 Pavol Prokop
Pavol Prokop Martina Zvaríková1
Martina Zvaríková1- 1Department of Environmental Ecology and Landscape Management, Faculty of Natural Sciences, Comenius University, Bratislava, Slovakia
- 2Institute of Zoology, Slovak Academy of Sciences, Bratislava, Slovakia
- 3Department of Nuclear Physics and Biophysics, Faculty of Mathematics, Physics and Informatics, Comenius University, Bratislava, Slovakia
- 4Department of Psychology, University of South Carolina Aiken, Aiken, SC, United States
Human–animal relationships have long been the subject of controversy because they are shaped by several cultural, inter-individual, and evolutionary factors. Understanding these relationships, however, is important to optimize conservation efforts. There is agreement that perceived similarity between animals and humans is associated with more positive attitudes. The human–animal similarity could be caused by phylogenetic closeness. We hypothesized that the bipedal posture of an animal may be perceived as a cue of phenotypic similarity with humans. We examined this topic by comparing perceived cuteness, fear, and willingness to protect animals differing in body posture, body size, and phylogenetic closeness with humans on a sample of N = 349 Slovak participants. We found that the bipedal posture enhanced perceived cuteness, but this effect was most pronounced in small-bodied animals, particularly those with direct eye contact. Phylogenetically close and small-bodied species (e.g., small mammals) received greater conservation support than phylogenetically distant species (e.g., invertebrates). However, anthropomorphic-looking animals received greater conservation support, suggesting that pictures of animals that more closely resemble humans can be used in conservation campaigns.
Introduction
We are not seeing animals as animals, but merely as reflections of ourselves, and if the mirror distorts too badly, we either bend it into shape or discard it.
Morris (1969, p. 200)
The complexity of human–animal relationships is the subject of controversy (Mullin, 1999) because animal roles in human lives are multidimensional (Kellert, 1980, 1983, 1993; Serpell, 2004). Throughout our evolutionary history, animals have been predators of our ancestors (Hart and Sussman, 2008), important sources of food (Ungar and Teaford, 2002), and vectors of zoonotic diseases (Kruse et al., 2004). Since the beginning of the Neolithic era, many animals have been treated as companions (Podberscek et al., 2005), with others being agricultural pests (Ordish, 1976). The enormous diversity of animal shapes, sizes, and behaviors heavily contributes to human evaluations of them (Serpell, 2004). Human preferences are of interest to conservationists because the low popularity of an animal, regardless of why it occurs, erodes public support for its conservation (Kellert, 1985; Houston et al., 2010; MacFarlane and Rocha, 2020).
Human activities contribute to the global loss of biodiversity comparable with the five previous mass extinctions of Earth’s history (Barnosky et al., 2011; Dirzo et al., 2014). More than 400 vertebrates became extinct since 1500 (IUCN, 2020). Although invertebrates received much less attention than vertebrates, their extinction rates seem to be similar. For instance, insect abundance in protected natural areas in Germany dropped by 75% over the last 27 years (Hallmann et al., 2017), and roughly 50% of butterfly species (Lepidoptera) were extirpated in Singapore since 1854 (Theng et al., 2020). The reduction of biodiversity can negatively influence ecosystem functioning and human wellbeing by means of disease spread, climate change, and decreased farm productivity (Newbold et al., 2019; van der Plas, 2019). Because all animals play important roles in ecosystems, it is necessary to investigate the factors underlying their preferences by the general public in order to make conservation more effective (Frynta et al., 2013).
The willingness of people to protect animals is influenced by a complex interplay between emotions and attitudes toward them (Castillo-Huitrón et al., 2020), resulting in non-random preferences for certain species (Driscoll, 1995; Bjerke and Østdahl, 2004; Frynta et al., 2013; Borgi and Cirulli, 2015; Prokop and Randler, 2018). Humans prioritize esthetically appealing animals (Serpell, 2004) characterized by bright colors (Marešová et al., 2009; Barua et al., 2012; Prokop and Fančovičová, 2013; Curtin and Papworth, 2020), large body size (Frynta et al., 2010; Knegtering et al., 2010; Smith et al., 2012; Macdonald et al., 2015; Curtin and Papworth, 2020), and with a non-aggressive appearance (Prokop and Fančovičová, 2017). Furthermore, these preferences are similar across genders (Prokop and Fančovičová, 2013).
Preferences for some animals over others are noteworthy because the perceived attractiveness of an animal species can be an important determinant of conservation support (Gunnthorsdottir, 2001; Tisdell et al., 2006; Martín-López et al., 2007; Knight, 2008; Wang et al., 2018; Curtin and Papworth, 2020). For instance, more popular species at the Paris Zoological Park can receive 46 times the funds of less popular species (Colléony et al., 2017).
Animals that are cognitively, behaviorally, physically, or phylogenetically similar to humans are perceived by humans in a more positive light than distant or dissimilar animals (Plous, 1993; Allen et al., 2002; Knight et al., 2003; Serpell, 2004). Humans express increased empathy and compassion with decreasing phylogenetic distance from humans (Miralles et al., 2019). It has been shown that our perception of “the others” may depend on the group to which we perceive that “the others” belong, that is, the more we perceive the members of the other person/group in need as being similar to us, the more we value their welfare (e.g., Batson, 2011). Małecki et al. (2020) found that the subjective perception of the level of kinship between humans and animals depicted in a narration had a significant influence on the improvement of attitudes toward animal welfare. However, the objective measure of the phylogenetic distance was not related to any changes in attitudes toward animal welfare.
Another largely overlooked cue of species similarity with humans is bipedal posture. Morris (1969) argues that the popularity of penguins, primates, bears, pandas, or dogs is significantly influenced by their ability to stand vertically because animals in bipedal postures resemble humans more than in quadrupedal postures. As far as we are aware, no research has investigated the degree to which the bipedal posture of an animal influences human perceptions and attitudes. We postulate that perceived cuteness, fear of animals, and willingness to protect animals would be worthwhile to investigate because these variables are related to the psychological aspects of animal conservation (Prokop and Fančovičová, 2013; Castillo-Huitrón et al., 2020).
We hypothesized that animals with bipedal posture would be perceived as cuter (and thus more likely to induce human willingness to protect them) than animals with quadrupedal posture. We also hypothesized that large animals would be perceived as more dangerous than small animals (Prokop et al., 2010; Staňková et al., 2021). Thus, the perceived cuteness of small animals in bipedal posture should be greater than the perceived cuteness of large animals in bipedal posture. Furthermore, we hypothesized that phylogenetically distant animals would be perceived as less cute than phylogenetically close animals (Miralles et al., 2019). Finally, both bipedal posture and phylogenetical closeness should be associated with a greater willingness to protect animals.
Materials and Methods
Participants
The research was carried out during the summer semester of 2020. Roughly 1,500 freshman students at a university in Slovakia were asked to voluntarily participate in the research. Furthermore, the participants were recruited through social networks. A total of 349 participants (272 females) comprised the final sample (age range = 18–67 years, mean = 25, SE = 0.45; Table 1).
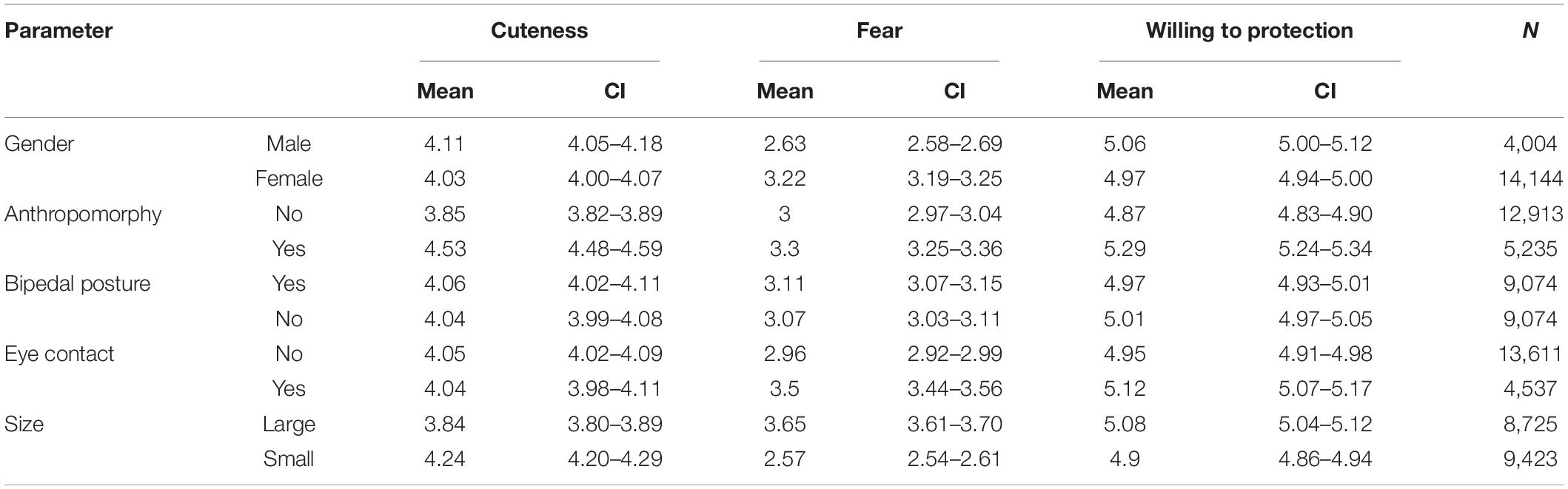
Table 1. Descriptive statistics for cuteness, fear, and willingness to protection ratings of animals.
Measurement of Cuteness, Perceived Fear, and Willingness to Protect Animals
The participants were asked to rate 52 pictures of 26 animal species for cuteness, perceived fear, and willingness to protect these animals on a seven-point scale (e.g., 1 = not at all cute, 7 = extremely cute). The ratings showed excellent reliabilities (Cronbach α = 0.97, 0.97, and 0.98, respectively).
