 Leonhard Sommer
Leonhard Sommer Yves P. Klinger
Yves P. Klinger Tobias W. Donath
Tobias W. Donath Till Kleinebecker
Till Kleinebecker Sarah Harvolk-Schöning
Sarah Harvolk-Schöning
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 16 January 2023
Sec. Conservation and Restoration Ecology
Volume 10 - 2022 | https://doi.org/10.3389/fevo.2022.1061484
This article is part of the Research Topic Origin, Conservation, and Restoration of the Threatened European Grassland Ecosystem in the Anthropocene View all 16 articles
Restoration of floodplain meadows remains a challenge, as many degraded sites suffer from seed limitation. The transfer of seed-containing plant material from species-rich donor sites is a widely used method to restore semi-natural grasslands. However, most studies on the success of such restoration projects comprise limited time frames. As factors determining restoration success may only become evident after many years, long-term observations are crucial. We re-investigated 20 restored grassland sites in the floodplain of the Northern Upper Rhine 13–16 years after plant material transfer with different soil preparation treatments. To this end, we carried out vegetation surveys on 254 permanent plots and studied the potential influence of soil preparation, soil nutrients, and hydrology on plant species composition, diversity, and transfer of target species. Since sustainable agricultural use is important to ensure the long-term stability of restored semi-natural grasslands, we further investigated biomass productivity and feeding value. While most target species increased in frequency or remained stable over time, we found no positive long-term effect of soil preparation on vegetation development and target species establishment. Instead, increased biomass yield and flooding frequency led to reduced restoration success, while higher soil C/N ratios had a positive effect. Overall, restoration measures did not affect the agricultural value of the restored grasslands, which had higher dry matter biomass yields compared with the donor sites. Our results indicate that the positive effect of soil preparation on the number and cover of target species, which is regularly reported in short-term studies, diminishes over time, and other factors such as site conditions become increasingly important. Furthermore, additional plant material transfer or manual seeding may be necessary to support target species establishment. Concerning agricultural usability, the integration of restored floodplain meadows in farming systems is possible and can ensure long-term management and thus stability of these ecosystems. Our study shows that long-term monitoring of restoration projects is necessary, as factors determining restoration success may only become evident in the long-term.
The worldwide degradation of ecosystems is one of the most urgent problems of our time (Díaz et al., 2019). Ecological restoration is a major tool to counteract ecosystem degradation, helping the health, integrity, and sustainability of ecosystems to recover (Society for Ecological Restoration, 2004; IPBES, 2019). The current UN “Decade on ecosystem restoration” (UN General Assembly, 2019) underpins the increasing importance of this field. Growing focus is given to the conservation and restoration of grasslands, as they cover a large proportion of land surface and provide high capacity to support biodiversity, multiple ecosystem services and are integral to human well-being (Bardgett et al., 2021). In Central Europe, semi-natural grasslands are of particular importance. They are the result of centuries of human activity, and low-intensive management by mowing or grazing is required to restore and maintain these semi-natural ecosystems and the services they provide (Bakker, 1989; Hejcman et al., 2013).
Floodplain meadows are outstandingly diverse grassland ecosystems with many rare and endangered plant species (Rodwell, 1992; Wesche et al., 2012). Historically, due to their high productivity, they served as an important source for forage provision for livestock (Rothero et al., 2016). Over the last decades, massive structural changes in agriculture resulted in severe consequences for floodplain meadows (Jefferson and Pinches, 2009). Conversion to arable fields, fertilisation, higher cutting frequencies, and alterations of the hydrological conditions have led to a drastic decline both in the amount of floodplain meadows and their ecological quality (Böger, 1991; Joyce and Wade, 1998; Bissels et al., 2004). The restoration of the plant diversity of floodplain meadows is therefore urgent, but is a challenging and long-lasting process (Engst et al., 2016).
The mere return to low-intensive management on degraded floodplain meadows often fails, as typical plant species hardly re-establish spontaneously. This is due to the transient soil seed bank of many typical floodplain meadow species (Bekker et al., 2000; Hölzel and Otte, 2004a) and lacking connectivity to the few species-rich remnant populations (Donath et al., 2003; Bissels et al., 2004). Therefore, active diaspore introduction is required to re-establish the typical vegetation within a reasonable timespan (Bissels et al., 2004; Vécrin et al., 2007; Jõgar and Moora, 2008; Ludewig et al., 2021). Research projects have shown the suitability of active species introduction for grassland restoration (Kiehl et al., 2010). Out of the available methods, the transfer of freshly cut plant material is considered particularly advantageous with respect to genetic diversity and autochthonism, and additionally enables the transfer of organisms other than plants, such as invertebrates (Harnisch et al., 2014; Stöckli et al., 2021).
Generally, the restoration of species-rich grassland using freshly cut plant material is more challenging on species-poor grassland sites compared to arable fields or raw soils (Kiehl et al., 2010; Hansen et al., 2022; Valkó et al., 2022). Soil preparation is commonly regarded as an important prerequisite for successful target species introduction. While it reduces competition by the existing grassland vegetation and creates niches for germination and successful establishment of seedlings with low competitive power (Schmiede et al., 2012), there is increasing evidence that its positive effects can diminish over longer time periods (Harvolk-Schöning et al., 2020; Freitag et al., 2021). This affirms the importance of long-term monitoring to evaluate the success of restoration measures (Resch et al., 2019), as well as considering a range of driving factors (Hölzel, 2019).
However, in addition to restoration, semi-natural grasslands require adapted management to create adequate disturbance regimes and to overcome seed limitation (Klinger et al., 2021). Typically, management of floodplain meadows consists of mowing, which was traditionally complemented by grazing in some areas (Kapfer, 2010). To ensure an adequate management, farmers often receive subsidies as part of agri-environment schemes (Donath et al., 2021; European Environment Agency, 2022). However, the acceptance for low-intensity management practices might be increased if the biomass produced on these sites could be used profitably. Thus it is desirable to keep floodplain meadows integrated in the regional farming systems (Tallowin and Jefferson, 1999; Donath et al., 2015). This was commonly the case until the middle of the last century, but with more possibilities to increase productivity, e.g., by fertiliser input, the interest of farmers to continue this practice decreased (Hejcman et al., 2013). If it could be shown that species diversity and composition had neutral or positive effects on fodder quantity and/or quality, this might increase the motivation of farmers to re-establish the management of sites with high nature conservation value (Donath et al., 2015). Donath et al. (2015) found that in comparison to sites with low nature conservation value, the fodder quality was comparable or even higher in sites with high nature conservation value, and that the harvested material could be integrated in farming systems. If this were the case also for restored semi-natural grasslands, a sustainable management of these sites and thus long-term restoration success could be ensured easier.
Soil conditions and productivity of floodplain meadows are linked to their agricultural value, which may consequently result in a conflict of goals for any restoration efforts (Donath et al., 2015). Increased nutrient levels can hamper the establishment of target species (Gough and Marrs, 1990; Pywell et al., 2006; Waldén and Lindborg, 2016), but relevant nutrients and respective thresholds vary between study systems. In addition, nutrient stoichiometry can modify restoration outcomes, as, e.g., limitation by nitrogen (N) has been shown to compensate for negative impacts of high P and K availability, restricting productivity and species competition (Pywell et al., 2002; Donath et al., 2007). Additionally, hydrological conditions such as flood and drought frequencies can strongly affect species composition in floodplains (Hölzel, 1999; Mathar et al., 2015), but their impact on restoration success has barely been studied so far.
In a large-scale floodplain meadow restoration experiment with plant material transfer at the Northern Upper Rhine in Germany, the effect of soil preparation and soil properties on species establishment on species-poor grassland had been investigated over the first 3 years (Schmiede et al., 2012). Here, we re-investigated the sites 13–16 years after the restoration to answer the following questions:
a. How have the target species developed on the restoration sites, and is the effect of soil preparation on species richness and composition still detectable after 13–16 years?
b. Do the restoration sites differ from the donor sites and the unrestored grassland in the close surrounding with respect to their ecological properties?
c. What is the agricultural value of the restoration sites, compared to unrestored reference grassland in the surrounding and the donor sites?
d. Which effect do soil properties, productivity, nutrient stoichiometry, and hydrological characteristics of the restoration sites have on the long-term restoration success?
The study area is located approximately 30 km southwest of Frankfurt in Hesse (Germany), in the floodplain of the Northern Upper Rhine. The mean annual temperature of 11.1°C marks the region as one of the warmest in Germany, and the mean annual precipitation is relatively low with 550 mm (HLNUG, 2022; stations Frankfurt (Main) Airport for temperature and Groß-Gerau-Wallerstädten for precipitation, 1992–2021). The fluctuating water level of the river Rhine results in both floods and droughts, with groundwater levels of up to 5 m below the surface (HLNUG, 2021). Soils are characterised by high clay contents often exceeding 50% (Burmeier et al., 2010), which adds to the alternating soil water conditions. The specific site conditions and low-intensive haymaking supported the development of species-rich floodplain meadows of the alliances Molinion and Cnidion (habitat types 6410 and 6440 according to the EU Fauna-Flora-Habitat directive), containing a high number of rare and endangered (alluvial) grassland species (Hölzel, 1999; Donath et al., 2003). However, intensification and conversion into arable fields caused massive habitat losses in the course of the 20th century, leaving only small isolated remnants of species-rich floodplain meadows (Böger, 1991; Hölzel and Otte, 2003).
After a series of major floods in 1983, 150 ha of arable fields were converted to non-intensively managed, unfertilized grassland in order to re-establish species-rich floodplain meadows (Böger, 1991; Bissels et al., 2004). However, typical species hardly re-immigrated, and the re-established grassland in the study area remained rather species-poor (Schmiede et al., 2012). From 2005 to 2008, freshly mown plant material (seed-containing green hay sensu Kiehl et al., 2010) was gained from eight donor sites to create 20 strips (“restoration sites”) on different species-poor grassland sites. The donor sites consisted of species-rich Molinion or Cnidion meadows. Each restoration site was 120 m long and 10 m wide and divided in three segments of 40 m length, each of which had been prepared 2–7 weeks before the plant material transfer. All three segments had been mown, and then treated as following:
• rotovated twice,
• ploughed and harrowed, or
• left untilled.
Rotovating broke up the soil surface, while ploughing turned over the topsoil, with subsequent harrowing breaking up the new surface and levelling it (Supplementary Figure A1). Both treatments left a fine-grained seedbed with close to no intact vegetation, but elimination was more complete after ploughing and harrowing. However, depending on the timespan until plant material application, modest regrowth occurred on both treatments. Treatments were randomly arranged on each restoration site. Plant material transfer took place between mid-September and the end of October, when most species on the donor sites carried ripe seeds. Harvest coincided with the first cut (two donor sites, five restoration sites) or second cut (six donor sites, 15 restoration sites), depending on the mowing regime at the donor sites. A detailed description of restoration measures and sites can be found in Schmiede et al. (2012).
