 Paulo Cesar Machado Andrade1*
Paulo Cesar Machado Andrade1* Paulo Henrique Guimarães de Oliveira2
Paulo Henrique Guimarães de Oliveira2 Aldeniza Cardoso de Lima3
Aldeniza Cardoso de Lima3 João Alfredo da Mota Duarte4
João Alfredo da Mota Duarte4 Sandra Helena da Silva Azevedo4
Sandra Helena da Silva Azevedo4 Anndson Brelaz de Oliveira5Carlos Dias de Almeida Júnior4Eleyson Barboza da Silva4
Anndson Brelaz de Oliveira5Carlos Dias de Almeida Júnior4Eleyson Barboza da Silva4 Jânderson Rocha Garcez5José Ribamar da Silva Pinto4
Jânderson Rocha Garcez5José Ribamar da Silva Pinto4 Liriann Chrisley Nascimento da Silva4
Liriann Chrisley Nascimento da Silva4 Midian Salgado Monteiro1Wander da Silva Rodrigues4Thiago Luiz Ferreira Anízio4Alfredo Luiz Belém Pontes4Ruth Lima Teixeira4Jefferson Moreira da Silva4
Midian Salgado Monteiro1Wander da Silva Rodrigues4Thiago Luiz Ferreira Anízio4Alfredo Luiz Belém Pontes4Ruth Lima Teixeira4Jefferson Moreira da Silva4 Wallice Luiz Paxiúba Duncan3Richard Carl Vogt6†
Wallice Luiz Paxiúba Duncan3Richard Carl Vogt6†- 1Laboratory Wild Animal, Faculty of Agricultural Sciences, Department of Animal and Plant Production, Federal University of Amazonas, Manaus, Brazil
- 2Institute of Social Sciences, Education and Animal Science, Federal University of Amazonas, Manaus, Brazil
- 3Institute of Biological Sciences, Federal University of Amazonas, Manaus, Brazil
- 4Community Chelonian Management Program—Pé-de-pincha Project, Manaus, Brazil
- 5Federal Institute of Education, Science and Technology of Amazonas, Manaus, Brazil
- 6Amazonian Chelonian Studies Center, National Institute of Amazonian Research, Manaus, Brazil
Chelonians represent an important resource in the Amazon, either as a source of protein at the base of the food chain of aquatic and transition ecosystems, or in the dispersion of seeds of plants from floodplains and flooded forests. The consumption and predatory exploitation of their meat and eggs by local populations has been, and still is, one of the main threats to these animals. Community-based conservation projects allied to official protection programs have been restoring populations of chelonians of the genus Podocnemis throughout the Amazon since 1974. In this study, we analyzed the historical time series of protection data of Podocnemis expansa, P. unifilis, P. sextuberculata and P. erythrocephala in areas protected by the government and communities in the Amazonas state and northwest of Pará state. Between 1974 and 2019, 230,444 nests and 21,350,201 hatchlings of P. expansa, 170,076 nests and 3,229,821 hatchlings of P. unifilis, 647,715 nests and 6,410,092 hatchlings of P. sextuberculata and 24,617 nests and 168,856 hatchlings of P. erythrocephala were protected. Community protection schemes emerged in 1990, and covered 80.7% of the areas and produced 64.2% of P. unifilis hatchlings and 44.6% of P. sextuberculata hatchlings. The areas with the highest production of P. expansa remain under government protection (57.4%). Using the time series of production of nests and hatchlings per beach, logistic growth curves were estimated, and the values of r and K were compared between the two protection systems (government and community). Beaches controlled by the government showed higher support capacity in the production of nests (1,910.7 ± 1,035) and hatchlings (211,513 ± 93,031) of P. expansa and P. sextuberculata (81,160 ± 34,924 hatchlings). However, the communities were more efficient in protecting nests (r = 0.102 ± 0.2315) and hatchlings (r = 0.282 ± 0.166) of P. unifilis. Community-based protection and monitoring programs are an important component that should be incorporated by the government’s environmental agencies for the management and conservation of turtles in the Amazon.
Introduction
At approximately 240 million years old, chelonians are one of the oldest reptile groups. Of the 360 species currently recognized worldwide, 56–61% are threatened (Rhodin et al., 2018; Stanford et al., 2020). The Amazon is considered one of the regions with the greatest wealth of chelonians in the world, comprising 18 species, 11 of which are aquatic, 5 semi-aquatic and two terrestrial species (Rueda-Almonacid et al., 2007; Ferrara et al., 2017). However, this diversity may be much greater (Carvalho et al., 2016; Cunha et al., 2019; Vargas-Ramírez et al., 2020). Five species of Podocnemididae are found in the region: the giant South American river turtle, Podocnemis expansa (Schweigger, 1812); the yellow-spotted river turtle, P. unifilis (Troschel, 1848); six-tubercled river turtle, P. sextuberculata (Cornalia, 1849); red-headed river turtle, P. erythrocephala (Spix, 1824); and the big-headed turtle, Peltocephalus dumerilianus (Vogt, 2008).
Chelonians play an important role in the ecosystem as the basis of food chains in aquatic, transitional and terrestrial environments (Campos-Silva et al., 2018) and assist in seed dispersal, mineral cycling and carbon storage (Jerozolimski et al., 2009; Nascimento et al., 2015; Lovich et al., 2018). They are a food source for a wide diversity of predators ranging from invertebrates such as ants, fly larvae, northern mole cricket, to vertebrates, such as fish, alligators, birds, lizards and even jaguars (Salera et al., 2009; Andrade et al., 2016; Erickson and Baccaro, 2016). Beaches where a large number of turtle nests occur provide positive co-benefits for other aquatic and terrestrial animal species that are attracted there (Campos-Silva et al., 2018).
Amazon turtles synchronize their life cycle with the variations in the water level of the rivers and lakes (Alho and Pádua, 1982; Vogt, 2008). During the flood season of the Amazonian rivers, these flooded areas are used as places of shelter and for feeding (Garcez et al., 2012; IBAMA, 2019a). Chelonians have mostly fruit-based (17–30.8%) and seed-based (17–18.9%) diets (Fáchin-Téran and Vogt, 2014; Garcez et al., 2020; Oliveira et al., 2020) thereby acting as potential seed dispersers and play an important role in the dynamics of the regeneration of floodplains and flooded forests (Eisemberg et al., 2017).
The turtles have developed features such as a protective shell and a life history characterized by seasonal and mass egg laying, producing many hatchlings (r strategists), with delayed maturity, prolonged reproductive life and great longevity that has allowed for adaptation in many ecosystems. These same features have made them vulnerable to extinction in the face of threats caused by man (Stanford et al., 2020).
Chelonians have always been an important food resource for the riverine populations of the Amazon (Pezzuti et al., 2010; Andrade, 2017). The native Indians, especially those ethnicities that lived in the floodplain areas, exploited this resource by consuming the eggs and meat of turtles (Smith, 1979; Prestes-Carneiro, 2013; Meza and Ferreira, 2015), with evidence that some of these species had areas of their current distribution influenced by human translocations in pre-and post-Colombian periods (Guix, 2020).
With the arrival of the Portuguese colonizers in the sixteenth century, this exploitation became even more intense, with millions of turtles of the Podocnemis genus being slaughtered. Millions of eggs from these turtles were also used for the production of oil for public lighting, in the preparation of food and even mixed with pitch to caulk sailing vessels (Schneider et al., 2011; Fiori and Santos, 2013, 2015; Andrade, 2017). It was only in 1849 that the first prohibition appeared in the Amazon region, Brazil, to protect the nesting beaches in the Solimões, Amazonas and Negro Rivers, since these species, mainly P. expansa, had begun to disappear (Andrade, 2015). In 1967, the Brazilian Federal Government, through law N° 5,197/67 (the Fauna protection law), prohibited the capture and marketing of wildlife, thus making it illegal to exploit turtles—a practice that for centuries had been carried out by the peoples of the Amazon, and as a result caused an impact on the extractive economy of the region (Benchimol, 1999; Antunes et al., 2016, 2019).
The main threats to the chelonians in the Amazon are the rampant overexploitation of adults and eggs (Pantoja-Lima et al., 2014; Morcatty and Valsecchi, 2015; IBAMA, 2019a; Charity and Ferreira, 2020), the loss and degradation of aquatic and terrestrial habitats (feeding, dispersal, reproduction, nesting) due to deforestation, the expansion of urban areas (Conway-Gomez, 2007; Bowne et al., 2018) and the implementation of hydroelectric dams, highways, mining, and even uncontrolled tourism on nesting beaches (Fagundes et al., 2018; ICMBio, 2018; IBAMA, 2019a).
The commercial capture of chelonians is one of the factors that, even today, contributes most to the decline of aquatic and terrestrial turtle populations throughout the Amazon (Hernández and Espín, 2003; Fachín-Terán et al., 2004; Pezzuti et al., 2010; Schneider et al., 2011; Norris and Michalski, 2013; Penaloza et al., 2013; Pantoja-Lima et al., 2014; Harju et al., 2017; Morcatty et al., 2020). In general, these resources are exploited by local communities for subsistence consumption or sold to nearby cities or large regional centers such as the cities of Manaus, Santarém and Belém (Canto et al., 1999; Andrade, 2008; Nascimento, 2009).
It is estimated, based on the seizure data of the environmental agencies and the police, that between 1992 and 2011, 86,949 chelonians (13,289 P. expansa; 3,933 P. unifilis; 19,279 P. sextuberculata, 474 P. erythrocephala, 49,583 Podocnemis sp., 184 Peltocephalus dumerilianus, 195 Chelonoidis sp. and 9 Chelus fimbriatus) and 42,941 eggs were seized in the Amazonas state alone (Nascimento, 2009; Andrade, 2015), which represents from 52 to 57% of all animals seized (Canto et al., 1999; Nascimento, 2009). Between 2012 and 2019, 11,894 chelonians (29% P. expansa, 27% P. unifilis, 7% P. sextuberculata, 5% P. erythrocephala, 1% P. dumerilianus and 31% Podocnemis sp.) and 16,090 eggs were seized (Charity and Ferreira, 2020).
With an annual average of seizures of 4,347 chelonians/year and 2,147 eggs/year, between 1992 and 2011, and decreasing to 1,487 animals/year and 2,011 eggs/year, between 2012 and 2019, this may indicate a reduction of enforcement and control actions in the region, but could also be because of improved protection.
In 1979, the project “Chelonians of the Amazon” emerged, through which the government began to protect the breeding areas of Podocnemididae that still existed in the Amazon. In addition, in the 1990s, several conservation activities were developed by the Amazonian riverine communities, especially to organize fisheries agreements (Pinto and Pereira, 2004). This participation in the process of implementation and monitoring the management of aquatic resource has been called co-management, community management or participatory management (Berkes, 2009; Freitas et al., 2009; Campos-Silva et al., 2020).
In 1999, in the middle of this period of changes, the Federal University of Amazonas (UFAM) created, in partnership with IBAMA and riverines of Terra Santa, a community-based chelonian conservation program in the lower Amazon, called “Pé-de-pincha,” has already returned 5,204,849 chelonian hatchlings back to nature (Andrade, 2017). Other actions of community conservation of chelonians have been recorded in the Brazilian Amazon (Miorando et al., 2013; Waldez et al., 2013) as well as in other countries of the Amazon Basin and Orinoco River (TCA, 1997; Soini, 1999; Towsend, 2008; Hernández et al., 2010; Harju et al., 2017).
Most of the protection programs for P. expansa and P. unifilis in the Amazon have sought to protect breeding females, nests and hatchlings, which seems to have contributed significantly to averting these species from the risk of extinction (Cantarelli et al., 2014; IBAMA, 2016; Pezzuti et al., 2018; Forero-Medina et al., 2019; Norris et al., 2019). However, turtles are long-living animals, and models that use demographic estimates of age, growth, fertility and survival are key to their management (Spencer, 2002; Zimmer-Shaffer et al., 2014). For Podocnemis expansa, there have been few studies aimed at estimating their population growth (Diniz and Santos, 1997; Norris et al., 2019; Rachmansah et al., 2020).
In order to adequately monitor conservation efforts, it is necessary to know if there is an increase in the populations of turtles through the work of protecting eggs and hatchlings in managed areas, both in areas protected by the State, and in areas protected by communities, and whether there are differences between these management systems. The systematization of the protection data of the nests and hatchlings of turtles in areas protected by the federal government or programs in community-based conservation in the state of Amazonas, would be one way. This, together with population attributes of the structure and the population dynamics of these stocks, will allow us to estimate population models for local turtles of the Podocnemis genus. Thus, we will able to evaluate the efficiency of these programs and predict the possible future impacts of these types of management systems on the conservation of these species.
The present study had the following objectives: (a) characterize and evaluate the different chelonian conservation and management systems for Podocnemis expansa; P. unifilis; P. sextuberculata; P. erythrocephala in the Amazonas state and in northwest Pará state, Brazil; (b) analyze the production data from the official federal environmental agency protection system of chelonian nesting beaches (1974–2019) and the conservation data from the community management system of chelonians (1999–2019) in these areas of the Amazon; (c) generate mathematical population models that simulate the evolution of the production of nests and hatchlings from the data of the management systems of chelonians analyzed; (d) estimate the intrinsic growth rate (r) and the support capacity (k) from the models generated.
Materials and Methods
Area of Coverage and Geographical Location
This study was developed from the analysis of the historical data series (1974–2019) of conservation of chelonians of the Amazon Chelonian project (PQA) of the Brazilian Institute of Environment and Renewable Natural Resources (IBAMA) and the National Center for Research and Conservation of Reptiles and Amphibians (RAN) at the Chico Mendes Institute of biodiversity (ICMBio). Data was obtained from the locations that participate in the Pé-de-pincha Program of the Federal University of Amazonas (UFAM) in the physiogeographic zone of the middle-lower Amazon River and on the Trombetas, Nhamundá, Uaicurapá, Andirá, Marau, Uatumã, Madeira, Negro and Tefé Rivers, for the period from 1999 to 2019; and also in the areas of PROBUC (Biodiversity Monitoring Program and the Use of Natural Resources) on the Juruá and Purus Rivers, and from the Center for Preservation and Research of Aquatic Chelonians (CPPQA), on the Uatumã River, over the same period. The map of the sites used in the analyzes is presented in Figure 1.
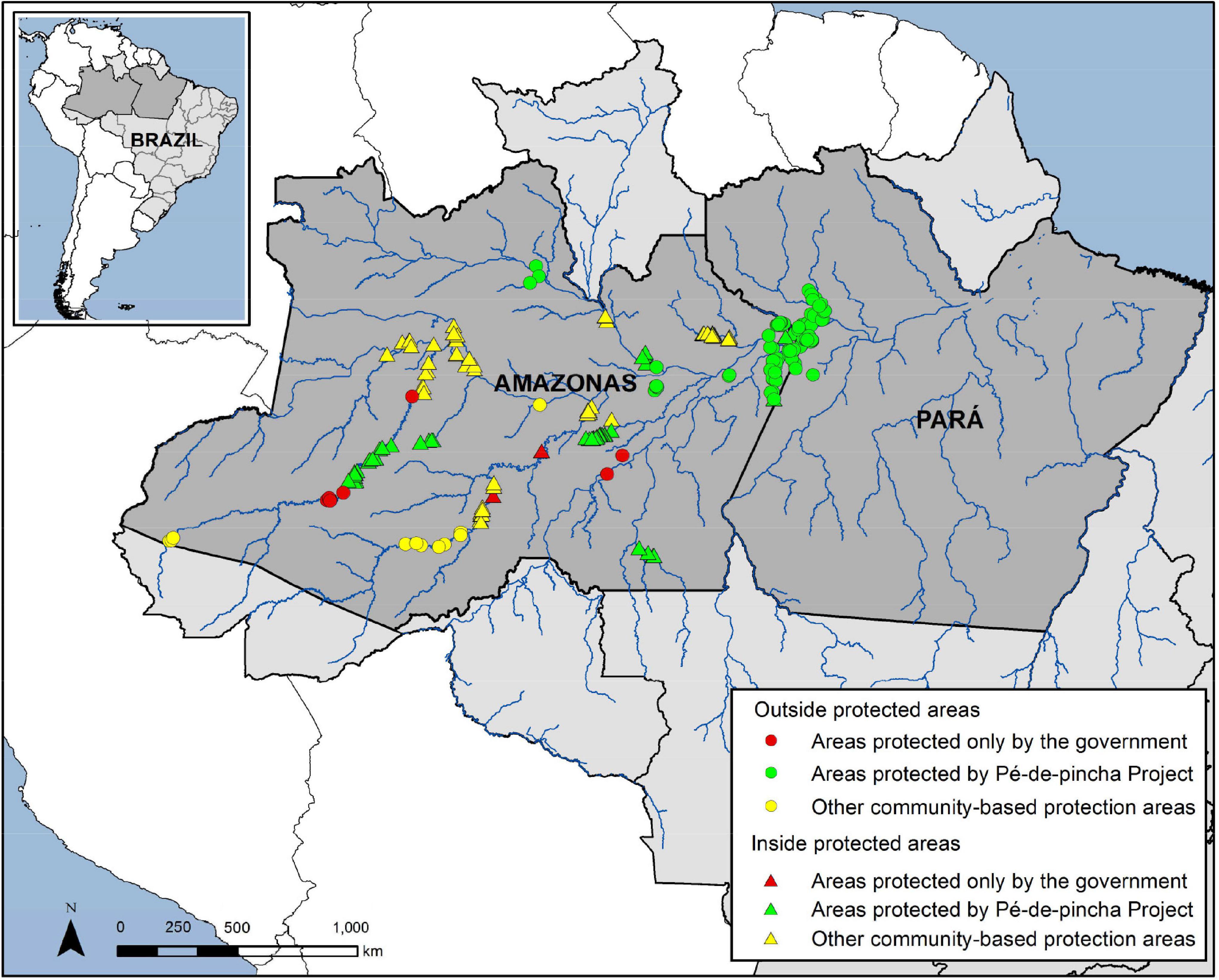
Figure 1. Chelonian conservation areas in the Amazonas state and the northwestern section of the Pará state, Brazil.