Measurement of Phylogenetic Distance From Humans
The phylogenetic divergence time from humans (in millions of years) was obtained for each species from timetree.org (Kumar et al., 2017).
Measurement of Direct Eye Contact
Animals on pictures were scored binomially (yes or no) according to direct or indirect eye contact. This procedure was used because it was not possible to obtain pictures of all animals with a standard gaze. This variable could be important because direct eye contact is a signal of potential threat (Emery, 2000).
Measurement of Anthropomorphism
Animals which looked like humans by their posture or activity (e.g., a basilisk running on the surface of water) were binomially classified as anthropomorphic or not. Two of us separately and independently scored the pictures for anthropomorphism. In the few cases where our scorings differed, we discussed the pictures of animals until we agreed on the score to be awarded. Anthropomorphism, in this sense, was not always associated with bipedal posture (e.g., standing Komodo dragon), and we therefore considered it as an additional independent variable.
Visual Stimuli
Colorful pictures, with their original background, of 14 vertebrate and 12 invertebrate species which at least occasionally use bipedal posture were downloaded from Google. The species were further selected according to their phylogenetical similarity and body size (Table 2). We used two pictures for each species: one in bipedal posture and one in quadrupedal posture. Throughout the paper, we used the term quadrupedal also for invertebrates with more than four legs. For invertebrates, pictures of bipedal animals were searched using common English or Latin names together with the word “bipedal” or “standing.” Birds were completely omitted because they are invariably bipedal.
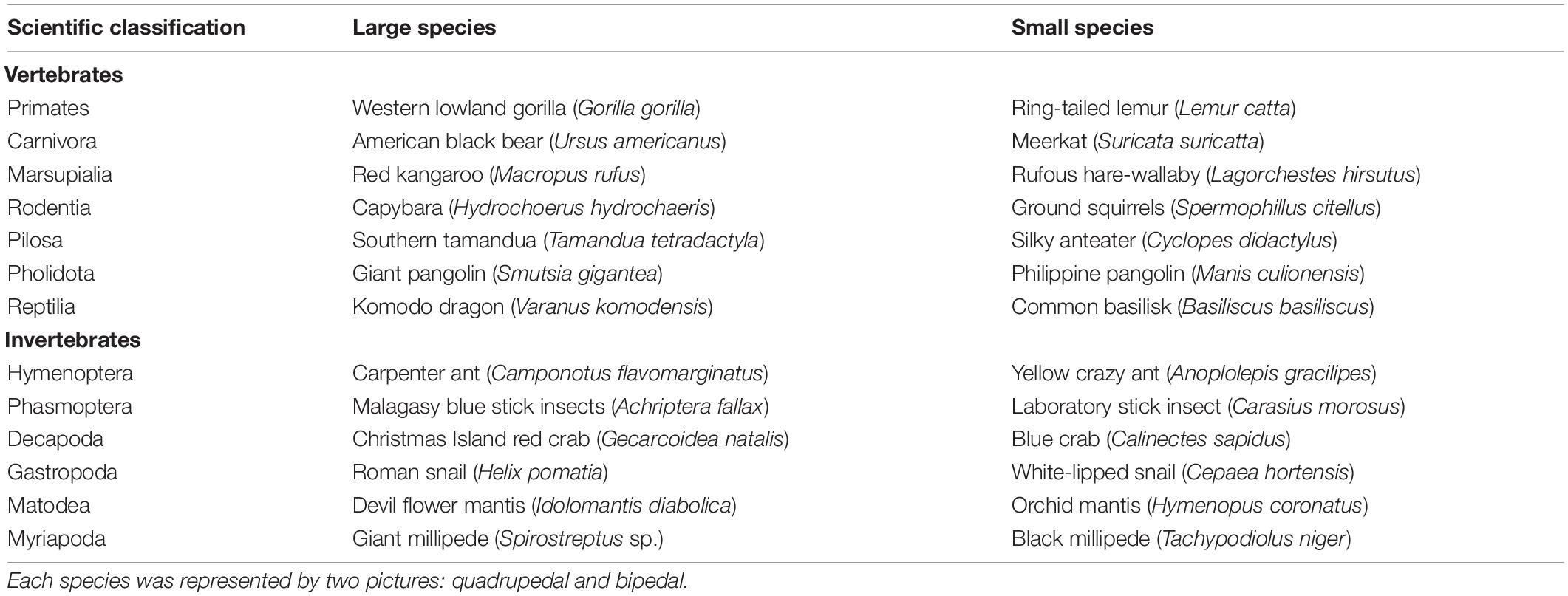
Table 2. List of species used in the research.
Statistical Analyses
The cuteness, fear, and protection scores were defined as dependent variables in a cumulative link mixed model (CLMM) in R software 3.6.3 (R Core Team, 2019) by applying the clmm function in an ordinal package (Christensen, 2019). Bipedal posture (bipedal or quadrupedal), anthropomorphic (yes or no), eye contact (yes or no), body size (large or small), and gender of the participant were categorical predictors, whereas phylogeny was a continuous predictor (fixed effects). The identity of the participants and the species of animal in the pictures were used as a grouping factor (random effects) to deal with correlations within the participant ratings. To select the particular factor, stepwise selection was used and evaluated by the Akaike information criterion (AIC). The syntax of the final CLMM models are as follows:
Cute.score ∼ Phylogeny + Anthropomorphy + Eye.contact + Biped + Gender + Size + Phylogeny:Gender + Phylogeny: Biped + Anthropomorphy:Eye.contact + Size:Gender + Size: Phylogeny + Eye.contact:Size + Anthropomorphy:Size + Size: Biped + (1 | ID.participant) + (1 | ID.species).
Protection ∼ Phylogeny + Anthropomorphy + Biped + Size + Size:Biped + Anthropomorphy:Phylogeny + (1| ID respondent) + (1| ID.species).
Fear ∼ Phylogeny + Anthropomorphy + Eye contact + Biped + Gender + Size + Phylogeny:Size + Phylogeny: Gender + Eye contact:Biped + Size:Gender + Biped:Phylogeny + (1| ID.respondent) + (1| ID.species).
Results
The best CLMM models for cuteness and fear ratings contain six fixed effects (phylogeny, anthropomorphy, bipedal posture, size, eye contact, and gender) and numerous interactive effects (eight for cuteness and five for fear; Tables 3, 4). In the case of the protection rating model, four fixed factors and two interactions were included (Table 5). The final CLMM model values of log likelihood ratio, AIC, and marginal, conditional, and pseudo r-squared are shown in Table 6. The likelihood ratio test showed the random effects, which have been included in all the CLMM models, to be significant (p < 0.001).
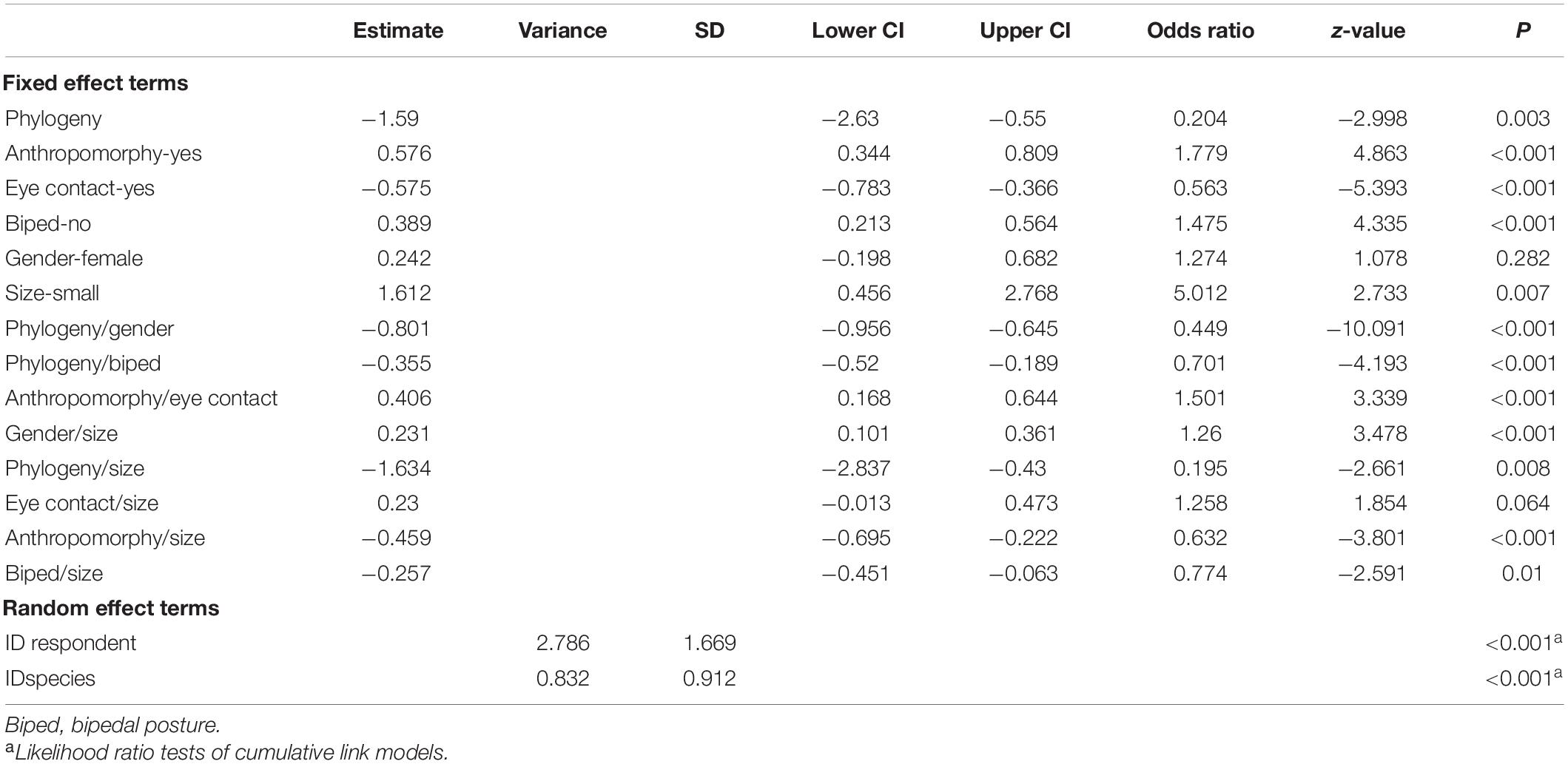
Table 3. Results of the cumulative link mixed model on the respondent’s cuteness ratings.
