 William Rendu1,2*
William Rendu1,2* Sylvain Renou3
Sylvain Renou3 Anastasiia Koliasnikova1
Anastasiia Koliasnikova1 Malvina Baumann4,5
Malvina Baumann4,5 Hugues Plisson5
Hugues Plisson5 Emmanuel Discamps6
Emmanuel Discamps6 Marie-Cécile Soulier6
Marie-Cécile Soulier6 Arthur Gicqueau1,6
Arthur Gicqueau1,6 Mathilde Augoyard5
Mathilde Augoyard5 Manon Bocquel5
Manon Bocquel5 Guillaume Guerin7
Guillaume Guerin7 Svetlana Shnaider1*
Svetlana Shnaider1* Kseniya Kolobova8
Kseniya Kolobova8- 1ZooSCAn (IRL 2013) CNRS - IAET SB RAS, Novosibirsk, Russia
- 2Institut français d'études sur l'Asie centrale (IFEAC), UAR3140 (CNRS), Bishkek, Kyrgyzstan
- 3HADES, Agence Atlantique, Bordeaux, France
- 4TRACEOLAB, University of Liège, Liège, Belgium
- 5UMR5199 De la Prehistoire A l'actuel Culture, Environnement et Anthropologie (PACEA), Pessac, Aquitaine, France
- 6UMR5608 Travaux de Recherches Archéologiques sur les Cultures, les Espaces et les Sociétés (TRACES), Toulouse, Midi-Pyrénées, France
- 7UMR6118 Geosciences Rennes, Rennes, Brittany, France
- 8Institute of Archaeology and Ethnography (RAS), Novosibirsk, Novosibirsk Oblast, Russia
During the MIS 4 in Southwestern France, Quina Neanderthal from the north of the Aquitaine was characterized by a hunting specialization on the reindeer and the lack of diversity in their diet. They developed task-specific locations dedicated to the capture, the butchery, and the consumption of reindeer, and the whole society seems, in this region, to be dependent on this food resource. In this context, the site of Chez-Pinaud at Jonzac (France) occupies a specific place. First, interpreted as a reindeer kill and butchery site, the recent recovery of the site underlines the importance of the large ungulate (horse and bison) to the faunal spectrum (30% of the NISP). Considering the quantity of meat and grease that these species can provide to hunters, the new zooarchaeological analyses suggest that at least the horse may have played a major role in the diet of the Neanderthal population. Since Jonzac is one of the largest sites for this period, these results relativize the importance of reindeer specialization of the Quina population and the lack of diversityl in their diet.
Introduction
In the last decades, considerable input on Neanderthal subsistence has been acquired on a particular period of the Middle Paleolithic record, the Quina Mousterian (Discamps and Royer, Costamagno et al., 2006). In southwestern France, the Quina Mousterian techno-complex, dated from the MIS 4 or early MIS 3 (Figure 1), is notably characterized by its recurrent association with reindeer hunting (Discamps and Royer, 2017; Rendu et al., 2022). In fact, out of the 32 Quina stratigraphic units that yielded sufficiently large faunal assemblages (i.e., with a total NISP of ungulates >100), 28 are dominated by the arctic deer's remains.
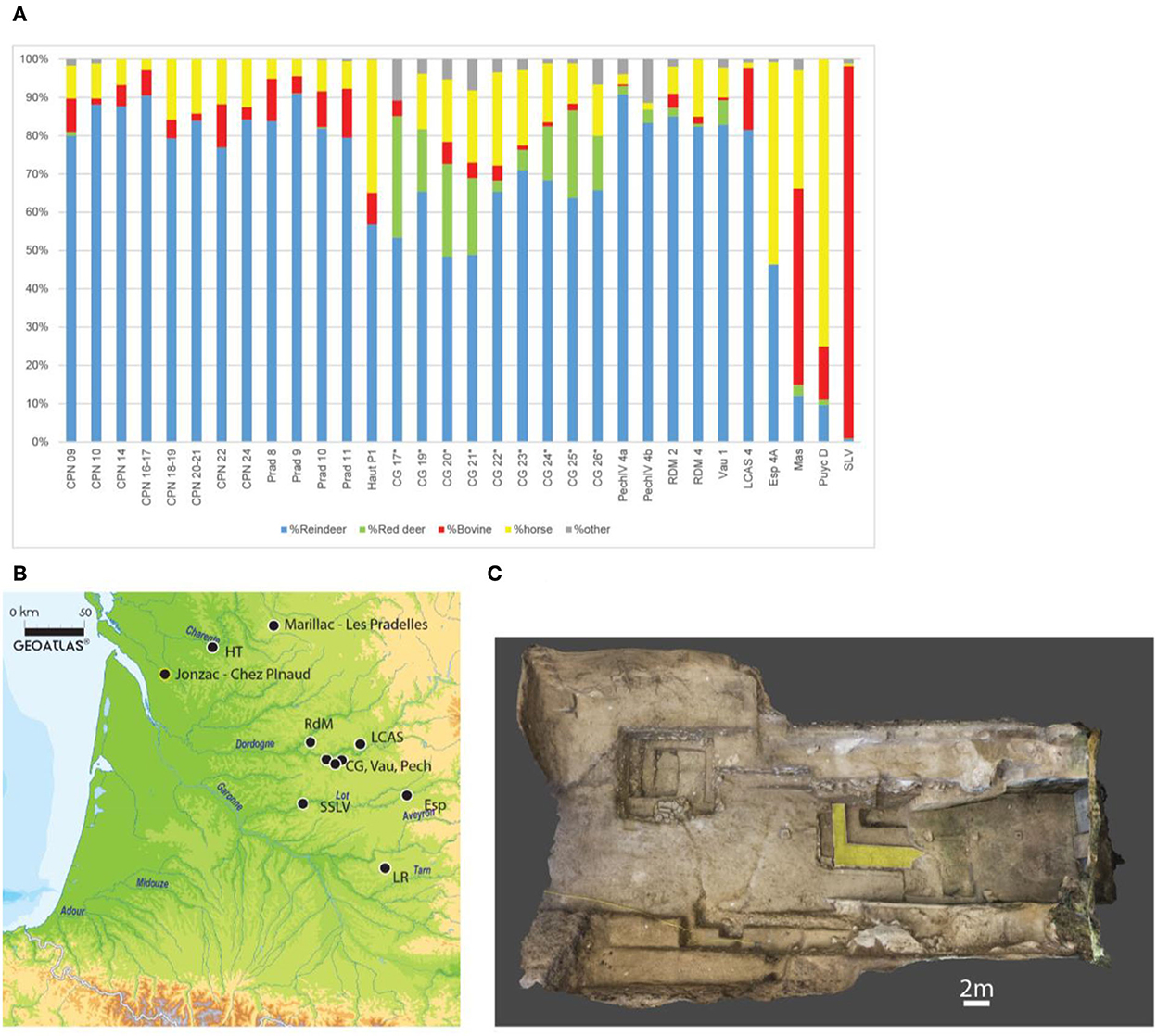
Figure 1. (A, B) Distribution of the main Quina sites yielding faunal assemblages in Southwestern France, and contribution of the main prey to the faunal spectra (in %NR; Blue, reindeer; green, red deer; Red, Bison; Yellow, Horse; Gray, other). (A) CG, Combe Grenal (Laquay, 1981; Guadelli, 1987; Vau, Vauffrey (Delpech, 1996); PechIV, Pech de l'Azé IV (Niven, 2013); RdM, Roc de Marsal (Castel et al., 2017); LP, Les Pradelles (Costamagno et al., 2006); CPN, Chez-Pinaud-Jonzac (Airvaux, 2004; Jaubert et al., 2008b; Niven, 2013); LCAS, La Chapelle-aux-Saints (Rendu et al., 2014); Haut, Hauteroche (Paletta, 2005); LR, La Rouquette (Rendu et al., 2011); SLV, Sous les Vignes (Turq et al., 1999); ESP, Espagnac (Jaubert, 2001). Numbers correspond to the different stratigraphic units. For Combe Grenal Reindeer was under-evaluated in the previous excavation due to selective sampling. Derived from Discamps and Royer, 2017. Map from Geoatlas. (C) Orthophotography of the site, extracted from the 3D model. In yellow, the 2019–2021 excavation area.
The probable abundance of reindeer in the environment during the Quina has been purportedly correlated to the major climatic pejoration of the Heinrich Stadial 6 (Discamps et al., 2011; Discamps and Royer, 2017). During this period, several indicators also point to a major drop in the ungulate biomass available for large game hunters (Discamps, 2014). In addition, sedentary prey that was present just before this event was replaced by reindeer, identified as a migratory species at that time (Britton et al., 2011).
This over-representation of reindeer in faunal spectra and the development of different task-specific locations (Binford, 1980) dedicated to subsistence activities [kill and first butchery sites (Niven et al., 2012) and secondary butchery sites (Costamagno et al., 2006)] conducted scholars to propose the strong dependence of Quina groups on reindeer, which would have occupied a central place in the Neanderthal diet at the time (Delagnes and Rendu, 2011; Rendu et al., 2022).
In this model, the Chez-Pinaud site at Jonzac plays a major role along with Les Pradelles (Costamagno et al., 2006), one of the best examples of these task-specific locations dedicated to the exploitation of reindeer herds (Niven et al., 2012; Rendu et al., 2022). We here report on new zooarchaeological data acquired on this key site, concerning notably the importance of reindeer and other prey in Neanderthal diets throughout the Quina period.
Material: The Chez-Pinaud Jonzac site
Excavated between 1999 and 2002 by a team led by Airvaux (Airvaux, 2004), and between 2004 and 2007 by Jaubert, Hublin, et al. (Jaubert et al., 2008a), Chez-Pinaud is situated at the bottom of a 10-m cliff. More than 24 stratigraphic units were identified, eight of them yielding artifacts attributed to the Quina Mousterian (Airvaux, 2004; Jaubert et al., 2008a; Niven et al., 2012).
Among them, Stratigraphic Unit (SU) 22 is a 1-m thick bone bed with excellent preservation of the bones and their spatial distribution (Jaubert et al., 2008a; Niven et al., 2012). During the Jaubert-Hublin excavations (2004–2007), a significant number of anatomical articulations were uncovered identifying “snapshots” on-site, allowing for discussing the carcass processing and the organization of activities within a specialized site dedicated to predation with high resolution (Rendu et al., 2022). Since 2019, the site is under a new excavation program focusing on the US22 bone bed directed by the CNRS, the IAET SB RAS, and Bordeaux University (dir. W. Rendu, K. Kolobova, and S. Shnaider). The excavation area covers a surface of ~8 m2. We applied the common “decapage” method consisting of removing the sediment over the complete excavated area without moving the artifacts to have a better view of their relative organization. Each decapage is followed by a photogrammetric model of the whole surface using a Canon EOS 600 D. Raw pictures were processed through Agisoft Metashape software to obtain a 3D model of the excavated area. Artifacts (lithics bigger than 1 cm and all the identifiable faunal remains or remains larger than 2 cm) were then piece-plotted using a NikonNivo total station. The different analytical databases were linked to the 3D model using ArcGIS for studying spatial distribution. In total, nine decapages (C1–C9) were realized between 2019 and 2021 by conducting the collection of almost 5,000 faunal remains (including 4% of teeth remains) and 2,000 lithics.
