
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 24 February 2022
Sec. Coevolution
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.711732
This article is part of the Research TopicEvolution and Function of Acoustic and Visual Signals in Avian Brood ParasitismView all 17 articles
 Antoine V. Turcotte-van de Rydt1*
Antoine V. Turcotte-van de Rydt1* Christina Petalas1,2
Christina Petalas1,2 Joanna M. Sblendorio3
Joanna M. Sblendorio3 Christopher A. Pearl4
Christopher A. Pearl4 Sharon A. Gill3
Sharon A. Gill3 Mélanie F. Guigueno1
Mélanie F. Guigueno1
Parental care can be costly to parents’ fitness. As such, abandonment of the current reproductive attempt may benefit potential future opportunities, maximising lifetime reproductive success. Obligate brood parasitism, a reproductive strategy in which parasites lay their eggs in the nests of other species and rely solely on them to raise the parasitic young, is an ideal system to study brood abandonment. Some parasitised host species have evolved anti-parasitic defences, notably clutch abandonment (egg burial and nest desertion), that may mitigate negative consequences of parasitism. Abandonment of clutches due to parasitism is not unlike abandonment of reproduction in times of stress, suggesting that host responses to parasitism could be triggered at least partly by elevated stress hormones that mediate individual decisions. Yet, the mechanistic basis for clutch abandonment remains unclear. Here, we experimentally parasitised clutches of yellow warblers (Setophaga petechia), a common host of the brown-headed cowbird (Molothrus ater), with model cowbird eggs to examine whether host circulating corticosterone (CORT) differed among females that accepted parasitic eggs or rejected them through clutch abandonment. We also assessed whether feather CORT, a measure of past corticosterone exposure, differed between accepters and abandoners. Finally, we investigated whether egg visual signals, specifically differences in maculation characteristics between model cowbird and host eggs, predicted abandonment of experimentally parasitised clutches. Circulating CORT was higher in females who abandoned their parasitised clutches, but not in those who accepted, relative to controls with no egg addition. Past stress and differences in maculation characteristics did not predict whether individuals accepted or abandoned experimentally parasitised clutches. Moreover, differences in maculation characteristics between the host and model cowbird eggs did not predict CORT levels or nest abandonment. Thus, parasitism with subsequent clutch abandonment may be associated with elevated circulating CORT, but neither past stress nor differences in maculation characteristics influenced abandonment. The combination of these results contributes to our understanding of the roles of corticosterone and egg visual signals in the context of clutch abandonment in brood parasitism specifically, and of parental care more broadly.
Brood abandonment (Wiggins et al., 1994) occurs in a variety of taxa from insects (Zink, 2003) to birds (Hosoi and Rothstein, 2000). Parental investment theory suggests that a parent should invest in its current brood to the extent that it maximises lifetime reproductive success, based on a trade-off between current and future reproduction (Klug and Bonsall, 2010; Davies et al., 2012). In some instances, it is beneficial for parents to abandon their current reproductive attempt (i.e., brood abandonment) for the benefit of potential future reproductive opportunities (Davies et al., 2012). In birds, cues that can elicit clutch abandonment vary between and within species, and may include predation (Ackerman et al., 2003; Lima, 2009), harsh weather conditions (Bottitta et al., 2003; Thierry et al., 2013), and poor body condition (Groscolas et al., 2008; Spée et al., 2010, 2011). Under these challenging circumstances, brood abandonment can be adaptive if parents are then able to reproduce under more favourable conditions (Sealy, 1995; Guigueno and Sealy, 2010). Although lifetime reproductive success may be maximised through brood abandonment, abandoners may face extensive costs associated with re-nesting and producing another brood later in the breeding season (Wiggins et al., 1994; Verboven and Tinbergen, 2002; Guigueno and Sealy, 2010). As such, brood abandonment represents a key reproductive decision, where abandoners must consider the trade-offs associated with investing in current versus future clutches (Verboven and Tinbergen, 2002; Johnston, 2011; Fokkema et al., 2016; Griesser et al., 2017).
Brood parasitism, in which parasites rely solely on other species to raise their young (Davies, 2000; Schulze-Hagen et al., 2009; Feeney et al., 2014; Soler, 2017), exploits parental care and imposes severe fitness costs on hosts. A parasitised host may incur fitness costs that decrease reproductive success, such as parasitic hatchlings evicting host eggs and nestlings (Soler et al., 1995; Kilner et al., 2004), outcompeting host offspring for food and space (Lorenzana and Sealy, 1997; Moskát et al., 2017), or causing carry-over effects which delay and decrease future nesting attempts in subsequent breeding seasons (Mark and Rubenstein, 2013). To combat these costs, many host species have evolved antiparasitic defences that reduce the impact of parasitism on lifetime reproductive success (Rothstein, 1990; Davies, 2000; Roldán and Soler, 2011; Medina and Langmore, 2015). For example, some hosts recognise the specific threat posed by brood parasites and strongly defend the nest against them during the egg-stage, when nests are most vulnerable to parasitism, employing front-line defences (Gill and Sealy, 2004; Lawson et al., 2021). If the parasite successfully lays an egg in the host nest, some hosts may eject it through grasp- or puncture-ejection to avoid raising the parasitic young while not having to abandon a clutch entirely (Servedio and Hauber, 2006). However, the ability to eject parasitic eggs is limited by the ability to visually recognise a parasitic egg (Soler et al., 2017) and morphological characteristics (Rasmussen et al., 2009; Guigueno and Sealy, 2011; Soler et al., 2015). Birds must have a bill of sufficient gape-size and strength to either grasp the parasitic egg or puncture it (Spaw and Rohwer, 1987; Picman, 1989), or risk incurring significant costs to their reproductive success by damaging their own eggs in the process (Peer et al., 2018). An alternative antiparasitic defence is clutch abandonment.
In clutch abandonment, hosts abandon clutches in response to brood parasitism. The abandoned clutch may consist of only parasitic eggs or it may include a mix of parasitic eggs and their own (Hosoi and Rothstein, 2000; Soler et al., 2015). Hosts abandon their clutches via either nest desertion followed by renesting at a new site or burial of the parasitised clutch followed by a new clutch laid in a superimposed nest (Guigueno and Sealy, 2010). Abandonment is costlier than egg ejection in terms of energy and time invested, as host eggs may be lost and nests need to be rebuilt (Clark and Robertson, 1981; Sealy, 1995; Moskát and Honza, 2002; Guigueno and Sealy, 2009; Soler et al., 2015). Clutch abandonment varies among species, as well as within and between conspecifics (Hosoi and Rothstein, 2000; Guigueno and Sealy, 2010). Only some host species reject parasitised clutches by abandonment, and within these species, individuals may vary in the forms of egg rejection they employ, using, for example, ejection or desertion (Servedio and Hauber, 2006; Honza et al., 2007) or nest desertion and egg burial (Sealy, 1995). The underlying mechanisms that influence within- and between-species variability in responses and the cues that trigger parents to abandon their nests remain unexplained (Abolins-Abols and Hauber, 2018; Avilés, 2018).
Hormones play a crucial role in modulating behaviour (Tata, 2005) and mediate many behavioural adjustments in birds, including antiparasitic responses (Bonier et al., 2009; Ruiz-Raya et al., 2018; Abolins-Abols and Hauber, 2020). Specifically, the glucocorticoid hormone corticosterone (CORT) is a principal mediator of the behaviours of birds as it increases in response to environmental stressors (Siegel, 1980). CORT is often used as an index of stress, although it is only one part of the integrated stress response and is an approximation of whole animal “stress” (MacDougall-Shackleton et al., 2019). Indeed, CORT has a role in a broad variety of functions unrelated to stress, such as regulating metabolism (Ouyang et al., 2013; MacDougall-Shackleton et al., 2019). Nevertheless, elevated CORT is associated with behavioural changes that promote individual survival (Wingfield, 2003; Angelier et al., 2009; Crossin et al., 2012), such as increasing vigilance while decreasing expression of reproduction-related behaviours (Wingfield et al., 1998; Wingfield, 2003; Schoenle et al., 2017). Therefore, CORT represents a promising candidate to explore as a proximate mediator of host responses to brood parasitism (Bókony et al., 2009). Simply the presence of a parasitic egg in the nest may increase the host’s circulating CORT levels (Ibáñez-Álamo et al., 2012; Mark and Rubenstein, 2013; Schoenle et al., 2017) and recent evidence suggests that antiparasitic defences may be partly mediated by CORT. Abolins-Abols and Hauber (2020) experimentally manipulated CORT levels in egg grasp-ejecting American robins (Turdus migratorius) and found that parasitised clutches were more likely to be accepted when potential increases in baseline CORT levels were inhibited. As baseline CORT changes in response to experimental parasitism, it thus can provide further insight into the mechanisms mediating a host’s response, including clutch abandonment (Landys et al., 2006; Ruiz-Raya et al., 2018; Scharf et al., 2021).
Individual allostatic load (one interpretation of “stress”) is related to CORT levels at both short- and long-time scales, with acute (relatively short-term) responses reflected in circulating levels. It is possible that stressful events earlier in the life of an individual influence current behaviours as well. Analyses of CORT in feathers provide a long-term measure of CORT, inferring past stress during feather growth over the moulting period (Bortolotti et al., 2008, 2009; Sherriff et al., 2011). During feather growth when cell differentiation is occurring, circulating CORT continually diffuses in a time-dependent manner through the blood quill into highly vascularised follicles along the entirety of the feather (days-to-weeks, Lattin et al., 2011; Jenni-Eiermann et al., 2015; Freeman and Newman, 2018; Aharon-Rotmann et al., 2021). Feather CORT can thus be used as a biomarker for an integrated measure of CORT secretion representing stress levels over a longer period, rather than to a specific environmental stressor as revealed by measures of circulating CORT from plasma (Fairhurst et al., 2013). Hence, feather CORT, representing general conditions from a different phase of the annual cycle, may be associated with how individuals respond to current reproductive decisions due to cumulative allostatic load over time (Martínez-Padilla et al., 2013; Hansen et al., 2016). This idea is analogous to the developmental stress hypothesis, which predicts that stress early in life can affect behaviour and stress responses later in life, such as a major reproductive decision like abandoning a clutch (Nowicki et al., 2002; Spencer and MacDougall, 2011).
For a CORT response to be initiated, a stressor must appear that causes the hypothalamus to initiate the hypothalamic-pituitary-adrenal cascade leading to CORT secretion. For brood parasite hosts, one such stressor may be the visual recognition of a host egg, as visual differences in colour or maculation between host and parasite eggs often drive host responses (Rothstein, 1974, 1990; Brooke and Davies, 1988; Lyon, 2003; Manna et al., 2017; Gómez et al., 2021). The ability of a parent to successfully recognise their own eggs or a discordant egg within their clutch and reject a parasitic egg or parasitised clutch should be adaptive, as the host would no longer incur the costs associated with brood parasitism (Hauber et al., 2019; Ducay et al., 2021). However, visual signal recognition ability may largely rely on the extent of differences in maculation between parasitic and host eggs (Rothstein, 1982; Lahti, 2006; Cassey et al., 2008; Spottiswoode and Stevens, 2010; Honza and Cherry, 2017; Hanley et al., 2019). Common cuckoo (Cuculus canorus) egg rejection increases with differences in appearance, such as extent of maculation and spot size, between cuckoo and host eggs (Davies, 2000). Appearance-based discrimination also occurs in brown-headed cowbird (Molothrus ater, cowbird hereafter) hosts (Segura et al., 2016; Dainson et al., 2017). Thus, the ability to recognise a parasitic egg may initiate rejection via egg ejection, but the role of differences in egg visual signals in the context of clutch abandonment is less clear (Davies and Brooke, 1989a; Langmore et al., 2005; Spottiswoode and Stevens, 2010). In addition to egg visual signals stimulating rejection, they may also affect circulating CORT levels. Indeed, Eurasian blackbirds (Turdus merula) have higher circulating CORT levels when parasitised with non-mimetic eggs than mimetic ones (Ruiz-Raya et al., 2018). Hence, clutch abandonment may then be mediated by increases in circulating CORT initiated by the extent of maculation characteristic differences between parasitic eggs and their own.
