 Mercedes Molina-Morales1*
Mercedes Molina-Morales1* Jesús Gómez2
Jesús Gómez2 Gustavo Liñán-Cembrano3
Gustavo Liñán-Cembrano3 Marta Precioso1
Marta Precioso1 Juan G. Martínez1
Juan G. Martínez1 Jesús M. Avilés4,5
Jesús M. Avilés4,5- 1Departmento of Zoología, Universidad de Granada, Granada, Spain
- 2Independent Research, Madrid, Spain
- 3Instituto de Microelectrónica de Sevilla (IMSE-CNM), Consejo Superior de Investigaciones Científicas (CSIC), Universidad de Sevilla, Seville, Spain
- 4Departamento de Ecología Funcional y Evolutiva, Estación Experimental de Zonas Áridas (EEZA) (CSIC), Almería, Spain
- 5Unidad Asociada (CSIC): Ecología en el Antropoceno, Facultad de Ciencias, Universidad de Extremadura, Badajoz, Spain
The existence of a coevolutionary process between avian brood parasites and their hosts predicts a lower intra-clutch variation in egg appearance of host eggs among rejecters as this would favor egg discrimination of parasite eggs by hosts once parasitic egg mimicry had evolved. So far empirical tests of this prediction have ignored the fact that different aspects of host egg phenotypes may differ in the relative role of environmental vs. genetic determination, and hence that the role of intra-clutch variation in egg rejection within a population cannot be invariant. Here, we estimated whether the intra-clutch variation in several aspects of host eggshell features is consistently associated to rejection of parasitic foreign eggs across years in a magpie host population parasitized by great spotted cuckoos. We innovatively estimated spottiness by means of the fractal dimension of eggs, which considers the homogeneity of spot pattern complexity in eggshells. Our results show that low intra-clutch variation in the blue-green coloration at the middle area of the eggs associated with a high chance of rejection, but only in one of the 3 years we conducted the study. In addition, females that rejected foreign eggs presented more homogenous spot patterns in their clutches as estimated by their fractal dimension than females that accepted experimental eggs, independently of the year of study. Finally, intra-clutch variation in egg volume of host eggs was not associated to rejection. Analyses at the individual level revealed that the relative role of genetic vs. environmental factors that determine egg phenotype would be feature-specific in magpies, females having a characteristic spottiness, but not color or volume, pattern. Our work stresses the importance of considering a holistic approach including several aspects of variation in host egg phenotype (size, color, and homogeneity of spot pattern), as some aspects might be more susceptible to selection through egg rejection than others, presumably because they are less influenced by variation in the environmental conditions. Moreover, our study highlights the importance of replication in studies on the adaptive value of host traits in egg rejection.
Introduction
Avian brood parasites impose dramatic costs on the reproductive success of their hosts, which has led to the evolution of counter-adaptations to circumvent them (Rothstein, 1990; Davies, 2000). One of the most widespread defenses against parasitism among hosts is the discrimination and rejection of parasite eggs from their nests. Egg discrimination reduces the harm caused by brood parasitism, and selects for egg mimicry in the parasite eggs to counter host defenses. Once brood parasites have evolved eggs that mimic those of their hosts, a further step in the arms race would be the evolution of a smaller degree of intra-clutch variation in the appearance of host eggs, as this could facilitate hosts to discriminate parasitic eggs (Victoria, 1972; Øien et al., 1995; Soler and Møller, 1996; Moskát et al., 2008). After more than two decades of empirical work examining the key prediction of this hypothesis (the intra-clutch variation hypothesis) in different host-brood parasite systems, the literature is equivocal regarding whether homogeneity in host egg appearance favors or not the discrimination of parasitic egg. Some studies have shown that females with lower intra-clutch variation were more likely to reject artificial foreign eggs as it would be expected (Stokke et al., 1999; Soler et al., 2000; Peer et al., 2010; Wang et al., 2016), but others found the opposite trend (Lotem et al., 1995; Avilés et al., 2004). Noteworthy, in some studies host intra-clutch variation was no associated with the probability of rejecting model foreign eggs (Karcza et al., 2003; Croston and Hauber, 2015). Discrepancy in the pattern also arises when the eggs used in experiments were conspecific or real cuckoo eggs (Procházka and Honza, 2003; Moskát and Lovászi, 2004; Stokke et al., 2004; Cherry et al., 2007; Landstrom et al., 2010; Polačiková et al., 2011; Abernathy and Peer, 2014).
Most of these studies have in common that they have been performed in one single population and study year providing a “snapshot” of the role of intra-clutch variation in rejection of foreign eggs. This approach implicitly ignores the fact that the different features of the avian egg phenotype, such as egg-shell color, degree and pattern of spottiness and egg volume, although partly genetically determined (Gosler et al., 2000; Liu and Cheng, 2010), also depend on climatic conditions, female condition or local availability of dietary components necessary for the synthesis of pigments included in the egg-shell (Moreno and Osorno, 2003; Avilés et al., 2007; Hubbard et al., 2010; Berkowic et al., 2015). The relative role of the genetic vs. the environmental component might hence be feature- or species- specific, which may have hampered our abilities to obtain reliable and reproducible patterns about the role of egg homogeneity for rejection. The effect of environmental conditions on egg phenotypes may even differ between brood parasites and their hosts rendering changes in mimicry that may potentially affect rejection (Avilés et al., 2007). Therefore, if, as it is common in terrestrial environments, conditions change from 1 year to another, the role of intra-clutch variation in egg rejection might differ between breeding seasons and for the different aspects of the egg’s phenotype. So far, very few studies have addressed changes in eggshell color or spottiness in time (Geltsch et al., 2017) in the context of the coevolution of host and parasite eggs.
Illustrating this issue, two studies on the role of intra-clutch variation in egg rejection in magpie (Pica pica) hosts have given opposite results (Soler et al., 2000; Avilés et al., 2004). Although both studies used different methods to estimate intra-clutch variation (photography vs. spectrophotometry), neither was replicated raising the possibility that differences between studies were merely due to environmental effects. Replication is important to draw general patterns and reach well-founded conclusions, and although replicates are not common in behavioral ecology research (Kelly, 2006, 2019; Nakagawa and Parker, 2015); they have often led to different and inconsistent results (Kelly, 2006). Replication has allowed the identification of factors that may affect egg rejection in magpies, such as female age (Molina-Morales et al., 2014; Martínez et al., 2020), that were previously discarded (Soler et al., 2000). Here, aiming to qualify the importance of replication in the assessment of cuckoo-host evolutionary hypotheses, we have estimated homogeneity (that is, intra-clutch variation) in host egg phenotype and tested egg rejection in a magpie population parasitized by the great spotted cuckoo (Clamator glandarius) during 3 consecutive years. We estimated the homogeneity of egg volume based on egg measurements, homogeneity of eggshell coloration using spectrophotometry, and, innovatively, estimated homogeneity of spottiness using the fractal dimension of eggshells calculated from pictures of the clutches. The fractal dimension (FD hereafter) is a parameter that measures complex patterns in objects whose value is influenced by properties that include number, length, turgidity, and connectivity of elements within a given object (Mandelbrot, 1983). FD is scale invariant providing a single value that summarizes the way the pattern “behaves” across scales (Mandelbrot, 1983; Jovani et al., 2013), and it has been used to describe complex color patterns or designs in living organisms, such as the complex color patterns of sexual ornaments in birds (Pérez-Rodríguez et al., 2013; Cantarero et al., 2018), the porosity of avian eggs (Zhang and Wang, 2012) or the homogeneity of spottiness in bird eggs and the amount of protoporphyrin deposited on the eggshell (Gómez et al., 2019, 2021).
Our goal is twofold: we examine the association between intra-clutch variation and egg rejection using different aspects of egg phenotype and multiple years, taking a more holistic approach to the problem. In order to better understand previous inconsistent results and the between-year variability in our results, we also study the variability of egg features across years, and analyze the repeatability of these in a subset of females that bred more than 1 year in our population, as a basis to determine trait-specific sensitivity of eggs to environmental variation. We predict that high homogeneity in egg appearance (low intra-clutch variation) is associated with rejection of foreign eggs when the traits are less plastic and more female-specific.
Methodology
Study Area and System
The study was conducted in La Calahorra (37° 10′ N, 3° 03′ W, Hoya de Guadix, Granada, Southern Spain) during the years 2016–2018. It is a patchy area of about 12 km2 where groves of almond trees (Prunus dulcis), in which magpies preferentially build their nests, are very common. Magpies are territorial, sedentary, and socially monogamous long-lived passerines (Birkhead, 1991). In our study area magpies lay one clutch during April-May, and are the main host of the great spotted cuckoo (Clamator glandarius).
