 Kôji Sasakawa
Kôji Sasakawa- Laboratory of Zoology, Department of Science Education, Faculty of Education, Chiba University, Inage-ku, Chiba, Japan
Male genitalia exhibit extreme morphological diversity among animals, but its diversification mechanism has not been fully elucidated. In insects, the intromittent membranous part has been barely studied. This study performed phylogenetic comparative analysis to examine the morphological associations between the intromittent membranous part (endophallus) and ejaculate characteristics in two Carabidae taxa with different endophallus diversification patterns (genus Amara and tribe Pterostichini). Endophallus morphology was analyzed using geometric morphometrics, whereby effects of shape and size components can be evaluated separately. In both taxa, no morphological associations were found between spermatophore and endophallus traits, but an association was detected between conjugated sperm length and endophallus shape. Considering the available information regarding functional morphology, the association between conjugated sperm length and endophallus shape may indicate a physical interaction between conjugated sperm and the endophallus through the spermatophore. The similarity of results between the two taxa implies functional equivalence among different endophallus types. Concerning individual sperm, several results were detected, but functional interpretation was difficult. Overall, these results imply that shape and size have different effects on ejaculate morphology, with a larger effect of shape than size. This pattern is consistent with previous results for non-intromittent and intromittent sclerotized parts, and the diversification mechanism of the intromittent membranous part may be similar to those of non-intromittent and intromittent sclerotized parts.
Introduction
Male genitalia exhibit extreme morphological diversity among animals. Their diversity has attracted the interest of evolutionary biologists, and numerous studies have investigated the mechanisms underlying this diversity (Arnqvist, 1998; Hosken and Stockley, 2004; Simmons, 2014). Single-species studies and comparative analyses of multiple species have shown that sexual selection is the primary driver of male genital diversity (e.g., Arnqvist and Danielsson, 1999; Simmons et al., 2009; Rowe and Arnqvist, 2012). However, several issues remain unresolved, partly because of methodological limitations.
One such issue is the insufficient examination of intromittent genital parts. Most empirical evidence for the diversification of male genitalia through sexual selection has been obtained from studies of insects. With respect to intromittent parts, these studies have exclusively examined sclerotized structures on the surface (Simmons, 2014). However, the intromittent genital parts of many species are largely membranous (Snodgrass, 1935). Thus, the most major structure of male genitalia remains unstudied. This is likely because of the difficulty of investigating the soft membranous parts, as well as an assumption that membranous parts are functionally less important than sclerotized parts.
Another unresolved issue is the insufficient distinction between shape and size components in morphological analyses. Many studies have analyzed male genital morphology using traditional morphometrics, in which linear measurements and often subsequent principal component analysis are employed (e.g., House and Simmons, 2003; Wenninger and Averill, 2006; Simmons et al., 2009). However, this method cannot adequately distinguish between shape and size components. This problem can be overcome by landmark-based morphometrics (Zelditch et al., 2012), which is a relatively new method in morphometric analysis. Several studies based on this method have been conducted for male genital sclerotized structures. Single-species studies based on mating experiments have revealed different effects between shape and size traits (e.g., Holwell et al., 2010; Simmons and Garcia-Gonzalez, 2011; House et al., 2021). A comparative study of multiple species (Rowe and Arnqvist, 2012) revealed diversification in shape components, which is a pattern opposite to the stabilizing selection in size components implied by negative allometry. Differences between shape and size components in terms of their functions and diversification patterns were highlighted in a recent review of male genital evolution (Simmons, 2014).
These issues must be addressed to fully elucidate the mechanisms underlying male genital diversification. This study focuses on the beetle family Carabidae. Because of its species richness (40,000 known species) and ecological and morphological diversities, Carabidae has been used in many ecological and evolutionary studies, including studies regarding the diversification of male genitalia and associated traits (e.g., Takami and Sota, 2007a; Janovska et al., 2013; Sasakawa, 2020). In Carabidae, a method has been established for observation of the male genital intromittent part of specimens under actual mating conditions (Janovska et al., 2013). The morphology of this genital structure, known as the endophallus, can be analyzed morphometrically (e.g., Takami and Suzuki, 2005; Schat et al., 2022). Thus, studies of Carabidae provide opportunities to address the focal issues of insufficient knowledge regarding intromittent membranous parts and the conflation of size and shape effects. Here, two carabid groups, namely the genus Amara and tribe Pterostichini, were studied. In these taxa, within-group morphological diversification has been recognized in both endophallus morphology (Kasahara, 1984; Sasakawa, 2006; Sasakawa and Kubota, 2007) and ejaculate characteristics (Sasakawa, 2006, 2007, 2020). In Pterostichini, a morphological association between the endophallus and spermatophore was reported based on categorical evaluation of morphology (Sasakawa, 2006). Considering these reports and because the endophallus makes physical contact with the ejaculate, the diversification of endophallus morphology may contribute to the diversification of ejaculate morphology. Furthermore, given previous reports regarding sclerotized structures, such contributions may differ between shape and size components. These hypotheses predict that, in phylogenetic comparative analyses among species, morphological traits of the endophallus are determinants of ejaculate traits; moreover, the effects differ between shape and size components. These predictions were tested in Amara and Pterostichini.
Materials and methods
Taxa examined and datasets
This study analyzed 8 Amara species in 4 subgenera, and 22 Pterostichini species in 4 genera and 15 subgenera (Figures 1, 2 and Supplementary Tables 2, 3). The endophallus of Amara is dorsoposteriorly straight, whereas the endophallus of derived lineages of Pterostichini is strongly ventrally bent (Sasakawa, 2006). Examinations of these two groups with contrasting endophallus diversification patterns are expected to provide numerous findings.
