 Áron Péter
Áron Péter- 1Department of Parasitology and Parasitic Diseases, University of Agricultural Sciences and Veterinary Medicine Cluj-Napoca, Cluj-Napoca, Romania
- 2Research Group Mycology, Department of Biology, Ghent University, Ghent, Belgium
- 3Faculty of Science, University of South Bohemia, České Budějovice, Czechia
- 4Biology Centre of the Czech Academy of Sciences, Institute of Entomology, České Budějovice, Czechia
- 5Department of Parasitology and Zoology, University of Veterinary Medicine, Budapest, Hungary
The tritrophic association of bats, bat flies, and Laboulbeniales microfungi is a remarkably understudied system that may reveal patterns applicable to community ecology theory of (hyper)parasites. Laboulbeniales are biotrophic microfungi, exclusively associated with arthropods, with several species that are specialized on bat flies, which themselves are permanent ectoparasites of bats. Several hypotheses were tested on biotic and abiotic traits that may influence the presence and prevalence of hyperparasitic Laboulbeniales fungi on bat flies, based on southeastern European data. We found a wide distribution of fungal infection on bat flies, with underground-dwelling bats hosting more Laboulbeniales-infected flies compared to crevice-dwelling species. Bat host behavior, sociality, roost selection (underground versus crevice), bat fly sex, and season all have significant effects on the prevalence of fungal infection. Laboulbeniales infections are more common on bat flies that are infecting bat species with dense and long-lasting colonies (Miniopterus schreibersii, Myotis myotis, Myotis blythii), which roost primarily in underground sites. Inside these sites, elevated temperature and humidity may enhance the development and transmission of Laboulbeniales fungi. Sexual differences in bat hosts’ behavior also have an effect on fungal infection risk, with densely roosting female bat hosts harboring more Laboulbeniales-infected bat flies.
Background
Although often neglected in biodiversity and conservation studies, parasites are among the most diverse modes of life. They are critical components of ecosystems, altering food webs, regulating population dynamics, and driving the evolution of other species (Windsor, 1995; Bush et al., 2001; Dougherty et al., 2016; Carlson et al., 2020). A special case of parasitism—and even more understudied—is hyperparasitism, the phenomenon of parasites exploiting other parasite species (Sullivan and Völkl, 1999). Hyperparasitism has an important role in regulating host–parasite cycles of natural populations, by shaping disease dynamics and other ecological connections between species, even their evolution (Parratt and Laine, 2016; Sandhu et al., 2021). Several studies on multitrophic relationships have targeted hyperparasites in biological control strategies (Verkerk et al., 1998; Tougeron and Tena, 2019). A recently resurfaced example of hyperparasitism is the tritrophic association found among bats (Mammalia: Chiroptera), their ectoparasitic bat flies (Diptera: Hippoboscidae: Nycteribiidae and Streblidae), and fungal biotrophic parasites of the dipterans (Ascomycota: Laboulbeniomycetes) (Haelewaters et al., 2017, 2018, 2021b; de Groot et al., 2020).
Bats represent the second-largest order of mammals, showing high species diversity and worldwide distribution (Burgin et al., 2018). Bats are the only mammalian order capable of active flight and possess a number of outstanding characteristics, like high resistance to a number of deadly pathogens (Brook and Dobson, 2015), a generally nocturnal lifestyle (Simmons and Conway, 2003), and high relative longevity (Wilkinson and South, 2002). They are one of the most important components of local ecosystems due to the high number of ecosystem services provided (Williams-Guillén et al., 2016). Owing in part to their unique characteristics, bats are regularly parasitized by a number of ectoparasites (Simmons and Conway, 2003), including ticks (Sándor et al., 2019, 2021a), mites, fleas (Zahn and Rupp, 2004), cimicid bugs (Hornok et al., 2017), and bat flies (Szentiványi et al., 2016).
Bat flies are a diverse group of ectoparasites exclusively infesting bats (Dick and Patterson, 2006). They show a series of morphological and lifestyle specializations to parasitic life, including loss of wings (all Nycteribiidae and some of Streblidae species), a bullet-shaped hard chitinous body plan, and ecomorphological adaptations (differences in morphology of posterior legs) to help overcome their hosts’ grooming behavior (Theodor, 1967; Dick and Patterson, 2006; Hiller et al., 2018). They are hematophagous parasites, feeding multiple times during their life. The reproductive mode of bat flies is unique, with the female depositing a 3rd instar larva in the proximity of the hosts on the wall of the roost. This larva immediately pupates and after about 3 weeks of development an adult emerges that seeks a host bat to feed (Meier et al., 1999; Dick and Patterson, 2006). Note that, while they are able to switch among hosts, male bat flies do not leave their bat hosts at any time (Dick and Patterson, 2006). There are 17 bat fly species present in Europe (Szentiványi et al., 2016), nine of which occur both in Bulgaria and Romania (Sándor et al., 2018). Bat flies show different levels of host specificity and are being increasingly recognized as vectors for multiple pathogens like Bartonella spp., or the human-malaria related Polychromophilus spp. (Hornok et al., 2012; Sándor et al., 2018, 2021b; Szentiványi et al., 2019a).
Laboulbeniales are biotrophic microfungi, living on the outer surface of living arthropods and producing three-dimensional multicellular structures called thalli (Blackwell et al., 2020; Haelewaters et al., 2021a). A thallus typically consists of the receptacle, which is attached to the host at the foot and carries structures that produce spermatia (antheridia) and ascospores (perithecia). The foot cell is the single point of attachment to the host and is either simple or rhizoidal, penetrating the host’s integument to make contact to the hemocoel (Haelewaters et al., 2021a,b). Infected hosts spread these parasitic fungi by direct contact, e.g., during mating; ascospores are released upon pressure on the perithecium and stick to the surface of the new host (Blackwell, 1980; De Kesel, 1995). The vast majority of Laboulbeniales species, around 80% percent of the currently known 2,325 species, infect insects from the order Coleoptera, whereas only some 10% are found on representatives of the order Diptera (Weir and Hammond, 1997). Currently two described species of Arthrorhynchus are known to be associated with nycteribiid flies in Europe (Haelewaters et al., 2017), and at least one of these taxa represents a species complex (Haelewaters et al., 2020). In recent years, the scientific interest in Laboulbeniales fungi of bat flies has increased, as this system represents a model to study hyperparasitic relationships. Haelewaters et al. (2021b) called for global collaborations to build non-biased datasets resulting from multitrophic fieldwork sensu Walker et al. (2018). Such datasets may be essential in answering community ecological questions (e.g., whether habitat destruction affects parasitism at multiple levels) and in studying disease transmission through bat flies (Haelewaters et al., 2017; de Groot et al., 2020).