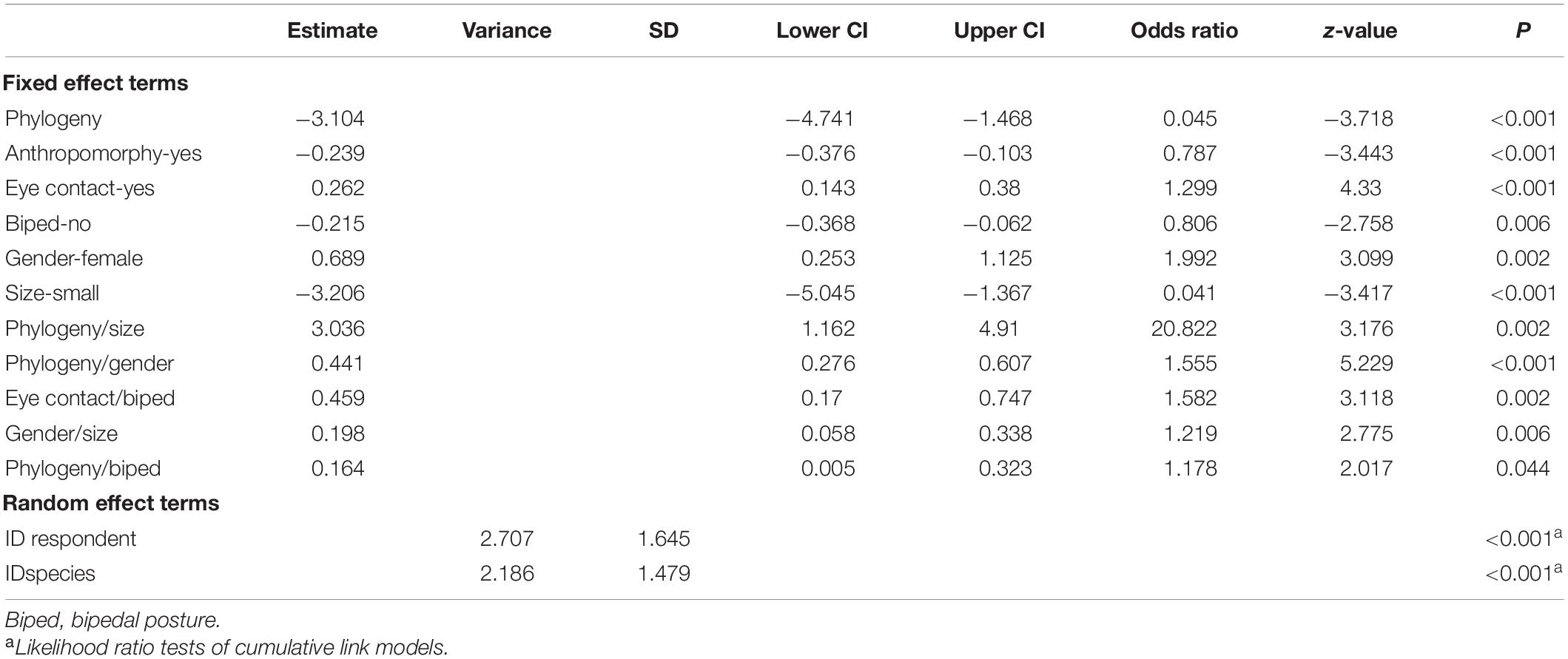
Table 4. Results of the cumulative link mixed model on the respondent’s fear ratings.
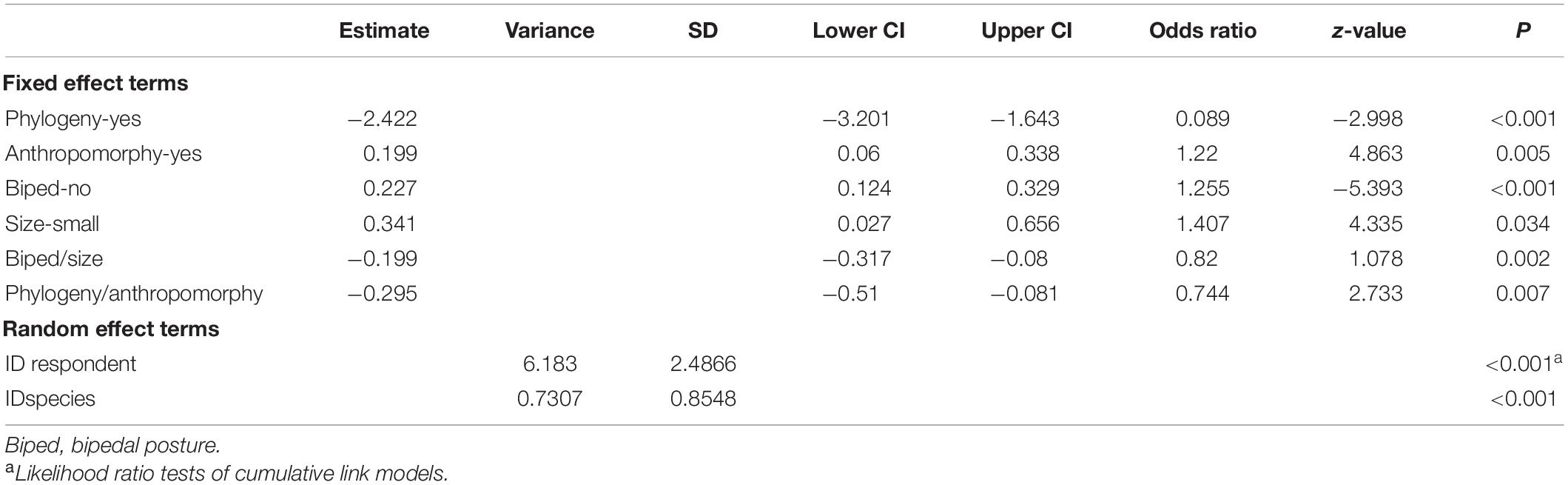
Table 5. Results of the cumulative link mixed model on the respondent’s willing to protection ratings.
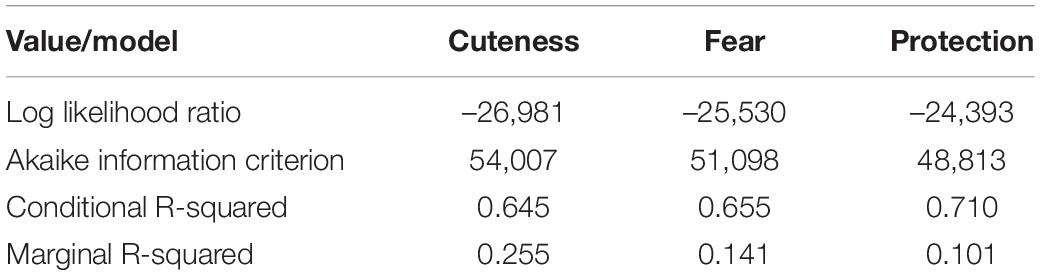
Table 6. Log Likelihood-ratio, Akaike information criterion, marginal, and conditional r-squared values of cumulative link mixed models.
Bipedal Posture
Animals with bipedal posture were rated as significantly less cute than animals with quadrupedal posture (Figure 1A). However, the combination of bipedal posture with other variables increased the perceived cuteness to a greater degree—for example, bipedals that are small in size were considered to be significantly cuter than larger ones (Figure 2A). Additionally, high cuteness scores were specifically associated with animals in bipedal posture that are phylogenetically closer to humans (e.g., bipedal mammals; Table 3).
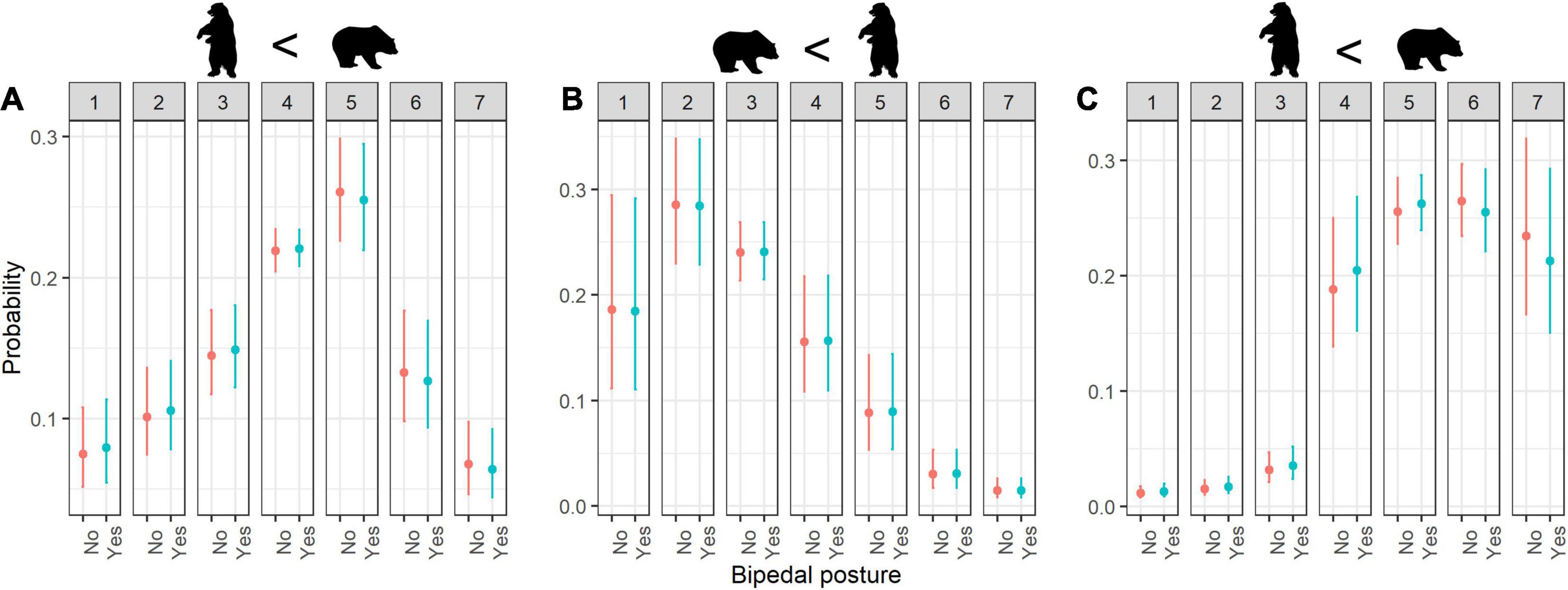
Figure 1. Estimated probability of ratings for (A) cuteness, (B) fear, and (C) willingness to protection according to Bipedal posture. Error bars indicate 95% confidence interval; numbers 1–7 refer to rating scores.