All the faunal remains were identified on the site (with the help of the portable comparative collection), and potential anatomical articulations were looked for by two zooarchaeologists (WR, SR) during the excavation and before any collecting session. Numerous anatomical articulations were identified during the excavations (NR = 46), some of them imply several bones such as a complete reindeer carp in articulation with its radio-ulna and metacarpal (Figure 2).

Figure 2. Remains 6,218–6,225 in anatomical articulation found during decapage 8.
Zooarchaeological data
Previous zooarchaeological analyses demonstrate that reindeer, which dominate largely the assemblage (>80% of total NISP, Supplementary Table 1), were killed in the direct vicinity of the site and partially processed there before exportation toward a secondary consumption camp (Beauval, 2004; Niven et al., 2012). The 18 individuals, including males, females, and juveniles (based on a dental MNI), present a catastrophic mortality profile. Based on cementum increment analyses, tooth eruption sequences, and fetal bone abundances, the exploitation of the site in winter has been proposed (Niven et al., 2012; Rendu et al., 2022).
However, while the zooarchaeological analyses of the Jaubert and Hublin collection brought significant information about reindeer exploitation, largely used to discuss the Quina diet (Discamps and Royer, 2017; Faivre et al., 2017; Rendu et al., 2022), the exploitation of large ungulates was not described in detail. Throughout, C. Beauval (Beauval, 2004) demonstrated on the large collection of Airvaux excavation (Airvaux, 2004) the fluctuation of the relative contribution of horse and bison through the thickness of US 22.
The exact place of large ungulates in the subsistence of Quina Neanderthals has been left mostly undiscussed. The importance of large ungulates might have been underestimated in previous studies, notably if one considers that their carcasses provide between 3 and 5 times more food than a reindeer carcass. At a regional scale, if we consider that Chez-Pinaud has been a central piece in our perception of the Quina diet, new results could lead us to ponder or even change our perception of the Neanderthal diet and subsistence at the time. Based on the material from the new excavation and our high-resolution control on the field, we propose to evaluate how a better inclusion of large ungulates in zooarchaeological interpretation and horse, in particular, can modify our perception of the use of the Chez-Pinaud site and the diet of the Quina population.
Methods
All the faunal remains were observed. For taxonomic and anatomical identifications, we used the reference skeletal collections from the IRL 2013 ZoSCAn (CNRS—IAET SB RAS) and the one from PACEA Laboratory (CNRS—Bordeaux University—MCC), sometimes complemented by the Virtual Faunal Comparative Collection from the Max Planck Institute (Niven et al., 2009). Pieces were identified at the most precise level and, when it was not possible to propose a specific attribution, ungulate size classes were used (adapted from Brain, 1981). With regard to the skeletal part profiles, all identifiable specimens (including shaft fragments) were taken into account and recorded following the “element, portion, segment” method (Gifford and Crader, 1977). Shaft fragmentation was evaluated using the shaft length and shaft circumference indexes (Villa and Mahieu, 1991). Analyses of the bone surfaces were conducted on all the identified remains and part of the non-identified ones. The bone surfaces were observed under a low-angled light systematically using a hand lens (enlargement: 20x) for the taphonomic and zooarchaeological observations. Weathering, root etching, and anthropogenic and carnivore modifications were systematically looked for (Behrensmeyer, 1978; Olsen and Shipman, 1988; Blumenschine et al., 1996; D'Errico and Villa, 1997; Pickering and Egeland, 2006). Oxide colorations of the bone cortical surfaces were also recorded. The proportion of preserved cortical surface was estimated per quartile (Rendu, 2010). When unclear modifications were detected, specimens were subjected to a more thorough evaluation with a 20–80x microscope. Percentage values were calculated based on the number of analyzed remains (NRa). Bones with unobservable surfaces were excluded from the calculation of the percentages of modified bones, thus NRa can change depending on the analysis type. Skeletal part representations were established for the reindeer and the horse using both %NNISP (Grayson and Frey, 2004) and %MAU index (Binford, 1978, 1981). Differential preservation has been tested for the reindeer and the horse by confronting frequencies of skeletal elements (in %NNISP) and their respective densities (Lyman, 1994; Lam et al., 1999). The possibility of a selective transport based on the nutritive value of the elements was tested using the SFUI (Metcalfe and Jones, 1988; Outram and Rowley-Conwy, 1998). Statistical tests of correlation (Spearman's rank rs) and Fisher exact tests were performed using the R stats package, and 95% confidence intervals for percentages were calculated using the BinomCI function of the R DescTools package using the Wilson method. Plots were carried out in QGIS (QGIS Development Team, 2022).
Results
Faunal spectrum
Due to good bone preservation (see infra) and a limited number of taxa identified in the faunal spectrum (7 species), 65% of the remains were taxonomically identified (Table 1). As expected, reindeer dominates largely the assemblages (76%), followed by horses (16%) and large bovids (8%). Red deer, leporids, foxes, and wolves complete the faunal spectrum.
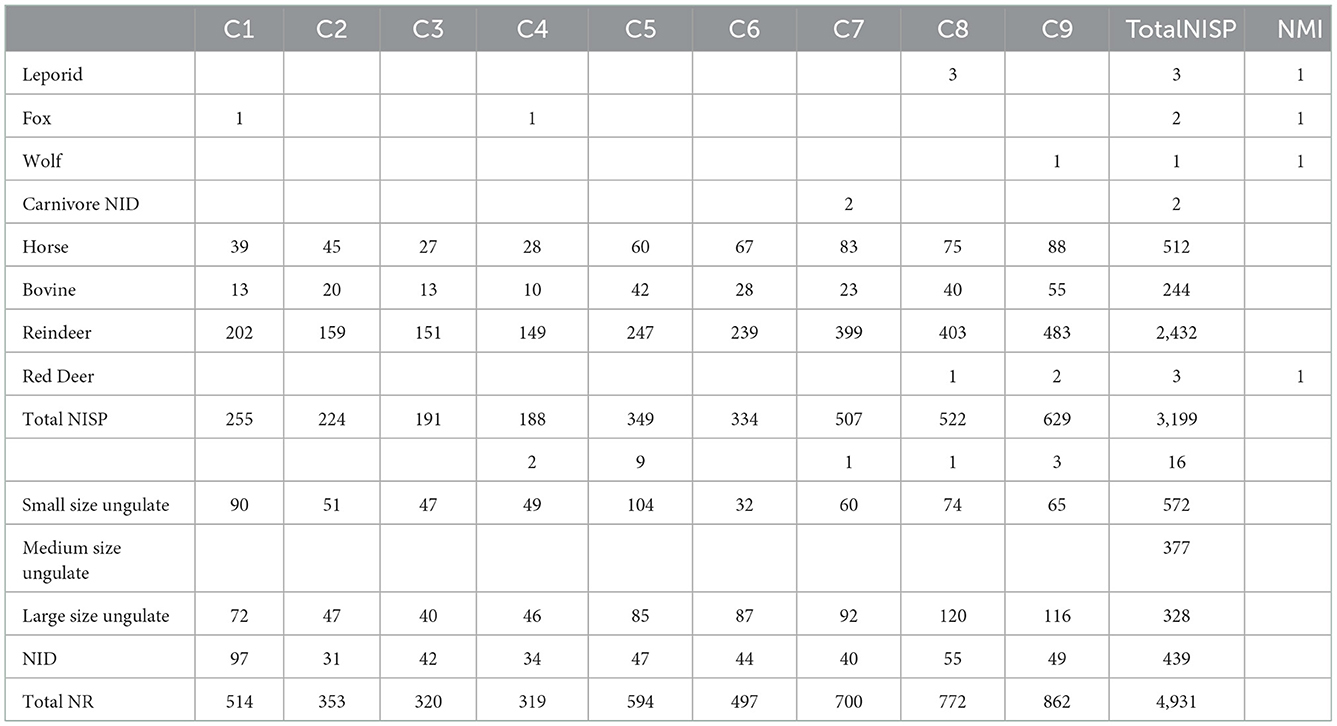
Table 1. Fauna spectrum per decapage (C1 to C9) expressed in number of remains (NR). MNIc was calculated on the combination of bone and teeth.
An evolution through the deposit can be observed with, in particular, the fluctuation of the Bovinae contribution as identified by C. Beauval on the Airvaux collections (Beauval, 2004). Fisher exact tests identify differences between the C4 and C5 decapages, and between the C6 and C7 decapages (Figure 3). The visualization of these fluctuations with confidence intervals (Figure 3), however, ponders these rather small differences between the decapages.
