
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 20 July 2021
Sec. Chemical Ecology
Volume 9 - 2021 | https://doi.org/10.3389/fevo.2021.676743
This article is part of the Research Topic Sensory Ecology of Plant-Pollinator Interactions View all 24 articles
 Carlos A. Martínez-Martínez1
Carlos A. Martínez-Martínez1 Guaraci D. Cordeiro2*
Guaraci D. Cordeiro2* Herbeson O. J. Martins1
Herbeson O. J. Martins1 Renan O. A. C. Kobal1
Renan O. A. C. Kobal1 Paulo Milet-Pinheiro3
Paulo Milet-Pinheiro3 Mariana A. Stanton4
Mariana A. Stanton4 Emanuella L. Franco5
Emanuella L. Franco5 Cristiane Krug6
Cristiane Krug6 Sidnei Mateus1
Sidnei Mateus1 Clemens Schlindwein7
Clemens Schlindwein7 Stefan Dötterl2
Stefan Dötterl2 Isabel Alves-dos-Santos8
Isabel Alves-dos-Santos8Crepuscular and/or nocturnal bees fly during the dusk, the dawn or part of the night. Due to their short foraging time and sampling bias toward diurnal bees, nocturnal bees are rarely collected and poorly studied. So far, they have been mostly sampled with light and Malaise traps. However, synthetic chemical compounds resembling floral volatiles were recently found to be a promising alternative to attract these bees. By reviewing available literature and collecting original data, we present information on the attraction and sampling of nocturnal bees with scent-baited traps. Bees were actively captured with entomological nets while approaching to filter papers moistened with distinct chemical compound, or passively caught in bottles with scent baits left during the night. So far, all data available are from the Neotropics. Nocturnal bees belonging to three genera, i.e., Ptiloglossa, Megalopta, and Megommation were attracted to at least ten different synthetic compounds and mixtures thereof, identified from bouquets of flowers with nocturnal anthesis. Aromatic compounds, such as 2-phenyletanol, eugenol and methyl salicylate, and the monoterpenoid eucalyptol were the most successful in attracting nocturnal bees. We highlight the effectiveness of olfactory methods to survey crepuscular and nocturnal bees using chemical compounds typically reported as floral scent constituents, and the possibility to record olfactory preferences of each bee species to specific compounds. We suggest to include this method in apifauna surveys in order to improve our current knowledge on the diversity of nocturnal bees in different ecosystems.
The nocturnal and/or crepuscular behavior in bees arose independently in four of the seven bee families: Andrenidae, Apidae, Colletidae, and Halictidae (Wcislo et al., 2004; Warrant, 2007; Danforth et al., 2019). There are about 250 described nocturnal bee species and they fly during the dusk, the dawn or part of the night. These bees can be obligatory nocturnal, such as the giant Indian bee Xylocopa tranquebarica (Burgett and Sukumalanand, 2000), or crepuscular, i.e., forage for pollen and nectar at dawn or dusk, such as Megalopta and Ptiloglossa (Warrant, 2007). Furthermore, under ideal moonlight and cloud-cover conditions, the crepuscular period is extended allowing “crepuscular bees” to search for food also during the night (Kerfoot, 1967; Somanathan et al., 2008; Liporoni et al., 2020). The main anatomical characteristics that indicate nocturnal and/or crepuscular behavior in these bees are the large size of their ocelli and compound eyes, as well as the high number of ommatidia (Kelber et al., 2006; Warrant et al., 2006; Berry et al., 2011), characteristics that improve visual orientation in low light conditions (Wcislo et al., 2004).
Besides visual adaptations for dim light, nocturnal pollinators often heavily depend on floral odors to find their host flowers (Borges et al., 2016). Indeed, nocturnal bees tend to visit and are attracted by flowers releasing a strong perfume at night, so far known to be mainly composed of aromatic (e.g., 2-phenylethanol), aliphatic (e.g., 1-octanol), and terpenoid (e.g., linalool) compounds (Cordeiro et al., 2017, 2019; Krug et al., 2018), all widespread among flower scents (Knudsen et al., 2006). Synthetic compounds that are broadly applied in male orchid bee (Euglossini) surveys were fortuitously found to also attract nocturnal bees (Carvalho et al., 2012; Knoll and Santos, 2012) and, more recently, nocturnal bees were effectively lured with compounds (presented individually or as blends) resembling floral volatiles of some night-blooming host plants of these bees (Cordeiro et al., 2017; Krug et al., 2018). Furthermore, during pollination studies at night, nocturnal bees are recorded on flowers (Hopkins et al., 2000; Somanathan and Borges, 2001; Franco and Gimenes, 2011; Krug et al., 2015; Cordeiro et al., 2017; Soares and Morellato, 2018, Cordeiro et al., 2021).
