins of ancestral hominin fish eating. J. Hum. Evol. 77, 50–63. doi: 10.1016/j.jhevol.2014.06.007
Russon, A. E., Kuncoro, P., Ferisa, A., and Handayani, D. P. (2010). How orangutans (Pongo pygmaeus) innovate for water. J. Comput. Psychol. 124, 14–28. doi: 10.1037/a0017929
Sachs, R., and Voelker, J. (1975). A primate, Mandrillus leucophaeus, as natural host of the African lung fluke Paragonimus africanus in West-Cameroon. Tropenmed. Parasitol. 26, 205–206.
Sakamaki, T. (1998). First record of algae feeding by a female chimpanzee at Mahale. Pan Africa News 5, 1–3. doi: 10.5134/143366
Salmi, R., Presotto, A., Scarry, C. J., Hawman, P., and Doran-Sheehy, D. M. (2020). Spatial cognition in western gorillas (Gorilla gorilla): an analysis of distance, linearity, and speed of travel routes. Anim. Cogn. 23, 545–557. doi: 10.1007/s10071-020-01358-3
Schoener, T. W. (1971). Theory of feeding strategies. Annu. Rev. Ecol. Syst. 2, 369–404. doi: 10.1146/annurev.es.02.110171.002101
Shabel, A. B. (2010). “Brain size in carnivoran mammals that forage at the land–water ecotone, with implications for robust australopithecine paleobiology,” in Human Brain Evolution, eds S. C. Cunnane and K. M. Stewart (Wiley Online Books), 173–187. doi: 10.1002/9780470609880.ch9
Shekelle, M., Stevenson, J., Kaufer, B., Stilwell, S., and Salim, A. (2019). “Mangrove: a possible vector for tarsier dispersal across open ocean,” in Primates in Flooded Habitats: Ecology and Conservation, eds A. A. Barnett, I. Matsuda, and K. Nowak (Cambridge: Cambridge University Press), 105–109. doi: 10.1017/9781316466780.016
Shultz, S., and Dunbar, R. I. M. (2010). Species differences in executive function correlate with hippocampus volume and neocortex ratio across nonhuman primates. J. Comput. Psychol. 124, 252–260. doi: 10.1037/a0018894
Shumaker, R. W., Walkup, K. R., and Beck, B. B. (2011). Animal Tool Behavior: The Use and Manufacture of Tools by Animals, Rev. and Updated. Baltimore, MD: Johns Hopkins University Press.
Stephens, D. W., and Krebs, J. R. (1986). Foraging Theory. Princeton, NJ: Princeton University Press.
Stewart, A.-M. E., Gordon, C. H., Wich, S. A., Schroor, P., and Meijaard, E. (2008). Fishing in Macaca fascicularis: a rarely observed innovative behavior. Int. J. Primatol. 29, 543–548. doi: 10.1007/s10764-007-9176-y
Stewart, K. M. (1994). Early hominid utilisation of fish resources and implications for seasonality and behaviour. J. Hum. Evol. 27, 229–245. doi: 10.1006/jhev.1994.1044
Stewart, K. M. (2010). “The case for exploitation of wetlands environments and foods by pre-sapiens hominins,” in Human Brain Evolution, eds S. C. Cunnane and K. M. Stewart (Wiley Online Books), 137–171. doi: 10.1002/9780470609880.ch8
Stiner, M. C., Munro, N. D., Surovell, T. A., Tchernov, E., and Bar-Yosef, O. (1999). Paleolithic population growth pulses evidenced by small animal exploitation. Science 283, 190–194. doi: 10.1126/science.283.5399.190
Stringer, C. B., Finlayson, J. C., Barton, R. N. E., Fernández-Jalvo, Y., Cáceres, I., Sabin, R. C., et al. (2008). Neanderthal exploitation of marine mammals in Gibraltar. Proc. Natl. Acad. Sci. U.S.A. 105, 14319–14324. doi: 10.1073/pnas.0805474105
Tan, A. W. Y. (2017). From play to proficiency: the ontogeny of stone-tool use in coastal-foraging long-tailed macaques (Macaca fascicularis) from a comparative perception-action perspective. J. Comput. Psychol. 131, 89–114. doi: 10.1037/com0000068
Tan, A., Tan, S. H., Vyas, D., Malaivijitnond, S., and Gumert, M. D. (2015). There is more than one way to crack an oyster: identifying variation in burmese long-tailed macaque (Macaca fascicularis aurea) stone-tool use. PLoS One 10:e0124733. doi: 10.1371/journal.pone.0124733
Tennie, C., O’Malley, R. C., and Gilby, I. C. (2014). Why do chimpanzees hunt? Considering the benefits and costs of acquiring and consuming vertebrate versus invertebrate prey. J. Hum. Evol. 71, 38–45. doi: 10.1016/j.jhevol.2014.02.015
Thompson, M. E., Muller, M. N., Machanda, Z. P., Otali, E., and Wrangham, R. W. (2020). The Kibale Chimpanzee Project: over thirty years of research, conservation, and change. Biol. Conserv. 252:108857. doi: 10.1016/j.biocon.2020.108857
Tipnee, S., Ramaraj, R., and Unpaprom, Y. (2015). Nutritional evaluation of edible freshwater green macroalga Spirogyra varians. Emergent Life Sci. Res. 1, 1–7.