Here, we studied traits that may influence the presence and prevalence of Laboulbeniales fungi on nycteribiid flies in southeastern Europe (Bulgaria and Romania). Traits studied included bat roosting habits, fly intensity on a given bat, fly population dynamics, and host specificity. We analyzed a dataset of bat flies collected from different bat species and diverse bat-roost environments and over different seasons (pre- and post-breeding).
Materials and Methods
Nycteribiid flies were collected from wild-captured bats during spring and autumn from 2009 to 2020 at 43 locations in Bulgaria (5) and Romania (38). Bats were captured with mist nets (D15 mesh, 5-shelf type, 3–12 m long; Ecotone, Gdynia, Poland) and harp traps at roost entrances (caves, mines, buildings) or in areas frequented by hunting bats. Bats were identified using morphological keys (Dietz et al., 2009) and their sex, age, forearm length, and weight were recorded. Bat flies were searched on the body of the host by visually scanning the bat’s fur and all visible flies were collected using fine forceps and then stored in 98% ethanol for further analyses (one tube/bat host). Flies were identified under binocular microscope using morphological keys (Theodor, 1967). Each bat fly was screened dorsally and ventrally for the presence of visible thalli of Laboulbeniales, which were counted and recorded in the database (Supplementary Material).
Captured bats were assigned to two main seasons: all bats captured before mid-May were assigned to spring (preceding birth and nursing season), while bats captured in the period of July–October, were assigned to autumn (after weaning). No bat was captured in the period between mid-May and mid-July to avoid disturbance in the critical period for females and young (pregnancy, and the period of intensive care before weaning) and neither during hibernation. Bat species were assigned to four groups according to their affinity to a particular roost-type in the non-hibernating period: (1) caves (large/wide underground spaces), (2) mines (narrow passages and reduced area available for colonies), (3) buildings, and (4) crevices (Kunz, 1982; Kunz and Lumsden, 2003; Patterson et al., 2007); and to two groups according to their social gatherings (1) large colonies (exceeding 50 females) and (2) small colonies (few females, roosting groups <50 individuals) (Dietz et al., 2009). Mean intensity (average number of bat flies per infested bat of a given species), prevalence (percentage indicating number of infected specimens relative to total studied specimens), and their 95% confidence interval (CI) were calculated using the R language and open-access environment for statistical computing (R Core Team, 2018). Confidence intervals were calculated with the R packages PropCIs (Scherer, 2018) for prevalences and Publish (Gerds et al., 2021) for mean intensity values. For comparing parasite prevalences, we used Fisher’s Exact tests. To test the importance of certain biotic factors (bat fly sex) and abiotic factors (roost type, season) on the presence versus absence of Laboulbeniales on bat flies, we used general linear models (GLM) under the assumption of a binomial distribution (absence/presence), with the R package stats (R Core Team, 2018). Intensity of infection values were compared with non-parametrical Wilcoxon Rank Sum tests. To assess the correlation between the intensity of infestation and Laboulbeniales infection of bat flies we used Spearman’s Rank Correlation. The bipartite network was constructed using the R package bipartite (Dormann et al., 2008). Differences were considered significant when p < 0.05.
Results
Our dataset of examined bat flies consisted of 4,122 specimens collected from 1,689 bat individuals. Fly specimens represented ten species: Basilia italica (n = 3), B. nana (n = 77), Nycteribia kolenatii (n = 238), N. latreillii (n = 182), N. pedicularia (n = 552), N. schmidlii (n = 1,144), N. vexata (n = 228), Penicillidia conspicua (n = 602), Pe. dufourii (n = 587), and Phthiridium biarticulatum (n = 509). We found 263 Laboulbeniales-infected bat flies out of the 4,122 examined specimens (prevalence 6.13%, CI 5.44–6.91; Table 1 and Supplementary Material). High prevalence was recorded for Pe. conspicua with 25.91% (CI 22.57–29.56), whereas lower prevalence was recorded for Pe. dufourii (8.85%, CI 6.80–11.44), N. vexata (5.70%, CI 3.27–9.59), Ph. biarticulatum (2.94%, CI 1.75–4.84), and N. schmidlii (2.01%, CI 1.32–3.01). The presence of Laboulbeniales was rare (prevalence <1%) for N. kolenatii, N. latreillii, and N. pedicularia, and no Laboulbeniales infection was found on two fly species (B. italica and B. nana). The majority (229 of 263, 87%) of infected flies were collected from three bat host species (Figure 1). Miniopterus schreibersii hosted the highest number of total bat flies (n = 1,829). This bat species also hosted the highest number of Laboulbeniales-infected bat flies (n = 186), with a prevalence of 10.16% (CI 8.86–11.64), followed by Myotis blythii (27 infected flies, 7.16%, CI 4.93–10.25) and My. myotis (16 infected flies, 4.08%, CI 2.47–6.57).
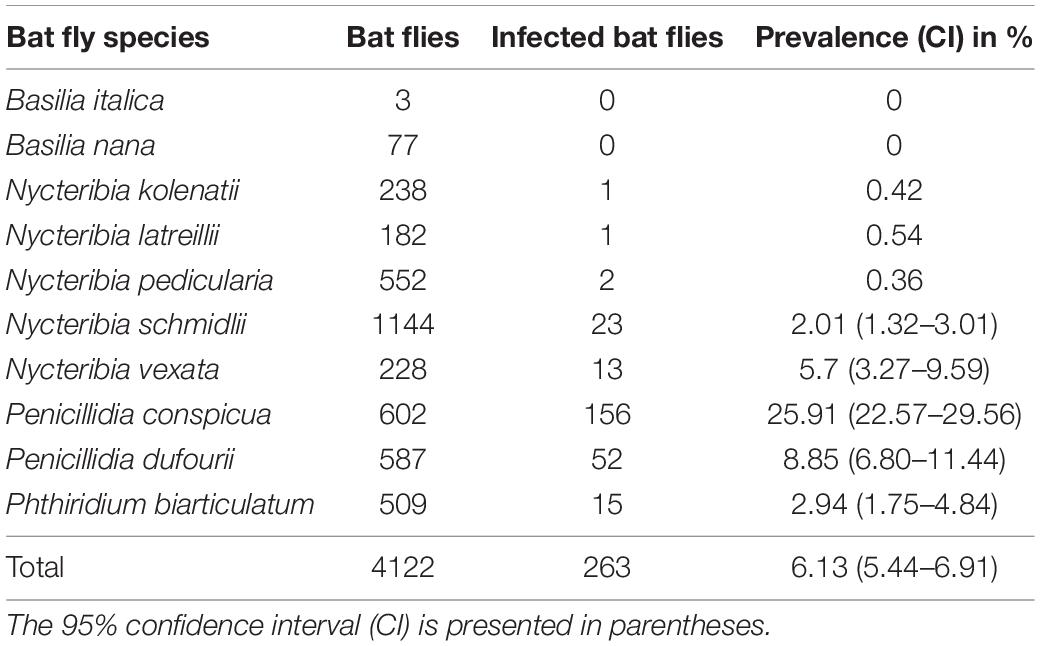
Table 1. Prevalence values of Laboulbeniales-infected bat fly species collected in this study.
