 Nan Jia1,2
Nan Jia1,2 Karl J. Niklas
Karl J. Niklas Buqing Yao
Buqing Yao Zhiqiang Wang
Zhiqiang Wang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 13 December 2022
Sec. Biogeography and Macroecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.1093474
This article is part of the Research Topic Ecosystem Nutrients Stoichiometry From Local to Global Scales View all 8 articles
Understanding the altitudinal patterns of plant stoichiometry in seeds is critical for characterizing important germination and dormancy strategies, soil seed bank composition, seed predation probability, efficiency of seed dispersal and seedling performance, and to predict how biodiversity might be influenced by climate change. However, our understanding of the altitudinal patterns of seed stoichiometry is extremely limited. In this study, we measured the concentrations of carbon (C), nitrogen (N) and phosphorus (P) in the seeds of 253 herbaceous species along an altitudinal transect (2,000–4,200 m) on the eastern Tibetan Plateau, China, and further to characterize seed C:N:P stoichiometry. The geometric means of C, N, and P concentrations were 569.75 mg/g, 34.76 mg/g, and 5.03 mg/g, respectively. The C:N, C:P, and N:P ratios were 16.39, 113.31, and 6.91, respectively. The seed C, N, and P concentrations and C:N:P ratios varied widely among major plant groups and showed significant altitudinal trends. In general, C, N, and P concentrations increased, whereas seed C:N:P ratios decreased with elevation. These results inform our understanding of the altitudinal patterns of seed stoichiometry and how to model ecosystem nutrient cycling.
Ecological stoichiometry provides a framework for understanding plant strategies dependent on or influenced by the differential allotment of multiple chemicals in different organs. This framework is important because it sheds light on ecosystem functionalities and is a valuable method for indirectly assessing the biogeochemical cycling of terrestrial ecosystems (Elser et al., 2000; Sterner and Elser, 2002; Wang et al., 2022a). Carbon (C), nitrogen (N), and phosphorus (P) are the three essential elements and play important roles in regulating plant growth and development (Aerts and Chapin, 2000; Vitousek, 2004). For example, the C:N:P ratios of plant organs have been used to evaluate potential terrestrial nutrient limitations (Güsewell, 2004; Reich and Oleksyn, 2004) and to characterize biogeochemical cycling of ecosystems (Güsewell and Gessner, 2009; Wang et al., 2015). In addition, the C:N:P ratios of plant organs vary significantly among different environmental gradients (McGroddy et al., 2004; Reich and Oleksyn, 2004; Yuan et al., 2011; Wang et al., 2020a). Therefore, evaluating the patterns of C:N:P ratios of plant organs along environmental gradients can improve our understanding the effects of climate change on global C, N, and P coupling cycles.
Changes in elevation are correlated with changes in a number of environmental factors including temperature, CO2 concentration, light, precipitation, and soil-type, which in turn can lead to differences in plant C, N, and P concentrations and their ratios (Sundqvist et al., 2011; Wang et al., 2020b). These environmental differences are also correlated with changes in the types of selection pressure on plant survival, growth and physiological function, thereby resulting in different adaptions (Lord, 1994). For example, leaf N and P concentrations decline and N:P ratios increase with increasing elevation as a result of low temperature suppression of soil nutrient mineralization and the decomposition of organic matter, both of which limit root nutrient uptake (Reich and Oleksyn, 2004). Moreover, increasing elevation is reported to affect the accumulation of inorganic P in leaves with an attending downward trend in N:P (Reich and Oleksyn, 2004). Similarly, fine root N and P concentrations decrease with increasing elevation, whereas fine root N:P ratio increasing (Zhao et al., 2016). Climate and soil variables jointly influence fine root C, N, and P concentrations and their ratios (Yuan et al., 2011; Wang et al., 2021a).
The effects of elevation on specific plant organs, such as leaves (Soethe et al., 2008; Hoch and Körner, 2012; Zhao et al., 2014), stems (Yao et al., 2015; Wang et al., 2018, 2022b) and roots (Yuan et al., 2011; Wang et al., 2022c) as well as soil microbes (He et al., 2019; Wang et al., 2021b) have been widely investigated and have significantly improved our understanding of plant responses to differences in elevation. However, our understanding of the effects (if any) on seed C:N:P stoichiometry is poor. Yet, during the earlier stages of plant ontogeny, many developmental and physiological processes depend on seed reserves (Bewley et al., 2013). For example, the C, N and P reserves in seeds are critical components affecting growth (Bu et al., 2018) because C is the most abundant component of dry matter, whereas N affects protein synthesis and P is essential for the synthesis of nucleobases (Elser et al., 2000; Sterner and Elser, 2002). Therefore, seed reserves might influence seed dormancy, germination, and survival with different nutrient elements having different effects on each stage. Given the importance of C, N and P to seedling growth and survival, it is important to evaluate see C:N:P stoichiometry, particularly along elevational gradients.
The goal of this paper was to evaluate the effects of elevation on seed stoichiometry in the eastern Tibetan Plateau, which represents one of the largest alpine meadows in the world, covering more than 60% of the area of the alpine steppe and alpine meadow (Yang et al., 2015). In this study, we compiled a dataset on C, N and P seed concentrations for 253 herbaceous species along an altitudinal gradient to determine the seed C, N, and P concentrations and C:N:P ratios across different plant groups, and examine the elevational patterns of the seed C, N, and P concentrations and C:N:P ratios.
This study area is located on the eastern edge of the Tibetan Plateau in China (100°44′-104°45′E, 33°06′-35°34′N; Figure 1). The climate is cold and humid, and the elevation rises from 2000 to 4,200 m. The mean annual temperature ranges from 1.2°C-4.6°C and the mean annual precipitation increased from 450 to 780 mm and growing season is short (from late May to late September; Bu et al., 2016, 2018). The main vegetation types in typical alpine meadow dominated by various graminoids (e.g., Cyperaceae and Poaceae) and herbs (e.g., Asteraceae, Fabaceae, Gentianaceae, Polygonaceae, Ranunculaceae, and Scrophulariaceae). All these factors make it an optimal site to explore the altitudinal patterns of seed C:N:P stoichiometry.
