 Fabiola Parra-Rondinel1
Fabiola Parra-Rondinel1 Alejandro Casas1,2*Domingo Begazo3Amalia Paco3Eusebia Márquez4Aldo Cruz5
Alejandro Casas1,2*Domingo Begazo3Amalia Paco3Eusebia Márquez4Aldo Cruz5 Jorge Segovia4
Jorge Segovia4 Ignacio Torres-García2,6Mariana Zarazúa2Luis Lizárraga4Juan Torres-Guevara1
Ignacio Torres-García2,6Mariana Zarazúa2Luis Lizárraga4Juan Torres-Guevara1- 1Departamento Académico de Biología, Facultad de Ciencias, Universidad Nacional Agraria La Molina (UNALM), Lima, Peru
- 2Instituto de Investigaciones en Ecosistemas y Sustentabilidad (IIES), Universidad Nacional Autónoma de México, Morelia, Mexico
- 3Centro Andino de Educación y Promoción José María Arguedas (CADEP), Cusco, Peru
- 4Universidad Nacional San Antonio Abad de Cusco (UNSAAC), Cusco, Peru
- 5Centro de Investigaciones de Zonas Áridas, Universidad Nacional Agraria La Molina, Lima, Peru
- 6Escuela Nacional de Estudios Superiores (ENES), Universidad Nacional Autónoma de México, Morelia, Mexico
The Andean region is one of the areas with the earliest signs of food production systems and highest agrobiodiversity of the world, which resulted from millennia of domestication in a context of high ecosystem heterogeneity and human cultures valuing diversity for risk management. FAO has reported nearly 4000 varieties of cultivated potatoes still grown in the Andes, 3000 of them currently occurring in Peru. Such diversity has enormous sources of variation in wild (atoq papa) and weedy (araq papa and k’ipa papa) potatoes that coexist with crops, but their variation, interactions and mechanisms influencing diversification processes still require studies. In order to have a panorama of the variation and mechanisms influencing it in a regional setting, we studied biocultural factors favoring potatoes diversity in communities of Cusco and Apurimac, Peru. Our study documented the regional variation of wild, weedy, and cultivated potatoes recognized by local Quechua people and conducted semi-structured interviews to document their use, cultural value, and strategies of gene flow management implemented. We also studied their phenology, floral biology, flower visitors, and conducted experimental crosses between the wild S. candolleanum and 30 varieties of cultivated potatoes. We identified the wild potatoes S. acaule, S. brevicaule and S. candolleanum and 53 varieties of araq papa used and managed by local people. The latter provide nearly one third of the annual consumption of tubers by people interviewed and are, therefore, highly valued, maintained and managed in crop fields (chacras). People recognized that crosses between wild, weedy, and cultivated potatoes occur, and identified flower visitors and frugivores consuming their berries. Overlap of blooming periods and flower visitors of wild, weedy, and cultivated potatoes was recorded. Almost all flower visitors are shared among the different potato species and varieties, the bumble bees being particularly relevant in pollination of all taxa studied. We recorded seed production in nearly 35% of the experimental crosses. K’ipa papas are sets of mixtures of plants resulting from remaining tubers of cultivated potatoes, but also those from seeds that may result from hybridization of wild, weedy, and cultivated potatoes. Since local people commonly use k’ipa papa varieties and some of them are kept for planting in chacras, sexual reproduction in k’ipa papas is possibly one main mechanism of variation and source of new varieties of crops. Maintaining wild and weedy potatoes, and the natural and cultural mechanisms of gene flow is crucial for in situ conservation and generation of potato variation.
Introduction
The Andean region has been recognized as one of the areas with the earliest signs of food production in the Americas and the highest agrobiodiversity of the world (Harlan, 1975; Hawkes, 1983; Vavilov, 1992; Brush et al., 1995; Zimmerer, 2003, 2016; Brush, 2004; Brack and Bravo, 2006; Clement et al., 2021). Agrobiodiversity is a term commonly associated to genetic diversity of crops, but it is a concept that includes the general richness and diversity of species of plants, animals, macromycetes, lichens, and microorganisms (bacteria, cyanobacteria, and fungi) that interact with crops in agroecosystems, as well as the heterogeneity of ecosystems in agricultural landscapes (FAO, 2016; Casas and Vallejo, 2019). In Peru, Brack and Bravo (2006) identified 180 native crop species, nearly 1700 plant species under some management type, most of them semi-domesticated, and nearly 4700 wild plant species whose products are used by people of the Andean, Amazonian, and the arid and semiarid zones of the Coastal region. Such diversity is representative of the remarkable agrobiodiversity of the area.
Domestication is an evolutionary process continually operating in the Andean region for more than 8,000 years, in contexts where ecosystems, technology and culture have also been dynamic. Human culture is one of the main drivers favoring crop attributes like forms, colors, textures, resistance against pests and other environmental challenges for food production, or that which confers adaptability to different elevations, soil types, rainfall regimes or conditions for storing the products harvested (Casas et al., 2016). Variation is the necessary condition for this process to operate, and such variation may be continually provided by wild and weedy potatoes when gene flow occurs (Casas et al., 2016). This study analyses the possible different sources of variation occurring in a particular area (the Andean region of southern Peru), the natural and human processes influencing on their state (ecological interactions and management techniques, respectively), and their possible role to explain the mechanisms favoring gene flow among such sources of variation. It is one approach to contextualize the processes influencing the population genetics of wild, weedy and crop potatoes in the area.
Native Andean crops have had an exceptional process of diversification, which can be explained in part due to the millenary history of food production and domestication in the region; but also, because local human cultures outstandingly value diversity as a basis to manage factors that put in risk the regional people’s subsistence. Management of diversity has been the keystone of what anthropologists have called risk management strategy, through the control of multiple vertical levels in Andean mountains (Murra, 1983). In addition, the outstanding Andean agrobiodiversity can be explained because of the also high heterogeneity of ecosystems and landscapes of the region (Brack and Bravo, 2006; Brack et al., 2008), which determines an extraordinary mosaic of microhabitats that in turn influences local adaptations and diversification of organisms through natural evolutionary processes. Finally, the extraordinary diversity is a result of reproductive interactions and gene flow between crops and their wild and weedy relatives. Potatoes and other Andean tubers stand among these exceptionally diverse components of agrobiodiversity, which are continually interacting with their non-crop relatives and have been domesticated by diverse cultures of the region. All wild, weedy, and cultivated potatoes are therefore important parts of the Andean agrobiodiversity.
Botanists and agronomists have identified and classified nearly 4,000 varieties of potato crops (Tobin et al., 2016), about 2800 to 3300 of them distributed in Peru (De Haan and Rodríguez, 2016). These varieties belong to the genus Solanum and have been considered to include 8 species (Hawkes, 1990), 7 species (Spooner and Salas, 2006), and, more recently, 4 species (Solanum tuberosum L., Solanum ajanhuiri Juz. and Bukasov, Solanum curtilobum Juz. and Bukasov and Solanum juzepczukii Juz., according to Spooner et al., 2014). Controversies about the correct status of classification of landraces, varieties and species involving the organisms recognized as potatoes have taken long time and continue until present, since the limits among these taxonomic units are rather fuzzy. Studies by taxonomists, throughout time, have gained new research tools to clarify their status and they still conduct research to understand the nature of potatoes, with increasing precision (Hawkes, 1990; Spooner and Salas, 2006; Spooner et al., 2014). Even fuzzier and complicated is the information documented from anthropological research. For instance, the project ‘in situ conservation of native crops and their wild relatives’ in Peru, documented from ethnographic approaches that people of the Peruvian Andes recognize and name nearly 9,000 potato varieties, even after a careful work for discarding synonyms (Velásquez-Milla et al., 2011; Torres-Guevara et al., 2017). These traditional varieties are recognized based on potato attributes and qualities that are difficult to measure through conventional methods used by botanists and agronomists like food quality, pulp texture, properties for processing, among others (Velásquez-Milla et al., 2011). However, the resulting ciphers from both botanical and ethnographic studies are equally impressive and both sources of information deserve deeper integral studies. The high number of varieties of potatoes in the Andean region reveals the existence of a great regional biocultural diversity, not only in terms of varieties adapted to particular environments and cultural contexts, but also in relation to different types of production systems, technological elements, economic relations, and social organization (Brush, 1992; Brush et al., 1992; Lastra et al., 2019; Zimmerer et al., 2020).
The great diversity of potatoes in the Andean region is influenced by the presence of species of wild relatives, which are grouped within the Quechua names sacha papa or atoq papa (meaning “potatoes of the mountain” and “fox potatoes,” respectively) sometimes used as medicine but that are not part of the food system (De Haan et al., 2007; Figure 1). Wild relatives of potatoes include nearly 200 species, according to Hawkes (1990) or about 100, according to Spooner et al. (2014), of the genus Solanum section Petota, distributed from southwestern United States to south central Chile (Huamán and Spooner, 2002; Spooner and Salas, 2006). Wild potatoes have the capacity to maintain reproductive interaction and interchange of genes with crop potatoes, as demonstrated by several studies (Jackson and Hanneman, 1999; Bradshaw et al., 2006; Jansky, 2006; Scurrah et al., 2008; Bethke et al., 2017; Maune et al., 2018). In the study area of Apurimac and Cusco, Spooner et al. (1999) reported the presence of 22 wild species, but in a more recent taxonomic treatment by Spooner et al. (2014) those taxa were grouped into the following 9 species: S. candolleanum Berthault, S. acaule Bitter, S. brevicaule Bitter, S. buesii Vargas, S. raphanifolium Cárd. and Hawkes, S. incasicum Ochoa, S. laxissimum Bitter, Solanum violaceimarmoratum Bitter, and S. pillahuatense Vargas. Solanum candolleanum is considered by Spooner et al. (2014) to include more than 30 taxa previously described as species now considered to be synonyms, among them S. bukasovii Juz., an important taxon of our study. Ahead, we will use the name S. candolleanum to refer to this taxon, following Spooner et al. (2014). Gene flow between wild and cultivated potatoes has been documented for some species and, although more research is needed, it has been identified as a primary source of variation in cultivated varieties. It is a continuous, silent process, which is possible because of the coexistence of wild and domesticated populations. But, besides atoq papas, different taxa called in Quechua araq papas and k’ipa papas also coexist and may interact with crop potatoes. However, their interactions have been scarcely studied.

Figure 1. Wild, weedy, and cultivated potatoes of the Andean region of southern Peru. (A) Solanum candolleanum, the atoq papa or sacha papa used in this study for experimental crosses with cultivated potatoes; the photograph shows from top to down the general aspect of the plant in a rocky environment, details of flowers, fruits and the small tubers; (B) araq papas, from top to down it is possible to see the aspect of the weedy plants associated to maize, and the size of tubers compared with the atoq papa; (C) k’ipa papas, the photos show aspects of some plants in fallow agricultural fields cultivated with potatoes the year before; plants emerged may derive from non-harvested tubers, as well as recruitment of plants from seeds.
The araq papas (approximately meaning “toward potatoes” in Quechua), are local taxa that behave like weedy plants and seem to have considerable intraspecific diversity. These plants are spread from Bolivia to Ecuador, where they are collected and consumed by Andean farmers, who name them as follows: Araq papa (central and southern Peru), chayka papa (central Peru), papa curao (central and northern Peru), lelekkoya (Bolivia), and semillu (Bolivia) (De Haan and Rodríguez, 2016). Araq papa is commonly associated with corn in chacras (crop fields), and their root systems typically have a long stolon and thick tuber skin (De Haan et al., 2007). Araq papas have been reported to be collected by local people, being an important source of food for them (Brush, 1992). Araq papas have been managed by Andean peoples from ancient times and are at present valuable resources; they coexist with wild and crop potatoes, and even planted or grown by farmers, maintaining their own cultural and biological role, as we document in this study. Araq papas were originally classified as Solanum aracc-papa by the Russian researchers Sergei Vasilievic Juzepczuk and Vladimire Alekseevich Rybin. More recently other authors considered them as Solanum tuberosum subsp. andigena; in the classification by Spooner et al. (2014) this taxon would correspond to the Andigenous group. However, their high morphological variation raises the question whether one single taxon is involved, rather suggesting the occurrence of polyploidy, hybridization and other processes that make araq papas to be a complex of taxa that deserves deeper studies.
Other local taxa are grouped under the name of k’ipa papa (meaning approximately “from potatoes”, in Quechua) whose exact nature remains unknown (Figure 1). According to Bonnave et al. (2015), k’ipa “is a Quechua term meaning volunteer plants of a crop species emerging in sites of former fields in the years following cultivation of this crop”. K’ipa papa are remains of potatoes cultivated during a previous cultivation period, that is, plants deriving from tubers of domesticated or cultivated potatoes (De Haan et al., 2007) that were not collected by the agriculturalists (Figure 1). However, it is common that people identify new varieties derived from k’ipa papa, which suggests that these may in addition result from seeds and, therefore, sexual reproduction, a similar system documented for mashua (Tropaeolum tuberosum Ruiz et Pavon; Velásquez-Milla et al., 2011) and oca Oxalis tuberosa Molina (Bonnave et al., 2015). How frequent the interaction is of atoq papas, araq papas and k’ipa papas with potato crops, and how people use and manage these plants to favor their interaction, are relevant questions to understand biological and cultural processes associated with potato diversification. Our study aspires to contribute to answer some of these questions. The main research question motivating our study was what natural and human cultural mechanisms are relevant to analyze gene flow among wild, weedy, and cultivated potatoes. In particular, what processes influence the movement of pollen and the effectiveness of interspecific and inter-varietal cross-pollination, and what factors participate in seed dispersal and movement of tubers in the space. We hypothesized that if wild, weedy, and crop potatoes have synchronized flowering period, share flower visitors and their crosses are successful, then gene flow among them would be effective. This general hypothesis allows visualizing the need of different sources of information related to pollination biology, seed dispersal and movement of vegetative propagules. Although some cultivated potatoes are sterile, and pollination and seed dispersal are irrelevant, wild and some weedy and cultivated potatoes’ seeds are effective propagules and their dispersal is relevant. We started our research on this topic by enquiring about what people know about flower visitors, and birds and mammals consuming fruits or the success of dispersal by simple fruit falling. The movement of tubers was expected to be particularly active in cultivated and weedy potatoes, influenced by their use value, which in turn influence the tolerance of weedy potatoes in crop fields, and propagation of tubers from desirable varieties from place to place, and other actions favoring their distribution and abundance in chacras.