Between 10 May and 13 June 2021, we investigated the vegetation on 254 plots (25 m2). From these, 180 were located on the 20 restoration sites (nine per site, three per treatment). For nine of the 20 restoration sites, the plots had been previously studied by Schmiede et al. (2010) annually in the first 3 years after restoration, enabling comparison over time.
Additionally, as a reference, we placed 40 plots (two per restoration site) on the unrestored grassland surrounding all restoration sites, with a distance > 15 m to the restoration sites. Furthermore, on the eight donor sites, 34 plots were surveyed (3–5 per site, depending on their size). To enable comparability with the data of Schmiede et al. (2012), species abundance was recorded using the modified Braun-Blanquet scale (van der Maarel, 1979). For data analysis, species abundance classes were transformed to percentage values following the approach of Schmiede et al. (2012). In addition to the vegetation plots, we recorded whole-site species lists for restoration and donor sites and estimated species abundances using a DAFOR scale (Norfolk Wildlife Trust, n.d.) with modifications (Supplementary Table A1).
In April and May 2021, we gathered soil samples from each of the 254 plots. To this end, composite samples of four topsoil cores (0–10 cm) were collected using a soil corer of 2.5 cm diameter. Samples were air-dried and sieved to 2 mm. Soil pH was measured in CaCl2 solution. The samples were extracted with calcium-acetate-lactate solution (CAL) for the determination of plant-available potassium (K) and phosphorus (P) (Blume et al., 2000). Total soil nitrogen (N) and carbon (C) were measured via elementary analysis (device “Unicube,” co. “elementar”; DIN EN 16168, 2012; DIN EN 15936, 2012), anorganic C was calculated from the CaCO3 content determined with the Scheibler method (Blume et al., 2000). The organic C content was calculated as the difference between total and anorganic C content, and the C/N ratio as the ratio between organic C and total soil N content (Kuntze et al., 1994).
For each plot, aboveground biomass was harvested in four randomly placed quadrats of 0.1 m2 at a height of 5 cm. Sampling took place between end of May and beginning of June, shortly before the regular first grassland cut at 8 June. Most of the donor sites are cut later in summer, but were sampled at the same time for comparability. Biomass samples were merged for each plot to one composite sample, dried at 60°C for 48 h, weighed, and milled to 0.5 mm. The acid detergent fibre (ADF), N, K, and P contents were determined via Near Infrared Spectroscopy (NIRS, details see Kleinebecker et al., 2011). As measures of nutrient stoichiometry, we calculated the N/K and N/P ratios. For feeding value assessment, we calculated the crude protein content (XP; Roth et al., 2011), the digestible energy (DE) for horses (National Research Council, 1999), the metabolisable energy (ME) for ruminants, and the net energy for lactation (Kirchgeßner and Kellner, 1982).
For calculation of hydrological variables, we used data from 33 groundwater wells (HLNUG, 2021) and daily water levels for 12 points of the river Rhine along the study area between 1 January 2001 and 31 December 2020. If data gaps for the groundwater wells were ≤ 30 days, we interpolated the groundwater levels (GWL) between adjacent time points to obtain daily groundwater water levels. The daily Rhine water levels were linearly interpolated between gauging stations Mainz, Nierstein-Oppenheim, and Worms (Wasserstraßen-und Schifffahrtsverwaltung des Bundes, 2021). The 45 groundwater points were used for daily Delaunay triangulation (Sinclair, 2016), including all points with an entry for the respective day. The daily groundwater level of each of the 254 plots was estimated as the inverse-distance weighted mean of the three nearest groundwater points. For each plot, we calculated three relevant hydrological predictors for species distribution (following Gattringer et al. (2019)):
• days per year with GWL > 0.7 m below ground (“Drought frequency”).
• days per year with inundation height > 0.5 m (“Flood frequency”).
• standard deviation of the GWL (“SD of GWL”).
The location in the fossil floodplain, which is protected from flooding by a dyke, and the recent functional floodplain is often used as a hydrological predictor for ecological properties of floodplain meadows (e.g., Bissels et al., 2004; Donath et al., 2007). For our restoration sites, it was well represented by the SD of GWL. The mean was 0.72 m ± SD of 0.14 m for sites located in the functional floodplain, and 0.37 m ± 0.11 m for sites located in the fossil floodplain, respectively. Thus, we focused on the SD of GWL for further analysis, instead of the floodplain compartment.
To assess the impact of soil preparation over time, we compared the number and cover of target species (Schmiede et al., 2012, slightly modified, Table 1) as well as the number of species of the three soil preparation treatments for each of the first 3 years after restoration and for 2021 separately for the previously studied restoration sites. If an ANOVA indicated significant differences, these were identified with a Tukey honest-significant difference test (HSD; α = 0.05). Data were ln-transformed to meet normality and homoscedasticity, and model assumptions were checked visually using diagnostic plots (Kozak and Piepho, 2018).
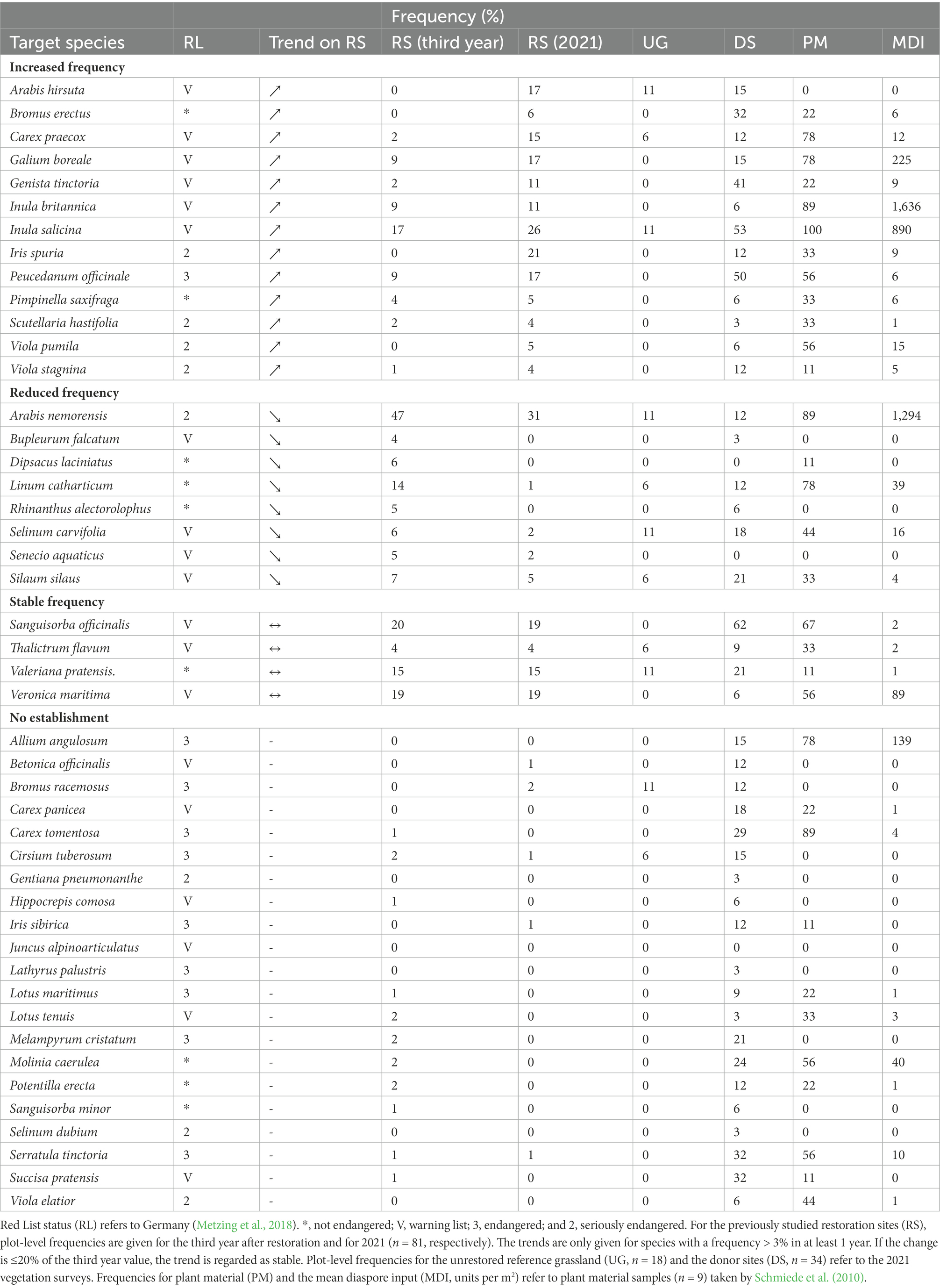
Table 1. Development of target species over time.
To assess temporal trends of the individual target species, we calculated their occurrence frequencies in the third year after restoration and in 2021 for the previously studied restoration sites on a plot basis. For comparison, occurrence frequencies in the plant material and mean diaspore input per target species for each restoration site were calculated from the plant material data of Schmiede et al. (2010). We performed analogous frequency calculations for the corresponding unrestored reference plots of the previously studied restoration sites and for the donor site plots from the 2021 data.
We explored the temporal development of the vegetation composition on the restoration sites using non-metric multidimensional scaling (NMDS) ordination for the previously studied restoration sites for (a) the first 3 years after restoration and (b) for 2021, including the corresponding unrestored reference and donor site plots. We performed a second NMDS for all restoration sites, unrestored reference plots, and donor sites. Ordinations were based on Bray-Curtis Distances, max. 100 iterations, and a random starting configuration. Three-dimensional solutions were chosen by visually checking the decrease of stress values with increasing number of dimensions, according to Leyer and Wesche (2008). To explore underlying ecological gradients, we included vectors for the number of species and target species, the proportions of plant life forms and life span groups, mean Ellenberg indicator values for light (L), temperature (T), continentality (K), moisture (F), nutrients (N), and soil reaction (R), the proportion of indicator species for alternating water levels (data from Klotz et al., 2002), as well as the soil, biomass, and hydrological variables described above. The package “vegan” was used for the ordination (Oksanen et al., 2020).
