 Ivette Marai Villa-Villaseñor1
Ivette Marai Villa-Villaseñor1 Beatriz Yáñez-Rivera2
Beatriz Yáñez-Rivera2 Rebeca Aneli Rueda-Jasso3
Rebeca Aneli Rueda-Jasso3 Ma. Antonia Herrera-Vargas4Rubén Hernández-Morales3
Ma. Antonia Herrera-Vargas4Rubén Hernández-Morales3 Esperanza Meléndez-Herrera4*Omar Domínguez-Domínguez3
Esperanza Meléndez-Herrera4*Omar Domínguez-Domínguez3- 1Programa Institucional de Doctorado en Ciencias Biológicas, Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Mexico
- 2CONACYT-CIAD, AC Mazatlán Unit for Aquaculture and Environmental Management, Mazatlan, Mexico
- 3Laboratorio de Biología Acuática, Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Mexico
- 4Laboratorio de Ecofisiología Animal, Instituto de Investigaciones Sobre Recursos Naturales, Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Mexico
Nitrate-nitrogen (NO3-N) pollution related to anthropogenic activities is increasing in freshwater ecosystems. Knowledge about NO3-N sensitivity in freshwater wild fish is needed to understand the differential tolerance between species. Goodeinae is a subfamily of 41 endemic fishes that inhabit central Mexico, with 33 species in the IUCN red list and three extinct. Distributional patterns suggest tolerant and sensitive goodeines related to the conservation gradient of freshwater ecosystems. Four species with a differential distribution and tolerance were selected to evaluate their physiological responses to NO3-N. Fish were exposed to different NO3-N concentrations for 96 h and the median lethal concentration (LC50) was determined. Swimming disorders plus gill and liver histopathological indexes were estimated and incorporated into an Integrated Biomarker Response (IBR) for each species. Skiffia lermae (LC50 = 474.332 mg/L) and Xenotoca variata (LC50 = 520.273 mg/L) were more sensitive than Goodea atripinnis (LC50 = 953.049 mg/L) and Alloophorus robustus (LC50 = 1537.13 mg/L). The typical histological damage produced by NaNO3-N exposure was fusion of secondary lamellae in gills. This was present in all species and cellular degeneration was observed at the highest concentrations. Secondary lamellae aneurysms were only observed in G. atripinnis. Liver alterations included vascular dilation in hepatic sinusoids, hyperemia and nuclear hypertrophy; higher concentrations produced hepatocyte cytoplasmic vacuolation and reduced frequency of cell nuclei. Behavioral and histopathological alterations could explain the differential species sensitivity. The results suggest that species which preserve gill function and transfer the task of detoxification to the liver might have the best chance of surviving in polluted environments. Moreover, species previously considered as tolerant may be highly susceptible to NaNO3-N exposure. Therefore, it is necessary to closely monitor NaNO3-N concentrations in freshwater ecosystems and, if possible, reduce their levels to avoid the loss of wild populations.
Introduction
One of the most representative fish groups in central Mexico are the goodeines. The major diversification within this group occurs in the Río Lerma-Santiago basin, one of the most polluted basins in the world (Domínguez-Domínguez et al., 2006, 2008; Lyons et al., 2019). The Goodeinae subfamily is composed of 41 species from 19 genera (Domínguez-Domínguez et al., 2008; Lyons et al., 2019). This group presents unique reproductive characteristics such as internal fertilization, matrotrophy, and viviparity (Iida et al., 2019), and is considered one of the most at-risk groups in the world (Duncan and Lockwood, 2001). During the last decades, their populations have decreased related to pollution and other anthropogenic activities (Lyons et al., 2019). According to the IUCN (2022), 13 species are critically endangered, 14 endangered, six vulnerable, two extinct in the wild, and one extinct. Most Goodeinae species are endemic or micro-endemic to a single spring, portion of a river or drainage, whereas others are widely distributed in the region (Foster and Piller, 2018). These distributional patterns suggest tolerant and sensitive goodeines related to the conservation gradient of freshwater ecosystems (Rueda-Jasso et al., 2017; Lyons et al., 2019). For example: Skiffia lermae and Alloophorus robustus have been classified as endangered, sensitive, and vulnerable species, respectively; since they only are found in spring areas, lake shorelines and small tributaries (Miller et al., 2009; Lyons et al., 2019). Xenotoca variata and Goodea atripinnis are considered as least concern and tolerant species, because some areas they inhabit are classified as highly polluted aquatic environments (Domínguez-Domínguez et al., 2008; Rueda-Jasso et al., 2017; Lyons et al., 2019).
Nitrates (NO3) constitute the most stable and abundant form of dissolved inorganic nitrogen in aquatic ecosystems (Galloway et al., 2004). Although nitrate-nitrogen (NO3-N) may be naturally present, they accumulate in the ecosystem due to human activities such as livestock and agricultural use of inorganic nitrogenous fertilizers. Municipal and industrial wastewater effluents also increase nitrogenous compounds (Du et al., 2019; Goeller et al., 2019). Despite the abundance of NO3-N, their impact on fish welfare has been underestimated since they are less toxic than other nitrogenous compounds (van Bussel et al., 2012; Kim et al., 2019; Presa et al., 2022). However, NO3-N negatively impact fish health (Camargo et al., 2005; Guillette and Edwards, 2005; McGurk et al., 2006; Gomez-Isaza et al., 2018, 2020, 2021; Presa et al., 2022).
The standard sensitivity evaluation to acute toxicity is the median lethal concentration (LC50). This index is usually complemented with other responses to increase the understanding of physiological trade-offs under acute exposure or predict the long-term permanence of fish in polluted environments (Kim et al., 2019; Dutra et al., 2020). Exposure bioassays to NO3-N in fish revealed more sensitivity during early life stages (Rueda-Jasso et al., 2017), likely due to interference with biological processes such as body growth and reproductive development or behavior (Yu et al., 2021). The gills are the main route of NO3-N absorption (Pereira et al., 2017). Physiological responses to pollutants aim to maintain gill activity preceding the loss of function related to the degree of damage (Emam et al., 2022). Inside the body, NO3-N causes physiological and behavioral alterations (Torno et al., 2018; Yang et al., 2019; Yu et al., 2021). The liver is the detoxifying organ and its condition indicates environmental quality (Corredor-Santamaría et al., 2021; Tramunt et al., 2021; Santos et al., 2022). Toxic pollutants cause functional alterations of the gills and liver, such as gas exchange failure and metabolic damage. These changes are associated with altered swimming behavior, such as loss of balance, top or bottom position in the water column and lethargy (Rodrigues et al., 2011; Schram et al., 2012; Pereira et al., 2017; Yang et al., 2019).
Given the conservation relevance of goodeines and based on the premise that differential persistence in freshwater bodies could be related to differential sensibility to environmental pollutants, the main objective of this study was to evaluate NO3-N sensitivity in offspring of S. lermae, X. variata, G. atripinnis, and A. robustus. Differential sensitivity was evaluated by LC50 values, histopathological indexes from the gills and liver, as well as swimming behavior alterations under acute exposure (96 h). Integrated Biomarker Response (IBR) analysis was performed to contrast their sensitivity.
Materials and methods
Ethical statements
Sampling and laboratory fish handling protocols were reviewed and approved (SEMARNATSGPA/DGVS/00012/19, PPF/DGOPA-014/20). The protocols also followed the Guide for the Care and Use of Laboratory Animals (1996). Toxicity was evaluated with the Fish Acute Toxicity Test (OECD, 2019).
Field collection and fish maintenance
Gravid females of S. lermae (n = 25), X. variata (n = 20), G. atripinnis (n = 7), and A. robustus (n = 5) were collected from Zacapu Lake, central Mexico (19°49′26.88″N; 101°46′37.32″W), during years 2020 and 2021, using 2 mm aluminum mesh minnow traps (Gee-minnow-traps® G-40, USA). Gravid females were transported to the laboratory with water from the collection site. Females were acclimatized for 15 days in 120 L aquariums (loading capacity: 6 L per fish), with dechlorinated water and 7 mg/L of dissolved oxygen, at 22°C, under a natural light photoperiod. They were fed two times each day with Artemia sp. and commercial Wardley fish flakes (Rueda-Jasso et al., 2017). On post-natal days 1–7, offspring were separated from their mothers, fed and housed under the same conditions previously described. Offspring remain in these conditions until post-natal 21, when they were subjected to the experiment (see below).
Water quality monitoring and solutions
Temperature, dissolved oxygen, pH, salinity, total dissolved solids and electric conductivity were recorded every 24 h using a multiparameter probe (YSI EXO2. Ohio, USA). The toxicity assay solution was prepared with sodium nitrate (NaNO3, 97%, Sigma-Aldrich) according to a stoichiometric calculation (Dutra et al., 2020). Concentrations of ammonium-nitrogen (NH4-N), nitrite-nitrogen (NO2-N), and NO3-N were measured following the methodologies described in the American Public Health Association (American Public Health Association [APHA], 2017).
Toxicity test
Each fry was acclimatized for 7 days, weighted with an analytical precision balance (OHAUS™ Adventurer d = 0.0001 g, China), their length was measured using a digital Vernier caliper (Thermo Fisher Scientific™ S/N 1366162, USA) and then randomly assigned to a treatment. Three replicates were used per treatment, with 10 fish in 10 L tanks (loading capacity: 1 L per fish). A control group was included for each species.
During NO3-N exposure, survival was monitored at 0.5, 1, 2, 3, 4, 6, 9, 12, 24, 36, 48, 72, and 96 h. Mortality was confirmed by the absence of response to stimulation. Swimming behavior (loss of balance and position in the bottom, middle, or surface of the water column) was recorded during 10 min by counting the animals that exhibited these actions at the same intervals mentioned above. Changes in swimming behavior were categorized as “low effect,” indicating an alteration in 10–33% of individuals; “moderate effect” 34–63% and “high effect,” 64–100%. Changes to swimming patterns were recorded only if they occurred in 10% of the fish within each aquarium (Ogueji et al., 2018). At the end of the experiment, surviving fish were anesthetized using clove oil (50 μl L-1) and processed for histology. Fish in the highest NO3-N concentrations were removed from the aquariums immediately after they became immobile.