Therefore, our study analyses social and ecological factors in the Andean area of southern Peru, in relation to the use and management of specific and intraspecific taxa of Solanum, considered to be potatoes and potatoes-relatives, which may contribute to explain mechanisms influencing potato diversification. Research on these topics is necessary not only to understand processes of potato diversification, but in addition for developing social and ecological bases for the conservation of those processes that dynamically generate agrobiodiversity and that are indispensable conditions to the in situ maintenance of this valuable biocultural heritage (Casas et al., 2016; Torres-Guevara et al., 2017).
Methods
Study Area
Ethnobotanical studies and collecting of plant specimens were conducted in the communities of Quiñota and Pallpa pallpa, province of Chumbivilcas, in Cusco, as well as in Haquira, Huancacalla Chico, San Juan de Llaccua, Cconchayoc, Ccocha and Patahuasi province of Cotabambas, in the Department of Apurimac, at the Sierra Sur of Peru (Figure 2). In each community we interviewed as much people as possible, according to their time availability and disposition to collaborate in the study. In this research, our effort was focused on inventorying the species and varieties of potatoes existing in the region, their use and management. Elevation, climate aspects and population of the sites studied are summarized in Table 1.
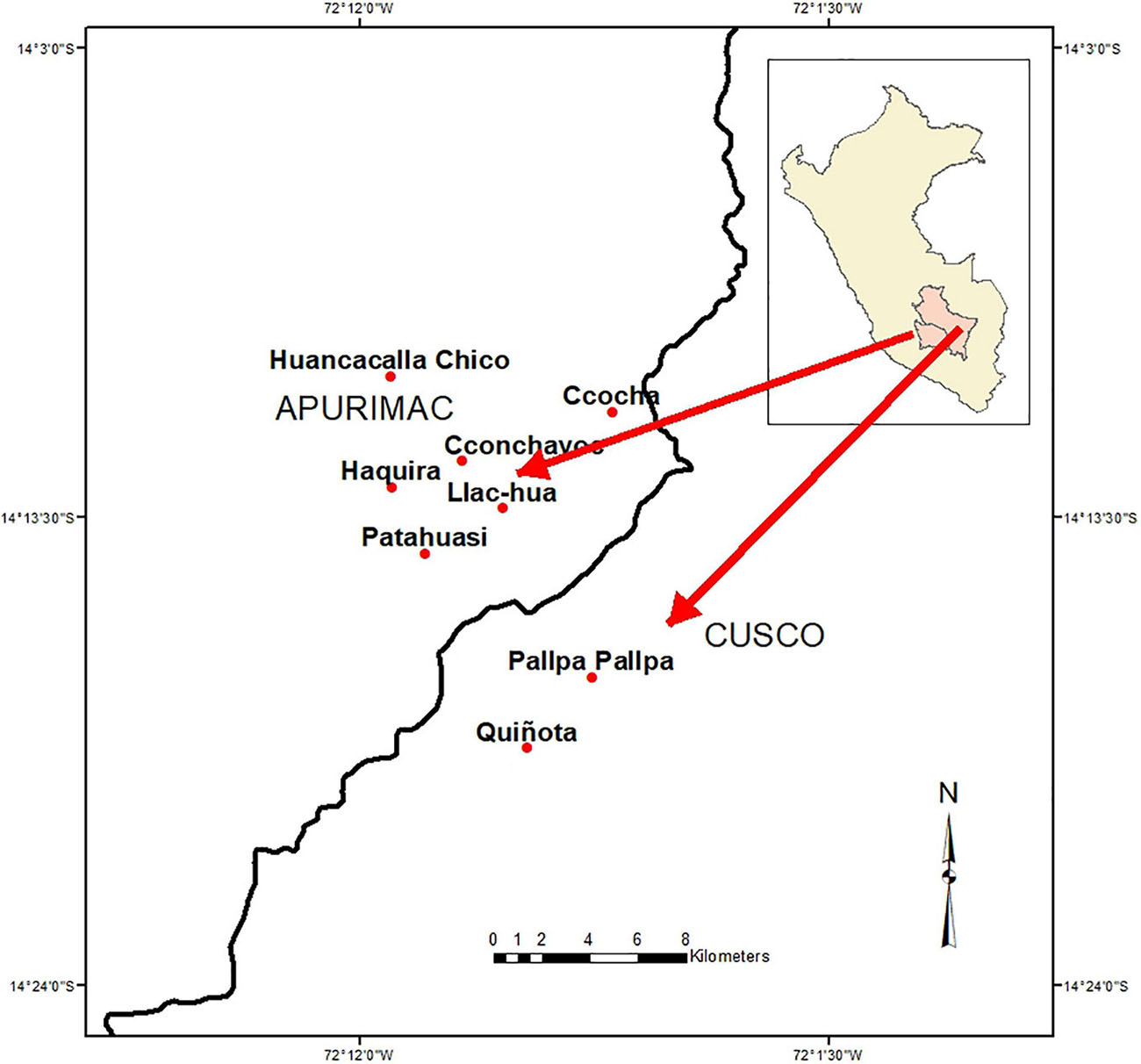
Figure 2. Map of the study area in Cusco and Apurimac regions at southern Peru. Sites Quiñota and Pallpa pallpa, in Cusco, as well as in Haquira, Huancacalla Chico, Llaccua, Cconchayoc, and Patahuasi in Apurimac (red dots).
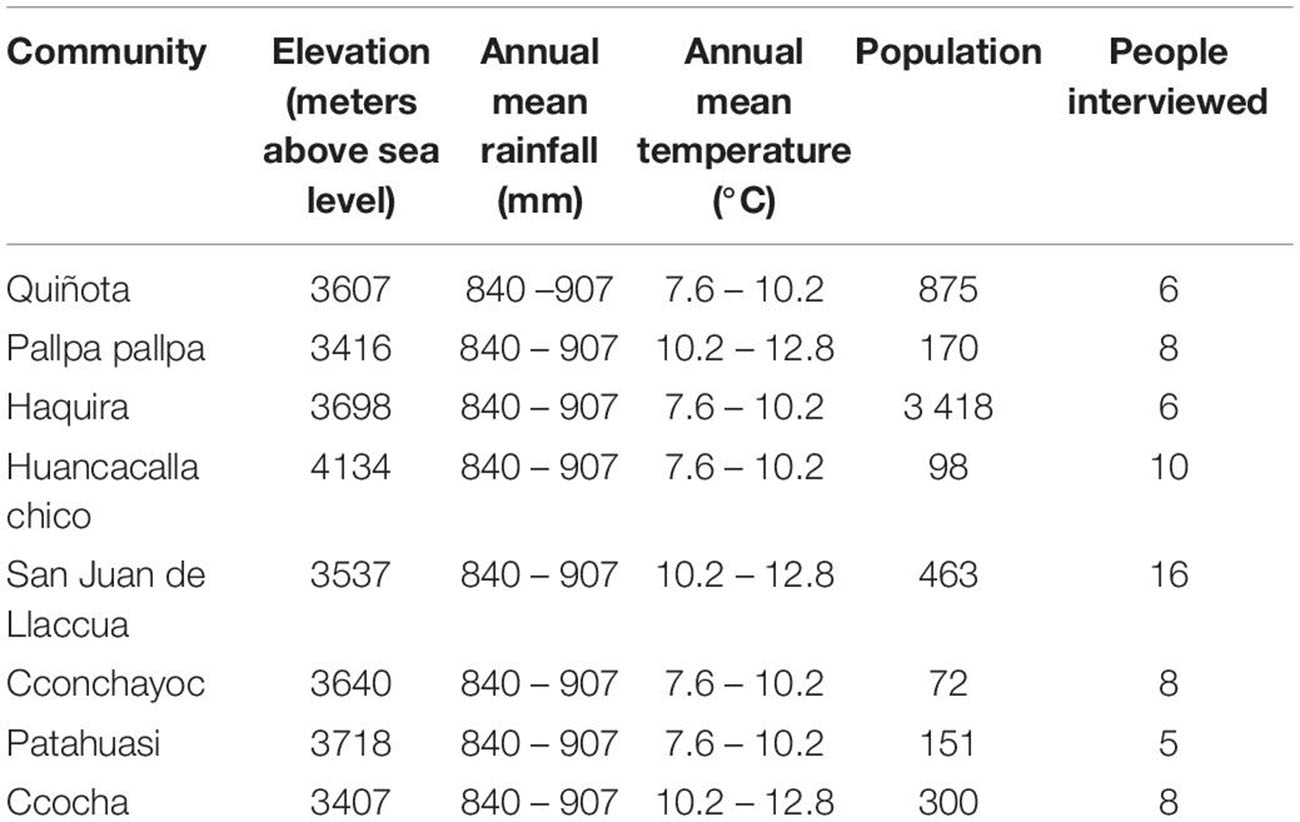
Table 1. Environmental and social aspects of the communities studied at the Andean regions of Apurimac and Cusco in southern Peru.
Ethnobotanical Studies
Semi-structured interviews were carried out with a total of 67 persons (42 women and 25 men). Interviews were performed individually and in focus groups 2-10 people each to allow the interchange of experiences and views among participants. We asked participants to elaborate oral free lists (Newing et al., 2011) of the varieties of potatoes (atoq, araq, and k’ipa papas) and then asked questions in relation to their names and attributes, their distribution, association, phenological and morphological aspects, period of availability of tubers, forms of use, preparation, management, commercialization, storing techniques and eventual rise of new variants (Supplementary Material 1).
We collected and prepared voucher specimens of all varieties of atoq, araq papas, and samples of k’ipa papas available in the crop fields of people interviewed, each specimen was documented with the information mentioned above when collected. Photographs in crop fields and in fresh collected material were taken, and the plant material herborized. The plants collected were labeled to further collecting tubers from them. Additional samples of leaves were collected and maintained in bags with silica-gel for further molecular studies. Samples of k’ipa papas were collected from random samples of 30 plants per chacra.
Preparation of Experiments in the Field
We established an experimental field in the community of Huancacalla Chico sector Patabamba, where we got the permit for planting 30 varieties of native crop potatoes, and where naturally occur the wild species S. candolleanum and S. acaule, which grow associated to or near the chacras. Sowing of crop potatoes started the 16th of November 2017, maintaining 0.40 m among plants and 0.90 m among rows. Weeding was unnecessary but hilling was conducted in January 2017, 64 days after tuber planting. We used cow dung to fertilize soil in plots (nearly 20 g for each tuber), and after 64 days we practiced an additional application.
Phenology
We recorded directly different events of wild, weedy, and cultivated potatoes. We recorded the time when cultivated potatoes were planted and when the terrains where araq papas grew were tilled. In the case of Solanum candolleanum we recorded the time of the emergence of the aerial plant parts. Later, the blooming starting, flowering peak and the end of the period. Finally, the time when fruiting started and finished.
Flower Visitors
For identifying the flower visitors and their possible role in pollination we established 6 videorecorders (three for wild individuals and three for cultivated potato varieties). We carried out continual videorecording for sets of 4 consecutive days before dawn (5:00 h) until nearly the sunset (17:00 h). Each camera videorecorded events occurring in one to four flowers. Then, the videorecorded tapes were reviewed to register the time and duration for every event of an insect visiting a flower. A given visit event was considered as an effective visit if the insect touched the reproductive structures of flowers (Dafni, 1992; Bumrungsri et al., 2008; Faegri and Van Der Pijl, 2013). During this period, our team collected all flower visitors using entomological nets, and the insects captured were preserved for further identification in the Laboratory of Entomology of the Faculty of Biology at the Universidad Nacional de San Antonio Abad de Cusco, following classification keys by Michener (2000), Silveira et al. (2002), and Fernández and Sharkey (2006). The specimens were deposited in the collection of the laboratory referred to, under the Márquez-Castellanos collection numbers.
Breeding Experiments
Experiments were conducted in the field, inter-crossing individuals from 30 different varieties of cultivated potatoes and the wild relative species Solanum candolleanum. Flowers of all crop varieties and the wild relative were pollen receptor and donors (female and male, respectively), each cross having from 3 to 5 repetitions. The fruit-set and seed number were calculated for all successful treatments. Seeds from all successful fruits were germinated and the percentage of germination was recorded. We emasculated 2-12 flower buds per plant. Then we removed other neighboring buds and flowers in the branches selected for the experiment and covered the flowers for the experiment with mosquito net bags. We obtained pollen from each cultivated variety and the wild relative from samples of flowers, which released pollen stimulated by vibration of a diapason. The pollen collected in Petri dishes was put in the stigma of the receptive flower and covered again with the bags, labeling in each case the source of pollen and date (Eijlander and Stiekema, 1994; Bamberg, 2020). Fruit harvesting was conducted 25, 40 and 45 days after the crosses, when fruits were mature. The successful fruits were transported to the National University San Antonio Abad de Cusco (UNSAAC) at Cusco, where we obtained the seeds produced by cutting the berries transversally. Seeds were then washed with clean water and counted. Seed germination was tested in a greenhouse at the Regional Centre of Research in Andean Biodiversity (CRIBA). The number of germinating seeds was registered after treating them with gibberellic acid for 24 h (see Supplementary Material 2).
