
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
SYSTEMATIC REVIEW article
Front. Ecol. Evol., 25 October 2021
Sec. Conservation and Restoration Ecology
Volume 9 - 2021 | https://doi.org/10.3389/fevo.2021.703220
This article is part of the Research TopicGrassland Conservation in Asia: Sustainability Under Climate ChangeView all 28 articles
 Oyunbileg Munkhzul1,2
Oyunbileg Munkhzul1,2 Khurelpurev Oyundelger3,4
Khurelpurev Oyundelger3,4 Naidan Narantuya1,5
Naidan Narantuya1,5 Indree Tuvshintogtokh1Batlai Oyuntsetseg2
Indree Tuvshintogtokh1Batlai Oyuntsetseg2 Karsten Wesche3,4,6*
Karsten Wesche3,4,6* Yun Jäschke4
Yun Jäschke4The Mongolian steppes with a long history of nomadic pastoralism cover a large area of the Palaearctic steppe biome and are still relatively intact. As livestock number has increased over the last two decades, grazing has been considered as the main reason of pasture degradation. However, the impact of grazing on vegetation dynamics, and its interaction with climate, is still not clear. We reviewed 44 publications in Mongolian language, covering 109 sites in five main steppe types, i.e., desert, dry, meadow, mountain, and high mountain steppe, with a mean annual precipitation and temperature range from 120 to 370 mm and from −6 to +5°C, respectively. We calculated relative changes in vegetation cover, species richness, and aboveground biomass from heavily grazed with respect to lightly/non-grazed conditions. Multiple linear regression models were used to test the impact of environmental factors, i.e., mean annual precipitation, coefficient of variation for precipitation, mean annual temperature and elevation. Grazing had a stronger effect on the vegetation of dry, desert and high mountain steppes, whereas its effect was less pronounced in the meadow and mountain steppes with mesic climate and high productivity. Vegetation cover, species richness and aboveground biomass were reduced by heavy grazing in the dry, desert and high mountain steppes. In the meadow steppes, grazing reduced vegetation cover, but increased richness and had nearly no effect on biomass. In the mountain steppe, richness and cover were not affected, but biomass was reduced by heavy grazing. Additionally, grazing effects on biomass tended to be more pronounced at sites with higher amounts of annual precipitation, and effects on cover changed from negative to positive as elevation increased. In conclusion, grazing effects in Mongolian steppes are overall negative in desert, dry and high mountain steppes, but no or even positive effects are found in meadow and mountain steppes. Especially, heavy grazing showed a detrimental effect on all vegetation variables in desert steppes, indicating the existence of combined pressure of climate and grazing in arid habitats, making them potentially sensitive to overgrazing and climate change. Grassland conservation and management should consider characteristics of different steppe types and give importance to local environmental conditions.
Grasslands cover 41% of the World's terrestrial surface (White et al., 2000), which sustain livelihoods for nearly 800 million people, and are a crucial source of livestock forage and habitat for diverse wildlife (White et al., 2000). Most of the world's grasslands are found in temperate regions, and these temperate grasslands have the distinction of being the most altered terrestrial ecosystems on the planet (Henwood, 2010) and the most threatened globally. Of these temperate grasslands, 19.6% (10.3 million km2) belong to the Palaearctic steppe biome, which forms a vast belt across the mid-latitudes of Eurasia, with nearly 1.3 million km2 occurring in Mongolia (Henwood, 2010; Wesche et al., 2016). The Mongolian grasslands are still relatively intact, representing the world's largest grasslands that are still healthy in terms of biodiversity and traditional land use (Batsaikhan et al., 2014). Hence in the last two decades, Mongolia has become a key region for studying the effects of land use to devise potentially sustainable land use strategies.
Mongolia has a long tradition of nomadic pastoralism, which has been the main agricultural sector in Mongolia for centuries. Approximately 83% of the territory (1.3 million km2) is rangeland (including grasslands, shrublands, forest steppe, and deserts that are grazed by domestic livestock; Angerer et al., 2008), supporting 71 million heads [118.7 million Sheep Units (SU)1; National Statistics Office of Mongolia, 2021]. Until 1990, the number of livestock was 25.8 million heads (54.3 million SU), however, after institutional and societal changes in the 90s, decentralization began and herders were allowed to privatize their herds. Since then, the number of livestock has increased dramatically, especially the number of sheep and goats due to the high market value of cashmere. Livestock density has increased more than three-fold, i.e., from 32 SU per 100 ha in 1961 to 99 SU in 2017 (National Statistics Office of Mongolia, 2018). According to a national report on the rangeland health of Mongolia, 58% of Mongolian rangeland areas are estimated to be degraded, of which 14, 21, 13, and 10% are slightly, moderately, heavily and fully degraded, respectively, based on field monitoring data (Densambuu et al., 2018).
Several global quantitative reviews (Milchunas and Lauenroth, 1993; Cingolani et al., 2005) revealed that grazing effects on vegetation interact with productivity. The latter in turn is controlled by abiotic factors, such as climate (precipitation, temperature) and topographic conditions. Mongolian grasslands often experience very high intra- and interannual rainfall variability, and growing season precipitation has the greatest effect on plant productivity (Liang et al., 2002). In wet regions with high productivity, moderate grazing can reduce the dominance of competitively superior species, thus increase plant diversity; while in dry grasslands with relatively low productivity, grazing effects tend to be neutral or moderately negative, with interannual variation of mean annual precipitation having a stronger effect than grazing on plant communities (Olff and Ritchie, 1998). Similar results were also found in Mongolian dry steppes, where grazing effects were smaller in drought years (Bat-Oyun et al., 2016). However, negative grazing impact on vegetation has been reported both in the forest-steppe in northern Mongolia (Takatsuki et al., 2018) and in the desert-steppe in northern China (Deng et al., 2013; Zhang et al., 2018). Contradictory results suggest that Mongolian grasslands are a mixture of equilibrium and non-equilibrium systems (Fernandez-Gimenez and Allen-Diaz, 1999; Zemmrich et al., 2010; von Wehrden et al., 2012; Ahlborn et al., 2020). In areas where mean annual precipitation (MAP) is <250 mm and the coefficient of variation of annual precipitation (cvP) is >33% (Ellis and Chuluun, 1993), the low and extremely variable annual precipitation has an overriding impact on livestock numbers, while in regions with higher and relatively constant precipitation, grazing should play a greater role in determining plant community structure and composition (Ellis and Swift, 1988). Furthermore, Mongolia has a typical continental climate with enormous daily and annual temperature fluctuations, and extreme climate events such as drought and cold harsh winter (dzud) also have huge effects on vegetation, animal husbandry, and thus economic situation of households. Local topographical conditions, for example location along a slope, might also interact with grazing effects, because soil moisture might decrease from valley bottom to upper slope (Fujita et al., 2009). Slope direction also affects nutrient availability and plant growth form, therefore interacts with grazing effects (Lkhagva et al., 2013).
Considering such substantial effects of grazing and environment on vegetation in Mongolian grasslands, dozens of studies have been conducted. However, publications on the effects of grazing on vegetation and its interaction with the local climate provide partly contradictory results, and thus, no consensus has arrived in the international literature. Hence, the current study aims to perform the first review of publications in Mongolian language, which reported grazing effects on the country's steppe vegetation from 1966 onwards. These publications provide valuable information, but are unfortunately not accessible without knowledge of the language, and many were not published in widely distributed outlets. Based on a thorough review of these publications, we addressed the following hypotheses:
• Heavy grazing by livestock negatively affects vegetation cover, species richness, and aboveground biomass. Species richness and aboveground biomass are supposed to be highest under moderate grazing intensity compared with light and heavy.
• The magnitude of the grazing effects differs among steppe types due to their specific environmental conditions. Grazing effects in meadow steppe and mountain steppe tend to be stronger, in line with classical equilibrium rangeland theory, as they receive usually more summer rainfall and the productivity of such grasslands is high. In contrast, size of grazing effects on desert steppe and dry steppe is expected to be smaller as a consequence of the overriding impact of precipitation patterns as implied by non-equilibrium dynamics.
Mongolia is a landlocked country of Central Asia, situated between 41°35′ and 52°08′ N, 87°44′-119°55′ E, far from any ocean. Its land area comprises 1.564 Mio. km2 (National Statistics Office of Mongolia, 2019), which is roughly equivalent to all countries of western and central Europe. Mongolia experiences an extreme continental climate characterized by long cold winters, short summers, and most precipitation falls during mid-June to August (Liu et al., 2013b). The climate is further characterized by high differentiation among four seasons and high diurnal temperature variation. Mongolia can be classified into six ecological zones (Figure 1), i.e., the high mountain, taiga, forest steppe, steppe, desert steppe and desert zones (Tuvshintogtokh, 2015). Among them, the forest steppe, steppe, desert steppe and parts of the desert zone are primarily rangelands. From the north to the south of Mongolia, the climate becomes drier (temperature increases and precipitation decreases).