Histological evaluation
Three individuals from three treatments (control, that closest to the LC50 and the most lethal concentration, LC100) per species were randomly selected and processed by routine histology. Gill and liver 5 μm transversal sections were obtained and stained with hematoxylin-eosin (Bancroft and Cook, 1994; Cano-Rocabayera et al., 2019). Slides were observed with a Leica microscope (DM3000, Germany), photo-documented using a camera (Leica DFC310 FX, Germany) and analyzed using NIH ImageJ software.
Gill and liver histopathological indexes
The histopathological condition index (HI) was calculated for each organism (Bernet et al., 1999; Antunes et al., 2017). Briefly, according to distinctive histological features, gill and liver alterations were classified into circulatory, progressive and regressive patterns. A pathological importance factor (w) was assigned for each type of damage (Supplementary Table 1). Similarly, a score value (a) was assigned for the occurrence of alterations from 0 to 5 (where 0 is no change; 1, changes in 1–20% of the sections; 2, 21–40%; 3, 41–60%; 4, 61–80%; 5, 81–100%, respectively). The HI for each rp was determined using the formula: HIrp org = Σalt (w × a) where HIrp org is the reaction index of the organ for each reaction pattern (rp) and alt is the alteration (Antunes et al., 2017).
Liver damage tissue index
For the liver, tissue damage was assessed by nuclear density (nD = number of nuclei/mm2) and nuclear area (nA = area of hepatocyte nuclei in μm2; Rodrigues et al., 2017). The average value of controls (nDc and nAc) was used as a reference to calculate a relative value for each feature. Since an increase in the nuclear area could indicate hypertrophy and a reduction could be related to pyknotic nuclei, the absolute value of the difference to 1 multiplied by 0.5 was used. Finally, the values were integrated into the following index: LDTI = (nD*1/nDc) + [| 1 – (nA*1/nAc)| × 0.5], where values near 1 indicate a normal or reference condition and values closest to 0 indicate major tissue damage.
Statistical analysis and integrated biomarkers response
All analyses were performed with R software (R Core Team, 2022, version 4.1.3). Variables were tested for normality and homoscedasticity with Shapiro-Wilk and Levene’s tests, respectively. The survival rate for each species was calculated by the Kaplan Meier method and compared with the log-rank test using the “survival” and “survminer” packages. Acute toxicity to NaNO3-N was determined by the LC50 for each species (95% confidence intervals after 96 h) using the Spearman-Karber method and considered significantly different when not overlapping using the “tsk” package. Relationships between swimming disorders and concentrations at each exposure time were analyzed with Spearman correlations using the “stats” package; graphs were produced using the “ggpubr” package. Histomorphometric analyses were performed using the “car” and “pgirmess” packages. LDTI data distribution was analyzed by a Kruskal-Wallis test. Tukey multiple range post-hoc tests were used to discriminate means (Torno et al., 2018). The values obtained for each parameter were expressed as mean ± standard error of the mean. Survival rates, LC50, swimming behavior, GHI, LHI, and LDTI were incorporated into an IBR to visualize and compare species-specific responses to acute NaNO3-N exposure using the “IBRtools” package (Bertrand et al., 2016).
Results
NaNO3-N exposure and water quality
Concentrations for acute toxicity assays were determined for each species. Thus, each species was exposed to slightly different ranges; S. lermae: 100, 200, 400, 800, and 1,200 mg NO3-N/L; X. variata: 200, 400, 800, 1,200, and 1,600 mg NO3-N/L; G. atripinnis: 400, 800, 1,200, 1,600, 2,000, and 2,400 mg NO3-N/L; and A. robustus: 400, 800, 1,200, 1,600, 2,000, 2,400, and 2,800 mg NO3-N/L. The body mass, length and total number of offspring for each species was: 0.028 ± 0.0004 g; 12.265 ± 0.058 mm; n = 180 for S. lermae, 0.072 ± 0.001 g; 15.656 ± 0.103 m; n = 180 for X. variata, 0.184 ± 0.001 g; 20.122 ± 0.047 mm; n = 210 for G. atripinnis and 0.182 ± 0.001 g; 21.382 ± 0.055 mm; n = 240 for A. robustus.
Water quality conditions (temperature: 22.059 ± 0.003°C; dissolved oxygen: 7.069 ± 0.001 mg/L; pH: 8.930 ± 0.004 log units; ammonium: 0.003 ± 4.88104E-06 mg/L and nitrite: 0.042 ± 0.002 mg/L) in the experimental tanks were similar for all groups and maintained in a range similar to the natural variations observed in Zacapu lake (Ramírez-García et al., 2022). Salinity (0.281 ± 0.001–12.041 ± 0.096 g/L), total dissolved solids (379.033 ± 1.913–13,017.266 ± 84.070 mg/L) and electric conductivity (551.290 ± 2.691–19,103.63 ± 141.191 μS/cm) increased as NaNO3-N concentrations increased.
Survival rate and lethal concentration 50%
Survival rates for the four Goodeinae species decreased as NaNO3-N concentrations increased and were related to exposure time. No mortality was observed at control concentrations (Figure 1). Survival rates for S. lermae and X. variata showed significant differences above 200 mg NaNO3-N/L vs. control [S. lermae: X2(5) = 306; p ≤ 0.001; X. variata: X2(5) = 149; p ≤ 0.001]; no fish survived after 96 h at 1,200 mg NaNO3-N/L for S. lermae and at 1,600 mg NaNO3-N/L for X. variata (Figures 1A,B). Survival rates for G. atripinnis showed significant differences above 800 mg NaNO3-N/L vs. control [X2(6) = 131; p ≤ 0.001] and no fish survived at 2,400 mg NaNO3-N/L (Figure 1C). For A. robustus significant differences were observed at 2,000 mg NaNO3-N/L vs. control [X2(7) = 940; p ≤ 0.001] and all fish died at 2,800 mg NaNO3-N/L (Figure 1D).
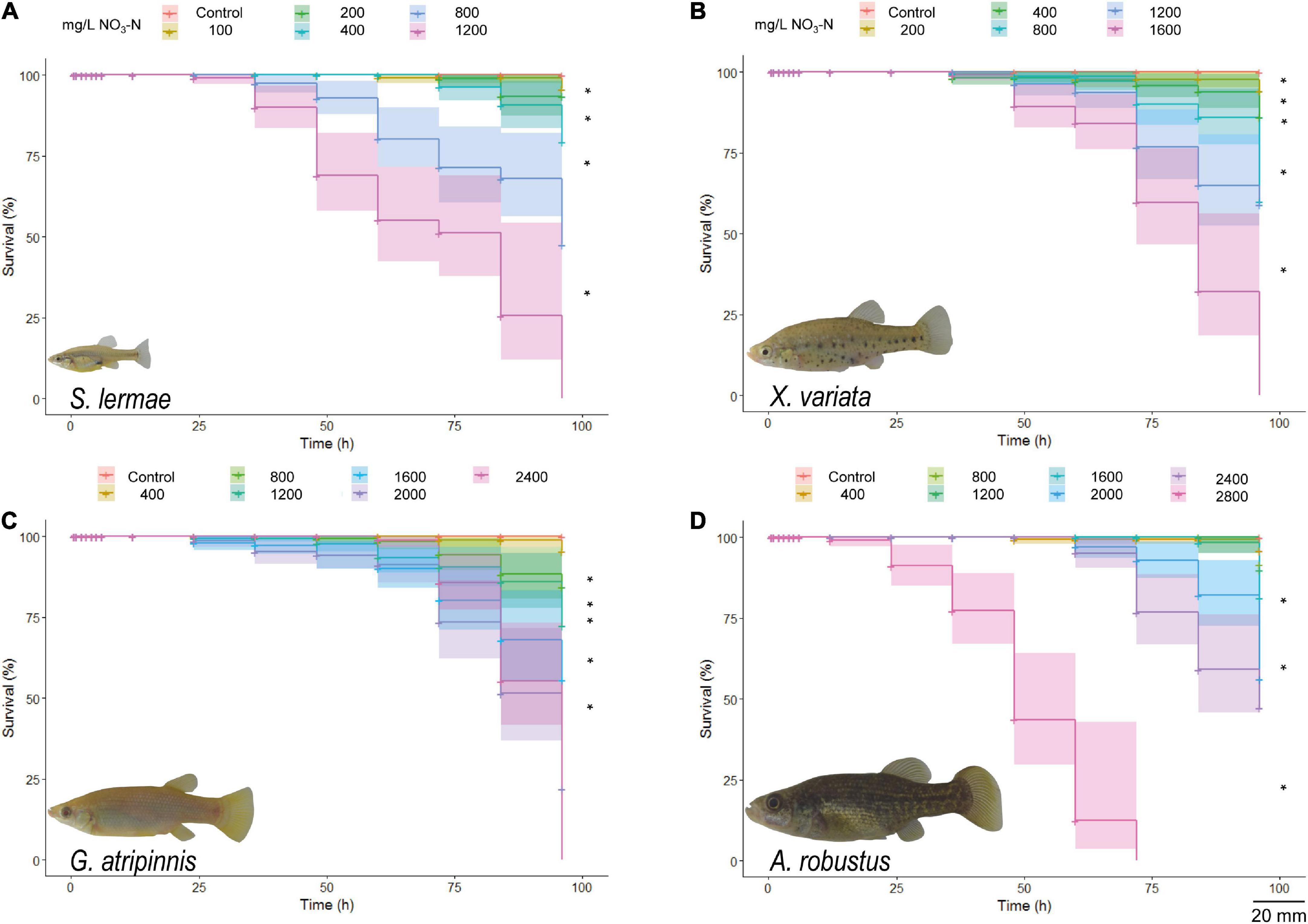
Figure 1. Kaplan-Meier survival curves for S. lermae (A), X. variata (B), G. atripinnis (C), and A. robustus (D) exposed to NaNO3-N. Log rank test indicated significant differences compared to the control groups (n = 30, *p < 0.001). Shading: 95% confidence intervals. Representative photograps of each species are shown (scale bar = 20mm).