The yellow warbler (Setophaga petechia, hereafter “warbler”) is a common North American passerine that is frequently parasitised by the brown-headed cowbird. They are the only host of the cowbird known to regularly reject parasitised clutches by two forms of clutch abandonment: nest desertion and egg burial (Clark and Robertson, 1981; Sealy, 1995; Mico, 1998; Hauber et al., 2006; Guigueno and Sealy, 2012). About 35% of female yellow warblers bury cowbird eggs, sometimes with their own eggs, and attempt to re-nest, whereas 12% of females desert the nest entirely and renest at a new site (Sealy, 1995). If parasitism occurs early in the laying period, roughly 50% of females bury (Clark and Robertson, 1981). However, warblers rarely desert when parasitised during incubation, possibly due to significant costs related to time lost and building an entirely new nest (Guigueno and Sealy, 2009). In addition, warblers are more likely to bury or desert parasitised clutches early in the breeding season when there is time to re-nest (Guigueno and Sealy, 2010). Due to high costs of clutch abandonment, warblers may rely on multiple stimuli when making their decision, such as interaction with the adult parasite at the nest (Guigueno and Sealy, 2011). Whereas the colours of the egg background (off-white) and maculation (dark brown) are similar between warbler and cowbird eggs (Guigueno and Sealy, 2009; Guigueno et al., 2014), the extent of differences in maculation characteristics (maculation cover and spot profile) between cowbird and warbler eggs have not been quantitatively measured despite obvious qualitative differences noted in the field.
Here, we experimentally parasitised nests of warblers with model cowbird eggs to investigate the relationship between CORT levels (current circulating via plasma and past via feathers), differences in maculation characteristics, and abandonment of parasitised clutches. As circulating CORT can change rapidly in response to environmental stressors (Wingfield, 2003), we predicted that circulating CORT would be higher in females who accepted (1a) model cowbird eggs, and (1b) those who abandoned experimentally parasitised clutches compared to controls. Given that accepters and clutch abandoners both received model cowbird eggs, we expected that individuals who experienced elevated past CORT would have an amplified current CORT in response to the presence of parasitic eggs (Martínez-Padilla et al., 2013; Hansen et al., 2016). Control individuals and individuals who received eggs each come from a random sample, but within the group of individuals who received eggs, abandoners and accepters would presumably split non-randomly if past stress played a role. Therefore, (2a) we reasoned that control individuals would have intermediate feather CORT levels between accepters and clutch abandoners with a greater variance than each of the two experimentally parasitised groups. We also explored two alternatives that (2b) increased feather CORT would be associated with increased circulating CORT and increase the probability of clutch abandonment and that (2c), if an individual experienced past stress and carry-over effects existed, hosts would not be able to pay the costs of abandonment and thus, individuals with higher feather CORT would be more likely to accept. As such, we predicted significant differences in means and variance among all three groups (control, accepters, and abandoners), with the greatest difference between abandoners and accepters. Finally, because differences in egg visual signals between cowbird and host eggs could potentially influence circulating CORT and the probability of abandoning a parasitised clutch (Ruiz-Raya et al., 2018), we assessed the extent of differences in egg maculation characteristics between warbler and model cowbird eggs added to clutches. We predicted that (3a) circulating CORT would be positively associated with the extent of differences in egg maculation characteristics between host and model cowbird eggs and (3b) accepters would have smaller differences in maculation characteristics compared to clutch abandoners. The combination of these analyses provides us with a fuller understanding of the effects of hormones and egg visual signals in the context of clutch abandonment in brood parasitism specifically, and of parental care more broadly.
The study was conducted within a radius of 35 km of Kalamazoo, MI, United States (42°17′24″N, 85°35′09″W), between May and June in 2010, 2011, and 2019. Nest sites were situated in habitats consisting of marshes and shrubs where yellow warblers were abundant and cowbirds were observed daily.
We found nests by observing singing males and females carrying nest-building materials. We sought to find nests as early in the breeding season as possible, ideally in the preliminary nest building stages, to ensure proper timing of experimental parasitism. We visited nests during the building stage every 2 days until nest-lining was complete. We then monitored the nests daily to identify the day of the first egg laid. Eggs were numbered in their laying order with a non-toxic permanent marker. Because natural parasitism by cowbirds is too infrequent at our field sites to provide a sufficient sample size, we made puncturable artificial eggs out of plaster of Paris, closely mimicking cowbird eggs in shape, colouration, weight and volume (see Guigueno et al., 2014). Two model cowbird eggs were added to each experimental nest, which does occur naturally (Kuehn, 2009), to increase the likelihood the females detected the clutch manipulation. On laying day 1, we added two cowbird eggs to the warbler’s clutch between 6:00 and 12:00 (Eastern Daylight Time), to limit the gap to the cowbirds’ egg laying window. The nests were monitored daily over the next 6 days to observe laying progression and to record acceptance or clutch abandonment via nest desertion or burial. Egg burial was recognised by the slight pushing down of the eggs and/or the addition of lining material over the eggs, while desertion was recognised when the female was not observed at the nest for 3 days in a row and eggs were cold (Guigueno and Sealy, 2010). One out of every three nests found were randomly chosen to act as controls and did not receive model cowbird eggs. Control nests were monitored at the same frequency as the experimental group.
Female warblers abandon on 2–3 days after parasitism, with burials occurring after 2.3 ± 0.1 days and nest desertion after 2.5 ± 0.3 days (mean ± SE, Guigueno and Sealy, 2010). Therefore, we collected a blood sample on laying day 4 to capture a snapshot of the female’s circulating CORT during this critical period. We captured females between 8:00 and 14:00 (Eastern Daylight Time). This time window was chosen to minimise time of day differences among individuals, while giving time to capture hard-to-capture birds and to sample multiple females on the same day. We sampled a small number of females after 12:00 pm to ensure that we collected blood samples on the appropriate day in the female’s laying cycle. On laying day 4, mist nets were placed near the focal host nest to passively capture the female. We then aimed to collect a blood sample from the alar vein within 3 min of capture, recording the time from capture to sample acquirement. In nine cases, sampling time exceeded 3 min, which would increase CORT due to acute stress of capture (Romero and Reed, 2005); therefore, we ran a correlation analysis between capture time and CORT level of all individuals and estimated at 3 min the CORT level for those females whose sampling time was prolonged. Each female was banded with a numbered United States Fish and Wildlife Service aluminium band and a unique combination of colour bands before being released to ensure we did not resample females within or between years. Blood samples were kept on ice until they were centrifuged to separate plasma from red blood cells. The separated plasma was pipetted into a separate labelled vial and frozen in a −20°C freezer for long-term storage.
Plasma corticosterone was determined using an enzyme-linked immunosorbent assay (ELISA) kit from ENZO Life Sciences, Inc. (Farmingdale, NY, United States), following kit instructions. This ELISA has been previously validated and used to measure plasma corticosterone in multiple songbirds including sparrows (Wada et al., 2007; Ouyang et al., 2021), wrens (Strange et al., 2016), finches (Kraft et al., 2021) and swallows (Sarpong et al., 2019). Plasma samples were diluted to fall within the range of the standard curve, generally 1:40 or 1:50, incubated with 1% steroid displacement reagent, and run in triplicate or duplicate depending on the sample volume. All samples from a single year were run on the same plate; the intra- and inter-assay coefficient of variation were less than 10%. Assay sensitivity was calculated to be 20.34 pg/ml and all samples read above that value.
During capture of the female warbler on the fourth laying day, the outermost rectrices (R6) on both sides of the tail were collected. We included both second-year and after second-year individuals within our study. Second-year individuals grew feathers during their juvenal moult in the nest, while after second-year birds grew rectrices feathers as part of their complete moult on the breeding grounds between June and September (Pyle et al., 1997). As such, birds within both age-classes grew sampled feathers within the previous breeding season, but, in non-overlapping timeframes. As such, we tested for differences between second-year and after second-year individuals using a linear model on R Statistical Package (R Core Team, 2020), with age as a predictor and feather CORT as the dependent variable. As there were no significant differences between age and feather CORT (p = 0.61), we merged the two groups for subsequent analyses.
Extraction of corticosterone from feathers was conducted following a procedure similar to that previously described in Bortolotti et al. (2008). The calamus was removed and the length and weight of the feather were recorded using a digital caliper and an analytical balance, respectively. The feather (vane and rachis) was cut and minced into small pieces and put into a 50 ml conical tube with 10 ml of methanol. The tubes were sonicated for 30 min in a sonicating bath and then incubated overnight at 50°C in a shaking water bath. The next morning, the methanol was filtered through a 70 μm cell strainer and further filtered through a 0.22 μm cellulose acetate syringe filter into a glass tube. Methanol was evaporated by placing tubes into a 50°C shaking water bath in a fume hood. Once evaporated, samples were reconstituted with 1 ml ELISA assay buffer with gentle rocking for 3 h. Samples were transferred to microcentrifuge tubes and frozen at −20°C until assayed no more than 7 days later.
Feather corticosterone was determined using an ELISA from ENZO Life-Sciences Inc., following the kit instructions with the extension of the standard curve by one value on the low end (total range: 20,000 – 6.4 pg/ml). All samples from a single year/season were run on the same plate in triplicate without dilution. Assay sensitivity was 6.4 pg/ml and all samples read well above this value. Intra- and inter-assay coefficients of variation were less than 10%. Corticosterone values were normalised to feather length.
On laying day 4, we collected the last laid egg. Some nests were inaccessible because of height or heavy vegetation and as a result, it was only possible to determine clutch size but not possible to either mark or sample the eggs, resulting in smaller sample sizes for eggs when compared to circulating CORT samples collected from females (Table 1).