Nocturnal and/or crepuscular bees (hereafter referred to as nocturnal bees) are usually undersampled, due to their short foraging time and sampling bias toward diurnal bees (Wcislo et al., 2004). As a consequence, representativeness of these bees in insect collections are normally scarce. So far, many of the nocturnal bees collected have been captured with light traps using white light tubes, modified Pennsylvania black light, ultraviolet light (UV), mercury vapor lamps (Chandler, 1961; Wolda and Roubik, 1986) or Malaise traps (Ferrari et al., 2016).
Light traps are efficiently used in the documentation of nocturnal bees, as well as in determining seasonal patterns of other insects (Wolda and Roubik, 1986; Abbas et al., 2019). However, one of their disadvantages is the high cost of batteries or power generators, and lamps with different types of lights being necessary for operating the trap. Likewise, these traps tend to be generalist, attracting various types of insects that are not the object of study, and trapping is strongly affected by abiotic variables such as moon phases and weather (Nowinszky and Puskás, 2017). Furthermore, these traps are fragile and pose danger to the collector, due to UV radiation emitted and toxicity of mercury (Price and Baker, 2016).
Due to the general scarcity of captured specimens of nocturnal bees in insect collections, our current knowledge about their diversity is still underestimated. In this study, we propose a methodological protocol to improve the sampling of nocturnal bees, an approach that might increase information on their diversity and on the olfactory preferences of the different species. In this study, we provide new records and a compilation of literature data on nocturnal bees lured with chemical compounds.
Nocturnal bees were sampled with synthetic chemical compounds from July 2019 to January 2020 in three localities of São Paulo State, southeastern Brazil: Osasco municipality (23°28′ S, 46°46′ W); Neblinas Park (23°44′ S, 46°09′ W); Municipal Reserve Serra do Japi (23°14′ S, 46°58′ W). The method was standardized by offering 1 ml of chemical compounds (or mixtures) added on filter papers (9 cm of diameter) and disposed on tree- or shrub branches at a height between 1.5 and 2 m above the ground. Bees were collected with an entomological handnet while approaching filter papers with chemical compounds (Figure 1A), between 04:00 and 6:00 am (before sunrise), and between 05:00 and 07:00 pm (by sunset), corresponding to the activity time of nocturnal bee species. The bees sampled were transferred to glass vials containing ethyl acetate. We offered compounds resembling floral volatiles previously identified as attractants to nocturnal bees (Cordeiro et al., 2017; Krug et al., 2018), i.e., 1-hexanol (Sigma-Aldrich, ≥99%), 2-phenylethanol (Acros Organics, 99%), 1-octanol (Sigma-Aldrich, ≥99%), and linalool (Sigma-Aldrich, 97%). Our sampling effort summed up to 87 h within 23 days. We are calling this sampling as active method, since the collector remains present during the exposure of the scent and actively collects the approaching bees.