The calculated LC50 values and corresponding 95% confidence intervals for each species followed the survival trend (Figure 2). Skiffia lermae was the most sensitive, LC50 = 474.332 (373.850–601.820) mg NaNO3-N/L, followed by X. variata LC50 = 520.273 (380.390–711.594) mg NaNO3-N/L, G. atripinnis LC50 = 953.049 (722.835–1256.584) mg NaNO3-N/L, and A. robustus LC50 = 1537.130 (1203.223–1963.699) mg NaNO3-N/L.
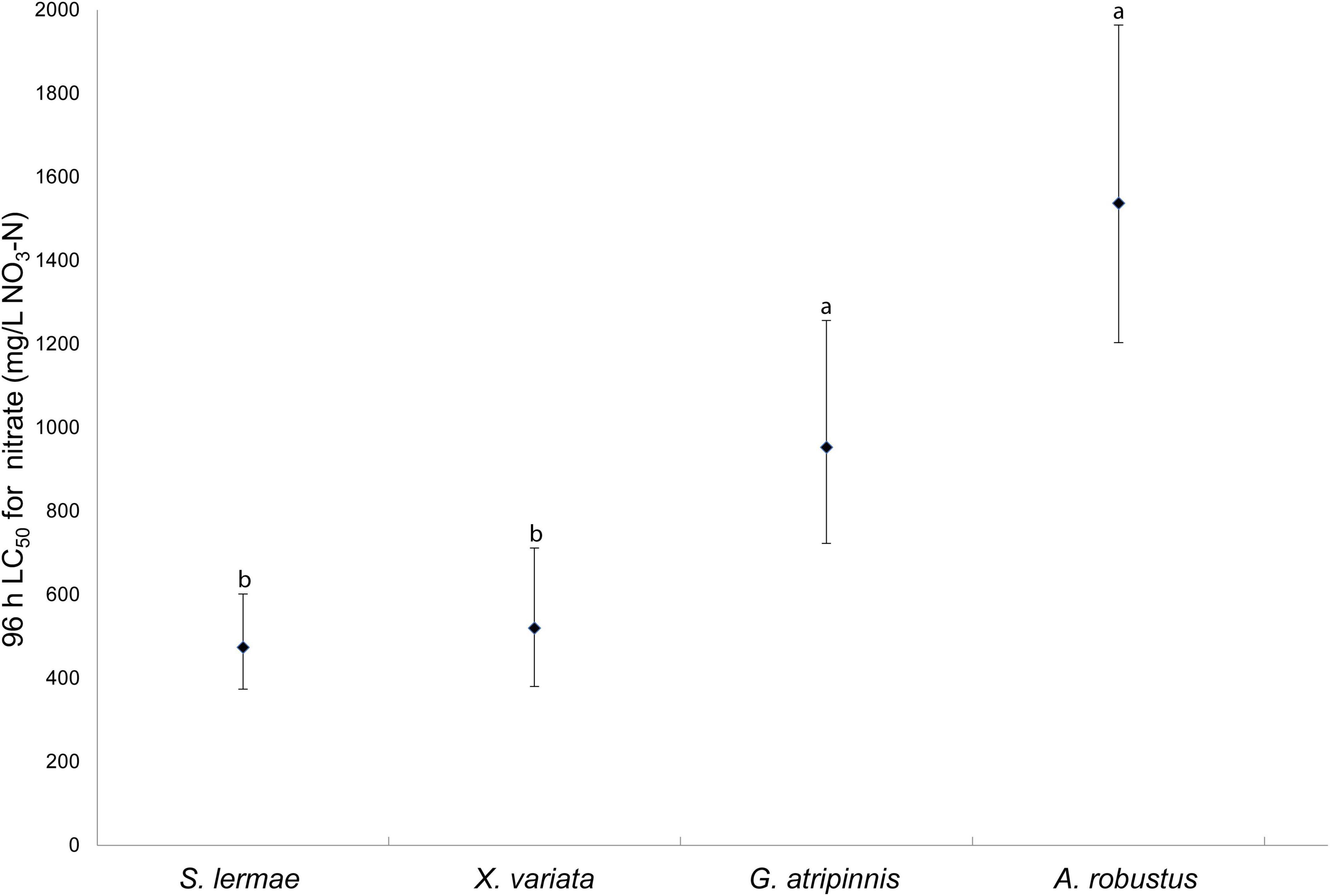
Figure 2. Lethal concentration 50% (LC50) and 95% confidence intervals for four species of freshwater fish exposed to NaNO3-N. LC50 were considered significantly different based on non-overlapping 95% confidence intervals. Different letters indicate significant differences (n = 30).
Swimming behavior
Swimming disorders were related to exposure time and concentration (Tables 1, 2 and Supplementary Figure 1). The first alteration in S. lermae offspring was that they remained on the bottom of the water column at 24 h of exposure to the lowest concentration (100 mg NaNO3-N/L). The other three species showed surface permanence as the first alteration at 48 h of exposure to 200 mg NaNO3-N/L in X. variata; while for A. robustus and G. atripinnis this occurred at 36 and 84 h of exposure to 400 mg/L, respectively (Table 1). The prevailing behavior during acute exposure was permanence at the bottom without exploring the aquarium, but some fish also showed surface preference.
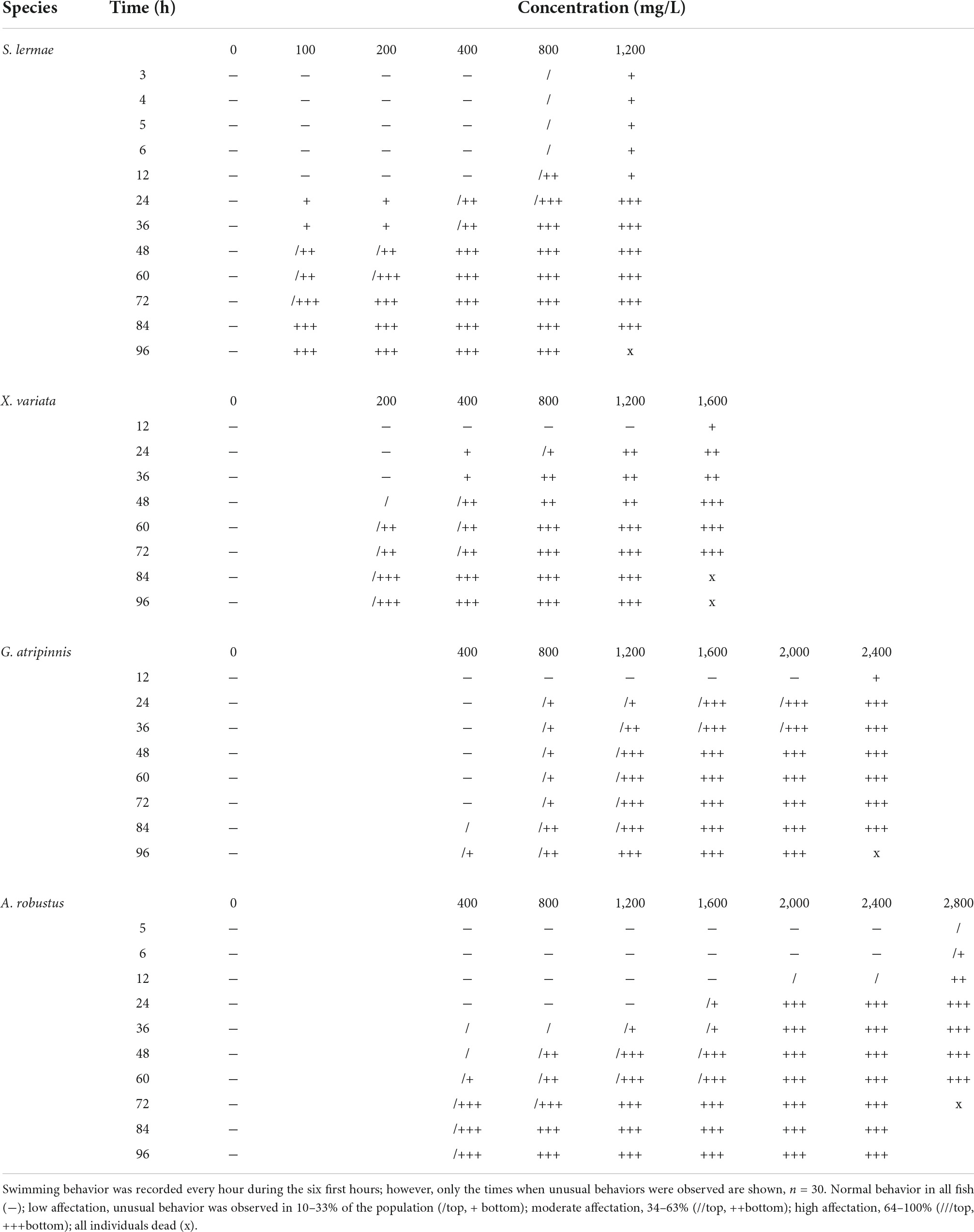
Table 1. Aquarium distribution.