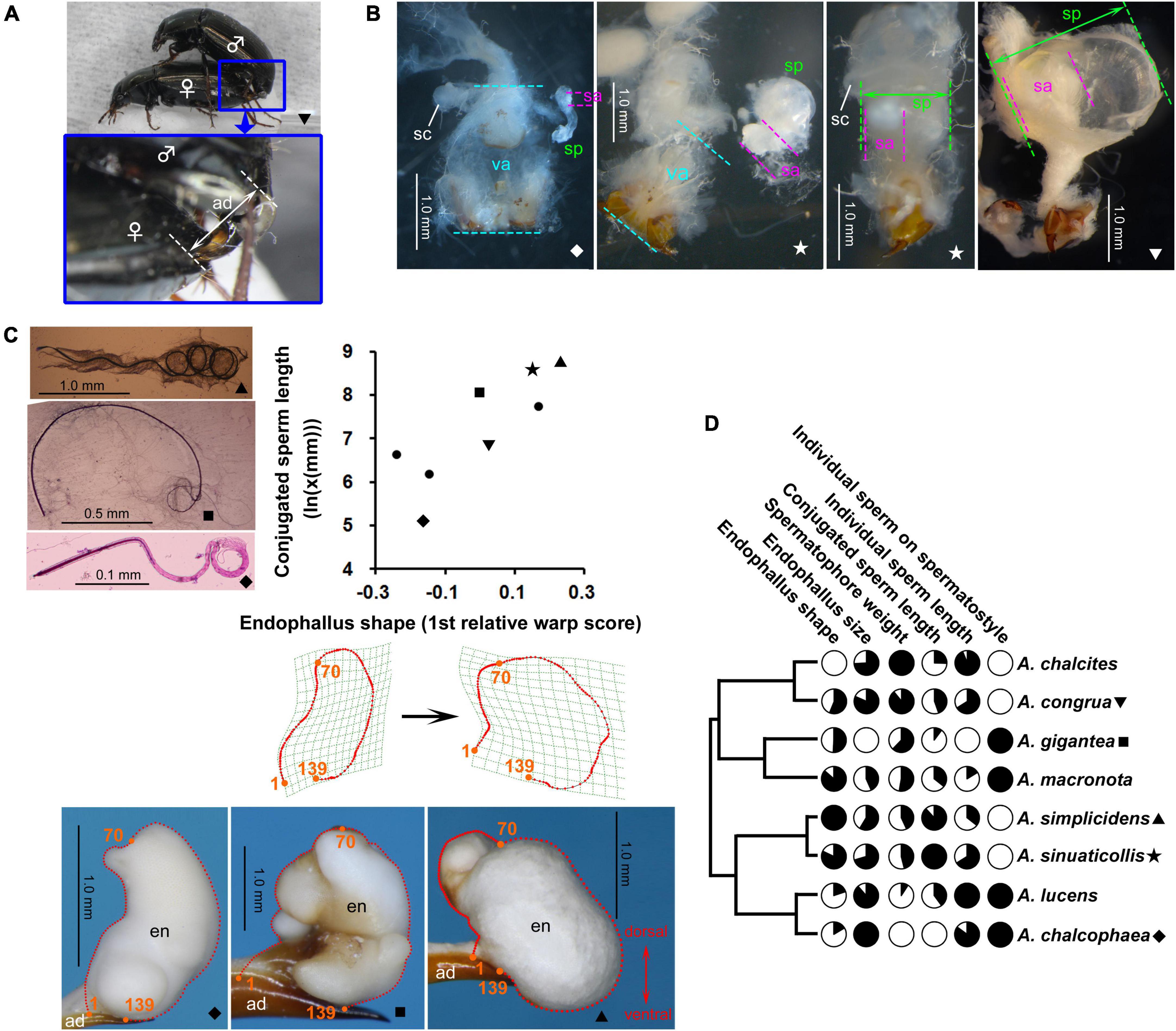
Figure 1. Genital and ejaculate morphologies of Amara. Symbols in panels (A–C) indicate that the trait is associated with the species represented by the same symbol in panel (D). ad, aedeagus; en, endophallus; sa, sperm ampulla; sc, spermatheca; sp, spermatophore; va, vagina. (A) Mating of Amara congrua, showing genital coupling during copulation. (B) Spermatophores and female genitalia, dorsal view; the vaginal wall was partly dissected except in the third image. (C) Giemsa-stained conjugated sperm (upper left), inflated endophallus, left lateral view (bottom), and scatterplot of conjugated sperm length and endophallus shape; thin-plate splines for the endophallus with the smallest and largest scores are visualized below. (D) Phylogeny used for comparative analysis, with variations of trait values (for quantitative size values, where each log-transformed trait value is divided by the log-transformed value of male body length) scaled so that fully white and black circles represent the minimum and maximum values, respectively (cf. Vahed et al., 2014).