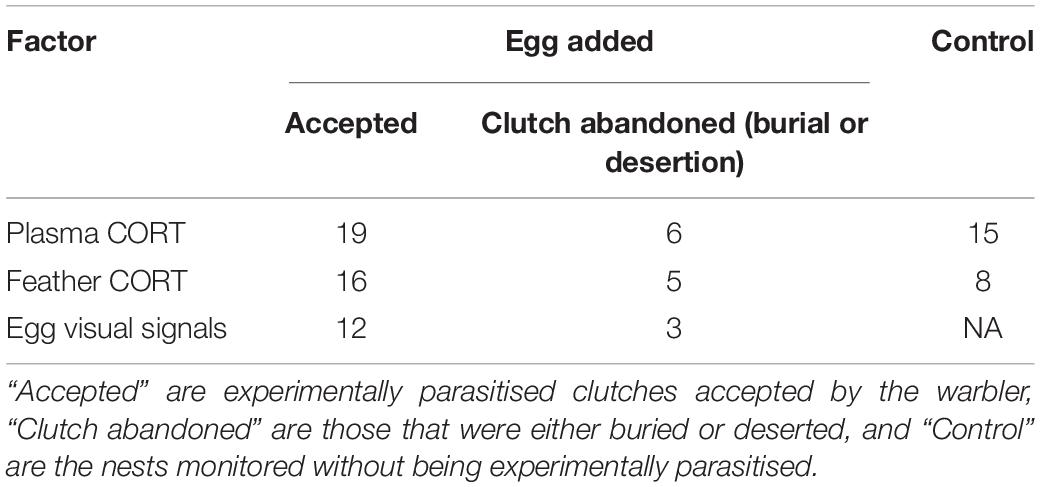
Table 1. Summary of CORT and egg visual signals measurements organised by treatment.
We followed a similar egg image analysis procedure as Hauber et al. (2018). The fourth laid warbler eggs were each compared to one standardised model cowbird egg. Eggs were placed on their side on a colour standard card with a ruler and were photographed with a digital camera mounted on a tripod. The eggs were each photographed four times, with a 90° rotation along the axis between photos. Image analysis on warbler and model cowbird eggs was done using Adobe Photoshop®. We first cropped each egg image into three sections along its long axis: (1) apex-end third, (2) middle-region third, and (3) blunt-end third. We quantified maculation coverage percentage within each third by converting the maculated area to black (RGB = 0,0,0) and the non-maculated area to white (RGB = 255,255,255). We then measured the percentage of black pixels within each third. We obtained the maculation coverage percentage for each third by finding the mean of the percentages between the four images for each egg. We calculated the absolute difference in maculation coverage percentage between the blunt and apex ends for all eggs, thus producing a measure of end-to-end maculation coverage variation. The model cowbird egg’s end-to-end maculation coverage value was then subtracted from the warbler egg’s maculation coverage, providing a measure of the end-to-end difference in maculation coverage between these two eggs. As such, a positive value signified that the host egg had greater variation than the cowbird model egg and a negative value would indicate that there was less variation.
We obtained an average spot length-to-width ratio of the apex and blunt thirds by randomly selecting five spots within each third of each egg image. The mean spot ratio within each of the apex and blunt thirds of the egg was then averaged across the four images for each egg, which we termed average spot profile. We then calculated the absolute difference in spot profile between the blunt and apex ends for all eggs, to have a consistent measure of within-egg end-to-end variation. We used the absolute spot profile difference for both the cowbird model egg and host egg. Finally, we subtracted the model cowbird egg’s absolute difference in within-egg spot profile variation from that of the host egg, producing either a positive or negative value. As such, positive value signified that the host egg had greater spot profile variation than the cowbird model egg and negative values indicated there was less variation.
All statistical analyses were conducted on R Studio (2021) with R Core Team (2020). Data were categorised into three treatment groups; controls, accepters, and abandoners, as detailed above. All graphs were constructed using “ggplot2” (Wickham, 2016) and “ggsignif” (Ahlmann-Eltze and Patil, 2019) packages for R. All data are presented as mean ± SE.
We took a two-analysis approach to explore the effects of experimental parasitism, testing whether egg addition was associated with elevated CORT and then testing whether egg addition and abandonment combined were associated with elevated CORT compared to controls. We reasoned that combining accepters and abandoners in a single experimental group would result in an inflated variance. Similarly, the statistical power for the comparison between accepters and abandoners would be low, and therefore we would expect lower ability to detect a difference if it existed. Before analysis, circulating CORT data were normalised using a log-transformation. There was one unusually small outlier that we corrected to one, which was still the lowest data point, to avoid deformations in the log-transformation. We had a priori directional predictions for (1a) and (1b), expecting higher CORT levels in experimental treatments compared to controls. As such, we used one-tailed Student’s t-test (using the “t.test” function) to detect differences between groups. To test the prediction that egg addition alone would result in higher CORT in accepters than in controls (1a), we excluded the outcome of clutch abandonment. To test the prediction that circulating CORT was associated with clutch abandonment (1b), abandoners were compared to controls. Because variance was similar between accepters and controls, but about half for abandoners versus controls, we ran t-tests with homogenous versus heterogeneous variances for predictions (1a) and (1b), respectively.
Length-normalised feather CORT data was subsequently log-transformed to meet the assumption of normality. Levene’s test was used to test for variance between the treatment groups using “leveneTest” function of the “car” package (Fox and Weisberg, 2019). To investigate the relationship between accepted, abandoned and controls clutches we used a generalised linear model using the “glm” function. If significant, we planned to do post hoc tests between all three groups to test predictions 2a and 2b.
For egg visual signals, analyses were conducted using only eggs from experimentally parasitised nests, where the warblers would have been confronted with the sight of the model cowbird eggs. We ran two linear models using the “lm” function: one associating corrected log-CORT with difference in within-egg maculation coverage variation, and a second pairing corrected log-CORT and within-egg spot profile variation between the host egg and the model cowbird eggs. Additionally, we used one-tailed Student’s two-sample t-tests (“t.test” function in R) to determine whether maculation coverage and spot profile variation were greater among warblers that abandoned their experimentally parasitised clutch relative to those that accepted.
Our sample consisted of a total of 40 yellow warbler nests, with 25 experimentally parasitised and 15 control nests (Table 1). Within the experimentally parasitised group, 19 females accepted the experimental parasitism (76%), and 6 abandoned their parasitised clutch (24%) through either desertion (n = 4) or burial (n = 2).
A total of 40 circulating CORT samples were collected (Table 1). Circulating CORT levels of accepters were not different than controls (t = 0.45, df = 33, p = 0.33; Figure 1A). In contrast, circulating CORT levels of abandoners were significantly higher than controls (t = 1.96, df = 15, p = 0.03; Figure 1B).
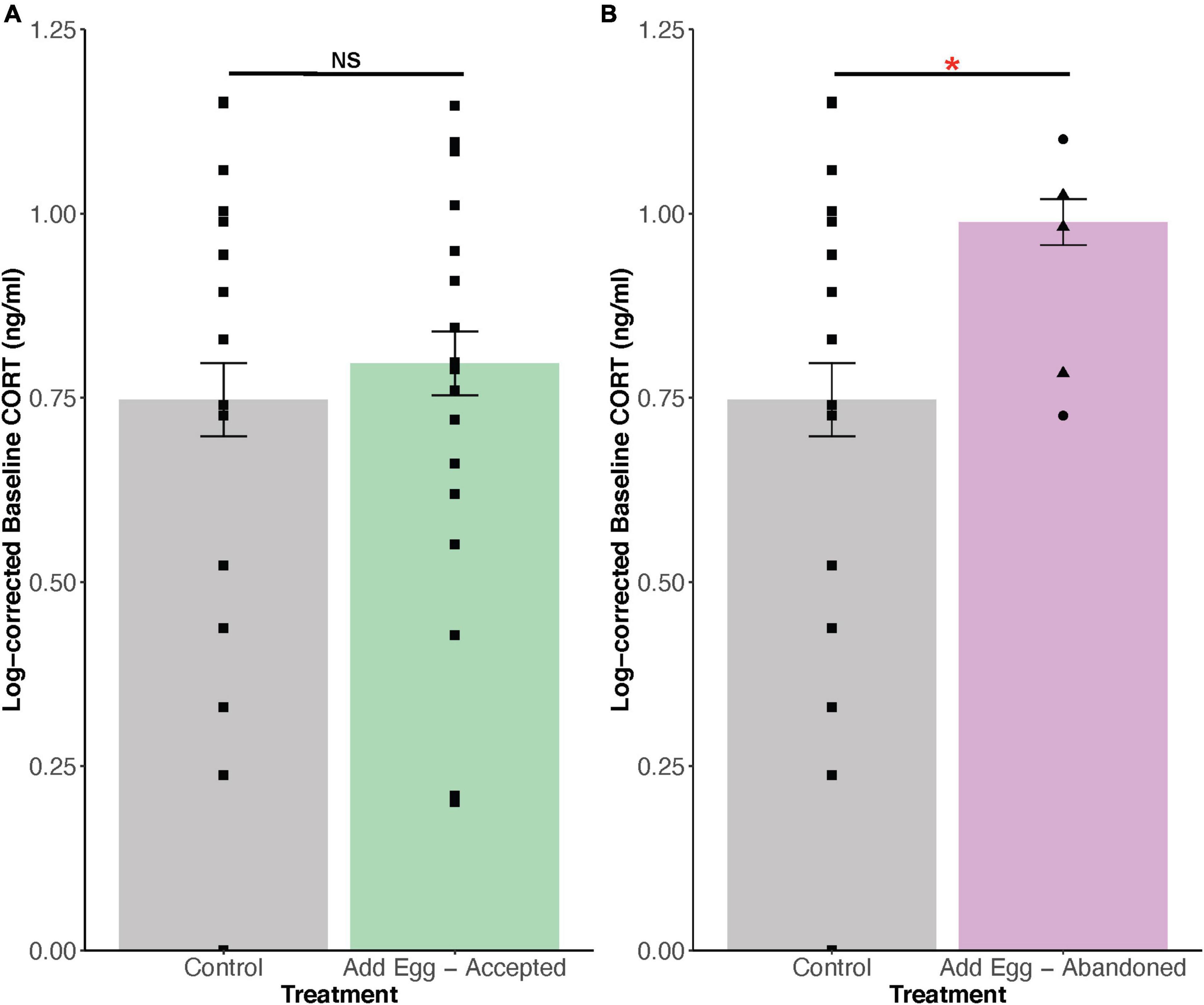
Figure 1. (A,B) Comparisons in log-corrected, circulating corticosterone (CORT) levels in plasma of controls (no model eggs added to clutches) and two experimental treatments (model eggs added). Among individuals with experimentally manipulated clutches that abandoned, abandoners by desertion are indicated with triangles, while burials are indicated with circles. (A) Individuals who accepted the cowbird model eggs did not have significantly higher circulating CORT than controls (t = 0.45, df = 33, p = 0.33). (B) In contrast, individuals who abandoned their experimentally parasitised clutches has higher circulating CORT than controls (t = 1.96, df = 14.77, p = 0.03). NS and * indicate “not significant” and p < 0.05, respectively. Mean ± SE.
Feather samples from 29 individuals were collected (Table 1). Variances were similar across treatments (F2,26 = 0.43, p = 0.65; Figure 2). Mean feather CORT concentrations were similar among treatment groups (F2,26 = 0.11, p = 0.89; Figure 2); as such, post hoc tests were not conducted.
