 Hui Zhang1,2†
Hui Zhang1,2† Chen Wang
Chen Wang
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 28 January 2021
Sec. Conservation and Restoration Ecology
Volume 9 - 2021 | https://doi.org/10.3389/fevo.2021.573948
This article is part of the Research Topic Trait-Based Plant Community Assembly, Ecological Restoration, and the Biocontrol of Invasive Exotic Plant Species View all 12 articles
Understanding functional traits that capture species response to overgrazing can help optimize ecosystem functioning. However, most studies have ignored the effects of life-form on the response of above- and below-ground traits to overgrazing. Here, we determined whether the above- and below-ground traits respond similarly to overgrazing; and does the response of above- and below-ground traits to overgrazing vary between the species of annual vs. perennial life-forms. We compared common grassland species between fenced and overgrazed meadows at four elevations (from 3000 to 3600 m), spanning an area of ∼200 square kilometers on the Qinghai-Tibetan Plateau. We found overgrazing resulted in large removal of perennial species at all elevations, which forced perennial species to alter their above- and below-ground traits so that they could shift from resource conservation to resource acquisition to avoid overgrazing. In contrast, overgrazing promoted annual species at all elevation, thus annual species did not make any response to overgrazing. These findings bridge the gap in the literature for the influence of annual vs. perennial life-forms on species-specific response at both above-, and below-ground levels during overgrazing. More important, these results can facilitate selecting species for sustainable ecosystem managements and restoration from overgrazing.
Optimizing ecosystem functions (e.g., productivity and nitrogen retention) has become a challenging task for agro-ecosystem management, as demands for food supply are ever-increasing (Orwin et al., 2018). Grazed land is one of the biggest agro-ecosystems that covers approximately 25% of the terrestrial surfaces on Earth (Asner et al., 2004). Overgrazing has been witnessed to have profound effects on ecosystem dynamics and species composition in grasslands ecosystems, as it leads to significant decrease in diversity, which in turn leads to increased greenhouse gas emissions and nitrogen loss, reduced productivity, water quality and lower soil carbon (C) storage (Ward et al., 2016; Zhou et al., 2017; Liu et al., 2018). Understanding of functional traits that capture responses of species to overgrazing can help in understanding vegetation composition dynamics. This in turn requires better understanding of above- and below-ground traits that respond to overgrazing as compared to landscapes that are not overgrazed (Orwin et al., 2010; Mouillot et al., 2011). Thus, use of the key functional traits that can capture species-specific response to overgrazing can help in selecting species to restoration from overgrazing (Orwin et al., 2018).
Above-ground traits, such as higher specific leaf area (SLA) and short plant height (H) that are associated with a resource acquisition vs. resource conservation trade-off, have widely been found as key traits that capture species-specific response to overgrazing (Briske, 1999; Westoby, 1999). In contrast, in the fenced condition, lower SLA and taller stature are regarded as a strategy for high resource conservation to adapt to resource limitation (Westoby, 1999). On the other hand, below-ground traits might also play a key role in mediating grazing-induced alterations to vegetation composition (Klumpp et al., 2009; McInenly et al., 2010). However, due to the notable lack of relevant data (Bergmann et al., 2017), it remains unclear whether below-ground traits also exhibit similar resource acquisition strategy (e.g., high root density, specific root length, specific root area, and low root tissue density) for avoiding overgrazing (McInenly et al., 2010; Kramer-Walter et al., 2016; Roumet et al., 2016; Feng et al., 2018).
Life-form plays a key role in influencing above-ground as well as below-ground traits. Usually annual species tend to have above- and below-ground traits that favor high resource acquisition, whereas perennial species tend to possess traits that promote a resource conservation strategy (Roumet et al., 2006, 2016; Zhang et al., 2018). Moreover, a meta-analysis has found that at the global scale, overgrazing promotes annual species over perennial species (Díaz et al., 2007). Thus, the response of above- and below-ground traits to overgrazing may vary between the species of annual vs. perennial life-forms. However, it remains unclear whether annual and perennial species differ in their above- and below-ground traits in response to overgrazing.
