 Lauryn Benedict
Lauryn Benedict Braelei Hardt
Braelei Hardt Lorraine Dargis
Lorraine Dargis- School of Biological Sciences, University of Northern Colorado, Greeley, CO, United States
To function effectively, animal signals must transmit through the environment to receivers, and signal transmission properties depend on signal form. Here we investigated how the transmission of multiple parts of a well-studied signal, bird song, varies between males and females of one species. We hypothesized that male and female songs would have different transmission properties, reflecting known differences in song form and function. We further hypothesized that two parts of male song used differentially in broadcast singing and aggressive contests would transmit differently. Analyses included male and female songs from 20 pairs of canyon wrens (Catherpes mexicanus) played and re-recorded in species-typical habitat. We found that male song cascades used in broadcast singing propagated farther than female songs, with higher signal-to-noise ratios at distance. In contrast, we demonstrated relatively restricted propagation of the two vocalization types typically used in short-distance aggressive signaling, female songs and male “cheet” notes. Of the three tested signals, male “cheet” notes had the shortest modeled propagation distances. Male and female signals blurred similarly, with variable patterns of excess attenuation. Both male song parts showed more consistent transmission across the duration of the signal than did female songs. Song transmission, thus, varied by sex and reflected signal form and use context. Results support the idea that males and females of the same species can show distinctly different signal evolution trajectories. Sexual and social selection pressures can shape sex-specific signal transmission, even when males and females are communicating in shared physical environments.
Introduction
Animal communication signals are shaped by many factors, including signaler and receiver morphologies, physiology, physical environments, and social environments (Brenowitz et al., 1997; Slabbekoorn and Smith, 2002; Podos et al., 2004; Bradbury and Vehrencamp, 2011). Signal evolution has been well studied in birds, with extensive research examining the form and function of male broadcast songs (Thorpe, 1961; Catchpole and Slater, 2008). Much less work has examined the properties of female songs (Odom and Benedict, 2018; Riebel et al., 2019). Broadcast songs, by their very nature, should be adapted to transmit long distances, leading many authors to emphasize that habitats select for increased song propagation distances and propagation consistency (Morton, 1975; Gil and Gahr, 2002; Boncoraglio and Saino, 2007; Apol et al., 2018). Signal efficacy, however, is not purely dependent on transmission distance, and many signals function effectively with short propagation distances (Endler, 1993; Higham and Hebets, 2013). Evaluating the transmission properties of different avian songs can therefore illuminate how evolution shapes signal form in conjunction with function, and will help to explain the diversity of bird songs found in nature. Males and females of the same species provide a particularly interesting test case for such comparisons because they occupy the same physical spaces but can be subject to different sexual and social selection regimes (West-Eberhard, 1983).
It is well established that environments can affect the propagation of animal sounds (Wiley and Richards, 1978, 1982; Forrest, 1994; Brumm and Naguib, 2009). In multiple systems, bird songs transmit well in their local environments (Blumenrath and Dabelsteen, 2004; Nemeth and Brumm, 2010; Ręk and Kwiatkowska, 2016). At a larger scale, however, it is becoming clear that not all songs have evolved for maximum transmission distance (Boncoraglio and Saino, 2007; Ey and Fischer, 2009). Factors including signaler identity, song function, and social context can shape bird songs in ways that override potential benefits of long-distance transmission (Greig et al., 2013; Mikula et al., 2020). Indeed, one branch of research emphasizes that many avian vocalizations have a form and delivery style that is adapted for short-distance transmission, often in aggressive contexts (Dabelsteen et al., 1998; Hof and Hazlett, 2010; Akçay et al., 2015; Zollinger and Brumm, 2015). Within-species comparisons have demonstrated that short-distance (quiet) songs show reduced transmission, even when played at the same amplitude as long-distance (loud) songs (Rek, 2013; Vargas-Castro et al., 2017; but see Niederhauser et al., 2018). Long- and short-distance vocalizations often differ in form, with short-distance vocalizations having higher frequencies and wider bandwidths and less energy concentrated in pure tones (Piza and Sandoval, 2016). In general, such buzzy broadband sounds are typical of animal aggressive signals (Morton, 1977; Blumstein and Récapet, 2009). Many short-distance bird songs function in agonistic contexts where reduced transmission allows for aggressive signaling to a nearby rival while simultaneously reducing eavesdropping by unwanted signal receivers (Akçay et al., 2015; Vargas-Castro et al., 2017).
Although long ignored in temperate areas of the world, female bird song is widespread (Langmore, 1998; Odom et al., 2014). Males and females, even when occupying the same territories, often exhibit differentiated sex roles and may experience distinct selective pressures on their communication, particularly in temperate latitudes (Slater and Mann, 2004; Price et al., 2009; Hall and Langmore, 2017). Comparisons of male and female song features, therefore, have the potential to illuminate different signal evolution trajectories within a single species. We are aware of only four previous studies comparing the transmission of male and female vocalizations from one species; together, their results show that sex-specific calls, songs, and duets can propagate differently through space (Mennill et al., 2009; Mouterde et al., 2014; Sandoval et al., 2015; Graham et al., 2017). In tropical rufous-and-white-wrens (Thryophilus rufalbus), the only species for which sex-specific song transmission has been demonstrated, male and female songs are generally similar in structure and are often used in similar contexts (Mennill and Vehrencamp, 2005). In other avian species, male and female songs have very different structures that reflect sex-specific functional differences.
In the study reported here we investigated the transmission properties of male and female bird songs with highly sex-specific structures and different known contextual use patterns. Canyon wrens (Catherpes mexicanus) are monomorphic, year-round residents in Western North America that live in monogamous territorial pairs (Jones and Dieni, 2020). Males use song as a broadcast signal of territory occupation and sing frequently throughout the breeding season (Benedict et al., 2013; Rose, 2013). Females almost never sing spontaneous broadcast song, but do use song to defend resources; they are quick to sing when challenged on their territories by conspecific female song (Hathcock and Benedict, 2018; Dargis et al., 2021). Thus, female songs are most often used in short-distance aggressive signaling. Male and female canyon wren songs have distinct structures, but similar durations (Figure 1). Male songs always include a descending cascade of pure-toned notes that are well-known among bird enthusiasts for ringing off canyon walls over long distances (Lopez, 2011; Jones and Dieni, 2020). Males often conclude songs by appending one or more buzzy “cheet” notes to song cascades. Female canyon wren songs always consist of a rising and then falling series of buzzy notes, sometimes with harmonics (Spencer, 2012). Both males and females vary note form within songs (Benedict et al., 2013, Supplementary Material).
