 Florina Stănescu
Florina Stănescu Rafael Márquez
Rafael Márquez Dan Cogălniceanu
Dan Cogălniceanu Federico Marangoni
Federico Marangoni- 1Black Sea Institute for Development and Security Studies, Ovidius University Constanţa, Constanţa, Romania
- 2Center for Morphological and Genetic Studies of Malignant Pathology (CEDMOG), Ovidius University Constanţa, Constanţa, Romania
- 3Fonoteca Zoológica, Departamento de Biodiversidad y Biología Evolutiva, Museo Nacional de Ciencias Naturales-CSIC, Madrid, Spain
- 4Faculty of Natural and Agricultural Sciences, Ovidius University Constanţa, Constanţa, Romania
- 5Departamento de Biología, Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste (FACENA-UNNE), Corrientes, Argentina
- 6Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Corrientes, Argentina
Courtship acoustic displays in anuran amphibians are energetically costly and risky, but have a major role in mating success since they encode relevant information regarding the caller’s identity and status. Age and size are essential traits shaping fitness, reproductive success and life-history strategies, and thus are expected to also have a role in courtship displays. We tested this assumption in a species of nest-building frogs, Leptodactylus bufonius, in northern Argentina. We conducted the first detailed quantitative description of the males’ mating calls and assessed the effects of biological traits (i.e., body size parameters and individual age) and local climate (i.e., air temperature and humidity) on the main acoustic features of these calls (i.e., call duration, inter-call duration, dominant frequency, and dominant frequency modulation). The calls were short (mean ± SE, 0.163 ± 0.004 s), whistle-like, single notes with harmonic structure. The dominant frequency (1381.7 ± 16.2 Hz) decreased with arm length (χ2 = 5.244, df = 1, p = 0.022) and had an upward modulation (456.4 ± 11.0 Hz) which increased with age (χ2 = 4.7012, df = 1, p = 0.030). Call duration and dominant frequency were the most static parameters at intra-individual level, indicating their role in individual recognition. Temperature and humidity shaped the temporal acoustic parameters, and the dominant frequency. Our findings suggest that the acoustic features of the mating calls in amphibians could promote female mate choice in relation to both size and age and open up new questions for future research: are females more attracted to older males, and what are the specific costs and benefits? We suggest that mating calls may direct female preferences toward males of certain size and age classes, ultimately shaping the life-history strategies in a given population. Finally, we found discrepancies in the mating calls of L. bufonius recorded from Corrientes and those previously described from other populations, which suggests that multiple species may have been recorded under the same name.
Introduction
Mating calls are among the most conspicuous components of courtship displays in anurans and are subject to high selective forces through male competition, predation, and female mate choice (Ryan, 1991; Duellman and Trueb, 1994; Gerhardt, 1994; Wells, 2007). The acoustic properties of the mating calls are of paramount importance to reproductive success (Gerhardt, 1994; Narins et al., 2006) and proved to be reliable indicators of heritable male traits that predict offspring fitness in terms of size, growth rates and efficiency, or performance (Woodward et al., 1988; Welch et al., 1998; Doty and Welch, 2001; Jaquiéry et al., 2010). Acoustic signaling is energetically costly (Ryan, 1991; Gerhardt, 1994; Taigen et al., 1996; Grafe and Thein, 2001; Brepson et al., 2013) and exposes the caller to higher predation risks, by revealing its location (Wells, 2007). Given these costs, it is expected that the acoustic features of the calls must encode the most important information regarding the caller’s traits in relation to its ability to yield successful offspring, and thus contribute to mating success (Ryan, 1988, 1991; Gerhardt, 1994; Welch et al., 1998; Narins et al., 2006; Achorn and Rosenthal, 2020). Age and size are essential traits in amphibians, governing fitness and life-history strategies (Blanckenhorn, 2000; Stearns, 2000), and are also major determinants of reproductive success (e.g., Manning, 1985; Kokko and Lindström, 1996; Wells, 2007). Thus, a relation between male sexual advertisement displays like mating calls and individual age and size is expected (Kokko, 1997; Felton et al., 2006; Kelleher et al., 2021).
Leptodactylus bufonius (Boulenger, 1894) is a common anuran in northern Argentina, also occurring in south-eastern Bolivia, most of Paraguay, and in the Pantanal region of Brazil (Heyer et al., 2004). It belongs to the Leptodactylus fuscus group, which includes terrestrial frogs where males construct mud nests at the edge of temporary ponds and other low-lying depressions (Heyer, 1969; Wells, 2007). L. bufonius males typically build cone-shaped nests that rise up from the ground, near semi-permanent or ephemeral bodies of water, and then call from inside or close to the nests, to attract the females (Philibosian et al., 1974; Crump, 1995; Reading and Jofré, 2003; Faggioni et al., 2017). Inside the mud nest, the female lays the eggs during the axillary amplexus, while the male produces a protective foam enveloping the eggs; the nest is usually sealed with a mud cap afterward (Cei, 1949; Philibosian et al., 1974; Crump, 1995). The eggs hatch within a few days, but the tadpoles remain within the closed mud nest until rain floods wash them into the nearest water body (Philibosian et al., 1974).