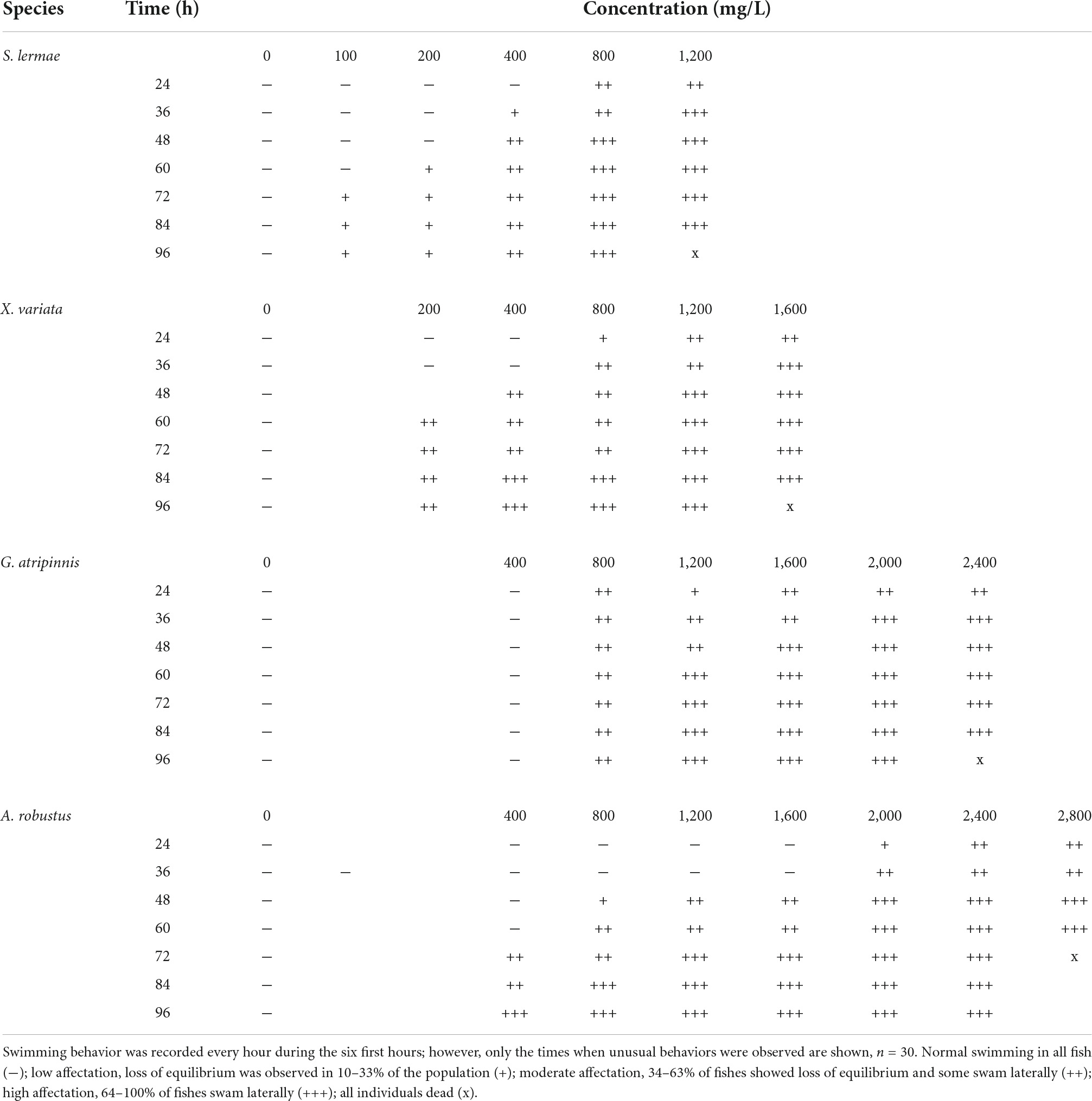
Table 2. Loss of equilibrium.
Loss of balance was related to exposure time and showed a similar trend, with S. lermae being more sensitive, followed by other species (Table 2 and Supplementary Figure 1). Goodea atripinnis did not show a loss of equilibrium at the lowest concentration.
Histological evaluation
Gill histopathological lesions integrated in the GHI showed a clear concentration-response pattern in all species (Table 3 and Figure 3). Circulatory index lesions were in a similar range; however, G. atripinnis was the only species that showed secondary lamellae aneurysms at the value closest to the LC50 (Figure 3). Lamellae aneurysms constitute the maximal circulatory damage associated with toxic exposure (w = 3 Supplementary Table 1). Secondary lamellae fusion was typical in S. lermae (Figure 3), where the progressive index reached the highest values (w = 3 Supplementary Table 1). Cellular degeneration was observed in the gills of all fish exposed to the lethal concentration. Thus, the regressive index showed the highest values at these concentrations for each species due to branchial degeneration resulting in death (w = 3 Supplementary Table 1).
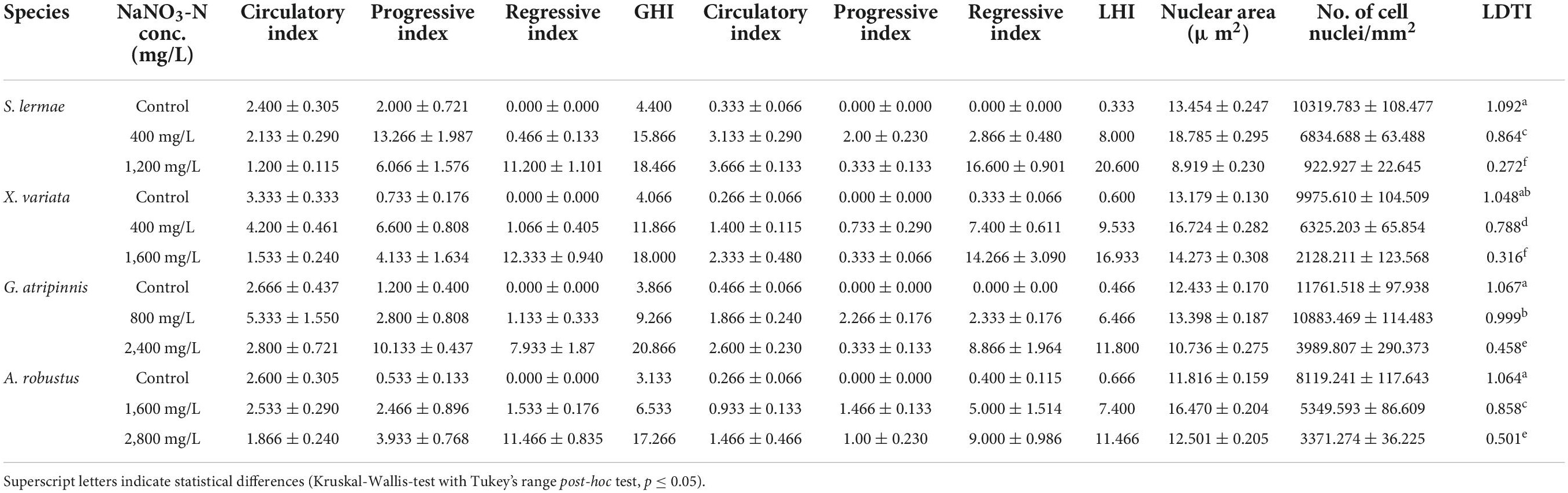
Table 3. Gill (GHI) liver histopathological (LHI) and tissue damage indexes (LTDI) of fish exposed to NaNO3-N (control, concentration closest to the LC50 and LC100), n = 3.
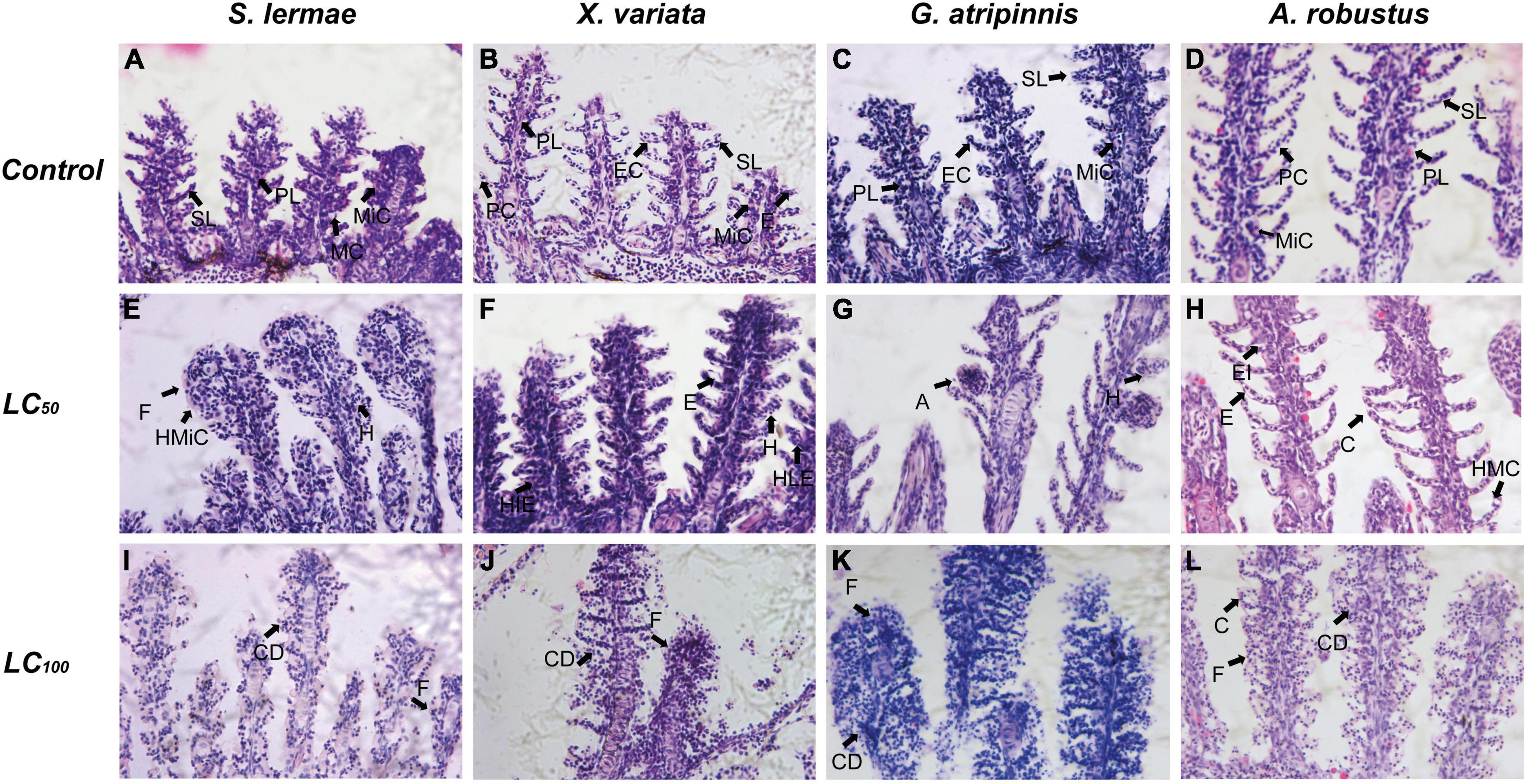
Figure 3. Effect of acute exposure to NaNO3-N on gill histology of four goodeine species. Representative photomicrographs of gill sections stained with hematoxylin and eosin from control (A–D), concentration closest to the LC50 (E–H) and LC100 (I–L) groups. A, Aneurysm of secondary lamellae; C, Change in curvature of secondary lamellae; CD, Cellular degeneration; E, Edema of secondary lamellae; EC, Epithelial cell; EI, Edema of interlamellar epithelium; F, Fusion of secondary lamellae; H, Hyperemia of secondary lamellae; HIE, Hyperplasia of interlamellar epithelium; HLE, Hyperplasia of lamellar epithelium; HMC, Mucus cell hyperplasia; HMiC, Mitochondria-rich cell hyperplasia; PL, Primary lamellae; SL, Secondary lamellae; MC, Mucus cell; MiC, Mitochondria-rich cell; PC, Pillar cell.