Ploidy Levels
For evaluating the ploidy level, we followed the technique developed by Huamán (1995). We collected terminal leaflets from plants of each variety planted in our experimental field, as well as from S. candolleanum. Leaflets were maintained in 70% ethanol for one hour, then dried and treated with one or two drops of a solution of 1 g of KI, 1 g of I and 100 ml of 80% ethanol for five minutes. Leaflets were cut to obtain epidermic tissue placed in a slide, a drop of glycerine, coverslip, and observed in a microscope. We counted the chloroplasts of the stomatic guard cells estimating the ploidy following the relations 2x:7- 8 chloroplasts, 3x: 9 -11, 4x:15 - 16 chloroplasts (Huamán, 1995).
Cultural Salience Analyses
Frequencies and order of mention of the varieties were recorded and the Smith and Sutrop salience indexes were calculated for each variety. The cognitive importance was estimated through free listing data with the index of Sutrop (S) with the formula S = F/(N mP), where F represents the frequency of the variety mentioned, N the total number of interviewed people, and mP is the medium position in which the variety was named (Sutrop, 2001; Rangel-Landa et al., 2016). We in addition calculated the index developed by Smith S = ((∑(L-Rj + 1))/L/N, where S is the average rank of an item across all lists in the sample, weighted by the lengths of the lists in which the item actually occurs; L is the length of (number of items in) a list; R the rank of the item in the list (first = 1); and N the number of lists in the sample (Smith and Borgatti, 1998). We calculated these indexes with the software FLAME v1.0 (Pennec et al., 2012). Use and management techniques recorded, as well as information on attributes recognized by people, were systematized following descriptive qualitative methods.
Results
Quechua Nomenclature, Use and Management of atoq papa, araq papa, and k’ipa papa
All people interviewed, said to know the wild relatives of potatoes, which are called atoq papa, as well as those of other tubers like oca (Oxalis tuberosa), mashua (Tropaeolum tuberosum) and olluco (Ullucus tuberosus Caldas). People mentioned that these plants grow in the rocks, fences, around or inside the chacras. These plants are removed to avoid competition with crops; according to people these plants emerge when the rainy season starts. They said that these plants are medicinal, fruits are used for curdling the milk to prepare cheese and added that their ancestors used their tubers during hard times of hunger and in rituals as payments to Pachamama, but no information about their preparation for consumption was remembered. They have seen that their fruits and seeds are consumed and dispersed by domestic animals, mainly horses and donkeys, as well as some regional birds like the zorzal chiguanco (Turdus chiguanco Lafresnaye and d’Orbigny) and deer (Odocoileus virginianus peruvianus Gray). They identified the waña papa (Solanum acaule), the atoq lisa or k’ita lisa (Solanum candolleanum), which is medicinal, used as a plaster for treating traumas, and the atoq añi or k’ita añi (Solanum brevicaule) whose flowers and leaves are used to prepare an infusion taken for illnesses of the prostate.
Another important group is araq papas. We recorded 53 varieties of these plants as shown in Table 2, all of them referred to in the interviews and focus groups and most of them occurring in the chacras of the households interviewed. In 2018, the winner of the “Araq papa Fair” in Quiñota, Cusco was a woman showing 76 varieties of araq papas, which indicates the great diversity of araq papas existing in the region (Figure 3). Araq papas grow at elevations from 3,000 m to 3,730 m, mainly in association with maize and fava beans (Vicia faba L.), but also with tarwi (Lupinus mutabilis Sweet), quinua (Chenopodium quinoa Willd.), kiwicha (Amaranthus caudatus Speg.) and other crops. Araq papas may be associated to potatoes, but people prefer to separate areas destined to these plants since the long stolon of araq papas competes with cultivated potatoes. People consider that harvesting of araq papas may cause damage to wheat and other crops since they have to remove the ground to collect tubers, and they avoid some associations that can be damaged by this way of harvesting.
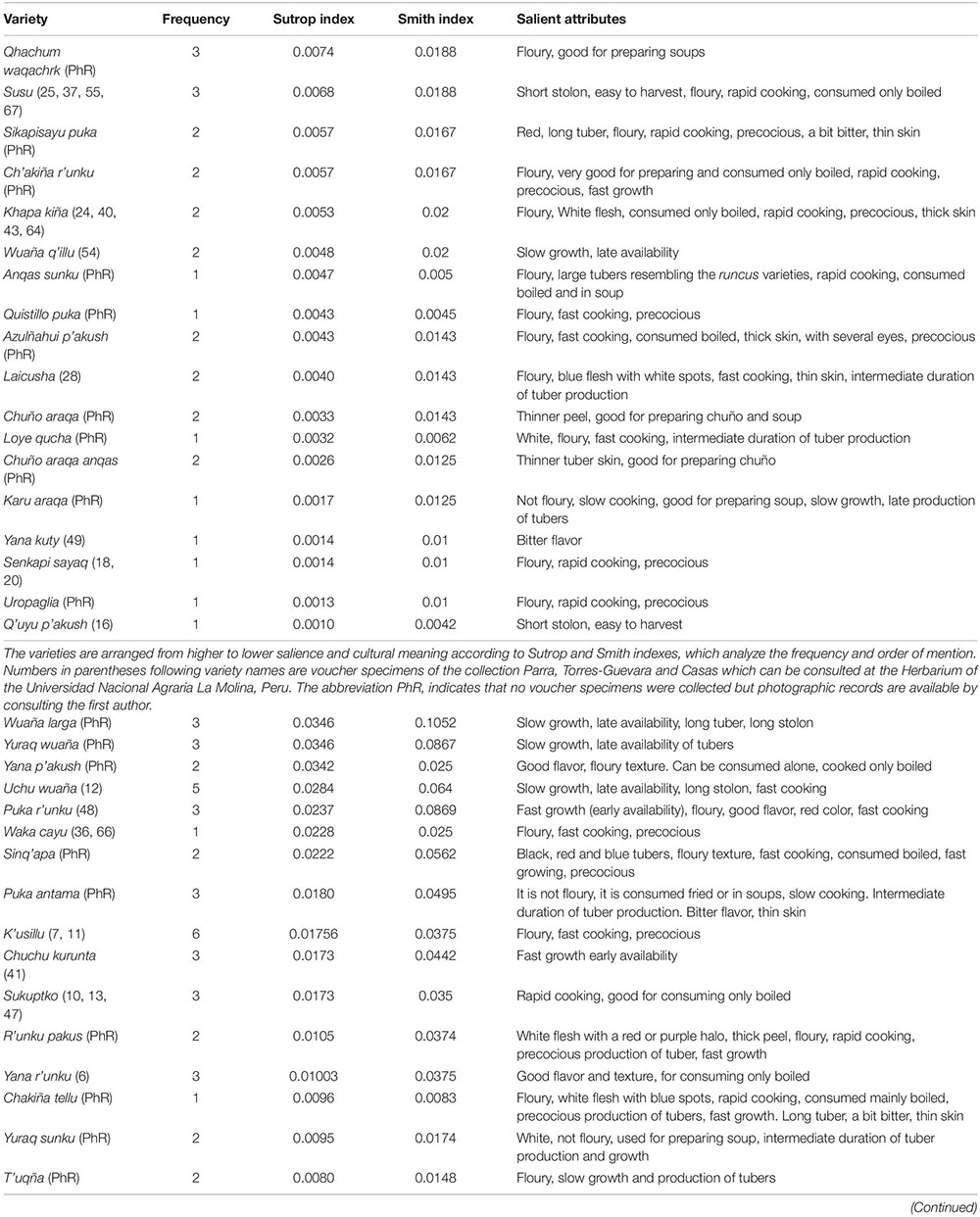
Table 2. From top to bottom, the most salient araq papa varieties mentioned by 67 persons and found in the chacras sampled (n = 20) and their attributes.

Figure 3. The Quiñota Araq papa Fair. Aspect of the fair showing the high variation of araq papa commercialized and interchanged among people of region. It can be noticed that some of the varieties recorded in this study are shown and commercialized in the fair.
With some exceptions referred to below, araq papas are not cultivated plants but grow “spontaneously” in chacras; however, if a person loses them or if he/she wants another interesting variant, he/she looks for recovering or having new varieties by obtaining tubers from other persons. People mention that during the ayllu, a collective work for helping to each other to harvest crops, there is a good opportunity to identify and get new varieties. Araq papas have characteristically thicker tuber skin and stolon much longer than potatoes (Figure 4). Their flower and leaf characters are highly variable, as well as colors, textures and other tuber attributes. Araq papas coexist with wild and domesticated potatoes, and their berries may result from sexual crosses; these berries remain in the chacras until their maturation and their seeds become established in the crop fields. The actual seed dispersal by domestic and wild frugivorous animals consuming their berries is yet to be tested. Araq papas depend on the land tilling, since according to local people, if a crop field is abandoned for 3 to 4 consecutive years, araq papas disappear from the fallows. This feature and its “spontaneous” emergence in tilled land led to the notion that araq papas are weedy plants.

Figure 4. Aspects of the morphology of araq papas. The upper row of images shows aspects of some of the different flower types recorded among the araq papas. The lower row of images shows a panorama of the fruits, the long stolon characterizing these plants (the person showing the stolon is Jorge Segovia, co-author of this study), and a transversal section of the tuber showing aspects of the thick skin and inner tissue, which is highly variable. The person appearing in the photograph of Figure 4 is Jorge Segovia, co-author of this manuscript, who agreed with both text and images contained in it.
Araq papas are outstandingly important for the Andean people food security. From people interviewed in this study we learned that their household’s food is based on these tubers from 2 to 4 (average 3.7 ± 1.1) months throughout the year, from December (some varieties producing tubers even before this month) to May. Potatoes, maize, fava beans, quinoa, and tarwi (Lupinus mutabilis), are the main staple crops for regional diet, but araq papas provide food when these crops are growing in the chacras, during the period of most critical scarcity of staple crops before their harvesting. In other words, nearly one third of the annual consumption of tubers of the regional people depends on araq papas.
Variation is high, but some varieties are the most frequent in chacras and were also mentioned through the free listing method, which allowed us identifying the most salient varieties. Table 2 indicates the most salient varieties and their attributes, according to local people. Salience indicates which varieties are preferred and more widely distributed in the region, which is important for further studies on their contribution on gene flow to crop populations. People practice artificial selection on araq papas when deciding what variety to promote or enhance in their chacras. They enhance those varieties producing larger tubers, with floury texture and good flavor, those that are faster to cook, those more resistant under storage conditions, and those producing shorter stolon since these are easier to harvest without damaging the associated crops. Other particular attributes are special colors and forms of tubers, how fast the tubers are produced during the production season and the ease to prepare the dried and frozen “chuño” (Table 2). Selection is not unidirectional since several attributes are valued for different purposes. For instance, the latter attribute, preparation of chuño, is related to a thinner tuber skin, which is therefore selected in some varieties, but in other varieties the thick skin is preferred. Commercialization of araq papas is important for some household, they sell one arroba (11.5 kg) for $30-40 Soles ($9.8 - 11.2 United States Dollars), while one arroba of cultivated potato has a value of $28-30 Soles ($7.8-8.4 United States Dollars) in the regional market.
K’ipas are considered by local people to be remains of tubers from potatoes cultivated the previous year in a chacra. However, as they say, “from time to time a new variety of potato may be found among the k’ipas”, a fact that suggests that together with plants derived from tubers, the fallows may contain plants derived from seed germination and, therefore, plants resulting from crosses among cultivated, weedy, and wild varieties are possible. K’ipas are valuable resources maintained and used by local people, complementing the main harvest of potatoes. Tubers of k’ipas are harvested and stored together with those of potatoes; therefore, their inclusion as part of the stock of “seeds” (the tubers to be planted are called “seeds”) for planting a new generation of potatoes. There is not a direct evaluation of hybridization and frequency of this process in k’ipas, but it would be particularly relevant.
Phenology
In cultivated potatoes, the vegetative growth from planting to blooming took 78 days, while in S. candolleanum it took 45 days and in araq papas 40 to 50 days from the emergence of the aerial plant part to the starting of flowering. The flowering peak was observed about 92 days after plating the cultivated varieties, 63 days after the emergence of S. candolleanum, and 76 days after tilling the ground in the case of araq papas. Blooming period is 27- 29 days in S. candolleanum, 26 to 29 days in araq papas and 25 to 29 days in crop potatoes. The general comparison of records of flowering monitored in wild and cultivated potatoes are shown in Figure 5. Although flowering peaks in wild and cultivated potatoes occur at different times, there is a general overlap of flowering that makes possible pollen interchange.