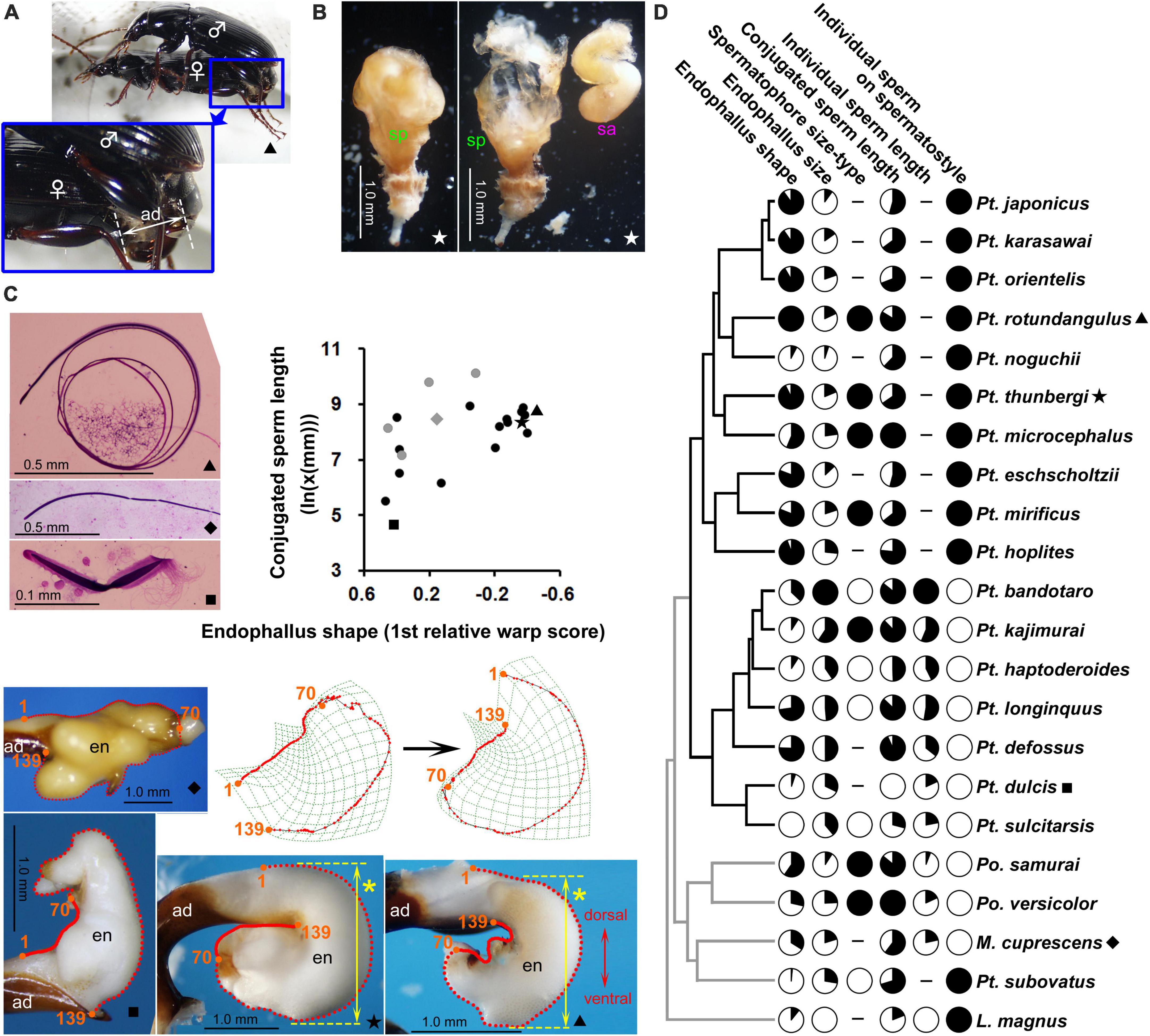
Figure 2. Genital and ejaculate morphologies of Pterostichini. Unless otherwise noted, explanations are identical to those for Figure 1. (A) Mating of Pterostichus rotundangulus. (B) Spermatophores with the right side dissected to show the sperm ampulla. (C) Conjugated sperm, endophallus, and scatterplot of conjugated sperm length and endophallus shape; for the ventrally bent endophallus shape, the dorsoventral width in the bent state is indicated with a yellow arrow and asterisk. (D) Phylogeny used for comparative analysis; considering its evolutionary polarity (Appendix A1), variations of endophallus shape were scaled so that fully white and black circles represent the minimum and maximum values, respectively. In the scatterplot (C) and phylogenetic tree (D), points and branches for species that were not included in the Pterostichus excluding P. subovatus dataset were marked with gray.
For both taxa, the dataset consisted of body size (male body length), endophallus shape, endophallus size, spermatophore size, conjugated sperm length (i.e., length of the axis known as the spermatostyle), individual sperm length, and condition (flagellate or non-flagellate) of individual sperm on the spermatostyle. All data were obtained either from samples newly prepared in this study, from the published literature, or from the identical samples used in previous studies. There were no data concerning the spermatophore size and individual sperm length of some Pterostichini because no data sources were available; such species were therefore excluded from the analyses of those traits. For Pterostichini, separate analyses were also performed for the dataset derived from Pterostichus excluding P. subovatus, for which monophyly was well supported, because uncertainties of evolutionary relationships in the basal lineages may affect results of comparative analyses [see Higginson et al. (2012)].
Examination of traits
New trait data were obtained in accordance with methods described in previous studies (Sasakawa, 2007, 2020). For the phylogenetic comparative analysis conducted here, all qualitative variables were required to be binary. Thus, Pterostichini spermatophore size, which had been classified into three types (Sasakawa, 2006), was converted into a binary variable by merging the two large types [P- and L-types sensu Sasakawa (2006)] into one category. The condition of individual sperm on the spermatostyle was classified as “flagellate type” or “non-flagellate type”; the two types differ mainly in whether the tails of individual sperm move freely or not, respectively.