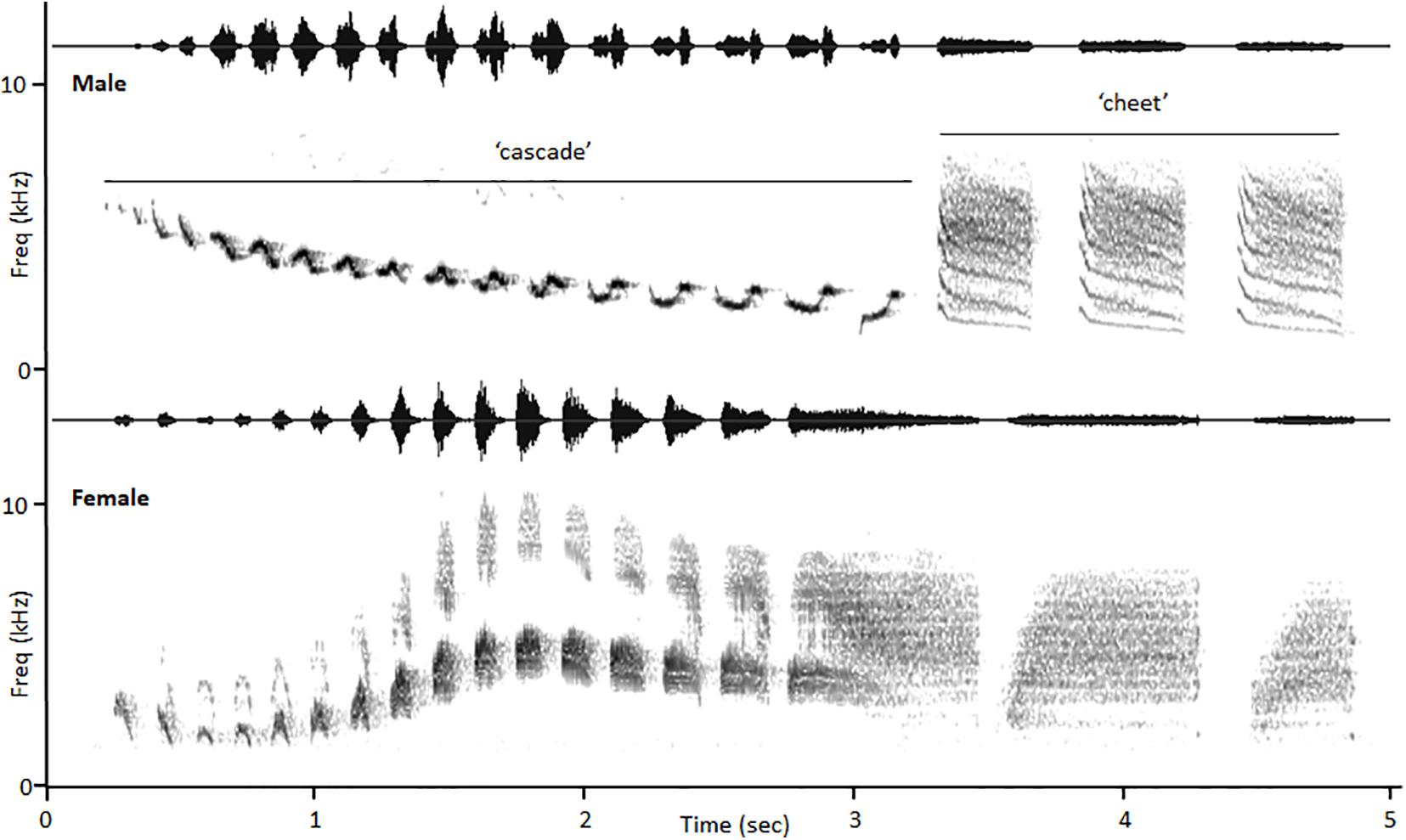
Figure 1. Spectrograms of male and female canyon wren songs, with the “cascade” and “cheet” portions of the male song indicated. Songs were recorded from a mated pair in Arizona (GIS: 31.908498, −109.188094) on June 7, 2019.
Theory suggests that buzzy female canyon wren songs should travel a limited distance, allowing signalers to communicate effectively with rivals and avoid eavesdropping (Vargas-Castro et al., 2017). The pure-toned notes of male canyon wren song cascades, in contrast, are expected to carry long distances, broadcasting male presence across widely spaced territories (Warning et al., 2015; Benedict and Warning, 2017). Interestingly, when male canyon wrens are challenged in aggressive confrontations, they adjust their songs to reduce the length of the cascade and to add or increase the number of terminal “cheet” notes (Benedict et al., 2012). Male “cheet” notes are much more broadband than cascade notes and female song notes (Figure 1). Thus, for our analyses we compared the transmission of female songs with the two parts of male song and we compared male cascades and “cheet” notes. We predicted that male cascades would propagate best over long distances. As all male songs include the cascade, we considered it to be the fundamental unit of male song for comparison with female songs and we focus on that male-female comparison. We also predicted that male cascades would propagate better than male “cheet” notes over distance.
For all comparisons, we assessed three measures of signal transmission often used in studies investigating acoustic adaptation in birds (Dabelsteen et al., 1993) as well as a novel metric describing the consistency of signal propagation by quantifying transmission variability across the beginning, middle and end portions of the sound. We predicted that male cascades, the main broadcast signal of this species, would show the highest signal consistency. We believe this metric offers a more complete picture of signal propagation in our study species, as it has been long hypothesized that birds living in open spaces should adapt their signals to improve propagation consistency instead of, or in addition to, transmission distance (Morton, 1975; reviewed in Boncoraglio and Saino, 2007; Hardt and Benedict, 2020).
Methods
Playback Tracks
Our canyon wren song playback exemplars (spectrograms in Supplementary Material) came from previous studies of this species conducted by our research group. For the male-female song comparisons we created 20 pairs of audio tracks representing male and female songs recorded from mated pairs during the same recording sessions. Most (n = 30) recordings were made by one author (LD) in Colorado in 2018 and in Arizona in 2019 during experiments where males and females were induced to approach a speaker playing conspecific song (Dargis et al., 2021). Remaining tracks (n = 8) came from a 2016 study with similar methods (Hathcock and Benedict, 2018). In both studies, males and females typically approached the playback speaker together, allowing recordings from both members of each pair to be made from a similar distance. Songs were recorded as .wav files using a Sennheiser MKH-60 or MKH-70 shotgun microphone connected to a Marantz PMD 661 MKII solid state digital recorder, with a sampling rate of 48 kHz, and a depth of 24 bits. For each pair we created a track containing the best quality recorded song for the male and another track containing the best quality recorded song for the female. This led to inclusion of seven different male song cascade types (Benedict et al., 2013). Using Audacity,1 we normalized each track to a peak amplitude of −0.5 dB, and filtered background noise using the “noise reduction” tool with a 20 dB reduction and a 1-s Noise Profile from immediately preceding the song in each recording (sensitivity = 6.0, 3 frequency smoothing bands). Noise Profile filtering approaches are effective at removing noise while preserving signal acoustic characteristics (Baker and Logue, 2007). We filtered out frequencies under 1,200 Hz in most tracks and under 1,600 Hz in a minority of tracks that had excess low-frequency background noise. Male and female song tracks from the same pair were always prepared with the same methods.