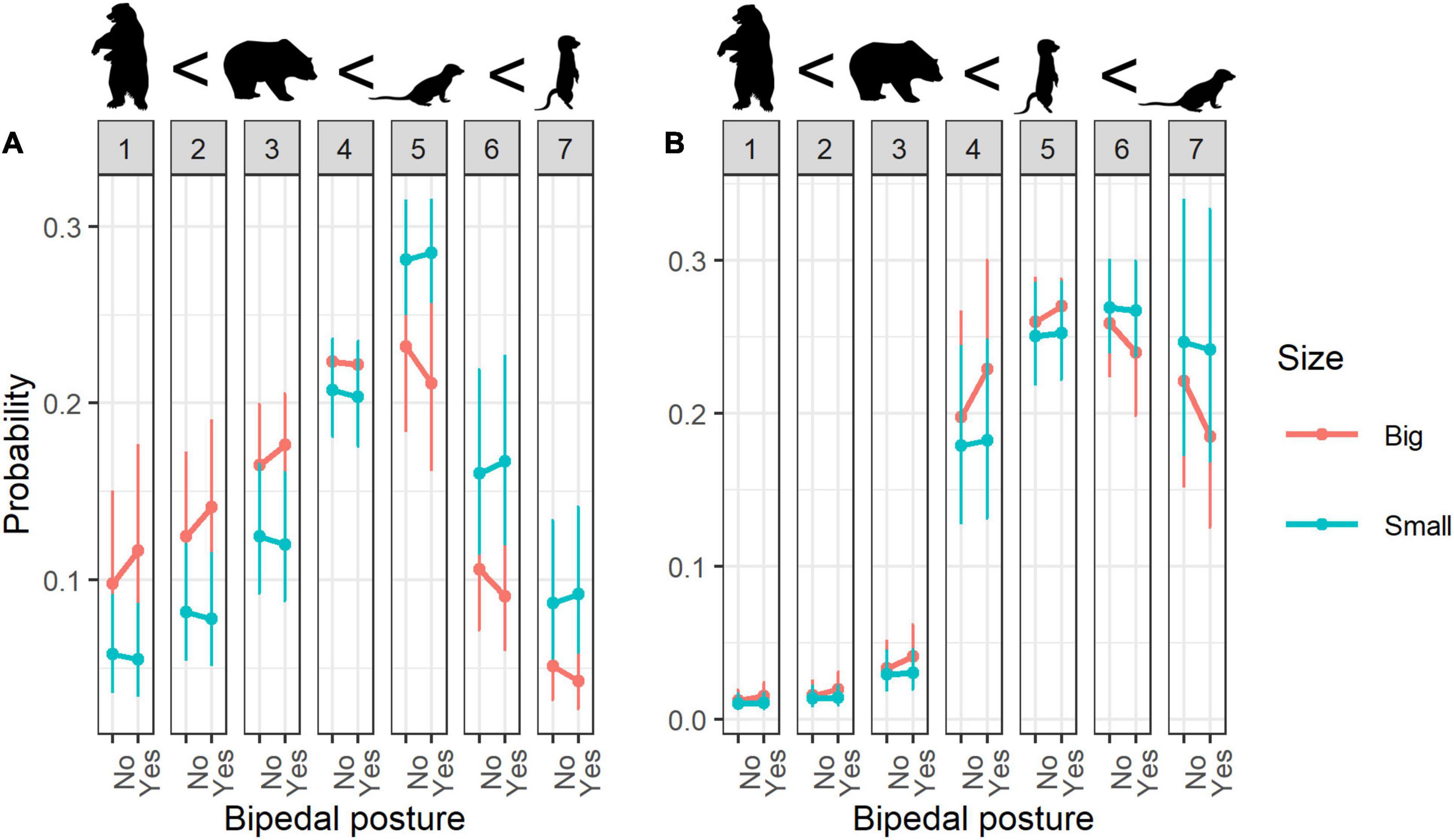
Figure 2. Interaction effects of Bipedal posture:Size for (A) cuteness, (B) willingness to protection. Error bars indicate 95% confidence interval; numbers 1–7 refer to rating scores.
Animals with bipedal posture received higher fear scores than animals with quadrupedal posture (Figure 1B). Interactive effects revealed that quadrupedal animals with direct eye contact were considered more menacing than other combinations of these factors (Table 4). Higher fear scores were associated with animals in bipedal posture that are phylogenetically close to humans, such as bipedal mammals (Table 4).
When considering the factor of bipedalism regarding willingness to protect, people were generally more willing to protect quadrupedal animals than animals with bipedal posture (Figure 1C). Animal size moderated this effect, such that willingness to protect was strongest toward small animals in bipedal posture (Figure 2B).
Phylogeny
The factor of phylogeny was very strong when assessing all dependent variables (cuteness, fear, and willingness to protect; Tables 3–5). Animals phylogenetically closer to humans (e.g., mammals) were perceived as cuter than phylogenetically distant animals (e.g., insects; Figure 3A). Small animals phylogenetically distant from human (e.g., small insects) received lower cuteness scores (Figure 3B). When rated by women, phylogenetically distant animals received significantly lower cuteness scores relative to the ratings provided by men (Table 3).
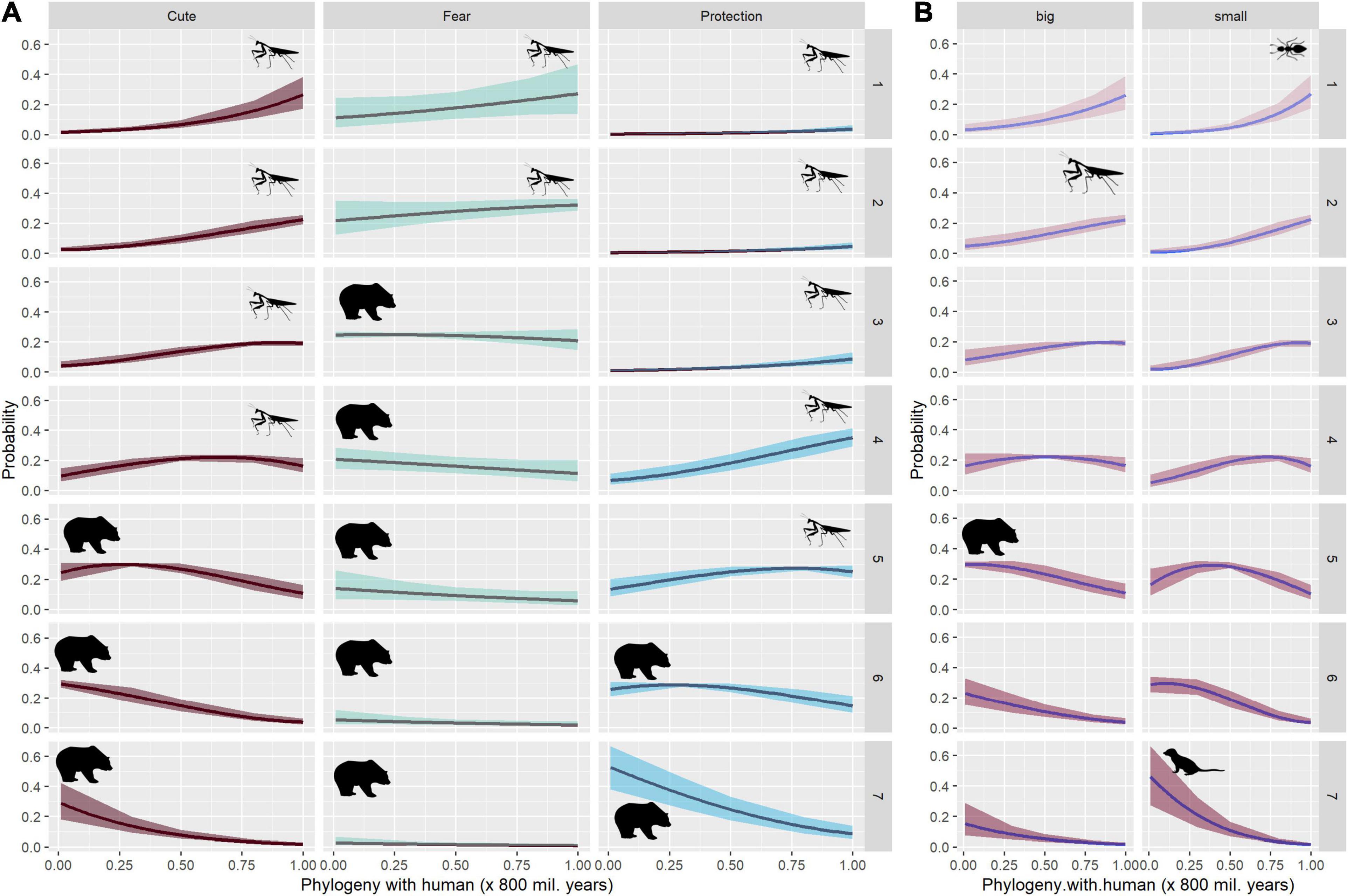
Figure 3. Estimated probability of ratings for cuteness, fear and willingness to protection according to Phylogeny (A), interaction effects of Phylogeny:Size for Cuteness (B). Error areas indicate 95% confidence interval; numbers 1–7 refer to rating scores.