We compared the different soil preparation treatments of all restoration sites with the unrestored reference grassland and the donor sites concerning soil C/N ratio, total soil N, plant available soil P and K, species and target species numbers, cover of target species, biomass yield, and energy content measures. Variables were pooled at the treatment level for the restoration sites and at the site level for the references. To this end, we performed an ANOVA, followed by a Tukey HSD test (α = 0.05). Data were ln-transformed if diagnostic plots indicated violations against model presumptions (Kozak and Piepho, 2018).
We calculated four indicators to quantify the long-term restoration success of the restoration sites at the site level, following Kiehl et al. (2010) (Supplementary Table A2). These were (a) absolute transfer rate of species, (b) absolute transfer rate of target species, (c) relative transfer rate of species, and (d) relative transfer rate of target species. We defined absolute and relative transfer rates for all species and for target species as the ratios between transferred and transferable species. For absolute transfer rates, species were regarded as transferable if their DAFOR abundance was R2 or higher on at least one corresponding donor site of a restoration site. For relative transfer rates, species found in the plant material from the respective restoration site (Schmiede et al., 2010) were regarded as transferable. A transferable species was regarded as transferred if recorded on a restoration site in 2021. Species from the corresponding unrestored reference plots were regarded as resident and excluded from the pool of transferable and transferred species for the respective restoration site. We opted for the calculation of both absolute and relative transfer rates as we had more data points for the absolute transfer rates (n = 20). However, since the relative transfer rates (n = 15) are based on the species composition of the plant material used for restoration, they are considered a more direct success measure. At plot level, we calculated the (e) increase in target species number and (f) increase in target species cover as the difference between the plot on the restoration site and the mean of the corresponding unrestored reference plots.
To identify factors determining the restoration success, linear regression models were used for the six success variables (a–f) separately at site level. Eleven explanatory variables were included in the model selection using the “dredge” function (R package MuMIn, Bartoń, 2020). These were the N/P and N/K ratio of the biomass, biomass yield, soil pH, plant-available P and K, soil organic C content, and the C/N ratio. Soil N content was not included due to high correlation with soil organic C (r = 0.995). Drought and flood frequency and the SD of GWL were included as hydrological variables. The explanatory variables were centred to a mean of 0 and scaled to a standard deviation of 1. The model with the lowest AIC that showed no multicollinearity (all variance inflation factors ≤2.5) was selected.
The development over time can only be assessed for the previously studied restoration sites, which were restored in 2005 and 2006, so the observation period is 15–16 years here. After that time, the differences between soil preparation treatments concerning target species number and cover vanished (Figure 1). Compared with the first years after restoration, in the long-term, both variables remained relatively stable for the ploughed and rotovated treatments but increased for the untilled treatment. In 2021, the mean number of target species per 25 m2 ranged from 2.2 ± 0.5 (ploughed, mean ± SE) to 3.0 ± 0.5 (rotovated), and mean target species cover ranged from 3.1 ± 0.8 (untilled) to 3.5 ± 0.9% (rotovated). The mean number of species per plot was around 25 for all treatments. For the untilled treatment, this marked a stable trend since the third year after restoration, whereas the species number per plot decreased from around 38 to 25 species for the treatments with soil preparation. This finding was supported by the NMDS ordination of the previously studied restoration sites indicating that species composition of soil disturbance plots became more similar to the unrestored reference plots until 2021 (Supplementary Figure A2).
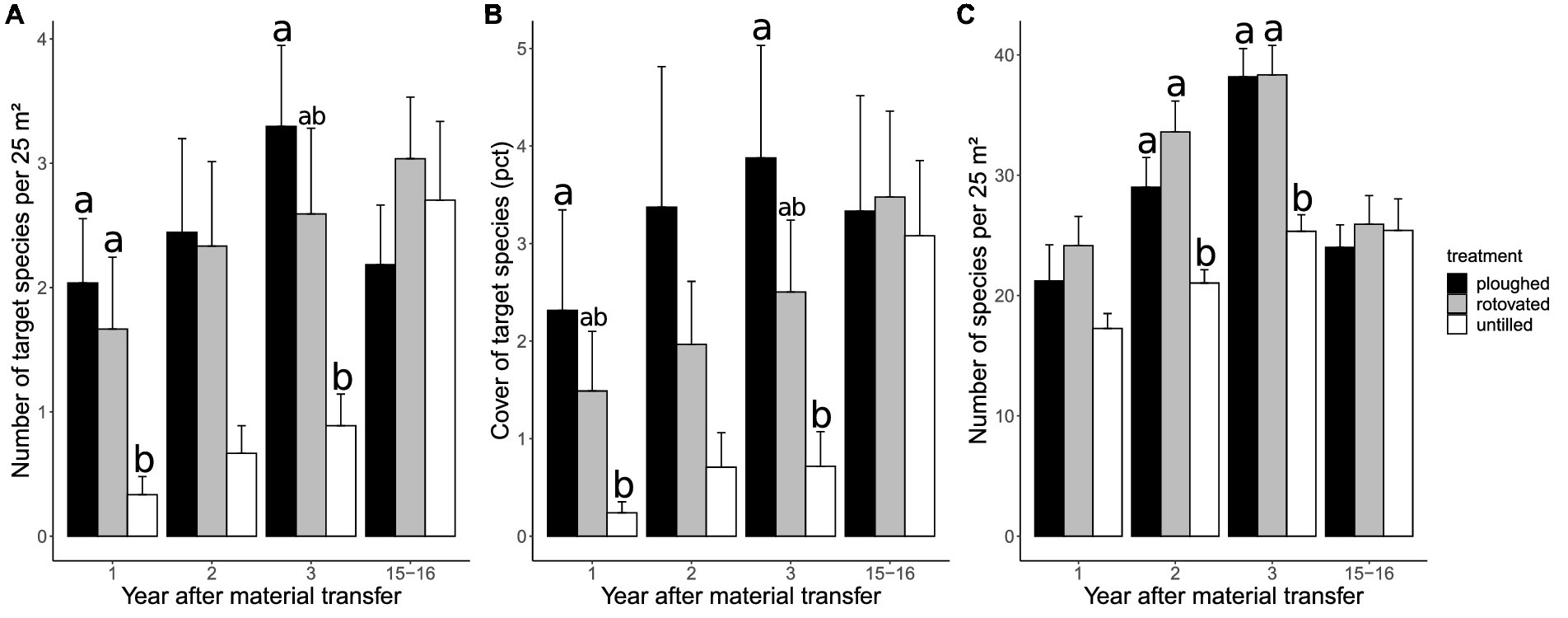
Figure 1. Development of the number (A) and cover (B) of target species and the number of species (C) per 25 m2 over time after the transfer of plant material on the previously studied restoration sites for the three different treatments ploughed, rotovated, and untilled (n = 9, respectively). Significant differences within years (p < 0.05, ln-transformed data) are indicated by different letters above the bars. Whiskers refer to the standard errors.
Out of the 46 target species, 13 showed a higher frequency at the restoration sites in 2021 compared to 3 years after restoration (Table 1). Among these, we found a range of Red List species, such as Carex praecox, Galium boreale, Genista tinctoria, Iris spuria, and Peucedanum officinale. During the investigation period, eight target species decreased in frequency. These had been mostly recorded with low frequencies by Schmiede et al. (2010) already, such as Bupleurum falcatum, Rhinanthus alectorolophus, and Selinum carvifolia. An exception was Linum catharthicum, which was recorded only in 1% of the restoration plots in 2021, compared to 14% 3 years after restoration. Sanguisorba officinalis and Veronica maritima remained relatively stable with a frequency of around 20%, respectively.
The NMDS of all restoration sites and the references for 2021 revealed that the donor sites were separated from the unrestored reference grassland and the restoration sites (Figure 2). While there was a wide overlap between the latter two groups, the centroid of the unrestored reference grassland was separated from the centroids of the restoration soil treatments, which were all slightly shifted towards the donor sites. The donor sites were characterised by higher target species and species numbers, energy contents, Ellenberg R values, and drought frequency compared to the other groups. Both the restored and unrestored sites were characterised by higher productivity levels, indicated by increased Ellenberg N values and biomass yields compared with the donor sites.
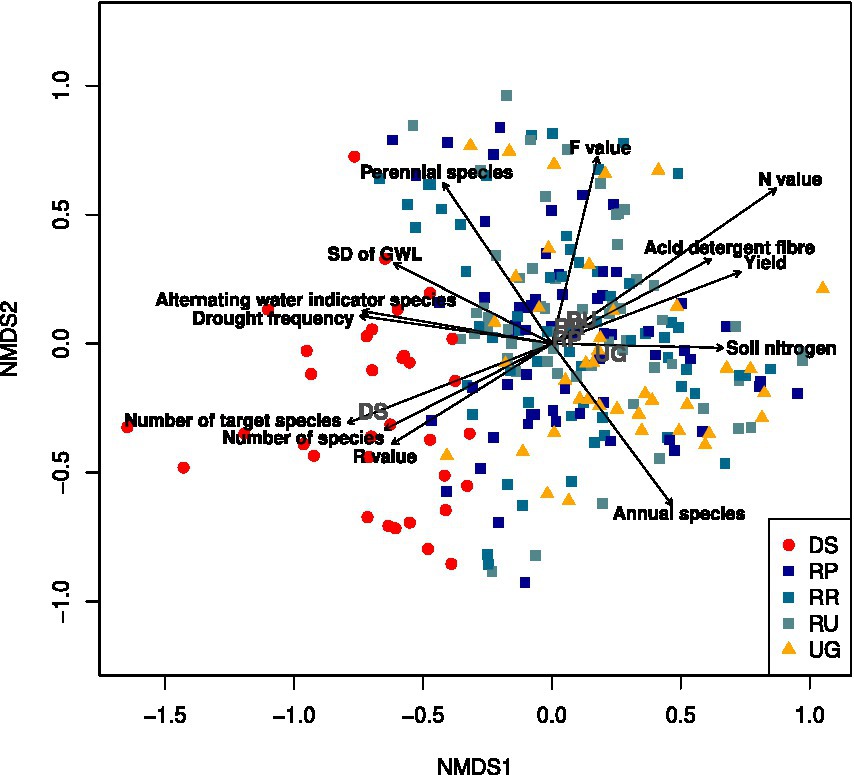
Figure 2. Non-metric multidimensional scaling (NMDS) of the vegetation plots for all restoration sites, the unrestored reference grassland and the donor sites in 2021 (axis 1 and 2 of the three-dimensional solution are shown). Final stress: 16.9. Plots are grouped by donor sites (DS), unrestored reference grassland (UG), and restoration sites with the treatments ploughed (RP), rotovated (RR), and untilled (RU). The group labels are located at the centroids of the groups. Vectors with r2 > 0.3 are displayed. For better readability, the cover of target species, the metabolisable energy (high correlation with number of species and target species) were removed despite r2 > 0.3.