We hypothesize that after overgrazing, perennial species might largely get removed, whereas annual species could largely get promoted. Thus, annual and perennial species may alter their above- and below-ground traits for responding to overgrazing. To test this hypothesis, we determined how parameters of above- and below-ground traits of species dwelling in fenced and overgrazed meadows differed across annual and perennial life-forms. We investigated an area of ∼200 square kilometers, across an elevation of 3000–3600 m in Qinghai-Tibetan plateau. Overgrazing has led to nearly half million square km of degraded grasslands that account for the one-third of the total alpine grassland cover on Qinghai-Tibetan Plateau (Cui and Graf, 2009). Finding out the key above- and below-ground traits that can classify vegetation composition between overgrazed and fenced meadows can help in effective sustainable ecosystem managements and restoration from overgrazing in Qinghai-Tibetan Plateau.
The maximum carrying capacity for cold season is approximately two yaks per hectare on Qinghai-Tibetan Plateau (Dong et al., 2015). Four overgrazed meadows in an area of ∼ 200 square kilometers were studied along an increasing elevation ranging from 3000 to 3600 m above mean sea level in the eastern part of the Qinghai-Tibetan Plateau. These meadows have undergone overgrazing at a comparable intensity as they have been overgrazed by approximately the same number of yaks (approximately four yaks per hectare) in winter season every year, for nearly 30 years. These four overgrazed meadows were distributed in Hezuo City, Gansu province (34°°5′N, 102°42′E; 3000 a.s.l.), Maqu County Gansu province (34°0′N, 102°44′E; 3400 and 34°5′N, 102°10′E 3600 a.s.l.), Ruergai County, and Sichuan province (34°1′N, 102°43′E; 