Methodology
We analyzed 82 technical reports and field records of the areas protected by PQA—IBAMA in the Amazonas state, as well as the data provided by the Chelonian Integrated Data System (SISQUELONIOS) of the RAN—ICMBio, for the period from 1974 to 2019. In addition, we analyzed 138 beach protection permits that were granted in the period between 1964 and 2004 by the respective environmental agencies: Department of Hunting and Fishing of the Ministry of Agriculture (until 1966), 58% of the permits; Brazilian Institute of Forest Development (IBDF) from 1967 to 1989, 27%; IBAMA between 1989 and 2004, 14%; and permits from the respective municipalities, 1%.
The chelonian management systems adopted in this period in different areas and rivers were identified and characterized. The existing data on the number of nests, eggs and hatchlings of giant South American turtles (Podocnemis expansa), yellow-spotted river turtles (P. unifilis), six-tubercled river turtles (P. sextuberculata) and red-headed river turtles (P. erythrocephala) and the time and resources applied in the conservation work of the chelonian nesting beaches by the federal government were inserted in a database and tabulated in a spreadsheet (Excel 2013) and then analyzed.
Similarly, data on the number of nests, eggs and hatchlings of the studied Podocnemis species and the time and resources applied in the work of community conservation of chelonians through the Pé-de-pincha program and in the PROBUC, for the period from 1999 to 2019 were also tabulated and analyzed. These data were analyzed and used for the formulation and simulation of mathematical models.
Mathematical Modeling, Model Validation and Statistical Analysis
The data were tabulated and two sample groups were considered: (1) the production of chelonians in areas with exclusive protection by the government (federal or municipal), and (2) in areas subject to community management. The production data of nests and hatchlings by species on each beach were related to the length of protection time each area had received. After the Pearson’s correlation analysis, regression analysis (linear, quadratic and polynomial) was performed, through which the existence of growth trends in the number of nests and hatchlings on each nesting beach was verified. These analyses were performed using the statistical programs MINITAB and STATISTICA v.7. The parameters of the regressions with the best fit were used in the population growth models: logistic and Gompertz (Barry, 1995; Gotelli, 2007). The curve estimates were made using the PAST 2.08 statistical program, and subsequently defined as a general model of analysis of the logistic curve:
- Logistic model:
where Nt = total number of nests/egg-laying females or hatchlings produced in time t; No = number of nests, egg-laying females or hatchlings produced in the first year of protection work; r = intrinsic population growth rate; E = Napier’s logarithmic constant = 2.717; K = carrying capacity (Gotelli, 2007).
With the curve models established for each breeding site for each species, the coefficients r and K of the growth models of the number of nests and hatchlings were defined. K values were determined by the maximum population density. The values of r and K were obtained directly from the models estimated by the PAST program, or through the first derivation of the logistic curve of population growth. The support capacity, K, was also estimated by regression between N and r, with K being equal to the value at which the line cut the X axis. The value of r was estimated from K and the size (N) of the population (Krebs, 1986; Brower et al., 1989). When the rate of intrinsic growth (r) was greater than zero, it meant that the population was growing; if it was equal to zero, it meant that it had stabilized; and, if it was less than zero, it meant that it was declining (Krebs, 1986). For each site, species and population indicator (nests or hatchlings) analyzed, we estimated the values of the instantaneous growth rate (r), the carrying capacity (K) of the nesting site and the equation of the estimated logistic curve, as well as performing the AKAIKE adjustment for the model tested.
To validate the model, several simulations were performed with the estimated models, which generated annual data that were compared with the actual data obtained from the historical series (1974–2019) of production of the number of nests/egg-laying females and chelonian hatchlings produced in the areas under community management. The robustness of the models was also evaluated using data that were collected directly in the field by the authors (1999–2019).
To evaluate the efficiency of the existing chelonian protection systems in the Amazon, the nesting beaches were divided into three groups (areas in the Juruá River, areas in the Purus River and areas of the Pé-de-Pincha program, in the middle-lower Amazon river) and two treatments or protection systems (areas of exclusive management by the government and areas of community management). Each protected beach was considered an observation, and each year was considered a repetition. The variables analyzed were the instantaneous growth rate r and the carrying capacity K of the number of nests/egg-laying females and the total number of hatchlings released on each beach.
A two-way analysis of variance was applied with factor 1 being the river or physiographic zone and factor 2 being the system of management (Sokal and Rohlf, 1990; Ferreira, 1991) in order to compare the means of the rates of growth r and carrying capacity K in the production of the nest and hatchlings of the systems of conservation considered (government and community) for the three different species of turtles studied (P. expansa, P. unifilis, and P. sextuberculata), whereafter a post hoc Tukey test was applied (Sokal and Rohlf, 1990; Zar, 1999). Statistical analyses were performed using the MINITAB program.
Results
Analysis of Historical Data Series on the Conservation of Turtles in the Amazon
A total of 2,318 records of annual production of chelonians, relating to 109 areas and 244 nesting beaches (207 in the Amazonas state and 37 in the northwest of Pará state) in 16 river channels of the western Amazon were analyzed in the survey of the historical data series (45 years of information). Of these areas, 52.9% are located in federal or state conservation units (CU), and 47.1% are outside of CUs. The Pé-de-pincha program was implemented in 76% of these areas (184 beaches), 56% of them were outside CUs (Figure 1).
Between 1974 and 2019, 230,444 Podocnemis expansa nests, 170,076 P. unifilis nests, 647,715 P. sextuberculata nests and 24,617 P. erythrocephala nests were protected, which produced 21,350,201 P. expansa hatchlings, 3,229,821 P. unifilis hatchlings, 6,410,092 P. sextuberculata hatchlings and 168,856 P. erythrocephala hatchlings (Supplementary Table 1 presents all the annual protection data of the chelonian nests and hatchlings from the 1974–2019 historical series; and Supplementary Table 2 shows a summary of the total number of nests, eggs and hatchlings protected, as well as the recorded number of defective hatchlings, stillbirths, unviable eggs, predated nests, nests damaged by rainfall or erosion and the seizures recorded in the Amazon state).
The increase in the number of protected Podocnemis expansa nests and hatchlings was related to the increase in number of protected areas (Figure 2 and Supplementary Figure B).
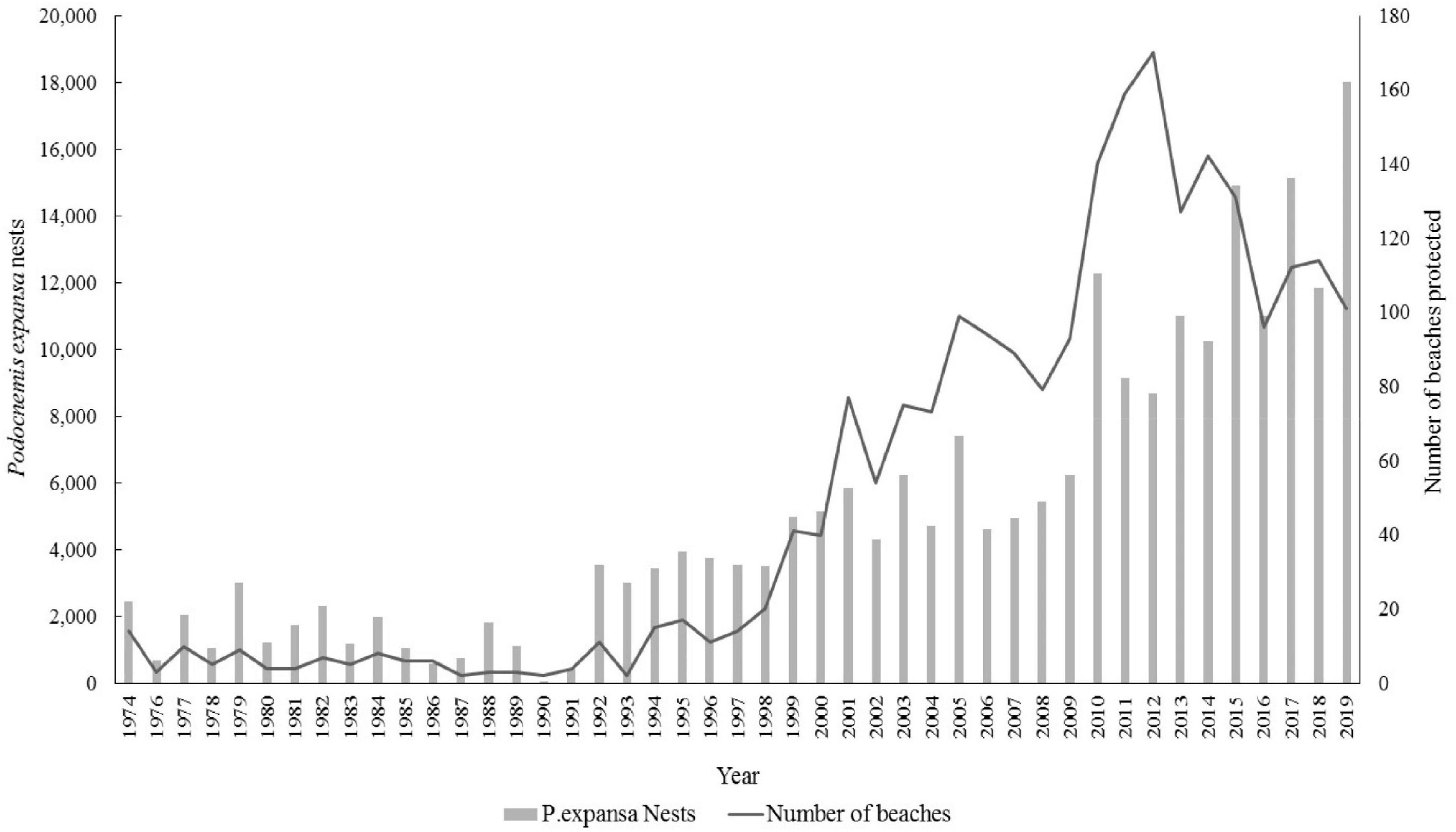
Figure 2. Nests of Giant South American turtle (P. expansa) protected in the Amazonas state and the northwestern section of the Pará state, Brazil, between 1974 and 2019. Source: PQA/IBDF/IBAMA CPPQA and Pé-de-pincha program technical reports.
From analysis only of the beach protection permits, it observed that were sent to 57 chelonian nesting beaches (locally known as “tabuleiros”) mainly in the Purus (36), Juruá (6) and Solimões (14) Rivers. Most of the permits were granted to the beaches managed by rubber plantation owners or their surviving heirs, mainly in the Purus (101). Before 1967, when the wildlife trade in Brazil was banned, the owners of beaches had an interest in this type of permit (34.5/year). Some of these authorized beach owners sold chelonians and their eggs to cover the expenses and earn income. After the ban, this number fell to 6.8/year.
For P. expansa, more consolidated information was available than for the other species, since this species is the main target of conservation efforts of environmental agencies. Not all reports presented data on nests and hatchlings of P. unifilis and P. sextuberculata, however, the data on P. unifilis, when recorded, were based on the marking of nests and counting of hatchlings, whereas the data on P. sextuberculata were based, for the most part, only on estimating the number of nests and hatchlings (Figure 3 and Supplementary Figure C).
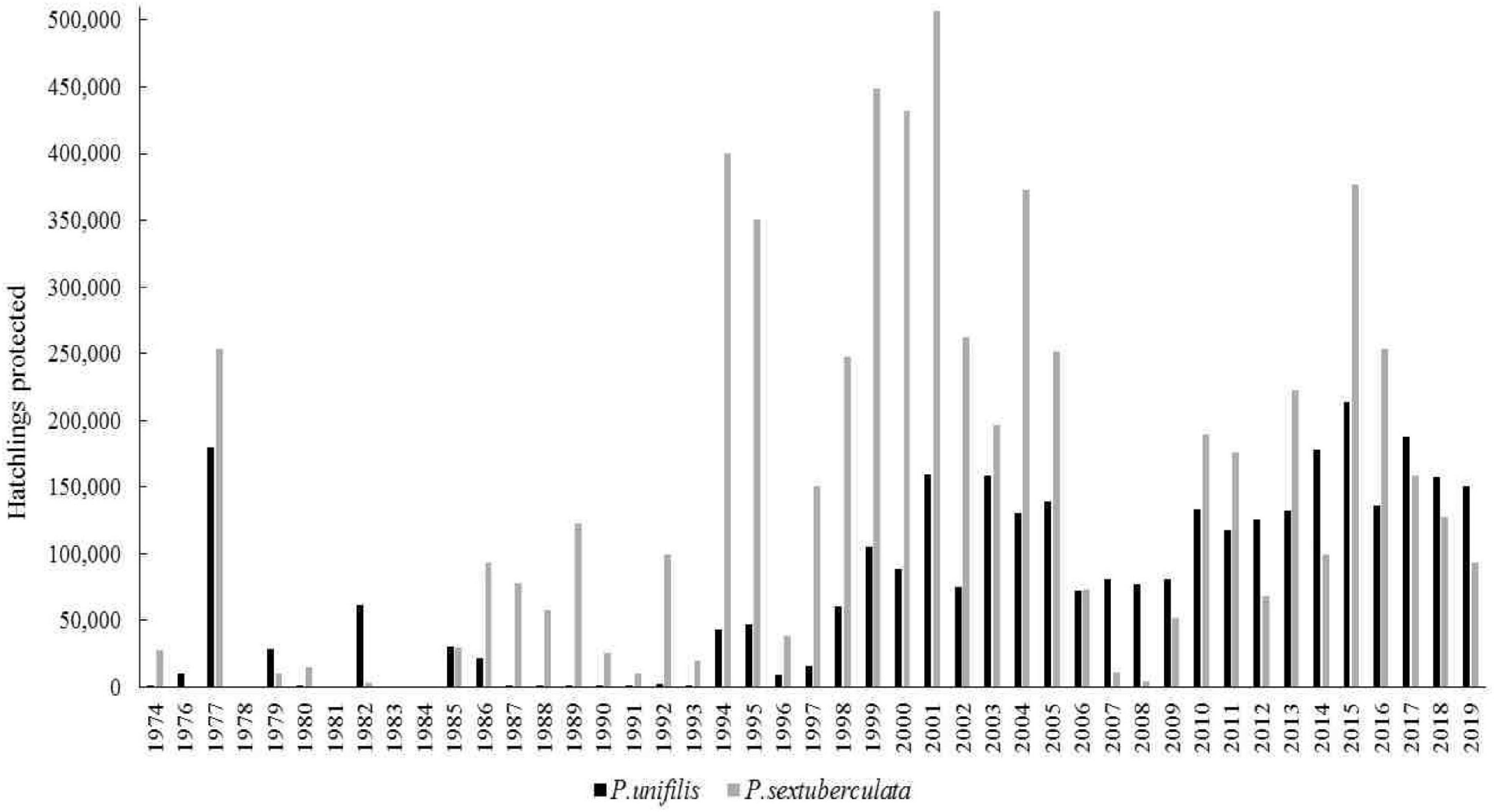
Figure 3. Yellow-spotted river turtle (P. unifilis) and six-tubercled river turtle (P. sextuberculata) hatchlings protected between 1974 and 2019 in the Amazonas state and the northwestern section of the Pará state, Brazil. Source: PQA/IBDF/IBAMA CPPQA and Pé-de-pincha program technical reports.
Data on Podocnemis erythrocephala only began to be systematically recorded from 1995 in communities in the Negro River and by the Pé-de-pincha program in the Nhamundá, Jamari, Andirá, Uaicurapá, Mamuru, Marau, Matupiri Rivers and Juruti Lake, and by CPPQA, on the Uatumã River (Supplementary Figure D). These groups protected 1,116 ± 942 nests and produced 7,675 ± 8,648 hatchlings per year.
With regard to the rivers where the protected beaches were located, initially, efforts were concentrated on the Purus, Juruá and Solimões Rivers. From 1999 onward, with the increase in the areas of community management, there was a diversification in the environments where the turtles were protected, and the expansion in the number of rivers covered, these being the middle-low Amazonas, Andirá, Sapucuá-Trombetas, Nhamundá, Negro, Madeira, Uatumã, Marau and Tefé Rivers.
The Purus River was responsible for 62.8% of the nests and 56.8% of the hatchlings of P. expansa protected over the 45 years in the Amazon, followed by the Juruá River (34.6% of the nests and 41.3% of the hatchlings). Across all other rivers, the production of nests and hatchlings reached 2.7 and 1.9% of the total, respectively (Supplementary Figure E).
Podocnemis unifilis hatchling production was 40.2% on the Juruá River, followed by the Purus River with 14.4% and the community protection areas of the Pé-de-pincha program for the middle-low Amazon River (14.2%), Sapucuá/Trombetas Lake (14.2%) and Andirá River (9.0%) (Figure 4).