Some authors (Philibosian et al., 1974; Reading and Jofré, 2003) noted that after construction, most of the nests are left to dry and thus decrease their permeability, with males returning to the nest site later, before starting calling activity. Thus, males can play an essential role in offspring survival through their choice of nest site location and the quality of nest construction. While the courtship behavior of this species was thoroughly described (e.g., Faggioni et al., 2017), this is not the case for the associated acoustic repertoire. Meanwhile, the mating success in this species depends on the males’ capacity to attract females to the nesting grounds through acoustic signals. Descriptions of the acoustic properties of the mating calls are mostly qualitative, with scant quantitative data restricted to one or few individuals (Cei, 1949; Barrio, 1965; Philibosian et al., 1974; Straughan and Heyer, 1976; Heyer, 1978; Heyer and Scott, 2006; Schalk and Leavitt, 2017). We provide here the first thorough quantitative description of the most important acoustic parameters of the mating calls in L. bufonius males from a population near Corrientes, northern Argentina, and assess how these parameters are shaped by biological traits (i.e., size parameters and individual age) and local climate conditions (i.e., air temperature and humidity). Given the complex reproductive behavior in this species (Faggioni et al., 2017), we expect that mating calls of L. bufonius males should encode information related to individual age and size-related parameters, as potential proxies of male vigor (i.e., older males have better survival skills, while larger males can outcompete smaller individuals), and experience or abilities in nest building and/or nest site choice (Reading and Jofré, 2003).
Materials and methods
Field work was carried out by FM during autumn 2007 – spring 2008, about 10 km northeast from Corrientes city, Argentina, within an area with numerous temporary and semi-permanent ponds called El Perichón (−27.434403, −58.747844). A more detailed description of the study area is provided by Marangoni et al. (2019). The mating calls were recorded in the field for 18 L. bufonius males during evening hours, from 20:00 to 22:00, using a Sony TCM-313 tape recorder. Air temperature and humidity were recorded at the beginning of each individual recording with a hand-held thermo-hygrometer. The corresponding individual age and body size measurements (i.e., body mass, snout-vent length, head width, right hind-limb length, arm length, and tibia length) for the 18 adult males were obtained from Marangoni et al. (2019). Briefly, snout-vent length and right hind-limb length were measured on laminated graph paper (accuracy ±1 mm), while head width, arm length, and tibia length were measured to the nearest 0.1 mm with digital calipers. Body mass was measured to the nearest 0.01 g with an electronic balance (Acculab Scales, Titusville, NJ, USA). Age was estimated through skeletochronology using the third toe of the right leg which was clipped and stored in 70% alcohol. The bone tissue was processed following a standard protocol for cryotomy (Marangoni et al., 2009). All applicable international, national, and/or institutional guidelines for the care and use of animals were strictly followed. All animal sample collection protocols complied with the current laws of Dirección de Recursos Naturales of Corrientes province (Argentina).
Recordings were normalized (peak −1.0 dB) with Audacity v. 2.1.2 software (Audacity Team, 2015). We performed the acoustic analyses using the software Raven Pro 1.5 (Bioacoustics Research Program, 2017; Cornell Lab of Ornithology). We measured call duration (CD) and inter-call duration (ICD) in the oscillograms with the function “Delta Time (s).” The spectral acoustic parameters were measured in spectrograms produced by fast Fourier Transformation (width, 512 points; 50% window overlap): the dominant frequency (DF) with the function “Max Frequency (Hz)”, and the dominant frequency modulation (DFmodulation) (Márquez et al., 1996; Bosch and de la Riva, 2004) as the difference between the maximum and minimum frequencies of the peak frequency contour, with the functions “Peak Frequency Contour Max Frequency” and “Peak Frequency Contour Min Frequency.” We determined the coefficient of variation (CV%=SD/mean×100) at inter-individual (CVinter) and intra-individual (CVintra) level, to differentiate between static (CVinter < 10%, CVintra < 5%) and dynamic (CVinter > 20%, CVintra > 12) acoustic parameters (Gerhardt, 1991; de Oliveira-Santos et al., 2022).
We conducted the statistical analyses in R v. 3.6.0 (R Core Team, 2019) with an a priori level of significance of 0.05. We fitted generalized least squares (GLS) models with the maximized log-likelihood method to assess the effects of biological traits and climate conditions on the acoustic parameters, with the package “nlme” (Pinheiro et al., 2019), function “gls.” We first assessed the collinearity among the potential predictor variables (i.e., age, all size parameters, air temperature, and humidity) and the potential correlation relationships between these predictors and the acoustic parameters by two-tailed Pearson’s r and Spearman’s rho tests with the function “chart.Correlation” in “PerformanceAnalytics” package (Peterson and Carl, 2020). We fitted GLS models for each of the acoustic parameters, using as predictors those biological and/or climate traits which appeared to be significantly correlated to the acoustic parameters (Supplementary Figures 1, 2). We also tested supplementary models with age and body mass, and combinations of air temperature, humidity and biological traits as predictors. Body mass was added as a predictor since it was previously demonstrated that it can influence the acoustic properties of calls (Ryan, 1988; Gillooly and Ophir, 2010; Ophir et al., 2010); body mass also had the highest correlation with the other size parameters (Supplementary Figures 1, 2). Since the sample size was small, we fitted the models with one predictor at a time and then grouped two by two. In total, we obtained 38 models, each representing a testing hypothesis regarding the effects of local climate and biological traits on the acoustic features of the mating calls (see Supplementary material). We then compared and selected the best fitted models based on Akaike’s information criterion corrected for sample size (AICc) following Burnham and Anderson (2002) and Mazerolle (2020), with the “aictab” function in the package “AICcmodavg” (Mazerolle, 2020). We assessed the significance of predictor variables from the best fitted models with Wald Chi-square tests (type III) with the function “Anova” in the package “car” (Fox and Weisberg, 2019). The goodness of fit of these models was assessed by inspecting the residuals.