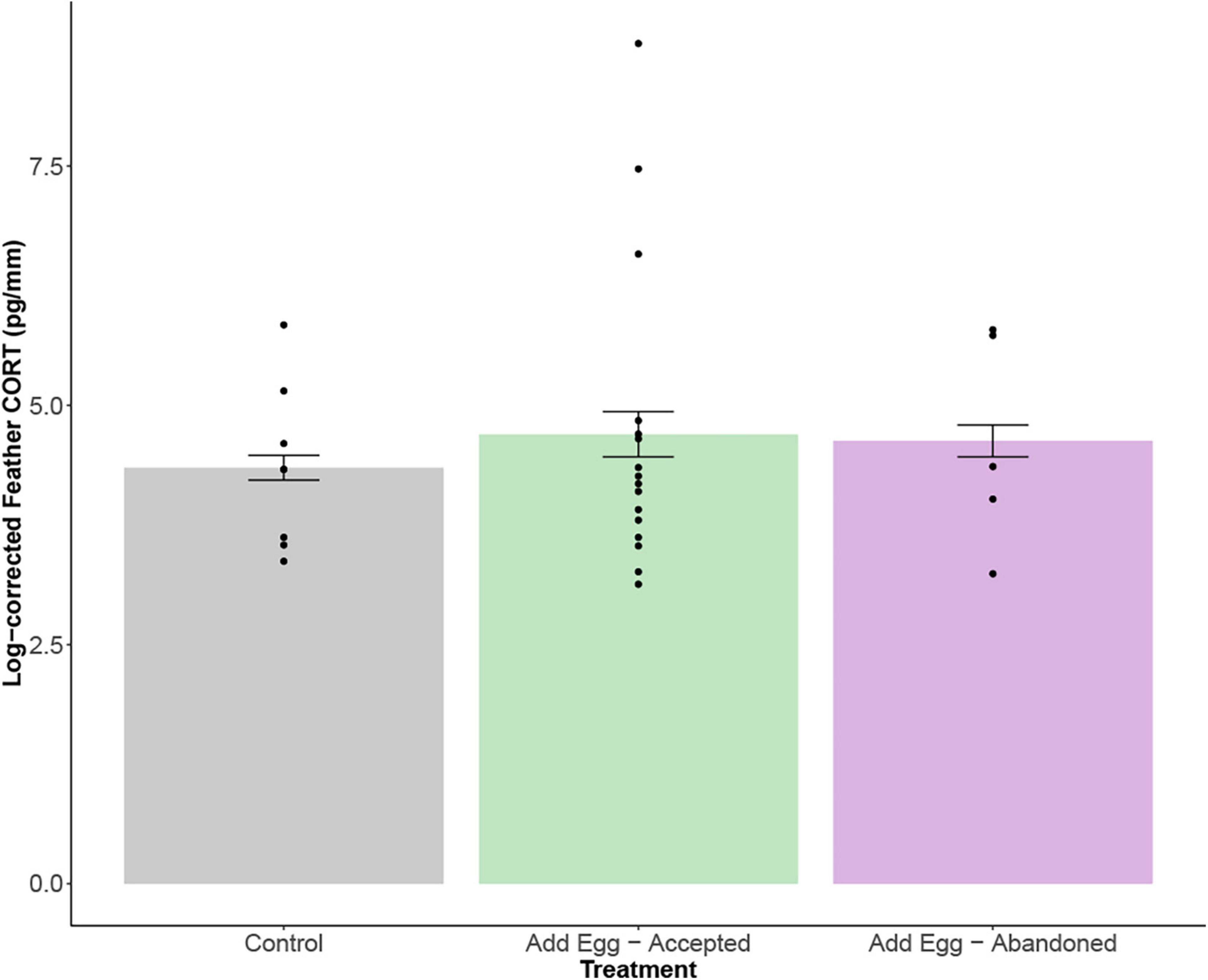
Figure 2. Log-corrected feather corticosterone (CORT) levels (pg/ml) in feathers of females across the three experimental groups. There was no significant difference in the variance (p = 0.65) or the mean (p = 0.89) between groups. A. F2,26 = 0.43, p = 0.65; B. F2,26 = 0.11, p = 0.89.
In total, 22 warbler eggs were analysed (Table 1). The model cowbird egg had an end-to-end maculation coverage variation of 36.6% and an end-to-end spot profile variation ratio of 0.04. Contrastingly, the warbler eggs had an average end-to-end maculation coverage variation of 33.3 ± 2.32% and an average end-to-end spot profile variation ratio of 0.16 ± 0.07.
Circulating CORT levels were not associated with differences in within-egg maculation coverage variation between cowbird model and host eggs (R2 = −0.05, p = 0.44; Figure 3A), nor were they associated with spot profile (R2 = 0.09, p = 0.27; Figure 3B). Separating the correlation between treatments (accepted versus abandoned) did not change the results. Further, differences in maculation coverage (df = 13, t = −0.43, p = 0.66) and spot profile (df = 13, t = −0.01, p = 0.50) variation between eggs were not greater in abandoners compared to accepters (Figures 4A,B).
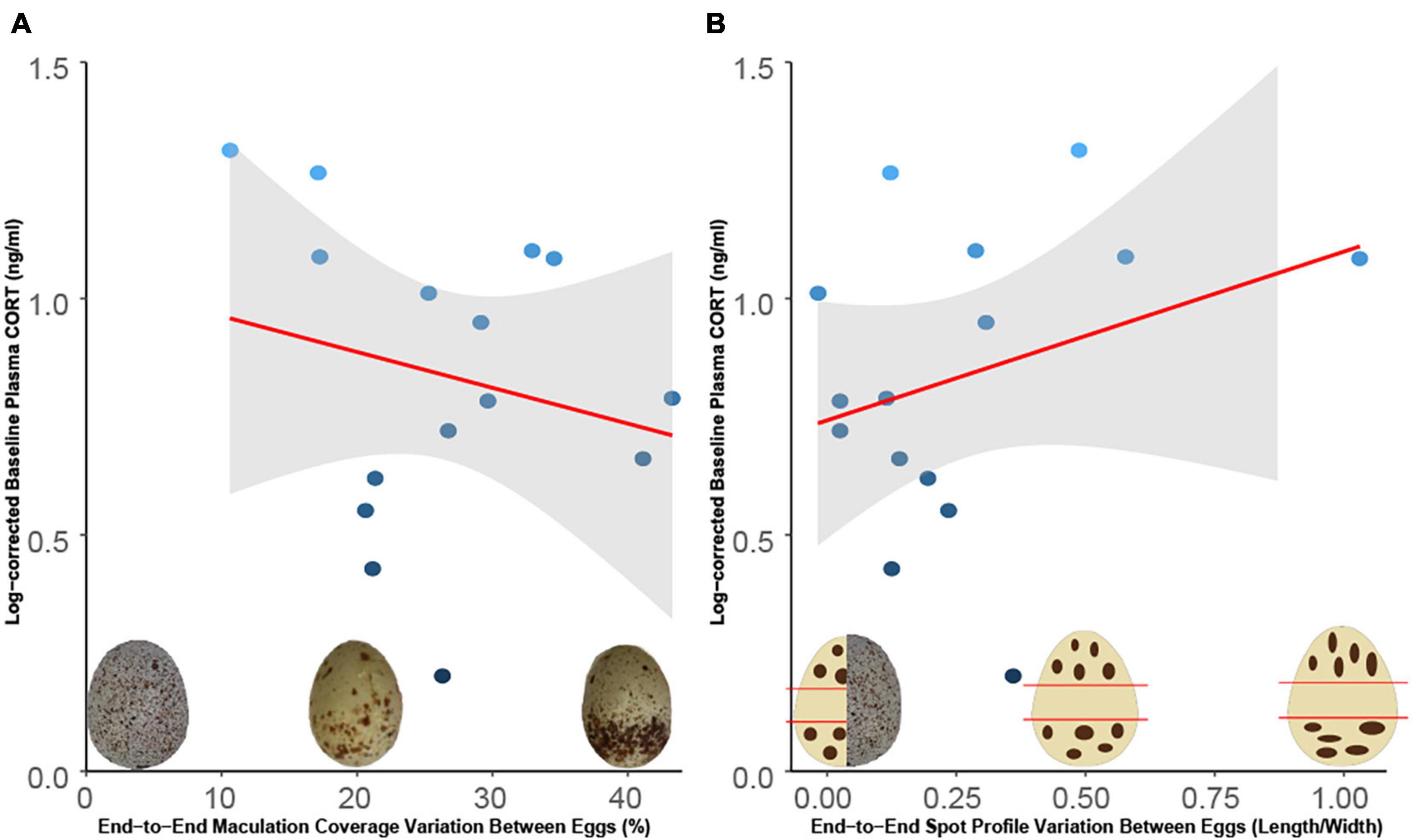
Figure 3. (A,B) Correlation between log-corrected circulating CORT levels from individual female warblers and increasing variation in (A) maculation coverage (p = 0.44) and (B) spot profile (p = 0.27) of their eggs relative to the model cowbird egg. The shaded area indicates 95% confidence interval of the mean slope. A. R2 = −0.05, B. R2 = 0.09
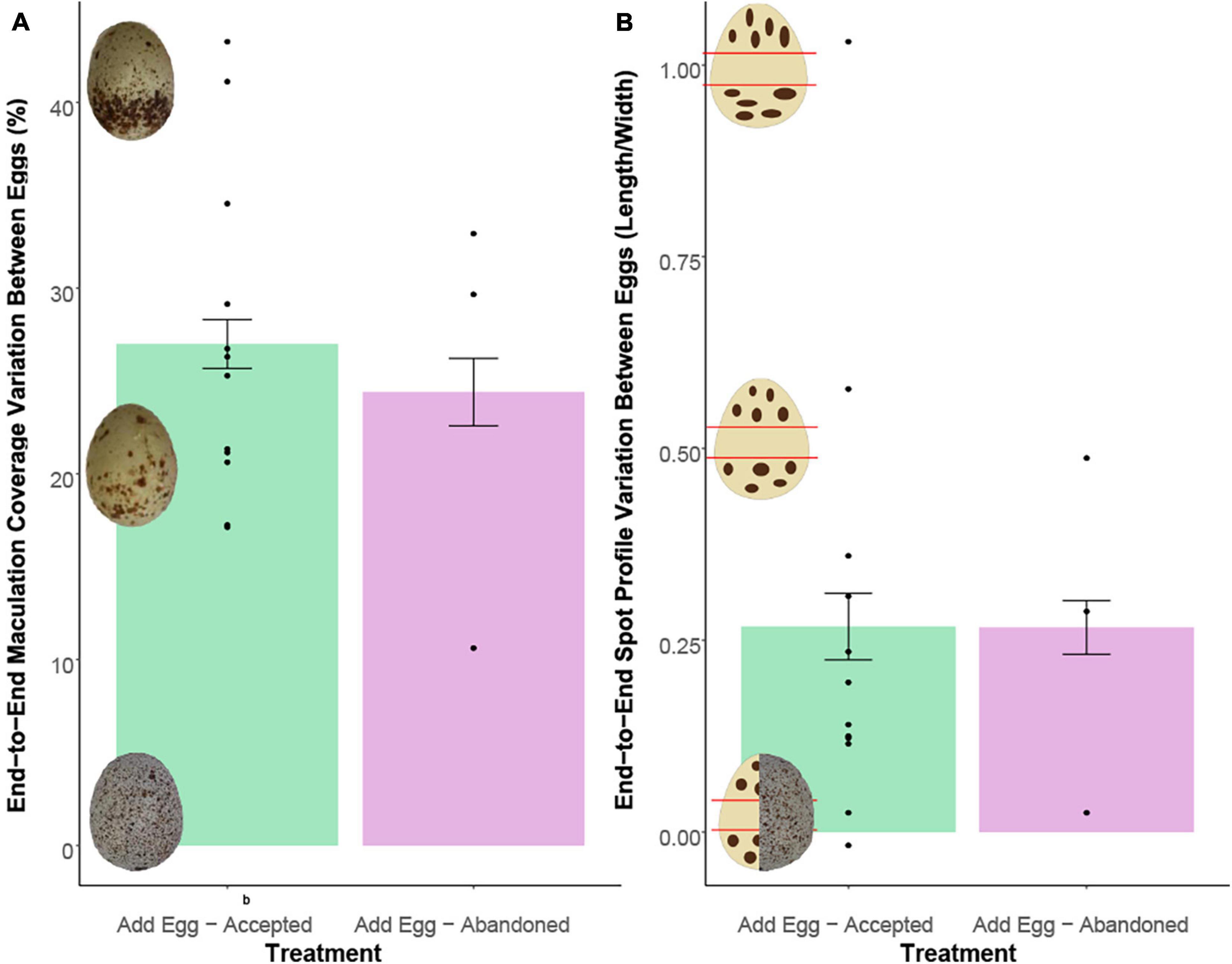
Figure 4. (A,B) Females who abandoned their experimentally parasitised clutch did not have eggs that varied more in (A) maculation coverage (p = 0.44) or (B) spot profile (p = 0.27) relative to the model cowbird egg.