Concerning soil nutrient status, the restoration sites and the unrestored reference grassland were very similar (Figure 3). Their C/N ratio averaged at 11.0 ± 0.1 (± SE), compared to 11.9 ± 0.3 for the donor sites. No significant differences between the groups were found for total nitrogen, plant-available P and K contents. However, soil nutrient contents of the donor sites were lower than those of the restoration sites and unrestored reference plots. Especially the low and very narrow plant-available P content of the donor sites (0.9 ± 0.1%) was noticeable.
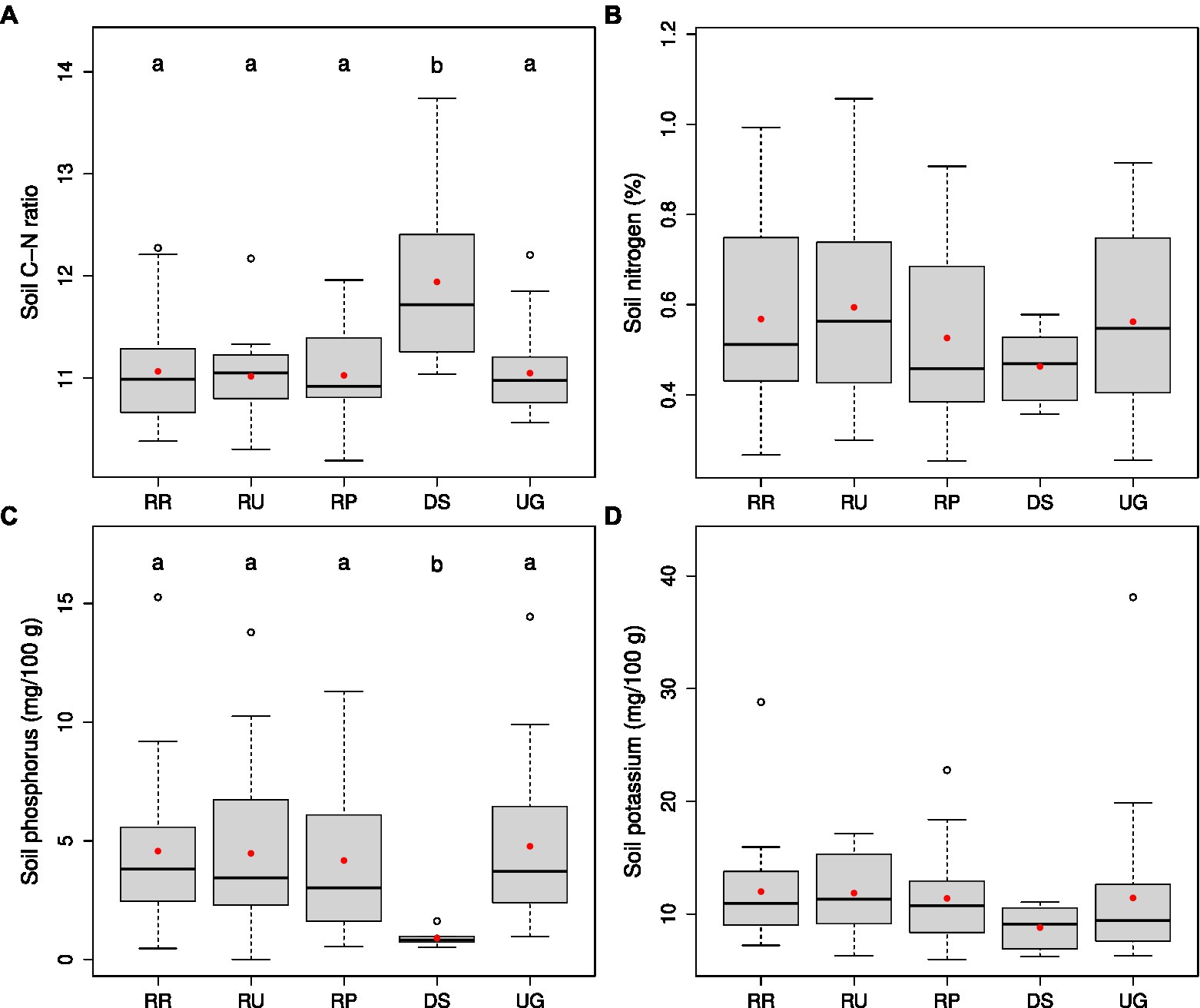
Figure 3. Box-whisker plots of the soil C/N ratio (A), total nitrogen (%; B), plant-available P (mg/100 g; C), and plant-available K (mg/100 g; D) in 2021 on the different site categories—donor sites (DS, n = 8), all restoration sites with treatments ploughed (RP), rotovated (RR), and untilled (RU), and unrestored reference grassland (UG; n = 20, respectively). Plot data were averaged on the treatment level or, in case of DS and UG, on the site level. Red dots display the mean values. Significant differences (p < 0.05) are indicated by different letters above the boxes (testing on ln-transformed data for soil P and soil K).
In 2021, over all restoration sites, the number of target species per plot was similar for all soil treatments with an average of 1.9 ± 0.3 (Figure 4A) and significantly higher than for the unrestored reference grassland (0.7 ± 0.2), but significantly lower than for the donor sites (7.0 ± 1.4). Although not significant, mean cover of target species was higher on the restoration sites (2.1 ± 0.4%) than on the unrestored reference grassland (1.3 ± 0.6%; Figure 4B). For the donor sites, however, target species cover was significantly and markedly higher (mean = 19.7 ± 5.0%). The same held true for the number of recorded plant species (Figure 4C).
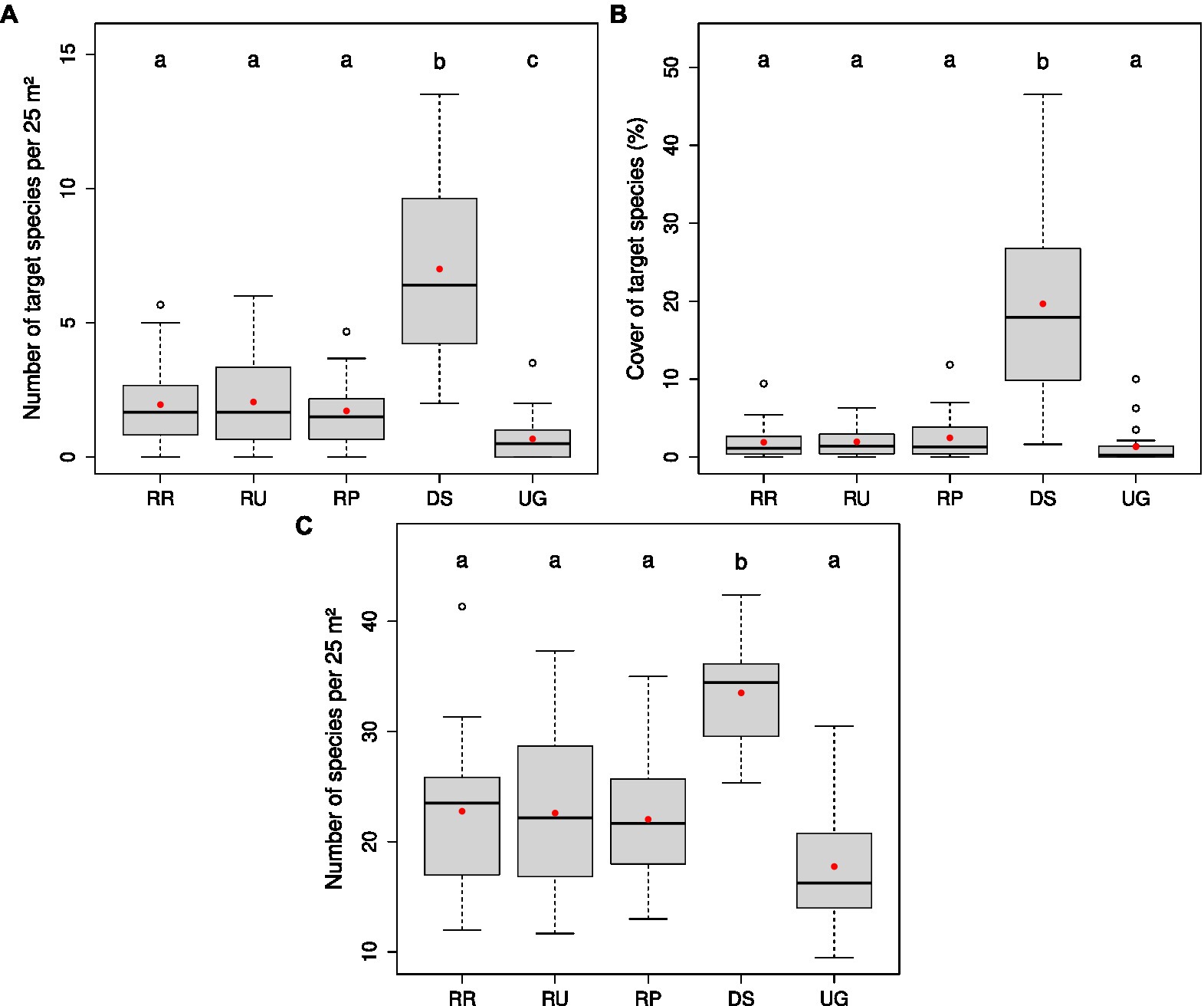
Figure 4. Box-whisker plots of the number of target species per plot (A), the cover of target species (%; B), and the number of species per 25 m2 plot (C) in 2021 on the different site categories—donor sites (DS, n = 8), all restoration sites with treatments ploughed (RP), rotovated (RR), and untilled (RU), and unrestored reference grassland (UG; n = 20, respectively). Plot data were averaged on the treatment level or, in case of DS and UG, on the site level. Red dots display the mean values. Significant differences (p < 0.05) are indicated by different letters above the boxes (testing on ln-transformed data for number and cover of target species).
The biomass yield levels of the restoration sites (407 ± 26 g/m2) and the unrestored reference grassland (421 ± 36 g/m2) did not differ but both were significantly higher compared to the donor sites, which had an average yield of 239 ± 29 g/m2 (Figure 5A). The energy content variables for cattle and horses were similar between restoration sites and the unrestored reference grassland, with those of the donor sites being 4–5% higher (Figures 5B–D).