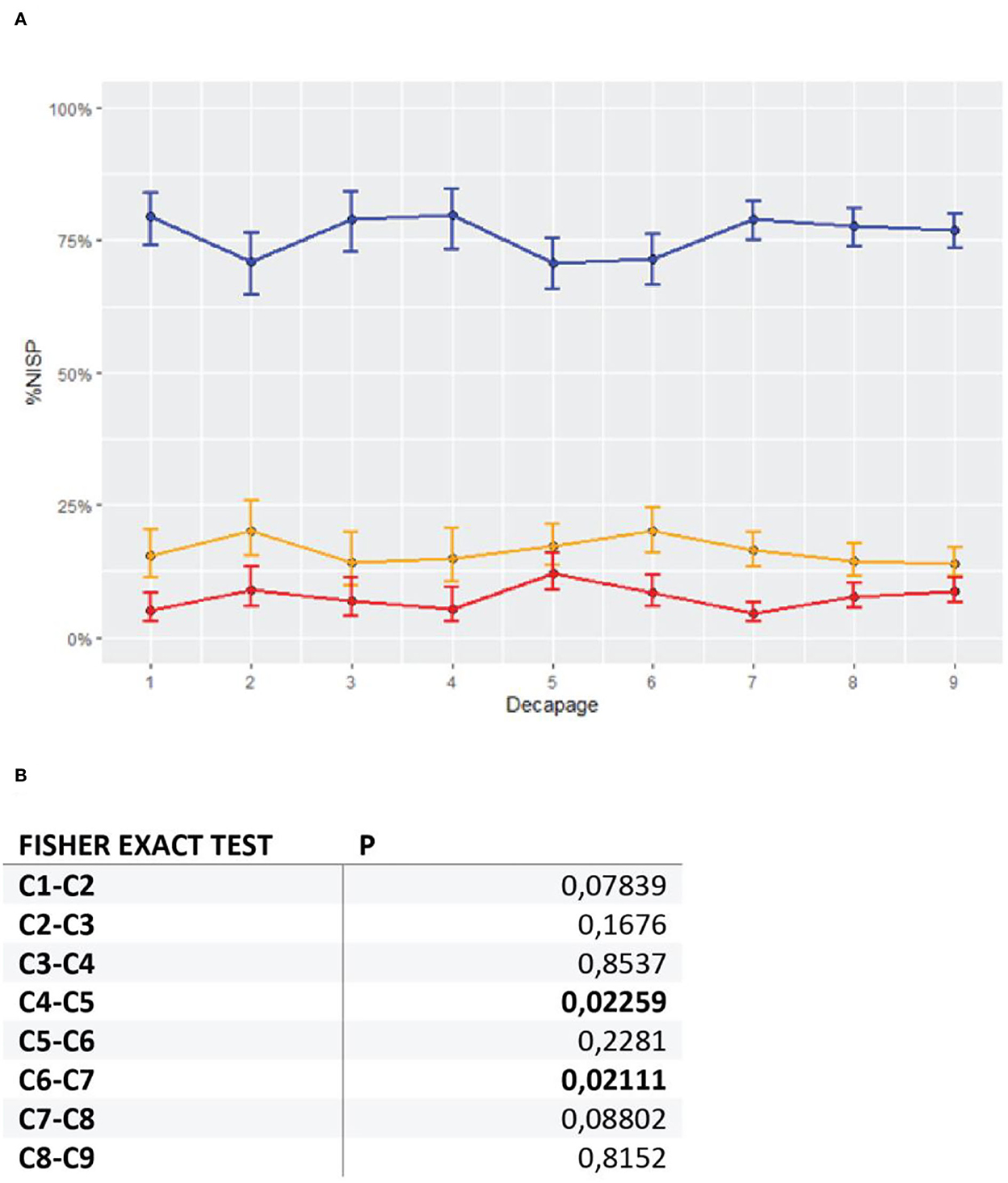
Figure 3. (A) Variations in Reindeer (blue), Bovine (red), and Horse (yellow) %NISP per decapage, with 95% confidence intervals; (B) Paired Fisher exact tests by decapage, performed on NISP of reindeer, large bovids, and horses. Statistically significant differences in values are highlighted in bold.
The contribution of the horse is however significantly higher in our assemblage than what Niven et al. (Niven et al., 2012) identified (16% of the NISP vs. 9%; Fisher exact test: p < 0,001), but closer to Beauval results (11.67% NISP; Beauval, 2004). If the MNI is taken into account, this trend is slightly less pronounced with reindeer (MNI = 28) dominating the assemblage followed by horse (MNI = 8) and bovinae (MNI = 4).
A detailed spatial analysis of the Chez-Pinaud dataset will be carried out in the future, but Figure 4 proposes a first general overview of the distribution and density of the three main taxa identified: no specific clustering by species is apparent. Remains do not tend to be clustered by species but, rather, mixed all together.
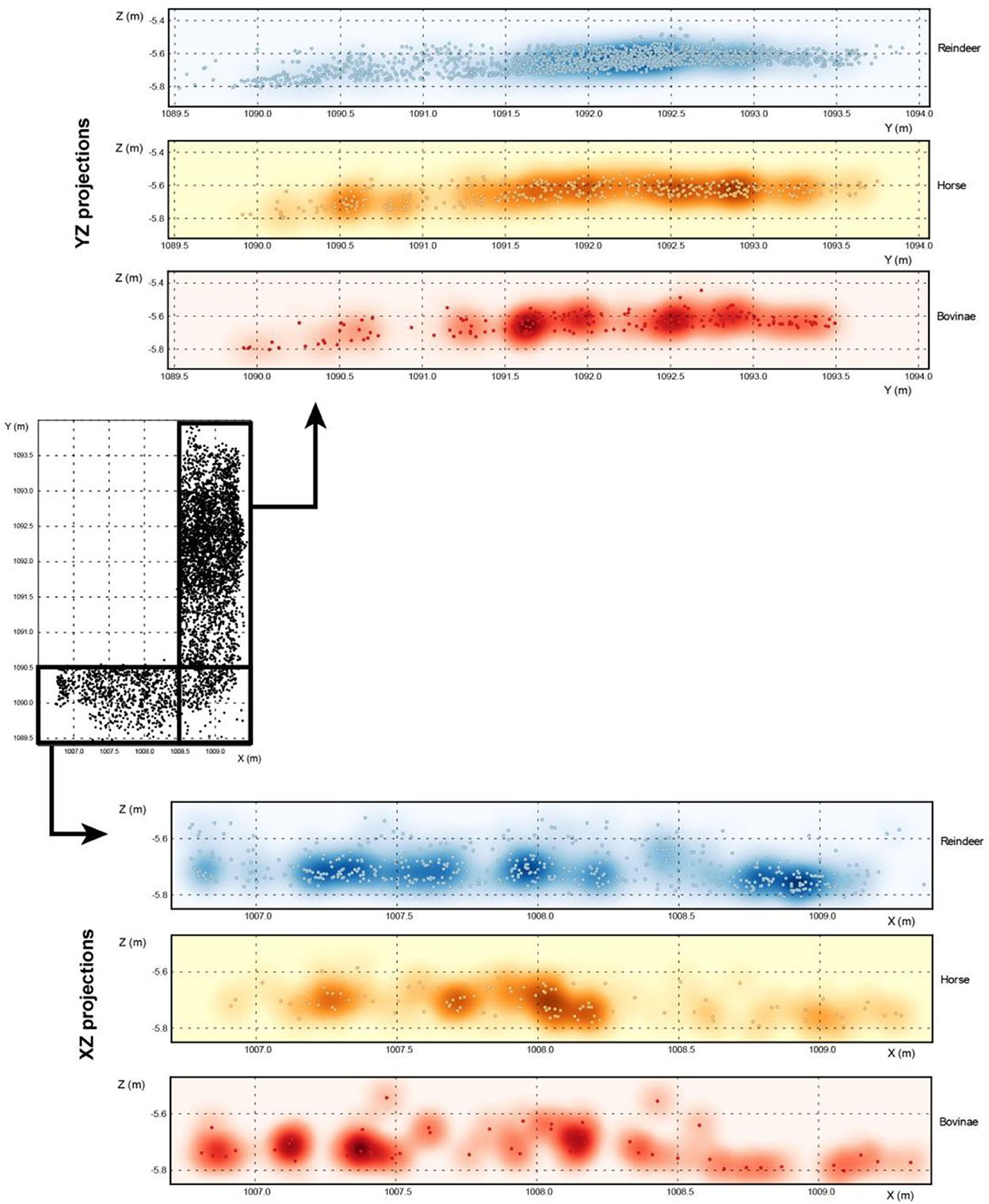
Figure 4. Spatial distribution of the three main taxa on sagittal (YZ) and frontal (XZ) projections (dots: identified remains, color background: density).
The comparison of ungulate size classes allows to overcome the greater difficulty of horse and bison identification. Indeed, the reindeer is almost the only member of the medium size ungulate category creating a bias in its advantage and leading to an overestimation of its remains and a tendency to attribute the anatomically identified remains of this category to this taxon. Table 2 clearly underlines that reindeer is overestimated in the faunal spectrum and that the large ungulates contribute to about one-third of the assemblage. This point is crucial: it tempers the presentation generally made of the deposit as being specialized [sensu (Mellars, 2004)] on reindeer exploitation (Jaubert et al., 2008a; Delagnes and Rendu, 2011; Niven et al., 2012; Rendu et al., 2022).
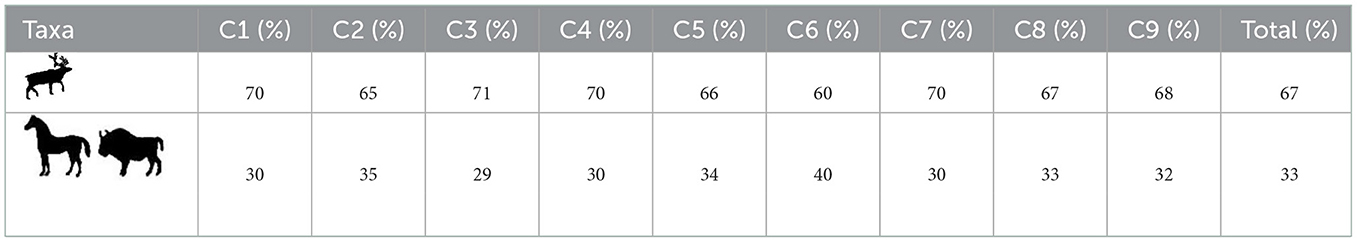
Table 2. Relative contribution of the different ungulate size classes to the Fauna spectrum per decapage (C1 to C9).
Taphonomy
The faunal stock is globally well preserved. The impact of weathering is particularly limited in intensity but not in frequency (Table 3). Nearly half of the material was affected by these changes. On the other hand, the advanced stages (stage 3 and stage 4) are visible on < 8% of the total number of bones. This supports the idea of rapid burial of the remains.
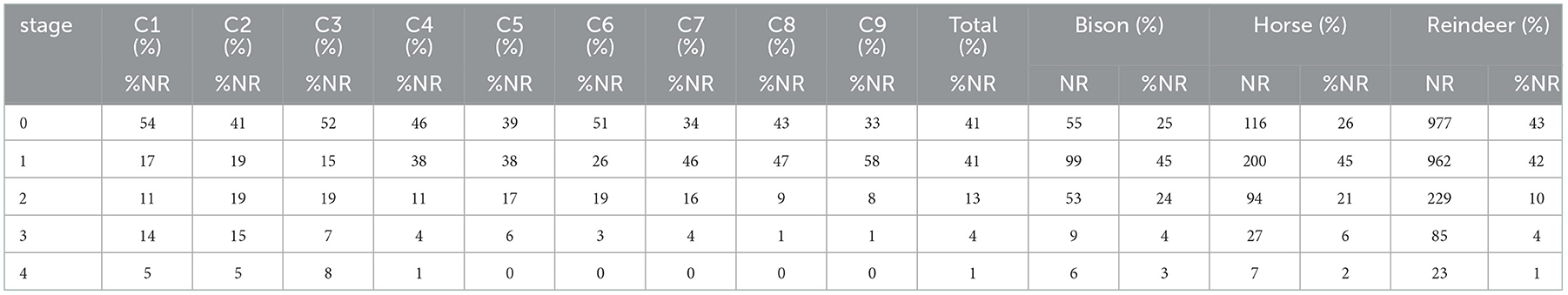
Table 3. Impact of the weathering (C1 to C9) on the three main taxa.
While there is no significant variation between bison and horse remains, the reindeer appears to be less affected by the weathering.