Fourteen of our 20 male song tracks included both the cascade notes and at least one terminal “cheet” note. To increase the sample size of the cascade vs. “cheet” note comparison, we added six additional male song tracks, created as described above. These recordings were made during the same studies as the female song tracks, but generally came from territories where only males were recorded. This provided us with a sample of 20 male songs for cascade vs. “cheet” transmission comparisons and for female song vs. “cheet” comparisons.
Playback Methods
We performed song transmission experiments on June 17th, 2019 between 8:30 and 9:30 a.m. in Northern Colorado at a site with appropriate rocky, arid canyon wren habitat (GIS coordinates: 40°26′5, −105°13′31) (Warning et al., 2015). Male and female songs were broadcast in a SSE direction along a cliff ledge at a SPL of 85 dB measured at 1 m with a sound level meter (Extech 407732); this song amplitude mimics natural canyon wren behavior (Dargis et al., 2021). The playback track was ordered by pair, with female and male songs alternating. Additional male songs with “cheet” notes were played at the end of the track. The playback session lasted just under 8 min. We chose our timing, location, and orientation to reflect natural singing behavior in this species (Jones and Dieni, 2020). Temperature and humidity during recording averaged 18.8°C and 32%, respectively, which reflect averages seen during male singing bouts on previous projects (Hardt, unpublished data). To avoid acoustic interference, we recorded playbacks on a clear day when average windspeed was below 8 kph.
Playback tracks were broadcast from a speaker positioned 1 m above the ground and re-recorded at an equal height 10, 50, and 100 m away to simulate potential listeners across a canyon wren’s territory. All three simultaneous recordings were made with Sennheiser MKH-60 shotgun microphones connected to Marantz PMD 661 MKII solid state digital recorders, with identical levels and settings; a sampling rate of 48 kHz, and a depth of 24 bits. Subsequently, we recorded each playback track in a featureless, open field at 1 m away from the speaker using the same equipment as above to generate a reference, non-degraded sound necessary for computation of degradation measures (Dabelsteen et al., 1993).
Acoustic Analysis
We analyzed each song using Raven Pro 1.6,2 selecting each of the non-degraded reference sounds (male cascade, male ‘‘cheet’’ notes or female song) in a box at the visible edges of time and frequency bounds using default spectrogram settings (Hanning type, DFT: 512 samples, grid spacing 86.1 Hz). For consistency, we used these bounds for every recorded version (10, 50, and 100 m) of each song. This was achieved by temporally aligning recordings using a loud tone at the start of the playback track which could be visibly detected on spectrograms at all distances. Once the tone was located on a test recording spectrogram, we copied the time and frequency bounds for each song from the reference recording to the test recordings. We used these selections to compute four degradation thought of as a measurement: blur ratio (BR), signal-to-noise ratio (SNR), excess attenuation (EA), and signal consistency (SC). Blur ratio can be thought of a measurement of the energy retained in a signal as it degrades over distance and was calculated with the package ‘‘baRulho’’ in R v4.0.3 Signal-to-noise ratio represents the relationship of the signal’s energy compared to background noise and was calculated with the equation provided by Dabelsteen et al. (1993). Background noise was sampled as a 1 s time slice preceding each song recording, as is standard in the field (Dabelsteen et al., 1993). At our location, background noise averaged 10.6 dB. Excess attenuation represents the retention of a signal’s clarity over distance while accounting for attenuation caused by atmospheric absorption and spherical spreading. For equations used, see Hardt and Benedict (2020).
Our fourth measure, Signal Consistency, is a novel metric that we used to assess propagation variability across the duration of the signal. To calculate it, we split each assessed signal into three equal time slices representing the beginning, middle, and end of that sound and determined the cross-correlation coefficient of the recording and the reference (Rmax) derived from Hilbert-transformed amplitude envelopes of each section. We determined Rmax with the function “corenv” from the R package “seewave.” We then took the inverse of the standard deviation of Rmax such that a high signal consistency indicates stable propagation over the three sections, and hence the entire signal.
Statistical Analysis
We analyzed data in R v 4.03 using the function “lm” (package: stats). To compare male cascades with female songs, we created a separate model for each of the four transmission measures, and used sex, recording distance, and pair identity for 20 pairs as fixed factors with interactions. For this paired analysis, we removed “cheet” notes from recordings when needed. To examine differences in propagation between the two parts of male song, we modeled each of the four measures for 20 songs with signal type (“cascade” or “cheet”) and distance as interacting fixed factors with song of origin as a covariate. We constructed similar models to compare female songs with male “cheet” notes, but because those comparisons do not use matched pairs, we consider them to be less robust (although still informative) and present them as Supplementary Material. We assessed model fit/adequacy with R squared values and residual vs. fitted value plots. For all models, adjusted R squared values exceeded 0.72 and residuals plots indicated sufficient fit. For each model, we report probability values derived from analysis of variance tables.
Results
Our models comparing male song cascades with female songs confirmed acoustic degradation, as all three measures of signal transmission varied with distance (BR: F = 81.83, p < 0.0001, EA: F = 103.75, p < 0.0001, SNR: F = 369.80, p < 0.0001) (Figure 2 and Table 1). Female songs had lower signal-to-noise ratios (F = 31.93, p < 0.0001) than male song cascades, but blur ratio (F = 1.05, p = 0.31) and excess attenuation were similar for the two sexes (F = 0.97, p = 0.33). Thus, male song cascades retained distinctness from background noise better than female songs at all distances (Figure 2). Signal-to-noise ratio (F = 8.23, p = 0.0006) also showed a significant interaction between sex and distance, with distance promoting more rapid degradation of female songs. Our signal consistency metric indicated that male cascades transmitted more consistently than female songs (F = 25.67, p < 0.0001), and that signal consistency did not vary with distance (F = 0.15, p = 0.70) (Figure 2). Three of four transmission metrics varied with pair identity (EA: F = 9.87, p < 0.0001, SNR: F = 27.16, p < 0.0001. SC: F = 49.09, p < 0.0001; Table 1).