Phylogenetically close animals were associated with higher fear scores when compared to phylogenetically distant animals (Figure 3A). The interaction terms showed that the fear scores were highest when considering animals phylogenetically close to humans and displaying a bipedal posture (e.g., mammals in bipedal posture; Table 4).
Phylogeny significantly affected the willingness to protect animals. People were willing to protect animals phylogenetically close to humans, such as mammals (Figure 3A), especially when it comes to small mammals (Table 5). The lowest protection scores were for animals that are phylogenetically distant from humans but looked anthropomorphic (e.g., insect with raised front legs; Table 5).
Anthropomorphism
Animals resembling humans in terms of posture or activity (e.g., a bipedally running reptile or standing gorilla) were perceived as cute (Figure 4A). Moreover, small animals resembling humans were evaluated as cuter than large anthropomorphic animals (Table 3). High cuteness scores were assigned to small anthropomorphic-looking animals with direct eye contact (Table 3).
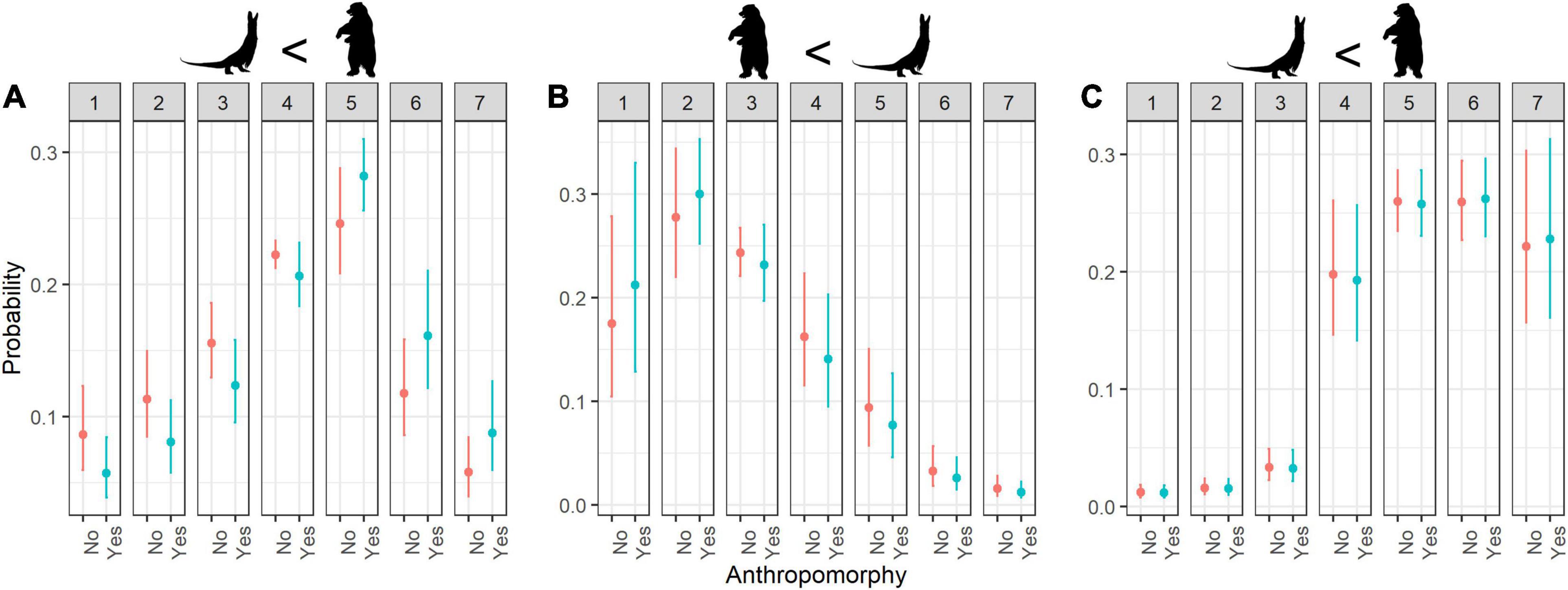
Figure 4. Estimated probability of ratings for (A) cuteness, (B) fear, and (C) willingness to protection according to Anthropomorphy. Error bars indicate 95% confidence interval; numbers 1–7 refer to rating scores.
In general, the respondents were not afraid of animals that looked anthropomorphic (lower fear scores; Figure 4B) and were willing to protect them (anthropomorphism increases the willingness to protect such animals; Figure 4C), but when considering the interaction with phylogeny, the willingness to protect decreased toward anthropomorphically looking animals that are phylogenetically less related to humans (e.g., insect with raised front legs; Table 5).
Size
Cuteness scores strongly depended on whether the animal is large or small and are biased in favor of small creatures (Figure 5A). In general, small animals that are phylogenetically closely related to humans were perceived as cuter than distant animals (Figure 3B), while small animals resembling humans with direct eye contact were rated cuter than large animals (Table 3). Women had a tendency to give higher cuteness ratings when considering small animals (Figure 6A).
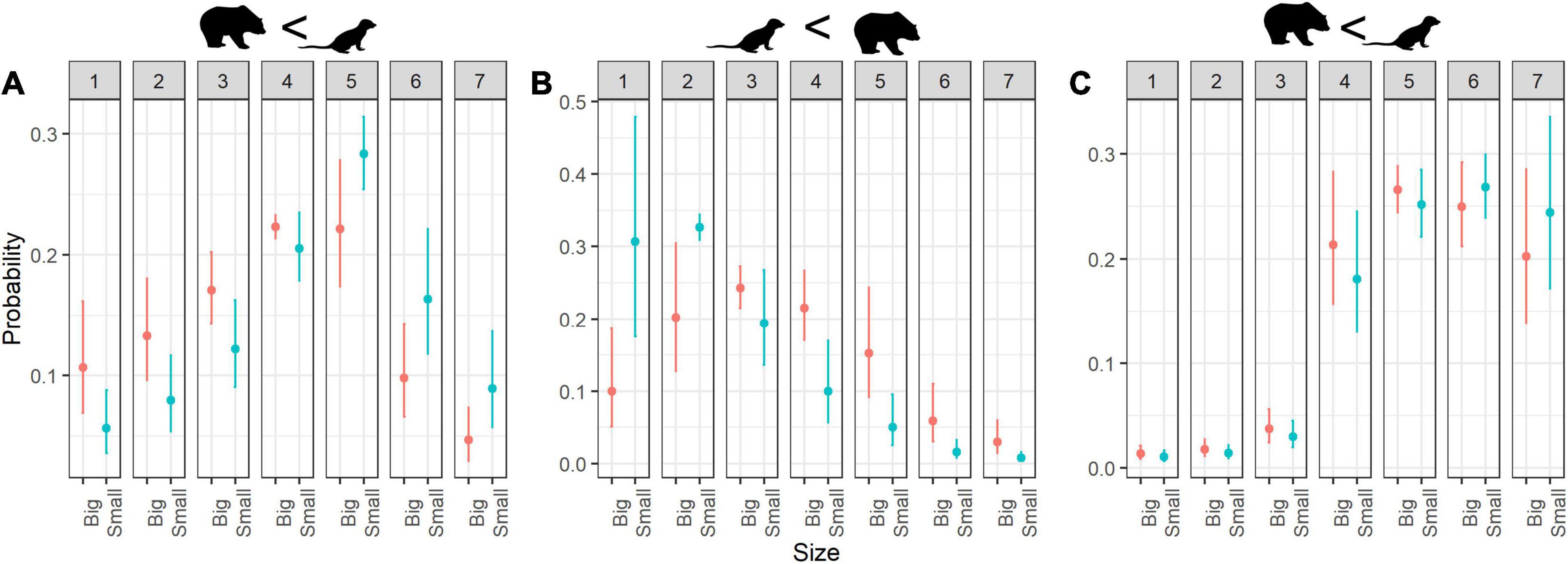
Figure 5. Estimated probability of ratings for (A) cuteness, (B) fear and (C) willingness to protection according to Size. Error bars indicate 95% confidence interval; numbers 1–7 refer to rating scores.

Figure 6. Interaction effects of Gender:Size for (A) cuteness and (B) fear. Error bars indicate 95% confidence interval; numbers 1–7 refer to rating scores.
As we expected, size modified the fear score as well. Small animals were rated as less menacing when compared to larger ones (Figures 5B, 6B).
Our results showed that people were willing to protect small animals (Figure 5C) and that small quadrupedals were evaluated with slightly higher protection scores than small bipedals (Figure 2B).
Eye Contact
When considering animals with eye contact, they were evaluated as less cute in comparison with no eye contact (Figure 7A), even though small anthropomorphic animals with eye contact were rated as cute (Table 3).