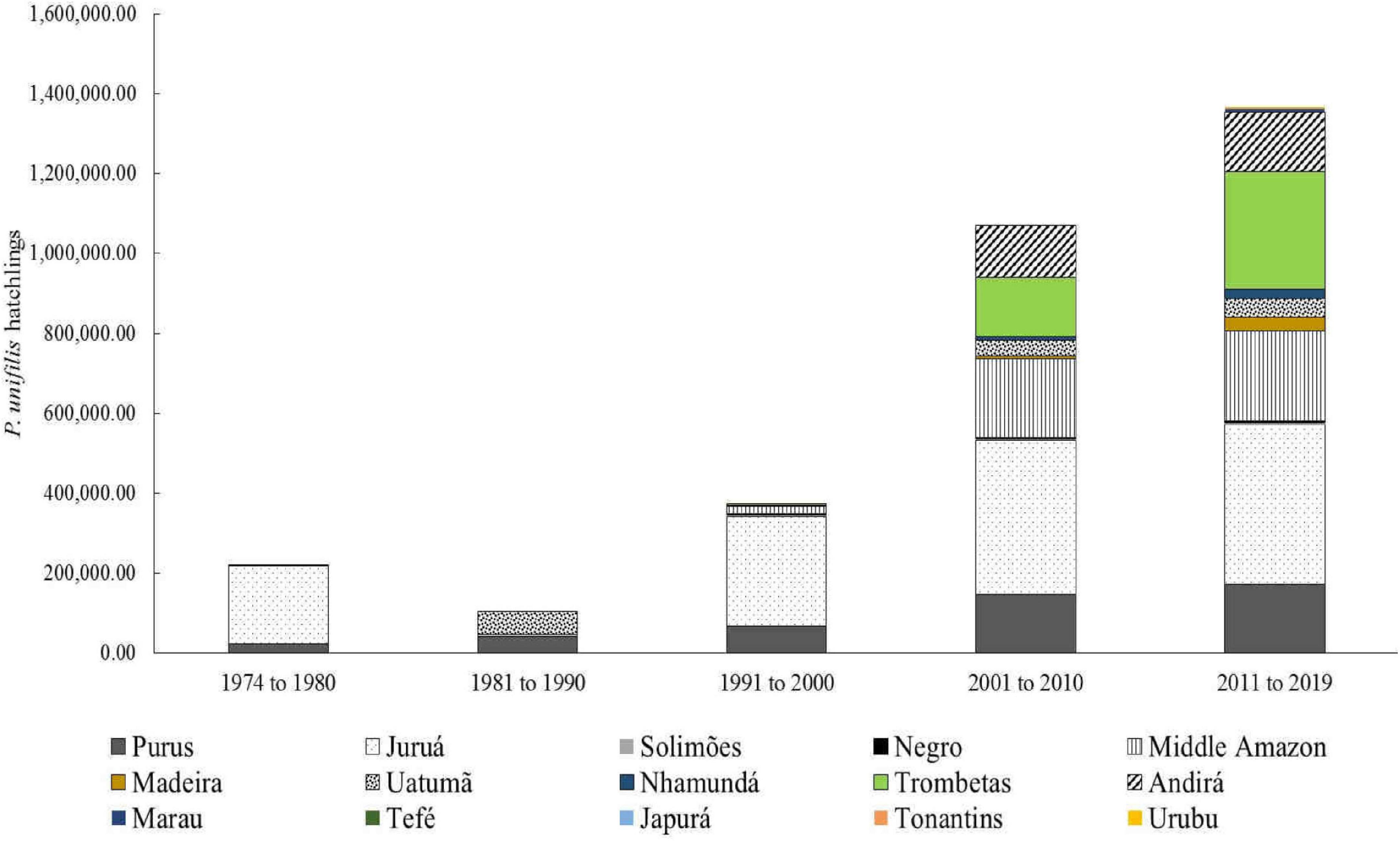
Figure 4. Number of P. unifilis hatchlings protected between 1974 and 2019 for different rivers in the Amazonas state and the northwestern section of the Pará state, Brazil.
For Podocnemis sextuberculata, 64.5% of the hatchlings were recorded in the Juruá River, 30.5% in the Purus, 1.6% in the middle Amazonas River and the rest in the other areas. In the case of this species, due to the large number of nests in the Purus and Juruá rivers, only estimates were performed. Only in community-based protection areas with few nests was it possible to check all nests and hatchlings.
In the Amazonas state, the following types of protection for the chelonian nesting beaches were identified:
(1) Owner of the beach or rubber plantation owner: The owner of the beach or “beach captain” (as he was locally known) that protected the nesting area by marking the beach with flags and preventing other people from removing eggs and adults from the area, and who had official authorization to do so. This system accounted for 100% of the protected areas from 1964 to 1976, when the federal government began to protect and monitor these areas. It ended in 2005. This system did not involve public funding and the protection work was carried out at the expense of the owner of the area (private initiative), but labor was provided by the local residents. Therefore, this work can be considered as a mixed form of participatory resource management and a precursor of community-based protection. It corresponded to 4% of all the analyzed protected areas.
(2) Government: These areas were considered as nesting areas protected by the federal government, and these involved protection carried out only by employees of the federal environmental agencies or by persons hired by them, without the participation of communities, and using federal resources. This system was used in the areas of large production of turtles in the Amazonas state (Purus and Juruá Rivers). It has accounted for 9.6% of all protected areas in the last 45 years.
(3) Community-based: The community-based chelonian protection system involved the communities of each region in all phases, from the perception and decision to protect these animals, the organization and labor for the execution of protection services and monitoring of the beach, recording of data (nests and hatchlings) and the holding of release events. The resources for these actions were obtained from the communities themselves or from partner institutions that supported these initiatives. This system may also involve the participation of environmental agencies providing logistical support, but the actions of protection are eminently community-based, and it is considered participatory management. Between 1974 and 2019, this system corresponded to 80.7% of the total protected areas.
(4) Municipal Authorities: In some municipalities, due to the absence of environmental agencies, the municipal authorities assumed the role of conservation of local turtles. Usually, they defined a municipal reserve area and made resources (financial, human and logistical) available for the execution of the monitoring and control of the nesting areas, but without the participation of the communities at any stage. This protection system corresponded to 2.9% of all evaluated protected areas.
(5) Specialized Centre: One specific case of protection by a specialized center was recorded. The Center for Preservation and Research of Aquatic Chelonians (CPPQA) belongs to a state company (Eletronorte/Eletrobrás) and began its chelonian conservation actions of as a form of environmental compensation, and then expanded them to the communities of the Uatumã River. The CPPQA worked in 2.8% of the areas analyzed from 1994 to 2019.
If we analyzed the number of beaches protected by the different systems of conservation of chelonians in the Amazonas state in the last four decades (Figure 5), it can be observed that from the 1990s there was an expansion of community-based protection systems. Which, in addition to replacing the system of the owner or captain of the beach, allowed for the exponential increase of the chelonian protection areas.
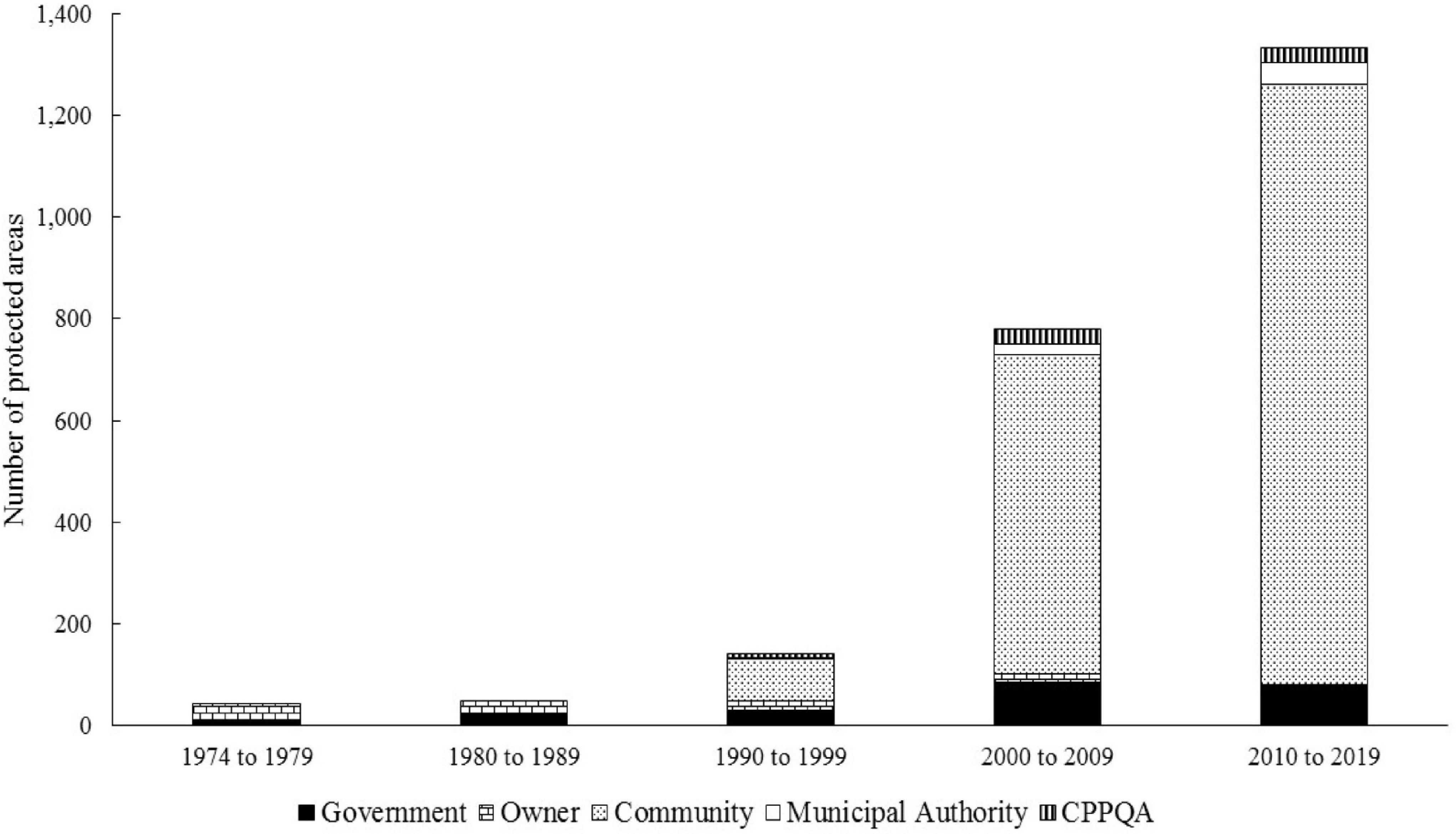
Figure 5. Number of chelonian nesting beaches protected by different management systems in the Amazonas state and the northwestern section of the Pará state, Brazil, between 1974 and 2019.
When analyzing the production of hatchlings in each protection system, it was found that although the beaches protected by the federal government correspond to only 10%, they account for the production of 13,732,700 hatchlings of P. expansa, or rather, 57.4% of the total protected turtles. The protected areas of the municipalities come in second place, with 23% of the total hatchlings produced. The community system accounted for 13.9% of protected turtle hatchlings (Figure 6). The increase in community-based protection of hatchlings of P. expansa between 1999 and 2019 was mainly due to the increase in the number of beaches involved in the Pé-de-pincha/UFAM program, which currently provides 93.9% of community protection of P. expansa.
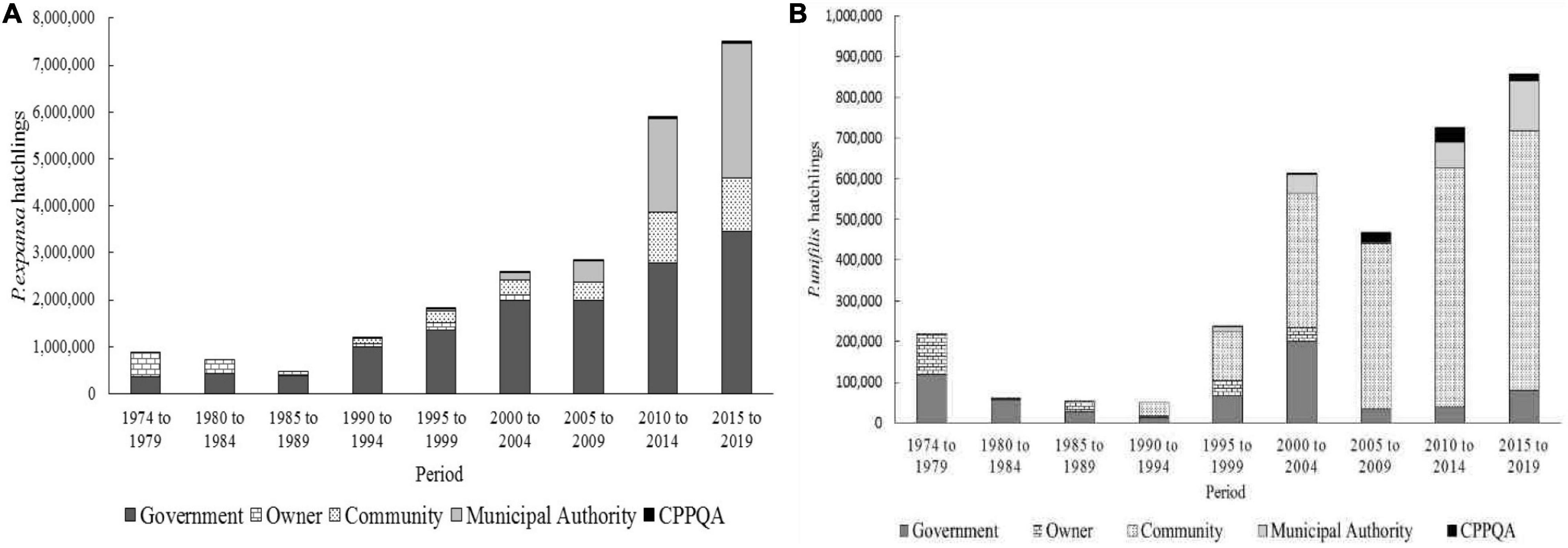
Figure 6. Participation of the different systems of protecting the production of chelonian hatchlings in the Amazonas state and the northwestern section of the Pará state, Brazil, between 1974 and 2019: (A) P. expansa; (B) P. unifilis.
When analyzing the production of Podocnemis uniflis hatchlings, it was found that the community conservation system is responsible for the protection of 2,112,070 hatchlings, or rather, 64.2% of the total production. Beaches protected by the federal government only come in second place with 19.4% of total production. Beach owners produced 6.3% and municipalities 7.6%.
The increase in the participation of the community-based conservation system from 1999 onward in the production of P. unifilis hatchlings, was mainly due to the increase in the areas of the Pé-de-pincha program, which has this species as its emblem. Initially, the program accounted for 15.9% of the community conservation of P. unifilis and, by 2019, it had reached 96.2% of the community production of hatchlings of this species.
For the production of hatchlings of P. sextuberculata, it was found that 44.6% of hatchlings were produced in areas of community management, 25% on beaches protected by federal environmental agencies, 20.1% by municipal authorities and 10.2% by beach owners (Supplementary Figure F). For P. erythrocephala, the first records of protection of the species only began in communities on the Negro River in 1995 and, from 1999, expanded to the Pé-de-pincha program areas. Of the hatchlings of P. erythrocephala protected, 58.8% were protected through the community conservation system, 40.3% by the government and 0.9% by the CPPQA. If we consider only the areas of community protection, the Pé-de-pincha program conserved 98.4% of the production of hatchlings of this species.
The different ways of managing the hatchlings of the protected nests was also evaluated (Figure 7). We categorized these according to the level of human intervention in the natural process of hatching and birth of offspring. Several levels of intervention were found then, the management of hatchlings was classified into three levels of intervention:
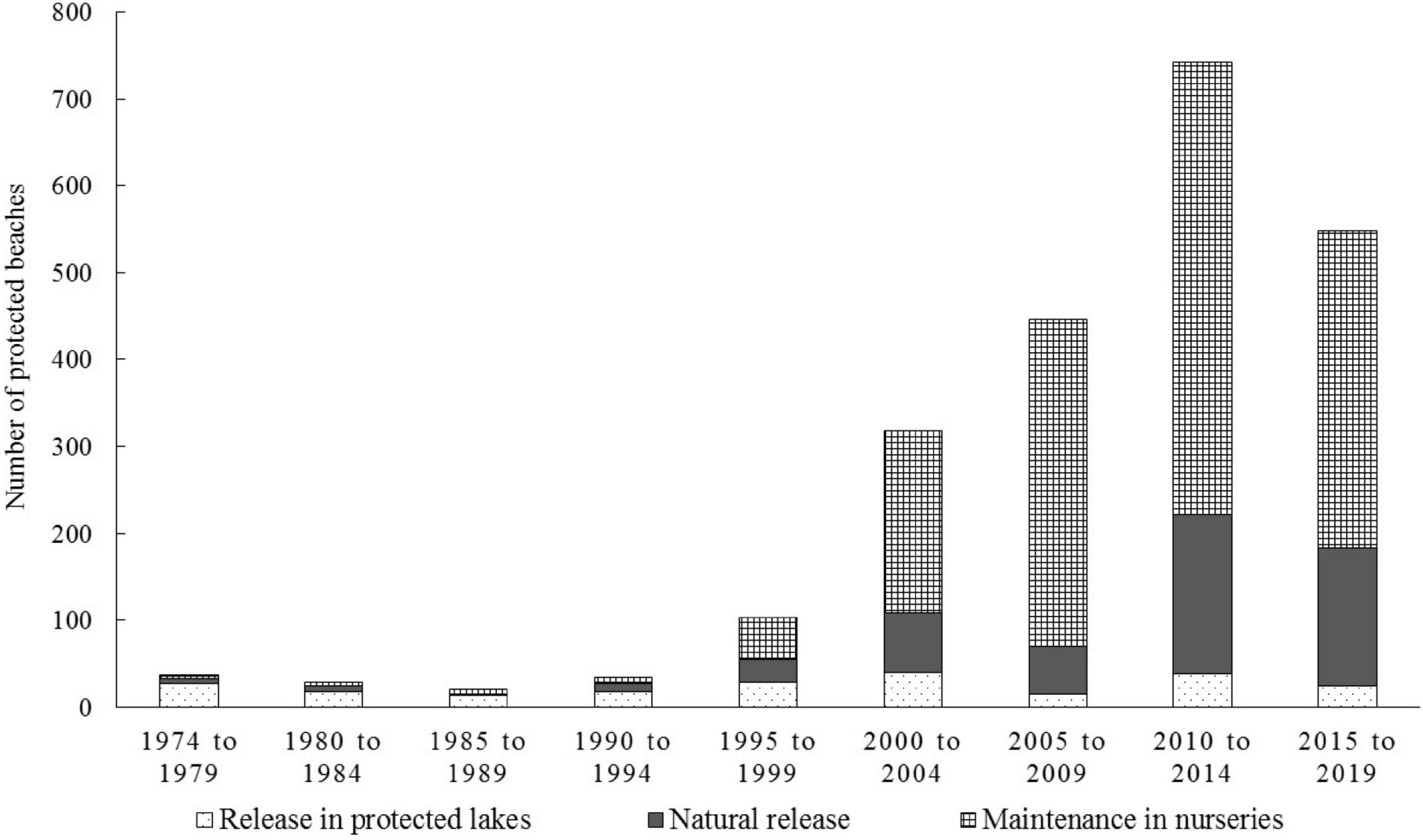
Figure 7. Number of protected beaches with different types of hatchling management: natural release; transport of hatchlings to lakes protected; and maintenance in nurseries or headstarting, for the period from 1976 to 2019, in the Amazonas state and the northwestern section of the Pará state, Brazil.