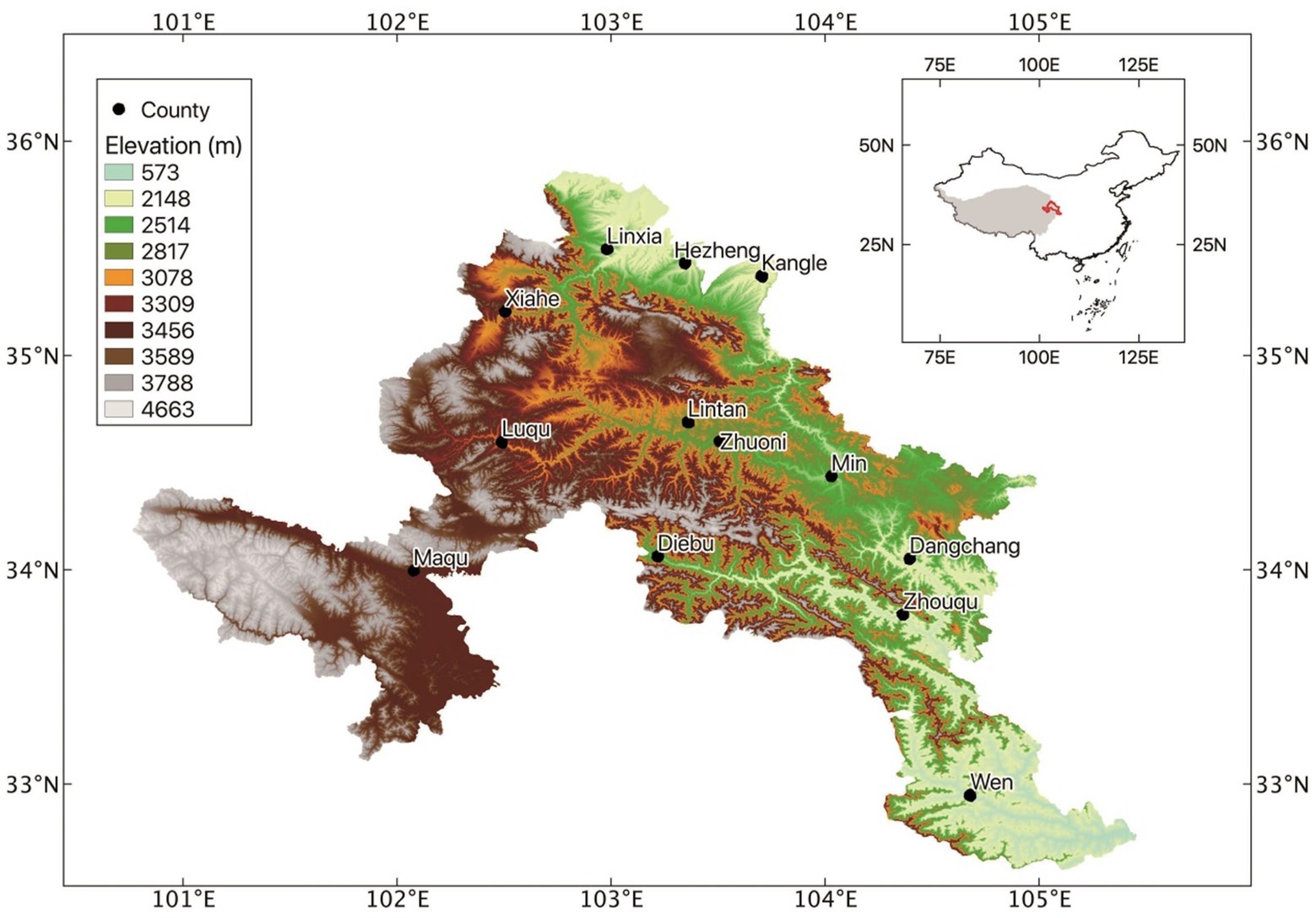
Figure 1. The study area and sampling sites along the elevation gradients.
The mature seeds of 253 species from 37 families were examined along an altitudinal gradient from late August to September in 2015. The time of collection was based on prior extensive field observations on seed development and dispersal time. See maturity was determined using a number of physical cues including see color, pericarp consistency, and the ease of detaching them. Mature seeds were sampled randomly from over 30 individual of each species from sites differing in 50 m across the elevational gradient. Intact seeds were stored in envelopes and oven-dried at 50°C to constant weight after which seeds were ground into a fine powder and C, N, and P concentrations were measured. Dry combustion in an elemental analyzer (Elementar TOC Vario, Germany) was used to measure C and N concentrations. The parts of fine powder were digested using microwave heating, and volumes of 5 ml of HNO3 65% and 1 ml of H2O2 30% were added for the digestion. The molybdenum blue method on an automatic flow injection analyzer (Lachat Quickchem 8500, United States) was used to measure P concentrations as described by Wang et al. (2020b). The stoichiometric ratios of C:N, C:P and N:P ratios were calculated on a mass basis.
The plant species were divided into two life-form groups (i.e., annual and perennial), two functional groups (i.e., forb and graminoid), two N-fixing groups (i.e., N-fixing and non-N-fixing), and two phylogenetic groups (i.e., monocotyledons and eudicots). We compared the differences in C, N, and P concentrations and C:N:P ratios of seeds across major functional groups using one-way analysis of variance (ANOVA) least-significant difference with (LSD) post hoc tests. The relationships between seed C, N, and P concentrations and C:N:P ratios and elevation were explored by ordinary least squares (OLS) regressions analysis after the seed C, N, and P, concentrations and C:N:P ratios were log-transformed to normalize frequency distributions. All statistical analyses were performed using R 2.15.2 (R Development Core Team, 2015).