In the liver, a concentration-response relationship for LHI also occurred (Table 3 and Figure 4). Vascular dilation in hepatic sinusoids occurred mainly in S. lermae (w = 1 Supplementary Table 1), while other species showed hyperemia as circulatory liver damage (Figure 4 and w = 3 Supplementary Table 1). Nuclear hypertrophy, as quantified in the progressive index (w = 1 Supplementary Table 1), was frequent at those concentrations close to the LC50 in all species. Hepatocyte cytoplasmic vacuolation and a relative reduction in nuclear frequency was observed in all species, especially in fish exposed to the lethal concentration, where the regressive index showed higher values than in other treatments (w = 3 Supplementary Table 1).
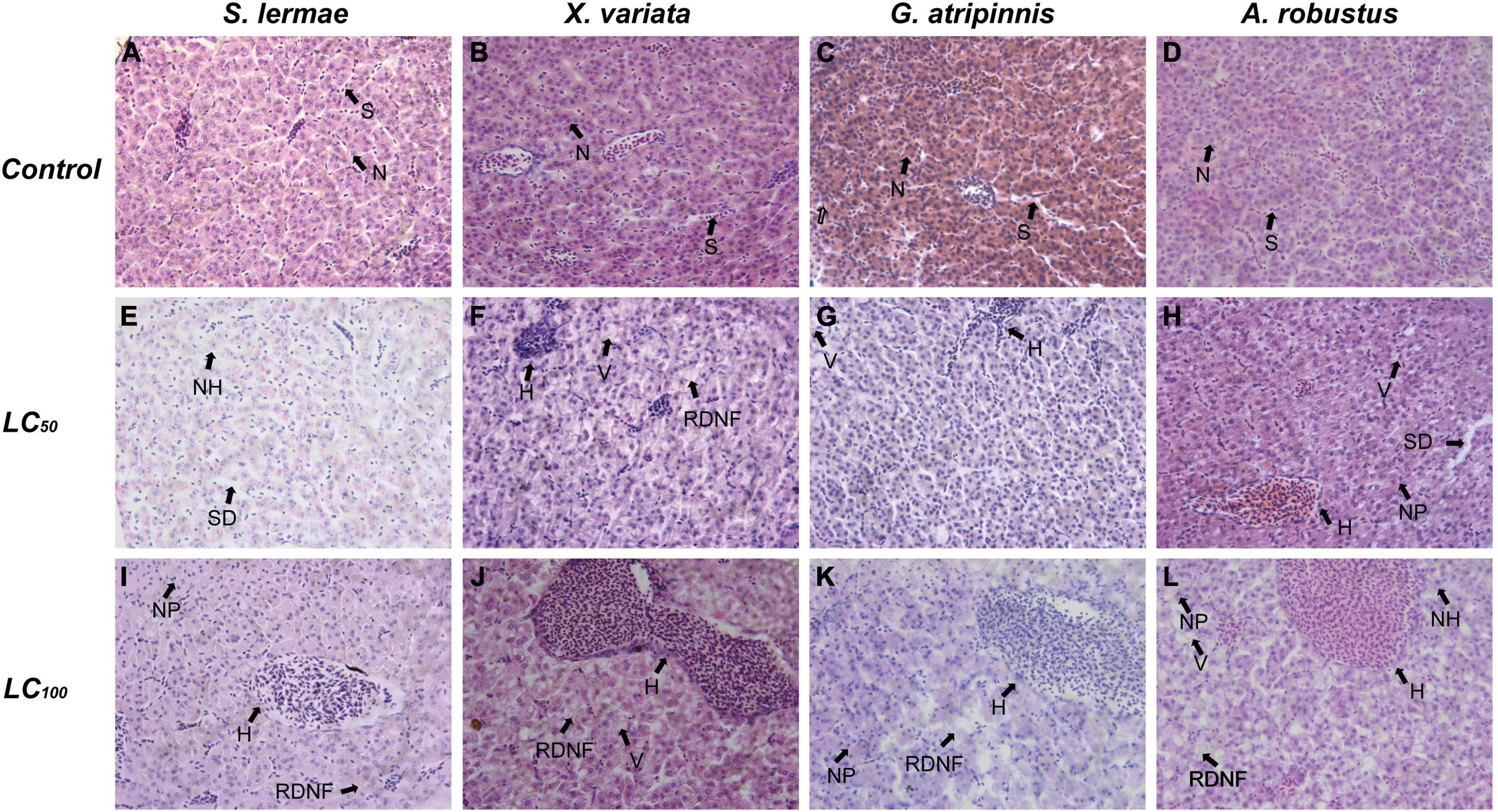
Figure 4. Effect of acute exposure to NaNO3-N on liver histology of four goodeine species. Representative photomicrographs of liver sections stained with hematoxylin and eosin from control (A–D), concentration closest to the LC50 (E–H) and LC100 (I–L) groups. H, Hyperemia; N, hepatocyte nucleus; NH, Nuclear hypertrophy; NP, nuclear pyknosis; RDNF, Relative decrease of nuclei frequency; S, sinusoid; SD, Blood sinusoid dilation; V, Vacuolation of the cytoplasm.
Quantitative hepatocyte density and nuclear area analysis were performed for the LTDI (values close to 0 indicate relevant tissue damage), which showed a concentration-response pattern (Table 3). The lowest hepatocyte density was observed at the lethal concentration, while hepatocyte nuclear area was higher at concentration closest to the LC50 in all species. However, the nuclear area in G. atripinnis showed similar values to their respective control group (Table 3).
Integrated biomarker response
For comprehensive evaluation, all traits analyzed were incorporated into an IBR for each species (Figure 5 and Supplementary Table 2). IBR values for each species were well weighted; control fish showed lower values and the lethal concentration had the highest score in all cases. Intercomparisons at doses closest to the LC50 indicated that the most sensitive species was S. lermae (IBR = 2.208), with the highest value, followed by X. variata (IBR = 2.124) at 400 mg NaNO3-N/L. A. robustus showed a higher IBR value than G. atripinnis; these were the most tolerant species to NaNO3. However, they differed by twice the concentration, in G. atripinnis it was 800 mg NaNO3-N/L (IBR = 0.977) and in A. robustus 1,600 mg NaNO3-N/L (IBR = 1.560). An important factor for the IBR in tolerant species was the liver (LHI), contrary to the most sensitive species where the important factor was gill affectation (GHI); swimming behavior was a promising biomarker in all species.