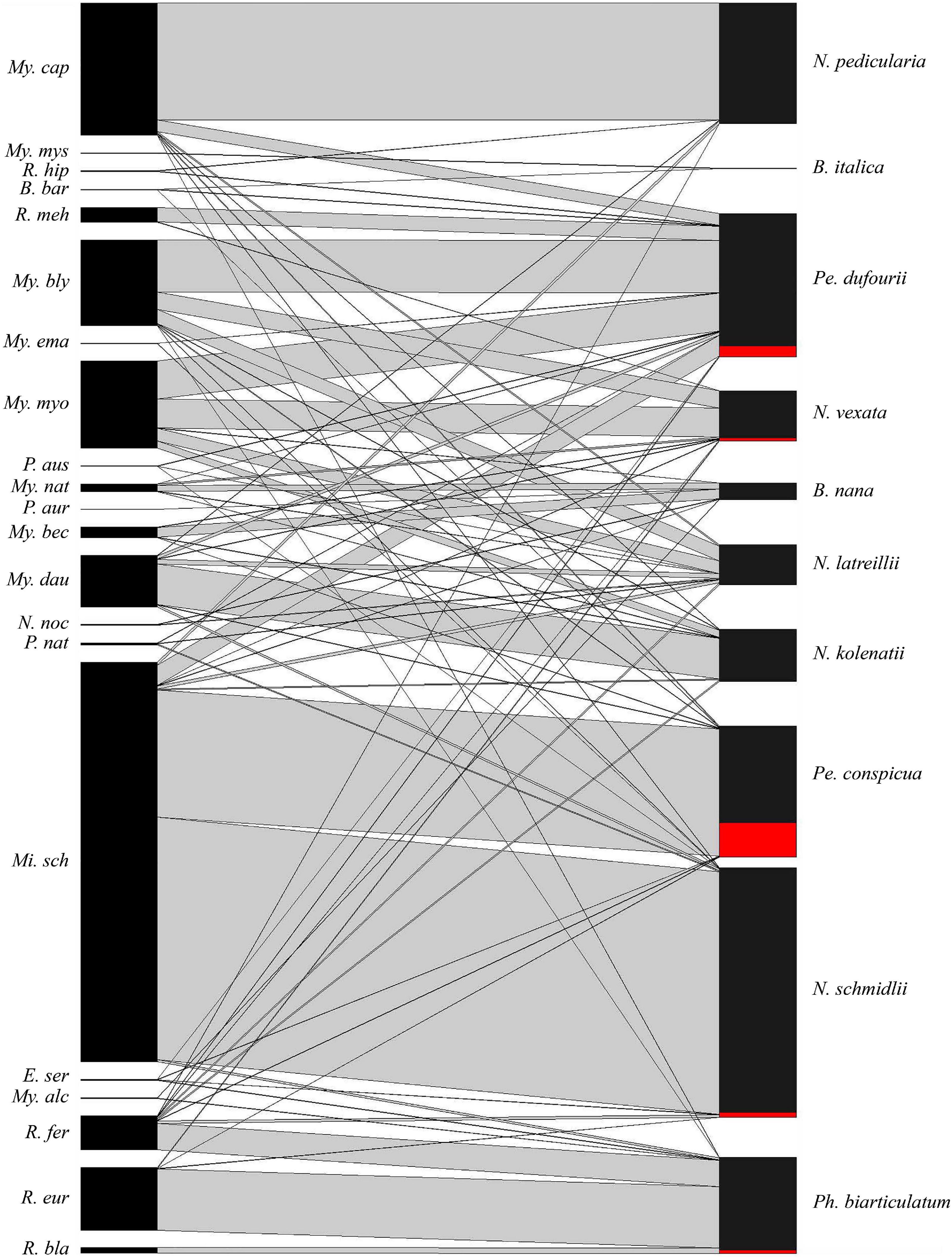
Figure 1. Bat–bat fly network. Shown is the association of bat hosts (left) with their bat fly parasites (right). Bar width represents the relative abundance of a species within each network level. The parasite prevalences of bat fly species with Laboulbeniales are shown in red, for Penicillidia dufourii (8.85%), Nycteribia vexata (5.70%), Penicillidia conspicua (25.91%), Nycteribia schmidlii (2.01%), and Phthiridium biarticulatum (2.94%).
The majority of bats were infested with multiple bat flies, the highest number of flies being 18 on a single bat individual of Myotis capaccinii (17 N. pedicularia and 1 Pe. dufourii). We found no correlation between the intensity of flies on bat hosts and the prevalence of fungal infection on flies. Also, there was no correlation between the intensity of flies and the prevalence of bat hosts with Laboulbeniales-infected bat flies present. However, when analyzing only the subset data of Pe. conspicua and Pe. dufourii (the number of flies on a given bat individual was 1–5 for these two species), we found a significant positive correlation between bat fly intensity and the prevalence of bat hosts with Laboulbeniales-infected flies (rho = 1, p = 0.016; Figure 2).