The geometric mean values of seed C, N, and P concentrations were 569.75 mg/g, 34.76 mg/g, and 5.03 mg/g, respectively (Table 1 and Figure 2). The C concentration ranged from 501.23 to 575.9 mg/g, N concentration ranged from 21.54–57.17 mg/g, whereas P concentration ranged from 2.87–5.18 mg/g (Table 1). The C:N:P ratios ranged from 8.77 to 24.65 for C:N, 99.11 to 185.27 for C:P, and 6.36 to 11.3 for N:P (Table 1). Seed C:N:P ratios differed significantly across different major plant groups. Seed C:N and C:P ratios were the highest in graminoids, whereas N-fixing species had the highest N:P ratios. In contrast, seed C:N and C:P ratios were the lowest in N-fixing species, whereas annual species had the lowest N:P ratios (Table 1).
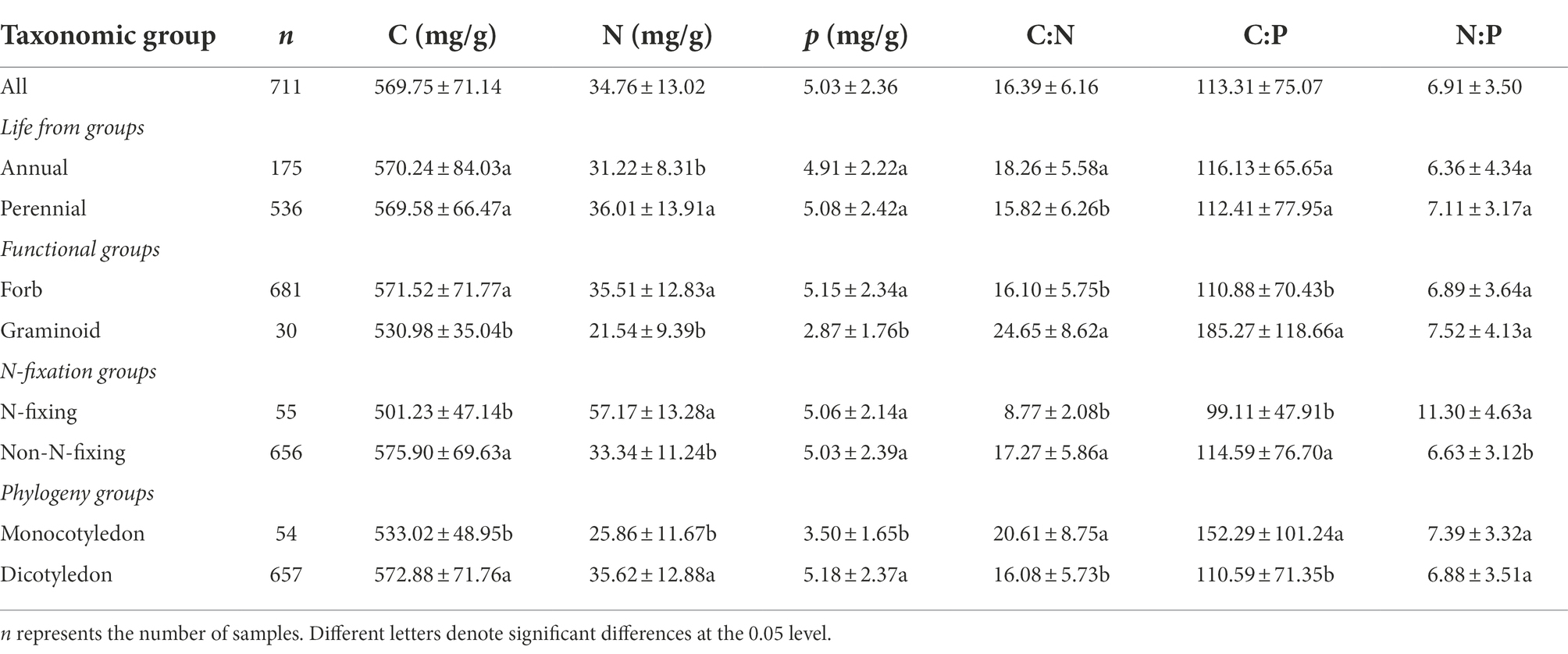
Table 1. Geometric means and standard deviations of seed C, N, and P concentrations and their ratios for major plant groups.
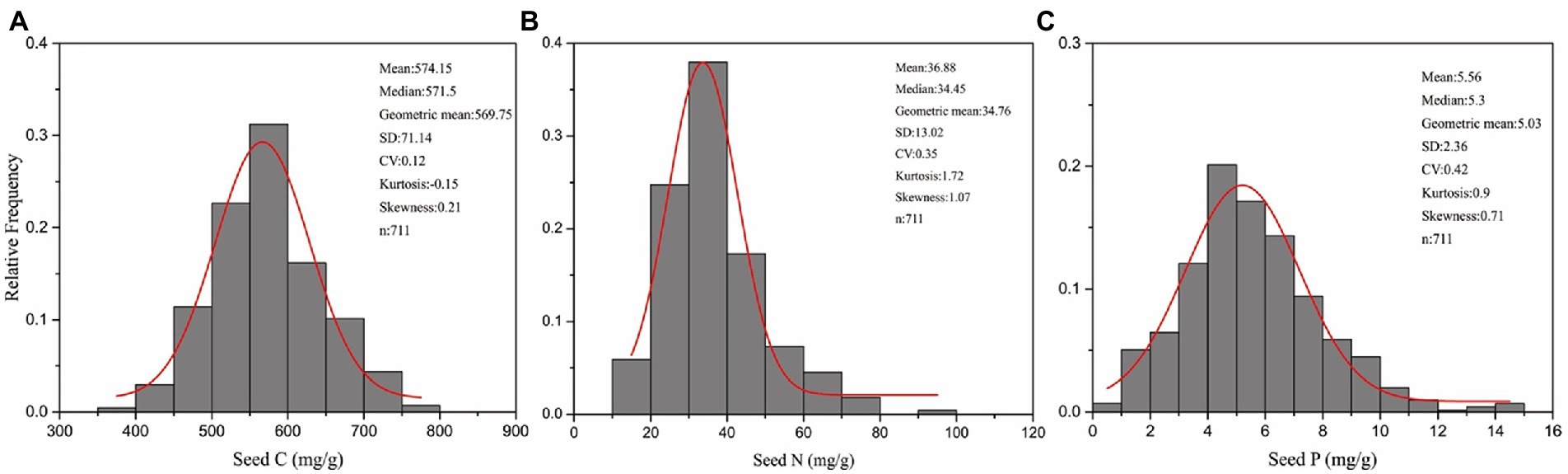
Figure 2. Frequency distribution of seed carbon (C) (A), nitrogen (N) (B) and phosphorus (P) concentrations (C) for all 253 herbaceous species on the eastern Tibetan Plateau.