Monitoring Magpie Individuals and Nests
We started to monitor the nests once the first pair starts nest building, usually at the beginning of March and continue until July when the breeding season ended. Once a nest is found we visit it once a week to detect egg laying. Nests were found by careful inspection of all trees in the area, GPS positioned and then visited at 5 days intervals. In order to determine whether nests were parasitized, during egg laying and hatching, they were visited every 2–3 days. We considered that nests were parasitized when at least one cuckoo egg was found in them. Magpies in our study area only reject about 5% of real cuckoo eggs (see Soler et al., 1995), and so the risk of not detecting parasitized nests (because magpies had rejected the cuckoo eggs quickly) is very low. The information recorded on each nesting attempt included laying date (that we expressed as the number of days from the first of April), number of cuckoo and magpie eggs, and number of cuckoo and magpie nestlings that fledged. Once the clutch was completed we registered several measures that have been used to calculate intra-clutch variation in egg appearance (see below). We monitored 13 nests in 2016, 53 nests in 2017 and 71 in 2018.
Egg Rejection Experiment
We tested magpie response to mimetic model eggs to classify females as acceptors or rejecters (e.g., Soler and Møller, 1990; Soler et al., 1999). Mimetic model eggs were made of plaster of Paris mixed with white glue and painted with acrylic paints. Model eggs were made to resemble the cuckoo eggs in appearance, size, and mass to the human eye, although they do not perfectly match the color of real great spotted cuckoo eggs when measured with a spectrophotometer (see Molina-Morales et al., 2014; Figure 1). We introduced one mimetic model egg during magpie egg laying and revisited the nest after 6–7 days. Previous work in our magpie population has shown that 75% of all rejection of artificial models occurs in the first 24 h after parasitism and that after 72 h all eggs have been rejected (Avilés et al., 2004). The response was coded as rejection if the model egg disappeared from the nest or acceptance if the model egg was incubated with the host’s clutch. It could be argued that the day on which the model egg is introduced into the clutch could affect females’ evaluation of intra-clutch variation if intra-clutch variation estimated on few host’s eggs was not representative of intra-clutch variation estimated on the entire host clutch. To rule out this possibility, we analyzed the level of correlation between intra-clutch variation estimated on three randomly selected eggs and intra-clutch variation estimated from the whole clutch across a sample of 90 clutches. Since intra-clutch variation in the three parts of the eggs is correlated (Supplementary Table 1) we performed the analyses using the measurements taken the sharp pole. The level of correlation was superior to 0.63 for the three PCs suggesting that intra-clutch variation for some eggs is similar to the one calculated for the entire clutch (PC1 r = 0.63, t90 = 7.70, P ≤ 0.001; PC2 r = 0.69, t90 = 9.04, P ≤ 0.001; PC3 r = 0.70, t90 = 9.19, P ≤ 0.001). Therefore, the fact that females may have partially or fully evaluated the clutch should not affect our conclusions.

Figure 1. Magpie clutches parasitized by Great spotted cuckoo. The picture shows magpie eggs, real cuckoo egg, and experimental model eggs. (A) Eight magpie eggs, one great spotted cuckoo egg and mimetic model egg in the lower right corner. (B) Seven magpie eggs, one great spotted cuckoo egg near to mimetic model egg in the lower right corner.
Intra-Clutch Variation in Egg Appearance
We considered intra-clutch variation in three aspects of avian egg phenotype known to influence foreign egg rejection in different brood-parasite hosts systems, namely egg volume (Marchetti, 2000), spottiness (Moskát et al., 2008), that we innovatively quantify using the Fractal dimension and, egg coloration (Rothstein, 1982; Soler et al., 2000; Lyon, 2003).
- Intra-clutch variation in egg volume was estimated as the standard deviation of egg volume (calculated on length and width of each egg applying Hoyt, 1979 formula) in each clutch.
- Intra-clutch variation in FD of spottiness. Once a magpie clutch was completed it was photographed using a CANON 350D digital camera (Canon Inc., Tokyo, Japan). Images were stored in jpeg format and a metric reference (pixel to mm conversion factor) was automatically obtained by extracting a red patch of known size present in all the images. Settings were adjusted automatically depending on light conditions. We were not able to work in the real color domain (equivalent reflectance images) because neither raw images were available nor gray targets of known reflectance were present in the images in order to linearize them. Therefore, we opted for transforming the images into their CIE L∗a∗b∗ 1976 [ISO 11664-4:2008(E), 2007] versions and operated only over the lightness channel L∗. To process these images, we modified the spot detection algorithm included in SpotEgg (Gómez and Liñán-Cembrano, 2017) by defining a local threshold for every pixel in the lightness channel. For every pixel, we created an adaptive threshold (Bradley and Roth, 2007) to determine whether the pixel belongs to the background or spot domains in which, instead of the most common 1/8th of the size of the image to determine the size of the windows that define the region where local thresholds are calculated, we considered 1/20th of the length of each egg, for a larger sensitivity. The obtained spot images were latter filtered in size using mathematical morphology operations to eliminate spots whose larger axis is smaller than 1/200 the length of the egg. Finally, the black and white spot images were passed to the Fractal Dimension (FD) calculation function in SpotEgg which employs the Minkowski–Bouligand definition (Schroeder, 1991) to determine FD, commonly known as the box-counting technique.
-Intra-clutch variation in egg coloration. At the same time we took photographs, we also measured egg coloration by spectrophotometry in the field. We obtained reflectance spectra in the 360–740-nm range from 888 eggs of 137 clutches, using a Konica Minolta CM250 spectrophotometer. Color was always measured five times; the first one in the sharp pole, three measures in the area in-between the two poles and another one in the blunt pole; each measure was circa 1 mm2. We calculated the mean value from the three measures in the area in-between poles (middle area hereafter). Measurements were relative and referred to a standard white reference (WS-2) and to the dark. Reference white and dark calibrations were made before the beginning of the measurements of each clutch.
We have used Principal Component Analyses (PCA) for describing variation in eggshell color based on reflectance data (Cuthill et al., 1999; Cherry and Bennett, 2001). A PCA was performed on raw reflectance data (five measures for each magpie egg) to reduce the number of correlated variables (39 variables as reflectance values were taken at 10 nm intervals between 360 and 740-nm range) into a few orthogonal variables summarizing color variation (see for instance Cuthill et al., 1999; Avilés et al., 2006). PCA allowed us to distinguish between achromatic “brightness” variation represented by the first principal component (PC1) and chromatic variation represented by PC2 and PC3 (Endler and Thery, 1996). Together these three first components explained 99.82% of the total variance in spectra of magpie eggs. PC1 was flat and described achromatic variation explaining 94.9% of the overall variation. PC2 and PC3 were not spectrally flat and together they accounted for 85.1% of the chromatic variance (see Figure 2). PC2 had high and positive loadings at short wavelengths and high negative loadings at long wavelengths and could therefore classify the eggs sampled along a gradient of long ultraviolet-red reflectance. PC3, however, had high positive loadings approximately at the blue-green (475–550 nm) wavelength and thus could be described as a bluish-greenness gradient. To estimate the degree of intra-clutch variation in coloration, we calculated the standard deviation of brightness (PC1 scores), ultraviolet-red color (PC2 scores) and blue-green color (PC3 scores) based on average values per egg in each magpie clutch.