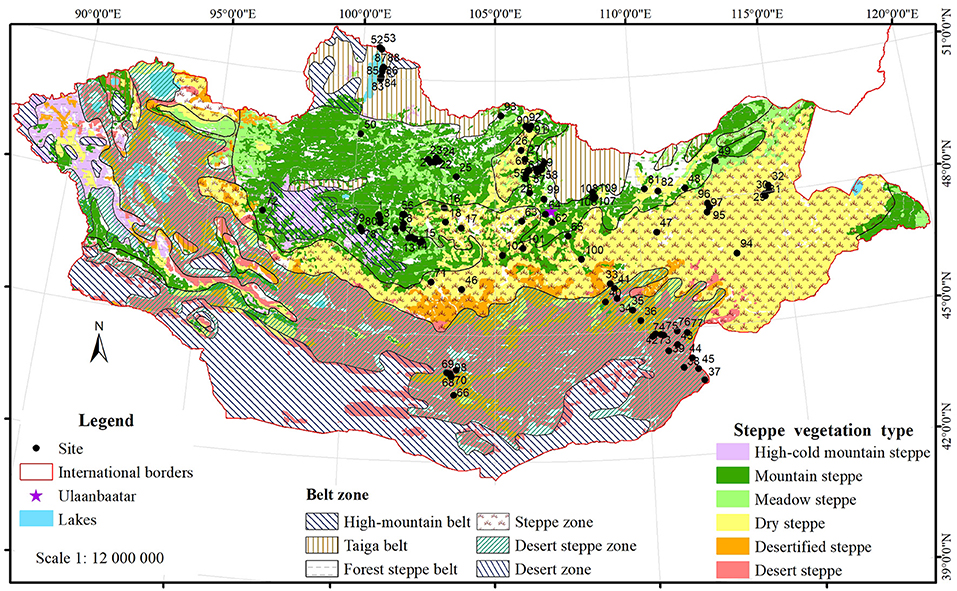
Figure 1. Ecological zones and steppe vegetation types of Mongolia, with 109 study sites extracted from the reviewed publications. Ecological belt zones are based on the adapted vegetation map of the People's Republic of Mongolia (Yunatov and Dashnyam, 1979; Ulziikhutag, 1985). Steppe vegetation types follow the Steppe Vegetation of Mongolia (Tuvshintogtokh, 2015), and the “dry steppe” here includes “typical steppe,” as it is commonly done in literature. The high-cold mountain steppe is abbreviated as the high mountain steppe in the following. The sites located in the “desertified steppe” are grouped into the “desert steppe” in the analyses. The site ID is the same as in Supplementary Table A1.
Between 1944 and 2014, mean annual temperature (MAT) has increased by 2.1°C on average, with the increase being stronger in winter (3.6°C) than in summer (1.8°C). In particular, MAT increased by 2.0–3.7°C in the forest steppe zone, 0.7–2.0°C in the steppe and desert zones (these two zones were considered together in the analysis; Dagvadorj et al., 2010; MARCC., 2014). As a result, the aridity index and the number of extreme warm days increased significantly (MARCC., 2014). The change in mean annual precipitation (MAP) varied strongly in both spatial and temporal perspective. For instance, MAP decreased by 8.7–12.5% in the central and southern regions and increased by 3.5–9.3% in the eastern and western regions of Mongolia (MARCC., 2014).
The total number of livestock in Mongolia has increased 2.7 times since 1990, when the democratic revolution took place and livestock was privatized. The magnitude of increment, however, differs among ecological zones: the total number increased 2.0 times in the high mountain belt, 3.2 times in the forest steppe and steppe zone, 2.5 times in the desert steppe zone, and 2.6 times in the desert zone (National Statistics Office of Mongolia, 2019). As a result, the steppe zone sustains the highest number of livestock, followed by the forest steppe and the desert steppe zone (Figure 2). The significant drops in livestock numbers in 2000–2002 and 2010 show the effects of dzud, i.e., severe winter, which caused the death of 28.5 and 17.6 million SU livestock, respectively, after the winters of 1999 and 2009. Still, and in contrast to the non-equilibrium theory, over the last decade, livestock units have shown a general increase, even in the dry and highly variable desert steppes.
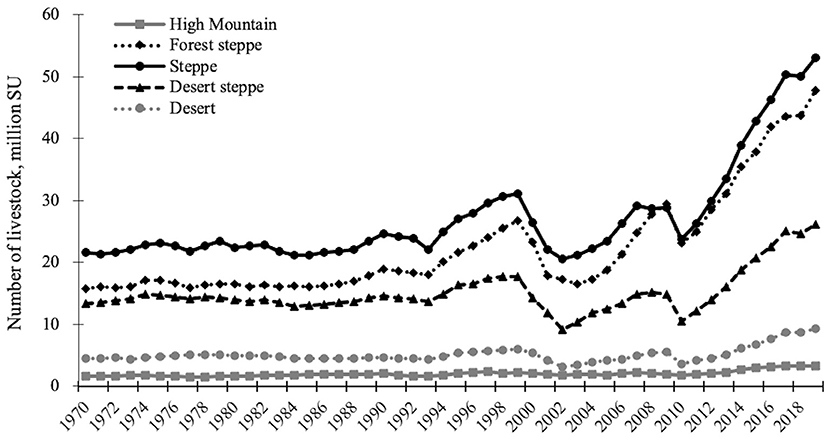
Figure 2. Change in livestock numbers in different ecological zones of Mongolia. The total number of livestock in a particular zone was calculated from data at the soum (small administrative unit within a province) level (330 soums and 9 districts of the capital city, in total, N = 339). A total of 216 soums had only one dominant ecological zone, so the livestock number in this zone is equal to the soum level. When a soum consists of more than one ecological zone, then each of the two (N = 111) or three (N = 12) most dominant zones were considered to have the same number at soum level, because livestock mobility is high even within a single soum. Therefore, there is an overestimation of total livestock numbers (the sum of sheep units from ecological zones are 11–20% higher than those from soums for the respective year range). Data at the soum level were provided by the National Statistics Office of Mongolia, 1970–2019. The Taiga zone is not included because this zone is a boreal forest, where hardly any pasture exists.
Considering that our focus was on overcoming language barriers and enhancing the international knowledge base, we searched for all available Mongolian publications on grazing effects on the rangeland vegetation in specialized databases, such as local libraries of the Mongolian Academy of Science, National University of Mongolia, and Mongolian University of Life Science (see detailed list from Supplementary Table A1). As search criteria, we used the Mongolian terms “urgamaljilt,” “belcheer,” “belcheerlelt,” “belcheerleltiin noloo,” and “hashsan talbai” which translate to “vegetation,” “rangeland,” “pastureland,” “grassland,” “grazing,” “grazing effect,” and “fenced area.” In particular, for literature, books and (conference) proceedings that were published before 1990, we first searched for the title, and checked the context and included the suitable studies. The main vegetation indicators we consider here are species richness (SR), vegetation cover (VC) and aboveground biomass (AGB) as they are the most frequently reported quantitative measurements of rangeland conditions. We included studies published between 1966 and 2019. In total, 44 publications were found (Supplementary Material 1), i.e., one dissertation, six master theses, five book sections, and 32 journal articles that together reported data from 109 sites (Supplementary Table A2). Among them, conditions of different grazing intensity levels (i.e., light, moderate and heavy grazing, indicated by the distance to river or that to winter/summer camp in the original publications) could be compared for 51 sites, by dividing the values of the response variables under heavy grazing to the same variables under light (or zero) grazing at the same site:
relative grazing effect of heavy grazing = (heavy–light)/light grazing, and similarly, relative grazing effect of moderate grazing = (moderate–light)/light grazing.
The dominant grazers in different steppe types are provided in Table 1. Sheep and goats mainly graze on all steppe types, which constitutes ca. 85% (ca. 58 million heads) of the total livestock of Mongolia (National Statistics Office of Mongolia, 2021). Hence, the livestock composition is largely similar in all steppe types. This way, trends are comparable between main rangeland types, in spite of differences in absolute values for all indicators being much lower in the desert steppes and high-mountain steppes. Multiple-year (≥2 years) monitoring data was available from another 32 sites, but grazing intensity was only mentioned at four sites, which had fences to exclude livestock grazing, thus we made a summary of fence effects on those sites only. We did not analyze the studies which reported either the results of 1 year at a single grazing level, or studies on seasonal vegetation dynamics within 1 study year. The workflow chart of the data extraction is summarized in Supplementary Figure A1.