(1) Natural: We considered natural management of hatchlings to be when there was only the counting of nests, and the hatchlings emerged from the nests and proceeded to the river without human intervention. Or in places where it was impossible count the nests and only the hatchlings were counted. We also included places where the nests were opened manually and the hatchlings counted and released immediately on the shores of the beach.
(2) Transport of hatchlings to lakes protected from natural predators: This system was widely used by beach owners, and was performed in almost 100% of the areas until 1976. It consisted of the manual removal of the hatchlings (especially P. expansa) from the marked nests. They were then transported to lakes, away from the beach and away from the main aquatic predators of the hatchlings (fish and caimans). These lakes usually had enough aquatic macrophytes in which the hatchlings could shelter, as well as being places where adult turtles would go during the flood season of the river.
(3) Maintenance in nurseries or headstarting: this classification comprises the places that maintained the hatchlings in captivity for different periods before release (from 7 to 90 days providing supplementary feeding) and using different types of nursery tanks (fenced natural lakes, wooden cages, net-tanks, water tanks, etc.).
Of the P. expansa hatchlings protected, 51.4% were released immediately; 24.1% were transported to protected lakes and 24.4% were kept in a nursery for a period before being released. Among the protected P. unifilis hatchlings, 28.1% were released naturally, 10.9% were transported to lakes and 61% were kept in nurseries before release. In the case of P. sextuberculata hatchlings, 39.5% naturally emerged from their nests, 37.5% were transported to the lakes and 23.1% were kept in a nursery before being released. For hatchlings of P. erythrocephala, 36.4% were released naturally and 63.6% were kept in a nursery.
It was estimated that, in the 45 years analyzed, US$2,340,410 were directly invested in the protection of turtles in the Amazonas state and northwest of Pará state by IBAMA and the Pé-de-pincha program (equivalent to US$8,000 to US$52,000/year). The Pé-de-pincha program accounted for 87% of these expenditures between 2010 and 2015. The average cost per protected hatchling was estimated at US$ 0.25 ± 0.2. In government protected areas, the average cost was US$0.13 ± 0.12 per hatchling, and in community-based conservation areas, this cost was US$0.33 ± 0.19 per hatchling. Even though community-based protection was almost all voluntary, the costs per hatchling were slightly higher due to the need for training and the meeting of some socio-economic demands of the communities, such as donations of basic food supplies.
Turtle Population Growth Models in the Amazon
The initial analysis of the data was performed by constructing column graphs of the time series and evaluating the trend lines of the graphs for each species and beach analyzed (Figure 8). Only sites with a historical series of more than 10 years of data were analyzed.
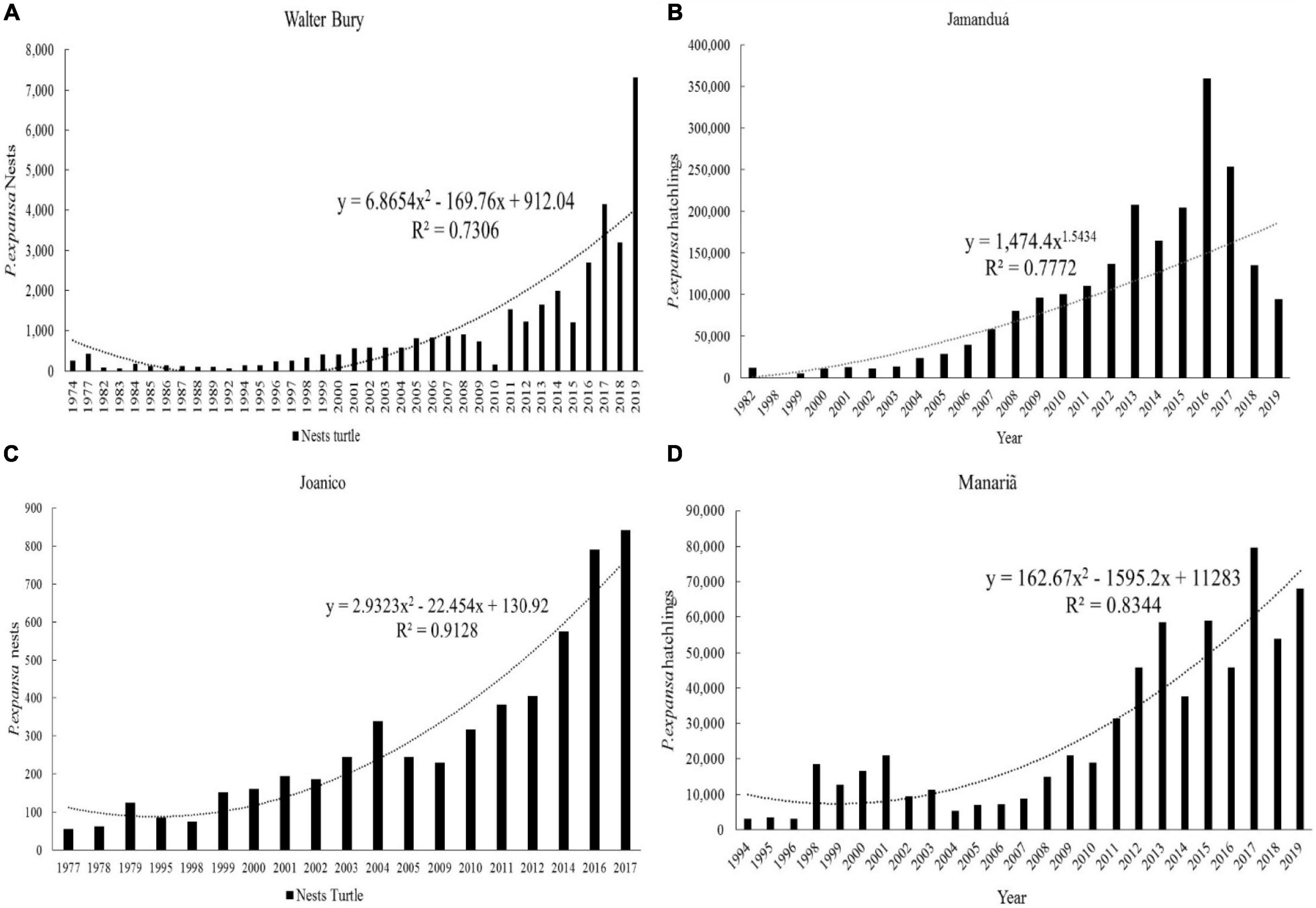
Figure 8. Time series of the production of P. expansa nests and hatchlings at nesting sites of the Juruá and Purus Rivers: (A) Nests—Walter Bury; (B) Hatchlings—Jamanduá; (C) Nests—Joanico; (D) Hatchlings—Manariã.
It was analyzed the time series of the annual production of nests and hatchlings of P. unifilis and P. sextuberculata in different nesting sites (Figures 9A–F).
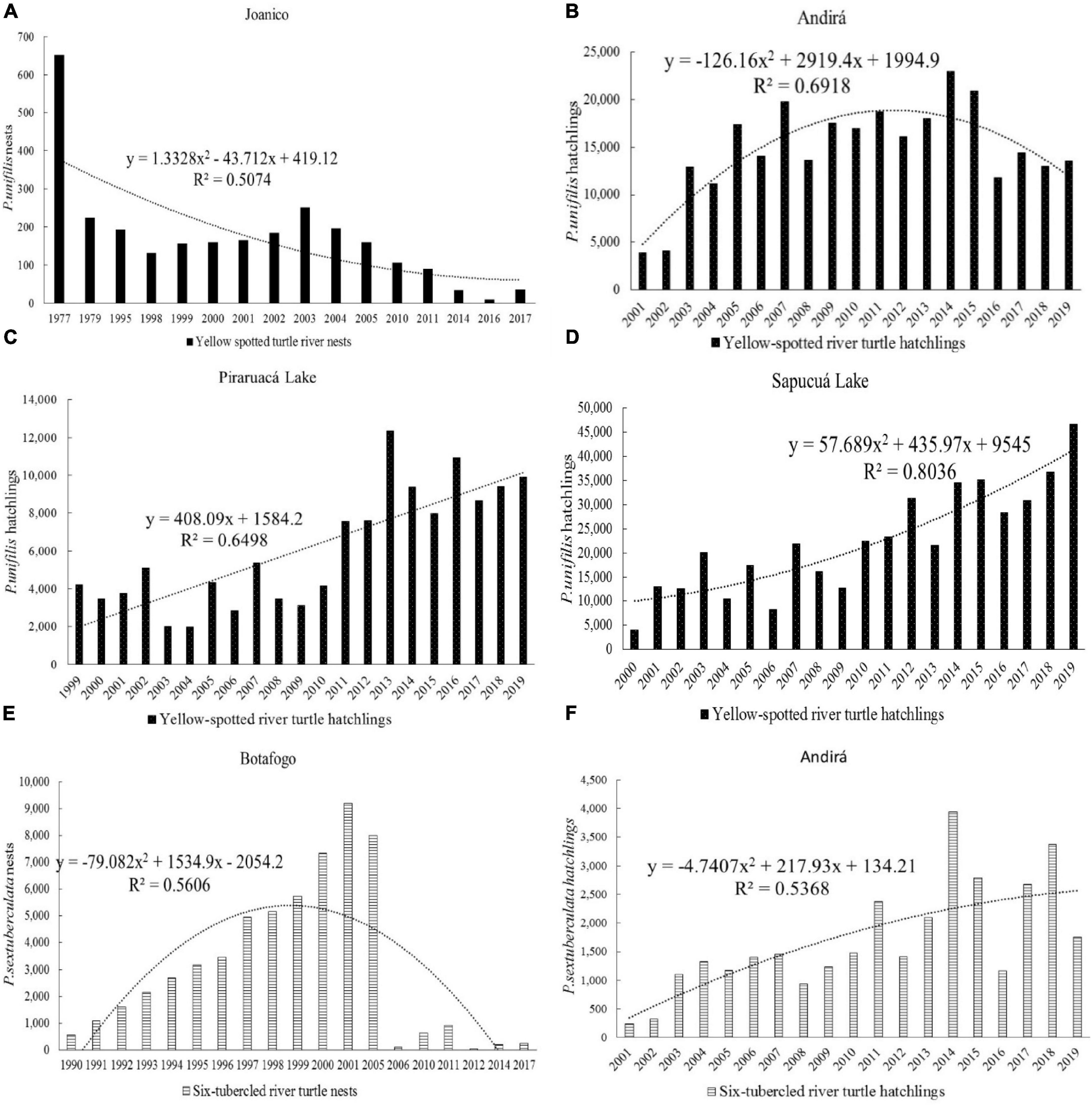
Figure 9. Time series for the production of P. unifilis and P. sextuberculata nests and hatchlings at different nesting sites: P. unifilis: (A) Nests in the Joanico, Juruá River; (B) Hatchlings in the Andirá River; (C) Hatchlings in the Piraruacá Lake; (D) Hatchlings in the Sapucuá Lake; P. sextuberculata: (E) Hatchlings in the Botafogo, Juruá River; (F) Hatchlings in the Andirá River.
After the analysis of the time series of the production of nests and hatchlings of the Podocnemis species studied for each beach, only those with R2 above 50% were selected to fit the logistic model. A total of 28 areas of the Juruá, Purus, middle-lower Amazon, Andirá, Nhamundá, Sapucuá/Trombetas and Negro Rivers were selected.
For each selected area, attempts were made to adjust the logistic growth to the curve model. For each area or nesting site analyzed, a curve relative to each species was generated (Figures 10A–D).
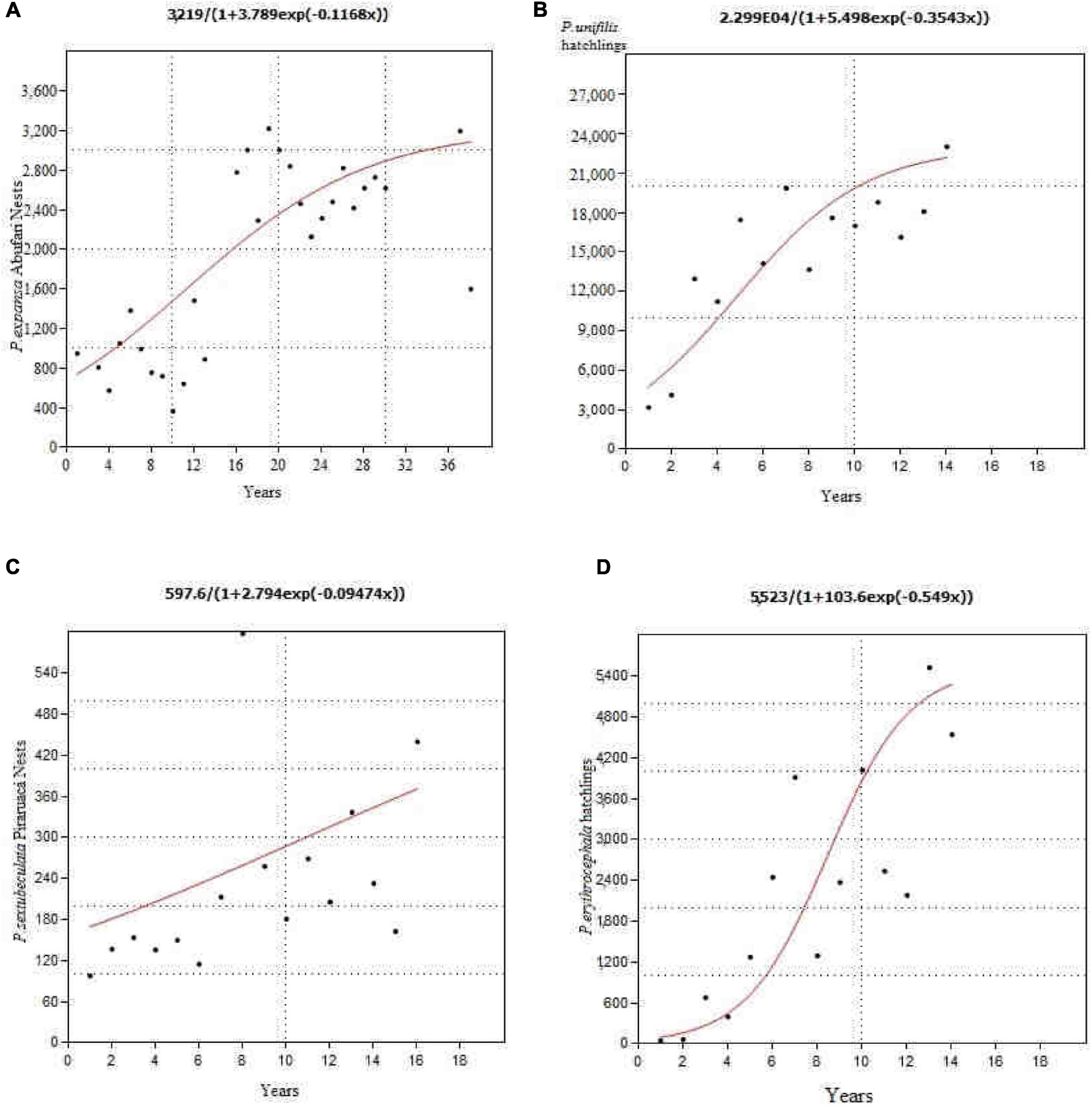
Figure 10. Logistic growth curves of the number of nests and hatchlings (A) nests of P. expansa, Abufari, Purus River; (B) hatchlings of P. unifilis, Andirá River; (C) nests of P. sextuberculata, Piraruacá Lake, middle Amazon; (D) hatchlings of P. erythrocephala, Andirá River.
In total, 160 logistic growth curves for number of nests/reproductive females and hatchlings were estimated and adjusted for the 28 breeding areas analyzed; 48 for P. expansa, 50 for P. unifilis, 50 for P. sextuberculata and 12 for P. erythrocephala.
There was a significant difference (F = 6.35, gl = 22, p = 0.02) between the growth rate r of the number of hatchlings of P. expansa from government protected areas (r = 0.172 ± 0.055) and those from communities (r = 0.085 ± 0.197), and also between the growth rate r of the number of nests/reproductive females (F = 4.97, gl = 22, p = 0.038)—Table 1. There was also a significant difference (F = 30.49, gl = 22, P < 0.0001) between the carrying capacity of the growth curves of the number of P. expansa nests and hatchlings in government protected areas (Knests = 1,910.7 ± 1,035 nests; Khatchlings = 211,513 ± 93,031 offspring), which was higher than those protected by the communities (Knests = 268.3 ± 303.5 nests; Khatchlings = 29,789 ± 35,716 hatchlings).