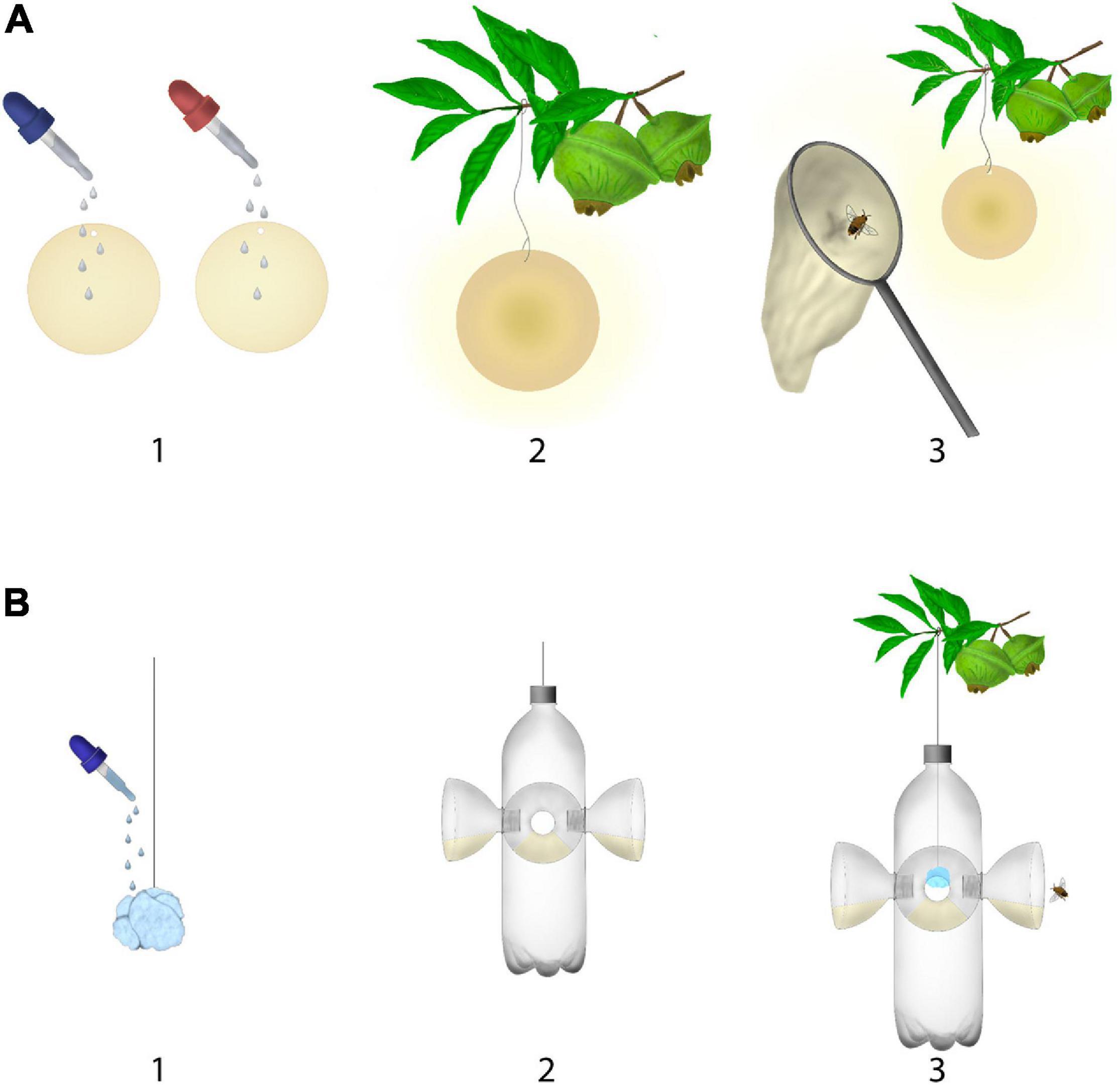
Figure 1. Diagram of the active method (A) and passive (B) methods used to collect nocturnal bees with chemical compounds. We recommend the application of about 1 ml chemical compounds to filter paper or a cotton ball.
The collected bees were mounted and identified with the taxonomic keys proposed by Moure (1945, 1964), Santos and Silveira (2009),Gonçalves and Santos (2010), Gonzalez et al. (2010) and Santos and Melo (2015), and deposited in the Paulo Nogueira-Neto Entomological Collection at the University of São Paulo (CEPANN), in São Paulo, Brazil.
A systematic review of the literature on nocturnal bees lured with chemical compounds was conducted on Google Scholar, JSTOR, NCBI, Scopus, and Web of science, with the following combinations of keywords: “Compounds” OR “Chemical lures” OR “Floral scent” OR “Floral volatile” OR “Nocturnal bees” OR “Nocturnal and Crepuscular bees” OR “Nocturnal anthesis” OR “Volatile organic compounds.” From the selected articles we recovered information about the localities, species, number of individuals attracted, and attractive compounds (Table 1 and Supplementary Table 1). In these studies, the nocturnal bees were mostly sampled with bottles scent traps (similar to those used in Euglossini bees sampling) left during the night. This trap is built with PET bottles, in which a cotton ball impregnated with a synthetic compound is inserted (Figure 1B). Chemical compounds used as lures in these previous studies were mostly: eucalyptol, eugenol and methyl salicylate. This sampling protocol is referred hereafter as passive method, since the traps are left in the field without direct supervision and removed in the day after.

Table 1. Number of individuals of nocturnal bee species lured with synthetic chemical compounds and collected with active and passive methods.