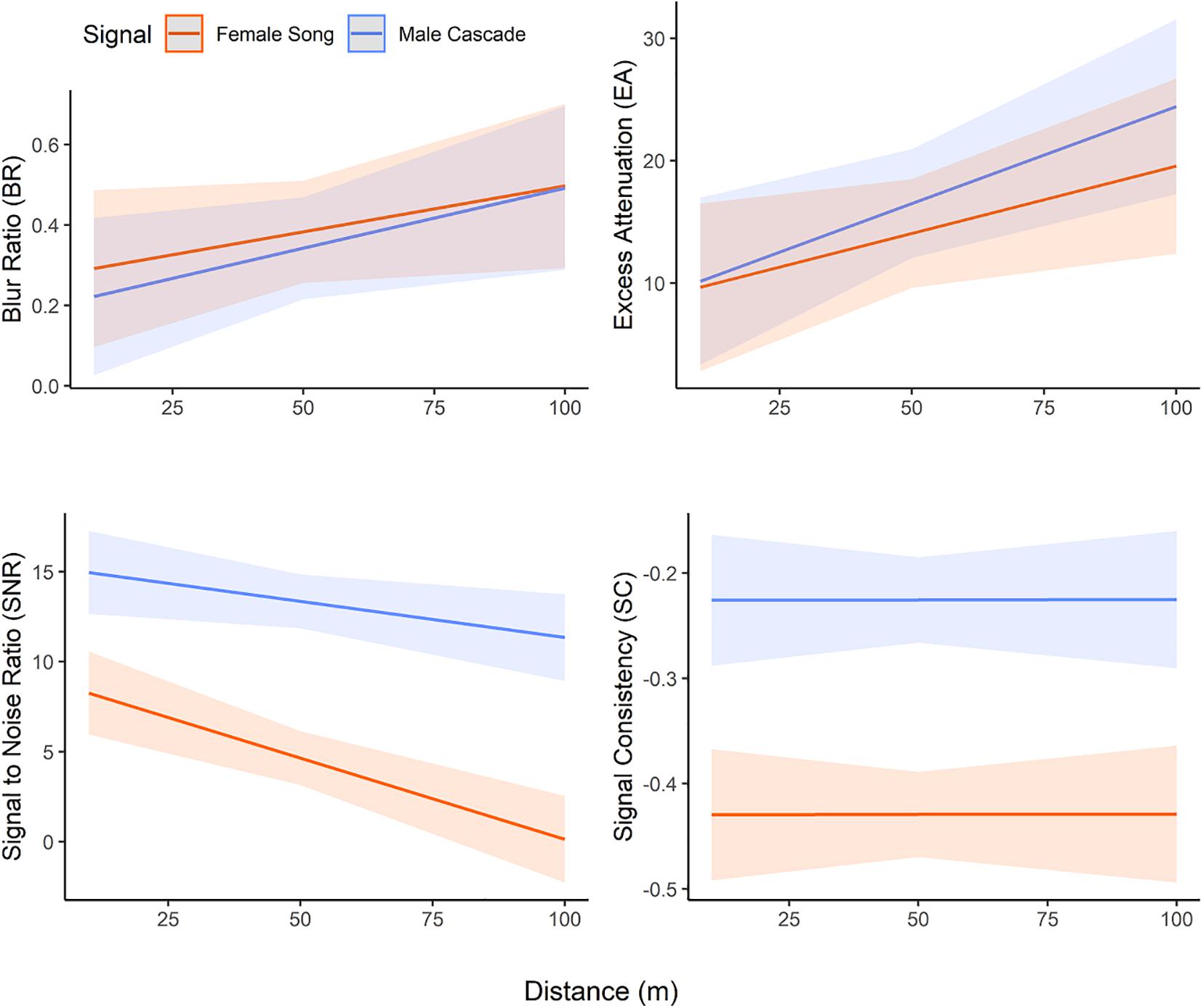
Figure 2. Modeled metrics of propagation quality and consistency for canyon wren female songs and male cascades over distance. Model includes 20 pairs of male and female songs, with sex, distance, and pair identity as fixed factors.
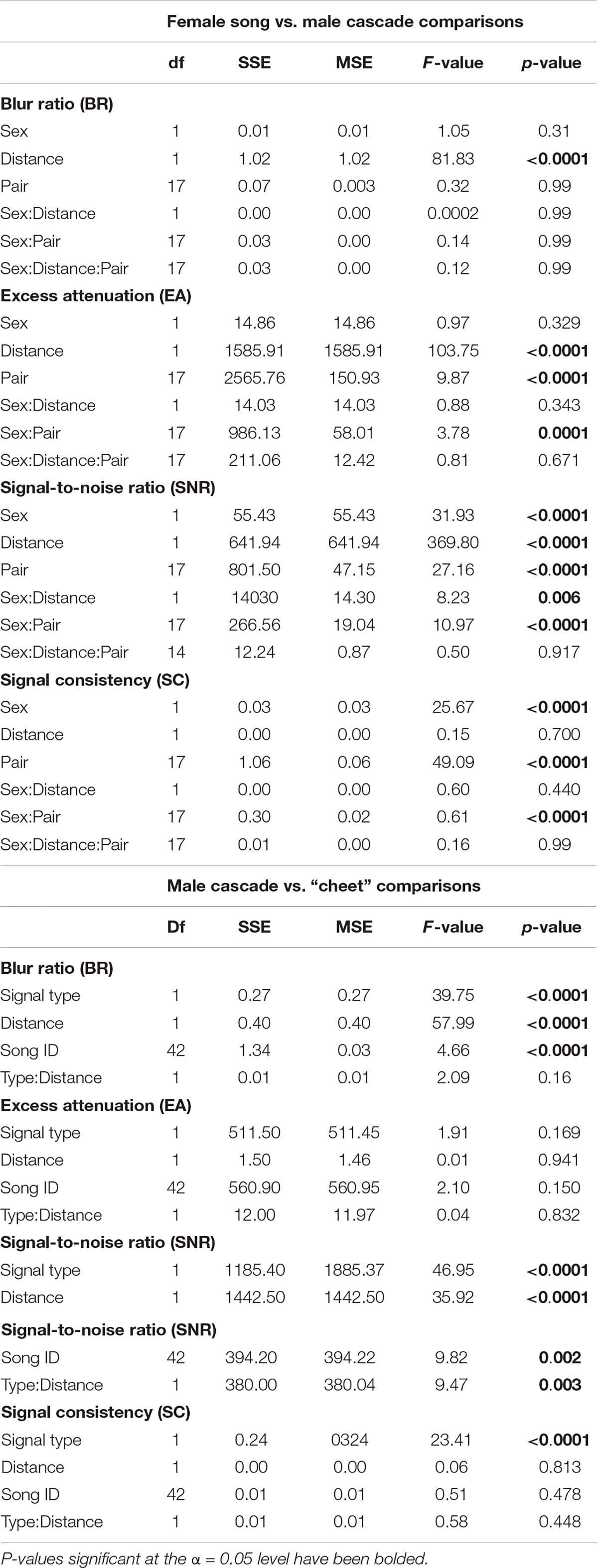
Table 1. ANOVA output for sex-based and within-male canyon wren song transmission comparisons of Blur Ratio (BR), Excess Attenuation (EA), Signal-to-noise Ratio (SNR), and Signal Consistency (SC) measurements.