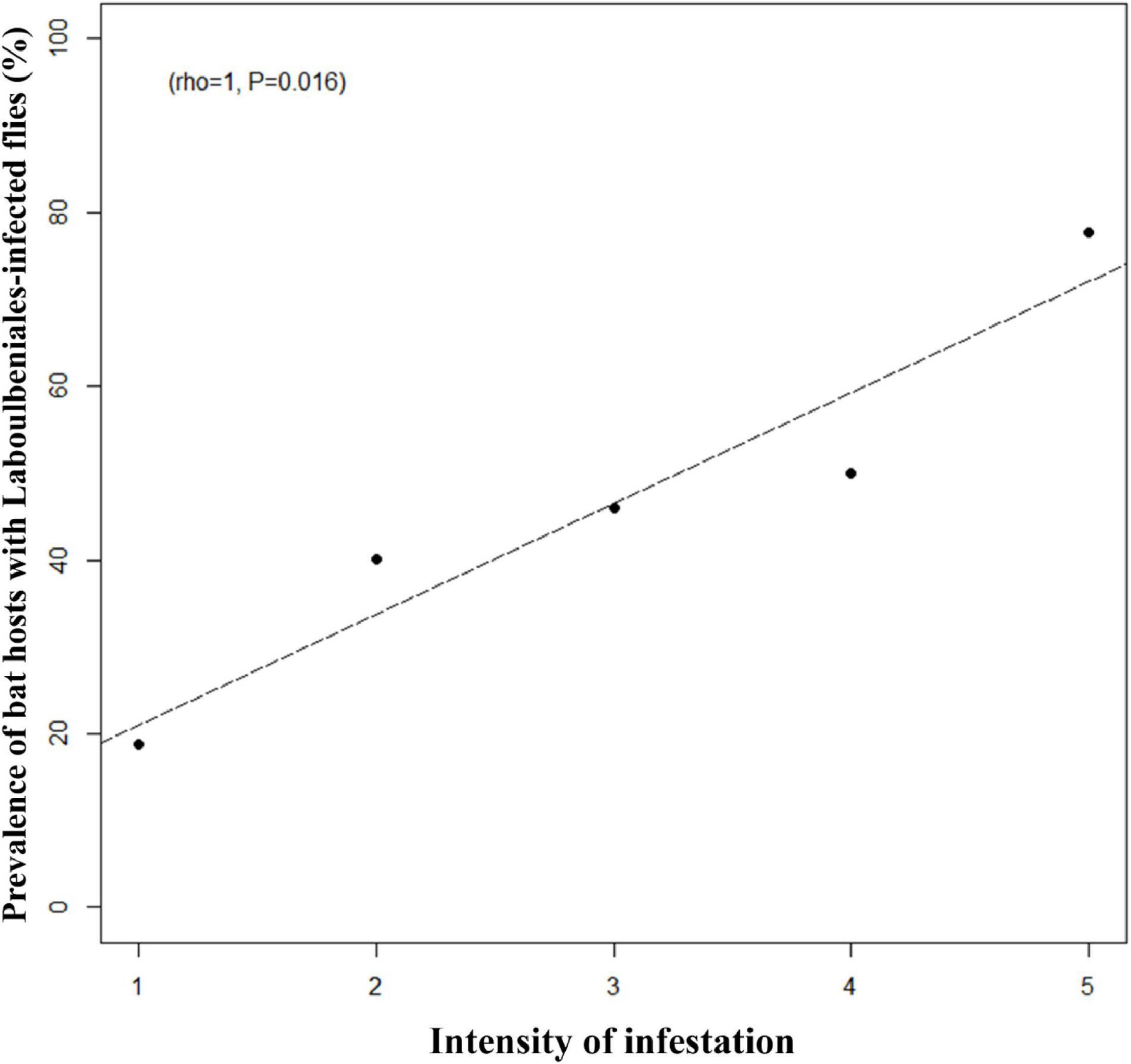
Figure 2. Correlation between bat fly intensity on bats hosts and the prevalence of bat hosts with Laboulbeniales-infected bat flies. Based on subset data for Penicillidia conspicua and Pe. dufourii.
Parasitism of bats with bat flies was significantly higher in autumn both in prevalence (x2 = 25.61, d.f. = 1, p < 0.001; spring 33.27%, CI 31.13–35.49; autumn 40.72%, CI 38.93–42.53) and in mean intensity (W = 310142, p < 0.001; spring 2.36%, CI 2.15–2.57; autumn 2.51%, CI 2.93–2.64). There was a significant difference (x2 = 13.05, p < 0.001) of parasite prevalence of bat flies with Laboulbeniales between the two seasons (spring 4.2%, CI 3.20–5.49; autumn 7.28%, CI 6.39–8.28; Table 2).
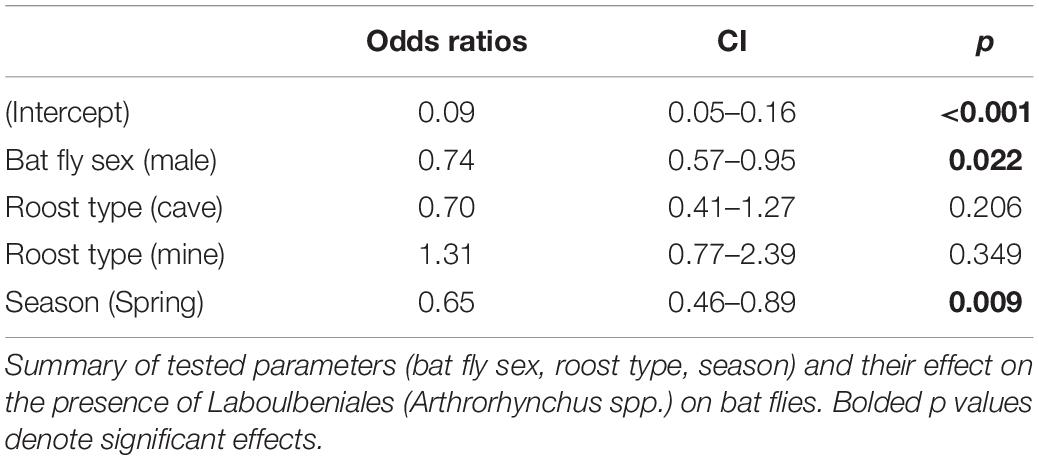
Table 2. Results of our modeling approach (GLM).
Bat fly sex was an important factor determining whether infection with Laboulbeniales was present (Table 2). Overall, we found significant differences (x2 = 4.16, p = 0.04) in parasite prevalence between female and male flies, with females being more infected by Laboulbeniales (females 7.1%, CI 6.11–8.23; males 5.48%, CI 4.53–6.62). When separately analyzing by bat fly species, no significant sex differences were found for Pe. conspicua (females 27.48%, CI 23.01–32.45; males 23.84%, CI 19.05–29.39). A significant difference in parasite prevalence of bat flies by Laboulbeniales was observed between female and male bat hosts: 132 infected flies out of 2,360 ones collected from female bats (5.59%, CI 4.73–6.59) versus 131 infected flies out of 1,762 ones collected from male bats (7.43%, CI 6.29–8.75; x2 = 5.42, p = 0.01). However, the difference was only marginally significant (p = 0.06) when separately analyzing flies collected from Mi. schreibersii, which is the only bat species in our dataset of which males and females roost together in mixed colonies.