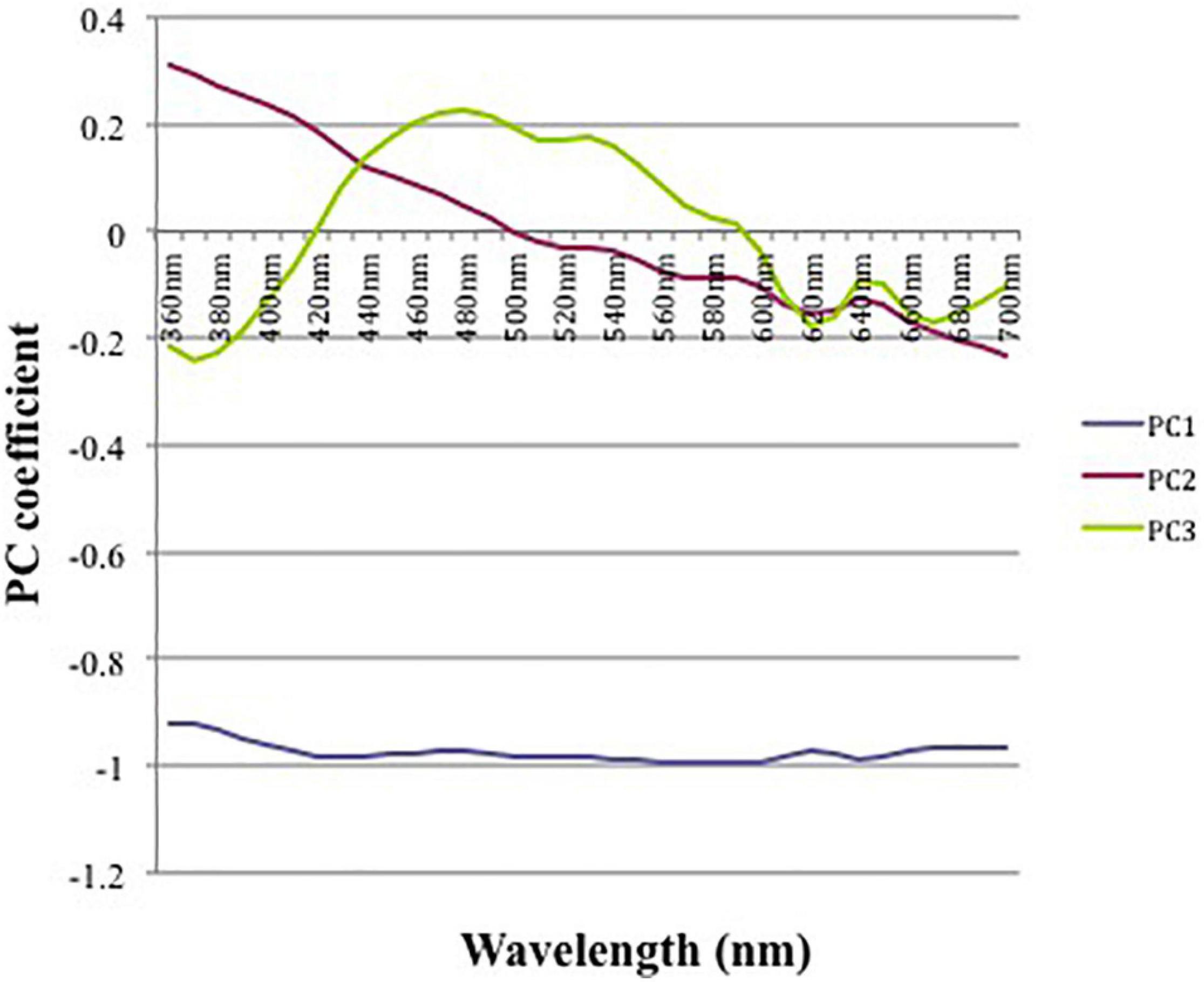
Figure 2. Principal components in relation to wavelength, derived from reflectance spectra from magpie eggs in each clutch. PC1 describes achromatic variation explaining 94.9% of the overall variation. PC2 and PC3 accounted for 85.1% of the chromatic variance. PC2 indicates ultraviolet-red colors and PC3 blue-green colors.
Statistical Analyses
Aiming to qualify the environmental nature of egg homogeneity we fitted four Mancova models. First, we considered intra-clutch variation in volume and FD as dependent variables and included clutch size and laying date as predictors and year as a fixed factor. In a second Mancova model we considered intra-clutch variation in all color (PCs) as dependent variables, and the predictors in the first model. Finally, we repeated the same model structure but using the mean values of egg volume and FD in a third Mancova model, and the mean values of color, in a fourth Mancova model, respectively. Moreover, we analyze within-individual patterns in intra-clutch variation on a subset of 21 females that bred more than once in our population and calculated repeatability of intra-clutch variation for each trait using a variance components analysis. All together these analyses aimed to help us to qualify the potential for selection of different egg features by estimating to what extent the different features are environmentally vs. genetically determined.
We fitted binomial generalized linear models (GLM) using SAS (SAS Institute, Cary, NC, United States) with rejection as dependent variable to analyse the role of intra-clutch variation in the different features considered. We run separated models for egg volume, FD of spottiness and color as predictors because a preliminary model including variation in these features, study year and their interactions did not converge. All models included study year as a fixed term and the interaction between study year and intra-clutch variation in the corresponding trait. We also entered standardized laying date as covariate, to control for possible differences in female quality (Thorley and Lord, 2015). In this system, parasitized nests have smaller clutch sizes because cuckoo females break some host eggs while parasitizing (Soler et al., 1996; Molina-Morales et al., 2013). Indeed, in this study the clutch size of parasitized nest was significantly smaller than that of non-parasitized ones [F(1, 135) = 17.56 P = 0.00005; Non-parasitized nests, mean = 6.4, SE = 0.11 n = 103; Parasitized nests mean = 5.38, SE = 0.26 n = 34). Besides be linked to parasitism, clutch size may per se affect intra-clutch variation (de Hierro and De Neve, 2010). Therefore, to avoid collinearity we opted to include clutch size, instead of parasitism, as covariate.
The model analyzing the influence of intra-clutch variation in the FD also included average spottiness as a covariate given that it is known that the FD depends on the degree of spottiness (Pérez-Rodríguez et al., 2013; Gómez et al., 2021). When dealing with color variation, we fitted separated models for the blunt pole, the sharp pole and the middle part to avoid collinearity issues, given that the degree of correlation between measures of intra-clutch variation in egg color at the three egg areas were highly positively correlated to each other (Polačiková et al., 2011; see Supplementary Table 1).
Results
Overall, we assessed the response toward model eggs in 137 magpie breeding attempts in the 3 years of study. In this study, 30 out of 137 (21.89%) mimetic model eggs were rejected (2016: 23.07%; 2017: 18.8%; 2018: 23.9%).
Yearly and Individual Variation in Egg Traits
Although intra-clutch variation in egg volume and FD [F(4, 248) = 0.443, P = 0.777] and color features (PCs) [F(18, 248) = 0.704, P = 0.805] did not change between the years (Supplementary Table 2), the average phenotype of magpie eggs in the population changed (Supplementary Table 3). Specifically, we found that the average coloration of eggs (ultraviolet-red reflectance of the sharp pole and the middle area and brightness of the blunt pole) [F(18, 248) = 2.655, P < 0.001], and the mean FD of spottiness of the eggshell, but not the mean volume, changed between years [F(4, 248) = 6.70, P < 0.001] (Supplementary Table 3 and Figure 3).
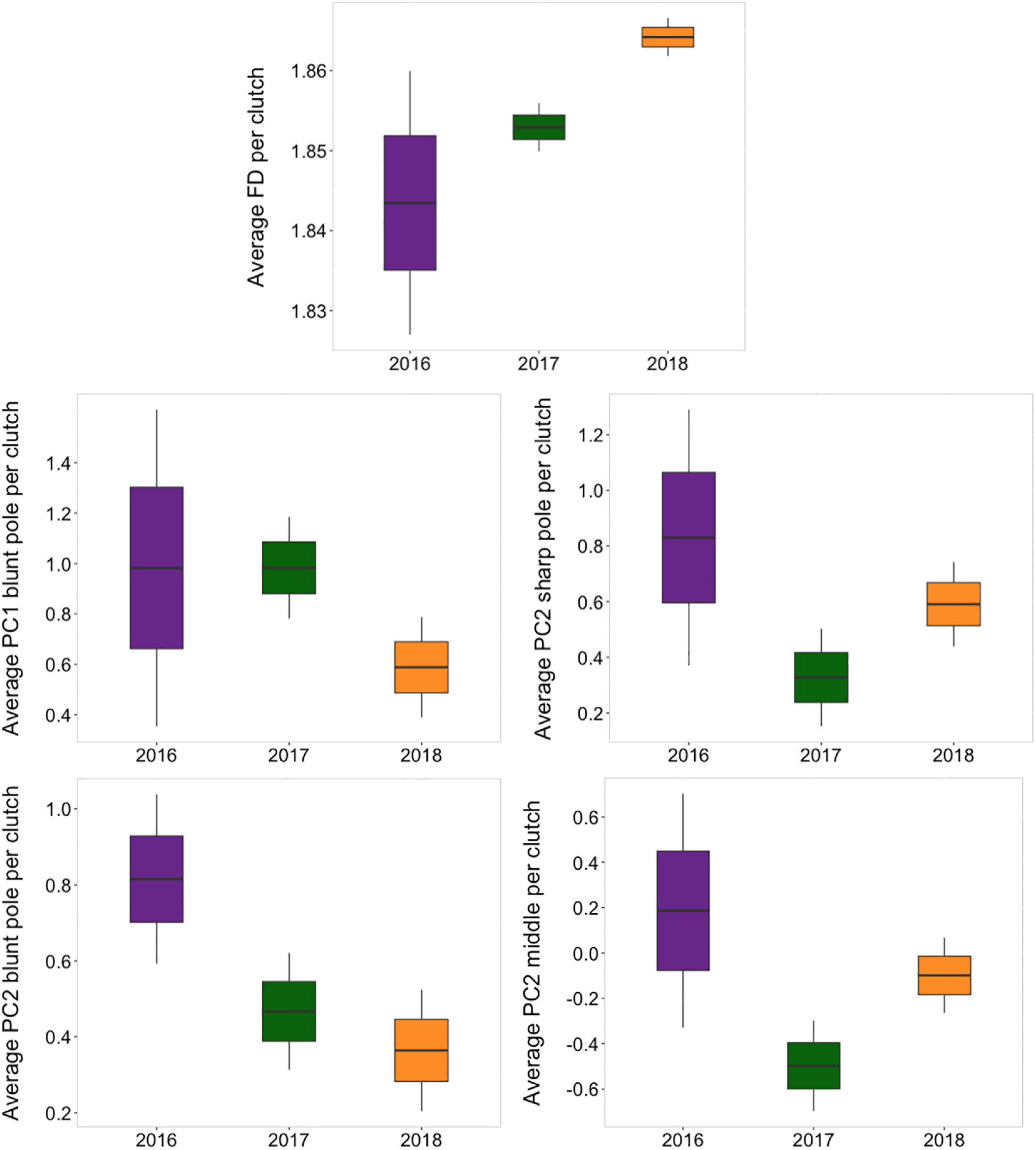
Figure 3. Yearly variation in average values per clutch of eggshell features. Horizontal line represents the mean value, boxes standard error and line bars that indicate confidence intervals (95%).