Comparisons of the two parts of male canyon wren song revealed transmission differences (Figure 3). The buzzy terminal “cheet” notes had significantly lower signal-to-noise ratios (F = 46.95, p < 0.0001), higher blur ratios (F = 39.75, p < 0.0001), and higher signal consistency (F = 28.01, p < 0.0001) than the pure toned song cascades, but did not differ in excess attenuation (F = 1.91, p = 0.17; Table 1). Further, the signal-to-noise ratio of “cheet” notes interacted with distance to decrease faster with distance than did the signal-to-noise ratio of the song cascade (F = 9.47, p = 0.003). Although “cheet” notes degraded more with distance than song cascades, the “cheet” notes showed more consistent transmission (F = 23.41, p < 0.0001) (Figure 3 and Table 1). We also found significant between-song variation in two metrics (BR: F = 4.66, p < 0.0001, SNR: F = 9.82, p = 0.002; Table 1).
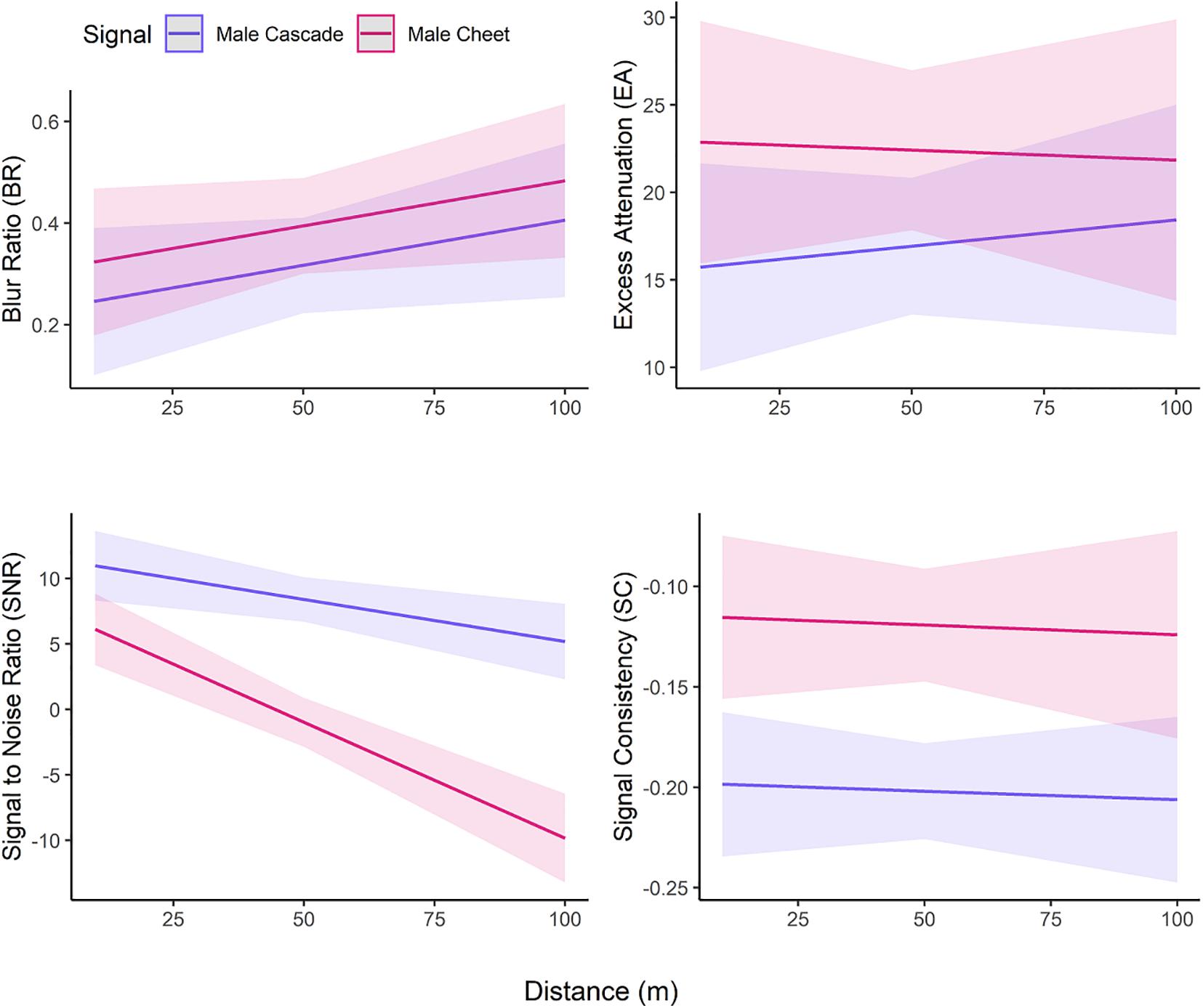
Figure 3. Modeled metrics of propagation quality and consistency for male canyon wren song cascades and “cheet” notes. Model includes 20 songs, with sex, distance, and song identity as fixed factors.
Supplementary male-female comparisons found that female songs propagated better than male “cheet” notes, with female songs having lower excess attenuation (F = 10.96, p < 0.001), and higher signal-to-noise ratios (F = 50.50, p < 0.0001) (Supplementary Table 1). Male “cheet” notes transmitted more consistently than female songs (F = 57.15, p < 0.0001).
Discussion
In this study, we demonstrated sex-based differences in bird song propagation, and we showed that different parts of a single bird song transmit as predicted by their functional and contextual use patterns. Our sex-based comparison found that male song cascades transmitted farther than female songs, supporting the hypothesis that these male signals have evolved for maximum broadcast efficacy. Additionally, female songs degraded faster with distance from the sound source. This result matches anecdotal observations in the field; male canyon wren song cascades are clearly distinct to the human ear over long distances, while female songs are harder to discern (Jones and Dieni, 2020). At the same time, female songs propagated farther and more clearly than male “cheet” notes, suggesting that female song has more broadcast function that those highly aggressive male notes. Given the similarities between human and avian sound detection thresholds, it is likely that birds have a similar perception of the effects of distance on male and female canyon wren song detectability (Dabelsteen et al., 1993; Martin, 2017; Zeyl et al., 2020).
Our results fit with hypothesized links between avian song structure, function, and transmission. The buzzy structure and aggressive function of female songs led us to predict that they would not propagate as far as the pure-toned male signals. Indeed, male canyon wren song cascades transmitted well over long distances, while female songs traveled less far. Interestingly, our two measures of song form degradation through space, blur ratio and excess attenuation, did not differ between male cascades and female songs. Thus, while male song cascades are detectable at longer distances, they do not retain structural clarity better than female songs. Previous work on a tropical wren species also found that signal-to-noise ratios varied with sex, but blur ratio did not, indicating that across species, female songs might consistently travel shorter distances than male broadcast song while retaining acoustic form (Mennill et al., 2009; Graham et al., 2017). These patterns make sense, given that canyon wren female songs are used almost exclusively in confrontations with female rivals, and are likely to be under similar selective regimes as other avian short-distance (quiet) songs adapted for use in contest situations (Morton, 1977; Akçay et al., 2015; Piza and Sandoval, 2016; Ręk and Kwiatkowska, 2016; Vargas-Castro et al., 2017). Like those short-distance songs, female canyon wren songs are expected to benefit if rapid degradation facilitates the transfer of aggressive intent to a rival but not to eavesdroppers.