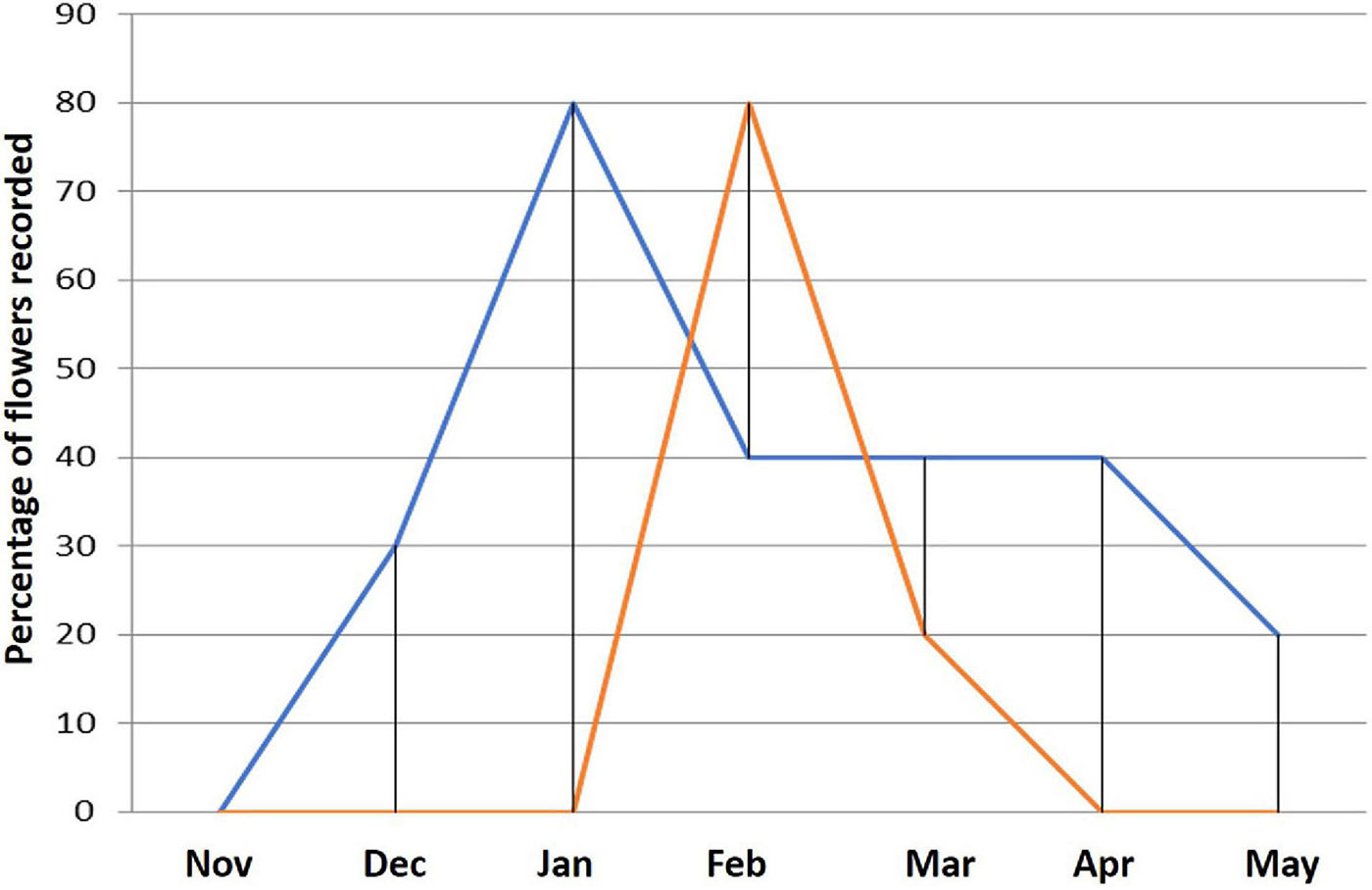
Figure 5. Floral phenology of wild and cultivated potatoes studied. The temporal production of flowers in the wild Solanum candolleanum (line blue) and varieties of cultivated potatoes (line red).
Pollination Biology
Duration of anthesis in wild and cultivated potatoes studied was 4-5 days, the process starting at dawn. During this period, we captured 38 specimens of flower visitors assigned to 11 species with some specimens unidentified (Table 3). Bombus opifex and Tetragonix sp. were collected but absent in all our videorecorded material, probably reflecting their scarcity.
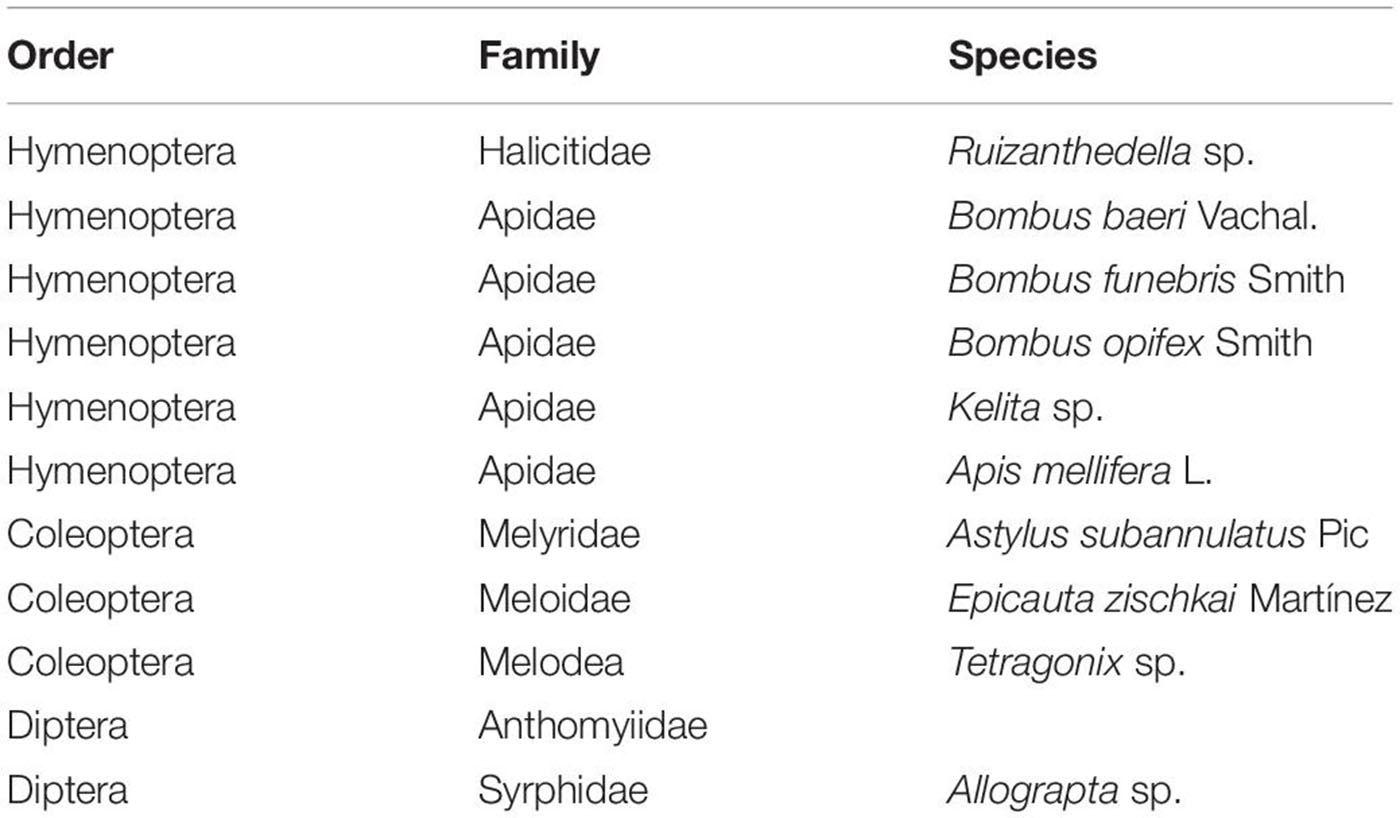
Table 3. Flower visitors identified in this study.
Kelita sp., Bombus baeri, and B. funebris are the species with the highest frequencies of visits all of them effective visits (Table 4, and Figure 6). Allograpta sp. and Anthomyiidae flies had high frequency of visits but nearly half and 3% of them were effective, respectively. Ruizathellia sp. and Astylus subannulatus had high proportion of effective visits but their visits frequency was low. Epicauta zischkai visited the flowers with low frequency and its visits were ineffective.

Table 4. Foraging behavior of the flower visitors identified (F, frequency of visits;%, percentage of visits).

Figure 6. Flower visitors shared by wild and cultivated potatoes studied. Some of the species identified visiting flowers of wild and cultivated potatoes sighted through the videorecording study of frequency, effectiveness, and duration of visits. (a) Kelita sp., (b1) Bombus opifex, (b2) detail of Bombus opifex© Alfredo F. Fuentes CC BY-NC 4.0. (Ueda, 2020) (c) Allograpta sp., (d1) Bombus funebris (d2) detail of Bombus funebris, (e) Anthomyiidae, (f) Astylus subannulatus.
As shown in Table 5, all flower visitors identified are shared among wild and cultivated potatoes. However, it is notorious that the highest frequencies of visits were recorded in Bombus funebris, Bombus baeri and Kelita sp. in the wild relatives, as well as in araq papas. Ruizanthellia sp., Allograpta sp. and Astylus subannulatus were more frequent in the cultivated potatoes with a medium effectiveness in their visits, as well as Anthomyiidae flies, which had low effective visits.
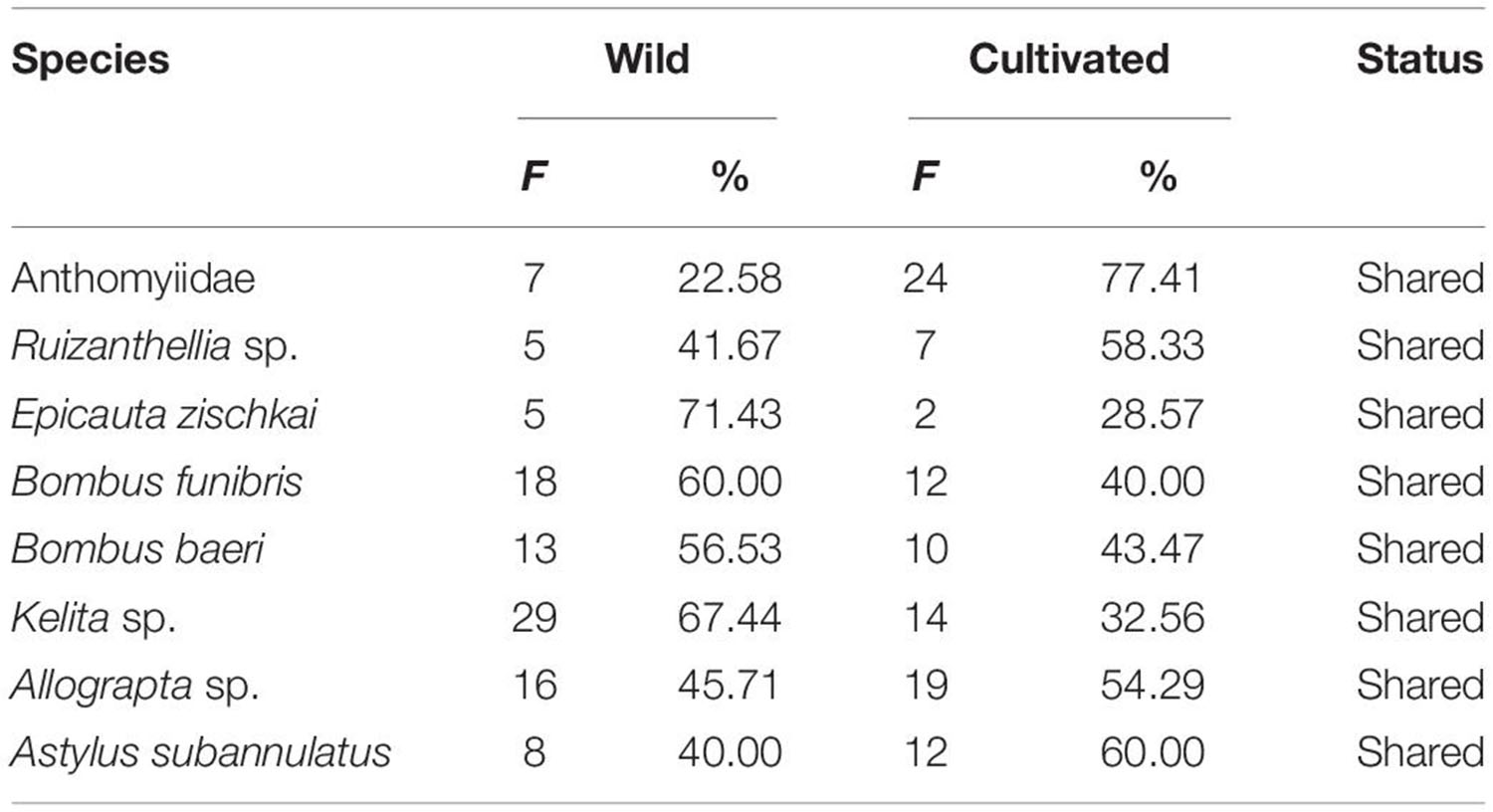
Table 5. Flower visitors recorded in flowers of wild and cultivated potatoes (F, frequency of visits;%, percentage of visits).
Visit to the same flower varied in four consecutive days (Table 6). In the wild potatoes, we recorded that from a total of 101 visits, the flowers received 10% of the visits during the first day, 64% during the second day, 15% during the third day, and 11% during the fourth day. In the cultivated potatoes, from a total of 100 visits, none was recorded the first day, while 54% of the visits were recorded during the second day, and 18% and 28% were recorded during the third and fourth day, respectively.