In the present and previous studies (literature review), 1115 individuals of 12 species of nocturnal bees were attracted to chemical compounds. The genus with the highest number of individuals and species registered was Megalopta (1050 individuals, 8 species) (Table 1).
The active sampling method attracted 103 individuals belonging to ten species of nocturnal bees, comprising our sampling (41 individuals, five species) and previous studies (62 individuals, six species). The bees most commonly recorded during the active methods were Megommation insigne (33 individuals) and Megalopta aeneicollis (25 individuals). Most individuals (39 individuals) were attracted by the aromatic compound 2-phenylethanol (Table 1).
The passive method, applied in all previous studies, attracted 1012 individuals, among them Megalopta amoena and M. guimaraesi, which accounted together for 994 individuals (Table 1).
The aromatic compounds eugenol and methyl salicylate trapped 530 individuals from 4 species and 233 individuals from 5 species, respectively (Table 1). The only monoterpene tested as single compound, eucalyptol, attracted 152 individuals of three Megalopta species. The aliphatic compound 1-octanol, attracted bees of the genera Megommation and Ptiloglossa. The different mixtures of compounds attracted 57 individuals from seven species, including some unique species, such as Megalopta cuprea and M. piraha which were only collected with specific mixtures, but not single compounds. Compounds from other chemical classes, such as irregular terpenes and nitrogen-bearing compounds, attracted only few bees (4 individuals) (Table 1).
The results demonstrate that chemical compounds are appropriate to sample nocturnal bees, and sampling of bees attracted by volatile organic compounds should be incorporated as an additional method to apifauna surveys. It is worthy to offer compounds identified in plants that serve as host for nocturnal bees and use them individually or as blends.
The capture of nocturnal bees has usually been done with black and fluorescent light traps. Chandler (1961) collected 392 individuals of Sphecodogastra texana in LaPorte Indiana between 1959 and 1960. However, according to the author there is a possible interference by the killing agent cyanide in the attraction of the bees. Likewise, Wolda and Roubik (1986) collected an astonishing number of individuals of two Megalopta species on the island of Barro Colorado, in Panama: 7,713 and 2,487 individuals of M. ecuadoria and M. genalis, respectively, were sampled. One possible reason that explains the high attraction of these two species of Megalopta to light traps on the island was the synchronization with the flowering of the Tachigalia versicolor where the light trap was installed (Wolda and Roubik, 1986). The floral volatiles emitted by this plant may have helped attracting the bees to the light trap. Another explanation is that a high abundance of nests of these two species might have been close to the light traps (Roulston, 1997; Wcislo et al., 2004).
Floral synthetic compounds almost exclusively attracted female nocturnal bees (Table 1). This differs with the sex of individuals attracted in the Euglossini tribe. Nemésio (2012) mentioned that attracting only males in this tribe has led to taxonomic problems that involve describing species based on male specimens and making it difficult to match males with females. This situation is similar in the nocturnal bees, especially of the genus Ptiloglossa, where the taxonomic identification keys were constructed only for male specimens (Moure, 1945). However, the increase in female nocturnal bees in collections, e.g., by using chemical attractants, allows the construction of taxonomic keys that include male and female specimens, as done by Velez-Ruiz (2015).
Our study suggests that chemical compounds sample a higher diversity of species than light traps. Overall 12 species of 3 genera (Megalopta, Megommation, and Ptiloglossa) were collected on chemical lures so far, while the surveys conducted with light traps recorded five species, most of them belonging to the genus Megalopta (Chandler, 1961; Kerfoot, 1967; Wolda and Roubik, 1986; Roulston, 1997). Although the sample design is not comparable in terms of time, area, climatic conditions, etc, the superiority in the number of species attracted by the chemical method suggests that this method is more effective in attracting high numbers of species.
The ability of chemical compounds according to the diversity and abundance of attracted nocturnal bees varied among the chemical classes used. However, this may be due to the differences in the design and duration of the sampling carried out by each author in the literary review. Nevertheless, most of the nocturnal bees were attracted to aromatic compounds, such as eugenol, methyl salicylate, 2-phenylethanol, and monoterpenes such as eucalyptol.