In detail (Supplementary Table 2), it is mainly the longitudinal cracking of the bones or cracks that are visible on the material with the exfoliations that correspond to the detachment of the outermost cortical layers. They are generally found on the same remains. The cracks within the thickness of the bones are rare. Since this taphonomic attack is generally associated with freeze alternations (Guadelli and Ozouf, 1994), and considering that the deposition took place in a peri-arctic climate (Jaubert et al., 2008b), it is possible that its low representation of frost modifications attests that the bone bed remained frozen most of the time (or at least did not undergo much freeze/thaw alternations) before its complete embedding.
Once again, it is possible to see a difference between the reindeer and the two large ungulates (Supplementary Table 3).
These different modifications had a limited impact on the preservation of the cortical surface of the bone. Indeed, more than 80% of the remains show preservation of at least 50% of their cortical surfaces (Figure 5), but reindeer remains appear to have been more altered.
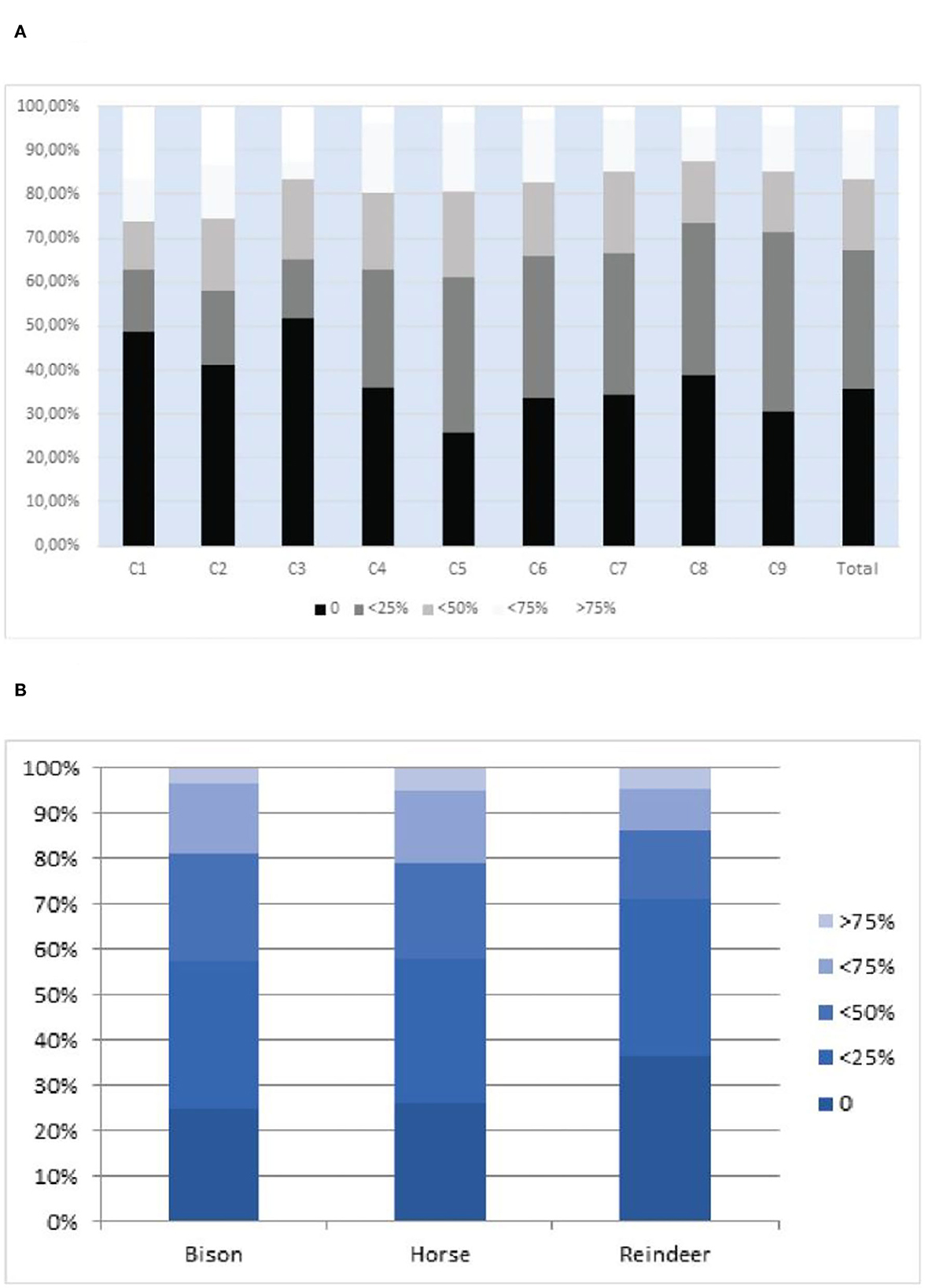
Figure 5. (A) Preservation of the cortical surfaces (per quartile) following (Rendu, 2010; Rendu et al., 2019): Stage 0: no destruction; Stage1: < 25% destroyed; stage2: < 50%; stage 3: < 75%; Stage 4: >75% destroyed. (B) Preservation of the cortical surfaces for the three main taxa, (per quartile) following (Rendu, 2010; Rendu et al., 2019) (Stage 0: no destruction; Stage1: < 25% destroyed; stage2: < 50%; stage 3: < 75%; Stage 4: >75% destroyed.
The carnivore damages are almost absent from the assemblage and affect only 1.2% of the remains. This low carnivore impact is coherent with what was observed previously in the Airvaux and Jaubert and Hublin collections (Beauval, 2004; Niven et al., 2012). Associated with their very limited presence in the faunal spectrum, their low impact strongly suggests that they had no major influence on bone accumulations. In addition, cut marks on the remains of the three carnivore taxa identified in the different collections evidence their exploitation by Neanderthals [a fox tibia (Niven et al., 2012); metacarpals of cave lion (Beauval, 2004); and a wolf tibia (this study)].
On the contrary, the human impact on the collection is particularly pronounced (Table 4), affecting 29% of the analyzed remains (Table 4).
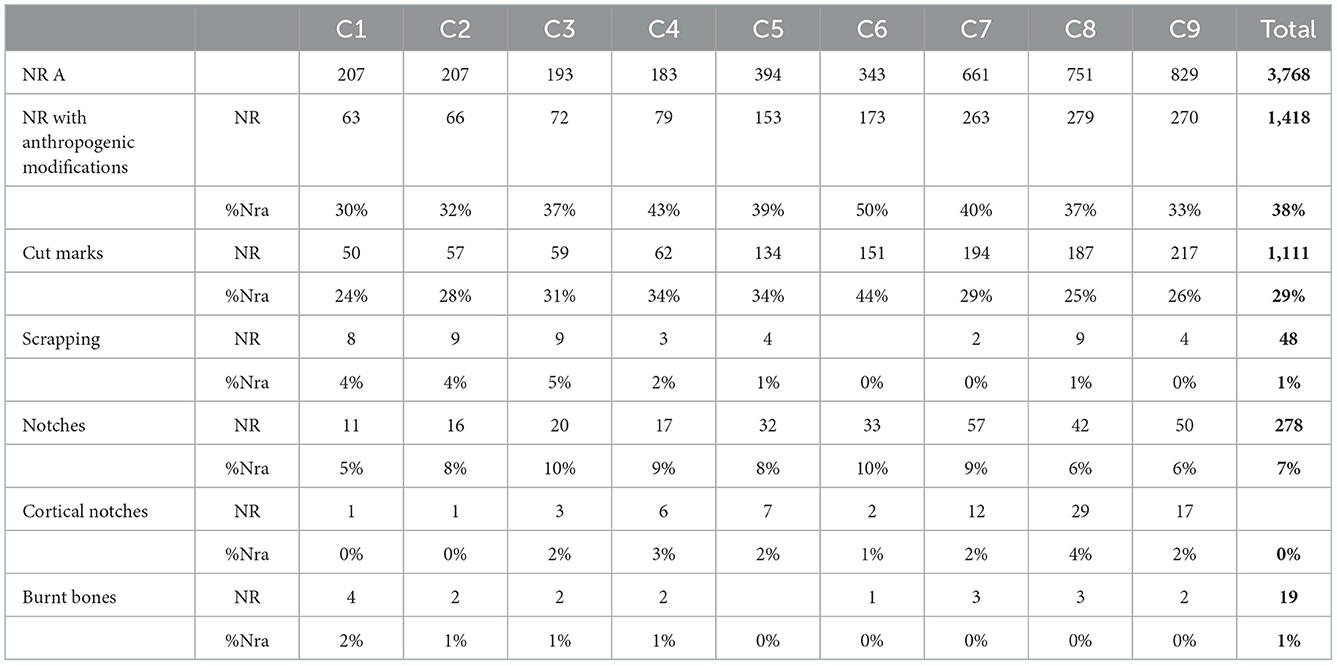
Table 4. Anthropogenic modifications. NRA: Number of analyzed remains.
The anthropogenic modifications affect all the taxa with the exception of the leporid (Table 5). If we did not identify any modification on the fox remains, Niven and collaborators identified cut marks on a distal extremity of a tibia (Niven et al., 2012). In addition, a longitudinal cut mark on a shaft fragment of a wolf tibia attests to defleshing activities on carnivores.
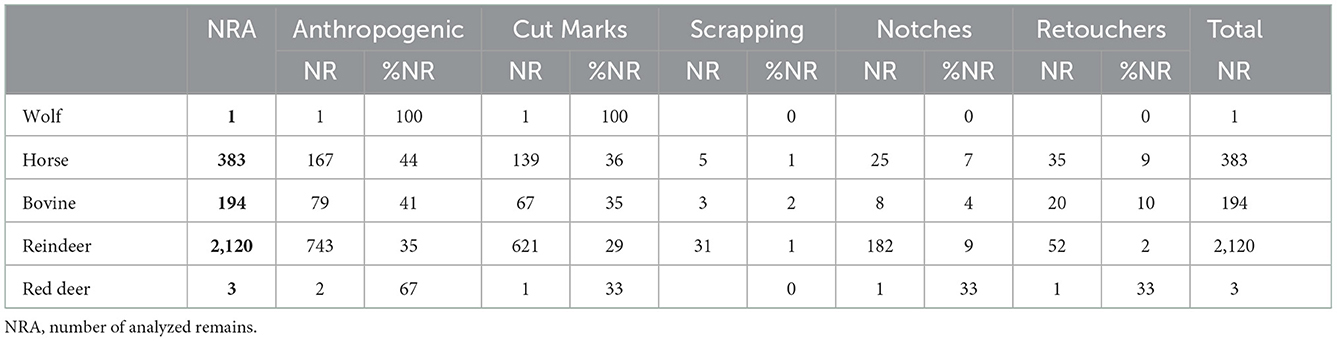
Table 5. Anthropogenic modifications par taxa and their details.
The very low quantity of burnt bones was already highlighted in the previous analysis (Niven et al., 2012) and is something very common in most Quina contexts, such as Les Pradelles or Roc de Marsal, for instance, where their frequency is largely below 1% (Costamagno et al., 2006; Castel et al., 2017).