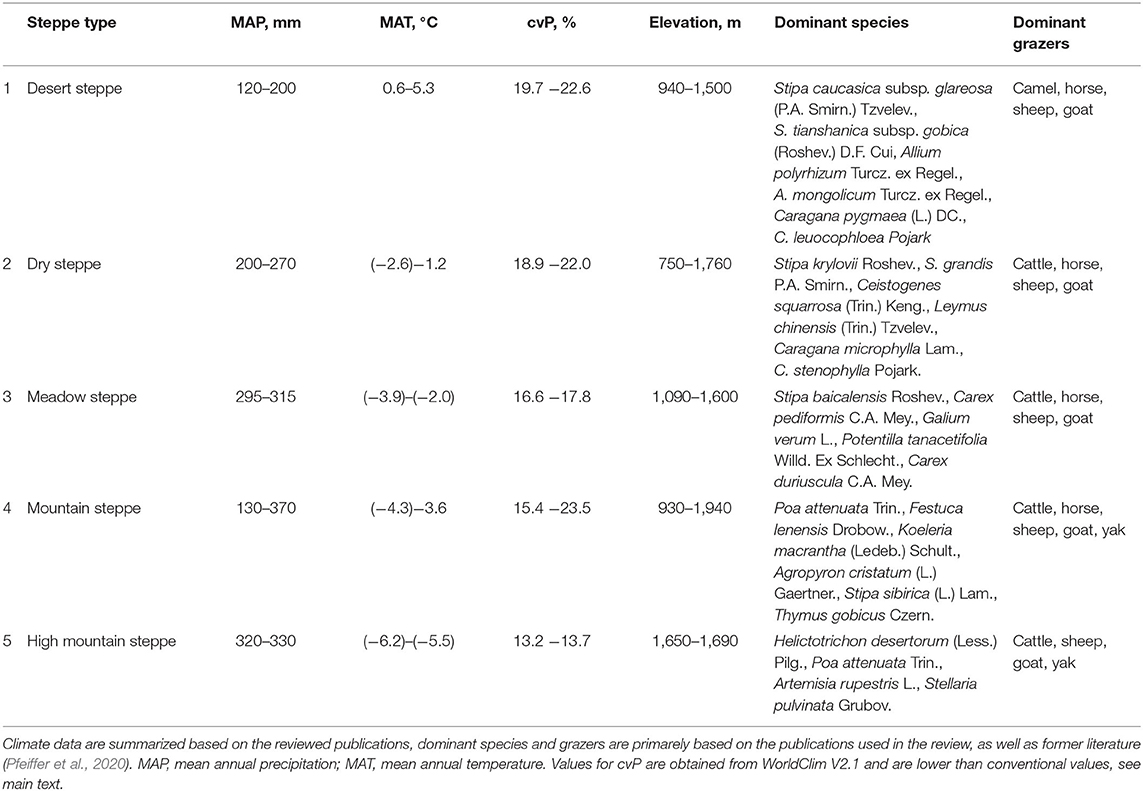
Table 1. Main environmental characteristics, dominant plant species and dominant grazers for five main steppe types in Mongolia.
We extracted quantitative values either directly from tables, or through the UN-SCAN-IT graph digitizer software (Silk Scientific, Inc.) from figures for the main response variables in the original publications. However, depending on the study design and purpose, surveying methods were inconsistent among studies. In particular, authors surveyed SR in different sizes of plots, which were usually large plots of 10 × 10 m2 (3–10 replicates), or occasionally small plots of 0.5 × 0.5 m2 or 1 × 1 m2. Some were sampled along a 50 or 100 m transect (3–5 replicates) and recorded the total species number in quadrats of 0.5 × 0.5 m2 or 1 × 1 m2 along the transect at an interval of 10 or 20 m for the 50 and 100 m transect, respectively. Moreover, whether estimates were based on the sum or the mean species number from subplots was not always clear. Reported VC was usually estimated from the same sample areas as for SR, and we calculated mean values of the repeated measures. AGB was estimated based on clipped standing biomass from 0.5 × 0.5 m2 to 1 × 1 m2 (with 3–10 replicates), and mostly from grazed sites rather than from grazing-exclusion plots; thereby providing only the residual biomass instead of primary productivity. When authors reported AGB values over the whole growing season, we selected the value of the peak biomass (late July or early August). Most biomass data were in units of centner/ha, so we recalculated them to g/m2.
Latitude, longitude, and elevation were usually directly reported in the publications. Else, we used Google Earth to estimate elevation based on the specific coordinates. Climate data was not always reported by authors, and even when it was, the timescale for the reported data was often missing. As the reviewed publication ranges between 1966 and 2000s, we extracted the available long-term climate data from Worldclim 2.1, i.e., monthly precipitation data from 1961 to 2018 and MAT for time range 1970–2000 (Harris et al., 2014; Fick and Hijmans, 2017). Data are compared with the data obtained from the National Agency for Meteorological and Environmental Monitoring (https://namem.gov.mn/eng/) for quality control. Annual total precipitation from Worldclim 2.1 data are largely realistic compared to station data and are much better than those from Chelsa V2 for our reviewed sites especially for recent years. Although the calculated cvP-values based on Worldclim 2.1 for 40 years seem to underestimate cvP-values in this region (Supplementary Material 2), the trend that cvP-values are higher in desert steppes than other steppe types still holds (Supplementary Figure A2), thus we opted Worldclim 2.1 climate data for further analyses.
Most of the publications reported the vegetation types for their study sites, which were typically five steppe types, i.e., desert steppe, dry or typical steppe, meadow steppe, mountain steppe, and high mountain steppe. Typical steppes mostly occur in eastern Mongolia and are dominated by tall feather grasses, while dry steppes, in a strict sense, occur in western and central Mongolia, as well as in the south of the typical steppe. Dry steppes are drier and regularly associated with shorter feather grassess and shrub species (Tuvshintogtokh, 2015; Pfeiffer et al., 2020). However, here we combine both steppe types into one, because the climatic and vegetation differences are relatively small compared to the differences among other steppe types. Furthermore, in earlier times, researchers often did not differentiate between these two steppe types and even mixed the two terms (Tuvshintogtokh, 2015). The main environmental characteristics and dominant plant species and dominant livestock types for five steppe types are summarized in Table 1, and the differences in the major environmental variables are illustrated in Supplementary Figure A2.
All measured response variables from the same location (i.e., with the same coordinates) were combined into one dataset. For some studies which were conducted at the same location and focused on the same response variables, but either (1) covering different study years, or (2) by different research groups, we calculated the mean to put them into one dataset.
We first calculated means and 95% confidence intervals from absolute values of each vegetation indicator at different grazing intensity levels for each steppe type and did not exclude outliers. Site-based paired t-tests were not performed due to the small number of samples in some cases. We also did not perform a strict formal meta-analysis, because most studies did not report sd/se values. We tested the impact of environmental factors (scaled to zero mean/unit variants) on relative grazing effects using multiple linear regression models. We only tested the main effects of environmental factors (i.e., MAP, cvP, MAT, and elevation) and did not include interaction terms because multiple environmental-grazing interactions are difficult to interpret due to the lack of specific ecological hypotheses. We fitted simple models without random factor, because only one value for each site was input for the model. We started with the most complex full model, including all environmental factors, and obtained estimates of effects for each. Then we simplified the initial full model in a stepwise procedure by progressively removing the least significant factor until we arrived at a null model. We compared the initial model with all subsequently reduced models in an ANOVA framework by comparing explained variance. Based on ANOVA results, we selected the final model in which all the remaining predictors were significant, and we reported the estimates for coefficients and standard errors for these predictors. For one of the studies on multiple year fence data, a two-way repeated measure ANOVA was performed to test the main effects of fencing and year on vegetation indicators with repeated subsites under different grazing regimes when such data were available. The significance level for all tests was p < 0.05 and all statistical analyses were conducted in R 3.1.0 (R Core Team, 2020). R packages “rstatix” (Kassambara, 2020a), “ggplot2” (Wickham, 2016), “tidyverse” (Wickham et al., 2019), “ggpubr” (Kassambara, 2020b), and “cowplot” (Wilke, 2020) were used.