Seed C, N, and P concentrations and C:N:P ratios were significantly correlated with altitude (p < 0.05; Figure 3). C, N, and P concentrations increased significantly with elevation (p < 0.05; Figures 3A–C), whereas seed C:N:P ratios decreased with increased elevation (p < 0.01; Figures 3D–F).
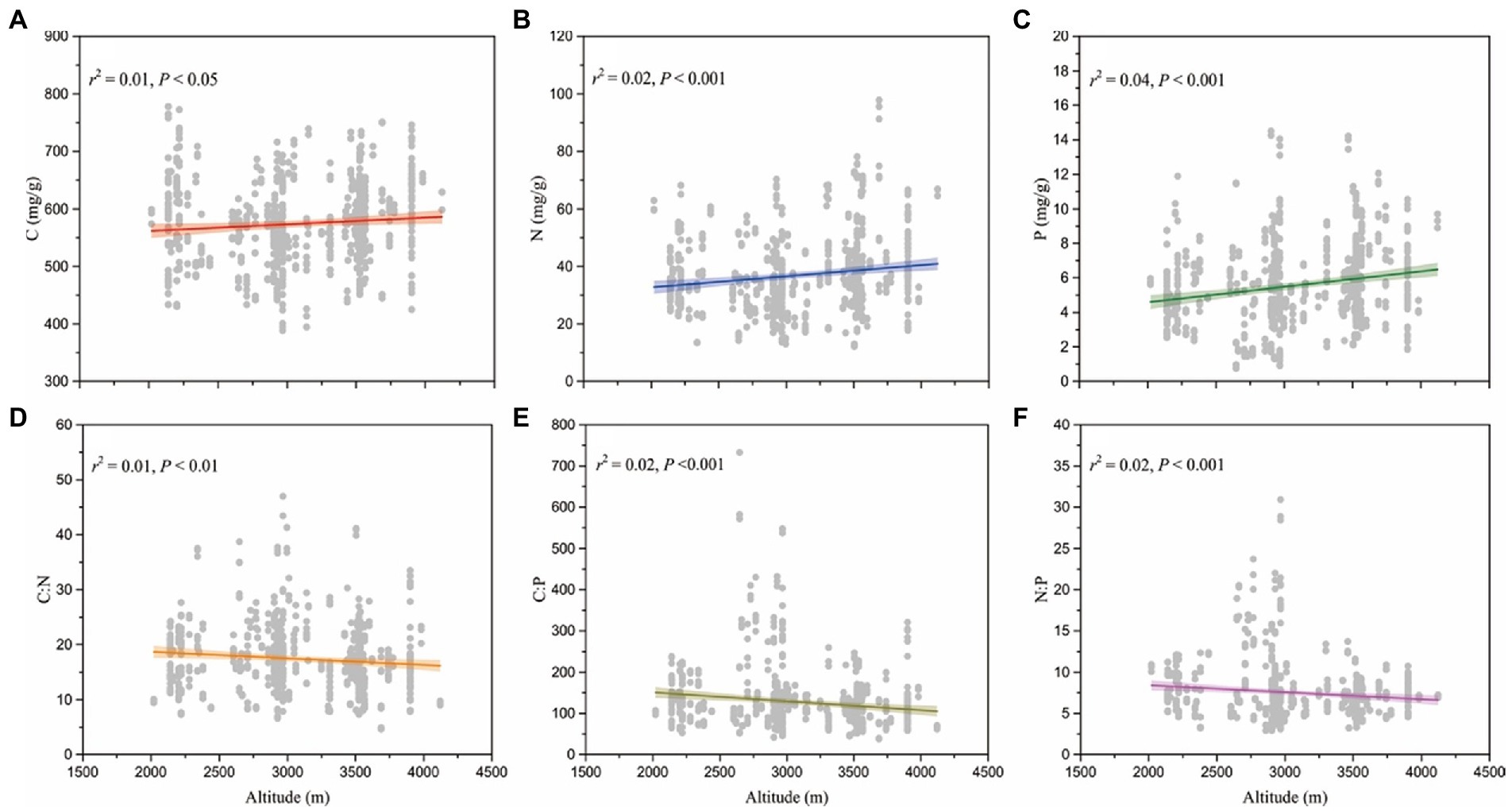
Figure 3. Changes in seed C, N, P (A-C) and C:N:P (D-F) ratios with the elevational gradients.