The skeletal profiles
The post-depositional fragmentation of the assemblage is limited and 91% of identified breaks were realized on green bone. In addition, more than 5% of the remains (teeth excluded) are found complete, including some of the ribs. This limited fragmentation is also highlighted by the relatively high frequency of long bones extremities: they represent more than 13% (217/1709) of the long bones NISP, though they are known to be usually under-represented on archaeological sites due to preservation issues or specific human or carnivore exploitation (Lyman, 1994; Marean and Assefa, 1999). For instance, at the contemporaneous Les Pradelles Quina site, their proportion is under 1% and it is interpreted as resulting from the destruction of these extremities by humans for recovering the grease within (Costamagno et al., 2006).
Skeletal profiles have been established for the reindeer and horse (Figure 6), and they show two different patterns: while all the reindeer skeletal parts are found at the site, the horse skeletal profile shows a greater discrepancy between meaty long bones and head than axial skeleton and griddles. Also, on the horse, it is worth noting the quasi-absence of the lower-leg elements.
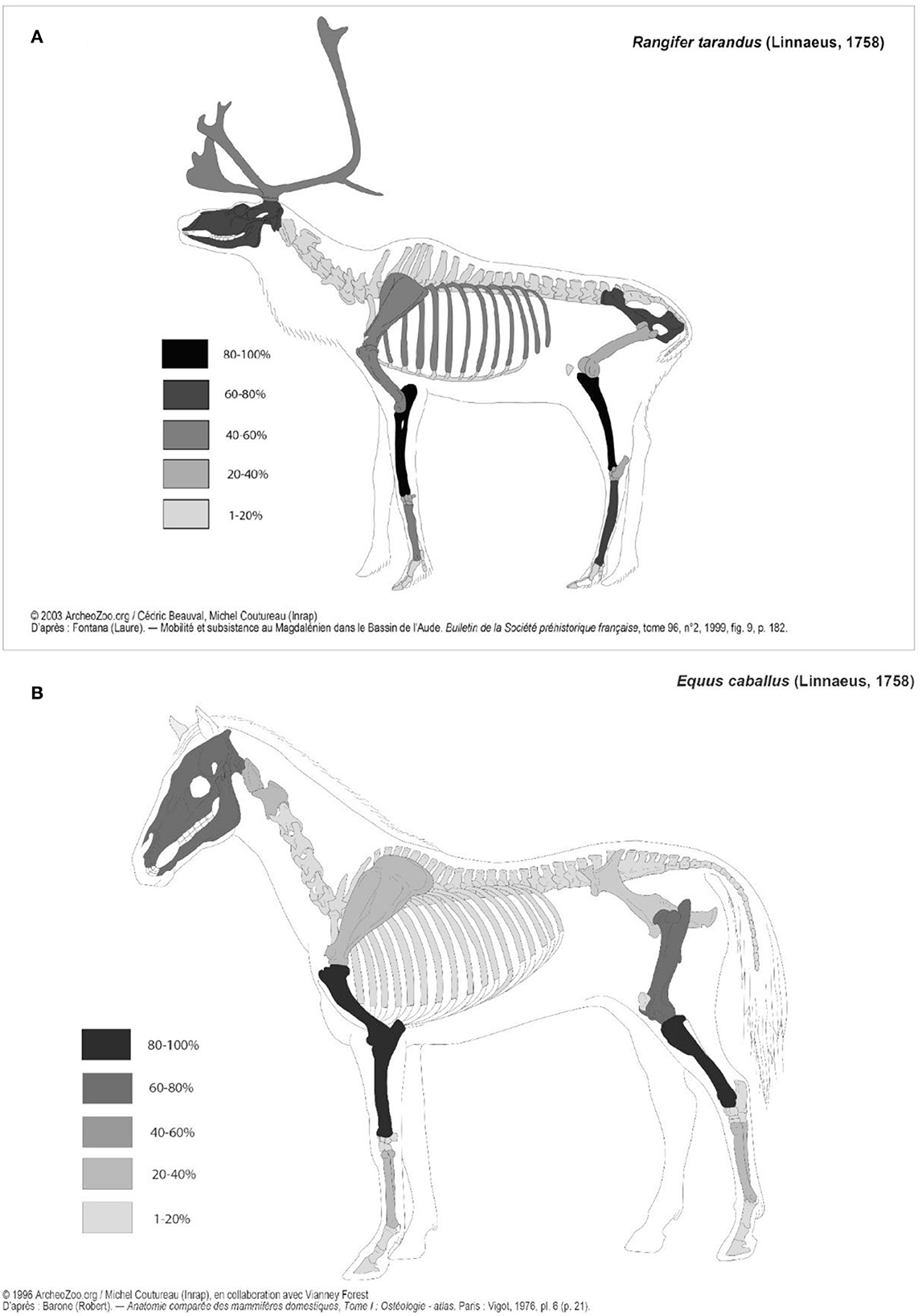
Figure 6. (A) Reindeer skeletal part representation of Chez-Pinaud horse remains expressed in %MAU, image modified from © 2003 ArcheoZoo.org / Cédric Beauval, Michel Coutureau (Inrap) D'après : Fontana (Laure). — Mobilité et subsistance au Magdalénien dans le Bassin de l'Aude. Bulletin de la Société préhistorique française, tome 96, n°2, 1999, fig. 9, p. 182. (B) Horse skeletal part representation of Chez-Pinaud horse remains expressed in %MAU, Modified from © 1996 ArcheoZoo.org / Michel Coutureau (Inrap), en collaboration avec Vianney Forest D'après : Barone (Robert). — Anatomie comparée des mammifères domestiques, Tome I : Ostéologie - atlas. Paris: Vigot, 1976, pl. 6 (p. 21).
As highlighted previously by Niven et al. (2012), there is a weak but significant correlation (rs = 0.4343, p < 0.001; ddl = 46, Figure 7) between the frequency of the reindeer skeletal elements and their relative density (Lyman, 1994; Lam et al., 1999) while none exists for the horse elements (rs = 0.0181; ddl = 46, Figure 7). This taxonomic difference in the preservation of the bone finds an interesting echo with the difference in the preservation of the cortical surfaces (cf. supra). It suggests that the existence of density-mediated destruction has influenced the skeletal part profile of the reindeer (such as proposed by Niven et al., 2012) but cannot explain the variations observed on the horse skeletal profile.
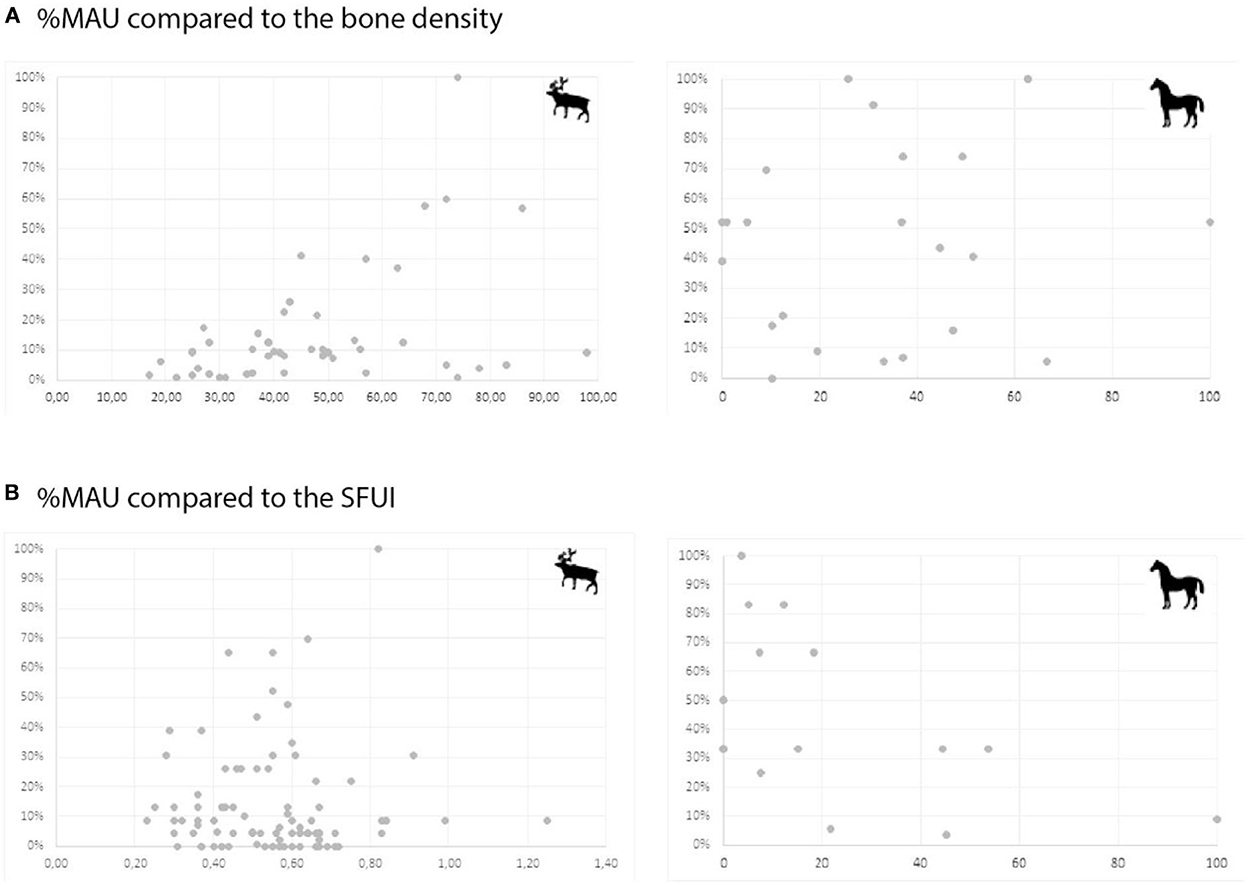
Figure 7. (A) Differential preservation test: for Reindeer (left) and horse (Right) relative skeletal representation (%NNISP) compared to the bone density (Lam et al., 1999). (B) reindeer (left) and horse (Right) skeletal part representation (y) [in %MAU see Binford (1978, 1981)] compared to their nutritive value (x) expressed in SFUI (Outram and Rowley-Conwy, 1998).
Simultaneously, there is no correlation (rs = 0.093) between the frequency of the reindeer skeletal elements (Supplementary Table 4) and their nutritive values expressed in SFUI (Metcalfe and Jones, 1988), as illustrated by Figure 7. This attests to the absence of evidence of selective transport by the human population for the cervid and confirms the results of Niven and collaborators, who concluded that the whole carcasses were brought to the site (Niven et al., 2012).