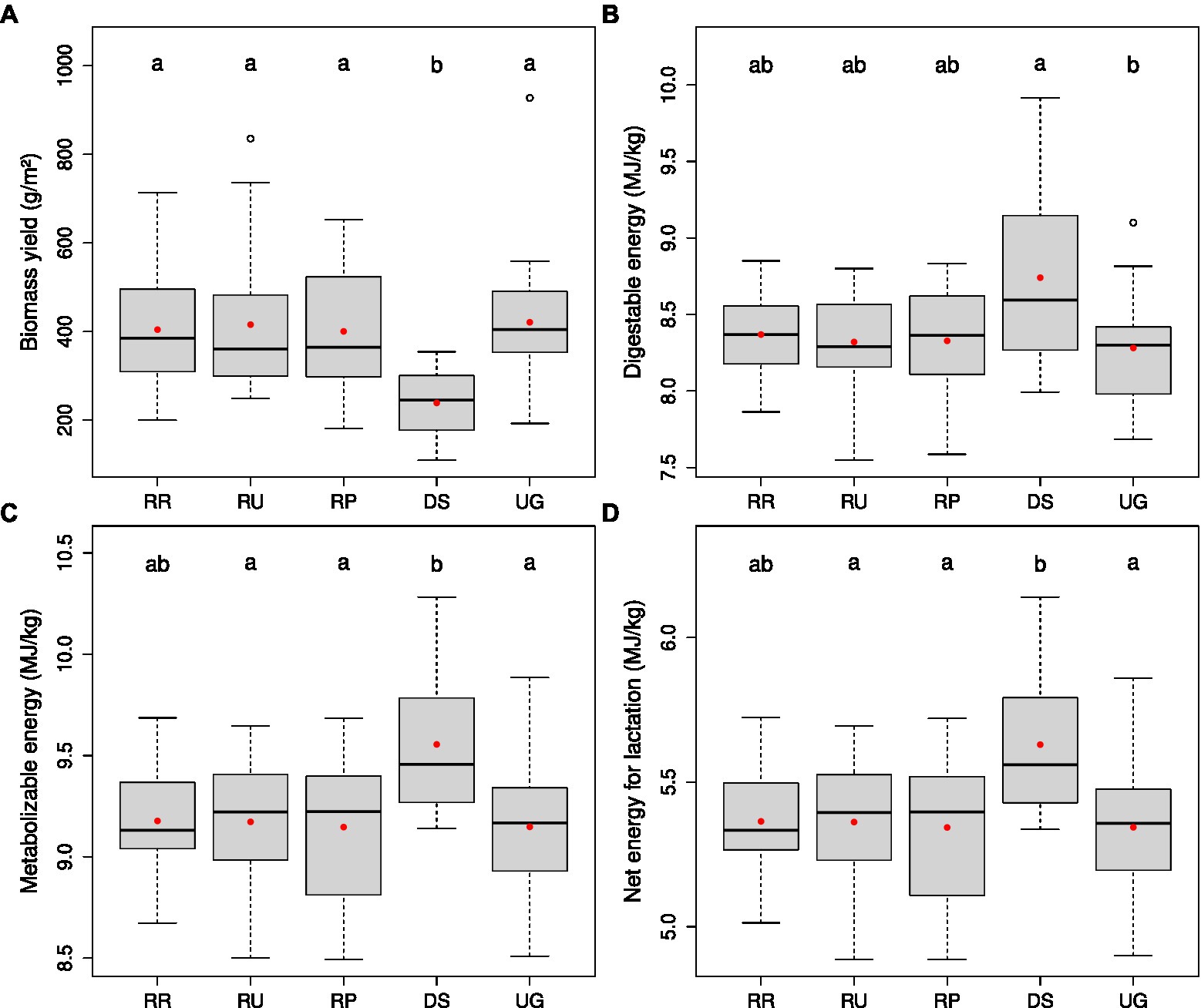
Figure 5. Box-whisker plots of the biomass yield (g/m2; A), the digestible energy for horses (MJ/kg; B), the metabolisable energy for ruminants (MJ/kg; C), and the net energy for lactation (MJ/kg; D; all referring to dry matter) in 2021 on the different site categories—donor sites (DS, n = 8), all restoration sites with treatments ploughed (RP), rotovated (RR), and untilled (RU), and unrestored reference grassland (UG; n = 20, respectively). Plot data were averaged on the treatment level or, in case of DS and UG, on the site level. Red dots display the mean values. Significant differences (p < 0.05) are indicated by different letters above the boxes (testing on ln-transformed data for biomass yield).
Absolute transfer rates of both all species and target species averaged at 40.1 ± 3.9 and 36.4 ± 6.2%, respectively (Supplementary Table A2). The corresponding relative transfer rates were 24.9 ± 1.6 and 34.7 ± 3.7%. Higher biomass yield was generally associated with lower numbers of target species and reduced absolute transfer rates of species and target species (Table 2). The C/N ratio was positively associated with both the number and the cover of target species. Out of the hydrological variables, higher flood frequency tended to reduce restoration success, while higher drought frequency and variation of the groundwater level tended to have positive effects. The R2 of the selected models ranged from 36 to 65%, except for the relative transfer rate of species, which could not be explained (R2 = 1%).
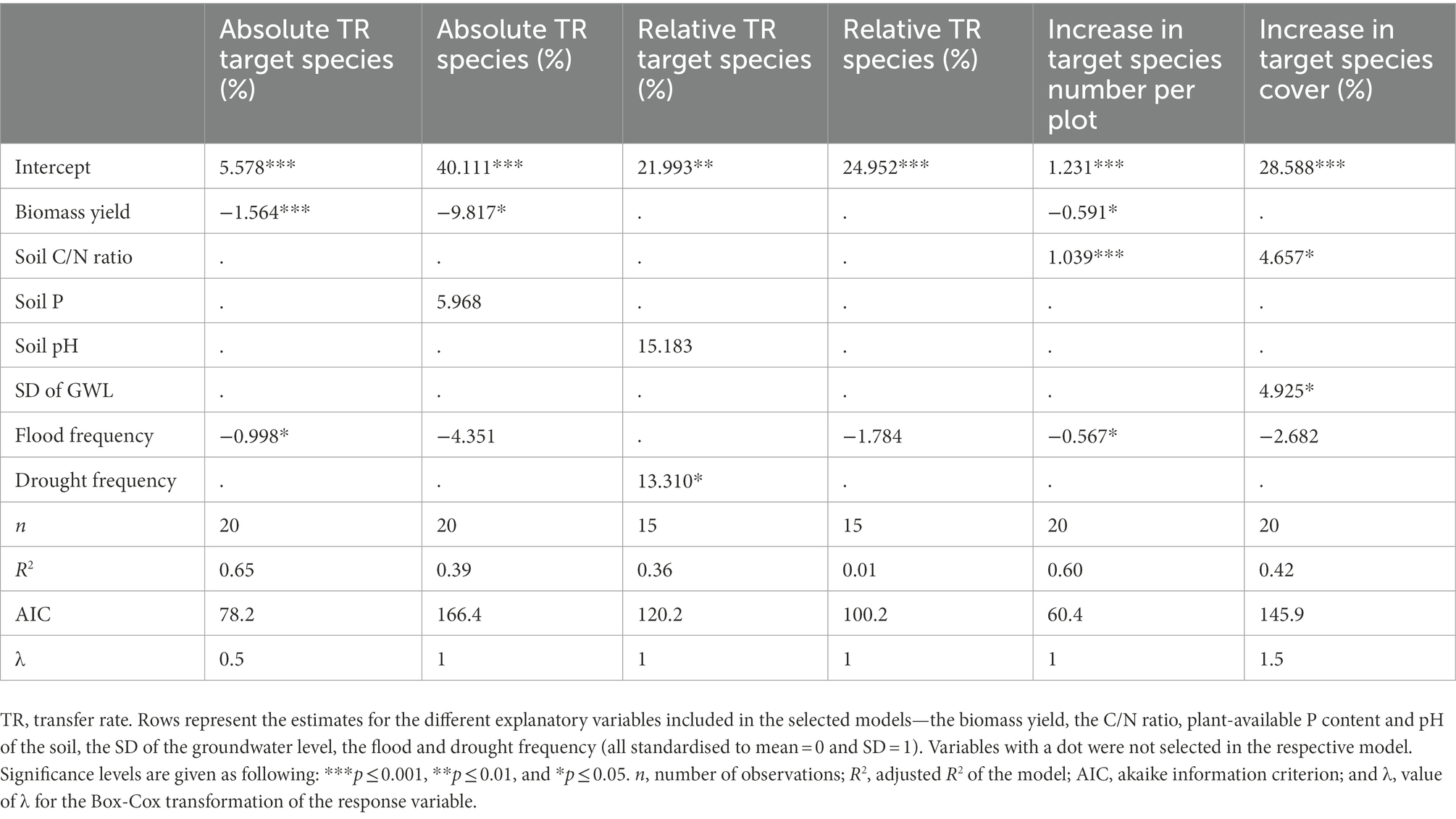
Table 2. Overview of the regression models selected by AIC criterion for the ecological restoration success variables on the site level.
We found no effect of soil preparation on vegetation development and target species establishment 13–16 years after restoration. This is surprising, as one of the main findings of Schmiede et al. (2012) was that soil disturbance, especially ploughing, enabled better (target) species establishment due to suppression of the existing grassland vegetation. This is a common observation among different grassland types, so that soil preparation prior to diaspore introduction in species-poor grassland is often recommended (Kiehl et al., 2010). However, studies deriving such advice from their findings mostly have short observation timeframes and to the best of our knowledge do not exceed 8 years (Edwards et al., 2007; Bischoff et al., 2018; Durbecq et al., 2021). In line with our findings, recent studies on floodplain meadow restoration by Harvolk-Schöning et al. (2020) and Heilscher (2020) indicate that the positive effect of soil preparation on the number and cover of introduced species diminishes in the long run.
Initially, soil disturbance creates micro-niches for germination and establishment of species from the plant material (Harvolk-Schöning et al., 2020), but also activates the soil seed bank (Schmiede et al., 2012; Ludewig et al., 2021). In the short term, this leads to promotion of ruderal species (Klaus et al., 2018). Accordingly, in our experiment, ruderal species, such as Cirsium arvense, Galium aparine, or Lactuca serriola emerged in high frequencies over the first 3 years after restoration, but receded in the long run. In contrast, some target species such as Inula britannica or Carex praecox emerged later or developed more slowly. This was presumably due to competition with the resident vegetation, but these species established in the long run even without tillage.