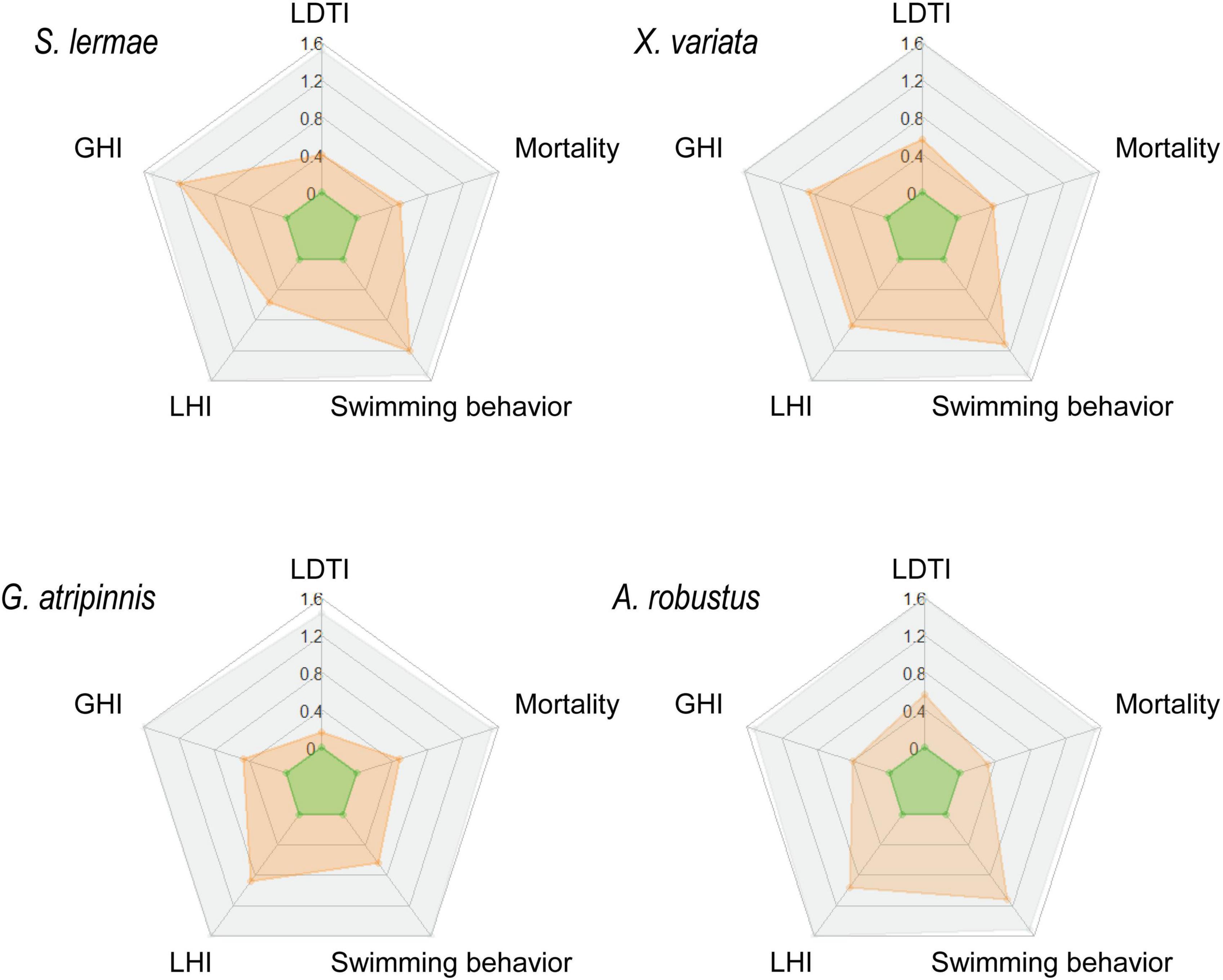
Figure 5. Integrated biomarker response (IBR) for four goodeine species exposed to NaNO3-N. Radar graphs show the calculated IBR for S. lermae, X. variata, G. atripinnis, and A. robustus from the control condition (green), exposed to the concentration closest to the LC50 (orange) and LC100 (gray). The spokes of the radar indicate all traits evaluated: mortality and swimming behavior, as well as gill (GHI), liver histopathological (LHI), and tissue damage indexes (LTDI).
Discussion
There was a clear concentration-response relationship, also dependent on time, in all exposed fish. This was evidenced by the Kaplan-Meier survival curves and their differential sensitivity. Low concentrations produced mortality in S. lermae and X. variata, while G. atripinnis and A. robustus were more tolerant to higher NaNO3-N concentrations. The level of affectation, due to NO3-N exposure, between goodeine species suggests differential tolerance. Reference parameters for NaNO3-N acute toxicity in freshwater fish offspring are consistent with the results of G. atripinnis and A. robustus (Supplementary Table 3). However, the values for S. lermae and X. variata suggest they are the most sensitive species currently reported.
Sensitivity, ontogenetic stage, and body size
Fish responses to NO3-N depend on adsorption (accumulation time in plasma), distribution, metabolism, and excretion (Spurgeon et al., 2020). A relationship between sensitivity and ontogenetic stage has been found. Hamlin (2006) suggested that offspring were more tolerant to elevated NO3-N than adults, contrary to other studies where earlier life stages were more sensitive (Adelman et al., 2009). Also, species with a bigger body size are less sensitive to NO3-N (Camargo et al., 2005). In fact, a recent study in relatively larger species found that low NO3-N concentrations did not alter growth and reproduction (Syed et al., 2022).
In this study, the four Goodeinae species had the same age during NaNO3-N exposure but differed in body size and developmental stage. Skiffia lermae was the smallest species, followed by X. variata, while G. atripinnis and A. robustus were the largest species. Likewise, S. lermae have earlier maturation and begin their reproductive life approximately 3 months after birth (body length: 29.5 ± 5.7 mm in females and 25.2 ± 5.4 mm in males. Ramírez-García et al., 2021). In contrast, G. atripinnis start its reproductive stage at 8 months (Silva-Santos et al., 2016), with a body length of 65.8 ± 17.6 mm in females and 65.6 ± 20.7 mm in males (Ramírez-García et al., 2021). Thus, the smallest and most precocious species were the most sensitive, while more tolerant responses were found in larger species.
Swimming behavior reflects differential sensitivity to nitrate-nitrogen
Swimming behavior alterations have been considered as an attempt to face unfavorable environmental conditions, from the regular swimming pattern that occurs in the middle of the water column (Gerhardt, 2007). The behavioral patterns observed in response to acute NaNO3-N were: location on the surface or bottom of the water column, loss of balance and lethargy. Skiffia lermae showed swimming behavior alterations earlier and at lower NaNO3-N concentrations, followed by X. variata, A. robustus, and G. atripinnis. Permanence in the surface of the water column is a compensatory response to inefficient oxygen uptake caused by damage to the gills or by conversion of oxygen-carrying pigments (i.e., hemoglobin) to forms that are incapable of carrying oxygen (i.e., methemoglobin Camargo and Alonso, 2006). Loss of balance and position at the bottom of the water column have been associated with a decreased capacity for respiratory gas exchange, as well as with the depletion of liver lipid reserves (Pereira et al., 2017). Other studies have reported a loss of balance and lethargy in fish exposed to NO3-N (Shimura et al., 2004; Hamlin, 2006; Rodrigues et al., 2011; Pereira et al., 2017).
Histopathological alterations of gills and liver could reflect different sensitivity to nitrate-nitrogen in goodeine species
Remarkably, the LC50 value for A. robustus is similar to C. clupeaformis and O. mykiss, two euryhaline species having proficient ion exchange regulation to tolerate changes between freshwater and marine environments (Camargo et al., 2005; Takvam et al., 2021). This is interesting given that the gills are one of the main organs involved in ion regulation and ammonia excretion, where specific glycoproteins are involved (Hwang et al., 2011). At the histological level, A. robustus showed preserved gill structure after NaNO3-N exposure. This suggests that gill function is maintained due to regressive, progressive and circulatory alterations restricted to the apical region. This was contrary to S. lermae which showed significant damage (highest GHI at minor concentrations), mainly lamellar fusion and cell hyperplasia; or X. variata and G. atripinnis which showed moderate gill alterations, like edema and aneurysms. Gill histopathological alterations in response to water conditions and pollutants, including NO2-N, have been described (Kroupova et al., 2005). The gills play a critical physiological role in nitrogenous waste detoxification; however, the response to NO3-N has not been fully explained (Gomez-Isaza et al., 2020). The branchial epithelium responds to non-optimal environmental changes through circulatory and progressive alterations oriented to preserve gill function. If ecological conditions remain unfavorable, degenerative mechanisms like lamellar fusion are triggered leading to loss of gill function (Antunes et al., 2017; Monsees et al., 2017). Thus, lamellar fusion observed in S. lermae fry could lead to ion-exchange failure and consequent fish anoxia. The different degrees of gill tissue damage observed in X. variata, G. atripinnis, and A. robustus suggest different mechanisms to face NO3-N exposure.
Liver histopathology also confirmed differential tolerance due to variation in circulatory, progressive and regressive alterations. These constitute the LHI, which showed similar values between species. Xenotoca variata and A. robustus showed the highest regressive index associated with hepatocyte cytoplasmic vacuolation, pyknosis, and a relative decrease in nuclear density. Meanwhile, S. lermae and G. atripinnis showed circulatory and progressive alterations, respectively. The liver has an essential role in metabolic pathways and pollutant detoxification (Bruslé and Anadon, 1996). Exposure to environmental pollutants could result in erythrocyte accumulation in sinusoids (hyperemia), vascular dilation and hemorrhage, as well as hepatocyte hypertrophy. The degenerative indicators were hepatocyte cytoplasmic vacuolation, pyknosis, and decreased nuclear density. An increased size of hepatocyte nuclei (nuclear hypertrophy) reveals higher transcriptional and metabolic activity in response to pollutants. In contrast, a decreased cell density is related to liver degeneration (Strüssmann and Takashima, 1990). Here the hepatocyte nuclear area and number of cell nuclei were quantified and integrated into the LDTI. All species showed an increased cell nuclear area and a decreased number of cell nuclei. However, X. variata presented the highest liver damage, followed by A. robustus and S. lermae, whereas G. atripinnis showed less liver damage.
Combined histopathological evaluation suggested that gill and liver damage were associated with alteration of critical metabolic processes, such as inhibition of protein synthesis, depletion of glycogen reserves and reduced ability to detoxify environmental pollutants (Mohamed, 2009). In this regard, S. lermae showed the most severe gill damage and a relatively well-preserved liver structure. On the contrary, X. variata showed a high GHI and the highest LHI, whereas G. atripinnis and A. robustus showed lower GHI and LHI. Interestingly, G. atripinnis showed a higher GHI than A. robustus but a lower LHI. This supports the hypothesis that species that preserve gill structure and function, letting the liver carry out detoxification, likely have better survival opportunities. It has been demonstrated that lamellar fusion in response to acute NO3-N exposure limits the absorption of toxic substances and inhibits ion exchange (Monsees et al., 2017).
The inclusion of all evaluated responses into IBR radar graphs allowed identifying the GHI, LHI, and swimming behavior as the main factors that contribute to explain the differential sensitivity of Goodeinae species to NO3-N. Sensitive species (S. lermae and X. variata) showed similar IBR values at the concentration closest to the LC50, the first species showed the most severe gill damage. A certain tolerance level to NO3-N exposure was found in G. atripinnis and A. robustus species which could be related to the activation of physiological detoxifying strategies for survival. In both tolerant species, gill damage was minor, suggesting that fish species showing gill NO3-N tolerance have higher survival opportunities in polluted freshwater bodies.