Analyses on the subset of females breeding more than once in the population revealed a low repeatability in the intra-clutch variation in egg volume and in the ultraviolet-red and blue-green colors of the eggshell (r < 0.6, p > 0.05). However, intra-clutch variation in brightness of the blunt pole was repeatable (r = 0.49, F = 2.92, df = 20, p = 0.009), as well as intra-clutch variation in FD (r = 0.58, F = 3.84, df = 20, p = 0.0017).
Intra-Clutch Variation in Egg Traits and Rejection in Different Years
Intra-clutch variation in the FD (Table 1) and in green-blue coloration in the middle area of the egg (Table 2) were associated with egg rejection of foreign eggs, and these patterns were in some cases affected by study year.
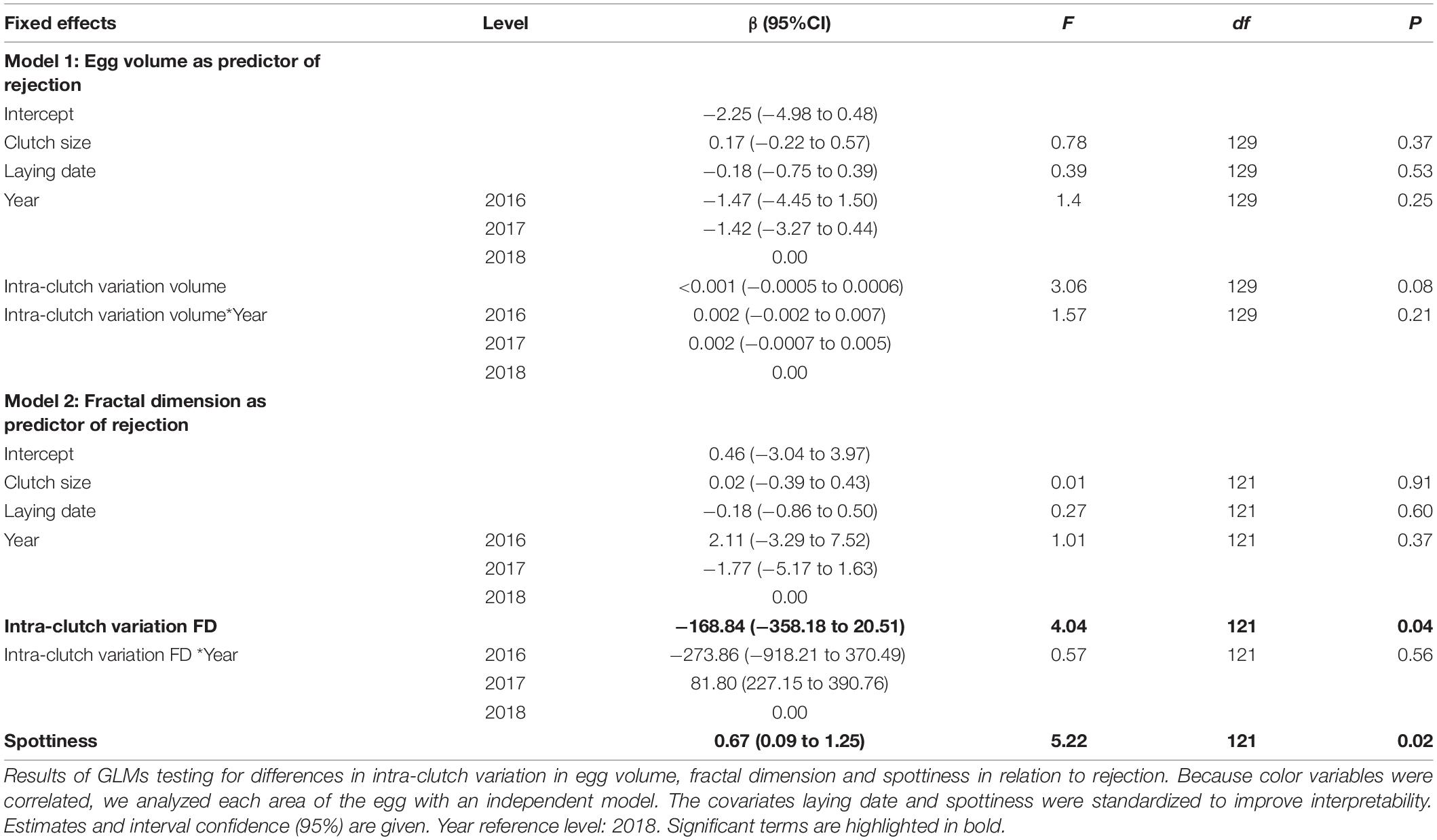
Table 1. Intra-clutch variation in egg appearance in relation to rejection of model eggs.
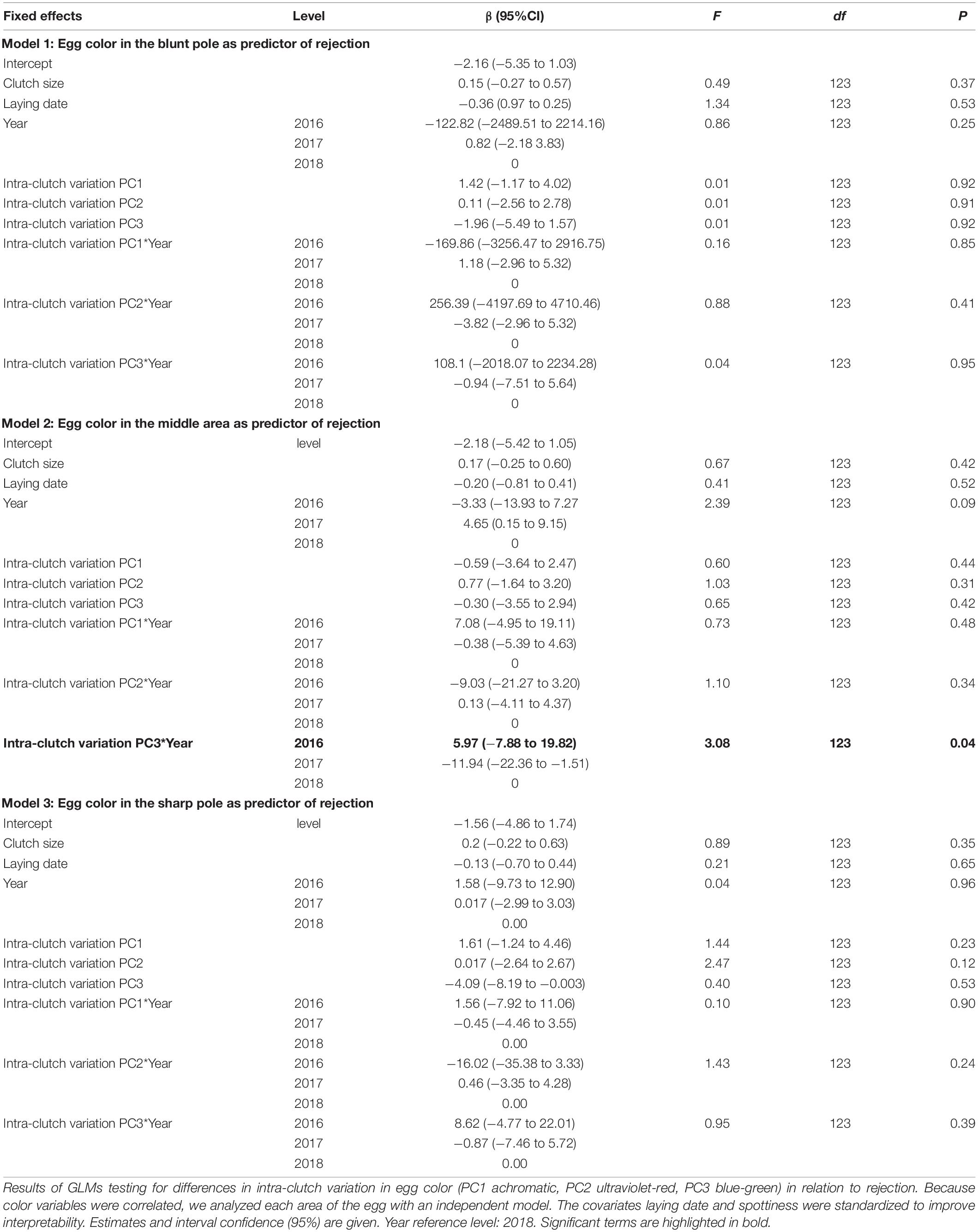
Table 2. Intra-clutch variation in egg appearance in relation to rejection of model eggs.