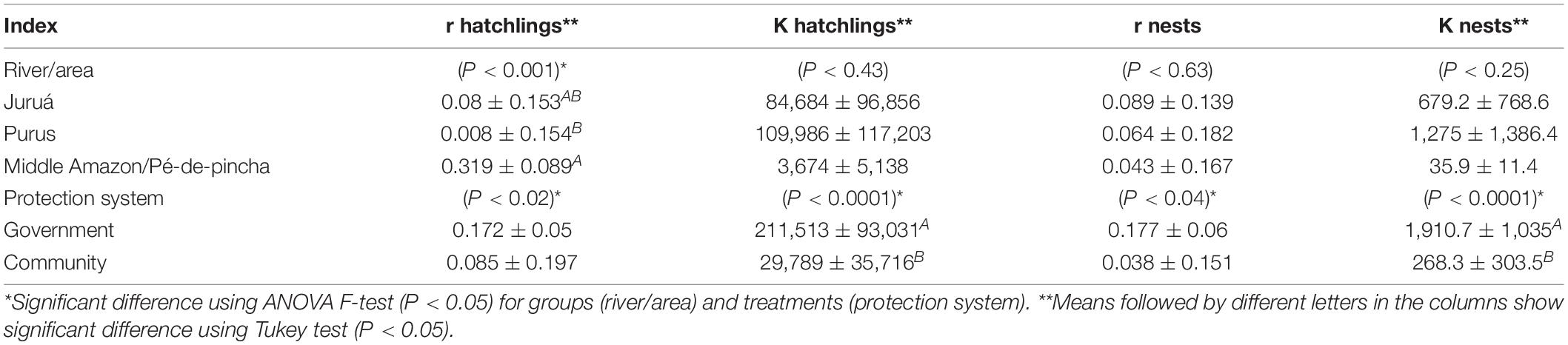
Table 1. Summary of the ANOVA of instantaneous growth rates r and carrying capacity K for number of nests/reproductive females and hatchlings of Podocnemis expansa.
The first two regions (Purus and Juruá) were older areas of protection of chelonians (30–45 years), which still had large populations of P. expansa, P. unifilis and P. sextuberculata. The last region covered the areas of action of the Pé-de-pincha community management program of chelonians, in the physiographic zone of the middle-lower Amazon, with an accumulated 20 years of protection.
It was found that the effect of rivers/areas was actually affecting the initial analysis. There is a significant difference between growth rates r in the number of hatchlings of P. expansa for river/nesting areas (F = 9.41, gl = 22, P = 0.001).
It was observed that, although there was a significant effect (P < 0.04 and P < 0.02) by the ANOVA, there is no significant difference between the means of nesting areas protected by the government and the communities for the instantaneous growth rate r of the number of nests and hatchlings of P. expansa when verified by the Tukey test (P < 0.05).
There was a significant difference (P < 0.001) between the rivers/protection areas for the r rate of the number of hatchlings of P. expansa. The rates (r) of the P. expansa hatchlings of the middle-lower Amazon/Pé-de-pincha project were higher (r = 0.319 ± 0.089) than those of the Purus River (r = 0.008 ± 0.154) and did not differ statistically from those of the Juruá River (r = 0.08 ± 0.153) when verified by the Tukey test (P < 0.05).
When the analysis on the K carrying capacity of P. expansa nests and hatchlings was performed, it was observed that there was no difference between the different areas. There was only a difference between the support capacity of the beaches administered by the government, with much higher averages (Tukey, P < 0.0001) of nests (1,910 ± 1,035) and hatchlings (211,513 ± 93,031) than the beaches of the communities. This is related to the fact that the federal government has chosen the areas with the largest populations and prioritized the protection of P. expansa, in addition to having greater power of supervision and control than the community has to protect these large stocks of P. expansa from being targeted by the traffickers of turtles.
For P. unifilis, there was no significant difference in the carrying capacity (K) of nests and hatchlings in the reproductive sites analyzed, neither by the different protection systems (F = 3.18, gl = 25, p = 0.089), nor between different areas (F = 1.12, gl = 25, p = 0.344)—Table 2. We found that the stocks of reproductive females nesting in sites protected by communities are similar to the average amount managed by government agencies. Acting in a greater number and diversity of areas (80.7%), the community system of protection of chelonians is responsible for most of the production of hatchlings of P. unifilis (64.2%), unlike P. expansa, where most of the production is in areas protected by the government.
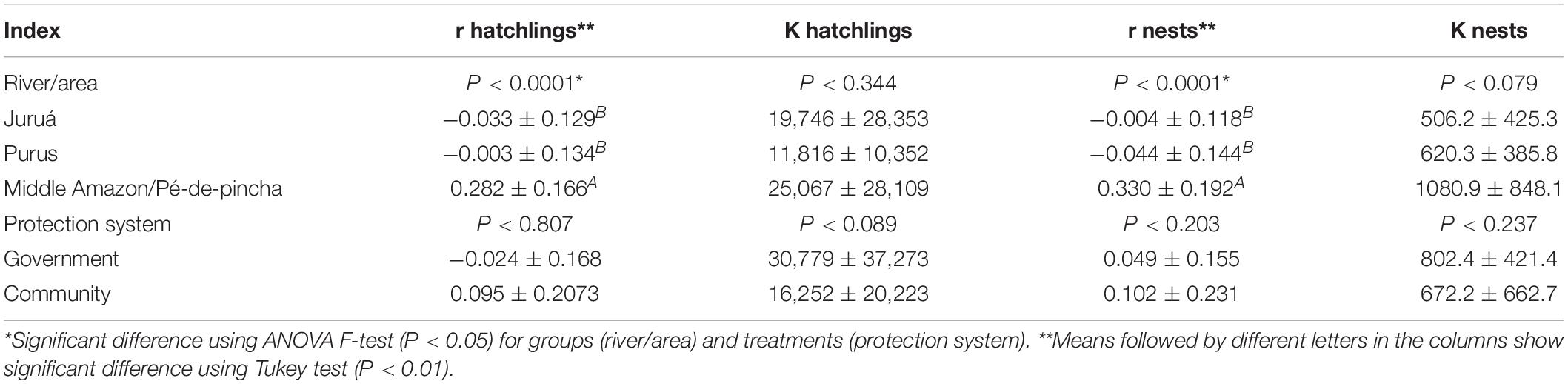
Table 2. Summary of ANOVA of instantaneous growth rates r and carrying capacity K for number of nests/reproductive females and hatchling of Podocnemis unifilis.
Significant differences for the rates of growth (r) of the number of nests and hatchlings (F = 11.52, gl = 25, P < 0.0001) were found between the rivers/areas evaluated. The areas of the middle-low Amazon where the Pé-de-pincha project operates showed growth rates of P. unifilis nests that were higher (0.33 ± 0.19) than the areas of the Juruá (−0.004 ± 0.12) and Purus (−0.044 ± 0.144) Rivers.
The negative r values found for most beaches of the Juruá and Purus Rivers (above 60%) could simply mean that the data for this species were underestimated by the lower number of records by beach monitors who would be favoring records of nests of P. expansa. However, when analyzing the historical production series of these areas, it was also found that, as the number of P. expansa females nesting on a beach increased, the number of nests of P. unifilis decreased in that area (Figure 11).

Figure 11. Comparison between the trend lines of the production of nests of P. expansa and P. unifilis: (A) Manariã beach, middle Juruá; (B) Joanico beach, lower Juruá.
Some of these beaches have been systematically monitored by the authors for more than 20 years, ensuring the authenticity and the same effort to record data for both P. unifilis and P. expansa, and, as such, this negates the possibility of underreporting of data for P. unifilis. This phenomenon had already been reported by the beach monitors of the Juruá and Purus Rivers, and was confirmed with the field data.
In the nesting areas analyzed, the number of P. expansa females laying eggs exceeds that of P. uniflis by around 13.7 ± 5.2 years of beach protection.
For P. sextuberculata, no significant differences were found between growth rates (r) of the number of hatchlings (F = 2.43, gl = 27, p = 0.132)—Table 3. However, there was an effect (F = 10.9, gl = 27, P = 0.004) of the protection system on the growth rate (r) of the P. sextuberculata nests, which was higher in the areas protected by the government (r = 0.219 ± 0.228). There was also an effect (F = 4.09, gl = 27, P < 0.03) of the protection system observed on the carrying capacity (K) of production of hatchlings, which was higher in the areas protected by the government (K = 81,160 ± 34,924 hatchlings), although there was no difference in the capacity of nests between beaches protected by the government and the communities.
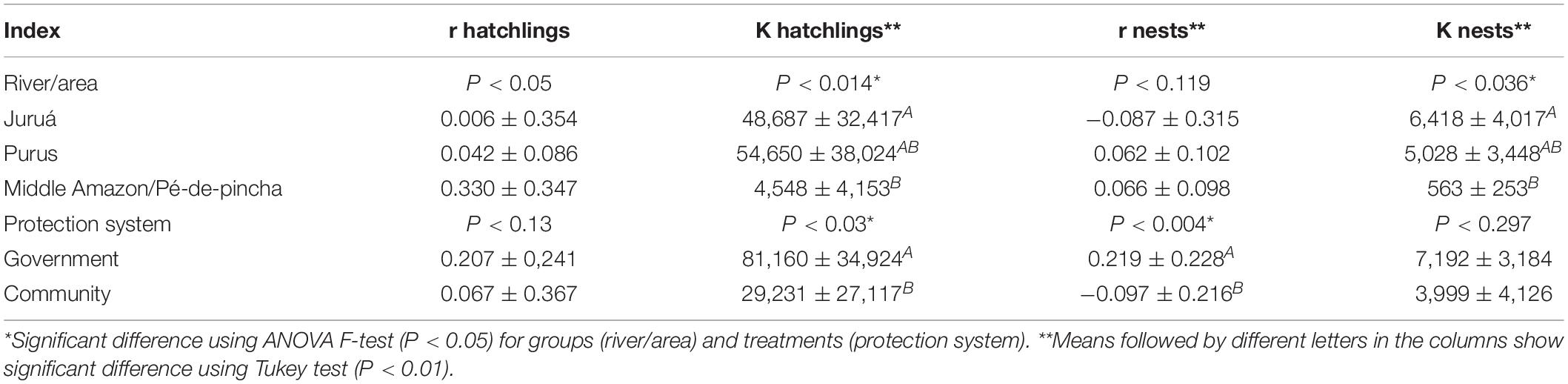
Table 3. Summary of the ANOVA of instantaneous growth rates (r) and carrying capacity (K) for number of nests/reproductive females and hatchlings of Podocnemis sextuberculata.
For carrying capacity (K) of nests and hatchlings of P. sextuberculata, there were differences between the rivers/areas analyzed (P < 0.04 and P < 0.01, respectively), and the beaches of the Purus and Juruá Rivers presented higher production of hatchlings and a greater number of nests than the areas of the middle Amazon River. In addition to the Purus and Juruá Rivers being white water rivers and, therefore, having greater distribution and abundance of this species, there seems to be a positive association between the increase in populations of P. sextuberculata and P. expansa, which is contrary to what occurs with P. unifilis. The competition for space on the beach for egg-laying seems to be eased by the difference in nesting periods, as P. sextuberculata usually reach their peak egg-laying before the P. expansa.
It was observed that in the Juruá River, even with its greater support capacity for P. sextuberculata nests, the populations presented negative instantaneous growth rates (r) for the nests and very low rates for the hatchlings, which may indicate the reduction of these stocks.
This could be just an underestimation of the data due to the lack of records of the number of nests and hatchlings of P. sextuberculata on each beach. In fact, due to the huge amount of simultaneous egg-laying of P. sextuberculata (sometimes more than 1,000 nests per night), this would require great effort to count the nests. In addition, the communities that monitor the beaches of the middle Juruá River have stopped estimating the production of nests and hatchlings since 2007, when the PROBUC protocol was implemented in the state conservation units, which prioritized only the registration of data for P. expansa and P. unifilis.
However, it should be noted that this specie was the most caught for consumption and sale (377 nests and 32,756 animals) according to the seizure data and consumption data collected by PROBUC and Andrade (2008, 2017), and it is possible that these negative rates actually indicate the reduction in stocks of P. sextuberculata in the Juruá River.
It was not possible to perform the same ANOVA for P. erythrocephala because curves were estimated for only 12 sites, that form of community protection systems. This species showed growth rates (r) for number of nests and hatchlings (r = 0.42 ± 0.3) that were higher than those for the other species analyzed.
To estimate a model of the general logistic curve for each species according to the protection system, 72 growth curves of nests/reproductive females on the beaches were selected, and the curves with growth rate (r) with negative values were eliminated. Simulations were performed with the selected curves up to 50 years of conservation, and the generated values were used to estimate a general growth curve of the number of nests/reproductive females of each species as a result of the protection system (Figure 12).
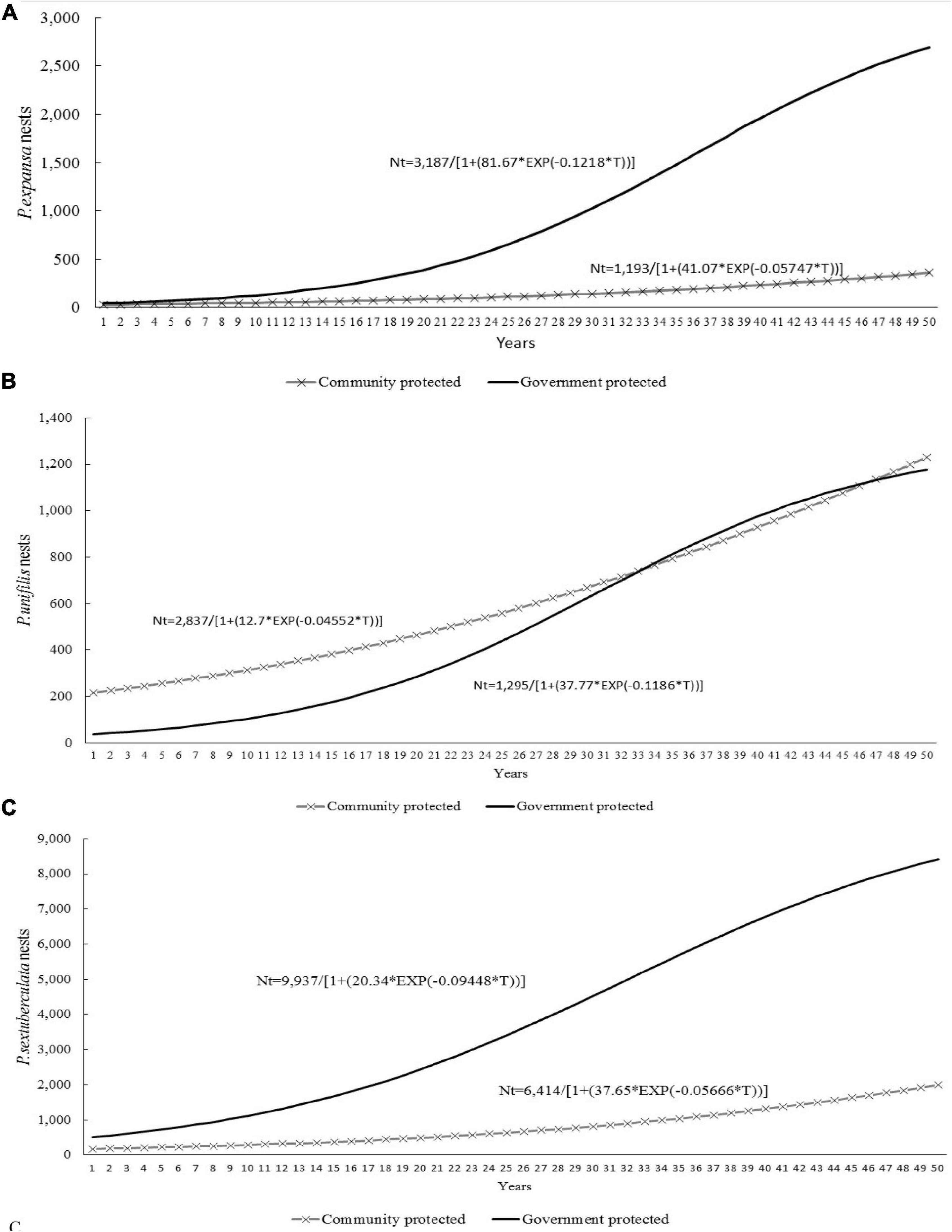
Figure 12. Logistic growth curves of the number of nests/reproductive females in the Amazonas state and the northwestern section of the Pará state, Brazil: (A) P. expansa; (B) P. unifilis; (C) P. sextuberculata.
After estimating the logistic growth model curves by species and by protection system, the proposed models were validated by comparing the estimated values to the real values using the Spearman correlation, and for most of the curves, the correlation was significant.
Discussion
Analysis of the Historical Data Series for Conservation of Chelonians in the Amazon
The analysis of the historical data series of chelonian protection in the Amazonas showed that an increase in the number of protected areas directly provided an increase in the number of protected nests and hatchlings. However, when comparing the 230,444 nests and 21,350,201 hatchlings of P. expansa protected for 45 years (5,121 ± 4,417 nests/year and 474,449 ± 411,488 hatchlings/year) with the estimates of 400–500,000 turtles females laying eggs in the Solimões and Madeira Rivers in the nineteenth century (Bates 1863 apud Smith, 1974 and 1979), it was noticed that these populations of P. expansa perhaps represent less than 1–2% of the original population in the Amazonas state.
However, it should be considered that in the surveys done in the 1970s by the IBDF in the 26 main beaches of the Solimões, Uatumã, Purus and Juruá Rivers, only between 1,750–2,767 egg-laying P. expansa females were recorded (Alfinito, 1978; Corrêa, 1978). In the first 10 years of protection, total production was estimated at 9,323 turtles nests in just two areas (Abufari and Walter Bury) in the Amazonas state (average 1,767 nests/year) (IBAMA, 1989). Cantarelli (2006) and Cantarelli et al. (2014) estimated that from 1975 to 2004, 6,163,521 hatchlings of P. expansa (2,732 nests/year) were protected on 12 beaches of the Purus and Juruá Rivers in the Amazonas state, with an increase of 25% per year, and the cost of protection per hatchlings was estimated at US$0.09.
In this study, when analyzing only the last 10 years (2009–2019), the average number of egg-laying P. expansa females in protected areas in the Amazonas state increased to 11,681 ± 3,321 animals/year, which represented an increase of 636% in 45 years (14% per year), with a cost of US$0.1 to 0.5 per protected hatchling.