Individual bird songs may have multiple component parts that function and evolve separately (Richards, 1981; Dabelsteen and Pedersen, 1988; Benedict and Bowie, 2012). This phenomenon is evident in male canyon wrens, where the cascade syllables used in all songs transmitted over long distances, while the “cheet” notes emphasized in aggressive contests degraded faster through space. Transmission differences were significantly reflected in signal-to-noise measurements and in the blurring of “cheet” notes. We conclude that “cheet” notes lose clarity at long distances, and become indistinguishable from background noise at distances of approximately 50 m (where SNR becomes negative in Figure 2). Multiple other studies have shown that different parts of a bird song can transmit with varying levels of clarity (Brenowitz, 1982; Dabelsteen and Pedersen, 1988; Graham et al., 2017; Grabarczyk and Gill, 2020). Such differences in transmission can reflect different functional roles for different song segments. In this case, the aggressive “cheet” notes indicate proximity, while cascades serve in long-distance communication. Because they contain these two elements with different transmission properties, male canyon wren songs are variable, and individuals can adjust song element use to maximize signal efficacy, given the context (Benedict et al., 2012). Female canyon wrens can also adjust song form by varying elements, but do not have a distinctly two-element song. This difference in song structure might be one reason why female song propagation distances fall between those of male cascades and male “cheet” notes.
In this study we report on a newly developed measure of acoustic transmission, signal consistency. We developed this measure because early work suggested that birds living in open spaces may face selective pressure to combat increased meteorological instability (i.e., wind, temperature) when compared to those living in closed spaces (Morton, 1975). Male canyon wren cascades and “cheet” notes propagated with more consistency than female songs, suggesting that male songs are more likely to be clearly detected at some point during signal transmission in the face of environmental fluctuations. Within male songs, however, cascades did not propagate with as much consistency as terminal “cheet” notes; this result ran counter to our predictions. We suspect that it may result from adaptive constraint, where cascades face tradeoffs between signal clarity and consistency for long-range use, while the temporally shorter, aggressive “cheet” notes may retain high consistency while remaining free of the requirement for clarity over distance. High “cheet” note transmission consistency may also reflect the fact that female songs and male cascades have relatively long durations, and more pronounced peak frequency variation than “cheet” notes. We suggest further investigation into the role of signal consistency in both short and long-range communication among animals living in open spaces.
Although we found support for our main predictions, close examination of the results (Table 1) reveals some study outcomes that were surprising. First, pair identity significantly predicted male and female song propagation for three metrics, with the songs of mated birds propagating similarly. This might result from the habitat, local recording conditions when we procured our song examples, or wind conditions during playback, but also raises the intriguing possibility that birds select mates based on similarity in signal transmission. We think that this speculation deserves further study. Second, male songs did not show the expected pattern of increased excess attenuation with increasing distance from the speaker. The lack of a pattern was apparently driven by high variability in the excess attenuation of male song, particularly the “cheet” notes. This variability and the inclusion of some different male song samples in the two comparisons (see section “Methods”) also apparently drove differences in modeled male cascade features between analyses (Figures 2, 3). Individual males, and even individual songs from one male, may differ substantially in their ability to reach distant audiences. We recommend follow up studies on individual variability and consistency in song transmission that include multiple songs and song types from each individual. These studies could also include measures of habitat and topography, as songs should transmit differently when certain individuals occupy habitats with different acoustic properties.
Our study demonstrated that the cascade portions of male canyon wren songs transmit farther than female songs and that male “cheet” notes have the shortest transmission distances. Thus, the typical broadcast song of this species has the best long-distance detectability, while two signals that are most often used in aggressive contests degrade more rapidly through space. These outcomes support a match between song form and function, and highlight sex differences in communication. Many of the songs of female birds in north temperate regions are similar in form to male songs of the same species (Hahn et al., 2013; Krieg and Getty, 2016; Rose et al., 2018). Canyon wrens provide a unique study system where male and female song forms have apparently evolved under different selective pressures and have different acoustic transmission properties. The sex-specific structures of these songs predict their acoustic propagation, highlighting the possibility that future within-species comparisons can use male and female signal structure to inform hypotheses about function and efficacy.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The animal study was reviewed and approved by the University of Northern Colorado IACUC 1606C-LB-Birds-19.
Author Contributions
LB and BH conceived the study. LB, BH, and LD conducted the field work. BH measured acoustic data and performed the statistical analyses. All authors assisted with writing, led by LB.
Funding
This work was supported by the Provost Research Dissemination and Faculty Development Grant from the University of Northern Colorado.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We are grateful to Alex Colpitts, Jake Lempka, and DeAnna Philpott for assistance in the field. We the Sylvan Dale Ranch and the Heart J Center for providing field access. We thank the members of the Benedict and McEntee labs for feedback on a draft of the manuscript. Two reviewers provided feedback that greatly improved the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.722967/full#supplementary-material
Footnotes
References
Akçay, Ç, Anderson, R. C., Nowicki, S., Beecher, M. D., and Searcy, W. A. (2015). Quiet threats: soft song as an aggressive signal in birds. Anim. Behav. 105, 267–274. doi: 10.1016/j.anbehav.2015.03.009
Apol, A., Sturdy, C. B., and Proppe, D. S. (2018). Seasonal variability in habitat structure may have shaped acoustic signals and repertoires in the black-capped and boreal chickadees. Evol. Ecol. 32, 57–74. doi: 10.1007/s10682-017-9921-4
Baker, M. C., and Logue, D. M. (2007). A comparison of three noise reduction procedures applied to bird vocal signals. J. Field Ornithol. 78, 240–253. doi: 10.1111/j.1557-9263.2007.00109
Benedict, L., and Bowie, R. C. K. (2012). Rattling cisticola song features and variability across sub-Saharan Africa. J. Zool. 287, 96–103. doi: 10.1111/j.1469-7998.2011.00890.x
Benedict, L., and Warning, N. (2017). Rock wrens preferentially use song types that improve long distance signal transmission during natural singing bouts. J. Avian Biol. 48, 1254–1262. doi: 10.1111/jav.01357
Benedict, L., Rose, A., and Warning, N. (2012). Canyon wrens alter their songs in response to territorial challenges. Anim. Behav. 84, 1463–1467. doi: 10.1016/j.anbehav.2012.09.017
Benedict, L., Rose, A., and Warning, N. (2013). Small song repertoires and high rate of song-type sharing among canyon wrens. Condor 115, 874–881. doi: 10.1525/cond.2013.130025
Blumenrath, S., and Dabelsteen, T. (2004). Degradation of great tit (Parus Major) song before and after foliation: implications for vocal communication in a deciduous forest. Behaviour 141, 935–958. doi: 10.1163/1568539042360152
Blumstein, D. T., and Récapet, C. (2009). The sound of arousal: the addition of novel non-linearities increases responsiveness in marmot alarm calls. Ethology 115, 1074–1081. doi: 10.1111/j.1439-0310.2009.01691.x
Boncoraglio, G., and Saino, N. (2007). Habitat structure and the evolution of bird song: a meta-analysis of the evidence for the acoustic adaptation hypothesis. Funct. Ecol. 21, 134–142. doi: 10.1111/j.1365-2435.2006.01207.x
Bradbury, J. W., and Vehrencamp, S. L. (2011). Principles of Animal Communication. Sunderland, MA: Sinauer Associates Incorporated.