The endophallus was everted by injecting toothpaste (White & White; LION, Tokyo, Japan) using an insulin syringe with a pre-attached 29-gauge needle (SS-10M2913; TERUMO, Tokyo, Japan), and its fully inflated state was analyzed using two-dimensional geometric morphometrics. Thus, species with a looped endophallus, the contour of which cannot be captured two-dimensionally (i.e., Pterostichus yoritomus, Pterostichus amagisanus, and Pterostichus adstrictus), were excluded from analysis. Scaled digital images of the left lateral side were acquired. The direction of the endophallus was adjusted by locating the following three points in the same horizontal plane: the more ventral of two points connecting the membranous endophallus to the chitinized aedeagus (landmark 1); the gonopore (landmark 70); and the more dorsal of two points connecting the membranous endophallus and chitinized aedeagus (for Amara), or the border between the endophallus and non-endophallus portion of the dorsal contour of the membranous area (for Pterostichini) (landmark 139). Using tpsDIG v2.17 (Rohlf, 2013b), these three points were plotted as landmarks; 136 points distributed along the contours between landmarks were plotted as semi-landmarks. tpsRelw v1.54 (Rohlf, 2013a) was used to convert the raw coordinates into Procrustes coordinates, slide the semi-landmarks along the contours, and perform relative warp analysis. The first relative warp score and centroid size were used as shape and size variables, respectively. In Amara, the endophallus was examined in 3–7 individuals of each species; intraspecific means were used as species values. Using this dataset of Amara, it was statistically confirmed that the effect of individual variations was negligibly small compared to the effect of interspecific variations (Appendix A1). Therefore, only one individual of each species was examined for Pterostichini, first because insufficient specimens were available for some species and second because preparation of the endophallus was difficult in small species.
Spermatophore size of Amara was evaluated by dry weight. Reproductive adults collected in the field and not mated for >2 days were allowed to mate. Immediately after mating, females were dissected; and spermatophore was observed, photographed, dehydrated with absolute ethanol for at >2 days, dried in a desiccator, and measured with a precision balance (UMX2; Mettler Toledo, Paris, France) to the nearest 0.1 μg.
New data regarding conjugated sperm length and individual sperm length were obtained through measurements to the nearest 0.01 μm; data from multiple individuals of a species were pooled after statistically confirming the validity of pooling data.
Quantitative values were averaged within species. Then, the spermatophore weights were converted to their cubic-root values to obtain a unit dimension equivalent to length; quantitative values other than endophallus shape variables, which were coordinates not measurements, were log-transformed.
Phylogenetic comparative analysis
Bayesian phylogenetic trees were constructed with BEAST v1.10.4 (Suchard et al., 2018). For Amara, two mitochondrial (cytochrome c oxidase I and 16S ribosomal RNA) and four nuclear (wingless, arginine kinase, elongation factor-1α, and 28S ribosomal RNA) genes were analyzed. For Pterostichini, wingless and 28S ribosomal RNA gene data were obtained from GenBank. Accession numbers of the new sequences are LC646255–LC646330. Ribosomal genes were aligned with MAFFT v7.273 (qinsi method) (Katoh and Standley, 2013). Optimal nucleotide substitution models were selected using Kakusan4 (Tanabe, 2011) under the BIC4 criterion.
Quantitative variables were analyzed through the phylogenetic generalized least squares method using the phylostep function in the R software package “phylolm” (Ho and Ané, 2014). Qualitative variables were analyzed through phylogenetic logistic regression using the phyloglmstep function in phylolm. These functions select the most parsimonious model based on the Akaike information criterion. In phylogenetic generalized least squares analysis, the λ value estimated using the maximum likelihood method was used as the phylogenetic parameter (“lambda” option). In phylogenetic logistic regression analysis, the “logistic_IG10” model was applied to all ejaculate traits because the phylogenetic parameter α showed non-convergence in all cases (»-4, indicating no phylogenetic signal). For each ejaculate trait, the initial model was constructed using endophallus shape and size as explanatory variables and body size as the covariate; because this study focused on the possible effects of male genital traits on ejaculate morphology, the genital traits were included as explanatory variables not as response variables (see also Higginson et al., 2012). In the model selections, both-direction stepwise selection (“both” option) was performed.
Detailed materials and methods, morphological descriptions, and raw data are available in the Supplementary materials.
Results and discussion
Morphologies of ejaculate and endophallus
The diversification patterns of ejaculate morphology were similar between Amara and Pterostichini (Figures 1, 2). Briefly, in each taxon: spermatophores occupy most of the female vagina or only the innermost half of the vagina; conjugated sperm are formed in all species, and their shapes are spiral, arcuate, or straight (only in Pterostichini); individual sperm are filamentous, without conspicuous interspecific morphological variations; and the condition of individual sperm on the spermatostyle can be classified into the two types described above.
For the endophallus, previously reported diversification patterns (Kasahara, 1984; Sasakawa, 2006; Sasakawa and Kubota, 2007) were confirmed by geometric morphometrics. In Amara, the first relative warp score constituted 47.4% of the total variance and was associated with the width of the endophallus; positive and negative values indicated wider and narrower shapes, respectively (Figure 1C). Because many related carabids have slender endophalli [Platynini: Habu (1978); Pterostichini: Sasakawa and Kubota (2007) and this study; Dolichina: Sasakawa (2006); Synuchina: Morita (2013) and Sasakawa and Itô (2018); Pristosiina and Calathina: Sasakawa et al. (2006)], the slender shape is considered ancestral with evolution toward becoming wider; this character state change corresponds to the switch from negative to positive on the axis of the first relative warp score. In Pterostichini, the first relative warp score constituted 61.8% of the total variance and was associated with bending of the endophallus; positive and negative values indicated straight and ventrally bent shapes, respectively (Figure 2C). Based on ancestral state reconstruction of endophallus shape using molecular phylogenetic trees in a previous study (Sasakawa and Kubota, 2007), the straight shape is considered ancestral and has evolved in the direction toward a strong ventral bend; this character state change corresponds to the shift from positive to negative on the axis of the first relative warp score.
Phylogenetic comparative analysis
Phylogenetic tree used for comparative analyses and trait states are shown in Figures 1D, 2D. Results of phylogenetic comparative analyses are shown in Table 1. In general, associations between traits are detected not only when one trait exerts selective pressure on the other, but also when traits evolve in response to a common selective pressure. Therefore, the results require careful interpretation. Nevertheless, some results imply a direct relationship between ejaculate and endophallus traits.