Our results indicate that seed C, N and P concentrations on the eastern Tibetan Plateau have geometric means that are 569.75 mg/g, 34.76 mg/g, and 5.03 mg/g, respectively, and thus higher than that of leaves (435 mg/g, 29.2 mg/g, and 2 mg/g, respectively) reported by He et al. (2006) and fine roots (447.6 mg/g, 11.09 mg/g, and 0.91 mg/g, respectively) reported by Geng et al. (2014) for herbaceous species in an alpine meadow. These findings support the fact that compared to leaves or fine roots, germinating seeds manifest higher metabolic activity and thus require higher amounts of nutrients for germination and primary growth (Bu et al., 2008) until seedlings become established and autotrophic (Soriano et al., 2011; Bewley et al., 2013; Cheng et al., 2015). For example, P reflects nucleic acid content, which is required for the synthesis of the sugar-phosphate intermediates of photosynthesis and respiration (Sterner and Elser, 2002). P content can provide sufficient ribosomes for seed germination and rapid initial growth (Elser et al., 2003). Higher C, N, and P content in seeds may also reflect an adaptive strategy for survival and preservation. In contrast, the seed C:N, C:P, and N:P ratios on the eastern Tibetan Plateau have a geometric mean of 16.39, 113.31, and 6.91, respectively, which are lower than that fine roots (40.36, 491.87, and 6.91, respectively) reported by Geng et al. (2014) for herbaceous species in an alpine meadow. The lower C:N:P ratios of seeds supports the growth rate hypothesis, which postulates that fast-growing species have higher P content and lower C:P and N:P ratios because higher growth rates requires more P supported by P-rich rRNA than N to ensure rapid protein synthesis (Elser et al., 2000; Sterner and Elser, 2002).
Seed C, N, and P concentrations and C:N:P ratios differ across major plant groups (Table 1). These differences may reflect different nutrient availability strategies among different plant groups. We found significantly higher N concentrations for perennial species compared with annual species. This result is consistent with a previous study (Bu et al., 2008), suggesting that higher seed N content is positively correlated with germination success (de Frenne et al., 2011). The lower C:N ratios in perennial species than annual species is likely mainly due to larger N concentrations in perennial species. Forb species have markedly higher C, N, and P concentrations than graminoid species. These results might be attributed to forbs generally having more rapid germination rates and more vigorous seedlings compared to the seeds of grasses (Ching and Rynd, 1978). Similarly, eudicots generally have higher seed C, N, and P concentrations than monocotyledon, perhaps as a consequence of differences in their ability to store nutrients (Bonfil, 1998). The N concentrations of seeds was significantly higher in N-fixing species than non-N-fixing species owing to the ability to absorb N in fine roots (McCormack et al., 2015) and also perhaps because of the high metabolic cost of N-fixation (Wardle and Greenfield, 1991). Interestingly, the seed N:P ratios in our study do not vary across the major plant groups, with the exception of N-fixers. This phenomenon may reflect a fundamental constraint on seed N and P concentrations and N:P ratios.
Seed C, N, and P concentrations and C:N:P ratios are closely correlated with the elevational patterns on the eastern Tibetan Plateau (p < 0.05). The variations of climate and the limitation of nutrient availability along an elevational gradient may interact to shape the elevational patterns of seed C, N, and P concentrations and C:N:P ratios. Seed C concentration increases significantly with altitude, supporting the notion that plant growth in higher elevations experiencing more stressed conditions require more non-structural C, including starch, low molecular weight sugars, and storage lipids to maintain cellular osmotic pressure and to resist freezing (Hoch et al., 2002; Millard et al., 2007; Hoch and Körner, 2012). Likewise, seed N and P concentrations increase with elevation, suggesting that for rapid initial growth, species from high elevation areas would take the opportunity to recruit seedlings. Thus, the high content of N and P in seeds of these species might be the result of natural selection for rapid initial growth. These patterns also support the Temperature-Plant Physiology and the Soil Substrate Age hypotheses (Reich and Oleksyn, 2004). Plants growing in high elevations tend to have greater N and P concentrations purportedly as a defense against the effects of low temperatures. This speculation is consistent with mechanisms that reduce N-rich enzymatic efficiency at low temperatures and P-rich RNA that can compensate for decreased rates of metabolic reactions (Weih and Karlsson, 2001; Reich and Oleksyn, 2004). In addition, our data indicate that seed C, N, and P concentrations are highly associated with one another (p < 0.01; Figure 4). This result indirectly supports the observation that seed C, N, and P concentrations follow the same trends along an elevational gradient.
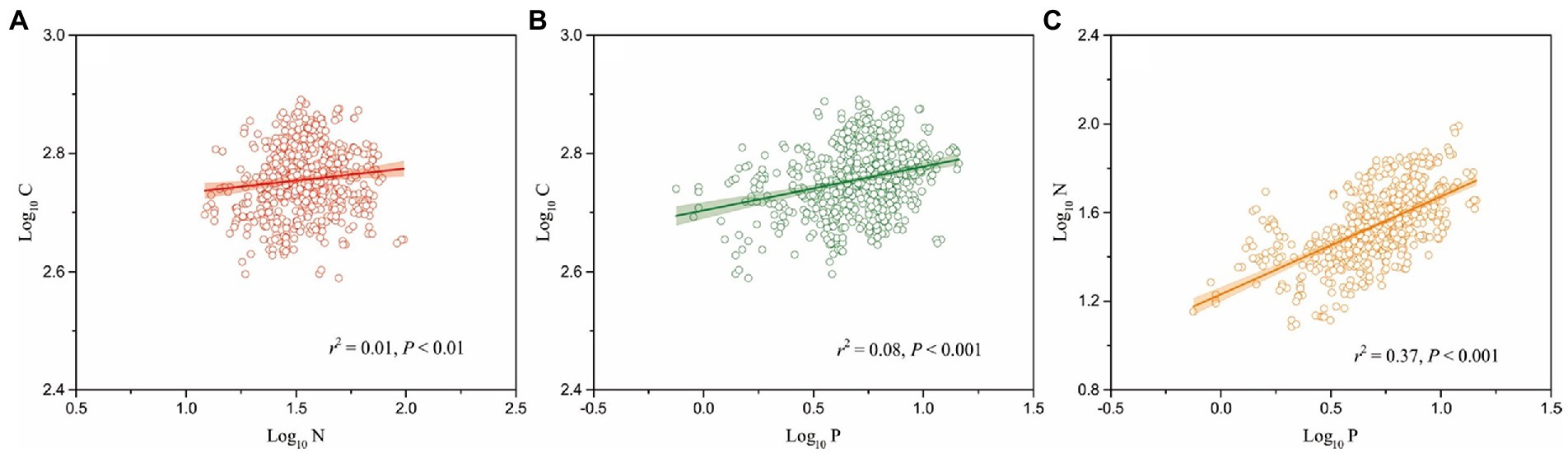
Figure 4. Relationships of seed carbon and nitrogen (A), carbon and phosphorus (B), nitrogen and phosphorus (C) concentrations for all 253 herbaceous species on the eastern Tibetan Plateau.