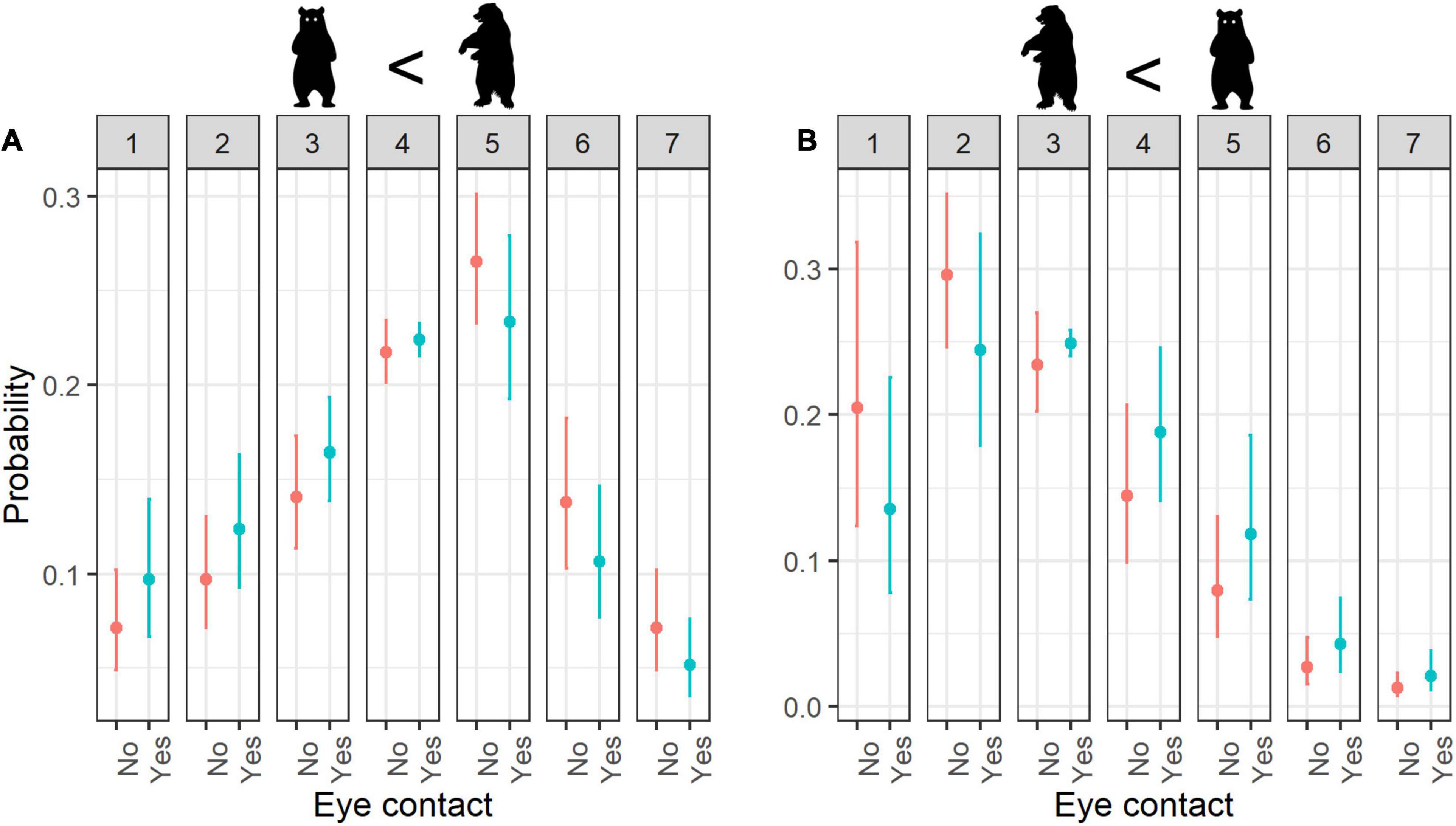
Figure 7. Estimated probability of ratings for (A) cuteness and (B) fear according to Eye contact. Error bars indicate 95% confidence interval; numbers 1–7 refer to rating scores.
With regard to fear, eye contact increased the fear scores (Figure 7B), especially together with quadrupedalism (Table 4).
Eye contact was not statistically significant for willingness to protect as an individual factor nor for interactions (Table 5).
Gender
Even though gender was not statistically significant as an individual factor for the cuteness evaluation, the interactions with other factors were significant (Table 3). As we have mentioned, animals that are phylogenetically distant to humans (e.g., insects) were rated as less cute than phylogenetically close animals (e.g., mammals; Figure 3A), but the cuteness scores decreased more when they were evaluated by women (Figure 8A). Small animals were assigned higher cuteness scores when rated by women compared to the ratings of men (Figure 6A).
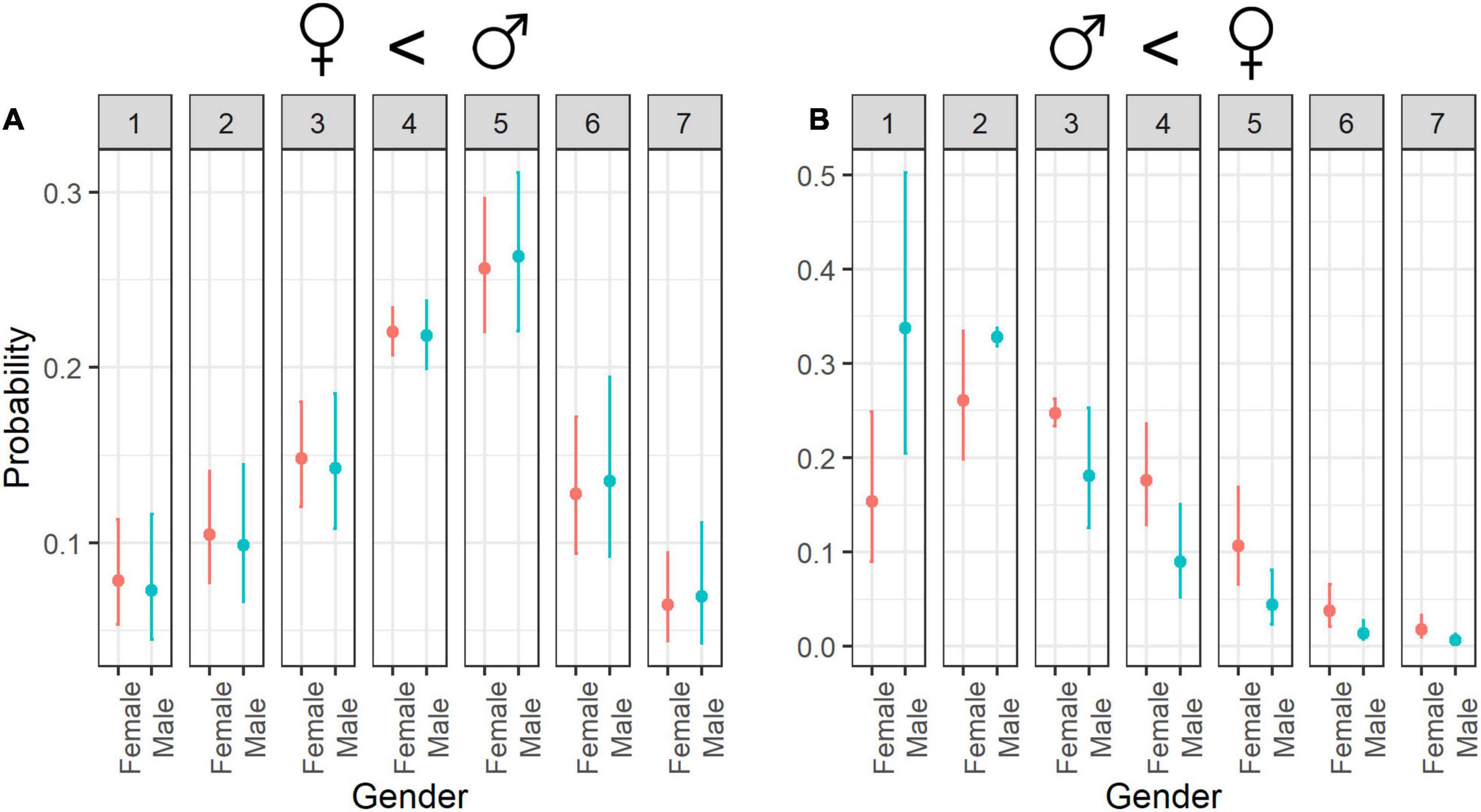
Figure 8. Estimated probability of ratings for (A) cuteness and (B) fear according to Gender. Error bars indicate 95% confidence interval; numbers 1–7 refer to rating scores.
For fear evaluations, gender was statistically significant as an individual factor, and it also interacted with several other variables. Women had a tendency to give higher fear scores relative to men (Figure 8B). The most fearful evaluations were phylogenetically close animals rated by women, and the least fearful evaluations were phylogenetically distant animals rated by men (Table 4).
Gender was not statistically significant for willingness to protection as an individual factor nor for interactions.
Relationship Between Cuteness, Fear, and Willingness to Protect Animals
Total cuteness scores positively correlated with willingness to protect animals and negatively correlated with perceived fear (Pearson r = 0.50 and –0.37, both P < 0.001, N = 349, respectively). Willingness to protect animals negatively correlated with perceived fear (Pearson r = –0.14, P = 0.01, N = 349).
Discussion
As far as we are aware, this is the first study which systematically investigated whether the bipedal posture of animals influences their perception by humans. We found that bipedal posture significantly influenced all three investigated domains of animal perception: perceived cuteness, fear, and willingness to protect them. Moreover, these associations were influenced by the size of the animal, phylogenetical distance from humans, direct eye contact, and perceived anthropomorphism.
We hypothesized that animals with bipedal posture would be perceived as cuter than animals with quadrupedal posture. In line with this hypothesis originally proposed by Morris (1969), bipedalism enhanced the perceived cuteness under certain contexts—for example, the cuteness of small animals with bipedal posture was enhanced more than the cuteness of large species with bipedal posture. With regard to fear, large animals, particularly carnivores, elicited stronger fear judgments than small animals (Prokop et al., 2010; Staňková et al., 2021). In addition, large, but not small, animals with bipedal posture induced greater fear. These results can be explained from an evolutionary perspective, as large carnivores were common predators of our ancestors in Africa (Treves and Palmqvist, 2007). Women are more frequently victims of predatory attacks than men (Treves and Naughton-Treves, 1999), which can explain their greater fear of large-bodied animals. Moreover, predation pressure on humans continues, although on a smaller scale than in our evolutionary past (e.g., Treves and Naughton-Treves, 1999; Löe and Røskaft, 2004; Packer et al., 2005).