The aromatic compounds eugenol and methyl salicylate and the monoterpene eucalyptol, widely used for the attraction of male euglossine bees (Nemésio, 2012), lured 527, 236, and 152 individuals of nocturnal bees, respectively (Carvalho et al., 2012; Knoll and Santos, 2012; Almeida et al., 2020), all from the genus Megalopta, with a single exception of one individual of Ptiloglossa sampled with methyl salicylate (Almeida et al., 2020). The species most abundantly attracted to the abovementioned three compounds were M. amoena and M. guimaraesi. Although little is known about the abundance of these compounds in plants with nocturnal anthesis, they are present in a wide variety of plants with diurnal and nocturnal anthesis (Knudsen et al., 2006; El-Sayed, 2021). We believe that the sampling of floral scents in a broader spectrum of plants visited by nocturnal bees might reveal these attractive compounds at least in some representatives.
Another aromatic worth mentioning is 2-phenylethanol. Unlike eugenol, methyl salicylate, and eucalyptol, 2-phenylethanol tends to be less specific and attracts more than one genus and family of nocturnal bees, including rare species such as P. pretiosa. Preliminary results also show that this compound is capable of eliciting physiological responses in electroantennography assays (EAG) with M. insigne (Supplementary Figure 1). This general efficiency of 2-phenylethanol in attracting nocturnal bees may be due to their widespread occurrence among floral scents (Knudsen et al., 2006), sometimes also as major component of bee-pollinated plants (Dobson, 2006), including some with nocturnal anthesis (Shaver et al., 1997; Cordeiro et al., 2017, 2019). This aromatic compound is also a known attractant for diurnal bees (Dötterl and Vereecken, 2010; Rocha-Filho and Garófalo, 2014).
Previous studies demonstrate that not only single compounds but also synthetic mixtures of compounds are capable of attracting nocturnal bee pollinators. The mixtures attracted 41 specimens of nocturnal bees of at least three genera and two families, including some Megalopta collected exclusively with these mixtures. Rare species such as M. cuprea and M. piraha were exclusively attracted to mixtures. Furthermore, unlike the individual compounds, these mixtures have the advantage of resembling the natural aroma emitted by the flowers and attracting potential pollinators (Cordeiro et al., 2017; Krug et al., 2018).
All methods used for apifauna surveys have advantages and disadvantages. The passive method was able to sample a high number of individuals. However, as Euglossini surveys, it can sample hundreds of individuals in one day (Viana et al., 2002; Nemésio and Vasconcelos, 2013), therefore it must be applied with care in fragmented forests with potentially small populations. The active sampling method attracted smaller number of individuals but more species. In addition, in the active sampling, it is possible (for some species) to determine the specimens directly in the field and avoid killing all attracted individuals. Finally, it enables isolating the caught bees in single vials, allowing pollen analyses of each individual. The passive sampling method allows the bottle traps with chemical compound to be left overnight. Thus it is less time consuming than active methods, where collectors spend hours in front of the baits.
To conclude, our study shows that synthetic chemical compounds lure a wide diversity of nocturnal bees. Nocturnal bee species are successfully attracted by aromatic compounds and monoterpenes such as eugenol, methyl salicylate, 2-phenylethanol and eucalyptol. Offering volatile compounds in an active and passive way should be included in nocturnal apifauna surveys, as this approach attracts species otherwise difficult to obtain, and helps clarifying taxonomic issues and the dynamics of their populations of these important pollinators.
The raw data supporting the conclusion of the article will be made available by the authors, without undue reservation.
CM-M, HM, IA-d-S, and RK: conceptualization. CM-M, CS, CK, EF, GC, HM, IA-d-S, RK, SD, and SM: investigation, methodology, and carried out collection of data. MS: electroantennographic assay. CM-M, GC, HM, IA-d-S, MS, PM-P, RK, and SD: writing – original draft preparation and review. IA-d-S: supervision. All authors have read, revised, and agreed the final version of the manuscript.
This research was funded by CNPq (Grant No. PVE 401466/2014-0) to IA-d-S, CS, and SD. CNPq scholarships to HM, IA-d-S, and CS. Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001 accordingto Art. 1°/n° 206 CAPES to CM-M and RK. FAPESP (Grant No. 2014/50316-7) to Massuo Jorge Katon.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We are thankful to the Ronaldo Pereira (Municipal Park Serra do Japi) and Ecofuturo Institute (Neblinas Park) for logistic support and the permission to work in the biological reserves. We are also grateful to Antônio Rogerio dos Santos for generously allowing collections to be made in his own residence. Likewise, we thank Prof. Massuo Jorge Kato for allowing us to use the infrastructure of the LQPN – Laboratório de Química de Produtos Naturais, University of São Paulo, where the EAG tests were conducted, the Laboratório de Biologia Comparada e Abelhas (LBCA), for the use its facilities for the identification of the collected specimens and Rita Isabel Velez Ruiz for the identification of Ptiloglossa species. We thank the Entomological Graduate Program of USP for support. Finally, we are grateful to the two reviewers for their suggestions.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.676743/full#supplementary-material
Abbas, M., Ramzan, M., Hussain, N., Ghaffar, A., Hussain, K., Abbas, S., et al. (2019). Role of light traps in attracting, killing and biodiversity studies of insect pests in Thal. Pak. J. Agric. Sci. 32, 684–690.