Our review of Mongolian publications between 1966 and 2019 allowed us to distinguish two major stages of research in Mongolia. The earliest investigations were conducted from 1970 to 1990, as a part of the Mongolian and Russian Joint Biological Expedition (MRJBE). During this expedition period, botanists aimed to study different vegetation types in various ecological zones, and thus established different monitoring stations in (i) mountain and forest steppe (1970-1985 at Tuvshruulekh soum in Arkhangai province), (ii) meadow steppe (1976–1990 at Shaamar soum in Selenge province), (iii) flooded meadow steppe (between 1987 and 1990 and since 2009 at Mungunmorit soum in Tuv province), (iv) typical steppe (from 1982 to present at Tumentsogt soum in Sukhbaatar province), (v) dry steppe (1973–1976 at Bayan-Unjuul soum in Tuv province), (vi) desert steppe (1970–1976 at Bulgan soum in Umnugovi province), and (vii) desert (1977–1990 at Ekhiin gol soum in Bayankhongor province). Studies from this period provided valuable qualitative information on plant community composition and flora. The herbarium collection also began with MRJBE and other expeditions. Moreover, some essential data on SR, VC, AGB, phenology and seasonal growth dynamics etc., have been collected not only at monitoring stations, but also during field trips across the whole range of steppes.
The second stage includes vegetation studies by Mongolian and international researchers in 2000–2019. During this period, researchers have focused on quantitative surveys based on different vegetation sampling methods (e.g., line-point intercept). Both study stages provided essential information about Mongolian vegetation. Although not all vegetation types were equally represented, the most commonly distributed steppe communities have been investigated.
Based on data from 51 sites, where three grazing intensity levels were studied, main indicators were compared among light, moderate and heavy grazing conditions from five steppe types (Table 2). In general, lightly grazed sites had higher VC in desert steppe and meadow steppe than heavily grazed sites, and AGB was higher under light grazing compared to heavy grazing across all steppe types, except in meadow steppe. Furthermore, changes in SR were generally not significant except in the desert and high mountain steppe. However, the absolute values of SR might show artifacts due to different sizes of sampling plots among studies as described in the method section, while comparisons within steppe types remain valid.
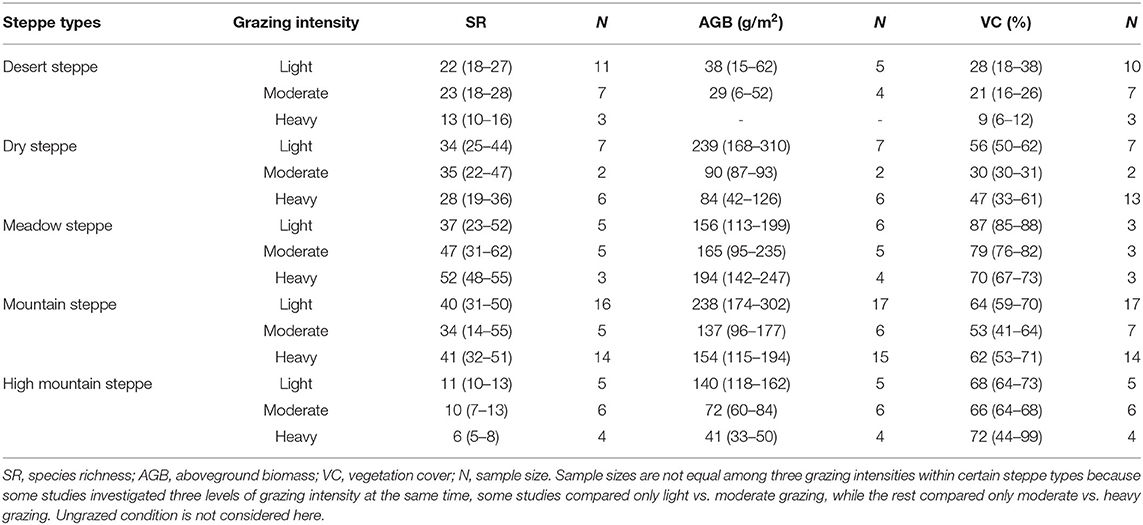
Table 2. Values (mean and 95% confidential interval) for indicators under different grazing intensities in different steppe types.
Relative grazing effects of heavy and moderate grazing differed among vegetation indicators and also among steppe types. Heavy grazing (Figure 3A) showed (1) negative effects on SR and VC in desert steppes; (2) negative effects on AGB and SR, yet neutral effects on VC in dry steppes; (3) positive effects on SR, yet negative effects on VC in meadow steppes; (4) negative effect on AGB, yet neutral effects on SR and VC in mountain steppes; (5) negative effects on AGB and SR, yet neutral effects on VC in high mountain steppes. Moderate grazing (Figure 3B) had relatively weaker negative effects, or neutral and even positive impacts compared to heavy grazing. Particularly, the effect of moderate grazing on SR was neutral except for meadow steppes.
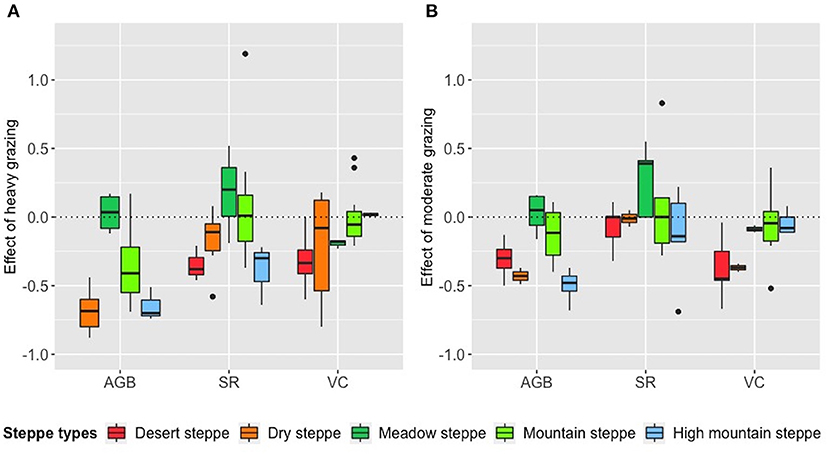
Figure 3. Relative grazing effects of (A) heavy and (B) moderate grazing on AGB, SR and VC in five different steppe types: Relative grazing effects were calculated as (heavy-light)/light grazing, and (moderate-light)/light grazing, respectively. There was no data on AGB under heavy grazing conditions in the desert steppes, so that box was not drawn. The abbreviations for vegetation indicators are the same as in Table 2.
Results from linear models showed that MAP and elevation had significant interactions with heavy grazing on AGB and VC, respectively (Table 3). Grazing effects on AGB were more negative when MAP was lower, while the magnitude of the effect decreased as MAP increased (Figure 4A); In addition, effects of heavy grazing on VC were neutral or even slightly positive at higher elevations, but became predominantly negative at lower elevations (Figure 4B). The relative effects of moderate grazing on VC interacted with MAP, i.e., moderate grazing showed neutral effects on VC when MAP was high (around 320 mm), but effects became strongerly negative when MAP was lower (Table 3; Supplementary Figure A3).
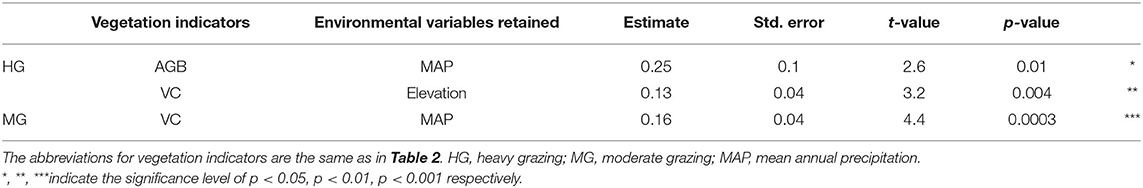
Table 3. Minimum adequate linear models for environmental interactions with relative grazing effects (heavy and moderate grazing) on AGB (N = 28), SR (N = 30), and VC (N = 21).
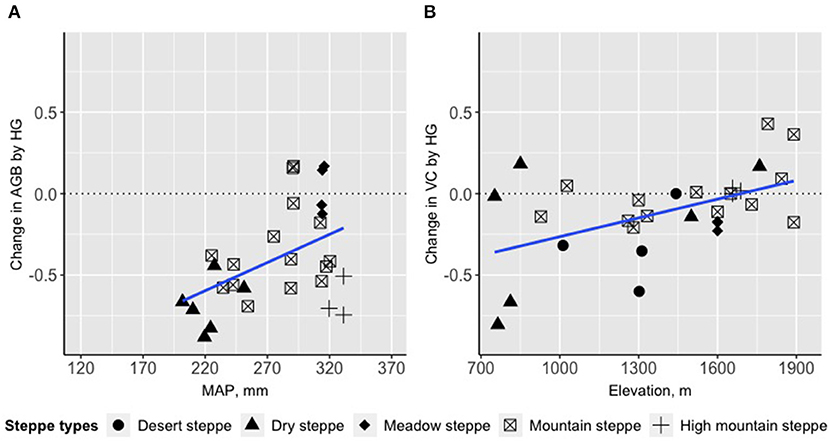
Figure 4. Change of the relative grazing effects (HG: heavy vs. light grazing) on vegetation indicators along major environmental gradients. (A) change of GE on AGB (N = 28; no data on desert steppe) along the MAP gradient, (B) change of GE on VC (N = 30) along the elevation gradient. Environmental factors are selected based on the results from Table 3.