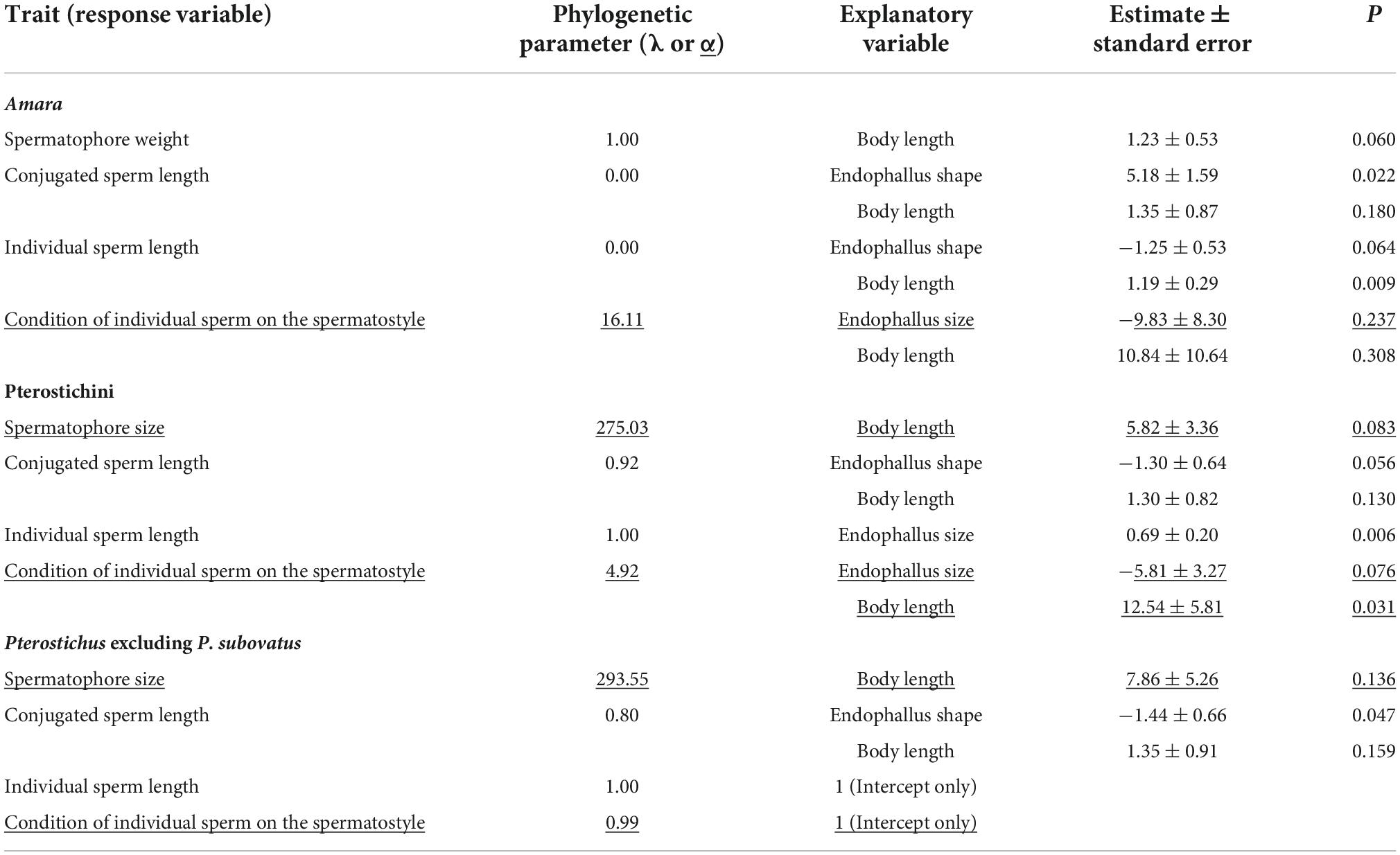
Table 1. Results of stepwise model selection for phylogenetic generalized least squares and phylogenetic logistic regression (underlined) analyses.
The relationship between spermatophore size and endophallus traits was similar in both taxa; neither the shape nor size of the endophallus showed a morphological association with spermatophore size. This finding is in contrast to the relationship between spermatophore size and the intromittent sclerotized part, which has been observed in Ohomopterus carabids and Callosobruchus brucids. In Ohomopterus, the male performs spermatophore displacement with a sclerite on the endophallus, known as the copulatory piece; a positive correlation between spermatophore weight and copulatory piece length was detected in phylogenetic comparative analysis (Takami and Sota, 2007a). In Callosobruchus, the male damages the female vagina with sclerotized spines on the endophallus, and through the resulting wound, the male injects substances that control female reproduction (Hotzy et al., 2012); rankings of spermatophore weight and spine harmfulness have been roughly consistent, although the results were based on species-level data (Rönn et al., 2007, 2011). Thus, morphological associations between the spermatophore and intromittent sclerotized part that might reflect the function of the sclerotized part have been reported. The present finding of no association between spermatophore and endophallus traits implies that the endophallus does not have functions such as spermatophore displacement or wounding of the female genitalia.