Across all treatments, target species that were already present 3 years after restoration mostly remained stable or increased in frequency until 2021. This is in accordance with the stable target species number on plots with soil disturbance and the observed increase for the untilled treatment. An especially encouraging case is Iris spuria, which was not detected by Schmiede et al. (2012) in the first 3 years after restoration, but was detected in considerable amounts on two restoration sites in 2021. The hard seed coat of this species can delay germination, so that establishment happens only after longer time periods (Hölzel and Otte, 2004b). Harvolk-Schöning et al. (2020) observed a similar pattern for Iris spuria on former arable fields. Our results clearly show that in the longer term, the establishment of this highly endangered species is possible on grasslands lacking typical floodplain meadow species.
Many target species were not successfully established, some of them despite frequent occurrence on the donor sites. For example, Succisa pratensis was barely captured in the plant material, which may be due to asynchronous fruit ripening with only a small proportion of ripe seeds when the plant material was harvested, as this species has a long flowering and seed shedding period (Adams, 1955). However, after-ripening of seeds may lead to increased germination even when they are harvested in an unripe state, as was shown for the non-native L. polyphyllus in mountain grasslands (Klinger et al., 2020). Diaspores of Allium angulosum, Selinum carvifolia, and Serratula tinctoria were captured in considerable amounts (in ≥44% of plant material samples and with ≥10 diaspores per m2 on average, respectively), but established poorly or not at all, with an occurrence frequency of 6% at maximum on the previously studied restoration sites over the whole observation period. This matches with observations by Harvolk-Schöning et al. (2020), and could be a consequence of specific germination requirements, e.g., characteristic temperature regimes (Hölzel and Otte, 2004a; Wagner et al., 2021).
Our findings on the nutrient levels for the grassland sites overall matched those of former studies in the region (Donath et al., 2007, 2015; Schmiede et al., 2012). Grasslands of high nature conservation value, often used as donor sites, consistently had much lower plant-available P contents and moderately lower plant-available K contents than species-poor grassland sites often chosen for restoration. For the N contents, no such pattern had been found in those studies. In our study, the tendency to increased N contents of the restoration sites and the untreated reference grasslands compared with the donor sites is probably due to seven restoration sites with high organic C contents of 8.6 ± 0.5% (mean ± standard error; vs. 4.9 ± 0.3% for the other 13 restoration sites). The higher average C/N ratios of the donor sites are mainly driven by two poor Molinion sites with very wide ratios of 12.9 and 13.7, respectively.
A range of rare and endangered plant species of floodplain meadows, many of which are listed in the Red Lists of Germany and Hesse, were successfully established on the restoration sites. However, our results confirm that the ecological restoration of grassland is challenging, even if the sward is disturbed prior to diaspore introduction (Kiehl et al., 2010; Harvolk-Schöning et al., 2020; Hansen et al., 2022). Thirteen to sixteen years after restoration, the vegetation composition of the restored plots was similar to the unrestored reference grassland plots, with only slight changes towards the composition of the donor sites. Nevertheless, the number of target species was significantly higher for restoration sites and also their cover increased, compared with the unrestored reference.
Different restoration measures affected neither the yield nor the energy content of the aboveground biomass. While a change in yield was not expected, a more diverse species composition with a higher proportion of forbs can be associated with higher energy contents of the biomass in floodplain meadows (Donath et al., 2004). The similar biomass energy contents of restored and unrestored reference grasslands can be explained by the marginal effects of restoration measures on the overall vegetation composition.
The yield levels of the restored and unrestored grasslands in our study system are mostly within the previously observed range of non-intensively managed grasslands of wet and mesotrophic sites (Tallowin and Jefferson, 1999; Donath et al., 2015). With dry matter yields of up to 705 g/m2, some sites exceeded the levels normally reached without fertilisation (Tallowin and Jefferson, 1999). Under these conditions and with regard to the current subsidy policy (EU area bonus and conservation contracts), haymaking is economically viable for the regional farmers. For lactating cows, the hay may be at best recommended as basic feed, as the net energy for lactation of 5.4 ± 0.2 MJ/kg dry matter (mean ± SD) would require supplementation with high-energy compounds (Donath et al., 2004, 2021; Schumacher, 2016). The metabolisable energy contents of 9.2 ± 0.3 MJ/kg dry matter indicate suitability as complete feed for non-lactating cows (Donath et al., 2004; Deutsche Landwirtschafts-Gesellschaft, 2009) and empty ewes or ewes in early pregnancy, as well as for integration in compound feed rations for calves (Bayerische Landesanstalt für Landwirtschaft (LfL), 2021). Practically, most of the hay harvested in the region is used for leisure horses. The hay from our restoration sites and their surroundings is suitable for this with regard to the observed digestible energy levels of 8.3 ± 0.3 MJ/kg dry matter (National Research Council, 1999; Donath et al., 2004).
Slightly higher energy contents of the biomass from highly species-diverse donor sites indicate that an increase in species diversity does not preclude the integration of species-rich swards into feeding rations (Tallowin and Jefferson, 1999; Donath et al., 2004). However, yield of the donor sites is on average 40% lower compared to the restoration sites, which makes it difficult for farmers to operate profitably. Thus, agri-environmental schemes obviously remain an important pre-requisite in the conservation of species-rich grasslands of high-nature value (Donath et al., 2021).
The transfer rates we observed were within the typical range for plant material transfer on species-poor grassland, but lower than on former arable fields (Kiehl et al., 2010). This holds true for both target species as well as total species numbers. While biomass yield and flood frequency had negative impacts, wider C/N ratios positively affected restoration success. These three predictors were identified as significant for at least two success variables.
Biomass yield levels are the result of complex interactions of biotic and abiotic factors (Doyle, 1982), with different nutrients being decisive in different locations and years (Fay et al., 2015). Beside generally relatively fertile soils in floodplains, we suspect that productivity in our restoration sites is partly increased by remnants of former fertilisation, which may be the reason for the very high yield levels of some of the sites. Due to the dominance of tall and highly productive grasses under fertile conditions (Honsova et al., 2007), high productivity reduced the suitability for the establishment of species-rich floodplain meadows.
Regular flooding events lead to nutrient deposition in close proximity to the river channel (Klaus et al., 2011; Poulsen et al., 2014) and increase the productivity of floodplain meadows by higher soil nutrient levels (Beltman et al., 2007). Apart from this, higher water availability increases mineralization and nutrient supply, which leads to highly variable biomass yield between years (Mathar et al., 2015), but also between sites (cf. Jakrlová, 1999). In our study, the positive relationship between flood frequency and biomass yield (r = 0.35, Supplementary Figure A3) could result from a mixture of the fertilising and the mineralizing effect of frequent flooding events. This may explain the identification of flood frequency as negatively affecting restoration success along with the biomass yield observed in 2021. Another reason for the adverse effect of flood frequency on restoration success might be that long flooding of seedlings emerged from transferred plant material impedes survival (Bao et al., 2018; Gattringer et al., 2018).
Increased soil C/N ratios were significantly associated with increases in target species number and cover. The C/N ratio in the soil as an indicator of N availability could be another long-term determinant of productivity. However, the positive correlation between C/N ratio and yield (r = 0.30) does not support this. Higher C/N ratios, although not reducing productivity, could facilitate the establishment of typical floodplain meadow species by reducing competition with generalist grassland species adapted to high and continuous N availability. Accordingly, adverse effects of soil nitrogen on target species establishment were observed in the floodplain of the river Elbe (Dullau et al., 2021).
While Schmiede et al. (2012) identified plant available P content in the soil to negatively affect target species numbers in our study system, our resurvey cannot confirm this for the long term. Considering an extended set of factors and sites, the restoration sites rich in organic C, which had not been covered by the study of Schmiede et al. (2012), were among the more productive ones, so that the overall effect of biomass yield might have masked the effect of soil P.
In our study, we found no long-term effect of soil preparation on vegetation development and target species establishment across a large dataset. This indicates that the positive effect of soil preparation on the number and cover of target species, which is regularly reported in short-term studies, diminishes over time, while the effects of local site conditions become more important. Therefore, soil preparation prior to seed introduction may not be necessary in floodplain meadow restoration. To increase restoration success, the productivity of restoration sites, soil C/N ratios, and flooding frequency should fit to the respective restoration goals. For practitioners, choosing restoration sites with productivity levels not greatly exceeding those of the donor sites may be most feasible. If restoration sites are too productive, management schemes that actively reduce site productivity are recommended.
Concerning biomass characteristics, we showed that despite considerable differences in yield, even restoration sites with low productivity provide biomass of sufficient amount and feeding value. Thus, the integration of restored grasslands in local farming systems is possible and can ensure long-term management and thus stability of these ecosystems. Furthermore, one-time introduction of target species showed only limited success. Thus, additional plant material transfer or manual seeding of target species is probably necessary. Further studies should investigate the potential of such supplementary measures. Overall, we strongly recommend long-term monitoring of restoration projects in other regions and grassland types, as factors determining restoration success may become evident only after longer time periods.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
TWD, TK, and SH-S conceived of the research idea and designed the study. LS gathered and analysed the data with the help of YPK and SH-S, and wrote the first draft of the manuscript. All authors contributed to the article and approved the submitted version.
The study was funded by the German Federal Environmental Foundation (grant number 35171/01).
We thank the German Federal Environmental Foundation for funding the study. We express our gratitude to J. Scholz vom Hofe for his exertion in soil and biomass sampling and sample processing, to J. Höhl for the lab analyses, and to S. Flecken for her input on the feeding value aspects of this manuscript. The authors thank M. Harnisch from the city of Riedstadt and R. Baumgärtel from the forestry administration Groß-Gerau for their professional and administrative help at the Northern Upper Rhine, as well as to the local farmers who patiently supported our investigations and are grateful to N. Hölzel from the University of Münster for his ideas on the sampling design and the interpretation of results, and to M. Hahn and K. Willkomm for their help during field work.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.1061484/full#supplementary-material
Bakker, J. P. (1989). Nature Management by Grazing and Cutting. Dordrecht: Kluwer academic publishers.
Bao, F., Elsey-Quirk, T., de Assis, M. A., Arruda, R., and Pott, A. (2018). Seasonal flooding, topography, and organic debris interact to influence the emergence and distribution of seedlings in a tropical grassland. Biotropica 50, 616–624. doi: 10.1111/btp.12550
Bardgett, R. D., Bullock, J. M., Lavorel, S., Manning, P., Schaffner, U., Ostle, N., et al. (2021). Combatting global grassland degradation. Nat. Rev. Earth Environ. 2:10. doi: 10.1038/s43017-021-00207-2
Bartoń, K. (2020). MuMIn: Multi-Model Inference (1.43.17). Available at: https://CRAN.R-project.org/package=MuMIn
Bayerische Landesanstalt für Landwirtschaft (LfL) (2021). Gruber Tabelle zur Fütterung der Milchkühe, Zuchtrinder, Schafe, Ziegen. 47th Edn.