Tórrez-Herrera, L. L., Davis, G. H., and Crofoot, M. C. (2020). Do monkeys avoid areas of home range overlap because they are dangerous? A test of the risk hypothesis in white-faced capuchin monkeys (Cebus capucinus). Int. J. Primatol. 41, 246–264. doi: 10.1007/s10764-019-00110-0
Trapanese, C., Meunier, H., and Masi, S. (2019). What, where and when: spatial foraging decisions in primates. Biol. Rev. Camb. Philos. Soc. 94, 483–502. doi: 10.1111/brv.12462
Tsuji, Y., and Kazahari, N. (2019). “Maritime macaques: ecological background of seafood eating by wild japanese macaques (Macaca fuscata),” in Primates in Flooded Habitats: Ecology and Conservation, eds A. A. Barnett, I. Matsuda, and K. Nowak (Cambridge: Cambridge University Press), 135–143.
Vanderpas, J. B., and Moreno-Reyes, R. (2017). Historical aspects of iodine deficiency control. Minerva Med. 108, 124–135. doi: 10.23736/S0026-4806.17.04884-4
Venturi, S., and Bégin, M. E. (2010). “Thyroid hormone, iodine and human brain evolution,” in Human Brain Evolution, eds S. C. Cunnane and K. M. Stewart (Wiley Online Books), 105–124. doi: 10.1002/9780470609880.ch6
Voelker, J., and Sachs, R. (1977). Affen als naturliche und experimentelle endwirte afrikanischer lungenegel (Paragonimus africanus, P. uterobilateralis). Tropenmed. Parasitol. 28, 137–144.
Walter, R. C., Buffler, R. T., Bruggemann, J. H., Guillaume, M. M., Berhe, S. M., Negassi, B., et al. (2000). Early human occupation of the Red Sea coast of Eritrea during the last interglacial. Nature 405, 65–69. doi: 10.1038/35011048
Whitehead, H., Laland, K. N., Rendell, L., Thorogood, R., and Whiten, A. (2019). The reach of gene-culture coevolution in animals. Nat. Commun. 10:2405. doi: 10.1038/s41467-019-10293-y
Whiten, A. (2011). The scope of culture in chimpanzees, humans and ancestral apes. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 366, 997–1007. doi: 10.1098/rstb.2010.0334
Whiten, A., and van de Waal, E. (2017). Social learning, culture and the “socio-cultural brain” of human and non-human primates. Neurosci. Biobehav. Rev. 82, 58–75. doi: 10.1016/j.neubiorev.2016.12.018
Whiten, A., Goodall, J., McGrew, W. C., Nishida, T., Reynolds, V., Sugiyama, Y., et al. (1999). Cultures in chimpanzees. Nature 399, 682–685. doi: 10.1038/21415
Will, M., Kandel, A. W., and Conard, N. J. (2019). Midden or molehill: the role of coastal adaptations in human evolution and dispersal. J. World Prehistory 32, 33–72. doi: 10.1007/s10963-018-09127-4
World Health Organization (2007). Assessment of Iodine Deficiency Disorders and Monitoring Their Elimination: A Guide for Programme Managers, 3rd Edn. Geneva: World Health Organization.
Wright, T. F., Eberhard, J. R., Hobson, E. A., Avery, M. L., and Russello, M. A. (2010). Behavioral flexibility and species invasions: the adaptive flexibility hypothesis. Ethol. Ecol. Evol. 22, 393–404. doi: 10.1080/03949370.2010.505580
Xu, S., Chen, Y., Cheng, Y., Yang, D., Zhou, X., Xu, J., et al. (2012). Positive selection at the ASPM gene coincides with brain size enlargements in cetaceans. Proc. Biol. Sci. 279, 4433–4440. doi: 10.1098/rspb.2012.1729
Keywords: foraging strategies, non-human primates, decision-making, spatial ecology, cognition, brain size
Citation: de Chevalier G, Bouret S, Bardo A, Simmen B, Garcia C and Prat S (2022) Cost-Benefit Trade-Offs of Aquatic Resource Exploitation in the Context of Hominin Evolution. Front. Ecol. Evol. 10:812804. doi: 10.3389/fevo.2022.812804
Received: 10 November 2021; Accepted: 31 January 2022;
Published: 21 February 2022.
Edited by:
Mingbo Yin, Fudan University, ChinaReviewed by:
Gottfried Hohmann, Max Planck Institute for Evolutionary Anthropology, GermanyJose Joordens, Naturalis Biodiversity Center, Netherlands
Copyright © 2022 de Chevalier, Bouret, Bardo, Simmen, Garcia and Prat. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gregorio de Chevalier, Z3JlZ29yaW8uZGUtY2hldmFsaWVyLWlsbG5lckBlZHUubW5obi5mcg==
†These authors have contributed equally to this work