Almeida, V. M. N., Franco, E. L., Carvalho, M. M., and Pereira, C. V. (2020). “Atração de abelhas crepusculares e diurnas por iscas-odores em duas áreas distintas na Chapada Diamantina-Bahia,” In A Interface Do Conhecimento Sobre Abelhas 2, Org, eds J. M. Barbosa and L. Batista (Belo Horizonte, Atenas: Editora), 18–33. doi: 10.22533/at.ed.3682001103
Berry, R. P., Wcislo, W. T., and Warrant, E. J. (2011). Ocellar adaptations for dim light vision in a nocturnal bee. J. Exp. Biol. 214, 1283–1293. doi: 10.1242/jeb.050427
Borges, R. M., Somanathan, H., and Kelber, A. (2016). Patterns and processes in nocturnal and crepuscular pollination services. Q. Rev. Biol. 91, 389–418. doi: 10.1086/689481
Burgett, D. M., and Sukumalanand, P. (2000). Flight activity of Xylocopa (Nyctomelitta) tranquebarica: a night flying carpenter bee (Hymenoptera: apidae). J. Apic. Res. 39, 75–83.
Carvalho, A. T., Maia, A. C. D., Ojima, P. Y., dos Santos, A. A., and Schlindwein, C. (2012). Nocturnal bees are attracted by widespread floral scents. J. Chem. Ecol. 38, 315–318. doi: 10.1007/s10886-012-0084-z
Chandler, L. (1961). Light trap collections of the nocturnal bee, Sphecodogastra texana (Cresson) (HymenIoptera, Halictidae). Proc. Indiana Acad. Sci. 71, 124–129.
Cordeiro, G. D., dos Santos, I. G. F., da Silva, C. I., Schlindwein, C., Alves-dos-Santos, I., and Dötterl, S. (2019). Nocturnal floral scent profiles of Myrtaceae fruit crops. Phytochemistry 162, 193–198. doi: 10.1016/j.phytochem.2019.03.011
Cordeiro, G. D., Liporoni, R., Caetano, C. A., Krug, C., Martínez-Martínez, C. A., Martins, H. O. J., et al. (2021). Nocturnal bees as crop pollinators. Agronomy 11, 1014.
Cordeiro, G. D., Pinheiro, M., Dötterl, S., and Alves-dos-Santos, I. (2017). Pollination of Campomanesia phaea (Myrtaceae) by night-active bees: a new nocturnal pollination system mediated by floral scent. Plant Biol. 19, 132–139. doi: 10.1111/plb.12520
Danforth, B. N., Minckley, R. L., Neff, J. L., and Fawcett, F. (2019). The Solitary Bees: Biology, Evolution, Conservation. Princeton: Princeton University Press.
Dobson, H. E. (2006). “). Relationship between floral fragrance composition and type of pollinator,” in Biology Of Floral Scent, eds N. Dudareva and E. Pichersky (Boca Raton: CRC Press/Taylor and Francis), 147–198. doi: 10.1201/9781420004007-8
Dötterl, S., and Vereecken, N. J. (2010). The chemical ecology and evolution of bee–flower interactions: a review and perspectives. Can. J. Zool. 88, 668–697. doi: 10.1139/z10-031
El-Sayed, A. M. (2021). The Pherobase: Database of Pheromones and Semiochemicals. https://www.pherobase.com
Ferrari, R. R., Arantes, R. C., and Silveira, F. A. (2016). A new species of Mydrosomella Michener (Hymenoptera: colletidae) from the semiarid caatinga domain, northeastern Brazil, with an identification key for the Species in the Genus. Ann. Zool. 66, 403–410. doi: 10.3161/00034541anz2016.66.3.006
Franco, E. L., and Gimenes, B. M. (2011). Pollination of Cambessedesia wurdackii in Brazilian campo rupestre vegetation, with special reference to crepuscular bees. J. Insect Sci. 11, 1–13. doi: 10.1673/031.011.9701
Gonçalves, R. B., and Santos, L. M. (2010). Notes and new species of the halictine genus Megommation Moure (Hymenoptera, Apidae, Augochlorini). Zootaxa 2685, 57–64. doi: 10.11646/zootaxa.2685.1.5
Gonzalez, V. H., Griswold, T., and Ayala, R. (2010). Two new species of nocturnal bees of the genus Megalopta (Hymenoptera: halictidae) with keys to species. Rev. Biol. Trop. 58, 255–263.