Cantarelli et al. (2014) estimated that there were 30,000 P. expansa egg-laying females in the Brazilian Amazon. Forero-Medina et al. (2019) estimated that there are 147,000 P. expansa females in six Amazonian countries (Brazil, Colombia, Bolivia, Peru, Venezuela and Ecuador) of which 109,473 females were in Brazil alone in 2014. In this study, the estimated number of protected P. expansa females in the Amazonas state would represent only 10.6–16.4% of the total estimated turtle females for the whole of the Amazon. According to IBAMA (2019a) and Fagundes et al. (2021), only between 9.4 and 10.7% of priority areas for conservation of this species receive protection in the Amazonas state.
For P. unifilis, the analysis of the historical series (1974–2019) showed a production of 170,076 nests and 3,229,821 hatchlings (mean of 3,779 nests/year and 71,774 hatchlings/year) and, for P. sextuberculata, a production of 647,715 nests and 6,410,092 hatchlings (mean of 14,394 nests/year and 142,446 hatchlings/year) was observed. Cantarelli (2006) reported that 9,240,264 P. unifilis and P. sextuberculata hatchlings (318,629 hatchlings of the two species/year) were protected by the PQA (1975–2004).
With regards to the production of P. expansa hatchlings, it was found that the federal environmental agencies prioritized the protection of the most productive areas of turtles, while the communities usually conserved areas with smaller populations of turtles (a result of intense human predation) or in environments where there is a greater occurrence of P. unifilis, thus prioritizing this second species.
It was also found that, for the smaller species of Podocnemididae (P. unifilis, P. sextuberculata and P. erytrocephala), the community conservation system managed to cover a larger number of areas and that produced larger quantities of hatchlings for the restocking of the areas, while for P. expansa, the protection system of the federal environmental agencies managed to protect larger quantities of nests and hatchlings, in a few high production nesting areas.
The community conservation of chelonians began to gain strength between 1990 and 1999, by which time it protected 58.2% of the areas, and expanded to 80.6% of the areas between 2000–2009, until it became responsible for the protection of 88–94% of the chelonian breeding areas in the Amazonas state between 2010 and 2019. The conservation actions have been developed by the Amazonian riverine populations, including the adoption of conservation management systems of turtles with the management of resources guided by various forms of social organization, in which local residents and all stakeholders are key (Pinto and Pereira, 2004; Lima et al., 2017).
The mobilization and organization of communities to protect the fishery resources in the lowlands of Solimões/Amazon Rivers eventually influenced the emergence of community initiatives for conservation of chelonians, as was the case of the Pé-de-pincha Program in the middle Amazon River (Batista et al., 2004; Pinto and Pereira, 2004; Andrade, 2015) and community management areas in Santarém (Miorando et al., 2013).
The Pé-de-pincha Program is one of the largest community management and volunteer programs in the world. The name originates from the footprints that yellow-spotted river turtles leave on the beach, which are similar to bottle cap imprints (that the riverine community call “pincha”). Thus, an extension program and community management of chelonians was born in which the community members are trained and work voluntarily to protect nests and take care of the hatchlings until they return to their natural habitat (Andrade, 2012, 2015, 2017; Lima et al., 2017). The Pé-de-pincha Program has been implemented in 123 communities in 18 municipalities of the Amazonas and west Pará states, helping to protect 184 nesting sites (56% outside of conservation units) in an area equivalent to 2.7% of the Amazon (Andrade, 2017).
In order to stimulate and maintain the mobilized communities, in addition to protecting the turtles, the Pé-de-pincha Program has been active in training volunteers and seeking sustainable economic alternatives. It has trained 219 volunteer environmental agents and 148 environmental managers in techniques related to conservation of chelonians and raised awareness with lectures to over 86,507 participants, and trained 1,350 teachers of rural schools in environmental education (Andrade, 2017). To encourage income generation, it has held courses in fish technology, poultry breeding, medicinal plants, community gardens and breeding of turtles for 5,798 participants. The total number of people involved were 28,379 (directly) and 314,845 (indirectly) and, as such, it is one of the largest volunteer programs in the country (Andrade, 2017).
In the various conservation units (extractive reserves and sustainable development reserves) of the Juruá and Purus River, there are also community management systems that have helped protect more than 30 nesting areas of turtles, during more than 40 years of conservation work. Unlike the areas of the Pé-de-pincha Program, these are areas that still have large stocks of P. expansa, P. unifilis and P. sextuberculata (Andrade, 2008, 2015; Fonseca et al., 2011; Campos-Silva et al., 2018).
However, there are other areas in which there has been work on community management of turtles such as in the Sustainable Development Reserve (RDS) of Mamirauá (Oliveira, 2006); the RDS Uatumã, RDS Piagaçú-Purus (Waldez et al., 2013), in Juruti and in Aritapera/Santarém in the lower Amazon (Miorando et al., 2013). This has also occurred in the states of Amapá, Rondônia and Acre (Silva et al., 2019), as well as in other Latin American countries such as Venezuela (Hernández et al., 2010); Colombia (TCA, 1997); Peru (Soini, 1999; Harju et al., 2017); and Ecuador (Towsend, 2008).
In 1996, thanks to this conservation work by the federal government, together with numerous community protection initiatives, P. expansa was removed from the list of endangered animals in Brazil (Cantarelli et al., 2014; Forero-Medina et al., 2019). But, since 2003, the federal government has been reducing the volume of resources devoted to the protection of this specie in the region, which has caused a reduction of between 15.5 and 46.8% in the production of hatchlings on some beaches protected only by the environmental agency without community support (Andrade, 2008).
In the state of Amazonas, which covers 28.4% of the Amazon, by 2015, 212 chelonian breeding areas in 15 rivers had received some protection, mainly for nests and hatchlings, with an average production of 1,077,768 ± 115,466 hatchlings/year: 35.3 ± 26.1% of P. expansa; 19.3 ± 25.4% of P. unifilis and 40.9 ± 32.6% of P. sextuberculata (Andrade, 2017). Most of these areas were protected through the work of local communities interested in the maintenance of this natural resource, and these areas increased from 69 (58.2%) in 2001 to 193 (93.4%).
Andrade et al. (2004) analyzed the effective costs for the production of hatchlings protected with community-based management in the middle Amazon, and estimated an average value of US$0.6–0.8/hatchling and a profitability of 120.1%, when considering the estimate of the value of environmental services provided by the communities. Campos-Silva et al. (2018) estimated that the value for maintaining all areas of community-based management of chelonians would be around US$833,000 annually for the Brazilian Amazon.
Miorando et al. (2013) evaluated the effects of community management on the protection of P. sextuberculata in the lowlands of the Amazon, and noted that areas with community-based management have a higher number of catches of animals per units of effort, which shows the importance of this protection system.
Initiatives involving local communities and institutions carried out in the Amazon have resulted in increased protection of nesting habitats and recovery of Podocnemididae populations (Cantarelli et al., 2014; Andrade, 2017; Lima et al., 2017; Campos-Silva et al., 2018; Forero-Medina et al., 2019). Participatory community monitoring programs (PCM) have become an important methodological innovation for the management of biodiversity conservation in protected areas (Costa et al., 2018).
The idea of decentralization in the management of natural resources and the involvement of local populations has gained a lot of credit in the formulation of public policies and regional development projects, and, as such, avoids the classic options of privatization of resources or exclusive control by the State (IPEA, 2010). These changes have been occurring worldwide and have a strong emphasis on communities and local impacts of policies based on community management (Campos-Silva et al., 2020). Conservation actions and sustainable management of natural resources require initiatives also integrated by participatory monitoring and management, in which the information collected by resource users helps guide local decision makers on conservation management (Kennet et al., 2015). However, it is necessary to fully integrate the diverse interactions between individuals, communities and institutions and the complex Amazon ecosystem in this type of long-term environmental monitoring (Oliveira Júnior et al., 2020).
The chelonian conservation programs, carried out for a long time through the government environmental agencies (IBAMA, ICMBio) or through community management (Pé-de-pincha Program), have shown that by maintaining the protection effort, there is a tendency to increase the number of females laying eggs and hatchlings being produced on each beach (Andrade, 2015; IBAMA, 2016; Campos-Silva et al., 2018). With the regional strengthening of protected areas and community-based management, it is possible to anticipate a recovery scenario of freshwater turtle stocks in the Amazon in the coming decades (Norris et al., 2019).
In 2017, the Amazonas state officially recognized the efforts of the communities by creating 265 chelonian protection areas and regulating a community system of chelonian breeding, and allowing the communities to carry out breeding and fattening of a portion of the hatchlings of P. expansa (10%) and P. unifilis (20%) that they protect (CEMAAM resolutions N°25 and 26/17-DOE, DOE, 2017). This possibility of ex situ management of turtles to rear them in semi-natural conditions to be commercialized seems to be a solution to generate income and cover part of the expenses for the protection of the species (Campos-Silva et al., 2018).
The Amazonas state has the largest number of registered chelonian breeders in Brazil (27), with more than 150 thousand animals in captivity (P. expansa and P. unifilis) and an annual supply of 2,623 ± 561 animals/year (15 tons/year) (Andrade, 2008; Garcez, 2009; IBAMA, 2019b). This seems to be promising, not only from an economic point of view, but also for its relevance in food security and culture of the peoples of the region (Dantas-Filho et al., 2020).
Turtle Population Growth Models in the Amazon
The tabulation of the historical series of data of protection of Podocnemis species permitted us to analyze the variation in the number of females nesting on each beach and the number of hatchlings produced over the years. Although the number of nests do not necessarily indicate the number of reproductive females on the beach (especially in the case of species with multiple layings, such as P. unifilis and P. sextuberculata), and also do not constitute the total number of individuals in a population, as males and females that did not reproduce, young and hatchlings were not counted, it was considered that their variation over time could be the only consistent indicator of population variation in those protected breeding sites. The same can be considered with regard to the number of hatchlings produced, which is directly related to the number of nests, and which allows us to estimate the rate of hatching or reproductive success of each species on each beach, over the years.
It should be taken into account that, in very few (9.8%) of the sites analyzed were larger and long-term surveys carried out with animal capture, marking and biometrics, which would have allowed the recording of basic information about the population structure (sex ratio, size classes, abundance).
Considering that for Podocnemis expansa there are few studies aimed at estimating population growth models (Corrêa, 1978; Diniz and Santos, 1997; Norris et al., 2019; Rachmansah et al., 2020), it is understood that it would be more robust to use the real data of variation of nests/reproductive females and hatchlings as indicators of population variation.
Thus, we chose to use a simpler model of population growth to verify that the data of nests and hatchlings fit the models of growth curve tested [logistic, as suggested by Hailey and Lambert (2002)]. A model of continuous growth was used rather than a discrete model and with delay in density-dependent response, because when the intrinsic growth rates (r) are less than 1.0 (as shown to be the case of the analyzed species), the behavior for the discrete model is similar to the continuous one (Gotelli, 2007).
In this study, it was possible to use the regression coefficients of the non-linear curve model (r and K) as explanatory variables of the temporal trend of each nesting site and compare them with the coefficients of equations of other areas.
The growth rate of the number of nests/reproductive females per unit of time (rt) is specific for each species, as well as for the nesting site analyzed, and for a given period of time of protection of the area. Therefore, it made no sense to obtain the mean values of r for each protection system used.
The same reasoning applies to the carrying capacity (K) of each nesting beach in relation to the number of nesting females. The competition for space for the nests in the sand of the beach can reach such a level that there will be overlapping of nests and the females will end up digging up and removing the eggs of the others. In addition, some females would end up looking for other places to nest. In other words, each breeding site has a support capacity (K) for nests and hatchlings produced that is limited, mainly, by the space factor. This constant is also specific for each species and laying site and, therefore, it would not make sense to use the average value of K to generate a unique model for the population growth curve.
However, the population parameters r and K are values that have units that allow comparison between one conservation area and another. Units of r signify individual by individual and unit of time (Gotelli, 2007). Thus, it was possible to compare the instantaneous growth rates between the different protection systems used over time in the breeding areas of chelonians in the Amazonas state.
Bence (1995) suggested the use of long time series to estimate prediction equations, from which the estimates of the mean, the regression intercept would be more reliable, as well as for when the explanatory variable had a temporal tendency. Barry (1995) reported that several studies consider the non-parametric estimation S functions of time variation through non-linear models as the most appropriate method for estimating growth curves.
We have followed all of the steps set out by Pereira and Arruda (1987) in order to carry out the practical adjustment of a time series of data to the various models of the growth curve: (a) first, we assessed the correlation and regression analysis between the data of the production of the population (number of nests, number of hatchlings) and years; (b) then we estimated regressions that best fitted the relationship between the variables; c) when the regressions were significant, the simulations were done in fit of the curve non-linear models (Logistic and Gompertz); d) and, finally, when the best models were identified, these were validated by comparison with the real distributions obtained from the data for each beach.
Podocnemis expansa lay eggs in synchrony with the flow of the Amazonian rivers (Alho and Pádua, 1982), and offspring births occur in the months from October to December. This would characterize, according to Gotelli (2007), a discrete population growth model that, moreover, would be subject to environmental stochasticity (variability associated with good and bad years of production of hatchlings, as for example, with the loss of nests in large floods) and with delays in time (turtles take a long time to become reproductively active). However, according to this same author, if the growth rate (r) is small (r < 2.0) the behavior of the discrete equation is similar to the continuous curve.
For growth models with delay, May (1976) defined that the response time of the population is inversely proportional to the growth rate (r) (t = 1/r), and therefore, slow-growing populations have long response time, but if this delay quotient (r.t) is also small (0 < rt < 0.368), the population will also grow smoothly until the support capacity is reached.
In the estimates of the r values for the logistic growth curves of nests and hatchlings analyzed in this study, the values were always below 1.0, and behaved as a continuous model with a low delay quotient, which may be explained by the fact that, although there is a very large initial pulse of births, compensatorily, there is also a large pulse of mortality in the initial phase of life of these turtles.
For P. expansa, the lower and often negative r rates of the Purus River nesting areas seem to be linked to the following two factors: (1) the vast majority of community nesting areas were initially managed in the system of beach owners, suffered exploitation of their resources and were subsequently abandoned and looted, which caused the drastic reduction in the number of nesting matrices when compared to the initial stocks. (2) The second factor may be that the population is close to reaching the carrying capacity of the beach.
In the middle Purus, only the nesting areas of P. expansa under government protection had positive r rates, even so, with low values, as in the case of Abufari, the largest nesting areas of P. expansa in the Amazonas state, which presented r rates equal to 0.134 for hatchlings and 0.117 for nests, which could indicate a dampening of the population growth rate due to the number of egg-laying females being close to the support capacity of the beach.
Apparently, the same situation did not occur in the Juruá River, especially in the middle Juruá, where the process of community organization seems to have been stronger. The communities of rubber tappers have replaced the old beach owners in the defense and protection of the nesting areas. In addition, the environmental groups joined forces with the rubber tappers, and together they sought to create extractivist reserves that eventually further protected P. expansa nesting areas of the Juruá River.
On the other hand, it was found that in the areas of the middle Amazon/Pé-de-pincha, the r rates were higher, which can be explained by the fact that in these areas the populations of P. expansa were drastically reduced, and therefore there is a significant amount of environmental resources available for just a few animals. In this region, the population growth curve would still be at the beginning, thus, with a much higher initial growth rate. This can be seen as a good indicator of the potential for recovery of chelonian populations through community work.
For P. unifilis, the negative or low instantaneous growth rates (r) on the protected beaches of the Purus and Juruá rivers could be related to underreporting of data. But, the most likely hypothesis that the increase in recruitment of females of P. expansa that reach reproductive age increases the competition with females of P. unifilis for space on the river banks and on the beach sand. Because they are smaller than the P. expansa, females of P. unifilis are probably forced to look for other spaces on the beach closer to vegetation or at the ends of the beach, or even migrate to other areas, such as ravines or other beaches. Since these other sites are not monitored, the records only show the main beaches and these end up indicating a reduction in the number of female P. unifilis nesting on that beach.
Another assumption would be that the actual reduction of female of P. unifilis on the Purus and Juruá beaches is a result of the capture of adults and the harvest of eggs for consumption and illegal sale. The analysis of data regarding predation of nests (558) of P. unifilis, specifically in the protected beach areas, revealed that this was in fact greater than that of the predation of nests (300) of P. expansa. But, the seizures of P. expansa (20,915) was greater than that of the P. unifilis (14,505).
The higher rates of growth r recorded in the areas of community management under the Pé-de-pincha Program are probably due to the fact that this program has prioritized P. unifilis as a key species (rather than P. expansa), and protects some areas where only P. unifilis occur.
Another aspect is the fact that these areas are also depredated areas, with reduced stocks of the species, which means that in these places the population growth curve of P. unifilis is still in its initial phase, for which the intrinsic growth rates r are usually higher.
In small chelonian populations, such as those in the middle Amazon, the Allee effect is believed to occur, with growth rates being reinforced by the increase in population size, probably due to the greater reproductive efficiency of the chelonian groups (Gotelli, 2007; Silva et al., 2009; Silva, 2020).
According to Crouse (1999), population models for chelonians should provide a tool to predict the likely response of a population over time and, above all, allow us to identify data gaps, which is the predictive capacity of the model.
We started with long time series (15–45 years) for the production of nests and hatchlings. Instead of using rates or estimated predictive values, we used real data regarding what occurred over time with the population of females nesting at each site. The proposed models are a simple tool, based on analyses of trends in the production of nests and hatchlings, but that do not lose their predictive character as a population model. This tool can be enriched by incorporating information on population structure and dynamics from long-term studies that are already being conducted with P. expansa, P. unifilis and P. sextuberculata in the Juruá, middle Amazon and Andirá Rivers (Andrade, 2012, 2015).
Models in matrices can be used in ecological models, but they represent a device where the reality of it is partly sacrificed for the benefit of using the special properties of the matrix in the formulation of the model. However, it should be considered that modeling and simulation basically demand five key elements that are actual data in the field, a systematic framework for observation and recording of data (the time series of the variables), a proposed basic model, a derived aggregate model, and simulations (Santos, 1986), all of these prerequisites were used in our study.