The relationships between conjugated sperm length and endophallus traits in Amara and Pterostichini were similar. In both taxa, no effect of endophallus size was detected. Significant and almost significant effects of endophallus shape were detected in Amara and Pterostichini, respectively, and the effects in Pterostichus excluding P. subovatus was significant. In Amara, the transition from a slender to a wide endophallus was associated with elongation of the sperm conjugate. In Pterostichini, the transition from a straight to a ventrally bent endophallus was associated with elongation of the sperm conjugate. Because the ventral bending of the Pterostichini endophallus broadens the dorsoventral width of its overall shape (Figure 2C, yellow arrows), the two taxa show the same associations between conjugated sperm length and endophallus width. Considering the three-dimensional structure and position within the spermatophore of conjugated sperm, one possible explanation for this morphological association is that it reflects physical interactions between conjugated sperm and the endophallus through the spermatophore, as discussed below. In both Amara and Pterostichini, the sperm conjugates of most species, especially long conjugates, have spiral structures (Figures 1C, 2C; Sasakawa, 2007, 2020). As in other insects, conjugated sperm are stored in sperm ampulla at the anterior end of spermatophore (sa in Figures 1B, 2B), in a position corresponding to the opening of the female spermatheca. To accommodate large-diameter spiral-conjugated sperm under the functional constraint that the sperm ampulla remains attached to the anterior end of the spermatophore, the sperm ampulla and spermatophore must be wide. This scenario, together with the observation that the spermatophore is formed by the inflated endophallus (Takami, 2002), can explain the observed association between conjugated sperm length and endophallus shape. Another possible explanation for this association is that conjugated sperm and endophallus shape are evolutionarily correlated because of a confounding factor. The most likely scenario is that the elongation of conjugated sperm and the transition from simple to elaborate endophallus shape occurred independently through sexual selection. Studies in other insects have demonstrated that long sperm and male genitalia of complex shape are favored by sexual selection (Simmons, 2001, 2014). In Ohomopterus, which belongs to the same family as the taxa studied, a positive correlation in length between the sperm conjugate and the copulatory piece–which do not directly interact–was detected in phylogenetic comparative analysis (Takami and Sota, 2007b); this result is similar to the present findings in Amara and Pterostichini.
The results for individual sperm differed from those for the spermatophore and conjugated sperm. In Amara, the association between shorter individual sperm and a wider endophallus was supported with marginal significance. In Pterostichini, endophallus size showed a positive correlation with conjugated sperm length. Thus, the patterns of morphological association differed between the studied taxa, and interpretation of these patterns in terms of functional morphology was difficult. This result is not surprising because the evolutionary patterns of individual sperm are highly variable and often differ among closely related lineages for various reasons. In Carabidae, the length (relative to body length) of individual sperm tends to differ according to conjugation type (Sasakawa, 2020). Thus, the differences in the conjugation type may produce the differences in role of individual sperm and resulting evolutionary patterns. Comparative studies between taxa with different conjugation types may provide some insights into this issue. In addition, the morphology of individual sperm may be influenced by female genitalia, rather than male genitalia. Previous studies have demonstrated that in some insects, individual sperm show correlated evolution with female spermatheca, which is the storage environment for individual sperm, because of their physical interactions (e.g., Morrow and Gage, 2000; Higginson et al., 2012). The taxa examined here also require analysis from this perspective.
Implications for the general mechanism of genital evolution and future studies
In terms of the general mechanism of genital evolution, the present results are notable in two respects. The first is the functional equivalence in male genitalia. The diversification patterns of endophallus shape greatly differed between Amara and Pterostichini. Nevertheless, the evolutionary responses of the spermatophore and conjugated sperm were similar. This similarity implies that the two different types of endophallus, namely wide in Amara and ventrally bent in Pterostichini, are functionally equivalent. No such evolutionary pattern has yet been explicitly reported for genital intromittent parts. The second and more important result is the similarity of evolutionary patterns among the non-intromittent part, intromittent sclerotized part, and intromittent membranous part. Many studies have examined the morphology of male genitalia, but few have distinguished shape and size components. Moreover, no studies have yet examined intromittent membranous parts. With respect to non-intromittent parts, studies based on mating experiments have been reported for four species: the dung beetle Onthophagus taurus (Simmons et al., 2009; Simmons and Garcia-Gonzalez, 2011), the praying mantis Ciulfina klassi (Holwell et al., 2010), the fruit fly Drosophila simulans (House et al., 2013, 2021), and the broad-horned flour beetle Gnatocerus cornutus (House et al., 2016; note that the organ regarded as the intromittent part by the authors is actually a non-intromittent part, the aedeagus). In O. taurus and C. klassi, effects on reproductive success were detected for shape but not size. In D. simulans and G. cornutus, effects were detected for both shape and size, but their direction and strength differed. Concerning the intromittent sclerotized part, mating experiment studies have been conducted in two species: the water strider Gerris lateralis (Arnqvist and Danielsson, 1999) and O. taurus (House and Simmons, 2003; Simmons et al., 2009). In G. lateralis, the only effects detected were for shape. In O. taurus, effects were detected for both shape and size, but their direction and strength differed. In the water strider genus Gerris, although no explicit comparison of shape and size was performed, the evolutionary patterns of shape and size of the non-intromittent and intromittent sclerotized parts (in the context of sexual selection) were examined through phylogenetic comparative analysis; based on quantitative analysis of shape and complexity, and allometric analyses of size, the authors concluded that shape is more diverse than size for both the non-intromittent part and the intromittent sclerotized part (Rowe and Arnqvist, 2012). Thus, these reports indicate that shape and size have different effects in both the non-intromittent part and the intromittent sclerotized part, and shape appears to be more important than size (note that shape was included as a factor in all cases). Importantly, these results are consistent with the present findings, in which the shape, but not size, of the intromittent membranous part was associated mainly with ejaculate morphology. The function and diversification mechanism of the intromittent membranous part may be similar to those of the non-intromittent part and the intromittent sclerotized part.
The present results provide new insights into the function and diversification of the intromittent membranous part of male genitalia; they demonstrate the importance of examining this genital component in future studies, but several methodological issues remain unresolved. One is the lack of single-species studies directly examining the selection pressure on traits through analysis of relationships between trait variations and reproductive success. Such analysis is essential for the verification of correlated evolution detected in phylogenetic comparative analysis (Rönn et al., 2007; Takami and Sota, 2007a); the present results for Amara and Pterostichini require confirmation through this approach. Single-species studies use either intraspecific genetic variations or artificial variations generated through methods such as partial ablation of genitalia (e.g., Hotzy et al., 2012). Generally, the artificial variation process is more convenient because it allows finer control of variations and the evaluation of various hypotheses using those variations (e.g., Wulff and Lehmann, 2016). However, this method cannot be applied to the intromittent membranous part because partial ablation would cause hemorrhage and subsequent death. New methods are needed to overcome this problem.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
KS designed the study, carried out the field and laboratory work, and wrote the manuscript.