Brenowitz, E. A. (1982). The active space of red-winged blackbird song. J. Comp. Physiol. 147, 511–522.
Brenowitz, E. A., Margoliash, D., and Nordeen, K. W. (1997). An introduction to birdsong and the avian song system. J. Neurobiol. 33, 495–500. doi: 10.1002/(sici)1097-4695(19971105)33:5<495::aid-neu1>3.0.co;2-#
Brumm, H., and Naguib, M. (2009). Environmental acoustics and the evolution of bird song. Adv. Study Behav. 40, 1–33. doi: 10.1016/s0065-3454(09)40001-9
Catchpole, C. K., and Slater, P. J. B. (2008). Bird Song. Cambridge: Cambridge University Press, doi: 10.1017/cbo9780511754791
Dabelsteen, T., and Pedersen, S. B. (1988). Song parts adapted to function both at long and short ranges may communicate information about the species to female blackbirds turdus merula. Ornis Scand. 19:195. doi: 10.2307/3676558
Dabelsteen, T., Larsen, O. N., and Pedersen, S. B. (1993). Habitat-induced degradation of sound signals: quantifying the effects of communication sounds and bird location on blur ratio, excess attenuation, and signal-to-noise ratio in blackbird song. J. Acoust. Soc. Am. 93, 2206–2220. doi: 10.1121/1.406682
Dabelsteen, T., McGregor, P. K., Lampe, H. M., Langmore, N. E., and Holland, J. O. (1998). Quiet song in song birds: an overlooked phenomenon. Bioacoustics 9, 89–105. doi: 10.1080/09524622.1998.9753385
Dargis, L., Benedict, L., and Najar, N. (2021). Female bird song rates do not covary with population density in a North American species. Ethology. doi: 10.1111/eth.13227
Endler, J. A. (1993). Some general comments on the evolution and design of animal communication systems. Philos. Trans. R. Soc. Lond. B Biol. Sci. 340, 215–225. doi: 10.1098/rstb.1993.0060
Ey, E., and Fischer, J. (2009). The “Acoustic Adaptation” hypothesis – a review of the evidence from birds, anurans, and mammals. Bioacoustics 19, 21–48. doi: 10.1080/09524622.2009.9753613
Forrest, T. G. (1994). From sender to receiver: propagation and environmental effects on acoustic signals. Am. Zool. 34, 644–654. doi: 10.1093/icb/34.6.644
Gil, D., and Gahr, M. (2002). The honesty of bird song: multiple constraints for multiple traits. Trends Ecol. Evol. 17, 133–141. doi: 10.1016/s0169-5347(02)02410-2
Grabarczyk, E. E., and Gill, S. A. (2020). Anthropogenic noise masking diminishes house wren (Troglodytes aedon) song transmission in urban natural areas. Bioacoustics 29, 518–532.
Graham, B. A., Sandoval, L., Dabelsteen, T., and Mennill, D. J. (2017). A test of the acoustic adaptation hypothesis in three types of tropical forest: degradation of male and female rufous-and-white wren songs. Bioacoustics 26, 37–61. doi: 10.1080/09524622.2016.1181574
Greig, E. I., Jordan Price, J., and Pruett-Jones, S. (2013). Song evolution in maluridae: influences of natural and sexual selection on acoustic structure. EMU Austral Ornithol. 113, 270–281. doi: 10.1071/mu12078
Hahn, A. H., Krysler, A., and Sturdy, C. B. (2013). Female song in black-capped chickadees (Poecile atricapillus): acoustic song features that contain individual identity information and sex differences. Behav. Processes 98, 98–105. doi: 10.1016/j.beproc.2013.05.006
Hall, M. L., and Langmore, N. E. (2017). Editorial: fitness costs and benefits of female song. Front. Ecol. Evol. 5:48. doi: 10.3389/fevo.2017.00048
Hardt, B., and Benedict, L. (2020). Can you hear me now? A review of signal transmission and experimental evidence for the acoustic adaptation hypothesis. Bioacoustics 30, 716–742. doi: 10.1080/09524622.2020.1858448
Hathcock, T. J., and Benedict, L. (2018). Conspecific challenges provoke female canyon wrens to sing but not to duet. Behav. Ecol. Sociobiol. 72:196. doi: 10.1007/s00265-018-2625-0
Higham, J. P., and Hebets, E. A. (2013). An introduction to multimodal communication. Behav. Ecol. Sociobiol. 67, 1381–1388. doi: 10.1007/s00265-013-1590-x
Hof, D., and Hazlett, N. (2010). Low-amplitude song predicts attack in a North American wood warbler. Anim. Behav. 80, 821–828. doi: 10.1016/j.anbehav.2010.07.017
Jones, S. L., and Dieni, J. S. (2020). “Canyon wren (Catherpes mexicanus),” in Birds of the World, eds A. F. Poole and F. B. Gill (Ithaca, NY: Cornell Lab of Ornithology), doi: 10.2173/bow.canwre.01
Krieg, C. A., and Getty, T. (2016). Not just for males: females use song against male and female rivals in a temperate zone songbird. Anim. Behav. 113, 39–47. doi: 10.1016/j.anbehav.2015.12.019
Langmore, N. E. (1998). Functions of duet and solo songs of female birds. Trends Ecol. Evol. 13, 136–140.