The probability of rejection was not associated with homogeneity in egg volume within the clutches (Table 1). However, the clutches of acceptors showed higher values of intra-clutch variation in the FD of their eggs than those of rejecters, irrespective of the year of study (Table 1 and Figure 4). Regarding eggshell color variation, rejection was not related to variation in the sharp and blunt poles (Table 2). However, intra-clutch variation in blue-green color (i.e., PC3 score) in the middle area of the eggs differed between acceptor and rejecter magpies in interaction with the year (Table 2 and Figure 5). Rejecters showed significantly lower intra-clutch variation in blue-green color than acceptors only in 2017 (Figure 5).
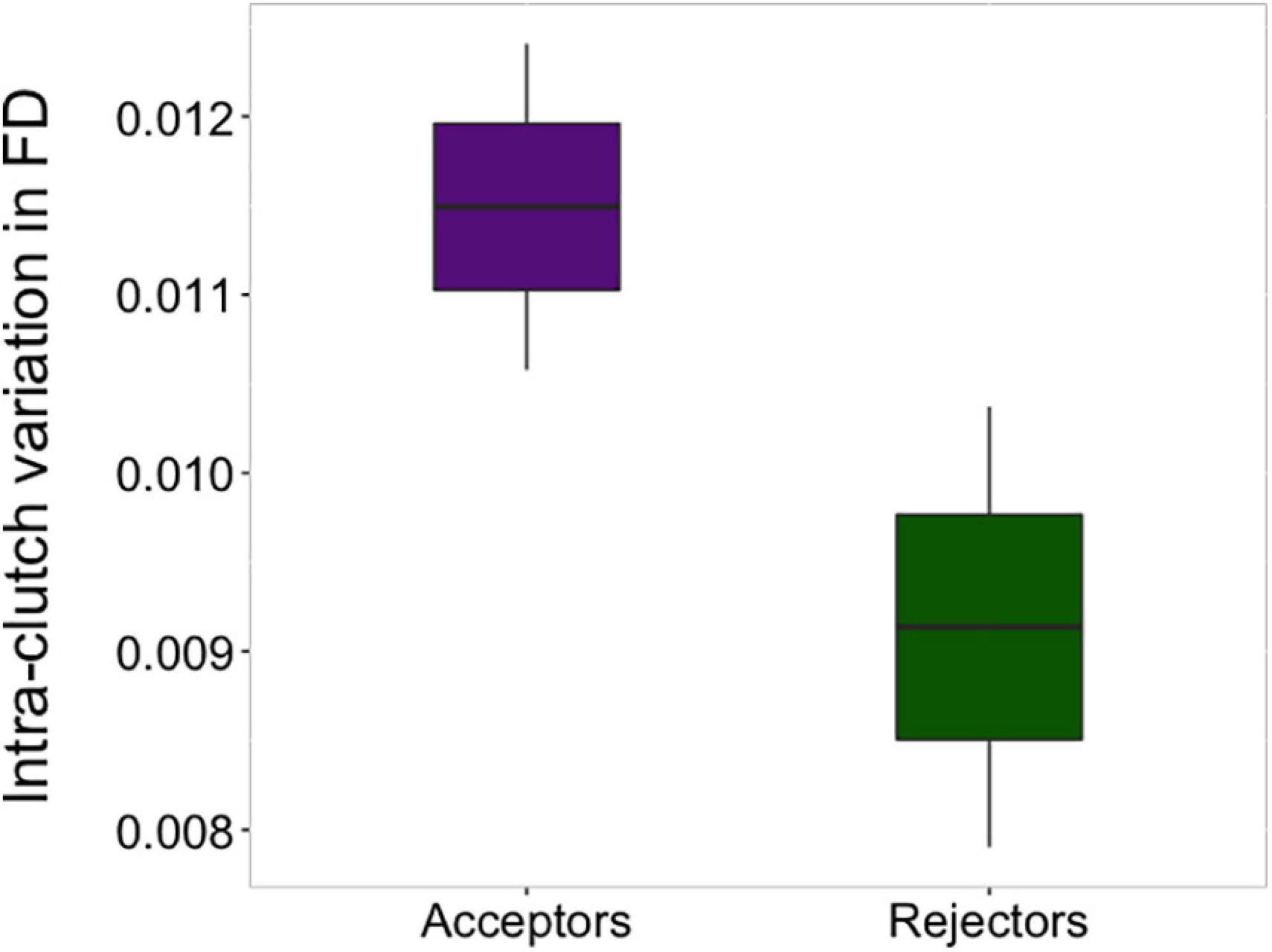
Figure 4. Differences in intra-clutch variation in the fractal dimension between acceptor and rejecter magpies. Horizontal line represents the mean value, boxes standard error and line bars that indicate confidence intervals (95%).