3200 a.s.l.) of China (Figure 1A). The histories for the overgrazed meadows were obtained by interviewing local farmers. All overgrazed meadows are characterized by a cold and dry climate; with a mean annual temperature of 2.4–3.2°C. Mean annual precipitation of 530 to 560 mm is primarily distributed from July to August. The local vegetation in overgrazed sites is dominated by herbaceous species, such as Ajania tenuifolia, Swertia tetraptera, and Taraxacum mongolicum (Zhang et al., 2013). Soils are classified as alpine meadow soils (Zhang et al., 2018).
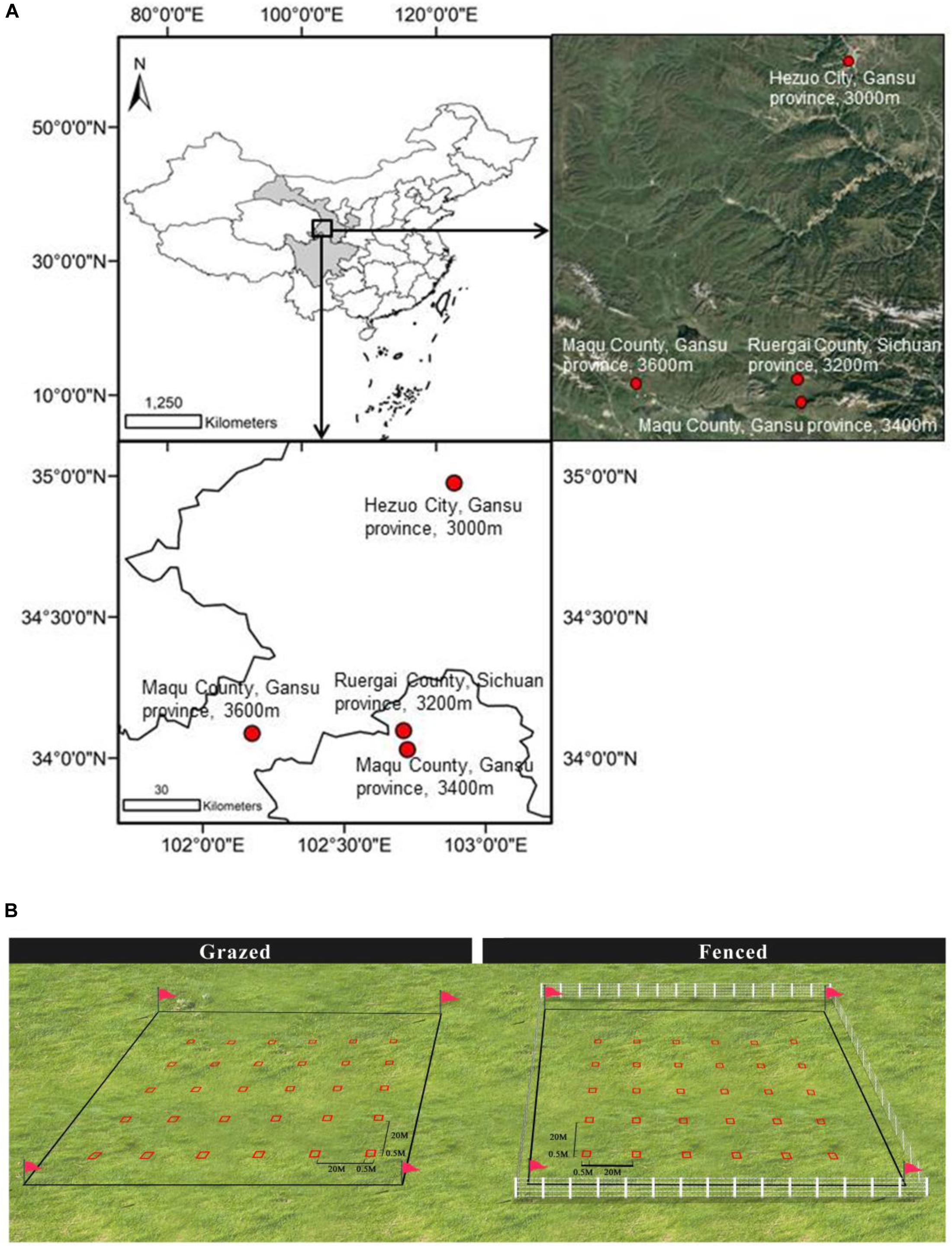
Figure 1. The topographic map of the four study sites along an increasing elevation ranging from 3000 m to 3600 m above mean sea level in the eastern part of the Qinghai-Tibetan Plateau, China (A), and the quadrat sampling design (B). At each site, there was one overgrazed and one matched fenced meadow and 30 0.5 m × 0.5 m quadrats ware randomly arranged in both overgrazed and fenced meadows (B).
Since year 2000, the Chinese government has established fences to protect some areas of overgrazed meadows. Thus, in each elevation, there are also fenced meadows and the fenced meadow has been protected from grazing for nearly 17 years. The climate and soil type of each fenced meadow are the same as each overgrazed meadow. However, the local vegetation in fenced sites in 3000 and 3200 m elevations (sub-alpine meadow) are dominated by herbaceous species, such as Elymus nutans, Kobresia pygmaea, Scirpus pumilus, and Thermopsis lanceolata (Zhang et al., 2018). In contrast dominating species in fenced sites in 3400 m and 3600 m elevations (alpine meadow) are Scirpus pumilus, Kobresia capillifolia, Festuca ovina, Poa poophagorum, Roegneria nutans, and Kobresia macrantha (Chu et al., 2007).