In this study, we examined mechanisms associated with clutch abandonment in a frequently parasitised host of the brown-headed cowbird, the yellow warbler, which regularly rejects naturally parasitised clutches via egg burial and nest desertion two forms of clutch abandonment (Sealy, 1995). Circulating CORT levels were higher in females who abandoned their parasitised clutches, either by egg burial or nest desertion, relative to females at control nests in which no model cowbird eggs were added (Figure 1B). Simply adding model cowbird eggs to clutches did not increase circulating CORT, as females that accepted cowbird eggs did not have elevated CORT relative to controls (Figure 1A). In addition, past stress, as approximated by feather CORT, did not predict whether females accepted or abandoned experimentally parasitised clutches (Figure 2). Finally, differences in egg visual signals, as measured by differences in maculation coverage and spot profile variation between the model cowbird egg and host eggs, did not predict abandonment nor were they correlated with circulating CORT levels of females who abandoned their nests (Figures 3, 4).
Several explanations for the increased CORT in abandoners relative to controls are possible. First, females with naturally higher circulating CORT may be more likely to abandon their nests, such that the presence of a parasitic egg might not alter host CORT levels. That is, individuals with high CORT are more predisposed to abandon for any reason, and the presence of a parasitic egg may tip them past the threshold of abandonment. Second, the presence of a parasitic egg might increase CORT, but only in some females, which then abandon the nest. Third, a parasitic egg might increase CORT in all females, but females whose CORT levels exceed a certain threshold may be more likely to abandon the nest than those whose CORT remains below the threshold. Fully resolving these differences would require a study design in which blood samples were collected before and after parasitism, preferably coupled with experimental manipulations of CORT via injections or implants. These approaches require capturing females multiple times during laying, which may substantially increase abandonment, thereby overwhelming any experimental effect due to experimental parasitism. Our data provides information to target appropriate hormone concentrations for future implant studies for testing these ideas.
Circulating glucocorticoids, such as CORT, vary in response to environmental stressors (Siegel, 1980; Wingfield, 2003). In our study, females who abandoned their parasitised clutch had higher circulating CORT, relative to females with control clutches that were not parasitised (Figure 1B). However, circulating CORT did not differ between control and accepter females. Females were sampled 3 days (laying day 4) after the clutch manipulation (laying day 1), suggesting that abandoners in the population may have had a more pronounced endocrine response than accepters (Figure 1A). Given that clutch abandonment is a generalised response, similar increases in circulating CORT occur in association with other abandonment-triggering factors, such as inclement weather (Romero et al., 2000; Thierry et al., 2013). Therefore, circulating CORT could provide us with a broader understanding of how environmental stressors, whether biotic or abiotic, affect nest abandonment in birds.
Brood parasitism per se does not appear to affect circulating CORT levels in female yellow warblers. Our results show that simply adding model cowbird eggs, without the host abandoning the clutch, was not associated with higher circulating CORT relative to control females. The absence of a difference between controls and accepters, contrary to our prediction, suggests that females who did not abandon experienced a comparable level of stress compared to unparasitised (control) individuals, a finding that differs from previous research. For instance, a common cuckoo host, the Eurasian blackbird, experienced increased circulating CORT following experimental parasitism (Ruiz-Raya et al., 2018). The difference in findings could depend on several factors including parasitism rates, predation rates, breeding season length, and other life history traits (Bókony et al., 2009). The fact that our study focused on a species with a variable abandonment response to parasitism may explain the different pattern of circulating CORT variation that we observed compared to species with more consistent responses such as Eurasian blackbirds (Ruiz-Raya et al., 2018). Our results are in line with the findings of Abolins-Abols and Hauber (2020), who found a causal link between stress-induced CORT and egg ejection rate in American robins, an egg ejecter species. Together, these findings suggest circulating CORT may mediate antiparasitic responses across different parasite-host systems and across rejection methods (Ibáñez-Álamo et al., 2012; Mark and Rubenstein, 2013; Schoenle et al., 2017).
Feather CORT, in contrast to circulating plasma CORT, indicates a long-term past “stress” history (Bortolotti et al., 2008), integrating levels of circulating CORT across the moult period. Our predictions (2a and 2b) that abandoners and accepters would have higher and lower past CORT than controls, respectively, focused on whether an individual’s prior state could predict a response to the future stressor of experimental parasitism. As such, these predictions resemble those of the developmental stress hypothesis, which proposes that stress early in life (e.g., nestling nutritional state) can affect behaviour and cognition later in life (e.g., adult song quality). Although this hypothesis focused primarily on song quality and nutrition (Nowicki et al., 2002), it was later applied to broader contexts including other sexually selected traits (Spencer and MacDougall, 2011). We proposed a similar reasoning to explain why abandoners and accepters co-exist in some host populations, including yellow warblers (Sealy, 1995; prediction 2a). However, feather CORT was not associated with the host’s response to the model cowbird eggs (Figure 2). In addition, variance in feather CORT among the three treatment groups (control, accepted, abandoned) was similar (Figure 2). These results contrast with our predictions that control individuals, who did not experience the stress of experimental parasitism, would display more variable stress histories and that experimental females would respond to parasitism according to their past stress histories (Bortolotti et al., 2008), thereby splitting the experimental parasitism groups into accepters and abandoners. Thus, feather CORT levels of female warblers did not predict responses to parasitism. This lack of an effect could occur if stress during moult was not high enough to induce carry-over effects (O’Connor et al., 2014). Overall, these results suggest that past stress, as measured through feather CORT, does not predict abandonment of parasitised clutches in yellow warblers within this study. Nonetheless, feather CORT only provides a proxy of CORT levels near the time of moult, and warblers may be responding to other time windows, such that an additional study is warranted.
The role of visual pattern differences in egg recognition has been widely established in other species, primarily in ejecters (Cassey et al., 2008; Spottiswoode and Stevens, 2010; Honza and Cherry, 2017; Hanley et al., 2019). In response to increased egg recognition by hosts, some specialist brood parasites, such as common cuckoos, have evolved eggs that mimic the colour and maculation of their host (Davies and Brooke, 1989a,b; Kilner, 2006; Kilner and Langmore, 2011; Spottiswoode and Stevens, 2011; Thorogood et al., 2019). However, in generalist brood parasites such as brown-headed cowbirds, egg visual signals differ from hosts to a greater extent compared to specialist parasites, and thus, the signals may be more noticeable and reduce the opportunity for the co-evolution of egg mimicry (Davies, 2000; Krüger, 2007). For instance, many cowbird hosts lay eggs that differ to varying degrees in colour and/or maculation from cowbird eggs, like those of yellow warblers (Abernathy and Peer, 2014). Warblers in our study had greater intra-egg maculation coverage and spot profile variation than the model cowbird egg (Figures 3, 4), and females could have potentially used this egg visual signal as a cue to abandon their parasitised clutch. Indeed, yellow warblers can determine their clutch has been manipulated using both tactile and visual cues (Guigueno and Sealy, 2012), but, prior to this study, the extent of differences in maculation were not quantitatively analysed.
Visual recognition of parasitism via differences in maculation could modulate CORT changes. As such, we examined whether differences in maculation and spot profile variation between warbler eggs and our model cowbird eggs correlated with circulating CORT (prediction 3a, Figure 3). Increased differences in visual signals could be associated with increases in circulating CORT and a decrease in parental care (Angelier et al., 2009). However, circulating CORT was not correlated with differences in maculation characteristics (Figure 3). Perhaps differences between cowbird and yellow warbler egg maculation were simply not large enough to stimulate a response. Indeed, some yellow warbler eggs look nearly identical to brown-headed cowbird eggs in maculation characteristics, and anti-parasitic behaviours in yellow warblers may be mediated primarily through tactile, rather than visual, cues (Guigueno and Sealy, 2009, 2012; Guigueno et al., 2014). Yet another possibility is that individuals that rejected experimental parasitism had higher baseline CORT, although we might still expect that abandonment would be associated with an interaction between CORT levels and visual signals.
Differences in egg maculation characteristics could be an important cue in eliciting egg rejection in brood-parasitic hosts, as has been shown in ejecters, but the same may hold true for clutch abandoners. Thus, we also examined whether differences in maculation characteristic variation predicted whether hosts accepted or abandoned their experimentally parasitised clutches (prediction 3b, Figure 4). However, abandoners did not show greater variation in maculation coverage or spot profile between their eggs and the model cowbird eggs, relative to accepters (Figure 4). Although yellow warblers from a previous study gazed longer at manipulated clutches, this increased gaze time only occurred between eggs that differed in both background colour and maculation (i.e., blue vs. off-white; brown maculation vs. none; Guigueno and Sealy, 2012). In addition, tactile cues, as measured by the warblers’ egg probing and body shifting during incubation, had a stronger effect than visual cues (Guigueno and Sealy, 2012). To abandon a clutch, females simply need to recognise they have been parasitised and use other cues such as tactile recognition of volumetric differences between the larger parasitic egg and the smaller host eggs (Guigueno and Sealy, 2012) and interaction with the adult parasite (Guigueno and Sealy, 2011). In sum, yellow warblers, as with other hosts that use clutch abandonment as an antiparasitic defence, seem to rely less on visual signals than egg-ejecting hosts to distinguish between their own eggs and the parasitic egg.
We explored the effects of current (plasma) CORT, past (feathers) CORT, and differences in egg maculation variation in the context of clutch abandonment in parasitised yellow warblers. We found evidence that current stress, as approximated by circulating CORT in plasma, was associated with clutch abandonment, but we did not find that maculation or past stress played a role. To further explore the action of CORT, future studies could experimentally increase CORT to determine whether this manipulation elicits abandonment by female warblers. This approach would be opposite of that of Abolins-Abols and Hauber (2020)’s, in which authors decreased ejection in an American robin (Turdus migratorius) by experimentally decreasing CORT synthesis with mitotane injections. Robins normally eject at a high rate (i.e., ∼ 90%; Croston and Hauber, 2014), so decreasing CORT would result in more individuals accepting cowbird eggs. In contrast, yellow warblers normally abandon 20–30% of experimentally parasitised clutches, but up to about 44% of naturally parasitised clutches (Sealy, 1995; Guigueno and Sealy, 2011), possibly because females interact with the adult parasite (Guigueno and Sealy, 2011). As such, in yellow warblers, increasing circulating corticosterone, via time-release pellets or silastic implants (Torres-Medina et al., 2018), for example, would potentially result in a larger number of abandoners than observed in our current study (i.e., a larger sample size) and would allow for causal, as opposed to correlational, conclusions. Experimentally increasing rates of abandonment through hormone manipulations would still reflect rates seen naturally, as cowbird parasitism and associated abandonment rates vary greatly geographically in yellow warblers (Kuehn, 2009). In sum, we conclude that circulating CORT was associated with abandonment of experimentally parasitised clutches and advocate for follow-up studies that manipulate hormone levels to explore causal mechanisms.