Phylogenetically distant animals were perceived as less cute than phylogenetically closer animals. This finding is consistent with the different methodological approaches previously documented (the capacity for an animal to feel pain: Plous, 1993; empathy toward animals: Miralles et al., 2019). The participants also indicated lesser willingness to protect phylogenetically distant species (e.g., invertebrates) than phylogenetically closer species (e.g., mammals), which suggests that our innate preference for cute animals (baby scheme, Borgi et al., 2014) and/or the ability to empathize with animals (Miralles et al., 2019) enhances our interest in the conservation of animals similar to us (Samples et al., 1986; Plous, 1993; DeKay and McClelland, 1996).
Although it is well known that people have negative attitudes toward invertebrates (e.g., Kellert, 1993; Schlegel and Rupf, 2010; Fukano and Soga, 2021), it is hard to establish powerful strategies regarding how these attitudes can be improved (Cardoso et al., 2011). We did not observe a positive trend toward willingness to protect invertebrates if they look more anthropomorphic (e.g., an ant with raised front legs). It seems that bipedal posture itself does not fully enhance anthropomorphism (Morris, 1969), though it makes a moderate contribution. We suggest that this result can be used in conservation campaigns, where certain unpopular invertebrates could be used as flagship species if they are photographed in a natural way. Fukano and Soga (2021), for instance, showed that when people saw photographs of insects with an indoor background, perceived disgust and danger for insects were higher compared to pictures with an outdoor background.
Contrary to our hypothesis, the participants were generally more willing to protect small quadrupedal animals compared with animals with bipedal posture. Small species could enhance perceived cuteness and, consequently, our interest in the conservation of these animals. However, human preferences and conservation efforts are directed toward rather large-bodied, charismatic mammals (Kontoleon and Swanson, 2002; Clucas et al., 2008; Sitas et al., 2009; Smith et al., 2012; Veríssimo et al., 2018). Albert et al. (2018) found that charismatic animals are beautiful/cute, dangerous/impressive, or rare/endangered. We used only two species (gorilla and bear) belonging to the list of charismatic animals of Albert et al. (2018). Thus, it seems that human willingness to protect is directed toward small, cute species over large-bodied but non-charismatic animals. It is still not clear, however, why bipedal posture did not contribute to willingness to protect animals. We speculate that the bipedal posture of certain large-bodied species could be perceived as too uncommon (e.g., capybara) and/or threatening, thus reducing perceived conservation needs. Perhaps the bipedal posture of small, cute species could enhance willingness to play rather than willingness to protect them. To support this idea, small animals in anthropomorphic posture with direct eye contact were rated as cute, and eye contact provides a foundation for communication and social interaction (Kleinke, 1986; Senju and Johnson, 2009).
Limitation
Since the perception of cuteness, fear, and willingness to protect an animal only according to a picture is a very complex process, we chose ratings (CLMM model) over simple pairwise comparisons. The main reason behind this methodological strategy is that the ratings for two items tell us not only which item is preferred but also the degree to which it is preferable, so ratings can be more informative than pairwise comparisons. Additionally, if we took into account the expected complex information from pairwise comparisons, we would need many more respondents. Heterogeneity and overparameterization of the model need to be solved in multifactorial dimensions.
Conclusion
Our research on how looking similar to humans influences the perception of animals showed that bipedal posture contributes to the perceived cuteness of (particularly small) animals. Cuteness is associated with willingness to protect animals, but bipedal posture itself did not enhance the conservation support for animals. It seems that the bipedal posture of large-bodied animals could be perceived as threatening or perhaps non-typical. Small bipedal animals, particularly those with direct eye contact, could initiate social interactions different from conservation needs. However, if phylogenetically distant and less cute animals look more anthropomorphic, they receive lesser intentions of willingness to protect. We did not experimentally manipulate perceived anthropomorphism; thus, more research in this field should be done before a definitive conclusion can be made.
Data Availability Statement
The data analyzed in this study is subject to the following licenses/restrictions: Data are available from authors upon request. Requests to access these datasets should be directed to corresponding author.
Ethics Statement
Ethical review and approval was not required for the study on human participants in accordance with the local legislation and institutional requirements. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
PP, MZa, and PF designed the study. MZi analyzed the data. AP commented on the manuscript and improved the English. PP, MZa, and PF collected the data. All authors contributed equally in writing the manuscript.
Funding
This project was funded by the grant VEGA (Nos. 1/0007/2021, 1/0286/2020, and APVV 20-0081) and by Consulat de Monaco a Bratislava (République slovaque).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Albert, C., Luque, G. M., and Courchamp, F. (2018). The twenty most charismatic species. PLoS One 13:e0199149. doi: 10.1371/journal.pone.0199149
Allen, M. W., Hunstone, M., Waerstad, J., Foy, E., Hobbins, T., Wikner, B., et al. (2002). Human-to-animal similarity and participant mood influence punishment recommendations for animal abusers. Soc. Anim. 10, 267–284. doi: 10.1163/156853002320770074
Barnosky, A. D., Matzke, N., Tomiya, S., Wogan, G. O., Swartz, B., Quental, T. B., et al. (2011). Has the Earth’s sixth mass extinction already arrived? Nature 471, 51–57. doi: 10.1038/nature09678
Barua, M., Gurdak, D. J., Ahmed, R. A., and Tamuly, J. (2012). Selecting flagships for invertebrate conservation. Biodivers. Conserv. 21, 1457–1476. doi: 10.1007/s10531-012-0257-7
Bjerke, T., and Østdahl, T. (2004). Animal-related attitudes and activities in an urban population. Anthrozoös 17, 109–129. doi: 10.2752/089279304786991783
Borgi, M., and Cirulli, F. (2015). Attitudes toward animals among kindergarten children: species preferences. Anthrozoös 28, 45–59. doi: 10.2752/089279315x14129350721939
Borgi, M., Cogliati-Dezza, I., Brelsford, V., Meints, K., and Cirulli, F. (2014). Baby schema in human and animal faces induces cuteness perception and gaze allocation in children. Front. Psychol. 5:411.
Cardoso, P., Erwin, T. L., Borges, P. A. V., and New, T. R. (2011). The seven impediments in invertebrate conservation and how to overcome them. Biol. Conserv. 144, 2647–2655. doi: 10.1016/j.biocon.2011.07.024
Castillo-Huitrón, N. M., Naranjo, E. J., Santos-Fita, D., and Estrada-Lugo, E. (2020). The importance of human emotions for wildlife conservation. Front. Psychol. 11:1277.
Clucas, B., McHugh, K., and Caro, T. (2008). Flagship species on covers of US conservation and nature magazines. Biodivers. Conserv. 17:1517. doi: 10.1007/s10531-008-9361-0
Colléony, A., Clayton, S., Couvet, D., Saint Jalme, M., and Prévot, A. C. (2017). Human preferences for species conservation: animal charisma trumps endangered status. Biol. Conserv. 206, 263–269. doi: 10.1016/j.biocon.2016.11.035
Curtin, P., and Papworth, S. (2020). Coloring and size influence preferences for imaginary animals and can predict actual donations to species-specific conservation charities. Conserv. Lett. 13:e12723.
DeKay, M. L., and McClelland, G. H. (1996). Probability and utility of endangered species preservation programs. J. Exper. Psychol. Appl. 2, 60–83. doi: 10.1037/1076-898x.2.1.60
Dirzo, R., Young, H. S., Galetti, M., Ceballos, G., Isaac, N. J., and Collen, B. (2014). Defaunation in the anthropocene. Science 345, 401–406. doi: 10.1126/science.1251817
Driscoll, J. W. (1995). Attitudes toward animals: species ratings. Soc. Anim. 3, 139–150. doi: 10.1163/156853095x00125
Emery, N. J. (2000). The eyes have it: the neuroethology, function and evolution of social gaze. Neurosci. Biobehav. Rev. 24, 581–604. doi: 10.1016/s0149-7634(00)00025-7
Frynta, D., Lišková, S., Bültmann, S., and Burda, H. (2010). Being attractive brings advantages: the case of parrot species in captivity. PLoS One 5:e12568. doi: 10.1371/journal.pone.0012568
Frynta, D., Šimková, O., Lišková, S., and Landová, E. (2013). Mammalian collection on Noah’s ark: the effects of beauty, brain and body size. PLoS One 8:e63110. doi: 10.1371/journal.pone.0063110
Fukano, Y., and Soga, M. (2021). Why do so many modern people hate insects? the urbanization–disgust hypothesis. Sci. Total Environ. 777:146229. doi: 10.1016/j.scitotenv.2021.146229
Gunnthorsdottir, A. (2001). Physical attractiveness of an animal species as a decision factor for its preservation. Anthrozoös 14, 204–215. doi: 10.2752/089279301786999355
Hallmann, C. A., Sorg, M., Jongejans, E., Siepel, H., Hofland, N., Schwan, H., et al. (2017). More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS One 12:e0185809. doi: 10.1371/journal.pone.0185809
Hart, D., and Sussman, R. W. (2008). Man the Hunted: Primates, Predators, and Human Evolution. Boulder, CO: Westview Press.
Houston, M. J., Bruskotter, J. T., and Fan, D. (2010). Attitudes toward wolves in the United States and Canada: a content analysis of the print news media, 1999–2008. Hum. Dimension Wildlife 15, 389–403. doi: 10.1080/10871209.2010.507563
Kellert, S. R. (1980). Knowledge, Affection, and Basic Attitudes Toward Animals in American Society: Phase III. Virginia: US Department of the Interior, Fish and Wildlife Service.