Ehrhart et al. (2014) performed a trend analysis of time series of nesting data and reproductive success of Caretta caretta in which they used simple methods of evaluation through descriptive statistics of the data (graphics of the number of nests produced annually and tables with the annual data of nests, hatchlings and hatching rates), as was done in this study with the production data for Podocnemis species.
Diniz and Santos (1997) were the first to propose a qualitative study of the population growth of Podocnemis expansa by using a mathematical model that employed the Leslie matrix, with some data from biotic parameters. However, based on the value obtained from the Kojima quota for the biotic parameters (β ≤ 0.745296820391 < 1), he predicted that the populations of Podocnemis expansa would be extinct in Brazil, unless it was possible to increase the survival rate of offspring to 20% in the first year of life. Before him, Corrêa (1978) had already tried to propose a projection model for the offspring of a female of P. expansa for 30 years (evolution of the herd until the 4th generation). For this, he considered the average of 100 eggs/nest, sexual maturity of the mother at 8 years of age, useful life of 30 years, probability of the sexual ratio of the offspring of 50% males and 50% females.
It should be considered, however, that the models of Corrêa (1978) and Diniz and Santos (1997) are only models of reproduction projection, based mainly on the mean values of reproductive indices, such as number of eggs/nest and hatch rate of offspring, which when inserted deterministically in a matrix model estimate simulated values not based on comparisons with the actual data of field the over time.
The protection of eggs and nests alone was not enough to replenish depleted stocks of sea turtles. The models showed that small reductions in annual survival of juveniles and adults can have a profound effect on population dynamics by reducing population growth. Thus, the most complete models built for the sea turtle population, as well as models for the other species of turtles, point to the need to maintain high annual survival of all phases of life in order to recover declining populations (Crouse, 1999).
Norris et al. (2019) used a population projection model of the Lefkovitch matrix based on stages (eggs, hatchlings, juveniles and adults) to evaluate P. unifilis management scenarios, and estimated that large areas could be recovered if headstarting of hatchlings is practiced more widely.
Rachmansah et al. (2020) analyzed the biological viability of sustainable management in species of tropical turtles using life history data (number of eggs, age to sexual maturity, annual survival), latitude and climatic variables that were examined using generalized additive models (GAMs) and also the Lefkovich matrix population model structured in stages. Based on the tested modeling, these authors concluded that harvesting of adult tropical turtles would not be viable, but that sustainable egg harvesting would be.
A simpler population trend analysis protocol, such as the model proposed in this study, can help environmental managers and community leaders to better understand the dynamics and efficiency of their protective actions, and assist in decision-making.
Future Perspectives and Recommendations
Environmental agencies are giving more and more space to community-based management, investing in the implementation of participatory monitoring programs for natural resources. It is necessary that environmental managers work to have greater community involvement in turtle conservation programs, no matter how long this process may be.
Training in management and environmental awareness is gradually changing communities’ perception of turtles. These changes in perception will not necessarily bring about changes in the use of these animals for subsistence consumption, but they will certainly influence the creation of rules and agreements for use.
With the advancement of technology, new ways for community members to collect data from nests and hatchlings of turtles on protected beaches should be adopted. To replace the old field sheets, applications on cell phones for recording monitoring data and sending them online to the database systems of environmental agencies have already begun to be tested. Also, the use of drones to estimate female turtles and their nests on a beach.
At the same time, new protocols for the monitoring of turtle populations, with the capture and recording of information about adult animals (and not just the nests and hatchlings), as well as the recording of data on the consumption of these species began to be implemented in protected areas in the Amazon. In addition, it is necessary that new applied ecology researches are carried out to obtain information about the structure and population dynamics of Podocnemidids in the different rivers of the region. This will help to build more robust population models that will help to simulate different scenarios for the conservation of these animals.
In areas with reduced turtle populations and where communities have an interest in trying to recover stocks, we suggest that other conservation tools can be adopted to ensure nest protection and improvement in hatching and hatch survival rates. Simple strategies historically used by beach monitors, such as the transfer of nests from threatened areas (by flooding or predation) to areas protected by the community, or the use of headstarting, to protect the hatchlings more by releasing them in protected natural lakes or keeping them in nurseries before release. Preferably, such strategies should be accompanied by research and used only until the population recovers.
Another important issue is that as local turtle stocks have increased, there has been, as a consequence, increased pressure from groups outside the community to illegally catch and sell these animals, and also increased community complaints about the lack of turtles. surveillance.
Faced with this situation of lack of control by environmental agencies and predatory outflow of resources, communities that protect turtles began to seek, with the environmental authorities, a form of remuneration for the environmental services they provide, or sustainable possibilities of income generation, through community creation of turtles (ex situ management) or quotas for extraction. Then, in 2017, in the state of Amazonas, the first authorized community agriculture units for turtles began operating. But other initiatives such as ecological tourism could be a good future option for these communities.
Conclusion
The riverside communities of the Amazon view the chelonians as an important food source, and it is one which has been managed and conserved by them with participation in different systems of use and protection throughout history.
The community system of protection of chelonians is responsible for most of the conservation areas of chelonians in the Amazonas state, and ensures that a great diversity of nesting environments are conserved in several rivers. In addition, the community management of chelonians has demonstrated that it is productive as a form of co-management, and is effective in increasing and protecting stocks of chelonians in the region, as well as being efficient, since it protects the species at a low operational cost.
Community-based chelonian conservation programs have their voluntary character and community mobilization as their strong point and, as such, have guaranteed the continuity of protection actions, even in the face of possible crises due to lack of support from environmental agencies. As tools for participatory management of natural resources, it is an important conservation strategy, complementary to the protection system created by the government.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Ethics Statement
Ethical approval was not required for our noninvasive study since we did not collect any biological samples nor interfere with the behavior of the study species. Permission to collect observational data from river turtle nesting areas was granted by research permit number IBAMA/SISBIO No.19232/2011.
Author Contributions
The present study was part of the doctoral thesis of PA, who carried out the compilation, tabulation and consolidation of all the data, as well as the statistical analyses, elaboration of graphs and discussion of the results. The other co-authors collaborated in collecting data in the field, tabulating annual data, recording and analyzing specific situations in different areas of chelonian protection, as well as helping to discuss the results obtained.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We would like to thank the Instituto Brasileiro do Meio Ambiente e Recursos Naturais Renováveis (IBAMA) and the Departamento de Mudanças Climáticas e Unidades de Conservação do Amazonas (DEMUC) for their partnership, logistical support and the release of the reports, documents and data used in this study. We also thank the Centro Nacional de Pesquisa e Conservação de Répteis e Anfíbios (RAN), the Centro de Preservação e Pesquisa de Quelônios Aquáticos (CPPQA) and our colleagues José Erickson, at the Instituto Piagaçu, and Marina Seco and Diogo de Lima, at the Instituto Mamirauá, and João Vitor Campos Silva, at the Instituto Juruá, for sharing the data from their programs. We are grateful for the grants and the financial support from the Fundação de Amparo à Pesquisa do Estado do Amazonas (FAPEAM), the administrative support from Fundação de Apoio Institucional Rio Solimões (UNISOL) and sponsorship from the Petrobras Ambiental Program, from the Mineração Rio do Norte and from Instituto Net-Claro-Embratel. We would also like to thank Camila Kurzmann Fagundes for elaborating the maps. We would also further like to thank all the community members who participate in the Pé-de-Pincha program for all the protection and monitoring work they do, especially to our friends Alfredo Luiz Belém Pontes, César Pontes, Francisco Mendes da Silva, Francisco Amâncio, Manoel Costa Neto, Ruth Néia Bentes, Maria José, and Antônio Costa for the data collection, shared information and their time.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.769328/full#supplementary-material
References
Alfinito, J. (1978). Identificação dos principais tabuleiros de tartarugas no rio Amazonas e seus afluentes. Boletim Técnico IBDF 5, 27–84.
Alho, C. J. R., and Pádua, L. F. M. (1982). Sincronia entre o regime de vazante do rio e o comportamento de nidificação da tartaruga da Amazônia Podocnemis expansa (Testudinata:Pelomedusidae). Acta Amazônica 12, 323–326. doi: 10.1590/1809-43921982122323
Andrade, P. C. M. (2008). Criação e Manejo de Quelônios no Amazonas. Projeto Diagnóstico da Criação de Animais Silvestres no Estado do Amazonas. Manaus: IBAMA, 537.
Andrade, P. C. M. (2012). Manejo Comunitário de Quelônios Projeto Pé-de-pincha. Manaus: Gráfica Moderna, 786.
Andrade, P. C. M. (2015). “Manejo Comunitário de Quelônios (Família Podocnemididae - Podocnemis unifilis,” in Médio Rio Amazonas e Juruá, eds P. Sextuberculata, P. Expansa, and P. Erythrocephala (Manaus: Gráfica Moderna), 324.
Andrade, P. C. M. (2017). “Manejo participativo de quelônios por comunidades na Amazônia,” in Olhares Cruzados Sobre as Relações Entre Seres Humanos e Animais Silvestres na Amazonia (Brasil, Guiana Francesa), eds G. Marchand and F. V. Velden (Manaus, BR: EDUA), 163–192.
Andrade, P. C. M., Garcez, J. R., Azevedo, S. H. S., Duarte, J. A. M., Barboza, E. S., Pound, M. C. D., et al. (2016). Projeto Pé-de-pincha: Técnicas de Conservação e Manejo de Quelônios - Manual do Monitor de Praia. Manaus: Gráfica Moderna, 117.
Andrade, P. C. M., Lima, A. C., Canto, S. L. O., Azevedo, S. H., Duarte, J. A. M., Oliveira, P. H. G., et al. (2004). “Pé-de-pincha: Manejo Sustentável de quelônios no baixo Amazonas,” in Ação Comunitária – Uma Outra Face do Ensino Superior Brasileiro Coleção Socializando Experiências, ed. A. I. Calderón (São Paulo, BR: Editora Olho D’água), 78–96.
Antunes, A. P., Fewster, R. M., Venticinque, E. M., Peres, C. A., Levi, T., Rohe, F., et al. (2016). Empty forest or empty rivers? A century of commercial hunting in Amazonia. Sci. Adv. 2:e1600936. doi: 10.1126/sciadv.1600936
Antunes, A. P., Rebêlo, G. H., Pezzuti, J. C. B., Vieira, M. A. R. M., Constantino, P. A. L., Campos-Silva, J. V., et al. (2019). A conspiracy of silence: Subsistence hunting rights in the Brazilian Amazon. Land Use Policy 84, 1–11. doi: 10.1016/j.landusepol.2019.02.045
Barry, D. (1995). A Bayesian model for growth curve analysis. Biometrics 51, 639–655. doi: 10.2307/2532951
Batista, V. S., Isaac, V. J., and Viana, J. P. (2004). “Exploração e manejo dos recursos pesqueiros da Amazônia,” in A pesca e os recursos pesqueiros na Amazônia Brasileira, ed. M. L. Rufino (Manaus, BR: IBAMA), 63–152.
Bence, J. R. (1995). Analysis of short time series: correcting for autocorrelation. Ecology 76, 628–639. doi: 10.2307/1941218
Berkes, F. (2009). Evolution of co-management: Role of knowledge generation, bridging organizations and social learning. J. Environ. Manag. 90, 1692–1702. doi: 10.1016/j.jenvman.2008.12.001
Bowne, R. D., Cosentino, B. J., Anderson, L. J., Bloch, C. P., Cooke, S., Crumrine, P. W., et al. (2018). Effects of urbanization on the population structure of freshwater turtles across the United States. Conserv. Biol. 32, 1150–1161. doi: 10.1111/cobi.13136
Brower, J., Zar, J., and Ende, C. (1989). Field and Laboratory Methods for General Ecology. Dubuque, IA: Wm.C.Brown Publishers, 112–145.
Campos-Silva, J. V., Hawes, J. E., Andrade, P. C. M., and Peres, C. A. (2018). Unintended multispecies co-benefits of an Amazonian community-based conservation programme. Nat. Sustainab. 1, 650–656. doi: 10.1038/s41893-018-0170-5
Campos-Silva, J. V., Hawes, J. E., Freitas, C. T., Andrade, P. C. M., and Peres, C. A. (2020). “Community-Based Management of Amazonian Biodiversity Assets,” in Participatory Biodiversity Conservation - Concepts, Experiences, and Perspectives, ed. C. Baldauf (Switzerland: Springer International Publishing), 99–111. doi: 10.1007/978-3-030-41686-7
Cantarelli, V. H. (2006). Alometria Reprodutiva da Tartaruga-da-Amazônia (Podocnemis Expansa): Bases Biológicas Para o Manejo. Ph.D thesis, Piracicaba, BR: Escola Superior de Agricultura Luiz de Queiroz/USP, 116.
Cantarelli, V. H., Malvasio, A., and Verdade, L. M. (2014). Brazil’s Podocnemis expansa Conservation Program: Retrospective and Future Directions. Chelonian Conserv. Biol. 13, 124–128. doi: 10.2744/CCB-0926.1
Canto, S. L. O., Oliveira, M. S., Rodrigues, E. C. P. G., Duarte, J. A. M., and Andrade, P. C. M. (1999). Consumo de Produtos da Fauna Silvestre no Estado do Amazonas. Abstract Retrieved From Anais do IV Congreso Internacional Sobre Manejo de Fauna Silvestre en Amazonia y Latino America. Asunción: University.Florida, 125.
Carvalho, V. T., Martínez, J. G., Hernández-Rangel, S. M., Astolfi-Filho, S., Vogt, R. C., Farias, I. P., et al. (2016). Giving IDs to turtles: SNP markers for assignment of individuals to lineages of the geographically structured Phrynops geoffroanus (Chelidae: Testudines). Conserv. Genetic Resour. 9, 157–163. doi: 10.1007/s12686-016-0626-8
Charity, S., and Ferreira, J. M. (2020). Wildlife Trafficking in Brazil. Cambridge: TRAFFIC International, 140.
Conway-Gomez, K. (2007). Effects of Human Settlements on Abundance of Podocnemis unifilis and P. expansa Turtles in Northeastern Bolivia. Chel. Conserv. Biol. 6, 199–205. doi: 10.2744/1071-8443(2007)6[199:EOHSOA]2.0.CO;2
Corrêa, H. B. (1978). Contribuição ao estudo dos quelônios amazônicos registrando casos de albinismo observados em Podocnemis expansa e Podocnemis sextuberculata. Boletim Técnico IBDF 5, 3–26.
Costa, D. C., Pereira, H. S., Marchand, G. A. E. L., and Silva, S. C. P. (2018). Challenges of Participatory Community Monitoring of Biodiversity in Protected Areas in Brazilian Amazon. Diversity 10:61. doi: 10.3390/d10030061
Crouse, D. T. (1999). Population modeling and implications for caribbean hawksbill sea turtle management. Chel. Conserv. Biol. 3, 185–188.
Cunha, F. A. G., Fernandes, T., Franco, J., and Vogt, R. C. (2019). Reproductive Biology and Hatchling Morphology of the Amazon Toad-headed Turtle (Mesoclemmys raniceps) (Testudines: Chelidae), with Notes on Species Morphology and Taxonomy of the Mesoclemmys Group. Chel. Conserv. Biol. 18, 2195–2209. doi: 10.2744/CCB-1271.1
DOE (2017). Diário Oficial do Estado do Amazonas - 14/09/2017, Manaus13-17. Available online at: https://imprensaoficial.am.gov.br [accessed on Sep 14, 2017].
Dantas-Filho, J., Pontuschka, R. B., Franck, K. M., and Gasparotto, P. H. G. (2020). Cultivo de quelônios promove conservação e o desenvolvimento social e econômico da Amazônia. Rev. Ciên. Saúde Anim. 2, 9–31.
Diniz, G. L., and Santos, C. I (1997). Crescimento populacional da tartaruga-da-Amazônia (Podocnemis expansa). Biomatemática 7, 128–133.
Ehrhart, L., Redfoot, W., Bagley, D., and Mansfield, K. (2014). Long-term trends in loggerhead (Caretta caretta) nesting and reproductive sucess at an important western Atlantic Rookery. Chel. Conserv. Biol. 13, 173–181. doi: 10.2744/CCB-1100.1
Eisemberg, C. C., Reynolds, S. J., Christian, K. A., and Vogt, R. C. (2017). Diet of Amazon river turtles (Podocnemididae): a review of the effects of body size, phylogeny, season and habitat. Zoology 120, 92–100. doi: 10.1016/j.zool.2016.07.003
Erickson, J., and Baccaro, F. (2016). Nest predation of the yellow-spotted amazon river turtle (Podocnemis unifilis. Troschel, 1848) by the fire ant (Solenopsis geminata, Fabricius, 1804) in the Brazilian Amazon. Herpetol. J. 26, 183–186.
Fáchin-Téran, A., and Vogt, R. C. (2014). Alimentação de Podocnemis sextuberculata (Testudines: Podocnemididae) na reserva Mamirauá. Amazonas Brasil. Rev. Colomb. Cien. Anim. 6, 285–298. doi: 10.24188/recia.v6.n2.2014.431
Fachín-Terán, A., Vogt, R. C., and Thorbjarnarson, J. B. (2004). “Patterns of Use and hunting of turtles in the Mamirauá Sustainable Development Reserve, Amazonas, Brasil,” in People in Nature – Wildlife Conservation em South and Central América, eds K. Silvius, R. E. Bodmer, and J. Fragoso (New York, NY: Columbia University Press), 363–377. doi: 10.7312/silv12782-022
Fagundes, C. K., Fath, F., Cortes, L. G., Uhlig, V., Andrade, P. C. M., Vogt, R. C., et al. (2021). A large scale analysis of threats to the nesting sites of Podocnemis species and the effectiveness of the coverage of these áreas by the Brazilian Action Plan for Amazon Turtle Conservation. J. Nat. Conserv. 61:125997. doi: 10.1016/j.jnc.2021.125997
Fagundes, C. K., Vogt, R. C., de Souza, R. A., and Jr, P. (2018). Vulnerability of turtles to deforestation in the Brazilian Amazon: Indicating priority areas for conservation. Biol. Conserv. 226, 300–310. doi: 10.1016/j.biocon.2018.08.009
Ferrara, C. R., Fagundes, C. K., Morcatty, T. Q., and Vogt, R. C. (2017). Quelônios Amazônicos – Guia de identificação e distribuição. Manaus: World Conservation Society, 182.