Four studies reported fence effects across multiple years (Table 4). Three of them were carried out in mountain steppes, one in meadow steppe (one site included two steppe types) and one in desert steppe. One of these studies, i.e., the 5-year fence experiment (1970–1974) in Arkhangai province in the Khangai mountain taiga range, reported AGB values under fenced vs. grazed conditions (with three subsites of three grazing regimes, i.e., light, moderate and heavy grazing). We considered “subsite” as a repeated measurement because there were no major difference in vegetation conditions among those subsites, fence and year as the two fixed factors. The results of two-way repeated measures ANOVA showed that year [F(4,16) = 9.78, p < 0.001] and fence [F(1,4) = 30.59, p = 0.005] both had significant effects on AGB. The interannual variation of AGB was evident from both fenced and grazed plots, and the difference between fenced and unfenced plots was larger for the first 3 years, but became smaller in the 5th year (Figure 5).
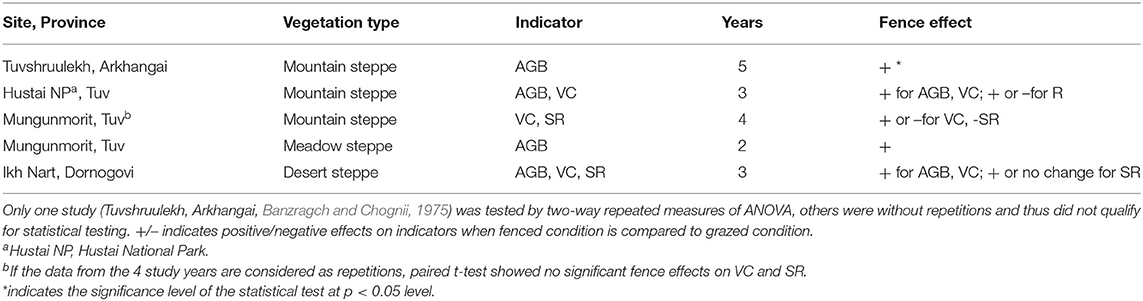
Table 4. Summary of the effects of fence/grazing intensity and year on AGB, VC, and SR in different vegetation types.
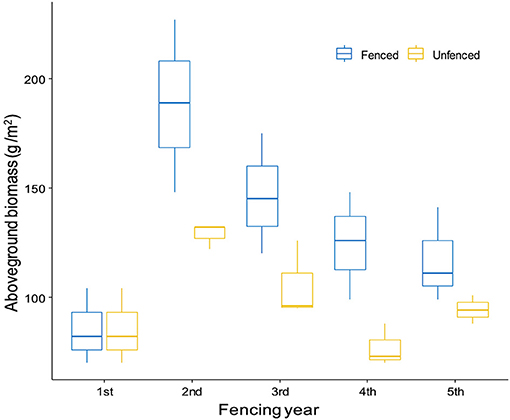
Figure 5. Fence effects on aboveground biomass across 5 years in the mountain steppe, Tuvshruulekh monitoring station in Arkhangai province (redrawn based on the data from Banzragch and Chognii, 1975). Three subsites under different grazing intensity levels were considered as repeated measurements here.
Our results demonstrated that vegetation indicators respond differently to the intensity of livestock grazing depending on the local environmental conditions. In line with our first hypothesis, heavy grazing generally had negative effects on AGB, SR and VC, and moderate grazing (compared to no/light grazing) mostly had positive effects on SR in accordance with the global reviews (Milchunas and Lauenroth, 1993; Gao and Carmel, 2020). Our results also support our second hypothesis that the magnitude of grazing effects differs among steppe types, but this also depends on indicators. Interestingly, heavy grazing had a strongly negative effect on dry steppe, desert steppe and high mountain steppes, whereas the negative effect was less pronounced, and even turned to positive or neutral in the meadow and mountain steppes. Notably, heavy grazing had no effect on (1) AGB in meadow steppes; (2) SR in mountain steppe; and (3) VC in dry steppe, mountain steppe and high mountain steppe, and a positive effect on SR in meadow steppe. Different sensitivities of steppe types reflect the interaction of large scale environmental factors with grazing effects. The pattern of moderate grazing effect is largely similar to that of heavy grazing. Thus, our further discussion will first focus on environmental interactions and then on the specific situations for certain steppe types.
A recent global meta-analysis on grazing effects on grassland vegetation showed that 41% of the variation in plant richness globally was explained by aridity, vegetation type and stocking rate together (Gao and Carmel, 2020). Several meta-analyses conducted in the grasslands of China showed that heavy grazing significantly decreased plant biomass (above and below ground biomass, also litter), but the extent of these responses was highly dependent on grazing intensity and environmental conditions, especially precipitation (Yan et al., 2013; Wang and Wesche, 2016; Hao and He, 2019). All of these studies emphasized the importance of the interplay between grazing intensity and environmental conditions. Grazing effects on plant species composition and ecosystem functions, i.e., forage provision, in grasslands largely depend on precipitation and its variability (Sloat et al., 2018).
Heavy grazing had mostly negative effects on AGB with exceptions at some sites from meadow steppe and mountain steppe, which is in line with equilibrium theories that emphasize closer feedback between vegetation and herbivores instead of climatic controls. The strong negative effects in dry steppes when MAP is around 220 mm indicate that non-equilibrium theory does not necessarily apply to this steppe type.
In semi-arid grassland systems, MAP and cvP play an important role in shaping vegetation dynamics (Vetter, 2005; von Wehrden et al., 2012). Our results find that MAP had interactions with grazing effects. This pattern could be partly explained by equilibrium theory, which suggests in humid and productive systems such as the meadow steppe and mountain steppe, grazing effects are more evident, while prediction from non-equilibrium theory is not supported. In systems with low and unstable rainfall, such as dry steppe and desert steppe, abiotic factors have overriding effects on livestock density, thereby alleviating the grazing effects on vegetation condition. In particular, negative grazing effects on AGB (Figure 4A) were weaker under higher MAP, but became stronger when MAP decreased, which might also indicate interactions with local soil nutrient conditions: AGB could be favored by grazing in nutrient-rich habitats like meadow and mountain steppes. However, reduced AGB only implies livestock removes forage, but does not indicate degradation. Grazing effects on AGB were more detrimental in nutrient-poor habitats, such as dry and high mountain steppes, where MAP was low and cvP-values were relatively high (Proulx and Mazumder, 1998; Fynn and O'Connor, 2000). However, we noticed that the long-term cvP-values based on Worldclim 2.1 for desert steppes were too low, with median of 22% (Supplementary Figure A2). However, in typical non-equilibrium systems, cvP-value should be usually >33%. Moreover, for recent years (2010–2018), cvP-values based on meteorological station data were ca. 37%, indicating stronger variation of precipitation in this steppe type. The difference of cvP-values from different time periods and different data sources might affect the results of the significance tests for cvP-values.
Elevation showed significant interactions with grazing, with grazing effects on VC being less pronounced or even slightly positive at the higher elevation. According to our linear models, elevation was more important than MAT and we thus assume that this is not a mere result of temperature declining with elevation. The change of VC was relatively constant between 1,500 and 1,900 m, but grazing effects were negative at the lower elevations in dry and desert steppes. The difference in plant community properties might explain this pattern. From low to high elevation, species composition changes from grass-sedge-forb to forb-shrub dominated vegetation (Lkhagva et al., 2013), and at higher elevation, plants tend to be smaller and/or shorter to adapt environmental stress (Halbritter et al., 2018), which may lead to less accessible phytomass and VC for livestock. Only a few studies have looked into the relationship between elevation and grazing on Mongolian rangeland vegetation. In particular, Zemmrich et al. (2010) found that elevation was not an important factor in grazing-induced vegetation change, although they pointed out that their analysis covered a limited altitudinal range. In addition, Lkhagva et al. (2013) compared lower and upper steppes on south- and north-facing slopes in northern Mongolia, and found variable responses of VC, AGB, and SR change depending on nutrient and water availability along the elevation gradient.