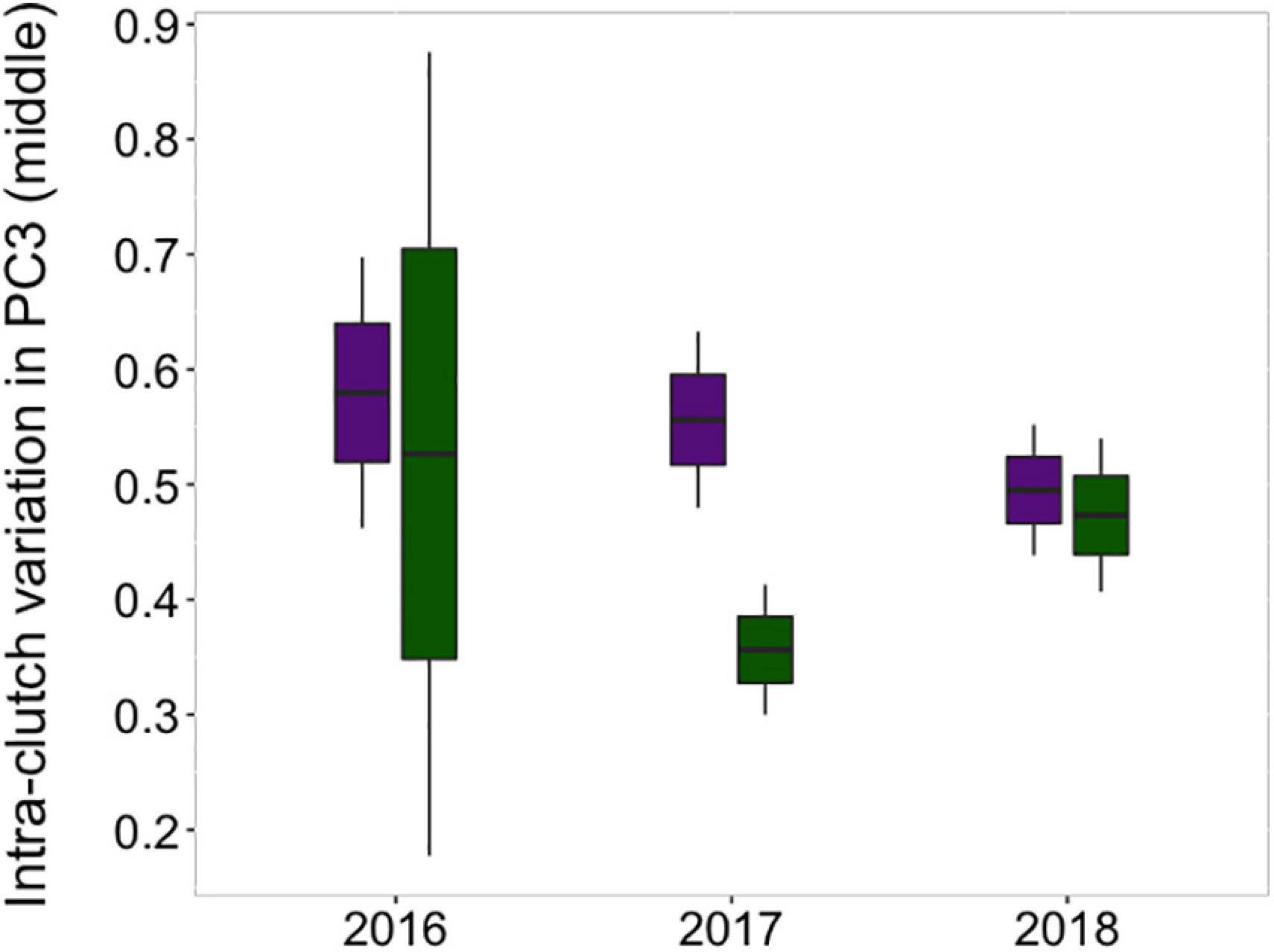
Figure 5. Differences in intra-clutch variation in blue-green egg coloration in the middle area of the eggs between acceptor and rejecter magpies in three different years. Green boxes correspond to rejecter individuals and purple ones correspond to acceptors.
Discussion
Our results show a clear, consistent relationship between intra-clutch variation in magpie egg phenotype and the probability of egg rejection by females. Moreover, we have demonstrated the importance of using different aspects of egg phenotype and of replicating the experiments over the years to better understand this association.
Victoria (1972) proposed that high homogeneity in the appearance of host eggs could be under strong selection in parasitized host populations because it would make easier the detection of noticeable features of parasite eggs by comparison of hosts own eggs with foreign ones (Øien et al., 1995; Soler and Møller, 1996). However, support for this hypothesis has been inconsistent, with studies showing opposite results (see section “Introduction”). Here we have analyzed whether intra-clutch variation in egg appearance changes and if this relates to host egg rejection in magpie hosts by considering variation in several traits and regions of host eggs. We have found that intra-clutch variation in egg appearance (volume, eggshell color, FD, see Supplementary Material) does not significantly differ between years but that the average values of some components of color and spottiness pattern varied, suggesting that variability of these eggshell features would have a strong environmental component. In addition, we also found that within-individual consistency differ for the different egg features, with brightness and patterns of spottiness being the features most typical of individuals. By using different host egg traits (size, color and Fractal Dimension) sampled across several years we have revealed that not all these traits show a consistent relationship with egg rejection, and that some of them have a higher potential to be subjected to selection pressures arising from rejection. In fact, it is intra-clutch variation in one of the more female-specific traits (patterns of spottiness as estimated by FD), which consistently affects egg rejection.
Our results confirm a key prediction of the co-evolutionary arms race hypothesis, namely that a low intra-clutch variation favors discrimination and rejection of foreign eggs in magpie for those traits that are more female-specific. Besides, these results are in accordance with a recent and innovative work using machine learning algorithms (as a proxy of a bird observer), which showed that birds may use primarily the most repeatable eggshell features (those with low intra-clutch variation) to recognize their own eggs and reject the foreign ones (Gómez et al., 2021). These findings stresses that the role of intra-clutch variation in rejection is contingent on the particular features of the egg phenotype considered, as some might show a higher degree of environmental determination whereas other are more female-specific.
Intra-clutch variation in blue-green color of the middle area of the eggs has an effect in the rejection behavior only one of the years, when rejecter females laid eggs more homogeneous in the blue-green coloring of their middle part. The importance of intra-clutch variation in blue-green coloration in the rejection of model eggs in magpies has been previously described (Avilés et al., 2004), although in that study higher intra-clutch variation in host egg appearance associated with rejection was found. The difference between the two studies may be due to several factors. First, it may be a consequence of methodological discrepancies, since we took measurements of the egg in three different areas, whereas the previous study considered the egg as a whole. Considering specific areas of the eggshell surface has been shown to be important in some studies; for example, both within clutch variation in the blunt pole (Spottiswoode and Stevens, 2010), or both egg poles (Polačiková et al., 2011). However, this possibility seems unlikely because in no case we found a negative association between homogeneity in blue-green color and rejection for the considered areas of the egg in this study. Secondly, Avilés et al. (2004) sampled the population only in one breeding season, whereas this study comprises three different breeding seasons. This is particularly important when host traits and behaviors may express plastically as it is known to happen for some host traits (Avilés et al., 2007; Hubbard et al., 2010; Molina-Morales et al., 2014; Berkowic et al., 2015; Martínez et al., 2020), and in particular for blue-green color of the eggshell (Soler et al., 2008). Alternatively, the inconsistency could be related with a different age structure in the population in different years. If intra-clutch variation covaries with female age, as it has been proposed for other species (Lotem et al., 1995; Siefferman et al., 2006), different age-structures in the population may render different rejection probabilities. Indeed, it has been shown that probability of rejection increases with age in magpies (Molina-Morales et al., 2014; Martínez et al., 2020). Although we do not know whether intra-clutch variation in blue-green color changes with age in magpies we have found that this feature was not repeatable. Another explanation is that differences in environmental conditions may favor or disfavor the ability to find food to obtain the pigmentation needed to color the eggs (Moreno and Osorno, 2003; Moreno et al., 2006; Morales et al., 2011). The intra-clutch variation in brightness is repeatable showing that this particular trait does not depend on environmental conditions.
In this work we have introduced a novel feature (in the context of foreign egg rejection experimental work) that we hypothesized can influence egg rejection in magpies, the fractal dimension of eggshell spottiness. The previous study that model host recognition used machine learning algorithms and characterized the patterning of spottiness using the FD of spottiness (Gómez et al., 2021), found that, among the several variables related with spottiness, the FD was the most important for recognition and almost the most repeatable within the clutch. Here, we have shown that this feature influences rejection in the same way in all the study years. Individuals that rejected the foreign eggs were those whose intra-clutch variation in fractal dimension was lower, which means that all the eggs within the clutch were similarly homogeneous. Fractal dimension characterizes the complexity of a spatial pattern in a given object or trait, and increases with surface complexity (Gómez et al., 2019). Fractal dimension of spottiness is indicative of the distribution of spots across the eggshell and is positive related with the amount of protoporphyrin (Gómez et al., 2019), the pigment that constitutes them. Previous studies have demonstrated that fractal dimension of coloration of the bib in partridges was related to body condition and immune responsiveness (Pérez-Rodríguez et al., 2013). Because intra-clutch variation in avian eggshell pigmentation has also been related to female quality (De Coster et al., 2013; Minias et al., 2020), it could be argued that rejecter females are individuals in a better condition or higher quality. In any case, our results agree with the expectation that the value of intra-clutch variation in determining egg discrimination should be larger for those traits, which are less environmentally driven.
Conclusion
Summing up, our work stresses the importance of considering a holistic approach including several aspects of egg phenotype and replication in studies on the adaptive value of host traits in egg rejection. Host may counteract parasite egg mimicry evolving very homogeneous eggs in such a way that intra-clutch variation in egg appearance is minimal.
Our results may suggest that this evolutionary process will not affect all egg traits with the same strength. Features such as color, or others, which may relate in their expression to the age and/or physical condition of the female at a given time, would be less likely used as cue for parasite egg recognition. However, the homogeneity of eggshell patterns (here quantified through FD) could have been favored by natural selection as a signature that facilitates the identification of own eggs. We must also acknowledge that the degree of difference between foreign and host eggs could affect the likelihood of rejection of foreign eggs. Nonetheless our study was based on rejection of mimetic models harboring little variation among them. Future experimental studies should ideally use real cuckoo eggs or models with variable levels of mimicry with host eggs to ascertain whether the role of intra-clutch variation in rejection is mediated by the degree of matching between parasite and host eggs. Plastic expression of the different aspects of the egg phenotype might, however, be species-specific or population-specific, and would need to be evaluated as a key premise in future tests of the intra-clutch variation hypothesis.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation. Also, a dataset is available at https://doi.org/10.6084/m9.figshare.16811494.v1.