The data presented here indicate that seed C:N:P ratios decrease along an elevational gradient. Previous studies have shown that low temperatures can depress soil microbe activity, resulting in the slow decomposition of organic matter, thereby reducing the availability of N and P in soils and thus limiting N and P uptake by roots (Körner, 1989; Wang et al., 2019; Long et al., 2020, 2022). Moreover, aggravated P leaching in soils caused by the increased precipitation at higher elevations can reduce P availability (Hedin et al., 2003). The decreasing seed N:P ratio along an elevational gradient indirectly supports the Temperature-Physiological hypotheses, which states that leaf N:P ratios decrease with mean annual temperature (MAT) at a global scale as a result of both physiological acclimation and adaptation to temperature (Reich and Oleksyn, 2004; Lin et al., 2021; Wang et al., 2022d). Collectively, these elevational patterns probably are the result of the collective influences of climate and soil drivers that lead to a gradual shift in seed C:N:P stoichiometry from low to high elevations. However, due to the lack of data of soil nutrients (e.g., soil total C, N, and P) in our dataset, our understanding of the mechanisms driving the elevational patterns of seed C:N:P stoichiometry on the eastern Tibetan Plateau is still severely limited and warrants further investigation.
In summary, this study provides a comprehensive documentation of the seed C:N:P stoichiometry across different plant groups in an alpine meadow on the eastern Tibetan Plateau. The results presented indicate that the seed C, N, and P concentrations and C:N:P ratios differ across major plant groups. Seed C, N, and P concentrations and C:N:P ratios also exhibited a statistically robust elevational pattern on the eastern Tibetan Plateau. These results advance our understanding of plant stoichiometry and have important implications for ecosystem functioning across large environmental gradients.
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
ZW conceived the idea and designed the research. NJ, KN, BY, and ZW performed the data analysis and wrote the paper. All authors contributed to the article and approved the submitted version.
This study was financially supported by the International Science and Technology Cooperation Project of Qinghai province of China (2022-HZ-817).
We are grateful to Jordi López-Pujol for the help with species identification.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aerts, R., and Chapin, F. S. (2000). The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns. Adv. Ecol. Res. 30, 1–67. doi: 10.1016/S0065-2504(08)60016-1)
Bewley, J. D., Bradford, K. J., Hilhorst, H. W. M., and Nonogaki, H. (2013). Seeds: Physiology of Development, Germination and Dormancy, 3rd Edn.. New York, Springer.
Bonfil, C. (1998). The effects of seed size, cotyledon reserves, and herbivory on seedling survival and growth in Quercus rugosa and Q. laurina (Fagaceae). Am. J. Bot. 85, 79–87. doi: 10.2307/2446557
Bu, H. Y., Du, G. Z., Chen, X. L., Xu, X. L., Liu, K., and Wen, S. J. (2008). Community-wide germination strategies in an alpine meadow on the eastern Qinghai-Tibetan plateau: phylogenetic and life-history correlates. Plant Ecol. 195, 87–98. doi: 10.1007/s11258-007-9301-1
Bu, H. Y., Jia, P., Qi, W., Liu, K., Xu, D. H., Ge, W. J., et al. (2018). The effects of phylogeny, life-history traits and altitude on the carbon, nitrogen, and phosphorus contents of seeds across 203 species from an alpine meadow. Plant Ecol. 219, 737–748. doi: 10.1007/s11258-018-0830-6
Bu, H. Y., Wang, X. J., Zhou, X. H., Qi, W., Liu, K., Ge, W. J., et al. (2016). The ecological and evolutionary significance of seed shape and volume for the germination of 383 species on the eastern Qinghai-Tibet plateau. Folia Geobot. 51, 333–341. doi: 10.1007/s12224-016-9271-y
Cheng, J. P., Cheng, X. X., Wang, L., He, Y. Q., An, C. F., Wang, X. F., et al. (2015). Physiological characteristics of seed reserve utilization during the early seedling growth in rice. Braz. J. Bot. 38, 751–759. doi: 10.1007/s40415-015-0190-6
Ching, T. M., and Rynd, L. (1978). Developmental differences in embryos of high and low protein wtheat seeds during germination. Plant Physiol. 62, 866–870. doi: 10.1104/pp.62.6.866
de Frenne, P., Kolb, A., Graae, B. J., Decocq, G., Baltora, S., de Schrijver, A., et al. (2011). A latitudinal gradient in seed nutrients of the forest herb Anemone nemorosa. Plant Biol. 13, 493–501. doi: 10.1111/j.1438-8677.2010.00404.x
Elser, J. J., Acharya, K., Kyle, M., Cotner, J., Makino, W., Markow, T., et al. (2003). Growth rate-stoichiometry couplings in diverse biota. Ecol. Lett. 6, 936–943. doi: 10.1046/j.1461-0248.2003.00518.x