We found a negative grazing effect throughout on AGB and SR, while there was no effect on VC in the high mountain steppes. Former studies reported that higher elevation mountain steppe pastures in the desert zone were more degraded than lower elevation vegetation receiving less rainfall but similar grazing pressure (Sheehy and Damiran, 2012). In our case, the high mountain steppes differ from all other types by their extremely low temperatures (MAT −6°C, Supplementary Figure A2a) and lower AGB, thus supporting the lowest number of livestock among all steppe types (Figure 1). It is surprising that SR decreased under grazing there because most plants in high mountains are well-adapted to cold and nutrient-poor environments, which often also is beneficial with respect to grazing avoidance or resistance (Miehe et al., 2011). For example, growth forms that stay close to the ground, i.e., rosette or cushion plants, are adapted to strong wind and low temperatures, but are also protected from grazing because livestock can hardly reach and graze them if they are palatable. The average VC in this steppe type is ca. 60%, in which cold-adapted cushion plants [e.g., Stellaria pulvinata Grubov, Androsace chamaejasme Wulfen and Eremogone meyeri (Fenzl.) Ikonn.] account for nearly 30% of the VC, while common caespitose grass species [e.g., Festuca lenensis Drobov, Koeleria altaica (Domin.) Krylov and Poa attenuata Trin.] account for just about 10% of the VC, and the rest are other forbs and semi-shrubs (Tuvshintogtokh, 2015). Therefore, heavy grazing might directly reduce the AGB of palatable graminoids and forbs, but grazing-tolerant plants might not be affected. The negative effects on SR might relate to local topographical conditions, such as the steepness of the slope, which lead to sensitivity to trampling and difficulty of seedling establishment.
It is also surprising that in the mountain steppes, VC and SR were not affected by heavy grazing, although AGB was reduced. This is contradictory to a previous English publication (Takatsuki et al., 2018), which may point to some publication bias that more pronounced results are internationally published while non-significant results remain local. Non-significant grazing effects might be attributed to a shift in community plant shift at species level and plant functional type levels, i.e., balances between decreasing palatable species, and increasing unpalatable species. Several studies noted substantial floristic and edaphic changes caused by grazing in mountain steppes. The proportion of grazing tolerant sedges increased and highly palatable grasses and shrubs decreased under heavy grazing (Zemmrich et al., 2010; Khishigbayar et al., 2015; Jamiyansharav et al., 2018). Lkhagva et al. (2013) found that heavy livestock grazing increased SR of wet and nutrient-rich sites in the boreal mountain steppe, which was, however, explained by local colonization of grazing-tolerant species.
Interestingly, SR and AGB responded positively to heavy grazing, although VC decreased in meadow steppes. Increased SR may be attributed to increase in free patches of bare soil, thus giving new seedlings, especially of annual species, chances to germinate (Frank, 2005). Additionally, plant density is highest in meadow steppes among all steppe types due to high levels of precipitation, soil moisture and nutrient (Pfeiffer et al., 2020), resulting in strong competition among species. Thus, heavy grazing can remove large amounts of AGB of dominant palatable species, and reduce competitive exclusion among species, which then gives less dominant species a chance to survive and grow. However, the increase in SR might not necessarily indicate the improvement of steppe condition in terms of forage quality, because the increase might only involve unpalatable species, as shown in the study by Yuan et al. (2016), where unpalatable annual species (i.e., Artemisia annua L. and A. sieversiana L.) increased in grazing-disturbed sites. The increase in AGB might also suggest compensatory growth when defoliated plants produce more biomass than non-defoliated plants (Oesterheld and McNaughton, 1991). Although no evidence of compensatory growth from meadow steppes in Mongolia is available, the grazing effects on cumulative growth of plants could be positive when the availability of leaf area, meristems, stored nutrients, and soil resources are high, and the frequency and intensity of defoliation are still suitable to stimulate plant regrowth (Noy-Meir, 1993). Moreover, the limited effects of grazing on AGB in meadow steppes also imply that the extend of biomass removal was not so large.
We noted negative grazing effects on AGB and SR, and a highly variable response of VC in dry steppes. The reduction of AGB and SR in dry steppes was reported in different studies in Mongolia (Bat-Oyun et al., 2016; Jamiyansharav et al., 2018), where plant species composition shifted, i.e., a decrease in palatable species due to the increase of grazing intensity. However, the effect was associated with local climate patterns, with grazing having a greater impact at drier sites than on wetter sites (Jamiyansharav et al., 2018). The highly variable response of the VC could be attributed to the combination of two steppe types (dry steppes comprise typical steppes as well), in which plant cover might remain invariant in the moister steppes, but in drier sites, variation in VC was pronounced due to variability in onset and total amount of annual precipitation. Thus, large inter-annual variations might also lead to large variations in the VC, such as in desert steppes (Table 1). Another possible explanation could be that grazing reduces plant height and sexual reproduction, so the plants increase their vegetative growth by horizontal growth (broadening) (Gao et al., 2015). In this way, VC may increase although biomass decreases. Finally, similar to other steppe types, lack of change in VC might be related to potential shifts in species composition (not studied).
In contrast to generally limited or even absent effects on aboveground parts that were found in many other studies in the desert steppes of Mongolia (Fernandez-Gimenez and Allen-Diaz, 2001; Wesche et al., 2010; Jamiyansharav et al., 2018) and Inner Mongolia (Bai et al., 2012), we found that SR and VC decreased by nearly 40% under heavy grazing (no data for AGB). The desert steppe vegetation is often considered to show non-equilibrium dynamics, which should be overridingly controlled by rainfall variability rather than grazing. Plants adapted to drought are also often tolerant of grazing, thus grazing effects on vegetation are often insignificant (Jäschke et al., 2020). However, soil moisture deficiency is the main driving factor in the desert steppe (Zemmrich et al., 2010), thus grazing-reduced litter and plant cover might intensify the water deficiency, make sandy soils more vulnerable to soil erosion (Li et al., 2008). Soil degradation then has subsequent negative feedback on vegetation, especially on VC. It is also important to note that the desert steppes of Mongolia and Inner Mongolian are under different ranges of MAP, i.e., 150–250 mm in China (Li et al., 2020), but 110–170 mm in Mongolia (Pfeiffer et al., 2020). The possible reasons for the inconsistency between our results and previous studies might be the differences in (1) moisture availability, which is closely related to soil texture, and (2) dominant plant community, i.e., presence/absence of subshrubs or shrubs, or ruderal species. For example, when ruderal forbs, particularly Chenopodium spp. become prevalent in heavily grazed sites, SR could be much lower compared to no/light grazing (Fernandez-Gimenez and Allen-Diaz, 2001). Additionally, when the percentage change of SR caused by grazing was around 10%, it might only involve the loss of 1–2 species compared to 8–19 species under ungrazed conditions in the southern Mongolian desert steppes (Wesche et al., 2010). The desert steppe has limited functional redundancy, thus its ecological functioning is sensitive to species loss (Liu et al., 2013a). Perhaps most important, although there are still interannual fluctuations in the total livestock number in the desert steppes, the general trend is toward increasing numbers (Figure 2), which indicates that non-equilibrium dynamics may not apply there anymore.
Our review shows that assessment of grazing effects depends on indicators being selected, and on steppe type under different abiotic environmental conditions. Elevation and precipitation patterns interact with grazing effects. According to our findings, meadow steppes demonstrated equilibrium dynamics, while responses of desert steppes did not support non-equilibrium dynamics. Although desert steppes have the highest cvP among all steppe types, the general increasing trend of livestock numbers (Figure 2) conclusively demonstrates that non-equilibrium dynamics simply do not hold anymore.
Responses in the mountain steppe might depend on local soil nutrient conditions. As such, further studies focused on vegetation should also include soil conditions. High mountain steppes share some characteristics with alpine steppes in other regions, but further information on topographical conditions might be helpful for clarifying grazing effects there. We also found support that moderate grazing promotes species richness in steppes where resource availability is relatively high and stable, i.e., meadow steppes. Therefore, differences in grazing impact among steppe types should be acknowledged for grassland management and conservation. Moreover, change in VC or AGB alone, without information on species composition change, especially the proportion of forage (palatable) species, may not be the best indicators of grassland condition change, thus grassland assessment on degradation should consider more relevant indicators.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
YJ and KW conceived the ideas and designed the methodology. OM collected the relevant data, established the database, and estimated the livestock numbers for different ecological zones of Mongolia (Figure 2). YJ and KO analyzed data and led the writing of the manuscript. NN, IT, and BO checked the database in correspondence to the publications of Mongolian and Russian Joint Biological Expedition and others. All authors contributed to the article and approved the submitted version.
OM was supported by the German Federal Ministry of Education and Research (BMBF) within the joint research program BioTip-Morestep (No. 01LC1820C, BMBF BiotTip program) for her doctoral study. KO is funded by PhD Scholarship program for the promotion of early-career female scientists of TU Dresden. The funding sources had no further involvement in the study design, data analyses, or paper submission.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The authors are greatful to the following libraries that allowed us to access archived materials: Mongolian Academy of Science, National University of Mongolia, and Mongolian University of Life Science. We thank the National Statistics Office of Mongolia and the National Agency for Meteorology and Environment Monitoring for providing statistical and meteorological data. We are also grateful to Manidari Damdindorj for drafting the map of the ecological zones and steppe vegetation types of Mongolia (Figure 1), and to Sainnemekh Sumjidmaa for her discussion and advice on creating the database.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.703220/full#supplementary-material
1. ^Horse = 7 SU, cattle = 6 SU, camel = 5 SU, sheep = 1 SU, goat = 0.9 sheep (National Statistics Office of Mongolia, 2019).