Ethics Statement
The animal study was reviewed and approved by Comité Etica de la Universidad de Granada.
Author Contributions
MM-M and JA conceived the idea and analyzed the data. MM-M led the writing of the manuscript. MP and JGM performed the experiment in the field work. JG and GL-C carried out the images processing and performed the analyzes related to fractal dimension. All authors contributed critically to the drafts and gave final approval for publication.
Funding
This study was made possible through funding by the Spanish Ministry of Science through different projects (CGL2011-27561 and CGL2014-55362-P), a grant (BES-2015-075675), the Juan de la Cierva–Incorporación Fellowship (IJC2018-036411-I), and the Junta de Andalucía through a project (P06-RNM-01862).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Brian Peer for inviting us to contribute to this special issue. We are also indebted to Daniel Hanley and Vanina Dafne Fiorini for comments that greatly improved the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.702637/full#supplementary-material
References
Abernathy, V. E., and Peer, B. D. (2014). Intraclutch variation in egg appearance of Brown-headed Cowbird hosts. Auk 131, 467–475. doi: 10.1642/auk-12-186-r1.1
Avilés, J. M., Soler, J. J., Pérez-Contreras, T., Soler, M., and Møller, A. P. (2006). Ultraviolet reflectance of great spotted cuckoo eggs and egg discrimination by magpies. Behav. Ecol. 17, 310–314. doi: 10.1093/beheco/arj031
Avilés, J. M., Soler, J. J., Soler, M., and Møller, A. P. (2004). Rejection of parasitic eggs in relation to egg appearance in magpies. Anim. Behav. 67, 951–958. doi: 10.1016/j.anbehav.2003.08.022
Avilés, J. M., Stokke, B. G., Moksnes, A., Røskaft, E., and Møller, A. P. (2007). Environmental conditions influence egg color of reed warblers Acrocephalus scirpaceus and their parasite, the common cuckoo Cuculus canorus. Behav. Ecol. Sociobiol. 61, 475–485. doi: 10.1007/s00265-006-0275-0
Berkowic, D., Stokke, B. G., Meiri, S., and Markman, S. (2015). Climate change and coevolution in the cuckoo–reed warbler system. Evol. Ecol. 29, 581–597. doi: 10.1007/s10682-015-9763-x
Bradley, D., and Roth, G. (2007). Adaptive thresholding using the integral image. J. Graph. Tools 12, 13–21. doi: 10.1080/2151237x.2007.10129236
Cantarero, A., Naranjo, J. C., Casas, F., Mougeot, F., Viñuela, J., and Alonso-Alvarez, C. (2018). The fractal dimension of a conspicuous ornament varies with mating status and shows assortative mating in wild red-legged partridges (Alectoris rufa). Sci. Nat. 105:45.
Cherry, M. I., Bennett, A. T., and Moskát, C. (2007). Host intra-clutch variation, cuckoo egg matching and egg rejection by great reed warblers. Naturwissenschaften 94, 441–447. doi: 10.1007/s00114-007-0216-4
Cherry, M. I., and Bennett, A. T. D. (2001). Egg colour matching in an African cuckoo, as revealed by ultravioletevisible reflectance spectrophotometry. Proc. R. Soc. Lond. Ser. B 268, 565–571. doi: 10.1098/rspb.2000.1414
Croston, R., and Hauber, M. E. (2015). Experimental shifts in intraclutch egg color variation do not affect egg rejection in a host of a non-egg-mimetic avian brood parasite. PLoS One 10:e0121213. doi: 10.1371/journal.pone.0121213
Cuthill, I. C., Bennett, A. T. D., Partridge, J. C., and Maier, E. J. (1999). Plumage reflectance and the objective assessment of avian sexual dichromatism. Am. Nat. 153, 183–200. doi: 10.2307/2463580
De Coster, G., De Neve, L., and Lens, L. (2013). Intra-clutch variation in avian eggshell pigmentation covaries with female quality. J. Ornithol. 154, 1057–1065. doi: 10.1007/s10336-013-0974-z
de Hierro, M. D. G. L., and De Neve, L. (2010). Pigment limitation and female reproductive characteristics influence eggshell spottiness and ground colour variation in the house sparrow (Passer domesticus). J. Ornithol. 151, 833–840. doi: 10.1007/s10336-010-0520-1
Endler, J. A., and Thery, M. (1996). Interacting effects of lek placement, display behavior, ambient light, and color patterns in three neotropical forest-dwelling birds. Am. Nat. 148, 421–452. doi: 10.1086/285934
Geltsch, N., Moskát, C., Elek, Z., Bán, M., and Stevens, M. (2017). Egg spotting pattern in common cuckoos and their great reed warbler hosts: a century perspective. Biol. J. Linn. Soc. 121, 50–62. doi: 10.1093/biolinnean/blw035
Gómez, J., Gordo, O., and Minias, P. (2021). Egg recognition: the importance of quantifying multiple repeatable features as visual identity signals. PLoS One 16:e0248021. doi: 10.1371/journal.pone.0248021
Gómez, J., and Liñán-Cembrano, G. (2017). SpotEgg: an image-processing tool for automatised analysis of coloration and spottiness. J. Avian Biol. 48, 502–512. doi: 10.1111/jav.01117
Gómez, J., Liñán-Cembrano, G., Castro, M., Pérez-Hurtado, A., Ramo, C., and Amat, J. A. (2019). The protoporphyrin content of Kentish Plover Charadrius alexandrinus eggshells is better predicted by the fractal dimension of spottiness than by colour. J. Ornithol. 160, 409–415. doi: 10.1007/s10336-018-1616-2
Gosler, A. G., Barnett, P. R., and James Reynolds, S. (2000). Inheritance and variation in eggshell patterning in the great tit Parus major. Proc. R. Soc. Lond. Ser. B Biol. Sci. 267, 2469–2473.
Hoyt, D. F. (1979). Practical methods of estimating volume and fresh weight of bird eggs. Auk 96, 73–77.
Hubbard, J. K., Uy, J. A. C., Hauber, M. E., Hoekstra, H. E., and Safran, R. J. (2010). Vertebrate pigmentation: from underlying genes to adaptive function. Trends Genet. 26, 231–239. doi: 10.1016/j.tig.2010.02.002
Jovani, R., Pérez-Rodríguez, L., and Mougeot, F. (2013). Fractal geometry for animal biometrics: a response to Kühl and Burghardt. Trends Ecol. Evol. 9, 499–500. doi: 10.1016/j.tree.2013.06.004
Karcza, Z., Moskát, C., Cherry, M. I., and Kisbenedek, T. (2003). Experimental manipulation of intraclutch variation in the great reed warbler shows no effect on rejection of parasitic eggs. Ethology 109, 15–22. doi: 10.1046/j.1439-0310.2003.00839.x
Kelly, C. D. (2006). Replicating empirical research in behavioral ecology: how and why it should be done but rarely ever is. Q. Rev. Biol. 81, 221–236. doi: 10.1086/506236
Kelly, C. D. (2019). Rate and success of study replication in ecology and evolution. PeerJ 7:e7654. doi: 10.7717/peerj.7654
Landstrom, M., Heinsohn, R., and Langmore, N. (2010). Clutch variation and egg rejection in three hosts of the pallid cuckoo, Cuculus pallidus. Behaviour 147, 19–36. doi: 10.1163/000579509x12483520922043
Lotem, A., Nakamura, H., and Zahavi, A. (1995). Constraints on egg discrimination and cuckoo-host co-evolution. Anim. Behav. 49, 1185–1209. doi: 10.1006/anbe.1995.0152
Lyon, B. E. (2003). Egg recognition and counting reduce costs of avian conspecific brood parasitism. Nature 422, 495–499. doi: 10.1038/nature01505
Marchetti, K. (2000). Egg rejection in a passerine bird: size does matter. Anim. Behav. 59, 877–883. doi: 10.1006/anbe.1999.1388
Martínez, J. G., Molina-Morales, M., Precioso, M., and Avilés, J. M. (2020). Age-related brood parasitism and egg rejection in magpie hosts. Am. Nat. 195, 876–885. doi: 10.1086/708155
Minias, P., Gómez, J., and Włodarczyk, R. (2020). Egg spottiness reflects female condition, physiological stress, and ornament expression in a common rallid species. Auk 137:ukaa054.