Elser, J. J., Sterner, R. W., Gorokhova, E., Fagan, W. F., Markow, T. A., Cotner, J. B., et al. (2000). Biological stoichiometry from genes to ecosystems. Ecol. Lett. 3, 540–550. doi: 10.1046/j.1461-0248.2000.00185.x
Geng, Y., Wang, L., Jin, D. M., Liu, H. Y., and He, J. S. (2014). Alpine climate alters the relationships between leaf and root morphological traits but not chemical traits. Oecologia 175, 445–455. doi: 10.1007/s00442-014-2919-5
Güsewell, S. (2004). N: P ratios in terrestrial plants: variation and functional significance. New Phytol. 164, 243–266. doi: 10.1111/j.1469-8137.2004.01192.x
Güsewell, S., and Gessner, M. O. (2009). N:P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms. Funct. Ecol. 23, 211–219. doi: 10.1111/j.1365-2435.2008.01478.x
He, X. J., Hou, E. Q., Veen, G. F., Farnon Ellwood, M. D., Dijkstra, P., Sui, X. H., et al. (2019). Soil microbial biomass increase along elevational gradients in the tropics and subtropics but not elsewhere. Glob. Ecol. Biogeogr. 29, 345–354. doi: 10.1111/geb.13017
He, J. S., Wang, Z. H., Wang, X. P., Schmid, B., Zuo, W. Y., Zhou, M., et al. (2006). A test of the generality of leaf trait relationship on the Tibetan plateau. New Phytol. 170, 835–848. doi: 10.1111/j.1469-8137.2006.01704.x
Hedin, L. O., Vitousek, P. M., and Matson, P. A. (2003). Nutrient losses over four million years of tropical forest development. Ecology 84, 2231–2255. doi: 10.1890/02-4066
Hoch, G., and Körner, C. (2012). Global patterns of mobile carbon stores in trees at the high-elevation tree line. Glob. Ecol. Biogeogr. 21, 861–871. doi: 10.1111/j.1466-8238.2011.00731.x
Hoch, G., Popp, M., and Körner, C. (2002). Altitudinal increase of mobile carbon pools in Pinus cembra suggests sink limitation of growth at the Swiss treeline. Oikos 98, 361–374. doi: 10.1034/j.1600-0706.2002.980301.x
Körner, C. (1989). The nutritional status of plants from high altitudes. A worldwide comparison. Oecologia 81, 379–391. doi: 10.1007/BF00377088
Lin, L., Long, N. N., Qiu, M., Liu, Y., Sun, F. H., and Dai, M. (2021). The inhibitory efficiencies of geraniol as an anti-inflammatory, antioxidant, and antibacterial, natural agent against methicillin-resistant Staphylococcus aureus infection in vivo. Infect Drug Resist. 14, 2991–3000. doi: 10.2147/IDR.S318989
Long, N. N., Tang, H. Q., Lin, L., Sun, F. H., Peng, C., and Dai, M. (2020). Activity of amomum tasaoko fruits essential oil against methicillin-resistant Staphylococcus aureus in vivo. J. Altern Complement Integr. Med. 6:126. doi: 10.24966/ACIM-7562/100126)
Long, N. N., Zhang, Y. J., Qiu, M., Deng, J. Z., Sun, F. H., and Dai, M. (2022). Dynamic changes of inflammatory response and oxidative stress induced by methicillin-resistant Staphylococcus aureus in mice. Eur. J. Clin. Microbiol. Indect. Dis. 41, 79–86. doi: 10.1007/s10096-021-04349-5
Lord, J. M. (1994). Variation in Festuca novae-zelandiae (Hace.) Cockayne germination behaviour with altitude of seed source. New Zeal. J. Bot. 32, 227–235. doi: 10.1080/0028825X.1994.10410369
McCormack, M. L., Dickie, I. A., Eissenstat, D. M., Fahey, T., Fernandez, C. W., Guo, D. L., et al. (2015). Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 207, 505–518. doi: 10.1111/nph.13363
McGroddy, M. E., Daufresne, T., and Hedin, L. O. (2004). Scaling of C:N:P stoichiometry in forests worldwide: implications of terrestrial Redfield-type ratios. Ecology 85, 2390–2401. doi: 10.1890/03-0351
Millard, P., Sommerkorn, M., and Grelet, G. A. (2007). Environmental change and carbon limitation in trees: a biochemical, ecophysiological and ecosystem appraisal. New Phytol. 175, 11–28. doi: 10.1111/j.1469-8137.2007.02079.x
R Development Core Team. (2015). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna.
Reich, P. B., and Oleksyn, J. (2004). Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. U. S. A. 101, 11001–11006. doi: 10.1073/pnas.0403588101
Soethe, N., Lehmann, J., and Engels, C. (2008). Nutrient availability at different altitudes in a tropical montane forest in Ecuador. J. Trop. Ecol. 24, 397–406. doi: 10.1017/S026646740800504X
Soriano, D., Orozco-Segovia, A., Márquez-Guzmán, J., Kitajima, K., Gamboa-de Buen, A., and Huante, P. (2011). Seed reserve composition in 19 tree species of a tropical deciduous forest in Mexico and its relationship to seed germination and seedling growth. Ann. Bot. 107, 939–951. doi: 10.1093/aob/mcr041
Sterner, R. W., and Elser, J. J. (2002). Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere. Princeton, USA: Princeton University Press.
Sundqvist, M. K., Giesler, R., and Wardle, D. A. (2011). Within-and across-species responses of plant traits and litter decomposition to elevation across contrasting vegetation types in subarctic tundra. PLoS One 6:e27056. doi: 10.1371/journal.pone.0027056
Vitousek, P.M. (2004). Nutrient Cycling and Limitation: Hawai’i as a Model System. Princeton, USA: Princeton University Press.