Kellert, S. R. (1983). “Affective, cognitive, and evaluative perceptions of animals,” in Behavior and the Natural Environment, eds I. Altman and J. F. Wohlwill. Boston, MA: Springer.
Kellert, S. R. (1985). Public perceptions of predators, particularly the wolf and coyote. Biol. Conserv. 31, 167–189. doi: 10.1016/0006-3207(85)90047-3
Kellert, S. R. (1993). Values and perceptions of invertebrates. Conserv. Biol. 7, 845–855. doi: 10.1046/j.1523-1739.1993.740845.x
Kleinke, C. L. (1986). Gaze and eye contact: a research review. Psychol. Bull. 100, 78–100. doi: 10.1037/0033-2909.100.1.78
Knegtering, E., Van Der Windt, H. J., and Schoot Uiterkamp, A. J. M. (2010). Public decisions on animal species: does body size matter? Environ. Conserv. 38, 28–36. doi: 10.1017/s0376892910000755
Knight, A. J. (2008). Bats, snakes and spiders, Oh my!” how aesthetic and negativistic attitudes, and other concepts predict support for species protection. J. Environ. Psychol. 28, 94–103. doi: 10.1016/j.jenvp.2007.10.001
Knight, S., Nunkoosing, K., Vrij, A., and Cherryman, J. (2003). Using grounded theory to examine people’s atti-tudes toward how animals are used. Soc. Anim. 11, 307–327. doi: 10.1163/156853003322796064
Kontoleon, A., and Swanson, T. (2002). The willingness to pay for property rights for the giant panda: can a charismatic species be an instrument for nature conservation? Land Econ. 79, 483–499. doi: 10.2307/3147295
Kruse, H., Kirkemo, A. M., and Handeland, K. (2004). Wildlife as source of zoonotic infections. Emerg. Infect. Dis. 10, 2067–2072. doi: 10.3201/eid1012.040707
Kumar, S., Stecher, G., Suleski, M., and Hedges, S. B. (2017). TimeTree: a resource for timelines, timetrees, and divergence times. Mol. Biol. Evol. 34, 1812–1819. doi: 10.1093/molbev/msx116
Macdonald, E. A., Burnham, D., Hinks, A. E., Dickman, A. J., and Malhi, Y. (2015). Conservation inequality and the charismatic cat: Felis felicis. Global Ecol. Conserv. 3, 851–866. doi: 10.1016/j.gecco.2015.04.006
MacFarlane, D., and Rocha, R. (2020). Guidelines for communicating about bats to prevent persecution in the time of COVID-19. Biol. Conserv. 248:108650. doi: 10.1016/j.biocon.2020.108650
Małecki, W., Sorokowski, P., Pawłowski, B., and Cieński, M. (2020). Human Minds and Animal Stories: How Narratives Make us Care About other Species. Milton Park: Routledge.
Marešová, J., Landová, E., and Frynta, D. (2009). What makes some species of milk snakes more attractive to humans than others? Theory Biosci. 128, 227–235. doi: 10.1007/s12064-009-0075-y
Martín-López, B., Montes, C., and Benayas, J. (2007). The non-economic motives behind the willingness to pay for biodiversity conservation. Biol. Conserv. 139, 67–82. doi: 10.1016/j.biocon.2007.06.005
Miralles, A., Raymond, M., and Lecointre, G. (2019). Empathy and compassion toward other species decrease with evolutionary divergence time. Sci. Rep. 9:19555.
Mullin, M. H. (1999). Mirrors and windows: sociocultural studies of human-animal relationships. Annu. Rev. Anthropol. 28, 201–224. doi: 10.1146/annurev.anthro.28.1.201
Newbold, T., Adams, G. L., Albaladejo Robles, G., Boakes, E. H., Braga Ferreira, G., Chapman, A. S., et al. (2019). Climate and land-use change homogenise terrestrial biodiversity, with consequences for ecosystem functioning and human well-being. Emerg. Top. Life Sci. 3, 207–219. doi: 10.1042/etls20180135
Ordish, G. (1976). The Constant Pest. a Short History of Pests and their Control. The Constant Pest. a Short History of Pests and their Control. USA: Macmillan Pub. Co.
Packer, C., Hilborn, R., Mosser, A., Kissui, B., Borner, M., Hopcraft, G., et al. (2005). Ecological change, group territoriality, and population dynamics in Serengeti lions. Science 307, 390–393. doi: 10.1126/science.1105122
Podberscek, A. L., Paul, E. S., and Serpell, J. A. (2005). Companion Animals and us: Exploring the Relationships Between People and Pets. Cambridge: Cambridge University Press.
Prokop, P., and Fančovičová, J. (2013). Does colour matter? the influence of animal warning coloration on human emotions and willingness to protect them. Anim. Conserv. 16, 458–466. doi: 10.1111/acv.12014
Prokop, P., and Fančovičová, J. (2017). Animals in dangerous postures enhance learning, but decrease willingness to protect animals. Eur. J. Math. Sci. Technol. Educ. 13, 6069–6077.
Prokop, P., and Randler, C. (2018). “Biological predispositions and individual differences in human attitudes toward animals,” in Ethnozoology: Animals in Our Lives, eds R. R. N. Alves and U. P. Albuquerque (London, UK: Academic Press), 447–466. doi: 10.1016/b978-0-12-809913-1.00023-5
Prokop, P., Usak, M., and Fančovičová, J. (2010). Risk of parasite transmission influences perceived vulnerability to disease and perceived danger of disease-relevant animals. Behav. Proc. 85, 52–57. doi: 10.1016/j.beproc.2010.06.006
R Core Team (2019). R: a Language and Environment for Statistical Computing. Vienna: R Foundation for statistical computing.
Røskaft, E. (2004). Large carnivores and human safety: a review. AMBIO J. Hum. Environ. 33, 283–288. doi: 10.1579/0044-7447-33.6.283
Samples, K. C., Dixon, J. A., and Gowen, M. M. (1986). Information disclosure and endangered species valuation. Land Econ. 62, 306–312. doi: 10.2307/3146394
Schlegel, J., and Rupf, R. (2010). Attitudes towards potential animal flagship species in nature conservation: a survey among students of different educational institutions. J. Nat. Conserv. 18, 278–290. doi: 10.1016/j.jnc.2009.12.002
Senju, A., and Johnson, M. H. (2009). The eye contact effect: mechanisms and development. Trends Cogn. Sci. 13, 127–1349. doi: 10.1016/j.tics.2008.11.009
Serpell, J. A. (2004). Factors influencing human attitudes to animals and their welfare. Anim. Welf. 13, S145–S152.
Sitas, N., Baillie, J. E. M., and Isaac, N. J. B. (2009). What are we saving? Developing a standardized approach for conservation action. Anim. Conserv. 12, 231–237. doi: 10.1111/j.1469-1795.2009.00244.x
Smith, R. J., Veríssimo, D., Isaac, N. J. B., and Jones, K. E. (2012). Identifying Cinderella species: uncovering mammals with conservation flagship appeal. Conserv. Lett. 5, 205–212. doi: 10.1111/j.1755-263x.2012.00229.x
Staňková, H., Janovcová, M., Peléšková, Š, Sedláčková, K., Landová, E., and Frynta, D. (2021). The ultimate list of the most frightening and disgusting animals: negative emotions elicited by animals in Central European respondents. Animals 11:747. doi: 10.3390/ani11030747
Theng, M., Jusoh, W. F., Jain, A., Huertas, B., Tan, D. J., Tan, H. Z., et al. (2020). A comprehensive assessment of diversity loss in a well-documented tropical insect fauna: almost half of Singapore’s butterfly species extirpated in 160 years. Biol. Conserv. 242:108401. doi: 10.1016/j.biocon.2019.108401
Tisdell, C., Wilson, C., and Nantha, H. S. (2006). Public choice of species for the ‘Ark’: phylogenetic similarity and preferred wildlife species for survival. J. Nat. Conserv. 14, 97–105. doi: 10.1016/j.jnc.2005.11.001
Treves, A., and Naughton-Treves, L. (1999). Risk and opportunity for humans coexisting with large carnivores. J. Hum. Evol. 36, 275–282. doi: 10.1006/jhev.1998.0268
Treves, A., and Palmqvist, P. (2007). “Reconstructing hominin interactions with mammalian carnivores (6.0–1.8 Ma),” in Primate Anti-Predator Strategies. Developments in Primatology: Progress and Prospects, eds S. L. Gursky and K. A. I. Nekaris (Boston, MA: Springer).
Ungar, P. S., and Teaford, M. F. (2002). Human Diet: its Origin and Evolution. Westport, CT: Greenwood Publishing Group.
van der Plas, F. (2019). Biodiversity and ecosystem functioning in naturally assembled communities. Biol. Rev. 94, 1220–1245.
Veríssimo, D., Campbell, H. A., Tollington, S., MacMillan, D. C., and Smith, R. J. (2018). Why do people donate to conservation? insights from a ‘real world’ campaign. PLoS One 13:e0191888. doi: 10.1371/journal.pone.0191888
Keywords: bipedal posture, animals, attitudes, conservation, willingness to protect
Citation: Prokop P, Zvaríková M, Zvarík M, Pazda A and Fedor P (2021) The Effect of Animal Bipedal Posture on Perceived Cuteness, Fear, and Willingness to Protect Them. Front. Ecol. Evol. 9:681241. doi: 10.3389/fevo.2021.681241
Received: 16 March 2021; Accepted: 07 June 2021;
Published: 02 July 2021.
Edited by:
Piotr Tryjanowski, Poznań University of Life Sciences, PolandReviewed by:
Daniel Frynta, Charles University, CzechiaPiotr Sorokowski, University of Wrocław, Poland
Copyright © 2021 Prokop, Zvaríková, Zvarík, Pazda and Fedor. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pavol Prokop, cGF2b2wucHJva29wQHNhdmJhLnNr