Mennill, D. J., and Vehrencamp, S. L. (2005). Sex differences in singing and duetting behavior of neotropical rufous-and-white wrens (Thryothorus rufalbus). Auk 122:175. doi: 10.1642/0004-80382005122
Mennill, D., Dabelsteen, T., and Barker, N. (2009). Degradation of male and female rufous-and-white wren songs in a tropical forest: effects of sex, perch height, and habitat. Behaviour 146, 1093–1122. doi: 10.1163/156853909x406446
Mikula, P., Valcu, M., Brumm, H., Bulla, M., Forstmeier, W., Petrusková, T., et al. (2020). A global analysis of song frequency in passerines provides no support for the acoustic adaptation hypothesis but suggests a role for sexual selection. Ecol. Lett. 24, 477–486. doi: 10.1111/ele.13662
Morton, E. S. (1975). Ecological sources s of selection on avian sounds. Am. Nat. 109, 17–34. doi: 10.1086/282971
Morton, E. S. (1977). On the occurrence and significance of motivation-structural rules in some bird and mammal sounds. Am. Nat. 111, 855–869. doi: 10.1086/283219
Mouterde, S. C., Theunissen, F. E., Elie, J. E., Vignal, C., and Mathevon, N. (2014). Acoustic communication and sound degradation: how do the individual signatures of male and female zebra finch calls transmit over distance? PLoS One 9:e102842. doi: 10.1371/journal.pone.0102842
Nemeth, E., and Brumm, H. (2010). Birds and anthropogenic noise: are urban songs adaptive? Am. Nat. 176, 465–475. doi: 10.1086/656275
Niederhauser, J. M., DuBois, A. L., Searcy, W. A., Nowicki, S., and Anderson, R. C. (2018). A test of the eavesdropping avoidance hypothesis as an explanation for the structure of low-amplitude aggressive signals in the song sparrow. Behav. Ecol. Sociobiol. 72:47. doi: 10.1007/s00265-018-2469-7
Odom, K. J., and Benedict, L. (2018). A call to document female bird songs: applications for diverse fields. Auk 135, 314–325. doi: 10.1642/auk-17-183.1
Odom, K. J., Hall, M. L., Riebel, K., Omland, K. E., and Langmore, N. E. (2014). Female song is widespread and ancestral in songbirds. Nat. Commun. :3379. doi: 10.1038/ncomms4379
Piza, P., and Sandoval, L. (2016). The differences in transmission properties of two bird calls show relation to their specific functions. J. Acoust. Soc. Am. 140:4271. doi: 10.1121/1.4971418
Podos, J., Huber, S. K., and Taft, B. (2004). Bird song: the interface of evolution and mechanism. Annu. Rev. Ecol. Evol. Syst. 35, 55–87. doi: 10.1146/annurev.ecolsys.35.021103.105719
Price, J. J., Lanyon, S. M., and Omland, K. E. (2009). Losses of female song with changes from tropical to temperate breeding in the New World blackbirds. Proc. Biol. Sci. 276, 1971–1980. doi: 10.1098/rspb.2008.1626
Rek, P. (2013). Soft calls and broadcast calls in the corncrake as adaptations to short and long range communication. Behav. Processes 99, 121–129. doi: 10.1016/j.beproc.2013.07.009
Ręk, P., and Kwiatkowska, K. (2016). Habitat complexity and the structure of vocalizations: a test of the acoustic adaptation hypothesis in three rail species (Rallidae). IBIS 158, 416–427. doi: 10.1111/ibi.12357
Richards, D. G. (1981). Alerting and message components in songs of rufous-sided towhees. Behaviour 76, 223–249.
Riebel, K., Odom, K. J., Langmore, N. E., and Hall, M. L. (2019). New insights from female bird song: towards an integrated approach to studying male and female communication roles. Biol. Lett. 15, 20190059. doi: 10.1098/rsbl.2019.0059
Rose, A. (2013). Changes in canyon wren vocalizations in advance of the breeding season. Ursidae 2:2.
Rose, E. M., Mathew, T., Coss, D. A., Lohr, B., and Omland, K. E. (2018). A new statistical method to test equivalence: an application in male and female eastern bluebird song. Anim. Behav. 145, 77–85. doi: 10.1016/j.anbehav.2018.09.004
Sandoval, L., Dabelsteen, T., and Mennill, D. J. (2015). Transmission characteristics of solo songs and duets in a neotropical thicket habitat specialist bird. Bioacoustics 24, 289–306. doi: 10.1080/09524622.2015.1076346
Slabbekoorn, H., and Smith, T. B. (2002). Bird song, ecology and speciation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 357, 493–503. doi: 10.1098/rstb.2001.1056
Slater, P. J. B., and Mann, N. I. (2004). Why do the females of many bird species sing in the tropics? J. Avian Biol. 35, 289–294. doi: 10.1111/j.0908-8857.2004.03392.x
Spencer, A. (2012). Female song in canyon wren, with notes on vocal repertoire. Colorado Birds 46, 268–276.
Thorpe, W. H. (1961). Bird-song: The Biology of Vocal Communication and Expression in Birds. Whitefish, MT: Literary Licensing, LLC.
Vargas-Castro, L. E., Sandoval, L., and Searcy, W. A. (2017). Eavesdropping avoidance and sound propagation: the acoustic structure of soft song. Anim. Behav. 134, 113–121. doi: 10.1016/j.anbehav.2017.10.008
Warning, N., Covy, N., Rose, A., Phan, X. M., and Benedict, L. (2015). Canyon wren territory occupancy and site attributes in northern colorado. Am. Midl. Nat. 174, 150–160. doi: 10.1674/0003-0031-174.1.150
West-Eberhard, M. J. (1983). Sexual selection, social competition, and speciation. Q. Rev. Biol. 58, 155–183. doi: 10.1086/413215
Wiley, R. H., and Richards, D. G. (1978). Physical constraints on acoustic communication in the atmosphere: implications for the evolution of animal vocalizations. Behav. Ecol. Sociobiol. 3, 69–94. doi: 10.1007/bf00300047
Wiley, R. H., and Richards, D. G. (1982). “Adaptations for acoustic communication in birds: sound transmission and signal detection,” in Acoustic Communication in Birds, Vol. 1, eds D. E. Kroodsma and E. H. Miller (New York, NY: Academic Press), 131–181. doi: 10.1016/b978-0-08-092416-8.50014-0
Zeyl, J. N., den Ouden, O., Köppl, C., Assink, J., Christensen-Dalsgaard, J., Patrick, S. C., et al. (2020). Infrasonic hearing in birds: a review of audiometry and hypothesized structure-function relationships. Biol. Rev. Camb. Philos. Soc. 95, 1036–1054. doi: 10.1111/brv.12596
Keywords: animal communication, acoustic propagation, signal transmission, signal adaptation, bird song, female song, Catherpes mexicanus
Citation: Benedict L, Hardt B and Dargis L (2021) Form and Function Predict Acoustic Transmission Properties of the Songs of Male and Female Canyon Wrens. Front. Ecol. Evol. 9:722967. doi: 10.3389/fevo.2021.722967
Received: 09 June 2021; Accepted: 19 October 2021;
Published: 10 November 2021.
Edited by:
Ann Valerie Hedrick, University of California, Davis, United StatesReviewed by:
Erin E. Grabarczyk, United States Department of Agriculture (USDA), United StatesJ. Roberto Sosa-Lopez, Instituto Politécnico Nacional (IPN), Mexico
Copyright © 2021 Benedict, Hardt and Dargis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lauryn Benedict, bGF1cnluLmJlbmVkaWN0QHVuY28uZWR1