Funding
This study was partly supported by JSPS KAKENHI (nos. 20-11227, 25830150, and 17K15171).
Acknowledgments
The author thank M. Ito and M. Shimada (University of Tokyo) for their laboratory facilities, J. Serrano (Universidad de Murcia, Spain) and J. Ogawa (Tokyo) for sample collections, H. Ikeda (Hirosaki University) for helpful advice on data analyses, and two reviewers for helpful comments.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.1032378/full#supplementary-material
References
Arnqvist, G. (1998). Comparative evidence for the evolution of genitalia by sexual selection. Nature 393, 784–786.
Arnqvist, G., and Danielsson, I. (1999). Copulatory behavior, genital morphology, and male fertilization success in water striders. Evolution 53, 147–156. doi: 10.2307/2640927
Habu, A. (1978). Fauna japonica. Carabidae: Platynini (Insecta: Coleoptera). Tokyo: Keigaku Publishing Co Ltd.
Higginson, D. M., Miller, K. B., Segraves, K. A., and Pitnick, S. (2012). Female reproductive tract form drives the evolution of complex sperm morphology. Proc. Natl. Acad. Sci. U.S.A. 109, 4538–4543. doi: 10.1073/pnas.1111474109
Ho, L. S. T., and Ané, C. (2014). A linear-time algorithm for Gaussian and non-Gaussian trait evolution models. Syst. Biol. 63, 397–408. doi: 10.1093/sysbio/syu005
Holwell, G. I., Winnick, C., Tregenza, T., and Herberstein, M. E. (2010). Genital shape correlates with sperm transfer success in the praying mantis Ciulfina klassi (Insecta: Mantodea). Behav. Ecol. Sociobiol. 64, 617–625. doi: 10.1007/s00265-009-0879-2
Hosken, D. J., and Stockley, P. (2004). Sexual selection and genital evolution. Trends Ecol. Evol. 19, 87–93. doi: 10.1016/j.tree.2003.11.012
Hotzy, C., Polak, M., Rönn, J. L., and Arnqvist, G. (2012). Phenotypic engineering unveils the function of genital morphology. Curr. Biol. 22, 2258–2261. doi: 10.1016/j.cub.2012.10.009
House, C. M., Lewis, Z., Hodgson, D. J., Wedell, N., Sharma, M. D., Hunt, J., et al. (2013). Sexual and natural selection both influence male genital evolution. PLoS One 8:e63807. doi: 10.1371/journal.pone.0063807
House, C. M., Lewis, Z., Sharma, M. D., Hodgson, D. J., Hunt, J., Wedell, N., et al. (2021). Sexual selection on the genital lobes of male Drosophila simulans. Evolution 75, 501–514. doi: 10.1111/evo.14158
House, C. M., Sharma, M. D., Okada, K., and Hosken, D. J. (2016). Pre and post-copulatory selection favor similar genital phenotypes in the male broad horned beetle. Integr. Comp. Biol. 56, 682–693. doi: 10.1093/icb/icw079
House, C. M., and Simmons, L. W. (2003). Genital morphology and fertilization success in the dung beetle Onthophagus taurus: An example of sexually selected male genitalia. Proc. R. Soc. Lond. B Biol. Sci. 270, 447–455. doi: 10.1098/rspb.2002.2266
Janovska, M., Anichtchenko, A. V., and Erwin, T. (2013). Significant new taxonomic tool for Carabidae (Insecta: Coleoptera): Endophallus inflation methods revised. Caucas. Entomol. Bull. 9, 39–42. doi: 10.23885/1814-3326-2013-9-1-39-42
Kasahara, S. (1984). On the differences between Amara chalcites Dejean and A. congrua Morawitz (Coleoptera: Carabidae). Coleop. News 64, 9–11.
Katoh, K., and Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Morita, S. (2013). A new Synuchus (Coleoptera, Carabidae) from the Kii Peninsula, central Japan, with a redescription of S. ventricosus. Elytra New Ser. 3, 1–7.
Morrow, E. H., and Gage, M. J. G. (2000). The evolution of sperm length in moths. Proc. R. Soc. Lond. B Biol. Sci. 267, 307–313. doi: 10.1098/rspb.2000.1001
Rohlf, F. J. (2013a). tpsDig, digitize landmarks and outlines, version 2.17 [software and manual]. Department of ecology and evolution. New York, NY: State University of New York.
Rohlf, F. J. (2013b). tpsRelw, relative warps analysis, version 1.53 [software and manual]. Department of ecology and evolution. New York, NY: State University of New York.