Finally, we found significant differences in the prevalence of Laboulbeniales infections between roost types. Bats roosting in mines had bat flies with higher parasite prevalence (9.28%, CI 7.86–10.92) compared to those roosting in caves (4.77%, CI: 3.99–5.68; x2 = 29.61, p < 0.001). Roost type, however, did not significantly contribute to the general GLM (Table 2). The prevalence of Laboulbeniales on flies collected from bats in buildings was 6.43% (CI 3.86–10.42), however, this number was not significantly different from those of flies collected from bats in caves or mines. Crevice-dwelling bats had no flies infected with Laboulbeniales.
Discussion
Most of the bat fly species collected in our study (8 out of 10) showed signs of Laboulbeniales infections (Table 1), although the parasite prevalence on three bat fly species is very low (<1%) and thus the infection may be considered rare. All infected bat fly species were already known to host Laboulbeniales fungi (Haelewaters et al., 2017; de Groot et al., 2020). The bat flies in our study were collected from a wide range of bat species in Bulgaria (nine bat species) and Romania (27 bat species). The overall prevalence of Laboulbeniales fungi on bat flies was generally low, similar to previous studies. Previously reported mean parasite prevalences of Laboulbeniales on nycteribiid flies ranged between 2.2 and 4.6% (Blackwell, 1980; Haelewaters et al., 2017, 2018; Jensen et al., 2019), with one study reporting an overall prevalence of 9.0% on bat flies collected from only Mi. schreibersii (Szentiványi et al., 2018). We found a relatively high prevalence, 6.13%, which is probably due to the higher number of Pe. conspicua bat flies in our samples. This bat fly species was previously identified as the main host for the fungus Arthrorhynchus nycteribiae (Haelewaters et al., 2017), and this is confirmed here (Table 1 and Figure 1).
Most bat fly species are highly host specific, and this is also the case for Pe. conspicua; this species was found parasitizing Mi. schreibersii with a few exceptions (Figure 1). Other fly species parasitizing Mi. schreibersii were Pe. dufourii and N. schmidlii. Penicillidia dufourii and N. schmidlii collected from Mi. schreibersii both showed a lower fungal parasite prevalence (8.85 and 2.01%, respectively), compared to Pe. conspicua from the same bat host. Our results are very similar to those found by Haelewaters et al. (2017): 33 of 142 (23.1%) Pe. conspicua collected from Mi. schreibersii infected by A. nycteribiae, 2 of 51 (3.9%) Pe. dufourii from My. myotis infected by A. nycteribiae, and 1 of 147 (0.68%) N. schmidlii from Mi. schreibersii infected by A. nycteribiae. Penicillidia dufourii mostly parasitizes My. myotis and My. blythii but occasionally also Mi. schreibersii because these three bat species frequently share roosts (Dietz et al., 2009). Penicillidia dufourii as well as N. schmidlii are probably accidental host species for A. nycteribiae, as a result of inter-specific encounters and interactions with Pe. conspicua on Mi. schreibersii bats. Nycteribia schmidlii is also host to another species of Laboulbeniales, Arthrorhynchus eucampsipodae. In the study by Haelewaters et al. (2017), parasite prevalence of N. schmidlii by A. eucampsipodae was 2.72% (4 of 147 bat flies infected). Arthrorhynchus eucampsipodae is much rarer than A. nycteribiae but has thus far been reported from twelve bat fly species in five genera (de Groot et al., 2020). However, these data are misleading because recent work has shown that this taxon consists of multiple species that are host specific (Haelewaters et al., 2020).
High prevalence values of bat flies by Laboulbeniales were observed on specimens collected from bats that often roost together in close proximity and in large colonies, such as Mi. schreibersii, My. myotis, and My. blythii. As mentioned above, these three bat species frequently use mixed colonies (Dietz et al., 2009), thus creating opportunities for continuous transmissions of ascospores from each others’ flies. The roosting behavior of these species was already suggested to be the main factor for the comparatively high success of parasitism of these bats by multiple parasite groups (Zahn and Rupp, 2004; Frank et al., 2015; Sándor et al., 2019).
Increased intensity of flies (= more bat fly specimens on a single host) implies increased levels of fly–fly interactions, thus increasing the opportunities for ascospore transmission. Indeed, we found a significant positive correlation between the number of individual bat flies on a single bat and the Laboulbeniales prevalence on these flies, for both Pe. conspicua and Pe. dufourii, the two most commonly found Laboulbeniales-infected bat fly species. This is likely caused by the high rate of fly–fly contacts on their bat hosts, due to increased competition among flies when bat fly intensity is high. Another factor that may contribute is a higher bat host response, as bats are known to react toward increased fly density by higher levels of scratching and grooming (Ter Hofstede and Fenton, 2005).
We found seasonal differences in the prevalence of Laboulbeniales infections on bat flies. In autumn, parasite prevalence on bat hosts was significantly higher than in spring. Bat flies are much more abundant in autumn (Lourenço and Palmeirim, 2008), again leading to increased opportunities for ascospore transmission among flies. Seasonal patterns of infection were also studied in Rickia wasmannii, a species of Laboulbeniales associated with Myrmica ants. It was found that prevalence was highest after winter and this was also in part explained by increased contacts (allogrooming) among ants that aggregate in dense clusters during wintertime (Haelewaters et al., 2015).
Bat fly sex had an impact on Laboulbeniales infections. Female bat flies were significantly more infected by Laboulbeniales than males. Similar results were already previously reported in Europe (Haelewaters et al., 2017; Szentiványi et al., 2018). The fact that Laboulbeniales appear to have a preference for female bat flies may be attributed to their longer lifespan resulting in build-up of inoculum, their larger size compared to males, and the accumulation of fat reserves during pregnancy (Haelewaters et al., 2017).
Bat flies collected from bats roosting in caves were found to be significantly more infected by Laboulbeniales fungi compared to flies from bats using either mines or buildings. This suggests that Laboulbeniales infection may be linked to the microclimatic factors present in these roosts (for example, elevated humidity levels in caves might enhance ascospore survival and development, as well as attachment to the integument of flies). Szentiványi et al. (2019b) found that the prevalence of bat fly-associated Laboulbeniales was negatively associated with annual mean temperature and humidity. However, their results were based on outside bioclimatic variables. We suggest to continuously measure temperature, humidity, and precipitation while surveying roosts for bats and their associated parasites and hyperparasites—to gain a better understanding of microhabitat conditions governing Laboulbeniales infections of flies collected from cave-dwelling bats.