Hopkins, M. J. G., Hopkins, H. F., and Sothers, C. A. (2000). Nocturnal pollination of Parkia velutina by Megalopta bees in Amazonia and its possible significance in the evolution of chiropterophily. J. Trop. Ecol. 5, 733–746. doi: 10.1017/s0266467400001681
Kelber, A., Warrant, E. J., Pfaff, M., Wallén, R., Theobald, J. C., Wcislo, W. T., et al. (2006). Light intensity limits foraging activity in nocturnal and crepuscular bees. Behav. Ecol. 17, 63–72. doi: 10.1093/beheco/arj001
Kerfoot, W. B. (1967). The lunar periodicity of Sphecodogastra texana, a nocturnal bee (Hymenoptera: halictidae). Anim. Behav. 15, 479–486. doi: 10.1016/0003-3472(67)90047-4
Knoll, F., and Santos, L. M. (2012). Orchid bee baits attracting bees of the genus Megalopta (Hymenoptera, Halictidae) in Bauru region, São Paulo, Brazil: abundance, seasonality, and the importance of odors for dim-light bees. Rev. Bras. Entomol. 56, 481–488. doi: 10.1590/s0085-56262012000400013
Knudsen, J. T., Eriksson, R., Gershenzon, J., and Ståhl, B. (2006). Diversity and distribution of floral scent. Bot. Rev. 72, 1–120. doi: 10.1663/0006-8101(2006)72[1:dadofs]2.0.co;2
Krug, C., Cordeiro, G. D., Schäffler, I., Silva, C. I., Oliveira, R., Schlindwein, C., et al. (2018). Nocturnal bee pollinators are attracted to guarana flowers by their scents. Front. Plant Sci. 9:1072. doi: 10.3389/fpls.2018.01072
Krug, C., Garcia, M. V. B., and Gomes, F. B. (2015). A scientific note on new insights in the pollination of guarana (Paullinia cupana var. sorbilis). Apidologie 46, 164–166. doi: 10.1007/s13592-014-0304-3
Liporoni, R., Cordeiro, G. D., Prado, P. I., Schlindwein, C., Warrant, E. J., and Alves-dos-Santos, I. (2020). Light intensity regulates flower visitation in Neotropical nocturnal bees. Sci. Rep. 10:15333. doi: 10.1038/s41598-020-72047-x
Moure, J. S. (1945). Contribuição para o conhecimento dos Diphaglossinae, particularmente Ptiloglossa (Hym. - Apoidea). Arq. Mus. Paranaense. 4, 137–178.
Moure, J. S. (1964). As espécies de Zikanapis, com a descrição de dois novos subgêneros e duas espécies novas (Hym. Apoidea). Stud. Ent. 7, 417–458.
Nemésio, A. (2012). Methodological concerns and challenges in ecological studies with orchid bees (Hymenoptera: Apidae: Euglossina). Biosci. J. 28, 118–135.
Nemésio, A., and Vasconcelos, H. L. (2013). Beta diversity of orchid bees in a tropical biodiversity hotspot. Biodivers. Conserv. 22, 1647–1661. doi: 10.1007/s10531-013-0500-x
Nowinszky, L., and Puskás, J. (2017). “Light-trap catch of insects in connection with environmental factors,” in Biological Control of Pest and Vector Insects, ed. V. D. C. Shields (Croatia, Rijeka: Editorial BoD – Books on Demand), 97–118.
Price, B., and Baker, E. (2016). NightLife: a cheap, robust, LED based light trap for collecting aquatic insects in remote areas. Biodivers. Data J. 4:e7648. doi: 10.3897/BDJ.4.e7648
Rocha-Filho, L. C., and Garófalo, C. A. (2014). Phenological patterns and preferences for aromatic compounds by male euglossine bees (Hymenoptera, Apidae) in two coastal ecosystems of the Brazilian Atlantic Forest. Neotrop. Entomol. 43, 9–20. doi: 10.1007/s13744-013-0173-x
Roulston, T. A. H. (1997). Hourly capture of two species of Megalopta (Hymenoptera: Apoidea; Halictidae) at black lights in Panama with notes on nocturnal foraging by bees. J. Kans. Entomol. Soc. 70, 189–196.