At each elevation, we adopted paired-sampling method to use the fenced meadow which has been constructed since 2000 for preventing grazing and its adjacent overgrazed meadow for comparison (Figure 1B). Sampling was conducted in August, during the peak of the growing season in 2017. At each of the four elevations, 30 plots, each of 0.5 m × 0.5 m, were arranged along four parallel transects in the fenced area and the adjacent overgrazed area of one meadow, with a 20 m spacing between the closest edges of the two areas (Figure 1B). The plots were spaced at 20 m intervals. For each of the 30 plots, species presence/absence was recorded. Total above-ground biomass of each species in each of the 30 plots was also measured to represent abundances (numbers of individuals) for each species to determine whether overgrazing change the dominance of annual and perennial species.
All common species shared between fenced and grazed meadows were sampled to measure three above-ground traits: leaf photosynthesis rate (A), specific leaf area (SLA) and above-ground maximum height (H). Further, five root traits which include root average diameter (RD), root biomass (RB), specific root length (SRL), root tissue density (RTD), and specific root area (SRA) were also recorded. All these traits could be good predictors of resource acquisition vs. resource conservation trade-off (Díaz et al., 2001; Roumet et al., 2006). Traits were measured from three to five mature individuals per species, following the procedures of measurements described in Bergmann et al. (2017) and Zhang et al. (2018) as well as mentioned in the Supplementary Material.
First, we utilized non-parametric analysis (a Wilcoxon signed-rank test) to test whether there were differences in the above- and below-ground traits and abundances for all common species shared between fenced and overgrazed sites at all four elevations using. Next, a Spearman correlation test was used to examine the bivariate relationships among all above- and below-ground plant traits at all elevations. Finally, a principal component analysis (PCA) was employed to evaluate which of the eight above-ground and root traits play an important role in species distribution between fenced and overgrazed meadows at all elevations. Given the unique set of traits between annual and perennial plant species (Roumet et al., 2006; Zhang et al., 2018), the above analyses were also performed separately for annuals and perennials to understand whether the response of above- and below-ground traits to overgrazing vary between annual and perennial species.
In total, we sampled 53–77 species in the fenced meadows from 3000 to 3600 m elevations, whereas only 40–56 species, could be identified in the sampling of the overgrazed meadows (Supplementary Figure 1). However, species composition was similar between the fenced and overgrazed meadows as the two meadows shared 28–55 species (Supplementary Figure 1). When dividing all species into annual and perennial species separately, we found annual species was promoted in the overgrazed meadow. That was because although overgrazing nearly has no influences on numbers of species for annual species (Supplementary Figure 1), but abundances for shared annual species in overgrazed meadows were approximately 4.3 times as those in the fenced meadows at all elevations (Supplementary Figure 2). However, perennial species were got largely removal in the overgrazed meadow, as overgrazing had not only removed 18–22 perennial species (Supplementary Figure 1), abundances for shared perennial species in the overgrazed meadows but also were only 1/3 of those in the fenced meadows (Supplementary Figure 2). For further analysis of below- and above-ground traits, we focused on these common species for direct comparisons of the parameters across overgrazed and fenced meadows at all the elevations.
All shared species had significantly lower root biomass (RB), root average diameter (RD), root tissue density (RTD), and above-ground maximum height (H) in the overgrazed meadows than those in the fenced meadows at all four elevations. In contrast, overgrazed meadows had higher specific root length (SRL), specific root area (SRA), leaf photosynthesis rate (A) and specific leaf area (SLA) than those in the fenced meadow (Figure 2). Consistent results could still be observed in perennial species, when we performed Wilcoxon signed-rank test separately for annuals and perennials (Figure 2). However, for annual species, differences in all above-ground and below-ground traits between fenced and overgrazed meadows were not significant (Figure 2).