Most bat species show sexual segregation in roost use, sociality, and mobility in their active periods, with females roosting in dense groups and males (even if present in the same location) roosting in smaller groups or as scattered individuals away from the females (Dietz et al., 2009). Moreover, female groups tend to be highly philopatric, whereas males show nomadic tendencies (Dietz et al., 2009; Jang et al., 2021). As a result, we hypothesized that these behavioral differences between sexes affect infection of their bat flies by Laboulbeniales. We found significant differences in the Laboulbeniales prevalence of bat flies collected from the two sexes, with flies originating from male bats having higher fungal prevalence compared to those from female bats. When Mi. schreibersii bats were analyzed separately, male bats still carried higher ratio of infected flies but the difference was marginally significant. Patterson et al. (2008) studied sex biases in parasitism of Neotropical bats and found that bat fly intensity and the number of bat fly species were higher for female bats. This was explained by higher survivorship rates and their larger size (= more “habitat”). The bat species and fly species studied by us are different, but no comparable data exist for European bats to our knowledge. Based on the findings by Patterson et al. (2008), we assumed that female bats are also parasitized by higher numbers of bat flies as well as more fly species. If that is indeed the case, bat flies on female bat hosts have more opportunities for encounters, which increases the likelihood of ascospore transmission among flies. Miniopterus schreibersii is the only bat species in our study with mixed male–female colonies (Rodrigues and Palmeirim, 2008; Ramos Pereira et al., 2009), and as a result the aforementioned overall female bias may be balanced out due to the mixture with males, allowing flies to move from one host (either male or female) to the other (either male or female).
Conclusion
We found that Laboulbeniales infections are more common on bat flies that are infesting bat species with dense and long-lasting colonies (Mi. schreibersii, My. myotis, My. blythii), roosting primarily in caves. Inside these cave ecosystems, abiotic traits (e.g., elevated humidity) may enhance the survival, development, and attachment of fungal ascospores. Finally, we observed sex-biases in parasitism of nycteribiid flies by Laboulbeniales with higher prevalence values of female flies compared to males and of flies collected from female bats. Only recently, this multitrophic and hyperparasitic system of bats, bat flies, and fungi started receiving more attention from an ecological perspective. The currently available data are too fragmented to fully understand interactions among the organisms in this system and biotic and abiotic traits affecting them (e.g., Haelewaters et al., 2018; Szentiványi et al., 2019b; de Groot et al., 2020, this article). There is a need to build large, non-biased, and standardized datasets for rigorous testing of hypotheses—several initiatives are being taken in this regard.
Data Availability Statement
The original data presented in this study are included in the Supplementary Material.
Ethics Statement
The animal study was reviewed and approved by the Research Bioethics Commission of USAMV CN.
Author Contributions
ÁP and AS conceived the study. ÁP, AS, and ADM collected specimens. ÁP, AS, and DH wrote the first draft of the manuscript. ÁP and DH prepared the figures. All authors contributed to revisions and approved the final version of the manuscript.
Funding
This research was supported in part by the National Research, Development and Innovation Office of Hungary (NKFIH-132794 to AS), the Collegium Talentum Programme of Hungary, and NTP-NFTÖ-20 (both to ÁP). DH was supported in part by the Research Foundation – Flanders (Junior Postdoctoral Fellowship No. 1206620N), and the Czech Science Foundation (Grant 21-06446S).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We wish to express our gratitude to the management authorities of Parcul Natural (PN) Portile de Fier, Parcul Naţional Domogled-Valea Cernei, Parcul Natural Apuseni, the Constanta Forestry Commission (Romsilva Constanta, Ocolul Silvic Baneasa), and the Bulgarian Ministry of Environment and Water for granting research permits.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.795020/full#supplementary-material
References
Blackwell, M. (1980). Developmental morphology and taxonomic characters of Arthrorhynchus nycteribiae and A. eucampsipodae (Laboulbeniomycetes). Mycologia 72, 159–168. doi: 10.1080/00275514.1980.12021164
Blackwell, M., Haelewaters, D., and Pfister, D. H. (2020). Laboulbeniomycetes: evolution, natural history, and Thaxter’s final word. Mycologia 112, 1048–1059. doi: 10.1080/00275514.2020.1718442
Brook, C. E., and Dobson, A. P. (2015). Bats as “special” reservoirs for emerging zoonotic pathogens. Trends Microbiol. 23, 172–180. doi: 10.1016/j.tim.2014.12.004
Burgin, C. J., Colella, J. P., Kahn, P. L., and Upham, N. S. (2018). How many species of mammals are there? J. Mammal. 99, 1–14. doi: 10.1093/jmammal/gyx147
Bush, A. O., Fernandez, J. C., Esch, G. W., and Seed, J. R. (2001). Parasitism: The Diversity and Ecology of Animal Parasites. Cambridge: Cambridge University Press.
Carlson, C. J., Hopkins, S., Bell, K. C., Doña, J., Godfrey, S. S., Kwak, M. L., et al. (2020). A global parasite conservation plan. Biol. Conserv. 250:108596. doi: 10.1016/j.biocon.2020.108596
de Groot, M. D., Dumolein, I., Hiller, T., Sándor, A. D., Szentiványi, T., Schilthuizen, M., et al. (2020). On the fly: tritrophic associations of bats, bat flies, and fungi. J. Fungi. 6:361. doi: 10.3390/jof6040361
De Kesel, A. (1995). Relative importance of direct and indirect infection in the transmission of Laboulbenia slackensis (Ascomycetes. Laboulbeniales). Belg. J. Bot. 128, 124–130.
Dick, C. W., and Patterson, B. D. (2006). “Bat flies: obligate ectoparasites of bats”, in Micromammals and Macroparasites: From Evolutionary Ecology to Management, eds R. P. S. Morand and B. R. Krasnov Poulin (Tokyo, Japan: Springer-Verlag), 179–194. doi: 10.1007/978-4-431-36025-4_11
Dietz, C., von Helversen, O., and Nill, D. (2009). Bats of Britain, Europe and northwest Africa. London: A&C Black.
Dormann, C. F., Gruber, B., and Fründ, J. (2008). Introducing the bipartite package: analysing ecological networks. R News 8, 8–11.