Molina-Morales, M., Martínez, J. G., Martín-Gálvez, D., Dawson, D. A., Burke, T., and Avilés, J. M. (2014). Cuckoo hosts shift from accepting to rejecting parasitic eggs across their lifetime. Evolution 68, 3020–3029. doi: 10.1111/evo.12471
Molina-Morales, M., Martinez, J. G., Martín-Gálvez, D., Dawson, D. A., Rodríguez-Ruiz, J., Burke, T., et al. (2013). Evidence of long-term structured cuckoo parasitism on individual magpie hosts. J. Anim. Ecol. 82, 389–398. doi: 10.1111/1365-2656.12022
Morales, J., Velando, A., and Torres, R. (2011). Biliverdin-based egg coloration is enhanced by carotenoid supplementation. Behav. Ecol. Sociobiol. 65, 197–203. doi: 10.1007/s00265-010-1025-x
Moreno, J., Lobato, E., Morales, J., Merino, S., Tomás, G., Martínez-de la Puente, J., et al. (2006). Experimental evidence that egg color indicates female condition at laying in a songbird. Behav. Ecol. 17, 651–655. doi: 10.1093/beheco/ark014
Moreno, J., and Osorno, J. L. (2003). Avian egg colour and sexual selection: does eggshell pigmentation reflect female condition and genetic quality? Ecol. Lett. 6, 803–806. doi: 10.1046/j.1461-0248.2003.00505.x
Moskát, C., Avilés, J. A., Bán, M., Hargitai, R., and Zölei, A. (2008). Experimental support for the use of egg uniformity in para- site egg discrimination by cuckoo hosts. Behav. Ecol. Sociobiol. 62, 1885–1890. doi: 10.1007/s00265-008-0618-0
Moskát, C., and Lovászi, P. (2004). Break-down of arms race between the red-backed shrike (Lanius collurio) and common cuckoo(Cuculus canorus). Behaviour 141, 245–262. doi: 10.1163/156853904322890843
Nakagawa, S., and Parker, T. H. (2015). Replicating research in ecology and evolution: feasibility, incentives, and the cost-benefit conundrum. BMC Biol. 13:88. doi: 10.1186/s12915-015-0196-3
Øien, I. J., Moksnes, A., and Røskaft, E. (1995). Evolution of variation in egg color and marking pattern in European passerines: adaptations in a coevolutionary arms race with the cuckoo, Cuculus canorus. Behav. Ecol. 6, 166–174. doi: 10.1093/beheco/6.2.166
Peer, B. D., Rothstein, S. I., and McCleery, R. A. (2010). Intraclutch variation in egg appearance constrains rejection of Brown-headed cowbird (Molothrus ater) eggs in common grackles (Quiscalus quiscula). Auk 127, 759–764. doi: 10.1525/auk.2010.10057
Pérez-Rodríguez, L., Jovani, R., and Mougeot, F. (2013). Fractal geometry of a complex plumage trait reveals bird’s quality. Proc. R. Soc. B Biol. Sci. 280:20122783. doi: 10.1098/rspb.2012.2783
Polačiková, L., Hauber, M. E., Procházka, P., Cassey, P., Honza, M., and Grim, T. (2011). A sum of its individual parts? Relative contributions of different eggshell regions to intraclutch variation in birds. J. Avian Biol. 42, 370–373. doi: 10.1111/j.1600-048x.2011.05202.x
Procházka, P., and Honza, M. (2003). Do common whitethroats (Sylvia communis) discriminate against alien eggs? J. Ornithol. 144, 354–363. doi: 10.1007/bf02465635
Rothstein, S. I. (1982). Mechanisms of avian egg recognition: Which egg parameters elicit responses by rejecter species?. Behav. Ecol. Sociobiol. 11, 229–239. doi: 10.1007/BF00299299
Rothstein, S. I. (1990). A model system for coevolution: avian brood parasitism. Annu. Rev. Ecol. Syst. 21, 481–508. doi: 10.1146/annurev.es.21.110190.002405
Schroeder, M. R. (1991). Minutes from an Infinite Universe: Fractals, Chaos, Power Laws. New York, NY: WH Freeman.
Siefferman, L., Navara, K. J., and Hill, G. E. (2006). Egg coloration is correlated with female condition in eastern bluebirds (Sialia sialis). Behav. Ecol. Sociobiol. 59, 651–656. doi: 10.1007/s00265-005-0092-x
Soler, J. J., Martinez, J. G., Soler, M., and Møller, A. P. (1999). Genetic and geographic variation in rejection behavior of cuckoo eggs by European magpie populations: an experimental test of rejecter-gene flow. Evolution 53, 947–956.
Soler, J. J., and Møller, A. P. (1996). A comparative analysis of the evolution of variation in appearance of eggs of European passerines in relation to brood parasitism. Behav. Ecol. 7, 89–94. doi: 10.1093/beheco/7.1.89
Soler, J. J., Navarro, C., Contreras, T. P., Avilés, J. M., and Cuervo, J. J. (2008). Sexually selected egg coloration in spotless starlings. Am. Nat. 171, 183–194. doi: 10.1086/524958
Soler, J. J., Soler, M., and Møller, A. P. (2000). Host recognition of parasite eggs and the physical appearance of host eggs: the magpie and its brood parasite the great spotted cuckoo. Etología 8, 9–16.
Soler, M., Martinez, J. G., and Soler, J. J. (1996). Effects of brood parasitism by great spotted cuckoo on the breeding success of the magpie hosts: an experimental study. Ardeola 43, 87–96.
Soler, M., and Møller, A. P. (1990). Duration of sympatry and coevolution between the great spotted cuckoo and its magpie host. Nature 343, 748–750. doi: 10.1038/343748a0
Soler, M., Soler, J. J., Martinez, J. G., and Møller, A. P. (1995). Magpie host manipulation by great spotted cuckoos: evidence for an avian mafia? Evolution 49, 770–775.
Spottiswoode, C. N., and Stevens, M. (2010). Visual modeling shows that avian host parents use multiple visual cues in rejecting parasitic eggs. Proc. Natl. Acad. Sci. U.S.A. 107, 8672–8676. doi: 10.1073/pnas.0910486107
Stokke, B. G., Moksnes, A., Røskaft, E., Rudolfsen, S., and Honza, M. (1999). Rejection of artificial cuckoo (Cuculus canorus) eggs in relation to variation in egg appearance among reed warblers (Acrocephalus scirpaceus). Proc. R. Soc. Lond. Ser. B Biol. Sci. 266, 1483–1488. doi: 10.1098/rspb.1999.0804
Stokke, B. G., Rudolfsen, G., Moksnes, A., and Røskaft, E. (2004). Rejection of conspecific eggs in chaffinches: the effect of age and clutch characteristics. Ethology 110, 459–470. doi: 10.1111/j.1439-0310.2004.00978.x
Thorley, J. B., and Lord, A. M. (2015). Laying date is a plastic and repeatable trait in a population of Blue Tits Cyanistes caeruleus. Ardea 103, 69–78. doi: 10.5253/arde.v103i1.a7
Victoria, J. K. (1972). Clutch characteristics and egg discriminative ability of the African village weaverbird Ploceus cucullatus. Ibis 114, 367–376. doi: 10.1111/j.1474-919X.1972.tb00834.x
Wang, L., Liang, W., Yang, C., Cheng, S. J., Hsu, Y. C., and Lu, X. (2016). Egg rejection and clutch phenotype variation in the plain prinia Prinia inornata. J. Avian Biol. 47, 788–794. doi: 10.1111/jav.00786
Keywords: brood parasitism, color, fractal dimensions, repeatability, egg phenotype and spottiness
Citation: Molina-Morales M, Gómez J, Liñán-Cembrano G, Precioso M, Martínez JG and Avilés JM (2021) The Role of Intra-Clutch Variation of Magpie Clutches in Foreign Egg Rejection Depends on the Egg Trait Considered. Front. Ecol. Evol. 9:702637. doi: 10.3389/fevo.2021.702637
Received: 29 April 2021; Accepted: 07 October 2021;
Published: 28 October 2021.
Edited by:
Cynthia Ursino, Princeton University, United StatesReviewed by:
Daniel Hanley, George Mason University, United StatesVanina Dafne Fiorini, University of Buenos Aires, Argentina
Copyright © 2021 Molina-Morales, Gómez, Liñán-Cembrano, Precioso, Martínez and Avilés. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mercedes Molina-Morales, bWVyY2hlQHVnci5lcw==