Santos, L. M., and Melo, G. A. R. (2015). Updating the taxonomy of the bee genus Megalopta (Hymenoptera: Apidae, Augochlorini) including revision of the Brazilian species. J. Nat. Hist. 49, 575–674. doi: 10.1080/00222933.2014.946106
Santos, L. M., and Silveira, F. A. (2009). Taxonomic notes on Megalopta Smith, 1853 (Hymenoptera: Halictidae: Augochlorini) with a synopsis of the species in the state of Minas Gerais, Brazil. Zootaxa 2194, 1–20. doi: 10.11646/zootaxa.2194.1.1
Shaver, T. N., Lingren, P. D., and Marshall, H. F. (1997). Nighttime Variation in Volatile Content of Flowers of the Night Blooming Plant Gaura drummondii. J. Chem. Ecol. 23, 2673–2682. doi: 10.1023/a:1022550607873
Soares, N. C., and Morellato, L. P. C. (2018). Crepuscular pollination and reproductive ecology of Trembleya laniflora (Melastomataceae), an endemic species in mountain rupestrian grasslands. Flora 238, 138–147. doi: 10.1016/j.flora.2016.12.005
Somanathan, H., and Borges, R. M. (2001). Nocturnal Pollination by the Carpenter Bee Xylocopa tenuiscapa (Apidae) and the Effect of Floral Display on Fruit Set of Heterophragma quadriloculare (Bignoniaceae) in India 1. Biotropica 33, 78–89. doi: 10.1646/0006-3606(2001)033[0078:npbtcb]2.0.co;2
Somanathan, H., Borges, R. M., Warrant, E. J., and Kelber, A. (2008). Visual ecology of Indian carpenter bees I: light intensities and flight activity. J. Comp. Physiol. A 194, 97–107. doi: 10.1007/s00359-007-0291-1
Velez-Ruiz, R. I. (2015). A Revision of the Bee Genus Ptiloglossa Smith (Hymenoptera: Colletidae: Diphaglossinae) Ph.D. thesis. Brookings SD: South Dakota State University.
Viana, B. F., Kleinert, A. M. P., and Neves, E. L. D. (2002). Comunidade de Euglossini (Hymenoptera, Apidae) das dunas litorâneas do Abaeté, Salvador, Bahia, Brasil. Ver. Bras. Entomol. 46, 539–545. doi: 10.1590/s0085-56262002000400008
Warrant, E. J., Kelber, A., Wallén, R., and Wcislo, W. T. (2006). Ocellar optics in nocturnal and diurnal bees and wasps. Arthropod Struct. Dev. 35, 293–305. doi: 10.1016/j.asd.2006.08.012
Wcislo, W. T., Arneson, L., Roesch, K., Gonzalez, V., Smith, A., and Fernández, H. (2004). The evolution of nocturnal behaviour in sweat bees, Megalopta genalis and M. ecuadoria (Hymenoptera: Halictidae): an escape from competitors and enemies? Biol. J. Linn. Soc. 83, 377–373. doi: 10.1111/j.1095-8312.2004.00399.x
Keywords: nocturnal bees inventory, crepuscular bees, apifauna survey, sampling method, floral scents, volatile organic compounds, 2-phenylethanol
Citation: Martínez-Martínez CA, Cordeiro GD, Martins HOJ, Kobal ROAC, Milet-Pinheiro P, Stanton MA, Franco EL, Krug C, Mateus S, Schlindwein C, Dötterl S and Alves-dos-Santos I (2021) Floral Volatiles: A Promising Method to Access the Rare Nocturnal and Crepuscular Bees. Front. Ecol. Evol. 9:676743. doi: 10.3389/fevo.2021.676743
Received: 05 March 2021; Accepted: 21 June 2021;
Published: 20 July 2021.
Edited by:
Sara Diana Leonhardt, Technical University of Munich, GermanyReviewed by:
Hannah Burger, University of Konstanz, GermanyCopyright © 2021 Martínez-Martínez, Cordeiro, Martins, Kobal, Milet-Pinheiro, Stanton, Franco, Krug, Mateus, Schlindwein, Dötterl and Alves-dos-Santos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guaraci D. Cordeiro, Z3VhcmFkY0BnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.