Wang, Z. Q., Bu, H. Y., Wang, M. C., Huang, H., and Niklas, K. J. (2020b). Allocation strategies for seed nitrogen and phosphorus in an alpine meadow along an altitudinal gradient on the Tibetan Plateau. Front. Plant Sci. 11:614644. doi: 10.3389/fpls.2020.614644
Wang, Z. Q., Gong, H. Y., Sardans, S., Zhou, Q. P., Deng, J. M., Niklas, K. J., et al. (2022c). Divergent nitrogen and phosphorus allocation strategies in terrestrial plant leaves and fine roots: a global meta-analysis. J. Ecol. 110, 2745–2758. doi: 10.1111/1365-2745.13985
Wang, Z. Q., Huang, H., Li, X. W., Mao, K. S., Ran, J. Z., and Deng, J. M. (2018). Allocation of nitrogen and phosphorus within and between needles, stems and roots of Picea seedlings. Nord. J. Bot. 36:e01952. doi: 10.1111/njb.01952
Wang, Z. Q., Huang, H., Wang, H., Peñuelas, J., Sardans, J., Niinemets, Ü., et al. (2022a). Leaf water content contributes to global leaf trait relationships. Nat. Commun. 13:5525. doi: 10.1038/s41467-022-32784-1
Wang, Z. Q., Huang, H., Yao, B. Q., Deng, J. M., Ma, Z. Q., and Niklas, K. J. (2021a). Divergent scaling of fine-root nitrogen and phosphorus in different root diameters, orders and functional categories: a meta-analysis. For. Ecol. Manag. 495:119384. doi: 10.1016/j.foreco.2021.119384
Wang, Z. Q., Ji, M. F., Deng, J. M., Milne, R. I., Ran, J. Z., Zhang, Q., et al. (2015). A theoretical framework for whole-plant carbon assimilation efficiency based on metabolic scaling theroy: a test case using Picea seedlings. Tree Physiol. 35, 599–607. doi: 10.1093/treephys/tpv030
Wang, Z. Q., Lv, S. Q., Song, H., Wang, M. C., Zhao, Q., Hung, H., et al. (2020a). Plant type dominates fine-root C:N:P stoichiometry across China: a meta-analysis. J. Biogeogr. 47, 1019–1029. doi: 10.1111/jbi.13791
Wang, Z. Q., Niklas, K. J., Ma, Z. Q., Jiang, D. C., and Deng, J. M. (2022b). 2/3 scaling of twig nitrogen to phosphorus in woody plants. For. Ecosyst. 9:100049. doi: 10.1016/j.fecs.2022.100049
Wang, Z. Q., Wang, M. C., Yu, K. L., Hu, H. F., Yang, Y. H., Ciais, P., et al. (2021b). Global synthesis for the scaling of soil microbial nitrogen to phosphorus in terrestrial ecosystems. Environ. Res. Lett. 16:044034. doi: 10.1088/1748-9326/abed78
Wang, Z. Q., Yu, K. L., Lv, S. Q., Niklas, K. J., Mipam, T. D., Crowther, T. W., et al. (2019). The scaling of fine root nitrogen versus phosphorus in terrestrial plants: a global synthesis. Funct. Ecol. 33, 2081–2094. doi: 10.1111/1365-2435.13434
Wang, Z. Q., Zhao, M. Y., Yan, Z. B., Yang, Y. H., Niklas, K. J., Huang, H., et al. (2022d). Global patterns and predictors of soil microbial biomass carbon, nitrogen, and phosphorus in terrestrial ecosystem. Catena 211:106037. doi: 10.1016/j.catena.2022.106037
Wardle, D. A., and Greenfield, L. G. (1991). Release of mineral nitrogen from plant root nodules. Soil Biol. Biochem. 23, 827–832. doi: 10.1016/0038-0717(91)90093-Y
Weih, M., and Karlsson, P. S. (2001). Growth response of mountain birch to air and soil temperature: is increasing leaf-nitrogen content an acclimation to lower air temperature? New Phytol. 150, 147–155. doi: 10.1046/j.1469-8137.2001.00078.x
Yang, Y. H., Ji, C. J., Chen, L. Y., Ding, J. Z., Cheng, X. L., and Robinsone, D. (2015). Edaphic rather than climatic controls over 13C enrichment between soil and vegetation in alpine grasslands on the Tibetan plateau. Funct. Ecol. 29, 839–848. doi: 10.1111/1365-2435.12393
Yao, F. Y., Chen, Y. H., Yan, Z. B., Li, P., Han, W. X., and Fang, J. Y. (2015). Biogeographic patterns of structural traits and C: N: P stoichiometry of tree twigs in China’s forests. PLoS One 10:e0116391. doi: 10.1371/journal.pone.0116391
Yuan, Z., Chen, H. Y. H., and Reich, P. B. (2011). Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus. Nat. Commun. 2:344. doi: 10.1038/ncomms1346
Zhao, N., He, N. P., Wang, Q. F., Zhang, X. Y., Wang, R. L., Xu, Z. W., et al. (2014). Invariant allometric scaling of nitrogen and phosphorus in leaves, stem, and fine roots of woody plants along an altitudinal gradient. PLoS One 9:e95196. doi: 10.1371/journal.pone.0095196
Keywords: alpine meadow, carbon, nitrogen, phosphorus, seed, stoichiometry
Citation: Jia N, Niklas KJ, Yao B and Wang Z (2022) Altitude patterns of seed C, N, and P concentrations and their stoichiometry in an alpine meadow on the eastern Tibetan Plateau. Front. Ecol. Evol. 10:1093474. doi: 10.3389/fevo.2022.1093474
Edited by:
Mauro Fois, University of Cagliari, ItalyReviewed by:
Megan McGroddy, Adnet Systems, United StatesCopyright © 2022 Jia, Niklas, Yao and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zhiqiang Wang, d2FuZ3pxQHN3dW4uZWR1LmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.