Dougherty, E. R., Carlson, C. J., Bueno, V. M., Burgio, K. R., Cizauskas, C. A., Clements, C. F., et al. (2016). Paradigms for parasite conservation. Conserv. Biol. 30, 724–733. doi: 10.1111/cobi.12634
Frank, R., Kuhn, T., Werblow, A., Liston, A., Kochmann, J., and Klimpel, S. (2015). Parasite diversity of European Myotis species with special emphasis on Myotis myotis (Microchiroptera, Vespertilionidae) from a typical nursery roost. Parasit. Vectors 8:101. doi: 10.1186/s13071-015-0707-7
Gerds, T. A., Torp-Pedersen, C., Holst, K. K., and Ozenne, B. (2021). Publish: Format Output of Various Routines in a Suitable Way for Reports and Publication. See. Available online at: https://cran.r-project.org/package=Publish (accessed on 10 Sep, 2021).
Haelewaters, D., Blackwell, M., and Pfister, D. H. (2021a). Laboulbeniomycetes: intimate fungal associates of arthropods. Ann. Rev. Entomol. 66, 257–276. doi: 10.1146/annurev-ento-013020-013553
Haelewaters, D., Boer, P., Gort, G., and Noordijk, J. (2015). Studies of Laboulbeniales (Fungi, Ascomycota) on Myrmica ants (II): variation of infection by Rickia wasmannii over habitats and time. Anim. Biol. 65, 219–231. doi: 10.1163/15707563-00002472
Haelewaters, D., Dick, C. W., Cocherán Pittí, K. P., Dittmar, K., and Patterson, B. D. (2021b). “Bats, bat flies, and fungi: exploring uncharted waters,” in 50 Years of Bat Research, eds B. K. Lim, M. B. Fenton, Brigham S. Mistry, A. Kurta, E. H. Gillam, et al. (Cham: Springer), 349–371. doi: 10.1007/978-3-030-54727-1-21
Haelewaters, D., Dima, B., Abdel-Hafiz, B. I. I., Abdel-Wahab, M. A., Abul-Ezz, S. R., Acar, I., et al. (2020). Fungal systematics and evolution: FUSE 6. Sydowia 72, 231–356. doi: 10.12905/0380.sydowia72-2020-0231
Haelewaters, D., Page, R. A., and Pfister, D. H. (2018). Laboulbeniales hyperparasites (Fungi, Ascomycota) of bat flies: Independent origins and host associations. Ecol. Evol. 8, 8396–8418. doi: 10.1002/ece3.4359
Haelewaters, D., Pfliegler, W. P., Szentiványi, T., Földvári, M., Sándor, A. D., Barti, L., et al. (2017). Parasites of parasites of bats: Laboulbeniales (Fungi: Ascomycota) on bat flies (Diptera: Nycteribiidae) in central Europe. Parasit. Vectors 10:395. doi: 10.1186/s13071-017-2022-y
Hiller, T., Honner, B., Page, R. A., and Tschapka, M. (2018). Leg structure explains host site preference in bat flies (Diptera: Streblidae) parasitizing neotropical bats (Chiroptera: Phyllostomidae). Parasitology 145, 1475–1482. doi: 10.1017/S0031182018000318
Hornok, S., Kovács, R., Meli, M. L., Gönczi, E., Hofmann-Lehmann, R., Kontschán, J., et al. (2012). First detection of bartonellae in a broad range of bat ectoparasites. Vet. Microbiol. 159, 541–543. doi: 10.1016/j.vetmic.2012.04.003
Hornok, S., Szõke, K., Boldogh, S. A., Sándor, A. D., Kontschán, J., Tu, V. T., et al. (2017). Phylogenetic analyses of bat-associated bugs (Hemiptera: Cimicidae: Cimicinae and Cacodminae) indicate two new species close to Cimex lectularius. Parasit. Vectors 10:439. doi: 10.1186/s13071-017-2376-1
Jang, J. E., Byeon, S. Y., Kim, H. R., Kim, J. Y., Myeong, H. H., and Lee, H. J. (2021). Genetic evidence for sex-biased dispersal and cryptic diversity in the greater horseshoe bat. Biodivers. Conserv. 30, 847–864. doi: 10.1007/s10531-021-02120-y
Jensen, K. M., Rodrigues, L., Pape, T., Garm, A., Santamaria, S., and Reboleira, A. S. P. S. (2019). Hyperparasitism in caves: bats, bat flies and ectoparasitic fungus interaction. J. Invertebr. Pathol. 166:107206. doi: 10.1016/j.jip.2019.107206
Kunz, T. H. (1982). “Roosting ecology,” in Ecology of Bats, ed. T. H. Kunz (New York, NY: Plenum Press), 1–55.
Kunz, T. H., and Lumsden, L. F. (2003). “Ecology of cavity and foliage roosting bats,” in Bat Ecology, eds T. H. Kunz and M. B. Fenton (Chicago: University of Chicago Press), 3–89. doi: 10.1002/ece3.8101
Lourenço, S., and Palmeirim, J. M. (2008). Which factors regulate the reproduction of ectoparasites of temperate-zone cave-dwelling bats? Parasitol. Res. 104, 127–134. doi: 10.1007/s00436-008-1170-6
Meier, R., Kotrba, M., and Ferrar, P. (1999). Ovoviviparity and viviparity in the Diptera. Biol. Rev. 74, 199–258. doi: 10.1017/S0006323199005320
Parratt, S. R., and Laine, A.-L. (2016). The role of hyperparasitism in microbial pathogen ecology and evolution. ISME J. 10, 1815–1822. doi: 10.1038/ismej.2015.247
Patterson, B. D., Dick, C. W., and Dittmar, K. (2007). Roosting habits of bats affect their parasitism by bat flies (Diptera: Streblidae). J. Trop. Ecol. 23, 177–189. doi: 10.1017/S0266467406003816
Patterson, B. D., Dick, C. W., and Dittmar, K. (2008). Sex biases in parasitism of neotropical bats by bat flies (Diptera: Streblidae). J. Trop. Ecol. 24, 387–396. doi: 10.1017/S0266467408005117
Ramos Pereira, M. J., Salgueiro, P., Rodrigues, L., Coelho, M. M., and Palmeirim, J. M. (2009). Population structure of a cave-dwelling bat. J. Hered. 100, 533–544. doi: 10.1093/jhered/esp032
R Core Team (2018). R: A Language and Environment for Statistical Computing. Vienna: Austria: R Foundation.