Results
A total of 194 calls (min–max = 7–16, mean = 10.8, SE = 0.5) were analyzed for 18 males. Temperature and humidity varied during call recordings: min–max = 17.5–29.4°C, mean = 23.1°C, SE = 0.7; min–max = 65–98%, mean = 89.9%, SE = 1.8. Skeletochronology was successful in all 18 individuals. Individual age varied from one to 4 years old, with the 2-year old age class being the best represented. The summary of age and body size data is provided in Supplementary Table 1 and Supplementary Figure 3. The raw dataset containing the individual age, morphometric and acoustic parameters is provided as a Supplementary Data Sheet.
Each call consisted of a single, short, whistle-like note with harmonic structure and an upward frequency modulation (Figure 1 and Table 1). Most of the energy was concentrated at the beginning of the note, producing a horizontal T-shaped pattern in the oscillogram (Figure 1), but note that the recording level control of the recorder may have acted in shaping the envelope. The DF was also the fundamental frequency. At the intra-individual level, the ICD was the most dynamic acoustic parameter, while CD and DF were the most static; at inter-individual level, all acoustic parameters were highly variable (Table 1).
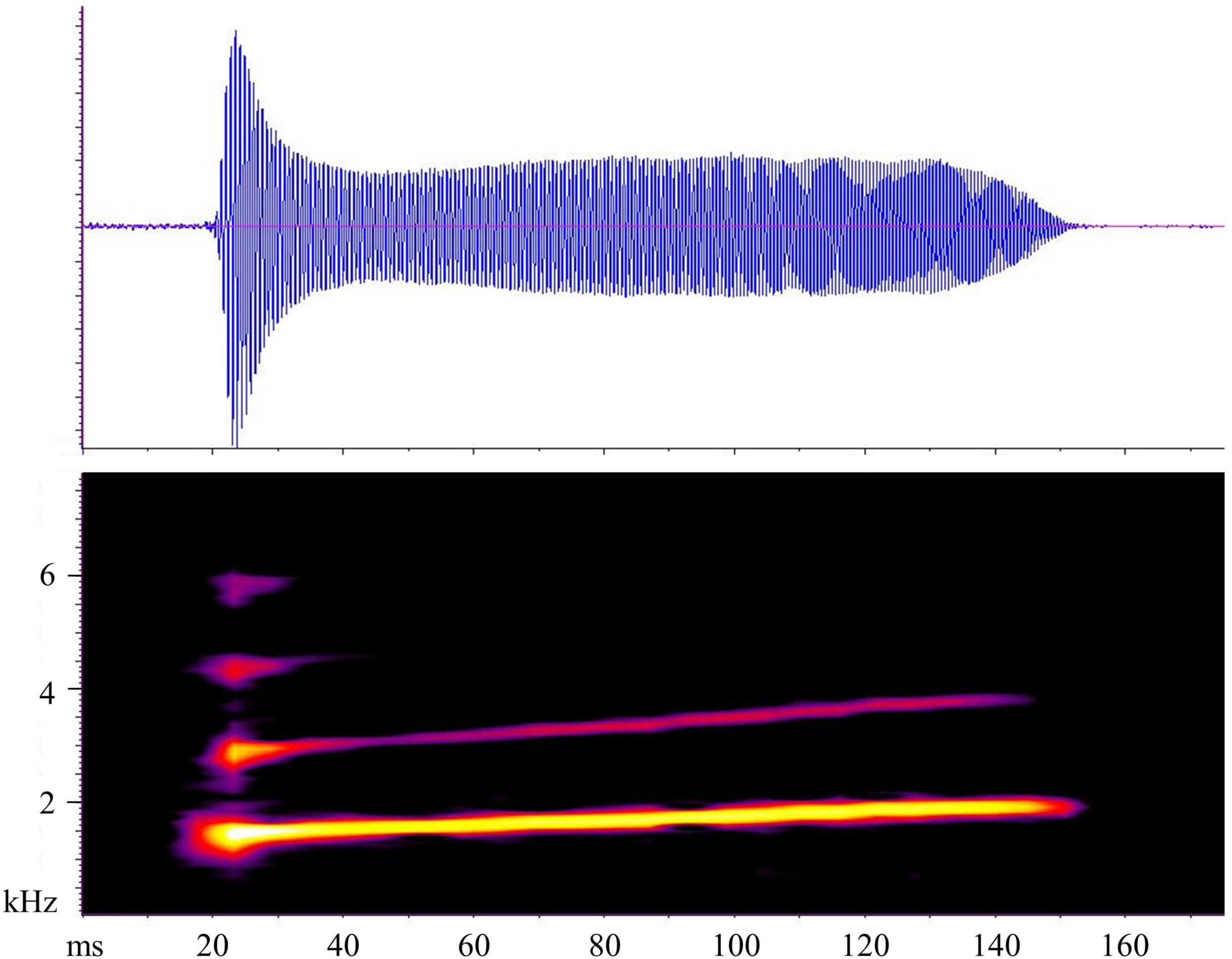
Figure 1. Graphical representation of one mating call of L. bufonius from Corrientes (Argentina) analyzed in this study. The upper part represents the oscillogram, and the lower part represents the audiospectrogram.