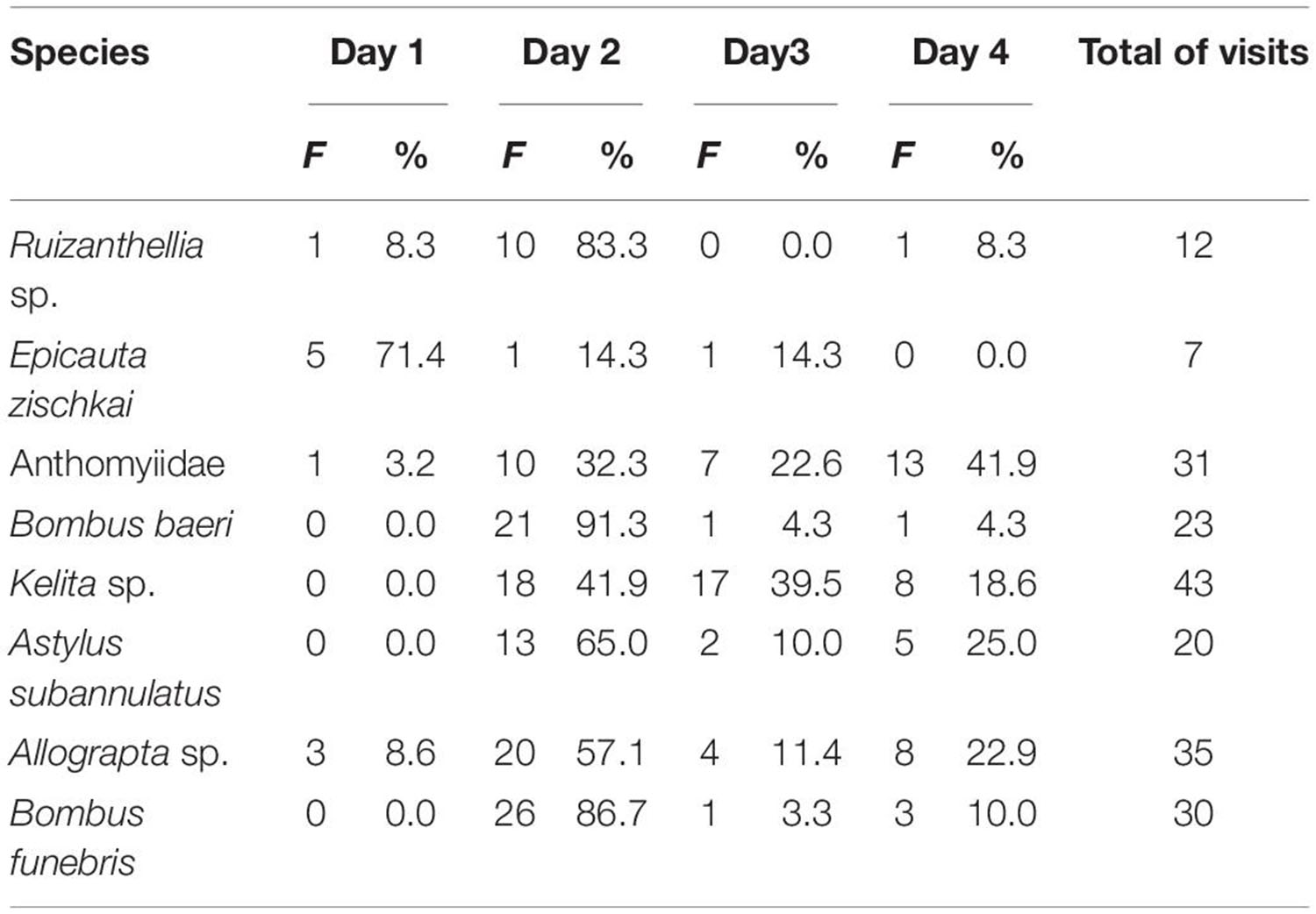
Table 6. Foraging activity of flowers visitor throughout four consecutive days.
Table 7 shows that the average duration of visits of all flower visitors was 27.84 ± 39.89 s, Astylus subannulatus being the species with the highest duration, although only 35% of their visits are effective. Most probably this beetle consumes flower parts. The most effective visitors, the Apidae bees conducted short visits, from 3 to 7 s on average, although Kelita sp. may have longer visits.
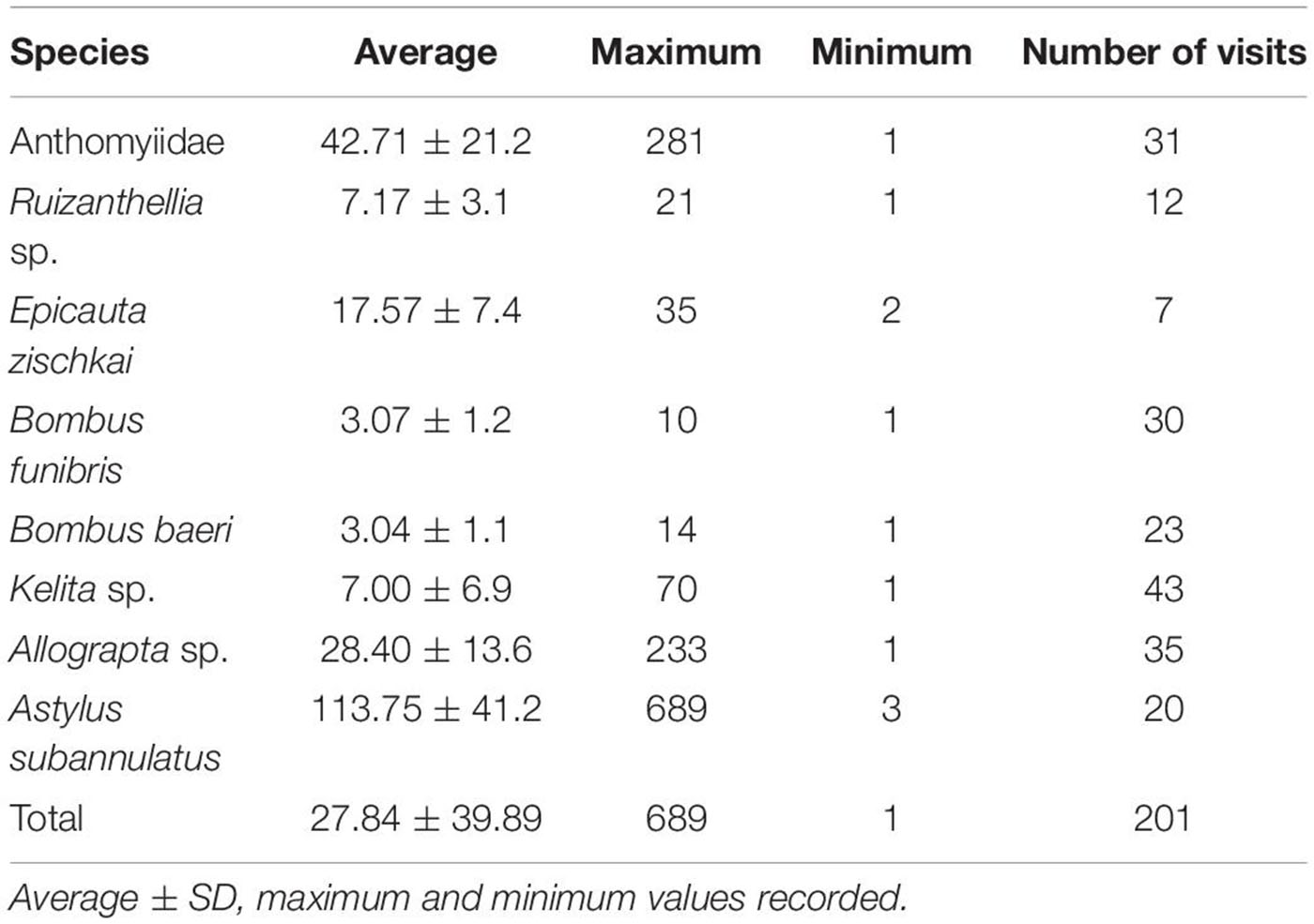
Table 7. Duration of visits (in seconds) of the species of insects visiting flowers of the potato species and varieties studied.
Activity of flower visitors was on average 31% between 9:00 and 12:00 h; it increased to nearly 60% (nearly 67% in wild populations) between 12:00 and 15:00 and decreased to nearly 11% from 15:00 to 17:00 (none in wild populations). All species of flower visitor had their highest activity between 12:00 and 16:00 h, but it is noteworthy that the most effective visitors, the Apidae bees visited flowers even from 15:00 to 17:00 h (Table 8).
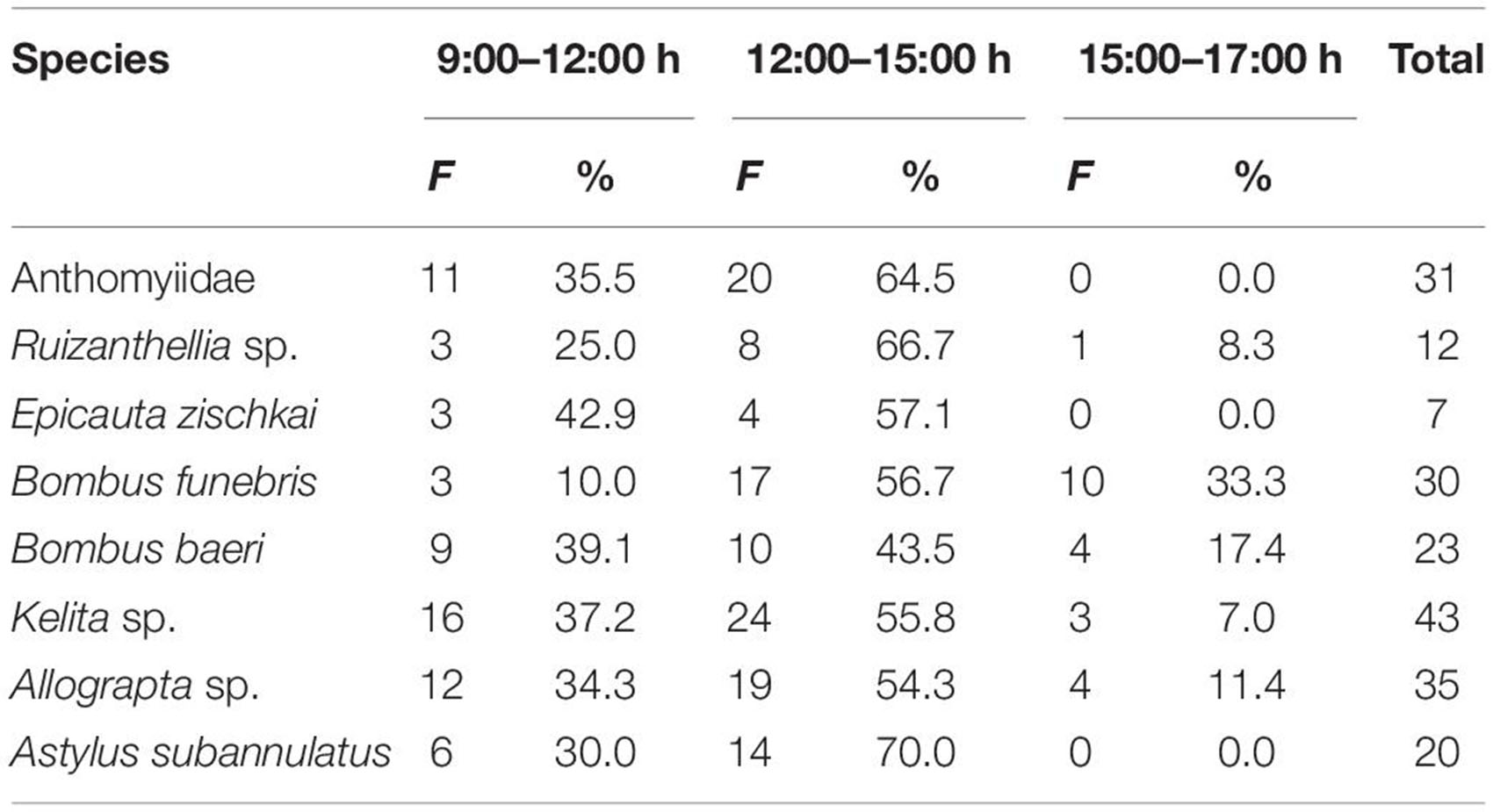
Table 8. Periods of higher activity of insects visiting flowers of the potato species and varieties studied.
Pollination is one of the main mechanisms of gene flow. One important condition for interchange of genes through pollination is synchronization of blooming and sharing of flower visitors, two aspects that, as described above, do occur. Another important aspect is the effectiveness of crosses among species and varieties. The reciprocal crosses among 30 varieties of cultivated potatoes and S. candolleanum were successful in nearly 35% of the cases, mainly in diploid and tetraploid varieties (Table 9). Also important, according to people interviewed, is the eventual dispersal of seeds by frugivores which currently is carried out by horses and donkeys and, of course, the active movement of tubers by humans. The latter is particularly important in the cases of cultivated varieties but also in the cases of araq papas and k’ipa papas.

Table 9. Cultivated crop varieties (CV) of potatoes (Solanum tuberosum subsp. tuberosum) and their ploidies reported by Márquez-Castellanos et al. (2019), used in the experimental crosses with the diploid Solanum candolleanum (S. c.) (Spooner et al., 2014).