The comparison of the horse skeletal part representation (Supplementary Table 5) with the Standardized Food Utility Index [SFUI; Figure 7, (Outram and Rowley-Conwy, 1998)] underlines a statistically significant negative correlation (rs = −0.54033; DDL = 14; p < 0.05). In other words, the poorest an element is, the more common it is in the assemblage. It would attest to the selective exportation of the richest skeletal parts of the horse carcasses to a consumption site and the discarding at Chez-Pinaud of the less interesting parts. Figure 7 underlines the existence of a gourmet strategy (Binford, 1981), which could explain the relatively weak correlation.
The site would have been used as an acquisition site for horse raw material, confirming a task-specific location dedicated to hunting activities (Jaubert et al., 2008a; Delagnes and Rendu, 2011; Niven et al., 2012).
Exploitation of the horse
Horse carcasses were intensively exploited during the Quina occupation. Indeed, 44% of the remains exhibit evidence of anthropogenic modifications, mostly cut marks (39% of the total remains, 43% of the remain with good preservation, and < 50% of the cortical surfaces destroyed) being largely more frequent than the exploitation marks observable on the reindeer remains. This difference is statistically highly significant (Khi2= 10.262, ddl = 1; p < 0.01).
The distribution of the cut marks on the horse skeleton attests to skinning, dismembering, and defleshing activities (Soulier and Costamagno, 2017; Soulier et al., 2022).
Skinning activities are identified for now by only circular marks at the base of two metatarsals, while this activity was documented on the reindeer elements (Beauval, 2004; Jaubert et al., 2008a; Claud et al., 2012; Niven et al., 2012).
The rib dismembering shows an interesting pattern: 9 out of the 17 articular heads exhibit the same transversal repetitive short disarticulations marks. The fact that these rib heads may come from the same individual and bear traces of the same gesture cannot be excluded. The two observable atlases attest to their separation from the cranium.
The defleshing activities are well identified on the collection, with at least 39 occurrences out of the 383 remains (teeth excluded; out of 247 remains if only the remains with very good preservation are considered [ < 25% of the cortical surface is altered)]. These occurrences are preferentially found on the axial skeleton (15 ribs and 7 vertebrae) compared to the long bone remains (NISP = 12), while these elements are more frequent. Associated with the exportation of the elements rich in meat, it shows the strong interest of the Quina Neanderthal from Chez-Pinaud for horsemeat. Two occurrences of tongue extraction have also been identified by cut marks on the inner part of mandibles. In addition, the gutting is attested by several cut marks in the inner part of the six ribs.
Notches have been recorded on 29% (22/74 NISP) of the long-bone shaft fragments, evidencing the intense long-bone breakage to recover the grease and marrow and probably also to extract blank for the bone tool industry (see below). It is noteworthy that horse bones are more difficult to break than reindeer bones and they contain proportionally less marrow because of the large amount of spongy tissue, characteristic of equids (Outram and Rowley-Conwy, 1998). On the other hand, horse marrow is richer in linoleic acid (Levine 1998), a substance of great interest to human groups living in cold environments.
The exploitation of the carcasses was not limited to the soft tissues, but the bone themselves were used as blanks for the production of bone tools. A large number of retouchers is produced on horse remains (NR = 35) and more generally on large ungulate remains (NR = 72), representing 8% of the number of remains of these taxa. This proportion is significantly higher than the frequency of retouchers (2.5%) made from medium-size ungulate blanks (Khi2 = 38.177, ddl = 1; p < 0.0001), confirming a strong selection on the nature of the support. This interest in large ungulates, in general, and horses, in particular, is clearly identified by the use of an upper horse incisor as a retoucher (Figure 8).

Figure 8. Piece #4061, decapage 6. Upper horse incisor used as retoucher. Photo and DAO: Malvina Baumann.
Discussion
A zooarchaeological analysis of material from new excavations at Chez-Pinaud brings new insights into the subsistence activities and the diet of the Quina Mousterian that exploited the site around 60,000 years ago.
First and foremost, the importance of large ungulates, and especially horses, seems higher than previously thought. When remains identified by ungulate size classes are taken into account, a large contribution of horse and bison can be highlighted (33% of the NISP). However, the dominance of the reindeer in the faunal spectrum does not necessarily imply its dominance in the Neanderthal diet. Indeed, medium and large size ungulates do not provide the same quantity of animal raw material to the hunters, and if we consider the quantity of meat available on reindeer [35–40 kg (Klokov, 2000)] and horse [150kg (Outram and Rowley-Conwy, 1998)], the ratio (around 4) is more or less equivalent to the reindeer/horse MNI ratio (28/8 = 3.5). Thus, considering that the site is in the direct vicinity of the kill site and consequently the whole carcasses were available, the Quina Neanderthal of Chez-Pinaud would have had access to the same quantity of reindeer and horse meat.
For now, while there are multiple pieces of evidence of seasonal winter hunting of the reindeer (Beauval, 2004; Niven et al., 2012; Rendu et al., 2022), no seasonal information are available for the horse, and we can only mention the absence of fetal remains. Thus, it is not possible to establish if the two taxa were hunted in the same season or not. The ongoing seasonal analyses should be able to solve this issue. Simultaneously, the limited number of individuals does not allow us to discuss deeply the hunted populations, but the presence of juvenile horses attests that matriarchal groups were exploited.
Part of the horse carcasses seems to have been intensively butchered, as was evidenced for the reindeer by Niven and colleagues. There is in addition a statistically significant difference in the frequency of cut marks between horse and reindeer remains (Khi2 = 3.873, ddl = 1; p < 0.05), an interesting pattern, even if such a distinction could be due to differences in handling larger carcasses during the butchering process (Soulier et al., 2022). Differences can also be seen in the anatomical articulations found during excavations: on the 44 bones found in articulated groups during the 2019–2021 excavation, 40 belong to reindeer, two to bison, and two to horse, the difference being statistically significant (Khi2 = 5.01, ddl = 3; p < 0.02). During the Jaubert and Hublin excavations, only reindeer connections were attested. This difference implies that the reindeer carcasses were dismembered more expediently, in larger parts, while the large ungulates would have been more systematically processed. This found an interesting echo in the fact that horse carcasses were more selectively transported than the reindeer ones, maybe due to the difference in weight between the two animals. Seasonal data on large ungulates will also provide discussion on this point, allowing access to the health status of prey, potentially different between reindeer and large ungulates (generating a more or less intense search for marrow for example).
The zooarchaeological analysis underlines a specific interest in the meat on the horse carcasses, whatever we consider the skeletal profiles or the human impact on the bones, confirming the specific place of this taxon in the Neanderthal diet. At the same time, the preferential use of horse bone remains a blank for the bone tool industry, suggesting that horses occupied a specific place in the whole Quina economy (Costamagno et al., 2018). At Les Pradelles, it has been suggested that the preference for large ungulate diaphysis as blanks for retouchers may result from the density constraints necessary to manufacture Quina scrapers (Costamagno et al., 2018); this selection toward large ungulates has also been noticed for several other Quina assemblages (e.g., Soulier, 2007; Daujeard et al., 2014; Jéquier et al., 2018). This particular place is notably underlined by the use of a horse incisor as a retoucher, a unique case in the Middle Paleolithic record.
Thus, the interpretation of the site as a site devoted to the capture and process of reindeer has to be reconsidered or, at least, pondered. The Neanderthals did not come specifically to hunt reindeer herds but rather to hunt reindeer and horses, at least (the place of bison remains to be explored in more detail). This has an important resonance in the discussion of the specialization of the Quina economy on the reindeer since US22 is the most important of the Quina unit from Chez-Pinaud, and Chez-Pinaud itself represents almost one-third of the units used in the different models to discuss the Quina population diet.
While the reindeer is the most common taxa in the Quina faunal spectra of Southwestern France, the horse is the second most common one, sometimes even dominating the spectra such as at La Rouquette L3 (Griggo in Rendu et al. (2011)] and Espagnac Level 2, 3, and 4a (Jaubert, 2001). Its frequency is also notable at Hauteroche (Paletta, 2005), Roc de Marsal Level 4 (Castel et al., 2017), and Combe Grenal Level 21 and 22 (Laquay, 1981; Guadelli, 1987). However, precautions have to be taken when dealing with old collections from Combe Grenal, as some selective sampling occurred during the 50–60s excavations creating a bias in favor of the large ungulates (Discamps and Faivre, 2017). Thus, if we reconsider the whole Quina spectrum through the lens of our current results, we can assume that the role of large ungulates in the Quina diet might have been under-evaluated. Figure 9 proposes the rebalancing of the faunal spectra from Figure 1 using the meat weight of the different taxa [for reindeer: 40 kg, after (Klokov, 2000); red deer: 55 kg, after (Varin, 1980); bison: 250 kg, after (Wheat, 1967; Berger and Cunningham, 1991); horse: 150 kg, (Outram and Rowley-Conwy, 1998)]. It underlines that while reindeer remains the dominant faunal spectrum in 28 out of 32 cases, it constituted the main resource of ungulate meat acquired by the Quina Mousterian in only 22 cases out of 32 and more than 50% in only 11 cases.