Rönn, J., Katvala, M., and Arnqvist, G. (2007). Coevolution between harmful male genitalia and female resistance in seed beetles. Proc. Natl. Acad. Sci. U.S.A. 104, 10921–10925. doi: 10.1073/pnas.0701170104
Rönn, J., Katvala, M., and Arnqvist, G. (2011). Correlated evolution between male and female primary reproductive characters in seed beetles. Funct. Ecol. 25, 634–640. doi: 10.1111/j.1365-2435.2010.01809.x
Rowe, L., and Arnqvist, G. (2012). Sexual selection and the evolution of genital shape and complexity in water striders. Evolution 66, 40–54. doi: 10.1111/j.1558-5646.2011.01411.x
Sasakawa, K. (2006). Morphological association between spermatophores and male genitalia in carabid beetles of the tribe Pterostichini (Coleoptera: Carabidae). Zool. Sci. 23, 587–591. doi: 10.2108/zsj.23.587
Sasakawa, K. (2007). Sperm bundle and reproductive organs of carabid beetles tribe Pterostichini (Coleoptera: Carabidae). Naturwissenschaften 94, 384–391. doi: 10.1007/s00114-006-0200-4
Sasakawa, K. (2020). Light-microscopic examinations of sperm in several lineages of Carabidae (Insecta: Coleoptera): Implications for the evolution of sperm conjugation. Entomol. Sci. 23, 338–348. doi: 10.1111/ens.12426
Sasakawa, K., and Itô, H. (2018). Taxonomic notes on the ground beetles in the genus Trephionus Bates, 1883 from central Honshu, Japan (Coleoptera, Carabidae, Sphodrini, Synuchina). Zookeys 742, 91–104. doi: 10.3897/zookeys.742.23752
Sasakawa, K., Kim, J. L., Kim, J. K., and Kubota, K. (2006). Descriptions of two new species of Pristosia (Coleoptera: Carabidae) from South Korea. Ann. Entomol. Soc. Am. 99, 1006–1011.
Sasakawa, K., and Kubota, K. (2007). Phylogeny and genital evolution of carabid beetles in the genus Pterostichus and its allied genera (Coleoptera: Carabidae) inferred from two nuclear gene sequences. Ann. Entomol. Soc. Am. 100, 100–109.
Schat, J., Weng, Y. M., Dudko, R. Y., Kavanaugh, D. H., Luo, L., and Schoville, S. D. (2022). Evidence for niche conservatism in alpine beetles under a climate-driven species pump model. J. Biogeogr. 49, 364–377. doi: 10.1111/jbi.14307
Simmons, L. W. (2001). Sperm competition and its evolutionary consequences in the insects. Princeton, NJ: Princeton University Press.
Simmons, L. W. (2014). Sexual selection and genital evolution. Austral Entomol. 53, 1–17. doi: 10.1111/aen.12053
Simmons, L. W., and Garcia-Gonzalez, F. (2011). Experimental coevolution of male and female genital morphology. Nat. Commun. 2:374. doi: 10.1038/ncomms1379
Simmons, L. W., House, C. M., Hunt, J., and Garcia-Gonzalez, F. (2009). Evolutionary response to sexual selection in male genital morphology. Curr. Biol. 19, 1442–1446. doi: 10.1016/j.cub.2009.06.056
Snodgrass, R. E. (1935). ). Principles of insect morphology. New York, NY: Cornell University Press.
Suchard, M. A., Lemey, P., Baele, G., Ayres, D. L., Drummond, A. J., and Rambaut, A. (2018). Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 4:vey016. doi: 10.1093/ve/vey016
Takami, Y. (2002). Mating behavior, insemination and sperm transfer in the ground beetle Carabus insulicola. Zool. Sci. 19, 1067–1073. doi: 10.2108/zsj.19.1067
Takami, Y., and Sota, T. (2007a). Rapid diversification of male genitalia and mating strategies in Ohomopterus ground beetles. J. Evol. Biol. 20, 1385–1395. doi: 10.1111/j.1420-9101.2007.01338.x
Takami, Y., and Sota, T. (2007b). Sperm competition promotes diversity of sperm bundles in Ohomopterus ground beetles. Naturwissenschaften 94, 543–550. doi: 10.1007/s00114-007-0225-3
Takami, Y., and Suzuki, H. (2005). Morphological, genetic and behavioural analyses of a hybrid zone between the ground beetles Carabus lewisianus and C. albrechti (Coleoptera, Carabidae): Asymmetrical introgression caused by movement of the zone? Biol. J. Linn. Soc. 86, 79–94. doi: 10.1111/j.1095-8312.2005.00527.x
Tanabe, A. S. (2011). Kakusan4 and Aminosan: Two programs for comparing nonpartitioned, proportional, and separate models for combined molecular phylogenetic analyses of multilocus sequence data. Mol. Ecol. Resour. 11, 914–921. doi: 10.1111/j.1755-0998.2011.03021.x
Vahed, K., Gilbert, J. D., Weissman, D. B., and Barrientos-Lozano, L. (2014). Functional equivalence of grasping cerci and nuptial food gifts in promoting ejaculate transfer in katydids. Evolution 68, 2052–2065. doi: 10.1111/evo.12421
Wenninger, E. J., and Averill, A. L. (2006). Influence of body and genital morphology on relative male fertilization success in oriental beetle. Behav. Ecol. 17, 656–663. doi: 10.1093/beheco/ark013
Wulff, N. C., and Lehmann, G. U. C. (2016). Function of male genital titillators in mating and spermatophore transfer in the tettigoniid bushcricket Metrioptera roeselii. Biol. J. Linn. Soc. 117, 206–216. doi: 10.1111/bij.12661
Keywords: conjugated sperm, functional morphology, geometric morphometrics, individual sperm, phylogenetic comparative analysis, spermatophore, endophallus, genital evolution
Citation: Sasakawa K (2022) Possible differences in effects on ejaculate morphology between shape and size components of the intromittent membranous part of insect male genitalia: Phylogenetic comparative analyses in Carabidae. Front. Ecol. Evol. 10:1032378. doi: 10.3389/fevo.2022.1032378
Received: 30 August 2022; Accepted: 12 October 2022;
Published: 28 October 2022.
Edited by:
Lenin Arias Rodriguez, Universidad Juárez Autónoma de Tabasco, MexicoReviewed by:
Sergio Pérez, La Salle Centro Universitario, SpainYoko Matsumura, Hokkaido University, Japan
Copyright © 2022 Sasakawa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kôji Sasakawa, a3Nhc2FAY2hpYmEtdS5qcA==