Bekker, R. M., Verweij, G. L., Bakker, J. P., and Fresco, L. F. M. (2000). Soil seed bank dynamics in hayfield succession. J. Ecol. 88, 594–607. doi: 10.1046/j.1365-2745.2000.00485.x
Beltman, B., Willems, J. H., and Güsewell, S. (2007). Flood events overrule fertiliser effects on biomass production and species richness in riverine grasslands. J. Veg. Sci. 18, 625–634. doi: 10.1111/j.1654-1103.2007.tb02576.x
Bischoff, A., Hoboy, S., Winter, N., and Warthemann, G. (2018). Hay and seed transfer to re-establish rare grassland species and communities: how important are date and soil preparation? Biol. Conserv. 221, 182–189. doi: 10.1016/j.biocon.2018.02.033
Bissels, S., Hölzel, N., Donath, T. W., and Otte, A. (2004). Evaluation of restoration success in alluvial grasslands under contrasting flooding regimes. Biol. Conserv. 118, 641–650. doi: 10.1016/j.biocon.2003.10.013
Blume, H. P., Deller, B., Leschber, R., Paetz, K., Furtmann, K., and Wilke, B. M. (2000). Handbuch der Bodenuntersuchung. Berlin: Beuth Verlag.
Burmeier, S., Eckstein, R. L., Otte, A., and Donath, T. W. (2010). Desiccation cracks act as natural seed traps in flood-meadow systems. Plant Soil 333, 351–364. doi: 10.1007/s11104-010-0350-1
Deutsche Landwirtschafts-Gesellschaft (2009). Empfehlungen zur Fütterung von Mutterkühen und deren Nachzucht. Available at: https://llg.sachsen-anhalt.de/fileadmin/Bibliothek/Politik_und_Verwaltung/MLU/LLFG/Dokumente/04_themen/fleischrind/10_fu-empfehl_mutterkuehe_nachzucht.pdf
Díaz, S., Settele, J., Brondízio, E. S., Ngo, H. T., Agard, J., Arneth, A., et al. (2019). Pervasive human-driven decline of life on earth points to the need for transformative change. Science 366:eaax3100. doi: 10.1126/science.aax3100
DIN EN 15936 (2012). Schlamm, behandelter Bioabfall, Boden und Abfall_-Bestimmung des gesamten organischen Kohlenstoffs (TOC) mittels trockener Verbrennung; Deutsche Fassung EN_15936: 2012. Beuth Verlag GmbH.
DIN EN 16168 (2012). Schlamm, behandelter Bioabfall und Boden_-Bestimmung des Gesamt-Stickstoffgehalts mittels trockener Verbrennung; Deutsche Fassung EN_16168: 2012. Beuth Verlag GmbH.
Donath, T. W., Bissels, S., Hölzel, N., and Otte, A. (2007). Large scale application of diaspore transfer with plant material in restoration practice—impact of seed and microsite limitation. Biol. Conserv. 138, 224–234. doi: 10.1016/j.biocon.2007.04.020
Donath, T. W., Daniel, V., and Schneider, S. (2021). Long-term development of fodder quantity and quality of non-intensively managed grasslands in South-Western Luxembourg [PDF].
Donath, T. W., Hölzel, N., Bissels, S., and Otte, A. (2004). Perspectives for incorporating biomass from non-intensively managed temperate flood-meadows into farming systems. Agric. Ecosyst. Environ. 104, 439–451. doi: 10.1016/j.agee.2004.01.039
Donath, T. W., Holzel, N., and Otte, A. (2003). The impact of site conditions and seed dispersal on restoration success in alluvial meadows. Appl. Veg. Sci. 6, 13–22. doi: 10.1111/j.1654-109X.2003.tb00560.x
Donath, T. W., Schmiede, R., and Otte, A. (2015). Alluvial grasslands along the northern upper Rhine – nature conservation value vs. agricultural value under non-intensive management. Agric. Ecosyst. Environ. 200, 102–109. doi: 10.1016/j.agee.2014.11.004
Doyle, C. J. (1982). Modelling the determinants of grassland stocking rates on dairy farms in England and Wales. Agric. Syst. 9, 83–95. doi: 10.1016/0308-521X(82)90024-5
Dullau, S., Rydgren, K., Kirmer, A., Jäger, U. G., Meyer, M. H., and Tischew, S. (2021). The Dessau grassland experiment—impact of fertilization on forage quality and species assembly in a species-rich alluvial meadow. Agriculture 11:4. doi: 10.3390/agriculture11040339
Durbecq, A., d’Ambly, M., Buisson, E., Jaunatre, R., Cluchier, A., and Bischoff, A. (2021). Seedling recruitment in mountain grassland restoration: effects of soil preparation and grazing. Appl. Veg. Sci. 24, 1–11. doi: 10.1111/avsc.12564
Edwards, A. R., Mortimer, S. R., Lawson, C. S., Westbury, D. B., Harris, S. J., Woodcock, B. A., et al. (2007). Hay strewing, brush harvesting of seed and soil disturbance as tools for the enhancement of botanical diversity in grasslands. Biol. Conserv. 134, 372–382. doi: 10.1016/j.biocon.2006.08.025
Engst, K., Baasch, A., Erfmeier, A., Jandt, U., May, K., Schmiede, R., et al. (2016). Functional community ecology meets restoration ecology: assessing the restoration success of alluvial floodplain meadows with functional traits. J. Appl. Ecol. 53, 751–764. doi: 10.1111/1365-2664.12623
European Environment Agency (EEA) (2022). Term: Agri-environment scheme. Available at: https://www.eea.europa.eu/help/glossary/eea-glossary/agri-environment-scheme
Fay, P. A., Prober, S. M., Harpole, W. S., Knops, J. M. H., Bakker, J. D., Borer, E. T., et al. (2015). Grassland productivity limited by multiple nutrients. Nat. Plants 1, 15080–15085. doi: 10.1038/nplants.2015.80
Freitag, M., Klaus, V. H., Bolliger, R., Hamer, U., Kleinebecker, T., Prati, D., et al. (2021). Restoration of plant diversity in permanent grassland by seeding: assessing the limiting factors along land-use gradients. J. Appl. Ecol. 58, 1681–1692. doi: 10.1111/1365-2664.13883
Gattringer, J. P., Ludewig, K., Harvolk-Schöning, S., Donath, T. W., and Otte, A. (2018). Interaction between depth and duration matters: flooding tolerance of 12 floodplain meadow species. Plant Ecol. 219, 973–984. doi: 10.1007/s11258-018-0850-2
Gattringer, J. P., Maier, N., Breuer, L., Otte, A., Donath, T. W., Kraft, P., et al. (2019). Modelling of rare flood meadow species distribution by a combined habitat surface water–groundwater model. Ecohydrology 12, 1–12. doi: 10.1002/eco.2122
Gough, M. W., and Marrs, R. H. (1990). A comparison of soil fertility between semi-natural and agricultural plant communities: implications for the creations of species-rich grassland on abondoned agricultural land. Biol. Conserv. 51, 83–96. doi: 10.1016/0006-3207(90)90104-W
Hansen, W., Klinger, Y. P., Otte, A., Eckstein, R. L., and Ludewig, K. (2022). Constraints in the restoration of mountain meadows invaded by the legume Lupinus polyphyllus. Restor. Ecol. 30:e13682. doi: 10.1111/rec.13682
Harnisch, M., Otte, A., Schmiede, R., and Donath, T. W. (2014). Verwendung von Mahdgut zur Renaturierung von Auengrünland: 10 Tabellen. Stuttgart: Ulmer.
Harvolk-Schöning, S., Michalska-Hejduk, D., Harnisch, M., Otte, A., and Donath, T. W. (2020). Floodplain meadow restoration revisited: long-term success of large scale application of diaspore transfer with plant material in restoration practice. Biol. Conserv. 241:108322. doi: 10.1016/j.biocon.2019.108322
Heilscher, C. (2020). Determinants of long-term restoration success in floodplain meadows. Master thesis, University of Münster, Germany.
Hejcman, M., Hejcmanová, P., Pavlů, V., and Beneš, J. (2013). Origin and history of grasslands in Central Europe—a review. Grass Forage Sci. 68, 345–363. doi: 10.1111/gfs.12066
HLNUG (2021). GruSchu—Hessen. GruSchu Hessen. Available at: https://gruschu.hessen.de/mapapps/resources/apps/gruschu/index.html?lang=de
HLNUG (2022). Wetterextreme. Wetterextreme. Available at: https://klimaportal.hlnug.de/wetterextreme
Hölzel, N. (1999). Flora und Vegetation der Auenwiesen im NSG Lampertheimer Altrhein—Eine aktuelle Zustandsanalyse mit Hinweisen zur zukünftigen Pflege und Entwicklung. Jahrbuch Naturschutz Hessen 4, 24–42.
Hölzel, N. (2019). “Limitierende Faktoren der Renaturierung,” in Renaturierungsökologie. eds. J. Kollmann, A. Kirmer, S. Tischew, N. Hölzel, and K. Kiehl (Berlin and Heidelberg: Springer), 35–52.
Hölzel, N., and Otte, A. (2003). Restoration of a species-rich flood meadow by topsoil removal and diaspore transfer with plant material. Appl. Veg. Sci. 6, 131–140. doi: 10.1111/j.1654-109X.2003.tb00573.x
Hölzel, N., and Otte, A. (2004a). Assessing soil seed bank persistence in flood-meadows: the search for reliable traits. J. Veg. Sci. 15, 93–100. doi: 10.1111/j.1654-1103.2004.tb02241.x
Hölzel, N., and Otte, A. (2004b). Ecological significance of seed germination characteristics in flood-meadow species. Flora-Morphol. Distribut. Funct. Ecol. Plants 199, 12–24. doi: 10.1078/0367-2530-00132
Honsova, D., Hejcman, M., Klaudisova, M., Pavlu, V., Kocourkova, D., and Hakl, J. (2007). Species composition of an alluvial meadow after 40 years of applying nitrogen, phospohorus and potassium fertilizer. Preslia 79, 245–258.
IPBES (2019). Global assessment report on biodiversity and ecosystem services of the intergovernmental science-policy platform on biodiversity and ecosystem services. Zenodo.
Jakrlová, J. (1999). Biomass Production of Floodplain Grasslands. Morava River Floodplain Meadows. DAPHNE, Centre for Applied Ecology, Bratislava, 139–146.