The results show that NO3-N are toxic since they produce behavioral, branchial, and hepatic alterations at a lower concentration in goodeines. Thus, it is recommended that their concentrations be monitored in freshwater bodies where goodeines inhabit. Their toxic effect may be increased by alterations in other water quality parameters like pH, hardness, and temperature. In this study, most water quality variables were close to Zacapu lake parameters; except salinity, TDS and conductivity, which are directly associated with the sodium ions in solution. Due to the large difference in progeny between species (S. lermae and X. variata females produce a mean of 12 and 38 hatchlings, respectively; whereas G. atripinnis and A. robustus females produce larger progeny of up to 150 offspring. Ramírez-García et al., 2021), this study evaluated offspring from a different number of gravid females. These differences in fecundity between goodeine species, in addition to differential sensitivity to NaNO3-N, could increase the ecological risk for S. lermae and X. variata in wild freshwater ecosystems. However, population comparisons were outside the scope of this study.
Conclusion
Goodeinae species show a differential NaNO3-N sensitivity. Since the most sensitive species showed higher gill damage, differential sensitivity could be related to different physiological mechanisms to face NO3-N. Thus, species that preserve gill function and transfer the task of detoxification to the liver might have the best chance of surviving in polluted environments. Future studies should evaluate ecologically relevant NO3-N concentrations to confidently predict goodeines persistence in polluted freshwater bodies.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was reviewed and approved by the Mexican Ministry of Environmental and Natural Resources (SEMARNATSGPA/DGVS/00012/19) and the Secretariat of Agriculture and Rural Development (SAGARPA: PPF/DGOPA-014/20).
Author contributions
IV-V: investigation, methodology, formal analysis, and writing—original draft. BY-R: formal analysis and writing—review and editing. RR-J: conceptualization and methodology. MH-V and RH-M: methodology. OD-D: funding acquisition, conceptualization, and writing—review and editing. EM-H: funding acquisition, data curation, writing—original draft, and writing—review and editing. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the Coordinación de la Investigación Científica-UMSNH to EM-H and OD-D and Chester Zoo Garden, Association Beauval Nature Pour la Conservation et la Recherche, Willema Zoo, Ostrava Zoo, The Mohammed Bin Zayed Species Conservation Found, Haus des Meeres—Aqua Terra Zoo, Poecilia Scandinavia, zoologische gesellschaft für arten- und populationsschutz, Goodeid Working Group, American Livebearers Association to OD-D. IV-V was Ph.D. fellow from CONACYT (grant no. 743476).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.1014814/full#supplementary-material
References
Adelman, I., Kusilek, L., Koehle, J., and Hess, J. (2009). Acute and chronic toxicity of ammonia, nitrite, and nitrate to the endangered topeka shiner (Notropis topeka) and fathead minnows (Pimephales promelas). Environ. Toxicol. Chem. 28, 2216–2223. doi: 10.1897/08-619.1
American Public Health Association [APHA] (2017). Standard methods for the examination of water and wastewater, 23rd Edn. Washington, DC: APHA.
Antunes, A., Rocha, T., Pires, F., de Freitas, M., Leite, V., Arana, S., et al. (2017). Gender-specific histopathological response in guppies Poecilia reticulata exposed to glyphosate or its metabolite aminomethylphosphonic acid. J. Appl. Toxicol. 37, 1098–1107. doi: 10.1002/jat.3461
Bancroft, J., and Cook, H. (1994). Manual of histological techniques and their diagnostic application. Edinburgh: Churchill Livingstone.
Bernet, D., Schmidt, H., Meier, W., and Wahli, T. (1999). Histopathology in fish: Proposal for a protocol to assess aquatic pollution. J. Fish Dis. 22, 25–34. doi: 10.1046/j.1365-2761.1999.00134.x
Bertrand, L., Asis, R., Monferrán, M., and Amé, M. (2016). Bioaccumulation and biochemical response in South American native species exposed to zinc: Boosted regression trees as novel tool for biomarkers selection. Ecol. Indic. 67, 769–778. doi: 10.1016/J.ECOLIND.2016.03.048
Bruslé, J., and Anadon, G. (1996). “The structure and function of fish liver,” in Fish Morphology, eds J. Munshi and H. Dutta (North-Holland: Science Publishers), 77–93. doi: 10.1016/j.ecolind.2016.03.048
Camargo, J., and Alonso, A. (2006). Ecological and toxicological effects of inorganic nitrogen pollution in aquatic ecosystems: A global assessment. Environ. Int. 32, 831–849. doi: 10.1016/j.envint.2006.05.002
Camargo, J., Alonso, A., and Salamanca, A. (2005). Nitrate toxicity to aquatic animals: A review with new data for freshwater invertebrates. Chemosphere 58, 1255–1267. doi: 10.1016/j.chemosphere.2004.10.044
Cano-Rocabayera, O., Sostoa, A., Padrós, F., Cárdenas, L., and Maceda-Veiga, A. (2019). Ecologically relevant biomarkers reveal that chronic effects of nitrate depend on sex and life stage in the invasive fish Gambusia holbrooki. PLos One 14:e0211389. doi: 10.1371/journal.pone.0211389
Corredor-Santamaría, W., Mora-Solarte, D., Arbeli, Z., Navas, J., and Velasco-Santamaría, J. (2021). Liver biomarkers response of the neotropical fish Aequidens metae to environmental stressors associated with the oil industry. Heliyon 7, 1–8. doi: 10.1016/j.heliyon.2021.e07458
Domínguez-Domínguez, O., Martínez-Meyer, E., Zambrano, L., and De León, G. (2006). Using ecological-niche modeling as a conservation tool for freshwater species: Live-bearing fishes in central Mexico. Conserv. Biol. 20, 1730–1739. doi: 10.1111/j.1523-1739.2006.00588.x
Domínguez-Domínguez, O., Zambrano, L., Escalera-Vázquez, L., Pérez-Rodríguez, R., and Pérez-Ponce de León, G. (2008). Cambio en la distribución de Goodeidos (Osteichthyes: Cyprinodontiformes: Goodeidae) en cuencas hidrológicas del centro de México. Rev. Mex. Biodivers. 79, 501–512.
Du, R., Cao, S., Peng, Y., Zhang, H., and Wang, S. (2019). Combined partial denitrification (PD)-anammox: A method for high nitrate wastewater treatment. Environ. Int. 126, 707–716. doi: 10.1016/j.envint.2019.03.007
Duncan, J., and Lockwood, J. (2001). Extintion in a field of bullets: A search for causes in the decline of the world’s freshwater fishes. Biol. Conserv. 102, 97–105. doi: 10.1016/S0006-3207(01)00077-5
Dutra, F., Cidemar Alab, J., Costa Gómez, M., Furtado, P., Valenti, W., and Cupertino-Ballester, E. (2020). Nitrate acute toxicity to post larvae and juveniles of Macrobrachium amazonicum (Heller, 1862). Chemosphere 242:125229. doi: 10.1016/j.chemosphere.2019.125229
Emam, M., Caballero-Solares, A., Xue, X., Umasuthan, N., Milligan, B., Taylor, R., et al. (2022). Gill and liver transcript expression changes associated with gill damage in Atlantic Salmon (Salmo salar). Front. Immunol. 13:806484. doi: 10.3389/fimmu.2022.806484
Foster, K., and Piller, K. (2018). Disentangling the drivers of diversification in an imperiled group of freshwater fishes (Cyprinodontiformes: Goodeidae). BMC Evol. Biol. 18:116. doi: 10.1186/s12862-018-1220-3
Galloway, J., Denterner, F., Capone, D., Boyer, E., Haowarth, R., Seitzinger, S., et al. (2004). Nitrogen cycles: Past, present, and future. Biogeochemistry 70, 153–226. doi: 10.1007/s10533-004-0370-0
Gerhardt, A. (2007). Aquatic behavioral ecotoxicology-prospects and limitations. Hum. Ecol. Risk Assess. 13, 481–491. doi: 10.1080/10807030701340839
Goeller, B., Febria, C., Warburton, H., Hogsden, K., Collins, K., Devlin, H., et al. (2019). Springs drive downstream nitrate export from artificially-drained agricultural headwater catchments. Sci. Total Environ. 671, 119–128. doi: 10.1016/j.scitotenv.2019.03.308
Gomez-Isaza, D., Cramp, R., and Franklin, C. (2018). Negative impacts of elevated nitrate on physiological performance are not exacerbated by low pH. Aquat. Toxicol. 200, 217–225. doi: 10.1016/j.aquatox.2018.05.004
Gomez-Isaza, D., Cramp, R., and Franklin, C. (2020). Living in polluted waters: A meta-analysis of the effects of nitrate and interactions with other environmental stressors on freshwater taxa. Environl Pol 261, 1–12. doi: 10.1016/j.envpol.2020.114091
Gomez-Isaza, D., Cramp, R., and Franklin, C. (2021). Exposure to Nitrate Increases Susceptibility to Hypoxia in Fish. Physiol. Biochem. Zool. 94, 124–142. doi: 10.1086/713252
Guide for the Care and Use of Laboratory Animals (1996). National research council of the National Academies, 8th Edn. (Washington, DC: The National Academies Press), 246.
Guillette, L., and Edwards, T. (2005). Is nitrate an ecologically relevant endocrine disruptor in vertebrates? Integr. Comp. Biol. 45, 19–27. doi: 10.1093/icb/45.1.19
Hamlin, H. (2006). Nitrate toxicity in Siberian sturgeon (Acipenser baeri). Aquaculture 253, 688–693. doi: 10.1016/j.aquaculture.2005.08.025
Hwang, P., Lee, T., and Lin, L. (2011). Ion regulation in fish gills: Recent progress in the cellular and molecular mechanisms. Am. J. Physiol. Regul. Integr. Comp. Physiol. 301, 28–47. doi: 10.1152/ajpregu.00047.2011
Iida, A., Arai, H., Someya, Y., Inokuchi, M., Onuma, T., Yokoi, H., et al. (2019). Mother-to-embryo vitellogenin transport in a viviparous teleost Xenotoca eiseni. Proc. Natl. Acad. Sci. U.S.A. 116, 22359–22365. doi: 10.1073/pnas.1913012116
IUCN (2022). The IUCN red list of threatened species. Version 2021-3. Available online at: https://www.iucnredlist.org (accessed May 15, 2022).