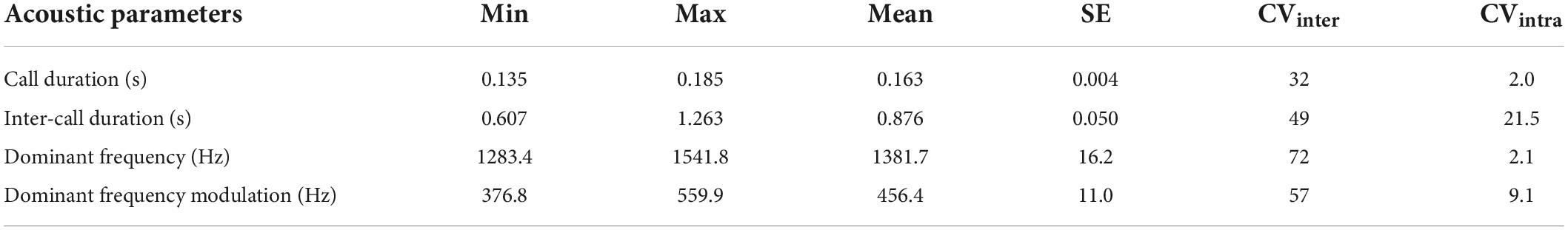
Table 1. Descriptive statistics of the acoustic traits of the mating calls in 18 male L. bufonius from Corrientes, Argentina.
The temporal acoustic properties were best predicted by air temperature and humidity, in contrast with the DF modulation, which was best explained by the individual age of the calling male; the DF was in turn best predicted by a combination of climate conditions and biological traits (i.e., humidity and arm length) (Supplementary Tables 1, 2). Air temperature had a significant negative effect on CD (χ2 = 8.524, df = 1, p = 0.003), while ICD was negatively affected by both air temperature (χ2 = 21.938, df = 1, p < 0.001) and humidity (χ2 = 7.180, df = 1, p = 0.007). Humidity (χ2 = 8.218, df = 1, p = 0.004) and arm length (χ2 = 5.244, df = 1, p = 0.022) had significant negative effects on the DF. The DF modulation significantly increased with age, older males producing calls with a higher frequency modulation (χ2 = 4.7012, df = 1, p = 0.030).
Discussion
Our study provides the first detailed quantitative description of mating calls in L. bufonius males and shows how local climate, age and size shape the acoustic properties of these calls. CD and DF are likely to be some of the most important features of mating calls (Duellman and Trueb, 1994; Welch et al., 1998; Wells, 2007). These traits were the most static at intra-individual level, followed by the DF modulation (Table 1), indicating their potential role in individual recognition and their potential for female preference. Individual characteristics may be the driving force for sexual selection; in some anurans, directional female preference may occur for static parameters related to male size (Márquez, 1995). Individual recognition is also important in chorus settings, allowing callers to discriminate between other neighboring males and intruders who might pose a risk in terms of maintaining the breeding territory or access to potential female partners (Wells, 2007).
The temporal acoustic parameters were positively correlated with size, indicating that larger males emitted longer calls, at slower rates (Supplementary Figures 1, 2). However, this relationship was weak compared to the effects of air temperature and humidity on the same acoustic features, which acted in the opposite direction (Supplementary Table 3). Previous female mate choice experiments showed that the effects of temperature on the acoustic parameters of the mating calls were canceled when the calling males and the females experienced the same ambient temperature, thus by temperature coupling of signal production and signal reception (Ryan, 1988). Previous research also showed that local climate can interfere with both sound production and reception in amphibians (Duellman and Trueb, 1994; Narins et al., 2006). In L. bufonius, higher temperature and humidity predicted shorter calls, emitted at higher rates (i.e., shorter ICD). Since amphibians are ectotherms, this is especially important under the forecasted global warming scenarios, which could lead to more energetically costly acoustic displays (i.e., calling at higher temperatures may increase the overall energy budget of the activity); this may have an effect on female acoustic preferences (Wells, 2007) and thus affect mating success.
Dominant frequency significantly decreased with increasing humidity and arm length. There is a well-established relationship between size and call frequency among species of anurans, and within species there are many examples of correlations between size and frequency (e.g., Márquez, 1995; Moser et al., 2022). In some species, males that emit calls with lower dominant frequencies appear to be more attractive to females and have a higher mating and fertilization success (Wells, 2007; Kelleher et al., 2022). In L. bufonius, a lower frequency is characteristic of larger males with longer arms. This is an expected result for this species, since arm length is an important trait for both nest building and for securing the amplexus. While males use their snouts to dig the nest, they have to use their arms as levers, to help push the mud away. A strong grip over the female is also important, since during the axillary amplexus, the male uses its hind limbs to froth the mucus mass released by the female into a protective foam for the eggs (Reading and Jofré, 2003).