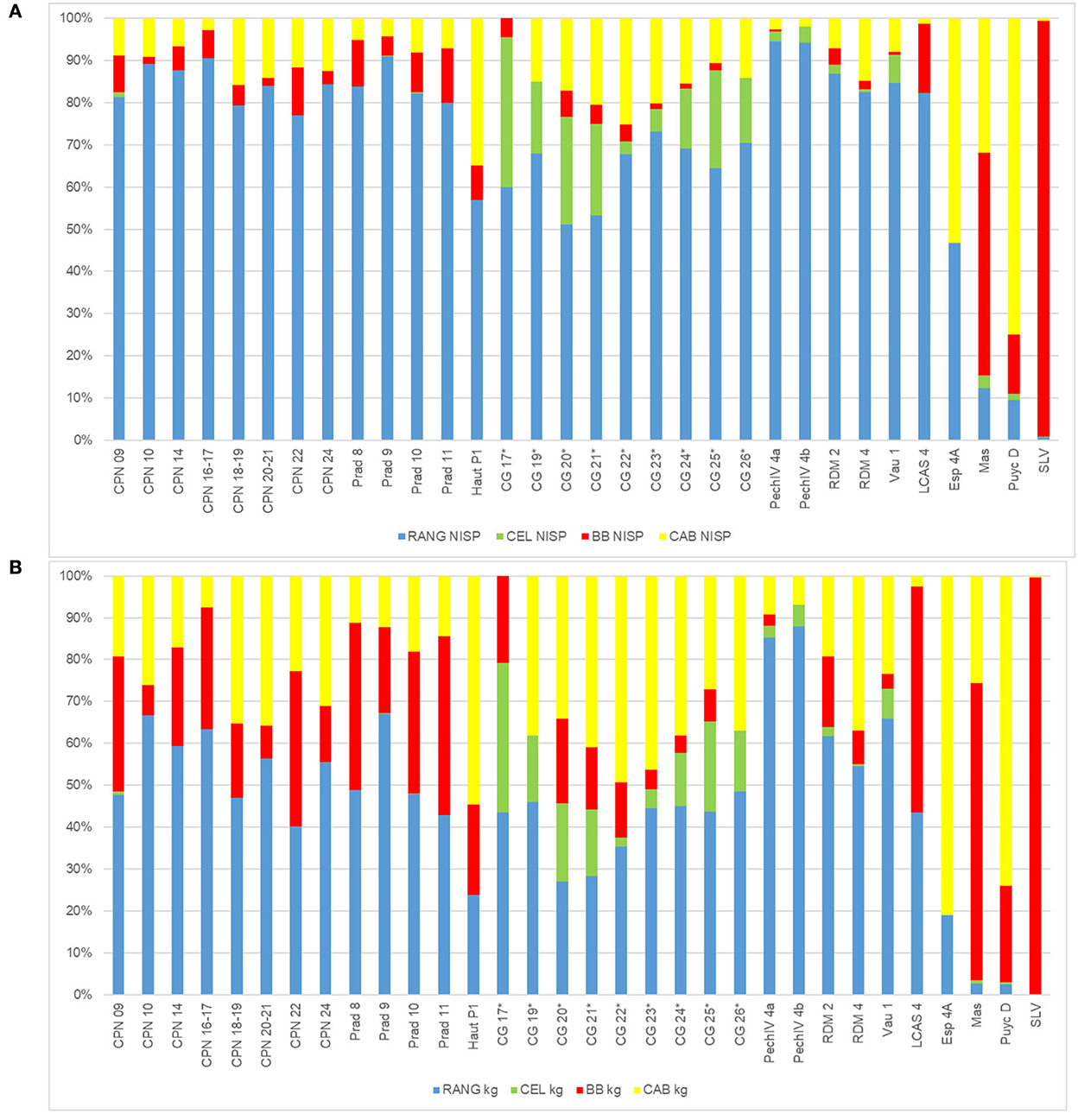
Figure 9. Spectra (A) in %NISP, (B) in kg of meat; Blue, reindeer; green, red deer; Red, bison; Yellow, horse; Gray, other). References used for data CG, Combe Grenal (Laquay, 1981; Guadelli, 1987); Vau, Vauffrey (Delpech, 1996); PechIV, Pech de l'Azé IV (Laquay, 1981; Niven, 2013); RdM, Roc de Marsal (Castel et al., 2017); LP, Les Pradelles (Costamagno et al., 2006); CPN, Chez-Pinaud-Jonzac (Airvaux, 2004; Jaubert et al., 2008b; Niven, 2013); LCAS, La Chapelle-aux-Saints (Rendu et al., 2014); Haut, Hauteroche (Paletta, 2005); LR, La Rouquette (Rendu et al., 2011); SLV, Sous les Vignes (Turq et al., 1999); ESP, Espagnac (Jaubert, 2001). Numbers correspond to the different stratigraphic units. *For Combe Grenal Reindeer was under-evaluated in the previous excavation due to selective sampling.
Naturally, numerous biases here are directly affecting these comparisons (the weight reference selected, the variation between males and females, the use of NISP and not MNI, the problem of transport strategies, the preservation, etc …). Keeping these different limitations in mind, it appears that although the reindeer might have dominated the number of animals killed by Quina Neanderthals (Discamps and Royer, 2017), other taxa might have significantly contributed to their diet.
Thus, the Quina subsistence pattern might have been more complex than previously described, and if Chez-Pinaud at Jonzac was also recurrently used as a horse kill and butchery site, it would suggest that the large ungulate predation, and more specifically the horse, played a role in the annual organization of the activities within the territory.
Conclusion
This article completes the data we have about the subsistence strategies developed at Chez-Pinaud during the late MIS4. The horse remains attest to the intense exploitation of the carcasses for the meat and the blank of the bone industry and confirms the use of the site as a kill site/primarily butchery site. Without changing the interpretation of the site function and its specific place in the Quina territory, it proposes a more accurate vision of the role of the Horse for the Neanderthals from Chez-Pinaud. In a broader view, by extrapolating our interpretations to the rest of the Quina records, we assume here that large ungulates (horses and bovines) were an important part of the protein resources for the Quina population, which has been presented as specialized by the reindeer. However, due to the lack of seasonal data for the horse, it is not possible for now to discuss a potential seasonal complementarity in the hunting between the horse and the reindeer. The ongoing project should soon clarify this issue and new data will be needed for the rest of the Quina record.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
WR, KK, and SS designed the research and directed the scientific team and the field research. AG, MA, MBo, and SR were responsible of part of the excavation. SS realized the spatial analysis of the site, the post field treatment of the data and the 3D modeling, and submitted the manuscript. ED made the spatial analysis of the faunal remains for this article. WR, SR, and AK collected the zooarchaeological data. WR, SR, AK, M-CS, and ED performed the zooarcaheological analyses. MBa and HP conducted the bone tool industry analysis. KK performed the lithic analysis. GG the discussion on the chronological attribution. WR wrote the article with the help of SS, SR, and AK. All the authors contributed to the editing and correction of it.
Funding
The excavation is part of the IRL 2013 ZooSCAn activity, a joint project between the CNRS and the Institute of Archaeology and Ethnography of the Siberian Branch of the Russian Academy of Sciences. The excavation is funded by the French Ministry of Cultural Heritage under the supervision of the Service Regional d'Archéologie de Nouvelle Aquitaine, Poitiers. We are grateful to the Conseil Général of the Charente-Maritime for its financial and logistic support. Our fieldwork would not have been possible without the major help of the Communauté de Commune de Haute Saintonge and Jonzac Municipality. The LIA Artemir and then the IRL Artemir were essential for their financial and logistic support. The study of the bone industry was supported by the Marie Sklodowska-Curie actions under the European Union's Horizon 2020 research and innovation programme (#839528, MBa). The HADES society supported us by funding part of the field and analytical time of SR. This article is part of the QuinaWorld project that has received funding from the European Research Council (ERC) under the European Union's Horizon 2020 research and innovation program ERC Grant agreement No. 851793.
Acknowledgments
We are thankful to Mr. Belot, Président of the Communauté de Commune de Haute Saintonge for his help and support. We thank Mrs. Annie Gendre for her help and support, and her friendship was essential; the team is really missing her. We are thankful to the UMR 5199 PACEA for its help during the field and the storage of the material.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.1085699/full#supplementary-material
References
Airvaux, J. (2004). Le site paléolithique de chez-Pinaud à Jonzac, Charente-Maritime. Prehistoire du Sud-Ouest. 8, 180p.
Beauval, C. (2004). La faune des niveaux Mousteriens de ‘Chez-Pinaud'(Jonzac, Charente-Maritime, France). Première analyse. Le Site Paléolithique de Chez-Pinaud à Jonzac, Charente-Maritime. Préhistoire du Sud Ouest 8, 125e156.
Behrensmeyer, A. K. (1978). Taphonomic and ecologic information from bone weathering. Paleobiology 4, 150–162. doi: 10.1017/S0094837300005820
Berger, J., and Cunningham, C. (1991). Bellows, copulations, and sexual selection in bison (Bison bison). Behav. Ecol. 2, 1–6. doi: 10.1093/beheco/2.1.1
Binford, L. R. (1980). Willow Smoke and Dogs' Tails: Hunter-Gatherer Settlement Systems and Archaeological Site Formation. Am. Antiquity 45, 4–20. doi: 10.2307/279653
Blumenschine, R. J., Marean, C. W., and Capaldo, S. D. (1996). Blind tests of inter-analyst correspondence and accuracy in the identification of cut marks, percussion marks, and carnivore tooth marks on bone surfaces. J. Archaeol. Sci. 23, 493–507. doi: 10.1006/jasc.1996.0047
Brain, C. K. (1981). The Hunter or the hunted. An Introduction to African Cave Taphonomy. Chicago, London: The University of Chicago press.
Britton, K., Grimes, V., Niven, L., Steele, T. E., McPherron, S., Soressi, M., et al. (2011). Strontium isotope evidence for migration in late Pleistocene Rangifer: Implications for Neanderthal hunting strategies at the Middle Palaeolithic site of Jonzac, France. J. Hum. Evol. 61, 176–185. doi: 10.1016/j.jhevol.2011.03.004
Castel, J. C., Discamps, E., Soulier, M-C., Sandgathe, D., Dibble, H. L., McPherron, S. P., et al. (2017). Neandertal subsistence strategies during the Quina Mousterian at Roc de Marsal (France). Quatern. Int. 431, 216–222. doi: 10.1016/j.quaint.2015.12.033
Claud, É., Soressi, M., Jaubert, J., and Hublin, J.-J.. (2012). Étude tracéologique de l'outillage moustérien de type Quina du bonebed de Chez-Pinaud à Jonzac (Charente-Maritime). Nouveaux éléments en faveur d'un site de boucherie et de traitement des peaux. Gallia Prehistoire 54, 3–32. doi: 10.3406/galip.2012.2492
Costamagno, S., Bourguignon, L., Soulier, M-C., Meignen, L., Beauval, C., Rendu, W., et al. (2018). Bone retouchers and site function in the Quina Mousterian: The case of Les Pradelles (Marillac-le-Franc, France), in The Origins of Bone Tool Technology, Mainz, Verlag des Römisch-Germanisches Zentralmuseums, dir, J. Hutson, 165–195.
Costamagno, S., Meignen, L., Beauval, C., Vandermeersch, B., and Maureille, B. (2006). Les Pradelles (Marillac-le-Franc, France): A mousterian reindeer hunting camp? J. Anthropol. Archaeol. 25, 466–484. doi: 10.1016/j.jaa.2006.03.008
Daujeard, C., Moncel, M-H., Fiore, I., Tagliacozzo, A., Bindon, P., Raynal, J-P., et al. (2014). Middle Paleolithic bone retouchers in Southeastern France: variability and functionality. Quatern. Int. 326–327, 492–518. doi: 10.1016/j.quaint.2013.12.022
Delagnes, A., and Rendu, W. (2011). Shifts in Neandertal mobility, technology and subsistence strategies in western France. J. Archaeol. Sci. 38, 1771–1783. doi: 10.1016/j.jas.2011.04.007
Delpech, F. (1996). L'environnement animal des Moustériens Quina du Périgord. Paléo 8, 31–46. doi: 10.3406/pal.1996.905
D'Errico, F., and Villa, P. (1997). Holes and grooves: the contribution of microscopy and taphonomy to the problem of art origins. J. Hum. Evol. 33, 1–31. doi: 10.1006/jhev.1997.0141
Discamps, E. (2014). Ungulate biomass fluctuations endured by Middle and Early Upper Paleolithic societies (SW France, MIS 5-3): The contributions of modern analogs and cave hyena paleodemography. Quat. Int. 337, 64–79. doi: 10.1016/j.quaint.2013.07.046
Discamps, E., and Faivre, J-P. (2017). Substantial biases affecting Combe-Grenal faunal record cast doubts on previous models of Neanderthal subsistence and environmental context. J. Archaeol. Sci. 81, 128–132. doi: 10.1016/j.jas.2017.03.009
Discamps, E., Jaubert, J., and Bachellerie, F. (2011). Human choices and environmental constraints: deciphering the variability of large game procurement from Mousterian to Aurignacian times (MIS 5-3) in southwestern France. Quatern. Sci. Rev. 30, 2755–2775. doi: 10.1016/j.quascirev.2011.06.009
Discamps, E., and Royer, A. (2017). Reconstructing palaeoenvironmental conditions faced by Mousterian hunters during MIS 5 to 3 in southwestern France: A multi-scale approach using data from large and small mammal communities. Quatern. Int. 433, 64–87. doi: 10.1016/j.quaint.2016.02.067
Faivre, J.-P., Gravina, B., Bourguignon, L., Discamps, E., and Turq, A. (2017). Late Middle Palaeolithic lithic technocomplexes (MIS 5–3) in the northeastern Aquitaine Basin: Advances and challenges. Quatern. Int. 433, 116–131. doi: 10.1016/j.quaint.2016.02.060
Gifford, D. P., and Crader, D. C. (1977). A computer coding system for archaeological faunal remains. Am. Antiq. 42, 225–238. doi: 10.2307/278983
Grayson, D. K., and Frey, C. F. (2004). Measuring Skeletal Part Representation in Archaeological Faunas. J. Taphon. 2, 27–42.