Kim, J., Kang, Y., Kim, K., Kim, S., and Kim, J. (2019). Toxic effects of nitrogenous compounds (ammonia, nitrite, and nitrate) on acute toxicity and antioxidant responses of juvenile olive flounder, Paralichthys olivaceus. Environ. Toxicol. Pharmacol. 67, 73–78. doi: 10.1016/j.etap.2019.02.001
Kroupova, K., Máchová, J., and Svobodova, Z. (2005). Nitrite influence on fish: A review. Vet. Med. 50, 461–471.
Lyons, J., Piller, K., Artigas-Azas, J., Dominguez-Dominguez, O., Gesundheit, P., and Köck, et al. (2019). Distribution and current conservation status of the Mexican Goodeidae (Actinopterygii, Cyprinodontiformes). ZooKeys 885, 115–158. doi: 10.3897/zookeys.885.38152
McGurk, M., Landry, F., Tang, A., and Hanks, C. (2006). Acute and chronic toxicity of nitrate to early life stages of lake trout (Salvelinus namaycush) and lake whitefish (Coregonus clupeaformis). Environ. Toxicol. Chem. 25, 2187–2196. doi: 10.1897/05-270r.1
Miller, R., Minckley, W., and Norris, S. (2009). Freshwater fishes of Mexico. Chicago, IL: University of Chicago Press.
Mohamed, F. (2009). Histopathological studies on Tilapia zillii and Solea vulgaris from Lake Qarun, Egypt. World J. Fish Mar. Sci. 1, 29–39.
Monsees, H., Klatt, L., Kloas, W., and Wuertz, S. (2017). Chronic exposure to nitrate significantly reduces growth and affects the health status of juvenile Nile tilapia (Oreochromis niloticus L.) in recirculating aquaculture systems. Aquac. Res. 48, 3482–3492. doi: 10.1111/are.13174
Ogueji, O., Nwani, C., Iheanacho, S., Mbah, C., Okeke, C., and Yaji, A. (2018). Acute toxicity effects of ibuprofen on behaviour and haematological parameters of African catfish Clarias gariepinus (Burchell, 1822). Afr. J. Aquat. Sci. 43, 293–303. doi: 10.2989/16085914.2018.1465393
OECD (2019). Test No. 203: Fish, acute toxicity test, OECD guidelines for the testing of chemicals, section 2. Paris: OECD Publishing.
Pereira, A., Carvalho, A., Cruz, C., and Saraiva, A. (2017). Histopathological changes and zootechnical performance in juvenile zebrafish (Danio rerio) under chronic exposure to nitrate. Aquaculture 473, 197–205. doi: 10.1016/j.aquaculture.2017.02.018
Presa, L., Neves, G., Maltez, L., Sampaio, L., Monserrat, J., Copatti, C., et al. (2022). Acute and sub-lethal effects of nitrate on haematological and oxidative stress parameters of juvenile mullet (Mugil liza) in freshwater. Aquac. Res. 53, 3346–3357. doi: 10.1111/are.15842
R Core Team (2022). R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
Ramírez-García, A., Domínguez-Domínguez, O., López-López, E., Moncayo-Estrada, R., and Cruz-Agüero, J. (2022). Temporal changes in the fish community structure of a small subtropical spring lake: Decadal and seasonal analysis. Ecol. Freshw. Fish. [Epub ahead of print]. doi: 10.1111/eff.12669
Ramírez-García, A., Moncayo-Estrada, R., González-Cárdenas, J. J., and Domínguez-Domínguez, O. (2021). Reproductive cycle of native viviparous fish species (Actinopterygii: Cyprinodontiformes: Goodeidae) in a subtropical Mexican lake. Neotrop. Ichthyol. 19, 1–20. doi: 10.1590/1982-0224-2021-0105
Rodrigues, R., Saturnino, K., and Fernandes, C. (2017). Liver histology and histomorphometry in hybrid sorubim (Pseudoplatystoma reticulatum × Pseudoplatystoma corruscans) reared on intensive fish farming. Aquac. Res. 48, 5083–5093. doi: 10.1111/are.13325
Rodrigues, R., Schwarz, M., Delbos, B., Carvalho, E., Romano, L., and Sampaio, L. (2011). Acute exposure of juvenile cobia Rachycentron canadum to nitrate induces gill, esophageal and brain damage. Aquaculture 322-323, 223–226. doi: 10.1016/j.aquaculture.2011.09.040
Rueda-Jasso, R., De los Santos-Bailón, A., and Campos-Mendoza, A. (2017). Nitrite toxicity in juvenile Goodeinae fishes Skiffia multipunctata (Pellegrin, 1901) and Goodea atripinnis (Jordan, 1880). J. Appl. Ichthyol. 33, 300–305. doi: 10.1111/jai.13292
Santos, R., Monteiro, S., Cortes, R., Pacheco, F., and Fernandes, L. (2022). Seasonal differences in water pollution and liver histopathology of Iberian Barbel (Luciobarbus bocagei) and Douro Nase (Pseudochondrostoma duriense) in an Agricultural Watershed. Water 14, 1–22. doi: 10.3390/w14030444
Schram, E., Roques, J., Abbink, W., Yokohama, Y., Spannings, T., de Vries, P., et al. (2012). The impact of elevated water nitrate concentration on physiology, growth and feed intake of African catfish Clarias gariepinus (Burchell 1822). Aquac. Res. 45, 1499–1511. doi: 10.1111/are.12098
Shimura, R., Ma, Y., Ijiri, K., Nagaoka, S., and Uchiyama, M. (2004). Nitrate toxicity on visceral organs of Medaka fish, Oryzias latipes: Aiming to raise fish from egg to egg in space. Biol. Sci. Space 18, 7–12. doi: 10.2187/bss.18.7
Silva-Santos, J., Martínez-Saldaña, M., Rico-Martínez, R., Gómez, J., and Arredondo-Figueroa, J. (2016). Reproductive biology of Goodea atripinnis (Jordan, 1880) (cyprinodontiformes: goodeidae) under controlled conditions. J. Exp. Biol. Agric. Sci. 4, 180–193. doi: 10.18006/2016.4(2)0.180.193
Spurgeon, D., Lahive, E., Robinson, A., Short, S., and Kille, P. (2020). Species sensitivity to toxic substances: Evolution, ecology and applications. Front. Environ. 8:588380. doi: 10.3389/fenvs.2020.588380
Strüssmann, C., and Takashima, F. (1990). Hepatocyte nuclear size and nutritional condition of larval pejerrey, Odontesthes bonariensis (Cuvier et Valenciennes). J. Fish Biol. 36, 59–65. doi: 10.1111/j.1095-8649.1990.tb03519.x
Syed, R., Masood, Z., Hassan, H., Khan, W., Mushtaq, S., Ali, A., et al. (2022). Growth performance, haematological assessment and chemical composition of Nile tilapia, Oreochromis niloticus (Linnaeus, 1758) fed different levels of Aloe vera extract as feed additives in a closed aquaculture system. Saudi J. Biol. Sci. 29, 296–303. doi: 10.1016/j.sjbs.2021.08.098
Takvam, M., Wood, C., Kryvi, H., and Nilsen, T. (2021). Ion transporters and osmoregulation in the kidney of teleost fishes as a function of salinity. Front. Physiol. 12:664588. doi: 10.3389/fphys.2021.664588
Torno, J., Einwächter, V., Schroeder, J., and Schulz, C. (2018). Nitrate has a low impact on performance parameters and health status of on-growing European sea bass (Dicentrarchus labrax) reared in RAS. Aquaculture 489, 21–27. doi: 10.1016/j.aquaculture.2018.01.043
Tramunt, B., Montagner, A., Tan, N., Gourdy, P., Rémignon, H., and Wahli, W. (2021). Roles of estrogens in the healthy and diseased oviparous vertebrate liver. Metabolites 11:502. doi: 10.3390/metabolo11080502
van Bussel, C., Schroeder, J., Wuertz, S., and Schulz, C. (2012). The chronic effect of nitrate on production performance and health status of juvenile turbot (Psetta maxima). Aquaculture 326-329, 163–167. doi: 10.1016/j.aquaculture.2011.11.019
Yang, X., Song, X., Peng, L., Hallerman, E., and Huang, Z. (2019). Effects of nitrate on aquaculture production, blood and histological markers and liver transcriptome of Oplegnathus punctatus. Aquaculture 501, 387–396. doi: 10.1016/j.aquaculture.2018.11.048
Yu, J., Wang, Y., Xiao, Y., Li, X., Zhou, L., Wang, Y., et al. (2021). Investigating the effect of nitrate on juvenile turbot (Scophthalmus maximus) growth performance, health status, and endocrine function in marine recirculation aquaculture systems. Ecotoxicol. Environ. Saf. 208:111617. doi: 10.1016/j.ecoenv.2020.111617
Keywords: ecotoxicology, LC50 96 h, histopathological changes, swimming behavior, gill, liver
Citation: Villa-Villaseñor IM, Yáñez-Rivera B, Rueda-Jasso RA, Herrera-Vargas MA, Hernández-Morales R, Meléndez-Herrera E and Domínguez-Domínguez O (2022) Differential sensitivity of offspring from four species of goodeine freshwater fish to acute exposure to nitrates. Front. Ecol. Evol. 10:1014814. doi: 10.3389/fevo.2022.1014814
Received: 09 August 2022; Accepted: 12 September 2022;
Published: 29 September 2022.
Edited by:
Georgina Alexandra Rivera-Ingraham, Universidad Católica del Norte, ChileReviewed by:
Wenyi Zhang, Nanjing Normal University, ChinaAhasan Habib, University of Malaysia Terengganu, Malaysia
Copyright © 2022 Villa-Villaseñor, Yáñez-Rivera, Rueda-Jasso, Herrera-Vargas, Hernández-Morales, Meléndez-Herrera and Domínguez-Domínguez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Esperanza Meléndez-Herrera, ZW1lbGVuZGV6QHVtaWNoLm14