In contrast with the other acoustic traits, the DF modulation was unaffected by air temperature and humidity, while also carrying information regarding the caller’s age. Some studies (Kokko and Lindström, 1996; Brooks and Kemp, 2001) suggest that older individuals may be more attractive to females simply by their ability to survive for a longer time. A longer survival can be the result of access to better resources, the ability to avoid or escape predators, or higher resistance to pathogens and/or parasites, and as such can also increase the chances of survival and for a better fitness of future offspring (Kelleher et al., 2021). While we do not know yet if females of this species actually prefer older males, we suggest that individual age in L. bufonius males could be an indicator of their experience in nest construction and/or in choosing the nest construction site, both of which are important determinants of offspring survival. For example, if the nest is too far from the future ponds, the tadpoles might either not make it into the water, and thus desiccate and die, or reach the water too late and be outcompeted by older and larger tadpoles already present in the water (Wells, 2007). Similarly, if the nest is not well constructed, the foam and eggs might desiccate or be exposed to predators. Since the DF modulation was not affected by the local climate and had a relatively low variation within the calling males, we suggest that this acoustic feature and the information encoded within should play a significant role in the mating success of this species.
Our findings suggest that the mating calls in L. bufonius could promote female mate choice in relation to both size and age. It is noteworthy that size and age were encoded separately within the two spectral parameters analyzed, which is in line with previous results that showed that age and size do not follow a linear relationship in this species, meaning that larger individuals are not necessarily older (Marangoni et al., 2019). In addition, both spectral parameters were static, favoring individual recognition (Gerhardt, 1991). Thus, females of this species are provided with the necessary information to opt for smaller/larger males from younger/older age classes, which may bring combined advantages stemming from both traits and ultimately favor males with certain life-history strategies (Table 2). Based on a previous study (Marangoni et al., 2019) regarding the life-history strategies of the same population of L. bufonius (Corrientes, Argentina), the prevalent strategy observed was “fast growth-early sexual maturity.” This strategy is in line with the reproductive behavior and ecology of the species, since the breeding events and reproductive success are highly dependent on the seasonal precipitation regimes and water bodies with high desiccation rates. Reaching a large size at sexual maturity is especially important in amphibians, since the growth rate decreases dramatically after this point, with most of the organism’s resources being redirected toward reproduction and survival (Heino and Kaitala, 1999). Thus, it seems more likely that females would favor large, young mates that may in turn contribute to produce offspring able to grow fast and mature early at larger sizes (Castellano and Cadeddu, 2011). Mating with large, old males could also be favored, but the life-history advantages are not so obvious, since the large size could have been attained at slower growth rates, at the cost of later sexual maturity, which may be beneficial only under certain circumstances (i.e., plentiful resources, little competition, and predation risk).

Table 2. Potential trait combination (i.e., body size and age) and associated life-history strategies encoded within the spectral properties of the mating calls of L. bufonius males (i.e., dominant frequency and dominant frequency modulation).
Of course, female mate choice based on call characteristics should also be related to the auditory abilities of females. For example, the size of some hearing organs like the tympanum usually increases with body size, while also becoming more receptive toward lower frequencies (Fox, 1995; Wells, 2007). Thus, larger females with larger tympanums might be more receptive toward larger males with lower DF, while smaller females might be more attracted to smaller males. This in turn would promote size-assortative mating, a reproductive feature not yet elucidated in this species.
Finally, when compared to the published call descriptions of L. bufonius from various populations (Supplementary Table 4), we noticed several inconsistencies related to the call structure, which could suggest either the existence of a geographic variation in the mating calls of this taxon and/or the existence of a more complex acoustic repertoire. Thus, the calls in our study are very similar to those described by Barrio (1965) for L. bufonius from Salinas Grandes (Argentina) and fit with the advertisement call type I (“single note”) according to Schalk and Leavitt (2017). However, in the description of the calls of L. bufonius from Santa Cruz (Bolivia) by Schalk and Leavitt (2017), the oscillogram of this single-note call type appears to have a different temporal structure, with two amplitude peaks, not visible in our sample and not mentioned by Barrio (1965). Furthermore, Schalk and Leavitt (2017) also describe a second type of call (“type II”), composed of three notes, which appears to have a different function. Harmonics are not visible in the spectrograms of any of the call types and are not mentioned by the authors.
Earlier descriptions by Straughan and Heyer (1976), and later by Heyer (1978), for the calls of L. bufonius from Embarcación (Argentina) are somewhat contradictory. While the graphic representations of the calls show a single-note structure of the calls, Straughan and Heyer (1976) describe a two-note structure based on the presence of amplitude modulation in time. Their description is similar to what can be observed in the oscillogram provided by Schalk and Leavitt (2017). Heyer (1978) later provides a graphic representation of type-I calls from the same location, with an oscillogram that shows a rather constant amplitude in time. These authors also reported the absence of a harmonic structure in the call (which was close to a pure tone). In contrast, the calls analyzed in our study clearly had a harmonic structure with two, up to four visible harmonics, similarly to what Barrio (1965) described, and most of the energy was concentrated in the first part of the note, which appears T-shaped in the oscillograms (Figure 1). In a later study, Heyer and Scott (2006) show a graphic representation of type-I calls of L. bufonius from Filadelfia (Paraguay) which consist of single notes, without a visible harmonic structure in the spectrogram, and no T-shape pattern in the oscillogram.