Discussion
Use and Management of Potatoes and Their Gene Flow
Since the classic reflexions by Darwin (1859), variation of organisms under domestication has been associated to human selection, but it is now clear that selection is not the only evolutionary force influencing this process. In fact, all evolutionary forces currently recognized by evolutionary biologists, either natural or human-influenced are acknowledged to be involved (Casas et al., 2016). Outstandingly, gene flow, which has been managed and driven for long time by humans has been discussed as a powerful force operating in domestication (Parra et al., 2012). This is especially the case in areas where crops coexist with their wild relatives, like potatoes in the Andean region where wild, weedy, and domesticated varieties concur, allowing the flow of genes among themselves, forming complex nets of continuous reproductive interactions. These interactions are increased where movement of propagules is conducted by humans in addition to natural means, and even more where wild and weedy relatives are used, valued, and managed by people, as in the case studied.
Wild atoq or sacha papas are currently recognized and used by the Andean people in general and in the study region in particular; these potatoes are mainly used as medicine, but in this and other studies people say that these potatoes (particularly S. candolleanum) were consumed during hunger times in the past (Rumold and Aldenderfer, 2016), but probably also at present. It is known that wild potatoes contain high levels of glycoalkaloids toxic for humans (Ralla et al., 2012), however, several studies by Timothy Johns in the 1980s documented detoxification mechanisms (since toxicity is not neutralized by heat or by removing the peel) practiced by several North American and South American peoples by using different types of phyllosilicate clays (Johns, 1986; Johns and Keen, 1986; Johns and Alonso, 1990). In the Andean region, these clays are prepared and consumed as a kind of sauce called “chaco” (Castillo-Contreras and Frisancho-Velarde, 2015), a suspension of clays and salt aimed at attenuating the bitter flavor of toxic tubers that also has detoxification effects. Such mechanisms of food processing have been described from the 16th Century by chroniclers of the Spanish Conquest and recently confirmed by several studies (Ralla et al., 2012). In addition, it has been documented that other forms of processing potatoes for consumption, like chuño and moray, involving grounding, freezing, and drying processes have detoxification effects (Melton et al., 2020). Also, some authors have reported that gene flow from cultivated/weedy to wild potatoes may confer attributes evolved through domestication (e.g., some aspects of weedy behavior for survival in disturbed areas), among them decrease of toxicity (see Kaiser et al., 2020), and this could explain that consumption of wild potatoes is feasible. In theory, wild potatoes occurring in the neighborhood of agricultural areas would be less toxic than those occurring far away, but this hypothesis must be tested. Documenting these aspects is relevant to explain the current consumption and cultural value of wild potatoes, as well as to analyze early stages of use and conditions that facilitated and enhanced domestication of potatoes (Johns, 1986, 1996; Rumold and Aldenderfer, 2016; Melton et al., 2020). Currently, atoq papas are potatoes respected by people in the study area and, although their abundance is not deliberately enhanced in chacras, atoq papas are not removed from crop fields except when people consider these plants compete with the cultivated potatoes.
Araq papas are deliberately tolerated in chacras, and their abundance is also enhanced by tilling the land, covering their tubers to protect them against herbivores, adding animal dung to favor their growth, and promoting their diversity by collecting and planting tubers from other chacras. In addition, these weedy potatoes are cultivated mainly in association with maize, although previously studies by De Haan et al. (2007) reported that this is a rare practice. We found that cultivation of araq papas is not rare, being commonly practiced in association with fava beans, tarwi, quinoa and other crops; moreover, we recorded some cases of araq papas being intensively cultivated as monocrops because of their high value in regional markets, which is even higher than that of cultivated potatoes. In Quiñota and Chumbivilcas, a local traditional fair is carried out where farmers show and sell their varieties of araq papas (Begazo et al., 2019). The fair shows the high cultural and economic value of araq papas in the region; people go the fairs to obtain tubers of varieties from different areas, which contribute to increase variation of araq papas managed by households and potentiates the gene flow among distant populations.
These forms of management are similar to those described for edible weeds of several species in Mesoamerica, Andes and Amazonia, which have been documented to involve processes of incipient domestication (Casas et al., 1996, 2007; Parra et al., 2010, 2012; Clement et al., 2021). Management of wild and weedy potatoes offer therefore the opportunity to analyze parallel similar processes of domestication in three of the most important settings of domestication in the Americas. This information has important consequences, one of them is that tolerance, enhancement, and cultivation may involve practices of human selection. People choose and favor particular varieties they like because of their differential attributes. But in addition, they choose particular phenotypes of each variety and favor their tolerance, care and propagation. We found this management patterns occurring with araq papas, people enhancing and removing desirable and undesirable individuals, respectively and selectively moving from place to place tubers of the varieties and individuals they prefer. This is human selection, operating through different mechanisms from those practiced on cultivated potatoes, but anyway human selection, related to a process of ongoing domestication occurring on weedy plants that maintain weedy behavior and ecological aspects. It is still necessary to evaluate the degree of phenotypic and genotypic differentiation of araq papas with respect to atoq papas and crop potatoes, but this fact illustrates that domestication can operate following several routes, not only crop cultivation.
Another important aspect documented in this study is that potatoes representing a gradient of management intensity are maintained and propagated in ways that favor gene flow among araq papa varieties, but also among araq papa, atoq papa and crop potatoes. People interviewed reported that fruits of the different types of potatoes produce seeds, new plants, and all of them form part of the chacras where araq papas are managed. We documented numerous varieties of araq papa and identified those with higher cultural value which are also the most frequent and widely distributed in chacras. We now can hypothesize that gene flow among the most salient and widely distributed araq papas (e.g., runcu, yuraq ch’akiña, p’akush, wuaña muyu, puca ch’akiña, q’abka and chakiña kulli) with cultivated varieties would be the higher in the region studied. Therefore, characterizing genotypes of araq papa varieties, analyzing their phylogenetic relationships, and studying the population genetics of those varieties that are culturally and ecologically more important, together with the atoq papas and crop potatoes neighboring their distribution would be the way of evaluating the amount of gene flow between wild, weedy and crop potatoes of the region.
Some studies report the presence of araq papas in several regions of Perú, but these are still scarce. Among the most relevant are those conducted in Cajamarca and Huánuco by Centro Internacional de la Papa [CIP], and The Federación Departamental de Comunidades Campesinas [FEDECH] (2006) and Meza (2014), in Cusco, by Domínguez (2017) and in Huancavelica by De Haan (2009) and Meza (2014). Our report for Cusco and Apurimac adds information to these efforts. In this study 53 varieties of araq papa were recognized by farmers in 8 villages, while in Huancavelica, De Haan (2009) recorded that few farmers mentioned 2 to 4 different varieties of araq papa, in only one of eight communities studied, whereas Domínguez (2017) documented that in Chumbivilcas, Cusco, there were 19 different clones of araq papa in one village (Pallpa pallpa, which was sampled in our study, where we recorded 15 varieties). Domínguez (2017) recorded that each farmer had on average 9.45 different clones (we recorded 9.33 in that village, and 8 to 12 in the whole region), showing that deeper research is needed at household unit, village and region levels. More studies are also needed at national scale for a diagnosis of the variation of these plants in Peru, identifying their distribution and the areas with the highest diversity. Such information is important to design policies of in situ conservation of these plants and for reaching a complete panorama of interactions and gene flow among populations of the complex atoq papa, araq papa, kipa papa and cultivated potatoes. This should be an urgent research priority since the traditional crop fields where araq papa are grown are under constant threats to disappear or being replaced by modern crops (SPDA et al., 2015).
K’ipa papas appear to have an exceptionally important role in the gene flow system and diversification of potato crops. These plants grow in fallow agricultural fields and sometimes emerge in chacras with other crop species as documented by Brush et al. (1981) and Tapia and Fries (2007), or as escaped plants as considered by De Haan (2009). These plants are generally considered to derive from cultivated potatoes and therefore their tubers are harvested and included in the stores where other potatoes are maintained. People of the Andean region tend to cultivate potatoes and other tubers through mixtures of varieties (Velásquez-Milla et al., 2011). Therefore, harvesting and storage places commonly include also mixtures of varieties, some of which are used for the following generation of potatoes. This fact favors the mechanism through which k’ipa papas derived from sexual means are included in the subsequent generations of potatoes propagated vegetatively, although farmers, as observed by Brush et al. (1981), are not generally aware about these mechanisms.
Based on this information, we hypothesize that fallow agricultural fields where potatoes were cultivated in the previous agricultural cycle should include k’ipa papas derived from the remaining non-harvested tubers but, in addition, individuals derived from seeds, which may in turn result from crosses among wild, weedy, and cultivated species and variants. A similar system was documented for another Andean tuber, Oxalis tuberosa or “oca” in Bolivia (Bonnave et al., 2014, 2015), where unconscious introduction of k’ipa oca, plants originated from seedlings and others derived from tubers (without distinction by the farmers). Moscoe and Emshwiller (2016) recognized this as a possible mechanism of diversification for “oca” in the Southern Peruvian Andes, and in other vegetatively propagated tropical crops (McKey et al., 2010). It is also the case of Tropaeolum tuberosa according to reports by farmers describing the rise of new varieties in field crops (Velásquez-Milla et al., 2011), and reports by Pissard et al. (2008) who discuss that intraspecific diversity in mashua fields could result from the incorporation of varieties generated by sexual reproduction identified as new, selected, and conserved by peasants. Also similar is the occurrence of cassava plants originated from hybrid seeds produced by crosses between cassava and wild relatives, as well as between different cassava varieties, as reported in “roças” of the Brazilian Amazon by Martins and Oliveira (2009).
For k’ipa papas, very few studies from Perú have been documented. Zimmerer and Douches (1991) described how farmers incorporated seeds of native potatoes to the gene pool of cultivated types from k’ipa papas. In Paucartambo Valley, in Cusco, there is a practice of rotation known as kutirpa, where after two years of potato crops in fields, farmers cultivate oca and mashua, and in harvest time, volunteer tubers of potato developed from botanical seeds, are collected with the other two crops. Also, there are few cases of volunteer tubers of potato harvested from barley fields, after crop rotation. This information suggests that use and management of k’ipas of several crop species are rather deliberate.
However, these processes must be more deeply studied. A main challenge for research is to evaluate the proportion of k’ipa papas derived from seeds and that derived from tubers. To evaluate it, we sampled at random plants of k’ipa papa in chacras, to be analyzed through molecular markers. These plants would also help to explain how crops that are traditionally propagated by vegetative means are highly diverse and able to show high levels of intraspecific variability and have been able to adapt to so many different and changing growing conditions like those from mountain regions as the Andes (Bonnave et al., 2014).
For potatoes, studies in Peru and Bolivia have documented how farmers practice selection of tubers from volunteers originated by sexual seeds from berries or tamborotoco dropping to the ground (Jackson et al., 1980; Johns and Keen, 1986; Quiros et al., 1992) and some anecdotal information has been reported for Cusco, near to Calca and Paucartambo (Quiros et al., 1992). In a particular case, in Chincheros, Cusco, Franquemont et al. (1990), mentioned that an important source of new varieties of potato is that resulting from artificial selection and cultivation of seed from the best fruits of potato plants. In any case, sexual hybrids could enter the complex array of landrace cultivars (Spooner et al., 2010) and represent an important amount of phenotypic variability available as new tubers for selection by farmer, as documented by Quiros et al. (1992) using isozyme analysis after outcrossing experiments with farmer’s preferred potatoes cultivar for sexual propagation. Zimmerer and Douches (1991) discussed from their own findings using isozymes, that a moderate to large number of genotypes within single potato landraces, could probably result from sexual recombination and cultivation of seedlings in potato fields, therefore, these authors questioned the assumption that vegetatively crops are originated only by clonal propagation.
Therefore, k’ipa papas are probably important reservoirs of new variation and direct source for maintaining gene flow from wild and weedy plants to crops. How much? It is again a relevant question, which should be tested by sampling fallow agricultural fields and identifying those plants derived from previously cultivated potatoes and those that are hybrids with other varieties.
Reproductive Biology
One of the years we conducted studies on reproductive biology (2016- 2017) was an ENSO year; however, our phenological data were similar to those reported by Román and Hurtado (2002); Ladrón de Guevara (2005), Sepúlveda-Cano (2013), and Meza et al. (2014). Our records of anthesis duration were also similar to those reported by Barrientos-Restrepo (2012); Sepúlveda-Cano (2013) and Meza et al. (2014). All this information shows the relative stability of temporal events of sexual reproduction, which could be an adaptation to long term periods of recurrent climate alteration events.
Flowers of the genus Solanum have poricidal anthers visited by numerous insects, including bees, beetles, and flies (De Luca and Vallejo-Marín, 2013). For Solanum tuberosum, several studies, including Bravo et al. (2002) and Sistema de Informacion de Organismos Vivos Modificados [SIOVM] (2017) reported bees of the genus Apis and Bombus as main flower visitors. We found a pattern similar to that characterized in Junín, Cusco and Puno, in Peru by Scurrah et al. (2008), who reported that the potato variety Yungay, and its wild relatives shared the following flower visitors: Astylus sp., Anthophora sp., Apis mellifera, Bombus baeri, Bombus funebris, Bombus opifex, Lonchopria sp., Megachile sp., most of which, and others, were recorded in this study. Although finer studies are required to test differences among flowers with different colors and fertile and sterile varieties, the information referred to above and the pattern we found indicating that wild, weedy, and cultivated potatoes share flower visitors suggest that domestication has not changed flower attributes affecting visitors as occurring in wild plants.
Our videorecording of flower visits allowed observing that most visitors visited flowers on the days 2 and 3 of the anthesis period. Most probably the first day the pollen production was incipient while in the fourth day the availability of pollen decreased, as suggested by De Luca and Vallejo-Marín (2013), or because of the number of flowers, as suggested by Pantoja et al. (2014).