In addition to manipulative studies, more research is needed to understand the role of other hormones and life history in mediating responses to brood parasitism. Hormones other than or in addition to CORT could influence clutch abandonment. Testosterone, progesterone, prolactin, and mesotocin may all play roles in abandonment of parasitised clutches, although this has primarily been examined in hosts that eject cowbird or cuckoo eggs, such as American robins and Eurasian blackbirds, respectively (Abolins-Abols and Hauber, 2018; Ruiz-Raya et al., 2020). In addition, stress is a mediator of life-history trade-offs (Bókony et al., 2009; Bonier et al., 2009; Abolins-Abols and Hauber, 2018), and it would be valuable to incorporate parasitism status and sympatry with cowbirds in a large-scale analysis to examine interactions between hormones, life history traits, and brood parasitism. There is a great opportunity for future research to take a comprehensive endocrine approach in examining the costliest form of rejection in brood-parasitic hosts, clutch abandonment.
This study provides initial findings pertaining to the endocrine correlates of clutch abandonment in brood-parasitic hosts and confirms that egg visual signals play a less important role in eliciting abandonment than tactile cues. The yellow warbler-cowbird system provides a strong model to address mechanistic questions of clutch abandonment, and our research invites future experimental studies incorporating additional species that abandon clutches from diverse host-parasite systems. Our study, along with the proposed follow-up studies, will contribute to a comprehensive understanding of the intricate links between brood parasitism, hormones, and clutch abandonment, and the fitness-related trade-offs between incurring versus avoiding costs associated with brood parasitism and clutch abandonment.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was reviewed and approved by this project was approved and conducted under the Banding Permit no. 23665, United States Fishes and Wildlife Service permit MB72878C-0 and Institutional Animal Care and Use Committee protocol # 19-05-02, all issued to SG.
MG, SG, and AT-VR developed the study design. MG, AT-VR, CP, SG, and JS conducted the data collection and field work. CP, CAP, SG, and JS completed the sample analysis. AT-VR and MG did statistical analysis. AT-VR, CP, and MG wrote the first draft of the manuscript. SG and CP did additional writing, with revisions provided by all other authors.
Funding was provided by a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant to MG (NSERC RGPIN-2019-05947), NSERC Canada Graduate Scholarship (Master’s) to AT-VR, and NSERC Undergraduate Summer Research Award to CP.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank the Southwest Michigan Land Conservancy for allowing us to conduct research on their properties: Sora Meadows and Augusta Floodplains, Jonathan Eiseman for providing accommodation during our stay, Amy Janik for her help with blood sampling and training, and Johnathan Eiseman, Michael Dreibelbis, and Amber Ng for helping with nest monitoring. Finally, we also thank the editor and two reviewers, for their thorough and detailed comments on a previous version of this manuscript that helped us improve the quality of this article.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.711732/full#supplementary-material
Supplementary Figure 1 | (A–F) The stages of the experiment conducted on yellow warbler nests near Kalamazoo, Michigan, from initial completion of building of the nest to laying day 4. (A) A completed nest in the final building stage; (B) Laying day 1, when we artificially parasitised the clutch; (C) A nest after we added two model cowbird eggs to the clutch on laying day 1; (D) Laying day 2; (E) Laying day 3; (F) Laying day 4, days of host blood and feather sampling.
Abernathy, V. E., and Peer, B. D. (2014). Intraclutch variation in egg appearance of brown-headed cowbird hosts. Auk 131, 467–475. doi: 10.1642/AUK-12-186-R1.1
Abolins-Abols, M., and Hauber, M. E. (2018). Host defences against avian brood parasitism: an endocrine perspective. Proc. R. Soc. B Biol. Sci. 285:20180980. doi: 10.1098/rspb.2018.0980
Abolins-Abols, M., and Hauber, M. E. (2020). Endocrine regulation of egg rejection in an avian brood parasite host. Biol. Lett. 16:20200225. doi: 10.1098/rsbl.2020.0225
Ackerman, J. T., Eadie, J. M. A., Yarris, G. S., Loughman, D. L., and McLandress, M. R. (2003). Cues for investment: nest desertion in response to partial clutch depredation in dabbling ducks. Anim. Behav. 66, 871–883. doi: 10.1006/anbe.2003.2283
Aharon-Rotmann, Y., Buttemer, W. A., Koren, L., and Wynne-Edwards, K. (2021). Experimental corticosterone manipulation increases mature feather corticosterone content: implications for inferring avian stress history from feather analyses. bioRxiv [preprint]. doi: 10.1101/2021.01.11.425815 *bioRxiv, 2021.01.11.425815,
Ahlmann-Eltze, C., and Patil, I. (2019). ggsignif: R package for displaying significance brackets for ‘ggplot2’. PsyArxiv [preprint]. doi: 10.31234/osf.io/7awm6
Angelier, F., Clément-Chastel, C., Welcker, J., Gabrielsen, G. W., and Chastel, O. (2009). How does corticosterone affect parental behaviour and reproductive success? A study of prolactin in black-legged kittiwakes. Funct. Ecol. 23, 784–793.
Avilés, J. M. (2018). Can hosts tolerate avian brood parasites? An appraisal of mechanisms. Behav. Ecol. 29, 509–519. doi: 10.1093/beheco/arx150
Bókony, V., Lendvai, Á. Z., Liker, A., Angelier, F., Wingfield, J. C., and Chastel, O. (2009). Stress response and the value of reproduction: are birds prudent parents? Am. Nat. 173, 589–598. doi: 10.1086/597610
Bonier, F., Martin, P. R., Moore, I. T., and Wingfield, J. C. (2009). Do baseline glucocorticoids predict fitness? Trends Ecol. Evol. 24, 634–642. doi: 10.1016/j.tree.2009.04.013
Bortolotti, G. R., Marchant, T., Blas, J., and Cabezas, S. (2009). Tracking stress: localisation, deposition and stability of corticosterone in feathers. J. Exp. Biol. 212, 1477–1482. doi: 10.1242/jeb.022152
Bortolotti, G. R., Marchant, T. A., Blas, J., and German, T. (2008). Corticosterone in feathers is a long-term, integrated measure of avian stress physiology. Funct. Ecol. 22, 494–500. doi: 10.1111/j.1365-2435.2008.01387.x
Bottitta, G. E., Nol, E., and Gilchrist, H. G. (2003). Effects of experimental manipulation of incubation length on behavior and body mass of common eiders in the canadian arctic. Waterbirds 26, 100–107.
Brooke, M., and Davies, N. B. (1988). Egg mimicry by cuckoos cuculus canorus in relation to discrimination by hosts. Nature 335, 630–632. doi: 10.1038/335630a0
Cassey, P., Honza, M., Grim, T., and Hauber, M. E. (2008). The modelling of avian visual perception predicts behavioural rejection responses to foreign egg colours. Biol. Lett. 4, 515–517. doi: 10.1098/rsbl.2008.0279
Clark, K. L., and Robertson, R. J. (1981). Cowbird parasitism and evolution of anti-parasite strategies in the yellow warbler. Wilson Bull. 93, 249–258.
Crossin, G. T., Trathan, P. N., Phillips, R. A., Gorman, K. B., Dawson, A., Sakamoto, K. Q., et al. (2012). Corticosterone predicts foraging behavior and parental care in macaroni penguins. Am. Nat. 180, E31–E41. doi: 10.1086/666001
Croston, R., and Hauber, M. E. (2014). High repeatability of egg rejection in response to experimental brood parasitism in the American robin (turdus migratorius). Behaviour 151, 703–718. doi: 10.1163/1568539X-00003164
Dainson, M., Hauber, M. E., López, A. V., Grim, T., and Hanley, D. (2017). Does contrast between eggshell ground and spot coloration affect egg rejection? Sci. Nat. 104:54. doi: 10.1007/s00114-017-1476-2.c
Davies, N. B., and Brooke, M. D. L. (1989a). An experimental study of co-evolution between the cuckoo, cuculus canorus, and its hosts. I. host egg discrimination. J. Anim. Ecol. 58, 207–224. doi: 10.2307/4995
Davies, N. B., and Brooke, M. D. L. (1989b). An experimental study of co-evolution between the cuckoo, cuculus canorus, and its hosts. II. host egg markings, chick discrimination and general discussion. J. Anim. Ecol. 58, 225–236. doi: 10.2307/4996
Davies, N. B., Krebs, J. R., and West, S. A. (2012). An Introduction to Behavioural Ecology. Hoboken, NJ: John Wiley & Sons.