Ahlborn, J., von Wehrden, H., Lang, B., Römermann, C., Oyunbileg, M., Oyuntsetseg, B., et al. (2020). Climate - grazing interactions in Mongolian rangelands: effects of grazing change along a large-scale environmental gradient. J. Arid. Environ. 173:104043. doi: 10.1016/j.jaridenv.2019.104043
Angerer, J., Han, G., Fujisaki, I., and Havstad, K. (2008). Climate change and ecosystems of Asia with emphasis on inner Mongolia and Mongolia. Rangelands 30, 46–51. doi: 10.2111/1551-501X(2008)3046:CCAEOA2.0.CO;2
Bai, Y. F., Wu, J. G., Clark, C. M., Pan, Q. M., Zhang, L. X., Chen, S. P., et al. (2012). Grazing alters ecosystem functioning and C:N:P stoichiometry of grasslands along a regional precipitation gradient. J. Appl. Ecol. 49, 1204–1215. doi: 10.1111/j.1365-2664.2012.02205.x
Banzragch, D., and Chognii, O. (1975). Productivity change of herbs feathergrass steppe in preservation regime. Proceeding Inst. Bot. 1, 63–69.
Bat-Oyun, T., Shinoda, M., Cheng, Y., and Purevdorj, Y. (2016). Effects of grazing and precipitation variability on vegetation dynamics in a Mongolian dry steppe. J. Plant Ecol. 9, 508–519. doi: 10.1093/jpe/rtv083
Batsaikhan, N., Buuveibaatar, B., Chimed, B., Enkhtuya, O., Galbrakh, D., Ganbaatar, O., et al. (2014). Conserving the world's finest grassland amidst ambitious national development. Conserv. Biol. 28, 1736–1739. doi: 10.1111/cobi.12297
Cingolani, A. M., Noy-Meir, I., and Díaz, S. (2005). Grazing effects on rangeland diversity: a synthesis of contemporary models. Ecol. Appl. 15, 757–773. doi: 10.1890/03-5272
Dagvadorj, D., Dorjpurev, J., and Namkhainyam, B. (2010). Climate Change Mitigation (Requirements and Opportunities to Reduce Greenhouse Gas Emissions). Ulaanbaatar: Ministry of Environment and Green Development.
Deng, L., Sweeney, S., and Shangguan, Z. P. (2013). Grassland responses to grazing disturbance: plant diversity changes with grazing intensity in a desert steppe. Grass Forage Sci. 69, 524–533. doi: 10.1111/gfs.12065
Densambuu, B., Sainnemekh, S., Bestelmeyer, B., and Budbaatar, U. (2018). National Report on the Rangeland Health on Mongolia. Ulaanbaatar.
Ellis, J. E., and Chuluun, T. (1993). “Cross-country survey of climate, ecology and land-use among Mongolian pastoralists,” in Conference on Grassland Ecosystems of the Mongolian Steppe (Racine, WI: Wingspread Center).
Ellis, J. E., and Swift, D. M. (1988). Stability of African pastoral ecosystems: alternate paradigms and implications for development. J. Range Manag. 41, 450–459. doi: 10.2307/3899515
Fernandez-Gimenez, M., and Allen-Diaz, B. (2001). Vegetation change along gradients from water sources in three grazed Mongolian ecosystems. Plant Ecol. 157, 101–118. doi: 10.1023/A:1014519206041
Fernandez-Gimenez, M. E., and Allen-Diaz, B. (1999). Testing a non-equilibrium model of rangeland vegetation dynamics in Mongolia. J. Appl. Ecol. 36, 871–885. doi: 10.1046/j.1365-2664.1999.00447.x
Fick, S. E., and Hijmans, R. J. (2017). WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315. doi: 10.1002/joc.5086
Frank, D. A. (2005). The interactive effects of grazing ungulates and aboveground production on grassland diversity. Oecologia 143, 629–634. doi: 10.1007/s00442-005-0019-2
Fujita, N., Amartuvshin, N., Yamada, Y., Matsui, K., Sakai, S., and Yamamura, N. (2009). Positive and negative effects of livestock grazing on plant diversity of Mongolian nomadic pasturelands along a slope with soil moisture gradient. Grassl. Sci. 55, 126–134. doi: 10.1111/j.1744-697X.2009.00147.x
Fynn, R. W. S., and O'Connor, T. G. (2000). Effect of stocking rate and rainfall on rangeland dynamics and cattle performance in a semi-arid savanna, South Africa. J. Appl. Ecol. 37, 491–507. doi: 10.1046/j.1365-2664.2000.00513.x
Gao, J., and Carmel, Y. (2020). A global meta-analysis of grazing effects on plant richness. Agric. Ecosyst. Environ. 302:107072. doi: 10.1016/j.agee.2020.107072
Gao, W., Angerer, J. P., Fernandez-Gimenez, M. E., and Reid, R. S. (2015). “Is overgrazing a pervasive problem across Mongolia? An examination of livestock forage demand and forage availability from 2000 to 2014,” in Proceedings of the Trans-disciplinary Research Conference: Building Resilience of Mongolian Rangelands, (Ulaanbaatar Mongolia), 35–41.
Halbritter, A. H., Fior, S., Keller, I., Billeter, R., Edwards, P. J., Holderegger, R., et al. (2018). Trait differentiation and adaptation of plants along elevation gradients. J. Evol. Biol. 31, 784–800. doi: 10.1111/jeb.13262
Hao, Y., and He, Z. (2019). Effects of grazing patterns on grassland biomass and soil environments in China: a meta-analysis. PLoS ONE 14:e0215223. doi: 10.1371/journal.pone.0215223
Harris, I., Jones, P. D., Osborn, T. J., and Lister, D. H. (2014). Updated high-resolution grids of monthly climatic observations - the CRU TS3.10 Dataset. Int. J. Climatol. 34, 623–642. doi: 10.1002/joc.3711
Henwood, W. D. (2010). Toward a strategy for the conservation and protection of the world's temperate grasslands. Gt. Plains Res. 20, 121–134. Available online at: http://digitalcommons.unl.edu/greatplainsresearch/1074
Jamiyansharav, K., Fernández-Giménez, M. E., Angerer, J. P., Yadamsuren, B., and Dash, Z. (2018). Plant community change in three Mongolian steppe ecosystems 1994-2013: applications to state-and-transition models. Ecosphere 9:e02145. doi: 10.1002/ecs2.2145
Jäschke, Y., Heberling, G., and Wesche, K. (2020). Environmental controls override grazing effects on plant functional traits in Tibetan rangelands. Funct. Ecol. 34, 747–760. doi: 10.1111/1365-2435.13492
Kassambara, A. (2020a). rstatix: Pipe-Friendly Framework for Basic Statistical Tests. R package version 0.6.0. Available online at: https://CRAN.R-project.org/package=rstatix
Kassambara, A. (2020b). ggpubr: 'ggplot2' Based Publication Ready Plots. R package version 0.4.0. Available online at: https://CRAN.R-project.org/package=ggpubr
Khishigbayar, J., Fernández-Giménez, M. E., Angerer, J. P., Reid, R. S., Chantsallkham, J., Baasandorj, Y., et al. (2015). Mongolian rangelands at a tipping point? Biomass and cover are stable but composition shifts and richness declines after 20 years of grazing and increasing temperatures. J. Arid Environ. 115, 100–112. doi: 10.1016/j.jaridenv.2015.01.007
Li, C., Hao, X., Zhao, M., Han, G., and Willms, W. D. (2008). Influence of historic sheep grazing on vegetation and soil properties of a Desert Steppe in Inner Mongolia. Agric. Ecosyst. Environ. 128, 109–116. doi: 10.1016/j.agee.2008.05.008
Li, F. Y., Jäschke, Y., Guo, K., and Wesche, K. (2020). “Grasslands of China,” in Encyclopedia of the World's Biomes, Vol. 3, eds M. I. Goldstein and D. A. DellaSala (Elsevier), 773–784.