Although we did not test self-compatibility, several previous studies have reported that most diploid species are self-incompatible (Spooner et al., 2014). It calls attention that most successful crosses in our experiments were those in which the cultivated varieties were female (pollen receptors), but not males (pollen donors), which appears to be consistent with the observation by Abdalla and Hermsen (1972) that self-compatible species can be crossed as a female, but not as a male, to self-incompatible species and most cultivars are male sterile (Spooner et al., 2014).
We found that crosses between atoq papa with cultivated potatoes are possible and successful. In theory, araq papas and crop potatoes are closer relatives, therefore, although we could not test the effectiveness of crosses among them, it is possible to hypothesize that these crosses would be more successful than between atoq papa and crop potatoes. The following step of our study will be testing this hypothesis and to evaluate through molecular markers how much gene flow occurs among populations.
The long history of evolution of wild populations and the great phenotypic variation of weedy potatoes allows suggesting that wild and weedy populations of potatoes are important reservoirs of genetic variation, and gene flow is a crucial process that allows these reservoirs to interchange and increase variation. Bees involved in pollen interchange and frugivores consuming fruits of wild, weedy, and cultivated potatoes, as well as the role of humans transferring vegetative propagules from place to place are all factors intervening in the movement of potatoes genes in the space. Maintaining populations of all plants and animals involved in these processes, including the human cultural motives, is crucial to maintain gene flow for ensuring potato diversification. Among the threats to bee populations are the use of insecticides, and the introduction of exotic parasites and the loss of floral resources (Goulson et al., 2015). But also, herbicides affecting populations of wild and weedy potatoes would interrupt crucial processes that explain the diversification of one of the most important crops sustaining human food security.
Conclusion
Atoq papas, araq papas and k’ipa papas are plant resources valued by the Andean people, either for medicinal purposes, or because of their complementary role in food security and sovereignty. At the same time, the information available suggests that these are important reservoirs of genetic variation which is in active gene flow through reproductive interaction and spatial movement of propagules, allowed and enhanced by human activities. Such variation and processes are crucial for the generation of the high diversity of potatoes characterizing the Andean region. Conserving this biocultural diversity is highly important for the Andean people and humanity, and conserving such diversity requires strategies for maintaining species and varieties of plants and animals involved, as well as the biological and cultural processes that have acted in this direction for thousands of years. Documenting all these aspects is crucial for understanding the mechanisms operating to explain the diversification of potatoes and the biocultural conditions for conservation. But, in addition, it would be equally important to understand what is happening with other Andean tubers.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author/s.
Ethics Statement
The studies involving human participants were reviewed and approved by local authorities, in particular Mr. Doroteo Condorimay Jauja, the mayor authority of the Community of Cocha, District of Haquira, Province of Cotabambas, region Apurimac. The participants provided their consent to participate in this study. Written informed consent was obtained from the relevant individual for the publication of any potentially identifiable images or data included in this article.
Author Contributions
FP-R did the conception and design of the research, fieldwork gathering data, data analysis, and writing the manuscript. ACa did the conception and design of the research, fieldwork gathering data, data analysis, writing the manuscript, and reviewing several versions of manuscript. DB, AP, and JS did the fieldwork gathering data. EM did the fieldwork gathering data, data analysis and interpretation, and writing the manuscript. ACr, IT-G, and MZ did the fieldwork gathering data, data analysis, and reviewing of manuscript. LL did the data analysis and reviewing of manuscript. JT-G did the conception and design of the research, fieldwork data gathering, and reviewing the manuscript. All authors contributed to the article and approved the submitted version.
Funding
The authors thank the Agrarian Innovation Programme (Programa de Innovación Agraria, PNIA) of Peru for financial support to the Project “Diversity, gene flow of Andean native crops and food security: actions against genetic erosion for facing climate change” (Research Project 027-2015-INIA-PNIA/UPMSI/IE), as well as projects A1-S-14306 and IN206520 supported by CONACYT and PAPIIT, UNAM, México, respectively, which allowed that the different studies were carried out in Mexico and Peru.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We specially thank the generosity of people from Chumbivilcas and Haquira in Cusco and Apurimac, Peru. We thank the crucial support of the Centro Andino de Educación y Promoción José María Arguedas (CADEP), especially Carmen Álvarez, who decidedly supported all steps of the research and helped with infrastructure, personnel, contacts with people in communities and eased equipment. We thank Alfredo F. Fuentes for the Bombus opifex photograph included in Figure 6 (b2). We also thank Justina Núñez and Domingo Begazo for their help in correcting Quechua writing and identifying synonymy in the terms recorded.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.617969/full#supplementary-material
References
Abdalla, M. M. F., and Hermsen, J. G. T. (1972). Plasmons and male sterility types in Solanum verrucosum and its interspecific hybrid derivatives. Euphytica 21, 209–220. doi: 10.1007/BF00036761
Bamberg, J. (2020). Emasculation technique reduces seedset in Solanum verrucosum. Am. J. Potato Res. 97, 111–113. doi: 10.1007/s12230-019-09754-3
Barrientos-Restrepo, E. (2012). Abejas Visitantes de Papa (Solanum tuberosum L.), en Tres Agroecosistemas de los Departamentos de Cundinamarca y Boyaca. Master’s thesis, Medellín: Universidad Nacional de Colombia.
Begazo, D., Torres, I., Márquez-Castellanos, E., Segovia, J., Zarazúa, M., Parra, F., et al. (2019). Sacha, aracca, k’ipas y cultura andina: las bases de la diversidad de papas. LEISA Revista de Agroecol. 35, 12–15.
Bethke, P. C., Halterman, D. A., and Jansky, S. H. (2017). Are we getting better at using wild potato species in light of new tools? Crop Sci. 57, 1241–1258. doi: 10.2135/cropsci2016.10.0889
Bonnave, M., Bleeckx, G., Rojas Beltrán, J., Maughan, P., Flamand, M.-C., Terrazas, F., et al. (2014). Farmers’ unconscious incorporation of sexually-produced genotypes into the germplasm of a vegetatively propagated crop (Oxalis tuberosa Mol.). Genetic Resources Crop Evol. 61, 721–740. doi: 10.1007/s10722-013-0068-z
Bonnave, M., Bleeckx, T., Terrazas, F., and Bertin, P. (2015). Effect of the management of seed flows and mode of propagation on the genetic diversity in an Andean farming system: the case of oca (Oxalis tuberosa mol.). Agricul. Hum. Values 33, 673–688. doi: 10.1007/s10460-015-9646-3
Brack, A., Aranda, C., Bernales, M., Bustamante, R., Campos, L., Capurro, F., et al. (2008). Diagnóstico Ambiental del Perú. Lima: Ministerio del Ambiente.
Brack, A., and Bravo, F. (2006). Peru-Millenary Legacy Lima. Chiclayo: Universidad San Martín de Porres.
Bradshaw, J. E., Bryan, G. J., and Ramsay, G. (2006). Genetic resources (including wild and cultivated Solanum species) and progress in their utilisation in potato breeding. Potato Res. 49, 49–65. doi: 10.1007/s11540-006-9002-5
Bravo, W., Franco, J., Main, G., Carrasco, E., and Gabriel, J. (2002). Evaluación de la dispersión de polen como medida de bioseguridad para la liberación de plantas transgénicas de papa a campo. Revista Latinoamericana De La Papa 13, 95–103.
Brush, S. B. (2004). Farmers’ Bounty: Locating Crop Diversity in the Contemporary World. London: Yale University Press.
Brush, S., Carney, H. J., and Huaman, Z. (1981). Dynamics of andean potato agriculture. Econ. Botany 35, 70–88. doi: 10.1007/BF02859217
Brush, S., Kesseli, R., Ortega, R., Cisneros, P., Zimmerer, K., and Quiros, C. (1995). Potato diversity in the Andean center of crop domestication. Conserv. Biol. 9, 1189–1198. doi: 10.1046/j.1523-1739.1995.9051176.x-i1
Brush, S. B. (1992). Ethnoecology, biodiversity, and modernization in Andean potato agriculture. J. Ethnobiol. 12, 161–185.
Brush, S. B., Taylor, E. J., and Bellon, M. (1992). Technology adoption and biological diversity in Andean potato agriculture. J. Dev. Econ. 39, 365–387. doi: 10.1016/0304-3878(92)90044-A
Bumrungsri, S., Harbit, A., Benzie, C., Carmouche, K., Sridith, K., and Racey, P. (2008). The pollination ecology of two species of parkia (Mimosaceae) in Southern Thailand. J. Trop. Ecol. 24, 467–475. doi: 10.1017/S026646740800521X
Casas, A., Otero-Arnaiz, A., Pérez-Negrón, E., and Valiente-Banuet, A. (2007). In situ management and domestication of plants in Mesoamerica. Annals Botany 100, 1101–1115. doi: 10.1093/aob/mcm126
Casas, A., Torres-Guevara, J., and Parra, F. (2016). Domesticación en el Continente Americano. Manejo de biodiversidad y evolución dirigida por las culturas del Nuevo Mundo. Lima: Universidad Nacional Autónoma de México/Universidad Nacional Agraria La Molina.
Casas, A., and Vallejo, M. (2019). “Agroecología y agrobiodiversidad,” in Crisis Ambiental en México, ed. L. Merino (Ciudad de México: UNAM), 99–118.
Casas, A., Vázquez, M. C., Viveros, J. L., and Caballero, J. (1996). Plant management among the Nahua and the Mixtec of the Balsas river basin: an ethnobotanical approach to the study of plant domestication. Hum. Ecol. 24, 455–478. doi: 10.1007/BF02168862
Castillo-Contreras, O., and Frisancho-Velarde, O. (2015). The “Chaco”: eatable medicinal clay in the Peruvian highlands and its properties in digestive diseases. Revista de Gastroenterología del Peru 35, 97–99.
Centro Internacional de la Papa [CIP], and The Federación Departamental de Comunidades Campesinas [FEDECH] (2006). Catálogo de Variedades de Papa Nativa de Huancavelica-Perú. Lima: Centro Internacional de la Papa.
Clement, C. R., Casas, A., Parra-Rondinel, F., Levis, C., Peroni, N., Hanazaki, N., et al. (2021). Disentangling domestication from food production systems in the neotropics. Quaternay 4:4. doi: 10.3390/quat4010004
De Haan, S. (2009). Potato Diversity and Height: Multiple Dimensions of Farmer-Driven-in situ Conservation in the Andes. PhD thesis, Wageningen University, Wageningen.
De Haan, S., Bonierbale, M., Ghislain, M., and Nuñez, J. (2007). Indigenous Biosystematics of Andean potatoes: folk taxonomy, descriptors and nomenclature. Acta Horticulturae 745, 89–134. doi: 10.17660/ActaHortic.2007.745.4
De Haan, S., and Rodríguez, F. (2016). “Potato origin and production,” in Advances in Potato Chemistry and Technology, eds J. Singh and L. Kaur (London: Elsevier Inc.), 1–32.
De Luca, P., and Vallejo-Marín, M. (2013). What’s the ‘buzz’ about? the ecology and evolutionary significance of buzz- pollination. Curr. Opin. Plant Biol. 16, 429–435. doi: 10.1016/j.pbi.2013.05.002
Domínguez, F. (2017). Variabilidad fenotípica y evaluación agrobotánica de araqa papas (Solanum tuberosum subespecie andigena) en la comunidad de Hatta Pallpa Pallpa del distrito de Quiñota-Chumbivilcas-Cusco. Bachellor thesis, Universidad Nacional San Antonio Abad de Cusco, Peru.
Eijlander, R., and Stiekema, W. J. (1994). Biological containment of potato (Solanum tuberosum): outcrossing to the related wild species black nightshade (Solanum nigrum) and bittersweet (Solanum dulcamara). Sexual Plant Reproduction 7, 29–40. doi: 10.1007/BF00241885
Fernández, F., and Sharkey, M. (2006). Introducción a los Hymenoptera de la región Neotropical. Bogotá: Sociedad Colombiana de Entomología and Universidad Nacional de Colombia.
Franquemont, C., Plowman, T., Franquemont, E., King, S. R., Niezgoda, C., Davis, W., et al. (1990). The ethnobotany of Chinchero, an Andean community in southern Peru. Fieldiana, Botany. News Series. Chicago, IL: Field Museum of Natural History.
Goulson, D., Nicholls, E., Botías, C., and Rotheray, E. L. (2015). Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 347, 1–9. doi: 10.1126/science.1255957
Hawkes, J. G. (1990). The Potato: Evolution, Biodiversity and Genetic Resources. London: Belhaven Press.
Huamán, Z. (1995). Técnicas Citológicas Para Determinar el Número Cromosómico y la Fertilidad de las Papas. Guía de Investigación. Lima: CIP (Centro Internacional de la Papa), 1–18.
Huamán, Z., and Spooner, D. M. (2002). Reclassification of landrace populations of cultivated potatoes (Solanum sect. Petota). Am. J. Bot. 89, 947–965. doi: 10.3732/ajb.89.6.947
Jackson, M. T., Hawkes, J. G., and Rowe, P. R. (1980). An ethnobotanical fi eld study of primitive potato varieties in Peru. Euphytica 29, 107–113. doi: 10.1007/BF00043180
Jackson, S. A., and Hanneman, R. E. (1999). Crossability between cultivated and wild tuber-and non-tuber-bearing Solanum. Euphytica 109, 51–67. doi: 10.1023/A:1003710817938
Jansky, S. (2006). Overcoming hybridization barriers in potato. Plant Breed. 125, 1–12. doi: 10.1111/j.1439-0523.2006.01178.x
Johns, T. (1986). Detoxification function of geophagy and domestication of the potato. J. Chem. Ecol. 12, 635–646.