Jefferson, R. G., and Pinches, C. E. (2009). The conservation of floodplain meadows in Great Britain: An overview. J. Nat. Conserv. 14, 11–24.
Jõgar, Ü., and Moora, M. (2008). Reintroduction of a rare plant (Gladiolus imbricatus) population to a river floodplain—how important is Meadow Management? Restor. Ecol. 16, 382–385. doi: 10.1111/j.1526-100X.2008.00435.x
Joyce, C. B., and Wade, P. M. (1998). European Wet Grasslands: Biodiversity, Management and Restoration. Chichester (New York): John Wiley and Sons Ltd.
Kapfer, A. (2010). Mittelalterlich-frühneuzeitliche Beweidung der Wiesen Mitteleuropas. Naturschut. Landschaftsplanung 42, 180–187.
Kiehl, K., Kirmer, A., Donath, T. W., Rasran, L., and Hölzel, N. (2010). Species introduction in restoration projects – evaluation of different techniques for the establishment of semi-natural grasslands in central and northwestern Europe. Basic Appl. Ecol. 11, 285–299. doi: 10.1016/j.baae.2009.12.004
Kirchgeßner, M., and Kellner, R. J. (1982). Schätzung des energetischen Futterwetes von Grün-und Rauhfutter durch die Cellulase-Methode. Landwirtsch. Forsch. 34, 276–281.
Klaus, V. H., Hoever, C. J., Fischer, M., Hamer, U., Kleinebecker, T., Mertens, D., et al. (2018). Contribution of the soil seed bank to the restoration of temperate grasslands by mechanical sward disturbance. Restor. Ecol. 26, S114–S122. doi: 10.1111/rec.12626
Klaus, V. H., Sintermann, J., Kleinebecker, T., and Hölzel, N. (2011). Sedimentation-induced eutrophication in large river floodplains—an obstacle to restoration? Biol. Conserv. 144, 451–458. doi: 10.1016/j.biocon.2010.09.031
Kleinebecker, T., Klaus, V. H., and Hölzel, N. (2011). Reducing sample quantity and maintaining high prediction quality of grassland biomass properties with near infrared reflectance spectroscopy. J. Near Infrared Spectrosc. 19, 495–505. doi: 10.1255/jnirs.957
Klinger, Y. P., Eckstein, R. L., Hansen, W., Kleinebecker, T., Otte, A., and Ludewig, K. (2021). Mowing machinery and migratory sheep herds are complementary dispersal vectors for grassland species. Appl. Veg. Sci. 24:e12579. doi: 10.1111/avsc.12579
Klinger, Y. P., Eckstein, R. L., Horlemann, D., Otte, A., and Ludewig, K. (2020). Germination of the invasive legume Lupinus polyphyllus depends on cutting date and seed morphology. Neo Biota 60, 79–95. doi: 10.3897/neobiota.60.56117
Klotz, S., Kühn, I., and Durka, W. (2002). BIOLFLOR–eine Datenbank zu biologisch-ökologischen Merkmalen der Gefäßpflanzen in Deutschland. Bundesamt für Naturschutz. Bonn: Schriftenreihe für Vegetationskunde
Kozak, M., and Piepho, H.-P. (2018). What’s normal anyway? Residual plots are more telling than significance tests when checking ANOVA assumptions. J. Agron. Crop Sci. 204, 86–98. doi: 10.1111/jac.12220
Kuntze, H., Roeschmann, G., and Schwerdtfeger, G. (1994). Bodenkunde: 188 Tabellen. 5 neubearb. und erw. 5th Edn. Stuttgart: Ulmer.
Leyer, I., and Wesche, K. (2008). Multivariate Statistik in der Ökologie: Eine Einführung. Berlin and Heidelberg: Springer.
Ludewig, K., Hansen, W., Klinger, Y. P., Eckstein, R. L., and Otte, A. (2021). Seed bank offers potential for active restoration of mountain meadows. Restor. Ecol. 29:e13311. doi: 10.1111/rec.13311
Mathar, W., Kleinebecker, T., and Hölzel, N. (2015). Environmental variation as a key process of co-existence in flood-meadows. J. Veg. Sci. 26, 480–491. doi: 10.1111/jvs.12254
Metzing, D., Hofbauer, N., Ludwig, G., and Matzke-Hajek, G. (2018). “Rote Liste gefährdeter Tiere, Pflanzen und Pilze Deutschlands,” in Band 7: Pflanzen (Bonn-Bad Godesberg: Bundesamt für Naturschutz).
National Research Council (1999). Nutrient Requirements of Horses. 5th Edn. Washington DC: National Academy Press.
Norfolk Wildlife Trust (n.d.). Guidance notes for recording DAFOR. Available at: https://www.norfolkwildlifetrust.org.uk/documents/downloads/cwa/handout-9-using-dafor (Accessed November 19, 2020).
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., et al. (2020). Vegan: Community ecology package (2.5–7). Available at: https://CRAN.R-project.org/package=vegan
Poulsen, J. B., Hansen, F., Ovesen, N. B., Larsen, S. E., and Kronvang, B. (2014). Linking floodplain hydraulics and sedimentation patterns along a restored river channel: river Odense, Denmark. Ecol. Eng. 66, 120–128. doi: 10.1016/j.ecoleng.2013.05.010
Pywell, R. F., Bullock, J. M., Hopkins, A., Walker, K. J., Sparks, T. H., Burke, M. J. W., et al. (2002). Restoration of species-rich grassland on arable land: assessing the limiting processes using a multi-site experiment. J. Appl. Ecol. 39, 294–309. doi: 10.1046/j.1365-2664.2002.00718.x
Pywell, R. F., Bullock, J. M., Tallowin, J. B., Walker, K. J., Warman, E. A., and Masters, G. (2006). Enhancing diversity of species-poor grasslands: an experimental assessment of multiple constraints: enhancing diversity of species-poor grasslands. J. Appl. Ecol. 44, 81–94. doi: 10.1111/j.1365-2664.2006.01260.x
Resch, M. C., Schütz, M., Graf, U., Wagenaar, R., van der Putten, W. H., and Risch, A. C. (2019). Does topsoil removal in grassland restoration benefit both soil nematode and plant communities? J. Appl. Ecol. 56, 1782–1793. doi: 10.1111/1365-2664.13400
Rodwell, J. S. (1992). “Grasslands and montane communities,” in British Plant Communities. Vol. 3. (Cambridge: Cambridge University Press).
Roth, F. X., Schwarz, F. J., Stangl, G. I., and Kirchgeßner, M. (2011). Tierernährung: Leitfaden für Studium, Beratung und Praxis. 13 Edn. Frankfurt (Main): DLG-Verlag.
Rothero, E., Lake, S., and Gowing, D. (2016). Floodplain Meadows: Beauty and Utility—A Technical Handbook. Milton Keynes: Floodplain Meadows Partnership, The Open University.
Schmiede, R., Handke, K., Harnisch, M., Donath, T. W., and Otte, A. (2010). Endbericht: Grundlagen für ein Handlungskonzept zur floristischen und faunistischen Anreicherung artenarmer Auenwiesen. Available at: https://www.dbu.de/OPAC/ab/DBU-Abschlussbericht-AZ-23329.pdf
Schmiede, R., Otte, A., and Donath, T. W. (2012). Enhancing plant biodiversity in species-poor grassland through plant material transfer—the impact of sward disturbance. Appl. Veg. Sci. 15, 290–298. doi: 10.1111/j.1654-109X.2011.01168.x
Schumacher, D., and Wolfgang, S. (2016). Naturschutzheu für Milchviehbetriebe? Land InForm 6, 36–39.
Sinclair, D. (2016). S-hull: a fast radial sweep-hull routine for Delaunay triangulation. arXiv [Preprint]. Available at: http://arxiv.org/abs/1604.01428
Stöckli, A., Slodowicz, D., Arlettaz, R., and Humbert, J.-Y. (2021). Transfer of invertebrates with hay during restoration operations of extensively managed grasslands in Switzerland. J. Insect Conserv. 25, 189–194. doi: 10.1007/s10841-020-00282-8
Tallowin, J. R. B., and Jefferson, R. G. (1999). Hay production from lowland semi-natural grasslands: a review of implications for livestock systems. Grass Forage Sci. 54, 99–115. doi: 10.1046/j.1365-2494.1999.00171.x
Valkó, O., Rádai, Z., and Deák, B. (2022). Hay transfer is a nature-based and sustainable solution for restoring grassland biodiversity. J. Environ. Manag. 311:114816. doi: 10.1016/j.jenvman.2022.114816
van der Maarel, E. (1979). Transformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio 39, 97–114. doi: 10.1007/BF00052021
Vécrin, M. P., Grévilliot, F., and Muller, S. (2007). The contribution of persistent soil seed banks and flooding to the restoration of alluvial meadows. J. Nat. Conserv. 15, 59–69. doi: 10.1016/j.jnc.2006.10.001
Wagner, M., Hulmes, S., Hulmes, L., Redhead, J. W., Nowakowski, M., and Pywell, R. F. (2021). Green hay transfer for grassland restoration: species capture and establishment. Restor. Ecol. 29:e13259. doi: 10.1111/rec.13259
Waldén, E., and Lindborg, R. (2016). Long term positive effect of grassland restoration on plant diversity—success or not? PLoS One 11:e0155836. doi: 10.1371/journal.pone.0155836
Wasserstraßen-und Schifffahrtsverwaltung des Bundes (WSV) (2021). Wasserstandsdaten der Rheinpegel Worms, Nierstein-Oppenheim und Mainz, bereitgestellt durch die Bundesanstalt für Gewässerkunde (BfG).
Keywords: conservation, farming, feeding value, green hay transfer, plant material transfer, seed limitation, soil disturbance
Citation: Sommer L, Klinger YP, Donath TW, Kleinebecker T and Harvolk-Schöning S (2023) Long-term success of floodplain meadow restoration on species-poor grassland. Front. Ecol. Evol. 10:1061484. doi: 10.3389/fevo.2022.1061484
Edited by:
Péter Török, University of Debrecen, HungaryReviewed by:
László Erdős, Hungarian Academy of Science, HungaryCopyright © 2023 Sommer, Klinger, Donath, Kleinebecker and Harvolk-Schöning. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Leonhard Sommer,  bGVvbmhhcmQuc29tbWVyQHVtd2VsdC51bmktZ2llc3Nlbi5kZQ==
bGVvbmhhcmQuc29tbWVyQHVtd2VsdC51bmktZ2llc3Nlbi5kZQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.