Overall, our study brings the first thorough quantitative description of the mating calls in L. bufonius, showing that the DF and DF modulation are probably the most important acoustic features of these calls, since they allow for individual recognition and carry information regarding the status of the emitter male (i.e., size and age). The DF modulation appeared as a more robust signal for the acoustic communication associated with courtship in this species, not affected by air temperature or humidity. We suggest that the acoustic properties of the mating calls could contribute to shaping life history strategies, by allowing females to select males of certain size and age classes. A review of previous descriptions of the mating calls in L. bufonius revealed that its acoustic repertoire is complex and yet not fully understood; in addition, we found discrepancies which suggest that multiple species may have been recorded under the name L. bufonius.
Data availability statement
The original contributions presented in this study are included in the article/Supplementary material, further inquiries can be directed to the corresponding authors.
Ethics statement
This animal study was reviewed and approved by the Dirección de Recursos Naturales, Argentina.
Author contributions
FM collected the data in the field. FS conducted the data analyses. All authors contributed to writing the manuscript.
Funding
This work was supported by Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina, through Postdoctoral fellowships to FM, by the Romanian Ministry of Research, Innovation and Digitization, CNCS – UEFISCDI, through project number: PN-III-P1-1.1-PD-2021-0260, within PNCDI III (FS), and by the SYNTHESYS Project ES-TAF6704, http://www.synthesys.info/, which was financed by European Community Research Infrastructure Action under the FP7 “Capacities” Program at the Museo Museo Nacional de Ciencias Naturales (CSIC) (FS and RM).
Acknowledgments
FM was grateful to A. I. Kehr for providing essential tools for field work as tape recorder, thermohygrometer, and digital scale. FM also was grateful to V. I. Gomez for invaluable help during fieldwork. We are grateful to AM and MS for their valuable comments on the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.1020613/full#supplementary-material
References
Achorn, A. M., and Rosenthal, G. G. (2020). It’s not about him: Mismeasuring ‘good genes’ in sexual selection. Trends Ecol. Evol. 35, 206–219. doi: 10.1016/j.tree.2019.11.007
Audacity Team (2015). Audacity®: Free audio editor and recorder. Version 2.1.2. Available online at: http://audacity.sourceforge.net/ (accessed June 1, 2022).
Barrio, A. (1965). Nuptial song affinities of cave-dwelling species of the genus Leptodactylus (Anura-Leptodactylidae) [in Spanish]. Physis 25, 401–410.
Bioacoustics Research Program (2017). Raven Pro: Interactive sound analysis software, Version 1.5. Ithaca, NY: The Cornell Lab of Ornithology.
Blanckenhorn, W. U. (2000). The evolution of body size: What keeps organisms small? Q. Rev. Biol. 75, 385–407. doi: 10.1086/393620
Boulenger, G. A. (1894). List of reptiles and batrachians collected by Dr. J. Bohls near Asuncion, Paraguay. Ann. Mag. Nat. Hist. 13, 342–348.
Bosch, J., and de la Riva, I. (2004). Are frog calls modulated by the environment? An analysis with anuran species from Bolivia. Can. J. Zool. 82, 880–888. doi: 10.1139/z04-060
Brepson, L., Voituron, Y., and Lengagne, T. (2013). Condition-dependent ways to manage acoustic signals under energetic constraint in a tree frog. Behav. Ecol. 24, 488–496. doi: 10.1093/beheco/ars189
Brooks, R., and Kemp, D. J. (2001). Can older males deliver the good genes? Trends Ecol. Evol. 16, 308–313. doi: 10.1016/S0169-5347(01)02147-4
Burnham, K. P., and Anderson, D. R. (2002). Model selection and multimodel inference: A practical information-theoretic approach, 2nd Edn. New York, NY: Springer.
Castellano, S., and Cadeddu, G. (2011). Does quality affect growth rate and age at maturity in species with indeterminate growth? Evol. Ecol. Res. 13, 797–812.
Cei, J. M. A. (1949). Courtship habits and reproduction of a characteristic Chacoan batrachian (Leptodactylus bufonius Boul.). Acta Zool. Lilloana 8, 105–110.
de Oliveira-Santos, J. R., Bressan Queiroz de Figueiredo, R. C., and Barbosa de Moura, G. J. (2022). The defense repertoire of males of Leptodactylus vastus Lutz 1930 in a fragment of the Atlantic Forest in northeastern Brazil. Ethol. Ecol. Evol. 1–12. doi: 10.1080/03949370.2022.2059784 [Epub ahead of print].
Doty, G. V., and Welch, A. M. (2001). Advertisement call duration indicates good genes for offspring feeding rate in gray tree frogs (Hyla versicolor). Behav. Ecol. Sociobiol. 49, 150–156. doi: 10.1007/s002650000291
Duellman, W. E., and Trueb, L. (1994). Biology of amphibians. Baltimore, MD: Johns Hopkins University Press.