Fiori, M. M., and Santos, C. F. M. (2013). Colonizadores portugueses, tartarugas e peixes-boi: uma história da busca por carne, gordura e combustível na Amazônia do século XVIII. Diálogos 17, 1247–1257. doi: 10.4025/dialogos.v17i3.805
Fiori, M. M., and Santos, C. F. M. (2015). A carne, a gordura e os ovos: colonização, caça e pesca na Amazônia, Série História, 63. Porto Alegre: EDIPUCRS, 109.
Fonseca, S. Jr., Marinelli, C. E., Carlos, H. S., Weigland, R., Fernandes, R. B., Campos-Silva, J. V., et al. (2011). Programa de monitoramento da biodiversidade e do uso dos recursos naturais – ProBUC: a experiência das unidades de conservação estaduais do Amazonas. Manaus: Centro Estadual de Unidades de Conservação, 42.
Forero-Medina, G., Ferrara, C. R., Vogt, R. C., Fagundes, C. K., Balestra, R. A. M., Andrade, P. C. M., et al. (2019). On the future of the giant South American river turtle Podocnemis expansa. Oryx 55, 73–80. doi: 10.1017/S0030605318001370
Freitas, C. E. C., Witkoski, A. C., Jacaúna, T. S., and Araújo, L. M. S. (2009). “Manejo de pesca em lagos da Amazônia Central - O lago Grande,” in A pesca na Amazônia Central - Ecologia, conhecimento tradicional e formas de manejo, eds T. J. P. Fraxe, A. C. Witkoski, and S. C. P. Silva (Manaus, BR: Editora da Universidade Federal do Amazonas), 351–378.
Garcez, J. R. (2009). Comercialização e reprodução da tartaruga-da-amazônia (Podocnemis expansa) e do tracajás (P. unifilis) em cativeiro no estado do Amazonas. [monography/graduate]. Manaus(BR): Federal University of Amazonas, 42.
Garcez, J. R., Andrade, P. C. M., and Soares, M. G. M. (2020). Composição da dieta de três espécies de quelônios no rio Juruá, Amazonas. Igapó Rev. Educ. Ciên. Tecnol. IFAM 14, 60–72.
Garcez, J. R., Andrade, P. C. M., Soares, M. G. M., Martins, K. P. M. De Oliveira, A. B., Duarte, J. A. M., et al. (2012). “Capítulo 9: Composição da dieta de Tracajá (Podocnemis unifilis), iaçá (P. sextuberculata) e tartaruga (P. expansa) no Rio Juruá e de tracajá (P. unifilis) no Médio Rio Amazonas” in Manejo Comunitário de Quelônios Projeto Pé-de-pincha, ed. P. C. M. Andrade (Manaus: Gráfica Moderna), 443–462.
Guix, J. C. (2020). Dispersión de quelonios por humanos en el neotrópico: implicaciones genéticas, ecológicas y biogeográficas - Projecte Neopangea. (Barcelona: Universitat de Barcelona). 10, 1–16.
Hailey, A., and Lambert, M. R. K. (2002). Comparative growth patterns in afrotropical giant tortoises (Reptilia, Testudinidae). Trop. Zool. 15, 121–139. doi: 10.1080/03946975.2002.10531170
’Harju, E., Sirén, A. H., and d Salo, M. (2017). Experiences from harvest-driven conservation: Management of Amazonian river turtles as a common-pool resource. New York, NY: Springer, doi: 10.1007/s13280-017-0943-5
Hernández, O., and Espín, R. (2003). Consumo ilegal de tortugas por comunidades locales en el río Orinoco medio, Venezuela. Acta Biol. Venez. 23, 17–26.
Hernández, O., Espinosa-Blanco, A. S., Lugo, M., Jiménez-Oraa, M., and Seijas, A. E. (2010). Artificial incubation of yellow-headed sideneck turtle Podocnemis unifilis eggs to reduce losses to flooding and predation, Cojedes and Manapire Rivers, southern Venezuela. Conserv. Evid. 7, 100–105.
IBAMA (2016). Manejo conservacionista e monitoramento populacional de quelônios amazônicos. Brasília: IBAMA, 136.
IBAMA (2019a). Plano de Ação Nacional para a Conservação dos Quelônios AmazônicosA. Brasília: IBAMA, 192.
IBAMA (2019b). Diagnóstico da Criação Comercial de Animais Silvestres no Brasil. Brasília: IBAMA, 56.
ICMBio (2018). Livro Vermelho da Fauna Brasileira Ameaçada de Extinção: Volume IV – Répteis. Brasília: ICMBio, 255.
IPEA (2010). “Recursos comuns e sustentabilidade ambiental: uma dimensão política, cap.8,” in Sustentabilidade Ambiental no Brasil: diversidade, economia e bem-estar humano, Livro 7, eds A. R. Alvarez and J. A. Mota (Brasília, BR: Instituto de Pesquisa Econômica Aplicada), 249–264.
Jerozolimski, A., Ribeiro, M. B. N., and Martins, M. (2009). Are Tortoises Important Seed Dispersers in Amazonian Forests? Oecologia 161, 517–528. doi: 10.1007/s00442-009-1396-8
Kennet, R., Danielsenn, F., and Silvius, K. M. (2015). Citizen science is not enough on its own. Nature 521:161. doi: 10.1038/521161d
Krebs, C. J. (1986). Ecologia – Análisis experimental de la distribución y abundancia. Madrid: Ed. Pirâmide, 165–215.
Lima, A. C., Silva, C. J., Mateus, W. D., Andrade, P. C. M., and Souza, A. Q. L. (2017). “Stakeholders in community management of turtle in Brazilian Amazon,” in Natural resources in wetlands: from Pantanal to Amazonia, eds M. A. Soares and M. A. G. Jardim (Belém, BR: Museu Paraense Emilio Goeldi/UFPA), 15–36.
Lovich, J. E., Ennen, J. R., Agha, M., and Gibbons, J. W. (2018). Where have all the turtles gone, and why does it matter? Bioscience 68, 771–791. doi: 10.1093/biosci/biy095
May, R. M. (1976). Theoretical Ecology: Principles and Applications. New York, NY: Oxford University Press.
Meza, E., and Ferreira, L. M. (2015). Agencia indígena y colonialismo: una arqueología de contacto sobre la producción de aceite de tortuga en el Orinoco medio, Venezuela (siglos XVIII y XIX). Amaz. Rev. Antropol. 7, 375–402. doi: 10.18542/amazonica.v7i2.3452
Miorando, P. S., Rebêlo, G. H., Pignati, M. T., and Pezzuti, J. C. B. (2013). Effects of community-based management on Amazon river turtles: A case study of Podocnemis sextuberculata in the Lower Amazon floodplain, Pará, Brazil. Chel. Conserv. Biol. 12, 143–150. doi: 10.2744/CCB-1011.1
Morcatty, T. Q., Santos, A., Nijman, V., and Valsecchi, J. (2020). Adapting a traditional hunting technique to improve capture rates for the endangered Yellow-Footed Tortoise (Chelonoidis denticulatus) during ecological surveys in Amazonia. J. Ethnobiol. 40, 252–267. doi: 10.2993/0278-0771-40.2.252
Morcatty, T. Q., and Valsecchi, J. (2015). Social, biological, and environmental drivers of the hunting and trade of the endangered yellow-footed tortoise in the Amazon. Ecol. Soc. 3:20. doi: 10.5751/ES-07701-200303
Nascimento, C. A. R. (2009). Histórico Oficial do Comércio Ilegal de Fauna no Amazonas. Ph.D theisi, Manaus: Universidade Federal do Amazonas, 53.
Nascimento, J. P. B., Meiado, M. V., Nicola, P. A., and Pereira, L. C. M. (2015). Germinação de sementes de Tacinga inamoena (K. Schum.) N.p. Taylor & Stuppy (Cactaceae) após endozoocoria por Chelonoidis carbonaria (Spix, 1824) (Reptilia: Testudinidae). Gaia Sci. 9, 9–14.
Norris, D., and Michalski, F. (2013). Socio-economic and spatial determinants of anthropogenic predation on Yellow-spotted River Turtle, Podocnemis unifilis (Testudines: Pelomedusidae), nests in the Brazilian Amazon: Implications for sustainable conservation and management. Zoologia 30, 482–490. doi: 10.1590/S1984-46702013000500003
Norris, D., Peres, C. A., Michalskia, F., and Gibbs, J. P. (2019). Prospects for freshwater turtle population recovery are catalyzed by pan-Amazonian community-based management. Biol. Conserv. 233, 51–60. doi: 10.1016/j.biocon.2019.02.022
Oliveira, P. H. (2006). Conservação de quelônios aquáticos e proteção de praias de nidificação com envolvimento comunitário na RDS Mamirauá. Tefé: Sociedade Civil Mamirauá, 68.
Oliveira, P. H. G., Castro, I. C., Andrade, P. C. M., Monteiro, M. S., and Gama-Neto, C. V. (2020). Alimentação de filhotes e juvenis de tracajás (Podocnemis unifilis) e tartarugas (Podocnemis expansa) na natureza e em sistemas de criação comunitária no Amazonas. Rev. Agroecoss. 12, 83–98. doi: 10.18542/ragros.v12i1.8956
Oliveira Júnior, J. G. C., Gamarra, N. C., Vieira, F. A. S., Dantas, I. F. V., Barros, E. L. S. F. C., Santos-Silva, M. R., et al. (2020). Four challenges of long-term socio-ecological research in Brazil. Oecol. Australis 24, 291–298. doi: 10.4257/oeco.2020.2402.05
Pantoja-Lima, J., Aride, P. H. R., Oliveira, A. T., Silva, D. F., Pezzuti, J. C. B., and Rebêlo, G. H. (2014). Chain of commercialization of Podocnemis spp. turtles (Testudines: Podocnemididae) in the Purus River, Amazon basin, Brazil: current status and perspectives. J. Ethnobiol. Ethnomed. 10, 1–10. doi: 10.1186/1746-4269-10-8
Penaloza, C. L., Hernandez, O., Espin, R., Crowder, L. B., and Barreto, G. R. (2013). Harvest of endangered sideneck river turtles (Podocnemis spp.) in the Middle Orinoco, Venezuela. Copeia 1, 111–120. doi: 10.1643/CE-10-158
Pereira, A. R., and Arruda, H. V. (1987). Ajuste Prático de Curvas na Pesquisa Biológica. Campinas: Fundação Cargill, 50.
Pezzuti, J. C. B., Castro, F., McGrath, D. G., Miorando, P. S., Barboza, R. S. L., and Romagnoli, F. C. (2018). Commoning in dynamic environments: community-based management of turtle nesting sites on the lower Amazon floodplain. Ecol. Soc. 23:36. doi: 10.5751/ES-10254-230336
Pezzuti, J. C. B., Lima, J. P., Silva, D. F., and Begossi, A. (2010). Uses and taboos of turtles and tortoises along rio Negro, Amazon basin. J. Ethnobiol. 30, 153–168. doi: 10.2993/0278-0771-30.1.153
Pinto, J. R. S., and Pereira, H. S. (2004). “Análise de incentivos institucionais no manejo participativo de fauna silvestre: o caso do projeto “Pé-de-pincha” no noroeste do Pará,” in Agricultura familiar- Pesquisa, formação e desenvolvimento, eds M. N. Angelo-Menezes and D. P. Neves (Belém: Universidade Federal do Pará), 165–184.
Prestes-Carneiro, G. B. (2013). Um cas de subsistance par pêche en Amazonie: le site archéologique de Hatahara (Amazonas, Brésil). Paris: Museum National D’Histoire Naturelle, 145.
Rachmansah, A., Norris, D., and Gibbs, J. P. (2020). Population dynamics and biological feasibility of sustainable harvesting as a conservation strategy for tropical and temperate freshwater turtles. PLoS One 15:e229689. doi: 10.1371/journal.pone.0229689
Rhodin, A. G. J., Stanford, C. B., Van Dijk, P. P., Eisemberg, C., Luiselli, L., Mittermeier, R. A., et al. (2018). Global conservation status of turtles and tortoises (Order Testudines). Chel. Conserv. Biol. 17, 135–161. doi: 10.2744/CCB-1348.1
Rueda-Almonacid, J. V., Carr, J. L., Mittermeier, R. A., Rodríguez-Mahecha, J. V., Mast, R. B., Vogt, R. C., et al. (2007). Las tortugas y los crocodilianos de los países andinos del trópico. Serie Guias Tropicales del campo N. 6. Bogotá: Conservación Internacional, 583.
Salera, G. Jr., Portelinha, T. C. G., and Malvasio, A. (2009). Predation on adult females of Podocnemis expansa Schweigger (Testudines, Podocnemididae) by Panthera onca Linnaeus (Carnivora, Felidae), in Tocantins State. Biota Neotropical 9, 387–391. doi: 10.1590/S1676-06032009000300033
Santos, J. E. (1986). Procedimentos básicos para modelagem ecológica. Rev. Ciên. Cultura 38, 983–992.
Schneider, L., Ferrara, C. R., Vogt, R. C., and Burger, J. (2011). History of turtle exploitation and management techniques to conserve turtles in the Rio Negro Basin of the Brazilian Amazon. Chel. Conserv. Biol. 10, 149–157. doi: 10.2744/CCB-0848.1
Silva, E. S. (2020). Modelagem de crescimento populacional por equações de diferenças: estabilidade local, bifurcação e discussão de um efeito particular de predação. Rev. Matem. Univ. Federal Ouro Preto 1, 51–66.
Silva, O. D., Delatorre, L. G., Rodrigues, L. A. D., and Mistro, D. C. (2009). Dinâmica espaço-temporal de uma população com efeito allee em um ambiente heterogêneo. Aplicada 2, 164–165.
Silva, T. L., Oliveira, M. N., Silva, L. L., Pinho, R. E. O., Lopes-Filho, I. I., Vasconcelos, V. S., et al. (2019). Manejo de quelônios por comunitários da Reserva Extrativista Riozinho da Liberdade – Acre – Brasil. South Am. J. Basic Educ. 6, 864–873.
Smith, N. (1974). Destructive exploitation of the South American river turtle association of Pacific coast geographers. Oregon State Univ. Press 36, 85–102. doi: 10.1353/pcg.1974.0000
Smith, N. (1979). Quelônios aquáticos da Amazônia: um recurso ameaçado. Acta Amazonica 9, 87–97. doi: 10.1590/1809-43921979091087
Soini, P. (1999). Un manual para el manejo de quelônios acuáticos en la Amazonía Peruana. Iquitos: IIAP, 68.
Spencer, R. J. (2002). Growth patterns of two widely distributed freshwater turtles and a comparison of common methods used to estimate age. Austr. J. Zool. 50, 477–490. doi: 10.1071/ZO01066
Stanford, C. B., Iverson, J. B., Anders, G. J., Rhodin, P. P. D., Mittermeier, R. A., Kuchling, G., et al. (2020). Turtles and Tortoises Are in Trouble. Curr. Biol. 30, 721–735. doi: 10.1016/j.cub.2020.04.088
Towsend, W. R. (2008). Experiencia del Pueblo indígena Cofán com la tortuga charapa (Podocnemis unifilis) en el rio Aguacarico, Ecuador. Quito: Fundación Sobrevivencia Cofan, 36.
Vargas-Ramírez, M., Caballero, S., Morales-Betancourte, M. A., Lasso, C. A., Amayad, L., Martínez, J. G., et al. (2020). Genomic analyses reveal two species of the matamata (Testudines: Chelidae: Chelus spp.) and clarify their phylogeography. Mole. Phyl. Evolut. 17:148.
Waldez, F., Gama, A., Adário, L., Marioni, B., Rossoni, F., and Erickson, J. (2013). Monitoramento participativo da caça de quelônios (Podocnemididae) por comunitários ribeirinhos no baixo rio purus e proteção de sítios de desova na RDS Piagaçu-purus, Brasil. Rev. Colomb. Cienc. Anim. 5, 4–23. doi: 10.24188/recia.v5.n1.2013.467
Keywords: freshwater turtles, participatory management, monitoring, population models, Podocnemis
Citation: Andrade PCM, de Oliveira PHG, de Lima AC, da Mota Duarte JA, da Silva Azevedo SH, de Oliveira AB, de Almeida Júnior CD, da Silva EB, Garcez JR, da Silva Pinto JR, da Silva LCN, Monteiro MS, da Silva Rodrigues W, Anízio TLF, Pontes ALB, Teixeira RL, da Silva JM, Duncan WLP and Vogt RC (2022) Community-Based Conservation and Management of Chelonians in the Amazon. Front. Ecol. Evol. 10:769328. doi: 10.3389/fevo.2022.769328
Received: 01 September 2021; Accepted: 02 February 2022;
Published: 22 March 2022.
Edited by:
Jean Boubli, University of Salford, United KingdomReviewed by:
Adriaan Engelbrecht, University of the Western Cape, South AfricaThomas George Bornman, South African Environmental Observation Network (SAEON), South Africa
Copyright © 2022 Andrade, de Oliveira, de Lima, da Mota Duarte, da Silva Azevedo, de Oliveira, de Almeida Júnior, da Silva, Garcez, da Silva Pinto, da Silva, Monteiro, da Silva Rodrigues, Anízio, Pontes, Teixeira, da Silva, Duncan and Vogt. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paulo Cesar Machado Andrade, cGNtYW5kcmFAeWFob28uY29tLmJy
†Deceased