Liang, C., Michalk, D. L., and Millar, G. D. (2002). The ecology and growth patterns of Cleistogenes species in degraded grasslands of eastern Inner Mongolia, China. J. Appl. Ecol. 39, 584–594. doi: 10.1046/j.1365-2664.2002.00735.x
Liu, G., Xie, X., Ye, D., Ye, X., Tuvshintogtokh, I., Mandakh, B., et al. (2013a). Plant functional diversity and species diversity in the Mongolian steppe. PLoS ONE 8:e77565. doi: 10.1371/journal.pone.0077565
Liu, Y. Y., Evans, J. P., McCabe, M. F., de Jeu, R. A. M., van Dijk, A. I. J. M., Dolman, A. J., et al. (2013b). Changing climate and overgrazing are decimating mongolian steppes. PLoS ONE 8:e57599. doi: 10.1371/journal.pone.0057599
Lkhagva, A., Boldgiv, B., Goulden, C. E., Yadamsuren, O., and Lauenroth, W. K. (2013). Effects of grazing on plant community structure and aboveground net primary production of semiarid boreal steppe of northern Mongolia. Grassl. Sci. 59, 135–145. doi: 10.1111/grs.12022
MARCC. (2014). Mongolia Second Assessment Report on Climate Change-2014, eds. D. Dagvadorj, Z. Batjargal, and L. Natsagdorj (Ulaanbaatar: Ministry of Environment and Green Development).
Miehe, G., Bach, K., Miehe, S., Kluge, J., Yang, Y. P., La, D., et al. (2011). Alpine steppe plant communities of the Tibetan highlands. Appl. Veg. Sci. 14, 547–560. doi: 10.1111/j.1654-109X.2011.01147.x
Milchunas, D. G., and Lauenroth, W. K. (1993). Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecol. Monogr. 63, 327–366. doi: 10.2307/2937150
National Statistics Office of Mongolia. (2018). Determine the Number of Livestock in Accordance with Pasture Capacity and Conditions. Ulaanbaatar: National Statistics Office of Mongolia. Available online at: https://downloads.1212.mn/1616736153165.pdf
National Statistics Office of Mongolia. (2019). Mongolian Statistical Yearbook-2019, eds. A. Ariunzaya, C. Bayanchimeg, A. Demberel, B. Batbayar, N. Munkhbayar, and T. Zaya (Ulaanbaatar). Available online at: http://www.1212.mn/Stat.aspx?LIST_ID=976_L01&type=description
National Statistics Office of Mongolia. (2021). Mongolian Statistical Yearbook-2020, eds. A. Ariunzaya, C. Bayanchimeg, A. Demberel, B. Batbayar, N. Munkhbayar, and T. Zaya (Ulaanbaatar). Available online at: http://www.1212.mn/stat.aspx?LIST_ID=976_L10_1
Noy-Meir, I. (1993). Compensating growth of grazed plants and its relevance to the use of rangelands. Ecol. Appl. 3, 32–34. doi: 10.2307/1941787
Oesterheld, M., and McNaughton, S. J. (1991). Effect of stress and time for recovery on the amount of compensatory growth after grazing. Oecologia 85, 305–313. doi: 10.1007/BF00320604
Olff, H., and Ritchie, M. E. (1998). Effects of herbivores on grassland plant diversity. Trends Ecol. Evol. 13, 261–265. doi: 10.1016/S0169-5347(98)01364-0
Pfeiffer, M., Dulamsuren, C., and Wesche, K. (2020). “Grasslands and shrublands of Mongolia,” in Encyclopedia of the World's Biomes, eds. M. I. Goldstein and DABT-E of the W. B. DellaSala (Oxford: Elsevier), 759–772.
Proulx, M., and Mazumder, A. (1998). Reversal of grazing impact on plant species richness in nutrient-poor vs. nutrient-rich ecosystems. Ecology 79, 2581–2592. doi: 10.1890/0012-9658(1998)0792581:ROGIOP2.0.CO;2
R Core Team (2020). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available online at: https://www.R-project.org/
Sheehy, D. P., and Damiran, D. (2012). Assessment of Mongolian Rangeland Condition and Trend (1997-2009). Final Report for the World Bank and the Netherlands-Mongolia Trust Fund for Environmental Reform (NEMO), 46.
Sloat, L. L., Gerber, J. S., Samberg, L. H., Smith, W. K., Herrero, M., Ferreira, L. G., et al. (2018). Increasing importance of precipitation variability on global livestock grazing lands. Nat. Clim. Chang. 8, 214–218. doi: 10.1038/s41558-018-0081-5
Takatsuki, S., Sato, M., and Morinaga, Y. (2018). Effects of grazing on grassland communities of the forest-steppe of northern Mongolia: a comparison of grazed versus ungrazed places. Grassl. Sci. 64, 167–174. doi: 10.1111/grs.12195
Tuvshintogtokh, I. (2015). The Steppe Vegetation of Mongolia, ed. C. Sanchir (Ulaanbaatar: Bembi San), 610.
Ulziikhutag, N. (1985). Identification Key of Plants in Pastures and Hayfields of the People's Republic of Mongolia, ed. C. Sanchir (Ulaanbaatar: Ulsiin hevleliin gazar), 556.
Vetter, S. (2005). Rangelands at equilibrium and non-equilibrium: recent developments in the debate. J. Arid Environ. 62, 321–341. doi: 10.1016/j.jaridenv.2004.11.015
von Wehrden, H., Hanspach, J., Kaczensky, P., Fischer, J., Wesche, K., von Wehrden, H., et al. (2012). Global assessment of the non-equilibrium concept in rangelands. Ecol. Appl. 22, 393–399. doi: 10.1890/11-0802.1
Wang, Y., and Wesche, K. (2016). Vegetation and soil responses to livestock grazing in Central Asian grasslands: a review of Chinese literature. Biodivers. Conserv. 25, 2401–2420. doi: 10.1007/s10531-015-1034-1
Wesche, K., Ambarli, D., Kamp, J., Török, P., Treiber, J., and Dengler, J. (2016). The Palaearctic steppe biome: a new synthesis. Biodivers. Conserv. 25, 2197–2231. doi: 10.1007/s10531-016-1214-7
Wesche, K., Ronnenberg, K., Retzer, V., and Miehe, G. (2010). Effects of large herbivore exclusion on southern Mongolian desert steppes. Acta Oecol. 36, 234–241. doi: 10.1016/j.actao.2010.01.003
White, R. S., Murray, S., and Rohweder, M. (2000). Pilot Analysis of Global Ecosystems Grassland Ecosystems. Washington, DC: World Resources Institute.
Wickham, H. (2016). ggplot2: Elegant Graphics for Data Analysis. New York, NY: Springer-Verlag. Available online at: https://ggplot2.tidyverse.org
Wickham, H., Averick, M., Bryan, J., Chang, W., McGowan, L. D. A., François, R., et al. (2019). Welcome to the tidyverse. J Open Sour Softw. 4:1686. doi: 10.21105/joss.01686
Wilke, C. (2020). cowplot: Streamlined Plot Theme and Plot Annotations for ‘ggplot2’. R package version 1.1.1. Available online at: https://CRAN.R-project.org/package=cowplot
Yan, L., Zhou, G., and Zhang, F. (2013). Effects of different grazing intensities on grassland production in China: a meta-analysis. PLoS ONE 8:e81466. doi: 10.1371/journal.pone.0081466
Yuan, Z. Y., Jiao, F., Li, Y. H., and Kallenbach, R. L. (2016). Anthropogenic disturbances are key to maintaining the biodiversity of grasslands. Sci. Rep. 6:22132. doi: 10.1038/srep22132
Yunatov, A. A., and Dashnyam, B. (1979). The vegetation map of People's Republic of Mongolia (1:1 500 000). Mong. Russ. Jt. Biol. Exped. (Ulaanbaatar).
Zemmrich, A., Manthey, M., Zerbe, S., and Oyunchimeg, D. (2010). Driving environmental factors and the role of grazing in grassland communities: a comparative study along an altitudinal gradient in Western Mongolia. J. Arid Environ. 74, 1271–1280. doi: 10.1016/j.jaridenv.2010.05.014
Keywords: steppes, grazing, diversity, biomass, climate, human impact
Citation: Munkhzul O, Oyundelger K, Narantuya N, Tuvshintogtokh I, Oyuntsetseg B, Wesche K and Jäschke Y (2021) Grazing Effects on Mongolian Steppe Vegetation—A Systematic Review of Local Literature. Front. Ecol. Evol. 9:703220. doi: 10.3389/fevo.2021.703220
Received: 09 June 2021; Accepted: 24 September 2021;
Published: 25 October 2021.
Edited by:
Balázs Deák, Hungarian Academy of Science, HungaryCopyright © 2021 Munkhzul, Oyundelger, Narantuya, Tuvshintogtokh, Oyuntsetseg, Wesche and Jäschke. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Karsten Wesche, a2Fyc3Rlbi53ZXNjaGVAdHUtZHJlc2Rlbi5kZQ==; a2Fyc3Rlbi53ZXNjaGVAc2VuY2tlbmJlcmcuZGU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.