Faggioni, G., Souza, F., Uetanabaro, M., Landgref-Filho, P., Furman, J., and Prado, C. (2017). Reproductive biology of the nest building vizcacheras frog Leptodactylus bufonius (Amphibia, Anura, Leptodactylidae), including a description of unusual courtship behaviour. Herpetol. J. 27, 73–80.
Felton, A., Alford, R. A., Felton, A. M., and Schwarzkopf, L. (2006). Multiple mate choice criteria and the importance of age for male mating success in the microhylid frog, Cophixalus ornatus. Behav. Ecol. Sociobiol. 59, 786–795. doi: 10.1007/s00265-005-0124-6
Fox, J., and Weisberg, S. (2019). An R companion to applied regression, 3rd Edn. Thousand Oaks, CA: Sage Publications.
Fox, J. H. (1995). Morphological correlates of auditory sensitivity in anuran amphibians. Brain Behav. Evol. 45, 327–338. doi: 10.1159/000113560
Gerhardt, H. C. (1991). Female mate choice in treefrogs: Static and dynamic acoustic criteria. Anim. Behav. 42, 615–635. doi: 10.1016/S0003-3472(05)80245-3
Gerhardt, H. C. (1994). The evolution of vocalization in frogs and toads. Annu. Rev. Ecol. Syst. 25, 293–324. doi: 10.1146/annurev.es.25.110194.001453
Gillooly, J. F., and Ophir, A. G. (2010). The energetic basis of acoustic communication. Proc. R. Soc. B Biol. Sci. 277, 1325–1331. doi: 10.1098/rspb.2009.2134
Grafe, T. U., and Thein, J. (2001). Energetics of calling and metabolic substrate use during prolonged exercise in the European treefrog Hyla arborea. J. Comp. Physiol. B 171, 69–76. doi: 10.1007/s003600000151
Heino, M., and Kaitala, V. (1999). Evolution of resource allocation between growth and reproduction in animals with indeterminate growth. J. Evol. Biol. 12, 423–429. doi: 10.1046/j.1420-9101.1999.00044.x
Heyer, W. R. (1969). The Adaptive ecology of the species groups of the genus Leptodactylus (Amphibia, Leptodactylidae). Evolution 23, 421–428. doi: 10.2307/2406697
Heyer, W. R. (1978). Systematics of the fuscus group of the frog genus Leptodactylus (Amphibia, Leptodactylidae). Nat. Hist. Mus. Los Angeles Cty. Sci. Bull. 29, 1–85. doi: 10.5479/si.00810282.301
Heyer, W. R., Reichle, S., Silvano, D., and di Tada, I. (2004). Leptodactylus bufonius. The IUCN Red List of Threatened Species 2004: e.T57115A11582955. Available online at: https://dx.doi.org/10.2305/IUCN.UK.2004.RLTS.T57115A11582955.en (accessed July 15, 2022). doi: 10.2305/IUCN.UK.2004.RLTS.T57115A11582955.en
Heyer, W. R., and Scott, N. J. Jr. (2006). The advertisement call of Leptodactylus laticeps (Amphibia, Anura, Leptodactylidae): Predatory aural luring? Herpetol. Nat. Hist. 9, 189–194.
Jaquiéry, J., Broquet, T., Aguilar, C., Evanno, G., and Perrin, N. (2010). Good genes drive female choice for mating partners in the lek-breeding European treefrog. Evolution 64, 108–115. doi: 10.1111/j.1558-5646.2009.00816.x
Kelleher, S. R., Scheele, B. C., Silla, A. J., Keogh, J. S., Hunter, D. A., Endler, J. A., et al. (2021). Disease influences male advertisement and mating outcomes in a critically endangered amphibian. Anim. Behav. 173, 145–157. doi: 10.1016/j.anbehav.2021.01.007
Kelleher, S. R., Silla, A. J., McFadden, M. S., Stares, M. G., Endler, J. A., and Byrne, P. G. (2022). Multiple phenotypic traits predict male mating success in a critically endangered frog. Behav. Ecol. Sociobiol. 76, 1–21. doi: 10.1007/s00265-021-03119-9
Kokko, H. (1997). Evolutionarily stable strategies of age-dependent sexual advertisement. Behav. Ecol. Sociobiol. 41, 99–107. doi: 10.1007/s002650050369
Kokko, H., and Lindström, J. (1996). Evolution of female preference for old mates. Proc. R. Soc. Lond. Ser. B Biol. Sci. 263, 1533–1538. doi: 10.1098/rspb.1996.0224
Manning, J. T. (1985). Choosy females and correlates of male age. J. Theor. Biol. 116, 349–354. doi: 10.1016/S0022-5193(85)80273-3
Marangoni, F., Courtis, A., Pineiro, J. M., Ingaramo, M. D. R., Cajade, R., and Stănescu, F. (2019). Contrasting life-histories in two syntopic amphibians of the Leptodactylus fuscus Heyer, 1978group. An. Acad. Bras. Ciênc. 91:e20180507. doi: 10.1590/0001-3675201920180507
Marangoni, F., Schaefer, E., Cajade, R., and Tejedo, M. (2009). Growth-mark formation and chronology of two Neotropical anuran species. J. Herpetol. 43, 546–550. doi: 10.1670/08-230R1.1
Márquez, R. (1995). Female choice in the midwife toads (Alytes obstetricans and A. cisternasii). Behaviour 132, 151–161. doi: 10.1163/156853995X00342
Márquez, R., De la Riva, I., and Bosch, J. (1996). Advertisement calls of three glass frogs from the Andean forests (Amphibia: Anura: Centrolenidae). Herpetol. J. 6, 97–99.