Johns, T. (1996). The Origins of Human Diet and Medicine: Chemical Ecology. Tucson, AZ: University of Arizona Press.
Johns, T., and Alonso, J. G. (1990). Glycoalkaloid change during the domestication of the potato, Solanum section petota. Euphytica 50, 203–210. doi: 10.1007/BF00023646
Johns, T., and Keen, S. L. (1986). Ongoing evolution of the potato on the altiplano of Western Bolivia. Econ. Botany 40, 409–424. doi: 10.1007/BF02859652
Kaiser, N., Douches, D., Dhingra, A., Glenn, K. C., Herzig, P. R., Stowe, E. C., et al. (2020). The role of conventional plant breeding in ensuring safe levels of naturally occurring toxins in food crops. Trends Food Sci. Technol. 100, 51–66. doi: 10.1016/j.tifs.2020.03.042
Ladrón de Guevara, O. (2005). Introducción a la climatología y la fenología agrícola. Cusco: Universitaria - UNSAAC.
Lastra, S., Parra, F., Casas, A., Torres-Guevara, J., Cruz, A., León, W., et al. (2019). “Uso de índices de diversidad para evaluar el cultivo de papa en una zona de agrobiodiversidad de los andes peruanos,” in De los Cultivos Nativos y el Cambio del Clima. Hallazgos (Huánuco y Apurímac), eds J. Torres-Guevara, F. A. Parra-Rondinel, A. Casas, and A. Cruz-Soriano (Lima: Universidad Nacional San Antonio Abad de Cusco, Universidad Nacional Agraria La Molina, Universidad Nacional Autónoma de México, Programa Nacional de Innovación Agraria, IDMA, CADEP), 119–144.
Márquez-Castellanos, E., Lizárraga, L., Parra, F., Casas, A., Torres-Guevara, J., Begazo, D., et al. (2019). “Conectividad entre poblaciones silvestres y cultivadas de papa,” in De los Cultivos Nativos y el Cambio del Clima. Hallazgos (Huánuco y Apurímac), eds J. Torres-Guevara, F. A. Parra-Rondinel, A. Casas, and A. Cruz-Soriano (Lima: Universidad Nacional San Antonio Abad de Cusco, Universidad Nacional Agraria La Molina, Universidad Nacional Autónoma de México, Programa Nacional de Innovación Agraria, IDMA, CADEP), 147–172.
Martins, P. S., and Oliveira, G. C. X. (2009). “Dinâmica evolutiva em roças de caboclos amazônicos,” in Diversidade Biológia E Cultural da Amazônia, eds I. C. G. Vieira, J. M. C. da Silva, D. C. Oren, and M. A. D’Incao (Belém: Museu Paraense Emílio Goeldi), 373–392.
Maune, J. F., Camadro, E. L., and Erazzu, L. E. (2018). Cross-incompatibility and self-incompatibility: unrelated phenomena in wild and cultivated potatoes? Botany 96, 33–45. doi: 10.1139/cjb-2017-0070
McKey, D., Elias, M., Pujol, B., and Duputié, A. (2010). The evolutionary ecology of clonally propagated domesticated plants. New Phytol. 186, 318–332. doi: 10.1111/j.1469-137.2010.03210.x
Melton, M. A., Biwer, M. E., and Panjarjian, R. (2020). Differentiating Chuño blanco and Chuño negro in archaeological samples based on starch metrics and morphological attributes. J. Archaeol. Sci. Rep. 34:102650. doi: 10.1016/j.jasrep.2020.102650
Meza, K. (2014). Capacidad de Establecimiento y Supervivencia de Poblaciones de Papa en Competencia con la Flora Pionera en Tres Localidades. Tesis Para Optar Por el Título de Ingeniera Agrónoma. Facultad de Agronomía. Peru: Universidad Nacional del Centro del Perú.
Meza, N., Daboin, B., and Coraspe, H. (2014). Aspectos de la biología floral en papa (Solanum tuberosum L.) bajo condiciones de invernadero. Rev. Fac. Agron. (LUZ). 2014, 75–84.
Moscoe, L. J., and Emshwiller, E. (2016). Farmer perspectives on OCA (Oxalis tuberosa; Oxalidaceae) diversity conservation: values and threats,”. J. Ethnobiol. 36, 235–256. doi: 10.2993/0278-0771-36.2.235
Newing, H., Eagle, C. M., Puri, R. K., and Watson, C. W. (2011). Conducting Research in Conservation. A Social Science Perspective. New York, NY: Taylor & Francis Group.
Pantoja, A., Pardo, A., García, A., Saenz, A., and Rojas, F. (2014). Principios y Avances Sobre Polinización Como Servicio Ambiental Para la Agricultura Sostenible en Países de Latinoamérica y el Caribe. Rome: FAO.
Parra, F., Blancas, J., and Casas, A. (2012). Landscape management and domestication of Stenocereus pruinosus (Cactaceae) in the Tehuacán Valley: human guided selection and gene flow. J. Ethnobiol. Ethnomed. 8:32. doi: 10.1186/1746-4269-8-32
Parra, F., Casas, A., Peñaloza-Ramírez, J. A., Cortés-Palomec, C., Rocha-Ramírez, V., and González-Rodríguez, A. (2010). Evolution under domestication: ongoing artificial selection and divergence of wild and managed Stenocereus pruinosus (Cactaceae) populations in the Tehuacán Valley, Mexico. Ann. Bot. 106, 483–496. doi: 10.1093/aob/mcq143
Pennec, F., Wencelius, J., Garine, E., Raimond, C., and Bohbot, H. (2012). FLAME v1.0: Free-list Analysis Under Microsoft Excel. Paris: CNRS.
Pissard, A., Arbizu, C., Ghislain, M., and Bertin, P. (2008). Influence of geographical provenance on the genetic structure and diversity of the vegetatively propagated Andean tuber crop, mashua (Tropaeolum tuberosum), highlighted by intersimple sequence repeat markers and multivariate analysis methods. Int. J. Plant Sci. 169, 1248–1260. doi: 10.1086/591979
Quiros, C. F., Ortega, R., van Raamsdonk, L., Herrera-Montoyo, M., Cisneros, P., Schmidt, E., et al. (1992). Increase of potato genetic resources in their centre of diversity: the role of natural outcrossing and selection by the Andean farmer. Genetic Resources Crops Evol. 39, 107–113. doi: 10.1007/BF00051229
Ralla, K., Sohling, U., Suck, K., Kasper, C., Ruf, F., and Scheper, T. (2012). Separation of patatins and protease inhibitors from potato fruit juice with clay minerals as cation exchangers. J. Separation Sci. 35, 1596–1602. doi: 10.1002/jssc.201200030
Rangel-Landa, S., Casas, A., Rivera-Lozoya, E., Torres-García, I., and Vallejo-Ramos, M. (2016). Ixcatec ethnoecology: plant management and biocultural heritage in Oaxaca. Mexico. J. Ethnobiol. Ethnomed. 12:30. doi: 10.1186/s13002-016-0101-3
Román, M., and Hurtado, G. (2002). Guía Técnica del Cultivo de Papa. El Salvador: CENTA Centro Nacional de Tecnología Agropecuaria y Forestal.
Rumold, C. U., and Aldenderfer, M. S. (2016). Late Archaic–Early formative period microbotanical evidence for potato at Jiskairumoko in the Titicaca Basin of southern Peru. Proc. Natl. Acad. Sci. U S A. 113, 13672–13677. doi: 10.1073/pnas.1604265113
Scurrah, M., Celis-Gamboa, C., Chumbiaca, S., Salas, A., and Visser, R. G. (2008). Hybridization between wild and cultivated potato species in the Peruvian Andes and biosafety implications for deployment of gm potatoes. Euphytica 164, 881–892. doi: 10.1007/s10681-007-9641-x
Sepúlveda-Cano, P. (2013). Diversidad de Abejas (Hymenoptera: Apoidea: Anthophila), en el cultivo de papa (Solanum tuberosum L.) y su efecto en la polinización. Master’s thesis, Medellín: Universidad Nacional de Colombia.
Silveira, F., Melo, G., and Almeida, E. (2002). Abelhas Brasileiras: Sistematica e Identificaçao. Belo Horizonte: Fernando A. Silveira.
Sistema de Informacion de Organismos Vivos Modificados [SIOVM] (2017). Recuperado el 12 de 01 de 2018, de. Philadelphia, PA: SIOVM
Smith, J. J., and Borgatti, S. P. (1998). Salience counts and so does accuracy: correcting and updating a measure for free-list-item salience. J. Linguistic Anthropol. 7, 208–202. doi: 10.1525/jlin.1997.7.2.208
SPDA, CCTA, and INIA (2015). Los Cultivos de la Sierra y el Cambio Climático Andino: Vulnerabilidad y Fortalezas. Lima: siete casos de la sierra centro y sur del Perú.
Spooner, D. M., Gavrilenko, T., Jansky, S. H., Ovchinnikova, A., Krylova, E., Knapp, S., et al. (2010). Ecogeography of ploidy variation in cultivated potato (Solanum sect. Petota). Am. J. Botany 92, 2049–2060. doi: 10.3732/ajb.1000277
Spooner, D. M., Ghislain, M., Simon, R., Jansky, S. H., and Gavrilenko, T. (2014). Systematics, diversity, genetics, and evolution of wild and cultivated potatoes. Bot. Rev. 80, 283–383. doi: 10.1007/s12229-014-9146-y
Spooner, D. M., López, A. S., Huamán, Z., and Hijmans, R. J. (1999). Wild potato collecting expedition in Southern Peru (Departments of Apurímac, Arequipa, Cusco, Moquegua Puno, Tacna in 1998: taxonomy and new genetic resources. Am. J. Pot. Res. 76, 103–119. doi: 10.1007/BF02853575
Spooner, D. M., and Salas, A. (2006). “Structure, biosystematics, and genetic resources,” in Handbook of Potato Production, Improvement, and Postharvest Management, eds J. Gopal and S. M. P. Khurana (New York, NY: Haworth’s Press, Inc.), 1–39.
Sutrop, U. (2001). List task and a cognitive salience index. Field Methods 13, 263–276. doi: 10.1177/1525822X0101300303
Tobin, D., Bates, R. M., Brennan, M. A. Jr., and Gill, T. (2016). Peru potato potential: biodiversity conservation and value chain development. Renewable Agriculture Food Systems 33, 19–32. doi: 10.1017/S1742170516000284
Torres-Guevara, J., Parra, F., and Casas, A. (2017). “Panorama de los recursos genéticos en Perú,” in Domesticación en el Continente Americano. Perspectivas de Investigación y Manejo Sustentable de Recursos Genéticos en el Nuevo Mundo, eds A. Casas, J. Torres-Guevara, and F. Parra (Morelia: Universidad Nacional Autónoma de México, Universidad Nacional Agraria La Molina, CONACYT), 103–134.
Ueda, K. (2020). iNaturalist Research-Grade Observations. Available online at: https://doi.org/10.15468/ab3s5x (accessed October 8, 2020).
Vavilov, N. I. (1992). Origin and Geography of Cultivated Plants. Cambridge: Cambridge University Press.
Velásquez-Milla, D., Casas, A., Torres-Guevara, J., and Cruz, A. (2011). Ecological and socio-cultural factors influencing in situ conservation of crop diversity by traditional Andean households in Peru. J. Ethnobiol. Ethnomed. 7:40. doi: 10.1186/1746-4269-7-40
Zimmerer, K. S. (2003). Geographies of seed networks for food plants (potato, ulluco) and approaches to agrobiodiversity conservation in the Andean countries. Soc. Nat. Resources 16, 583–601. doi: 10.1080/08941920309185
Zimmerer, K. S. (2016). Spell of the Urubamba: anthropogeographical essays on an Andean valley in space and time. AAG Rev. Books 4, 127–130. doi: 10.1080/2325548x.2016.1187490
Zimmerer, K. S., de Haan, D., Jones, A. D., Creed-Kanashiro, H., Tello, M., Plasencia-Amaya, F., et al. (2020). Indigenous smallholder struggles in Peru: nutrition security, agrobiodiversity, and food sovereignty amid transforming global systems and climate change. J. Latin Am. Geography 19, 74–111. doi: 10.1353/lag.2020.0072
Keywords: Andean tubers, biocultural diversity, crop evolution, domestication, gene flow, genetic resources, in situ conservation, potato diversity
Citation: Parra-Rondinel F, Casas A, Begazo D, Paco A, Márquez E, Cruz A, Segovia J, Torres-García I, Zarazúa M, Lizárraga L and Torres-Guevara J (2021) Natural and Cultural Processes Influencing Gene Flow Among Wild (atoq papa), Weedy (araq papa and k’ipa papa), and Crop Potatoes in the Andean Region of Southern Peru. Front. Ecol. Evol. 9:617969. doi: 10.3389/fevo.2021.617969
Received: 15 October 2020; Accepted: 30 April 2021;
Published: 24 May 2021.
Edited by:
Ima Célia G. Vieira, Museu Paraense Emílio Goeldi, BrazilReviewed by:
C. Robin Buell, Michigan State University, United StatesSebastián Cordero, Pontificia Universidad Católica de Valparaíso, Chile
Copyright © 2021 Parra-Rondinel, Casas, Begazo, Paco, Márquez, Cruz, Segovia, Torres-García, Zarazúa, Lizárraga and Torres-Guevara. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alejandro Casas, YWNhc2FzQGNpZWNvLnVuYW0ubXg=