Rodrigues, L., and Palmeirim, J. M. (2008). Migratory behaviour of the Schreiber’s bat: when, where and why do cave bats migrate in a Mediterranean region? J. Zool. 274, 116–125. doi: 10.1111/j.1469-7998.2007.00361.x
Sandhu, S. K., Morozov, A. Y., Holt, R. D., and Barfield, M. (2021). Revisiting the role of hyperparasitism in the evolution of virulence. Am. Nat. 197, 216–235. doi: 10.1086/712351
Sándor, A. D., Corduneanu, A., Péter, Á, Mihalca, A. D., Barti, L., Csõsz, I., et al. (2019). Bats and ticks: host selection and seasonality of bat-specialist ticks in eastern Europe. Parasit. Vectors 12:605. doi: 10.1186/s13071-019-3861-5
Sándor, A. D., Földvári, M., Krawczyk, A. I., Sprong, H., Corduneanu, A., Barti, L., et al. (2018). Eco-epidemiology of novel Bartonella genotypes from parasitic flies of insectivorous bats. Microb. Ecol. 76, 1076–1088. doi: 10.1007/s00248-018-1195-z
Sándor, A. D., Mihalca, A. D., Domşa, C., Péter, Á, and Hornok, S. (2021a). Argasid ticks of palearctic bats: distribution, host selection, and zoonotic importance. Front. Vet. Sci. 8:684737. doi: 10.3389/fvets.2021.684737
Sándor, A. D., Péter, Á, Corduneanu, A., Barti, L., Csõsz, I., Kalmár, Z., et al. (2021b). Wide distribution and diversity of malaria-related haemosporidian parasites (Polychromophilus spp.) in bats and their ectoparasites in eastern europe. Microorganisms 9:230. doi: 10.3390/microorganisms9020230
Scherer, R. (2018). Package ‘PropCIs’. Available online at: http://ftp5.gwdg.de/pub/misc/cran/web/packages/PropCIs/PropCIs.pdf (accessed on 10 Sep, 2021)
Simmons, N. B., and Conway, T. M. (2003). “Evolution of ecological diversity in bats,” in Bat Ecology, eds T. Kunz and M. B. Fenton (Chicago, IL: University of Chicago Press), 493–535.
Sullivan, D. J., and Völkl, W. (1999). Hyperparasitism: multitrophic ecology and behavior. Ann. Rev. Entomol. 44, 291–315. doi: 10.1146/annurev.ento.44.1.291
Szentiványi, T., Estók, P., and Földvári, M. (2016). Checklist of host associations of European bat flies (Diptera: Nycteribiidae, Streblidae). Zootaxa 4205, 101–126. doi: 10.11646/zootaxa.4205.2.1
Szentiványi, T., Haelewaters, D., Pfliegler, W. P., Clément, L., Christe, P., and Glaizot, O. (2018). Laboulbeniales (Fungi: Ascomycota) infection of bat flies (Diptera: Nycteribiidae) from Miniopterus schreibersii across Europe. Parasit. Vectors 11:395. doi: 10.1186/s13071-018-2921-6
Szentiványi, T., Christe, P., and Glaizot, O. (2019a). Bat flies and their microparasites: current knowledge and distribution. Front. Vet. Sci. 6:115. doi: 10.3389/fvets.2019.00115
Szentiványi, T., Haelewaters, D., Rádai, Z., Mizsei, E., Pfliegler, W. P., Báthori, F., et al. (2019b). Climatic effects on the distribution of ant- and bat fly-associated fungal ectoparasites (Ascomycota, Laboulbeniales). Fungal Biol. 39, 371–379. doi: 10.1016/j.funeco.2019.03.003
Ter Hofstede, H. M., and Fenton, M. B. (2005). Relationships between roost preferences, ectoparasite density, and grooming behaviour of neotropical bats. J. Zool. 266, 333–340. doi: 10.1017/S095283690500693X
Theodor, O. (1967). An Illustrated Catalogue of the Rothschild Collection of Nycteribiidae in the British Museum (Natural History), with Keys and Short Descriptions for the Identification of Subfamilies, Genera, Species and Subspecies. London: British Museum (Natural History).
Tougeron, K., and Tena, A. (2019). Hyperparasitoids as new targets in biological control in a global change context. Biol. Control 130, 164–171. doi: 10.1016/j.biocontrol.2018.09.003
Verkerk, R. H. J., Leather, S. R., and Wright, D. J. (1998). The potential for manipulating crop-pest-natural enemy interactions for improved insect pest management. Bull. Entomol. Res. 88, 493–501. doi: 10.1017/S0007485300026018
Walker, M. J., Dorrestein, A., Camacho, J. J., Meckler, L. A., Silas, K. A., Hiller, T., et al. (2018). A tripartite survey of hyperparasitic fungi associated with ectoparasitic flies on bats (Mammalia: Chiroptera) in a neotropical cloud forest in Panama. Parasite 25:19. doi: 10.1051/parasite/2018017
Weir, A., and Hammond, P. M. (1997). Laboulbeniales on beetles: host utilization patterns and species richness of the parasites. Biodivers. Conserv. 6, 701–719. doi: 10.1023/A:1018318320019
Wilkinson, G. S., and South, J. M. (2002). Life history, ecology and longevity in bats. Aging Cell 1, 124–131. doi: 10.1046/j.1474-9728.2002.00020.x
Williams-Guillén, K., Olimpi, E., Maas, B., Taylor, P. J., and Arlettaz, R. (2016). “Bats in the anthropogenic matrix: challenges and opportunities for the conservation of Chiroptera and their ecosystem services in agricultural landscapes,” in Bats in the Anthropocene: Conservation of Bats in a Changing World, eds C. C. Voigt and T. Kingston (Cham, Switzerland: Springer), 151–186. doi: 10.1007/978-3-319-25220-9_6
Windsor, D. A. (1995). Equal rights for parasites. Conserv. Biol. 9, 1–2. doi: 10.1046/j.1523-1739.1995.09010001.x
Keywords: biotrophic fungi, Chiroptera, host-parasite interactions, Nycteribiidae, sex-biased infection, symbiosis
Citation: Péter Á, Mihalca AD, Haelewaters D and Sándor AD (2022) Focus on Hyperparasites: Biotic and Abiotic Traits Affecting the Prevalence of Parasitic Microfungi on Bat Ectoparasites. Front. Ecol. Evol. 10:795020. doi: 10.3389/fevo.2022.795020
Received: 14 October 2021; Accepted: 17 January 2022;
Published: 17 February 2022.
Edited by:
Pavel Kindlmann, Charles University, CzechiaReviewed by:
Lien Luong, University of Alberta, CanadaGustavo Graciolli, Federal University of Mato Grosso do Sul, Brazil
Copyright © 2022 Péter, Mihalca, Haelewaters and Sándor. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Danny Haelewaters, danny.haelewaters@gmail.com