Mazerolle, M. J. (2020). AICcmodavg: Model selection and multimodel inference based on (Q)AIC(c). R package version 2.3-1. Available online at: https://cran.r-project.org/package=AICcmodavg (accessed June 1, 2022).
Moser, C. F., Schuck, L. K., Olmedo, G. M., and Lingnau, R. (2022). Individual variation in the advertisement call of Aplastodiscus albosignatus (Anura: Hylidae) is correlated with body size and environmental temperature. Zoologia 39:e21008. doi: 10.1590/s1984-4689.v39.e21008
Narins, P., Feng, A. S., Fay, R. R., and Popper, A. N. (2006). Hearing and sound communication in amphibians. Berlin: Springer. doi: 10.1007/978-0-387-47796-1
Ophir, A. G., Schrader, S. B., and Gillooly, J. F. (2010). Energetic cost of calling: General constraints and species-specific differences. J. Evol. Biol. 23, 1564–1569. doi: 10.1111/j.1420-9101.2010.02005.x
Peterson, B. G., and Carl, P. (2020). PerformanceAnalytics: Econometric tools for performance and risk analysis. R package version 2.0.4. Available online at: https://CRAN.R-project.org/package=PerformanceAnalytics (accessed June 1, 2022).
Philibosian, R., Ruibal, R., Shoemaker, V. H., and McClanahan, L. L. (1974). Nesting behavior and early larval life of the frog Leptodactylus bufonius. Herpetologica 30, 381–386.
Pinheiro, J., Bates, D., DebRoy, S., and Sarkar, D., and R and Core Team (2019). nlme: Linear and nonlinear mixed effects models. R package version 3.1-143. Available online at https://CRAN.R-project.org/package=nlme (accessed June 1, 2022).
R Core Team (2019). R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
Reading, C. J., and Jofré, G. M. (2003). Reproduction in the nest building vizcacheras frog Leptodactylus bufonius in central Argentina. Amphib. Reptil. 24, 415–427. doi: 10.1163/156853803322763882
Ryan, M. J. (1988). “Constraints and patterns in the evolution of anuran acoustic communication,” in The evolution of the amphibian auditory system, eds B. Fritzsch, M. J. Ryan, W. Wilczynski, T. E. Hetherington, and W. Walkowia (New York, NY: John Wiley & Sons), 637–677.
Ryan, M. J. (1991). Sexual selection and communication in frogs. Trends Ecol. Evol. 6, 351–355. doi: 10.1016/0169-5347(91)90224-L
Schalk, C. M., and Leavitt, D. J. (2017). Leptodactylus bufonius. Cat. Am. Amphibians Reptil. 2017, 905.1–905.22. doi: 10.1007/s12045-017-0549-z
Schneider, C., Rasband, W., and Eliceiri, K. (2012). NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675. doi: 10.1038/nmeth.2089
Stearns, S. C. (2000). Life history evolution: Successes, limitations, and prospects. Naturwissenschaften 87, 476–486. doi: 10.1007/s001140050763
Straughan, I. R., and Heyer, W. R. (1976). A functional analysis of the mating calls of the Neotropical frog genera of the Leptodactylus complex (Amphibia, Leptodactylidae). Papéis Avulsos Zool. 29, 221–245.
Taigen, T. L., O’Brien, J. A., and Wells, K. D. (1996). The effect of temperature on calling energetics of the spring peeper (Pseudacris crucifer). Amphib. Reptil. 17, 149–158. doi: 10.1163/156853896X00180
Welch, A. M., Semlitsch, R. D., and Gerhardt, H. C. (1998). Call duration as an indicator of genetic quality in male gray tree frogs. Science 280, 1928–1930. doi: 10.1126/science.280.5371.1928
Wells, K. D. (2007). The ecology and behaviour of amphibians. London: The University of Chicago Press. doi: 10.7208/chicago/9780226893334.001.0001
Keywords: neotropical, acoustic parameters, skeletochronology, sexual selection, life-history strategies
Citation: Stănescu F, Márquez R, Cogălniceanu D and Marangoni F (2022) Older males whistle better: Age and body size are encoded in the mating calls of a nest-building amphibian (Anura: Leptodactylidae). Front. Ecol. Evol. 10:1020613. doi: 10.3389/fevo.2022.1020613
Received: 16 August 2022; Accepted: 13 September 2022;
Published: 29 September 2022.
Edited by:
Ann Valerie Hedrick, University of California, Davis, United StatesReviewed by:
Adolfo Martino, Universidad Nacional de Río Cuarto, ArgentinaMirco Solé, Universidade Estadual de Santa Cruz, Brazil
Copyright © 2022 Stănescu, Márquez, Cogălniceanu and Marangoni. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Florina Stănescu, ZmxvcmluYV9zdGFuZXNjdUB5bWFpbC5jb20=; Rafael Márquez, cm1hcnF1ZXpAbW5jbi5jc2ljLmVz