Ducay, R. L., Luro, A. B., Hansen, E. S., and Hauber, M. E. (2021). Multicomponent shell traits are consistent with an individual recognition function of the appearance of common murre (uria aalge) eggs: a biological replication study. Ecol. Evol. 11, 2402–2409. doi: 10.1002/ece3.7264
Fairhurst, G. D., Marchant, T. A., Soos, C., Machin, K. L., and Clark, R. G. (2013). Experimental relationships between levels of corticosterone in plasma and feathers in a free-living bird. J. Exp. Biol. 216, 4071–4081. doi: 10.1242/jeb.091280
Feeney, W. E., Welbergen, J. A., and Langmore, N. E. (2014). Advances in the study of coevolution between avian brood parasites and their hosts. Annu. Rev. Ecol. Evol. Syst. 45, 227–246. doi: 10.1146/annurev-ecolsys-120213-091603
Fokkema, R. W., Ubels, R., and Tinbergen, J. M. (2016). Great tits trade off future competitive advantage for current reproduction. Behav. Ecol. 27, 1656–1664. doi: 10.1093/beheco/arw097
Freeman, N. E., and Newman, A. E. M. (2018). Quantifying corticosterone in feathers: validations for an emerging technique. Conserv. Physiol. 6:51. doi: 10.1093/conphys/coy051
Gill, S. A., and Sealy, S. G. (2004). Functional reference in an alarm signal given during nest defence: seet calls of yellow warblers denote brood-parasitic brown-headed cowbirds. Behav. Ecol. Sociobiol. 56, 71–80. doi: 10.1007/s00265-003-0736-7
Gómez, J., Gordo, O., and Minias, P. (2021). Egg recognition: the importance of quantifying multiple repeatable features as visual identity signals. PLoS One 16:e0248021. doi: 10.1371/journal.pone.0248021
Griesser, M., Wagner, G. F., Drobniak, S. M., and Ekman, J. (2017). Reproductive trade-offs in a long-lived bird species: condition-dependent reproductive allocation maintains female survival and offspring quality. J. Evol. Biol. 30, 782–795. doi: 10.1111/jeb.13046
Groscolas, R., Lacroix, A., and Robin, J. P. (2008). Spontaneous egg or chick abandonment in energy-depleted king penguins: a role for corticosterone and prolactin? Horm. Behav. 53, 51–60. doi: 10.1016/j.yhbeh.2007.08.010
Guigueno, M. F., and Sealy, S. G. (2009). Nest sanitation plays a role in egg burial by yellow warblers. Ethology 115, 247–256. doi: 10.1111/j.1439-0310.2008.01597.x
Guigueno, M. F., and Sealy, S. G. (2010). Clutch abandonment by parasitized yellow warblers: egg burial or nest desertion? Condor 112, 399–406. doi: 10.1525/cond.2010.090135
Guigueno, M. F., and Sealy, S. G. (2011). Aggression towards egg-removing cowbird elicits clutch abandonment in parasitized yellow warblers, dendroica petechia. Anim. Behav. 81, 211–218. doi: 10.1016/j.anbehav.2010.10.005
Guigueno, M. F., and Sealy, S. G. (2012). Increased investigation of manipulated clutches suggests egg recognition without rejection in a brown-headed cowbird (molothrus ater) host, the yellow warbler (setophaga petechia). Auk 129, 17–25. doi: 10.1525/auk.2011.11138
Guigueno, M. F., Sealy, S. G., and Westphal, A. M. (2014). Rejection of parasitic eggs in passerine hosts: size matters more for a non-ejecter. Auk 131, 583–594. doi: 10.1642/AUK-14-36.1
Hanley, D., López, A. V., Fiorini, V. D., Reboreda, J. C., Grim, T., and Hauber, M. E. (2019). Variation in multicomponent recognition cues alters egg rejection decisions: a test of the optimal acceptance threshold hypothesis. Philos. Trans. R. Soc. B Biol. Sci. 374:20180195. doi: 10.1098/rstb.2018.0195
Hansen, W. K., Bate, L. J., Landry, D. W., Chastel, O., Parenteau, C., and Breuner, C. W. (2016). Feather and faecal corticosterone concentrations predict future reproductive decisions in harlequin ducks (histrionicus histrionicus). Conserv. Physiol. 4:cow015. doi: 10.1093/conphys/cow015
Hauber, M. E., Dainson, M., Luro, A., Louder, A. A., and Hanley, D. (2019). When are egg-rejection cues perceived? A test using thermochromic eggs in an avian brood parasite host. Anim. Cogn. 22, 1141–1148. doi: 10.1007/s10071-019-01306-w
Hauber, M. E., Luro, A., McCarty, C. J., Barateli, K., Cassey, P., Hansen, E. S., et al. (2018). Interannual repeatability of eggshell phenotype in individual female common murres (uria aalge). Can. J. Zool. 97, 385–391. doi: 10.1139/cjz-2018-0172
Hauber, M. E., Moskát, C., and Bán, M. (2006). Experimental shift in hosts’ acceptance threshold of inaccurate-mimic brood parasite eggs. Biol. Lett. 2, 177–180. doi: 10.1098/rsbl.2005.0438
Honza, M., and Cherry, M. I. (2017). “Egg characteristics affecting egg rejection”,” in Avian Brood Parasitism: Behaviour, Ecology, Evolution and Coevolution, ed. M. Soler (Cham: Springer International Publishing), 401–419. doi: 10.1007/978-3-319-73138-4_22
Honza, M., Požgayová, M., Procházka, P., and Tkadlec, E. (2007). Consistency in egg rejection behaviour: responses to repeated brood parasitism in the blackcap (sylvia atricapilla). Ethology 113, 344–351. doi: 10.1111/j.1439-0310.2007.01340.x
Hosoi, S. A., and Rothstein, S. I. (2000). Nest desertion and cowbird parasitism: evidence for evolved responses and evolutionary lag. Anim. Behav. 59, 823–840. doi: 10.1006/anbe.1999.1370
Ibáñez-Álamo, J. D., De Neve, L., Roldán, M., Rodríguez, J., Trouvé, C., Chastel, O., et al. (2012). Corticosterone levels in host and parasite nestlings: is brood parasitism a hormonal stressor? Horm. Behav. 61, 590–597. doi: 10.1016/j.yhbeh.2012.02.008
Jenni-Eiermann, S., Helfenstein, F., Vallat, A., Glauser, G., and Jenni, L. (2015). Corticosterone: effects on feather quality and deposition into feathers. Methods Ecol. Evol. 6, 237–246. doi: 10.1111/2041-210X.12314
Johnston, J. C. (2011). Effects of stage in incubation, time in season, and proportion of original clutch remaining on nest desertion by house sparrows, Passer domesticus. Behaviour 148, 1121–1135. doi: 10.1163/000579511X596589
Kilner, R. M. (2006). The evolution of egg colour and patterning in birds. Biol. Rev. 81, 383–406. doi: 10.1017/S1464793106007044
Kilner, R. M., and Langmore, N. E. (2011). Cuckoos versus hosts in insects and birds: adaptations, counter-adaptations and outcomes. Biol. Rev. 86, 836–852. doi: 10.1111/j.1469-185X.2010.00173.x
Kilner, R. M., Madden, J. R., and Hauber, M. E. (2004). Brood parasitic cowbird nestlings use host young to procure resources. Science 305, 877–879. doi: 10.1126/science.1098487
Klug, H., and Bonsall, M. B. (2010). Life history and the evolution of parental care. Evolution 64, 823–835. doi: 10.1111/j.1558-5646.2009.00854.x
Kraft, F. L. H., Crino, O. L., and Buchanan, K. L. (2021). Developmental conditions have intergenerational effects on corticosterone levels in a passerine. Horm. Behav. 134:105023. doi: 10.1016/j.yhbeh.2021.105023
Krüger, O. (2007). Cuckoos, cowbirds and hosts: adaptations, trade-offs and constraints. Philos. Trans. R. Soc. B Biol. Sci. 362, 1873–1886. doi: 10.1098/rstb.2006.1849
Kuehn, M. J. (2009). Persistence Versus Decline of Host Defenses Against Brood Parasitism: A Model System for Studies of Relaxed Selection and Phenotypic Plasticity? Ph. D, Thesis.
Lahti, D. C. (2006). Persistence of egg recognition in the absence of cuckoo brood parasitism: pattern and mechanism. Evolution 60, 157–168. doi: 10.1111/j.0014-3820.2006.tb01090.x
Landys, M. M., Ramenofsky, M., and Wingfield, J. C. (2006). Actions of glucocorticoids at a seasonal baseline as compared to stress-related levels in the regulation of periodic life processes. Gen. Comput. Endocrinol. 148, 132–149. doi: 10.1016/j.ygcen.2006.02.013
Langmore, N. E., Kilner, R. M., Butchart, S. H. M., Maurer, G., Davies, N. B., Cockburn, A., et al. (2005). The evolution of egg rejection by cuckoo hosts in Australia and Europe. Behav. Ecol. 16, 686–692. doi: 10.1093/beheco/ari041
Lattin, C. R., Reed, J. M., DesRochers, D. W., and Romero, L. M. (2011). Elevated corticosterone in feathers correlates with corticosterone-induced decreased feather quality: a validation study. J. Avian Biol. 42, 247–252. doi: 10.1111/j.1600-048X.2010.05310.x
Lawson, S. L., Enos, J. K., Mendes, N. C., Gill, S. A., and Hauber, M. E. (2021). Pairing status moderates both the production of and responses to anti-parasitic referential alarm calls in male yellow warblers. Ethology 127, 385–394. doi: 10.1111/eth.13139
Lima, S. L. (2009). Predators and the breeding bird: behavioral and reproductive flexibility under the risk of predation. Biol. Rev. 84, 485–513. doi: 10.1111/j.1469-185X.2009.00085.x
Lorenzana, J. C., and Sealy, S. G. (1997). A meta-analysis of the impact of parasitism by the brown-headed cowbird on its hosts. Stud. Avian Biol. 18, 241–253.
Lyon, B. E. (2003). Egg recognition and counting reduce costs of avian conspecific brood parasitism. Nature 422, 495–499. doi: 10.1038/nature01505
MacDougall-Shackleton, S. A., Bonier, F., Romero, L. M., and Moore, I. T. (2019). Glucocorticoids and “stress” are not synonymous. Integrat. Organ. Biol. 1:obz017. doi: 10.1093/iob/obz017
Manna, T., Moskát, C., and Hauber, M. E. (2017). “Cognitive decision rules for egg rejection,” in Avian Brood Parasitism: Behaviour, Ecology, Evolution and Coevolution, ed. M. Soler (Cham: Springer International Publishing), 437–448. doi: 10.1007/978-3-319-73138-4_24
Mark, M. M., and Rubenstein, D. R. (2013). Physiological costs and carry-over effects of avian interspecific brood parasitism influence reproductive tradeoffs. Horm. Behav. 63, 717–722. doi: 10.1016/j.yhbeh.2013.03.008
Martínez-Padilla, J., Mougeot, F., García, J. T., Arroyo, B., and Bortolotti, G. R. (2013). Feather corticosterone levels and carotenoid-based coloration in common buzzard (buteo buteo) nestlings. J. Raptor Res. 47, 161–173. doi: 10.3356/JRR-12-41.1
Medina, I., and Langmore, N. E. (2015). The costs of avian brood parasitism explain variation in egg rejection behaviour in hosts. Biol. Lett. 11:20150296. doi: 10.1098/rsbl.2015.0296
Mico, M. A. (1998). Yellow Warbler Nests, Structure, Building Materials and Cowbird Parasitism. Ph. D, Thesis. Winnipeg: University of Manitoba.
Moskát, C., Hauber, M. E., and Louder, M. I. M. (2017). “The evolution of nest sharing and nest mate killing strategies in brood parasites,” in Avian Brood Parasitism: Behaviour, Ecology, Evolution and Coevolution, ed. M. Soler (Cham: Springer International Publishing), 475–492. doi: 10.1007/978-3-319-73138-4_26
Moskát, C., and Honza, M. (2002). European cuckoo cuculus canorus parasitism and host’s rejection behaviour in a heavily parasitized great reed warbler Acrocephalus arundinaceus population. IBIS 144, 614–622. doi: 10.1046/j.1474-919X.2002.00085.x
Nowicki, S., Searcy, W., and Peters, S. (2002). Brain development, song learning and mate choice in birds: a review and experimental test of the “nutritional stress hypothesis.”. J. Comp. Physiol. A 188, 1003–1014. doi: 10.1007/s00359-002-0361-3
O’Connor, C. M., Norris, D. R., Crossin, G. T., and Cooke, S. J. (2014). Biological carryover effects: linking common concepts and mechanisms in ecology and evolution. Ecosphere 5:28. doi: 10.1890/ES13-00388.1
Ouyang, J. Q., Muturi, M., Quetting, M., and Hau, M. (2013). Small increases in corticosterone before the breeding season increase parental investment but not fitness in a wild passerine bird. Horm. Behav. 63, 776–781. doi: 10.1016/J.YHBEH.2013.03.002
Ouyang, J. Q., Macaballug, P., Chen, H., Hodach, K., Tang, S., and Francis, J. S. (2021). Infrared thermography is an effective, noninvasive measure of HPA activation. Stress 24, 584–589. doi: 10.1080/10253890.2020.1868431
Peer, B. D., McCleery, R. A., and Jensen, W. E. (2018). Resistance is futile: prohibitive costs of egg ejection in an obligate avian brood parasite host. Anim. Behav. 144, 45–51. doi: 10.1016/j.anbehav.2018.08.002
Picman, J. (1989). Mechanism of increased puncture resistance of eggs of brown-headed cowbirds. Auk 106, 577–583. doi: 10.1093/auk/106.4.577
Pyle, P., and Howell, S. N. G., Institute for Bird Populations, and Point Reyes Bird Observatory. (1997). Identification Guide to North American Birds: A Compendium of Information on Identifying, Ageing, and Sexing “Near-Passerines” and Passerines in the Hand. Bolinas, Calif: Slate Creek Press.