Guadelli, J.-L.. (1987). Contribution a l'etude des zoocenoses prehistoriques en Aquitaine (Würm ancien et interstade würmien) (PhD thesis). Universite Bordeaux 1.
Guadelli, J. -L., and Ozouf, J. -C. l. (1994). “Étude expérimentale de l'action du gel sur les restes fauniques. Premiers résultats,” in Outillage peu élaboré en os et bois de cervidés. IV Table ronde taphonomie/bone modification no 6 (Paris: Artefact), 47–56.
Jaubert, J. (2001). Un site moustérien de type Quina dans la vallée du Célé: Pailhès à Espagnac-Sainte-Eulalie. Gallia Préhistoire 43, 1–100. doi: 10.3406/galip.2001.2312
Jaubert, J., Hublin, J.-J., McPherron, S. P., and Soressi, M. (2008a). CHEZ-PINAUD, JONZAC (CHARENTE-MARITIME) Fouille programmée triennale 2005-2007 3ème année 2007, Service régional de l'archéologie de Poitou-Charentes, Poitiers. ed.
Jaubert, J., Hublin, J.-J., Mcpherron, S. P., Soressi, M., Bordes, J.-G., Claud, É., et al. (2008b). Paléolithique moyen récent et Paléolithique supérieur ancien à Jonzac (Charente-Maritime).
Jéquier, C., Livraghi, A., Romandini, M., and Peresani, M. (2018). “Same but different: 20,000. years of bone retouchers from northern Italy A diachronologic approach from Neanderthals to anatomically modern humans,” in The Origins of Bone Tool Technology,Mainz, Verlag des Römisch-Germanisches Zentralmuseums, dir. J. Hutson, 269–285.
Klokov, K. B. (2000). Nenets reindeer heders on the lower Yenisei River: Traditional economy under current conditions are responses to economic change. Polar Res. 19, 39–47. doi: 10.1111/j.1751-8369.2000.tb00326.x
Lam, Y., Chen, X., and Pearson, O. (1999). Intertaxonomic variability in patterns of bone density and the differential representation of bovid, cervid, and equid elements in the archaeological record. Am. Antiq. 64, 343–362. doi: 10.2307/2694283
Laquay, G. (1981). Recherches sur les faunes du Würm I en Périgord (PhD thesis). Universite Bordeaux 1.
Marean, C. W., and Assefa, Z. (1999). Zooarcheological evidence for the faunal exploitation behavior of Neandertals and early modern humans. Evol. Anthropol. Issues News Rev. 8, 22–37. doi: 10.1002/(SICI)1520-6505(1999)8:1<22::AID-EVAN7>3.0.CO;2-F
Metcalfe, D., and Jones, K. T. (1988). A Reconsideration of Animal Body-Part Utility Indices. Am. Antiq. 53, 486–504. doi: 10.2307/281213
Niven, L. (2013). “A diachronic evaluation of Neanderthal cervid exploitation and site use at Pech de l'Azé IV, France,” in Zooarchaeology and Modern Human Origins, eds. J. L. Clarck, J.D. Speth, 151–161.
Niven, L., Steele, T. E., Finke, H., Gernat, T., and Hublin, J.-J.. (2009). Virtual skeletons: using a structured light scanner to create a 3D faunal comparative collection. J. Archaeol. Sci. 36, 2018–2023. doi: 10.1016/j.jas.2009.05.021
Niven, L., Steele, T. E., Rendu, W., Mallye, J-B., McPherron, S. P., Soressi, M., et al. (2012). Neandertal mobility and large-game hunting: the exploitation of reindeer during the Quina Mousterian at Chez-Pinaud Jonzac (Charente-Maritime, France). J. Hum. Evol. 63, 624–635. doi: 10.1016/j.jhevol.2012.07.002
Olsen, S. L., and Shipman, P. (1988). Surface modification on bone: Trampling versus butchery. J. Archaeol. Sci. 15, 535–553. doi: 10.1016/0305-4403(88)90081-7
Outram, A., and Rowley-Conwy, P. (1998). Meat and Marrow Utility Indices for Horse (Equus). J. Archaeol. Sci. 25, 839–849. doi: 10.1006/jasc.1997.0229
Paletta, A. (2005). L'évolution des comportements de subsistance des hommes du Moustérien au Solutréen dans la région Poitou-Charentes (France) (PhD thesis). Muséum national d'histoire naturelle, Paris.
Pickering, T. R., and Egeland, C. P. (2006). Experimental patterns of hammerstone percussion damage on bones: implications for inferences of carcass processing by humans. J. Archaeol. Sci. 33, 459–469. doi: 10.1016/j.jas.2005.09.001
QGIS Development Team. (2022). QGIS Geographic Information System, and the Open Source Geospatial Foundation Project. Available online at: http://qgis.osgeo.org.
Rendu, W. (2010). Hunting behavior and Neanderthal adaptability in the Late Pleistocene site of Pech-de-l'Azé I. J. Archaeol. Sci. 37, 1798–1810. doi: 10.1016/j.jas.2010.01.037
Rendu, W., Beauval, C., Crevecoeur, I., Bayle, P., Balzeau, A., Bismuth, T., et al. (2014). Evidence supporting an intentional Neandertal burial at La Chapelle-aux-Saints. Proc. Natl. Acad. Sci. U. S. A. 111, 81. doi: 10.1073/pnas.1316780110
Rendu, W., Bourguignon, L., Costamagno, S., Meignen, L., Soulier, M-C., Armand, D., et al. (2011). “Mousterian hunting camps: Interdisciplinary approach and methodological considerations,” in Hunting Camps in Prehistory. Current Archaeological Approaches, Proceedings of the International Symposium, eds F. Bon, S. Costamagno, and N. Valdeyron (University Toulouse II - Le Mirail), 61–76.
Rendu, W., Pubert, E., and Discamps, E. (2022). “Using cementochronology to discuss the organization of past neanderthal societies,” in Dental Cementum in Anthropology, eds, S. Naji, W. Rendu, L. Gourichon (Cambridge: Cambridge University Press), 275–287.
Rendu, W., Renou, S., Soulier, M-C., Rigaud, S., Roussel, M., Soressi, M., et al. (2019). Subsistence strategy changes during the Middle to Upper Paleolithic transition reveals specific adaptations of Human Populations to their environment. Sci. Rep. 9, 15817. doi: 10.1038/s41598-019-50647-6
Soulier, M.-C.. (2007). Étude archéozoologique du carré M16 de la couche 2 du gisement moustérien du Roc-de-Marsal (Dordogne), Mémoire de Master I, Université de Toulouse-Le Mirail 110p.
Soulier, M.-C., Costamagno, S., Claud, E., and Deschamps, M. (2022). “Tracing the past: butchering a bison with Middle Palaeolithic stone tools,” in Recreating Artefacts and Ancient Skills: From Experiment to Interpretation, Targoviste, Cetatea de Scaun Publishing, eds. M. Mărgărit, and A. Boronean? 13–31.
Soulier, M. C., and Costamagno, S. (2017). Let the cutmarks speak! Experimental butchery to reconstruct carcass processing. J. Archaeol. Sci:. Rep. 11, 782–802. doi: 10.1016/j.jasrep.2016.12.033
Turq, A., Guadelli, J.-L., and Quintard, A. (1999). “A propos de deux sites d'habitat moustérien de type Quina à exploitation du bison: l'exemple du Mas-Viel et de Sous-les-Vignes,” in J. P.Brugal, F. David, J. G. Enloe, J. Jaubert, Antibes, eds. Le Bison: Gibier et Moyen de Subsistance Des Hommes Du Paléolithique Aux Paléoindiens Des Grandes Plaines, 143–158.
Varin, E. (1980). Chevreuil, cerf, sanglier: Etudes et récits d'un chasseur. les Editions de l'Orée. 270p.
Villa, P., and Mahieu, E. (1991). Breakage patterns of human long bones. J. Hum. Evol. 21, 27–48. doi: 10.1016/0047-2484(91)90034-S
Keywords: Neanderthal, hunting strategy, zooarchaeology, Middle Paleolithic, site function
Citation: Rendu W, Renou S, Koliasnikova A, Baumann M, Plisson H, Discamps E, Soulier M-C, Gicqueau A, Augoyard M, Bocquel M, Guerin G, Shnaider S and Kolobova K (2023) Neanderthal subsistence at Chez-Pinaud Jonzac (Charente-Maritime, France): A kill site dominated by reindeer remains, but with a horse-laden diet? Front. Ecol. Evol. 10:1085699. doi: 10.3389/fevo.2022.1085699
Received: 31 October 2022; Accepted: 30 November 2022;
Published: 20 January 2023.
Edited by:
Eduardo Jiménez-Hidalgo, University of the Sea, MexicoReviewed by:
Ruth Blasco, Institut Català de Paleoecologia Humana i Evolució Social (IPHES), SpainMarie-Hélène Moncel, Director of Research CNRS-MNHN, France
Copyright © 2023 Rendu, Renou, Koliasnikova, Baumann, Plisson, Discamps, Soulier, Gicqueau, Augoyard, Bocquel, Guerin, Shnaider and Kolobova. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: William Rendu,  d2lsbGlhbS5yZW5kdUBjbnJzLmZy; Svetlana Shnaider, c3ZldGEuc2huYXlkZXJAZ21haWwuY29t
d2lsbGlhbS5yZW5kdUBjbnJzLmZy; Svetlana Shnaider, c3ZldGEuc2huYXlkZXJAZ21haWwuY29t