R Core Team (2020). R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing.
Rasmussen, J. L., Sealy, S. G., and Underwood, T. J. (2009). Video recording reveals the method of ejection of brown-headed cowbird eggs and no cost in american robins and gray catbirds. Condor 111, 570–574. doi: 10.1525/cond.2009.090019
Roldán, M., and Soler, M. (2011). Parental-care parasitism: how do unrelated offspring attain acceptance by foster parents? Behav. Ecol. 22, 679–691. doi: 10.1093/beheco/arr041
Romero, L. M., and Reed, J. M. (2005). Collecting baseline corticosterone samples in the field: is under 3 min good enough? Comput. Biochem. Physiol. Mol. Integr. Physiol. 140, 73–79. doi: 10.1016/j.cbpb.2004.11.004
Romero, L. M., Reed, J. M., and Wingfield, J. C. (2000). Effects of weather on corticosterone responses in wild free-living passerine birds. Gen. Comput. Endocrinol. 118, 113–122. doi: 10.1006/gcen.1999.7446
Rothstein, S. I. (1974). Mechanisms of avian egg recognition: possible learned and innate factors. Auk 91, 796–807. doi: 10.2307/4084731
Rothstein, S. I. (1982). Mechanisms of avian egg recognition: which egg parameters elicit responses by rejecter species? Behav. Ecol. Sociobiol. 11, 229–239. doi: 10.1007/BF00299299
Rothstein, S. I. (1990). A model system for coevolution: avian brood parasitism. Annu. Rev. Ecol. Syst. 21, 481–508.
Ruiz-Raya, F., Ibáñez-Álamo, J. D., Parenteau, C., Chastel, O., and Soler, M. (2020). Prolactin mediates behavioural rejection responses to avian brood parasitism. bioRxiv [preprint]. doi: 10.1101/2020.10.28.358994 *bioRxiv, 2020.10.28.358994,
Ruiz-Raya, F., Soler, M., Abaurrea, T., Chastel, O., Roncalli, G., and Ibáñez-Álamo, J. D. (2018). Hormonal responses to non-mimetic eggs: is brood parasitism a physiological stressor during incubation? Behav. Ecol. Sociobiol. 72:153. doi: 10.1007/s00265-018-2565-8
Sarpong, K., Madliger, C. L., Harris, C. M., Love, O. P., Doucet, S. M., and Bitton, P. P. (2019). Baseline corticosterone does not reflect iridescent plumage traits in female tree swallows. Gen. Comput. Endocrinol. 270, 123–130. doi: 10.1016/j.ygcen.2018.10.015
Scharf, H. M., Abolins-Abols, M., Stenstrom, K. H., Tolman, D. T., Schelsky, W. M., and Hauber, M. E. (2021). Exposure to a mimetic or non-mimetic model avian brood parasite egg does not produce differential glucocorticoid responses in an egg-accepter host species. Gen. Comput. Endocrinol. 304:113723. doi: 10.1016/j.ygcen.2021.113723
Schoenle, L. A., Dudek, A. M., Moore, I. T., and Bonier, F. (2017). Red-winged blackbirds (Agelaius phoeniceus) with higher baseline glucocorticoids also invest less in incubation and clutch mass. Horm. Behav. 90, 1–7. doi: 10.1016/j.yhbeh.2017.02.002
Schulze-Hagen, K., Stokke, B. G., and Birkhead, T. R. (2009). Reproductive biology of the european cuckoo cuculus canorus: early insights, persistent errors and the acquisition of knowledge. J. Ornithol. 150, 1–16. doi: 10.1007/s10336-008-0340-8
Sealy, S. G. (1995). Burial of cowbird eggs by parasitized yellow warblers: an empirical and experimental study. Anim. Behav. 49, 877–889. doi: 10.1006/anbe.1995.0120
Segura, L. N., Di Sallo, F. G., Mahler, B., and Reboreda, J. C. (2016). Red-crested cardinals use color and width as cues to reject shiny cowbird eggs. Auk 133, 308–315. doi: 10.1642/AUK-15-181.1
Servedio, M. R., and Hauber, M. E. (2006). To eject or to abandon? Life history traits of hosts and parasites interact to influence the fitness payoffs of alternative anti-parasite strategies. J. Evol. Biol. 19, 1585–1594. doi: 10.1111/j.1420-9101.2006.01124.x
Sherriff, M. J., Dantzer, B., Delehanty, B., Palme, R., and Boonstra, R. (2011). Measuring stress in wildlife: techniques for quantifying glucocorticoids. Oecologia 166, 869–887. doi: 10.1007/s00442-011-1943-y
Soler, M. (2017). “Begging behaviour, food delivery and food acquisition in nests with brood parasitic nestlings,” in Avian Brood Parasitism: Behaviour, Ecology, Evolution and Coevolution, ed. M. Soler (Cham: Springer International Publishing), 493–515. doi: 10.1007/978-3-319-73138-4_27
Soler, M., Martinez, J. G., Soler, J. J., and Møller, A. P. (1995). Preferential allocation of food by magpies Pica pica to great spotted cuckoo Clamator glandarius chicks. Behav. Ecol. Sociobiol. 37, 7–13. doi: 10.1007/BF00173893
Soler, M., Ruiz-Raya, F., Roncalli, G., and Ibáñez-Álamo, J. D. (2015). Nest desertion cannot be considered an egg-rejection mechanism in a medium-sized host: an experimental study with the common blackbird turdus merula. J. Avian Biol. 46, 369–377. doi: 10.1111/jav.00571
Soler, M., Ruiz-Raya, F., Roncalli, G., and Ibáñez-Álamo, J. D. (2017). Relationships between egg-recognition and egg-ejection in a grasp-ejector species. PLoS One 12:e0166283. doi: 10.1371/journal.pone.0166283
Spaw, C. D., and Rohwer, S. (1987). A comparative study of eggshell thickness in cowbirds and other passerines. Condor 89, 307–318. doi: 10.2307/1368483
Spée, M., Beaulieu, M., Dervaux, A., Chastel, O., Le Maho, Y., and Raclot, T. (2010). Should I stay or should I go? Hormonal control of nest abandonment in a long-lived bird, the Adélie penguin. Horm. Behav. 58, 762–768. doi: 10.1016/j.yhbeh.2010.07.011
Spée, M., Marchal, L., Lazin, D., Le Maho, Y., Chastel, O., Beaulieu, M., et al. (2011). Exogenous corticosterone and nest abandonment: a study in a long-lived bird, the Adélie penguin. Horm. Behav. 60, 362–370. doi: 10.1016/j.yhbeh.2011.07.003
Spencer, K. A., and MacDougall, S. A. (2011). Indicators of development as sexually selected traits: the developmental stress hypothesis in context. Behav. Ecol. 22, 1–9. doi: 10.1093/beheco/arq068
Spottiswoode, C. N., and Stevens, M. (2010). Visual modeling shows that avian host parents use multiple visual cues in rejecting parasitic eggs. Proc. Natl. Acad. Sci. U.S.A. 107, 8672–8676. doi: 10.1073/pnas.0910486107
Spottiswoode, C. N., and Stevens, M. (2011). How to evade a coevolving brood parasite: egg discrimination versus egg variability as host defences. Proc. R. Soc. B Biol. Sci. 278, 3566–3573. doi: 10.1098/rspb.2011.0401
Strange, M. S., Bowden, R. M., Thompson, C. F., and Sakaluk, S. K. (2016). Pre- and postnatal effects of corticosterone on fitness-related traits and the timing of endogenous corticosterone production in a songbird. J. Exp. Zool. Part A Ecol. Genet. Physiol. 325, 347–359. doi: 10.1002/jez.2022
Tata, J. R. (2005). One hundred years of hormones. EMBO Rep. 6, 490–496. doi: 10.1038/sj.embor.7400444
Thierry, A. M., Massemin, S., Handrich, Y., and Raclot, T. (2013). Elevated corticosterone levels and severe weather conditions decrease parental investment of incubating Adélie penguins. Horm. Behav. 63, 475–483. doi: 10.1016/j.yhbeh.2012.12.011
Thorogood, R., Spottiswoode, C. N., Portugal, S. J., and Gloag, R. (2019). The coevolutionary biology of brood parasitism: a call for integration. Philos. Trans. R. Soc. B Biol. Sci. 374:20180190. doi: 10.1098/rstb.2018.0190
Torres-Medina, F., Cabezas, S., Marchant, T. A., Wikelski, M., Romero, L. M., Hau, M., et al. (2018). Corticosterone implants produce stress-hyporesponsive birds. J. Exp. Biol. 221:173864. doi: 10.1242/jeb.173864
Verboven, N., and Tinbergen, J. M. (2002). Nest desertion: a trade-off between current and future reproduction.y Anim. Behavor 63, 951–958. doi: 10.1006/anbe.2001.1971
Wada, H., Hahn, T. P., and Breuner, C. W. (2007). Development of stress reactivity in white-crowned sparrow nestlings: total corticosterone response increases with age, while free corticosterone response remains low. Gen. Comput. Endocrinol. 150, 405–413. doi: 10.1016/j.ygcen.2006.10.002
Wiggins, D. A., Pärt, T., and Gustafsson, L. (1994). Correlates of clutch desertion by female collared flycatchers Ficedula albicollis. J. Avian Biol. 25, 93–97. doi: 10.2307/3677025
Wingfield, J. C. (2003). Control of behavioural strategies for capricious environments. Anim. Behav. 66, 807–816. doi: 10.1006/anbe.2003.2298
Wingfield, J. C., Maney, D. L., Breuner, C. W., Jacobs, J. D., Lynn, S., Ramenofsky, M., et al. (1998). Ecological bases of hormone—behavior interactions: the “emergency life history stage”1. Am. Zool. 38, 191–206. doi: 10.1093/icb/38.1.191
Keywords: avian brood parasitism, egg rejection, egg visual signals, clutch abandonment, Setophaga petechia, Molothrus ater, corticosterone
Citation: Turcotte-van de Rydt AV, Petalas C, Sblendorio JM, Pearl CA, Gill SA and Guigueno MF (2022) Clutch Abandoning Parasitised Yellow Warblers Have Increased Circulating Corticosterone With No Effect of Past Corticosterone or Differences in Egg Maculation Characteristics. Front. Ecol. Evol. 10:711732. doi: 10.3389/fevo.2022.711732
Received: 19 May 2021; Accepted: 18 January 2022;
Published: 24 February 2022.
Edited by:
Brian Peer, Western Illinois University, United StatesReviewed by:
James Frederick Hare, University of Manitoba, CanadaCopyright © 2022 Turcotte-van de Rydt, Petalas, Sblendorio, Pearl, Gill and Guigueno. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Antoine V. Turcotte-van de Rydt, YW50b2luZS50dXJjb3R0ZS12YW5kZXJ5ZHRAbWFpbC5tY2dpbGwuY2E=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.