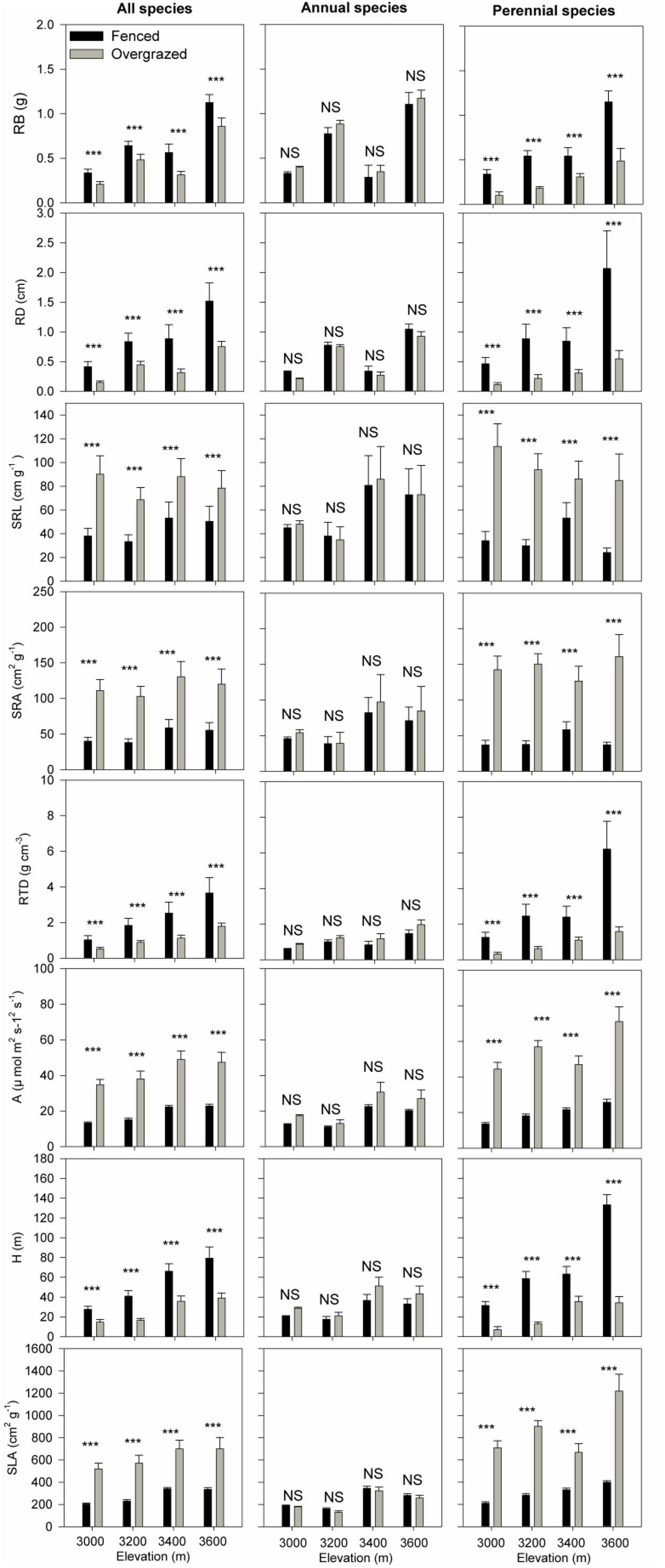
Figure 2. The differences in above-ground traits [maximum height (H), leaf maximum photosynthesis rate (A) and specific leaf area (SLA)] and below-ground traits [root average diameter (RD), root biomass (RB), specific root length (SRL), specific root area (SRA) and root tissue density (RTD)] between fenced and overgrazed meadows for all species (left panel) and separately for annual (middle panel) and perennial (right panel) species from 3000 to 3600 m elevations. *** and NS (non-significant) indicates P < 0.001 and P > 0.05 based on Wilcoxon signed-rank tests.
We observed strong correlations amongst the five root traits, as well as between the three above-ground traits, for all common species in both, fenced and grazed meadows at all four elevations (Figure 3). For example, RD, H, RB, and RTD are all significantly positively correlated (Figure 3). Similarly, SRL, A, SRA, and SLA are also all significantly positively correlated (Figure 3). However, RD, H, RB, and RTD are significantly negatively associated with SRL, A, SRA, and SLA (Figure 3). However, when we tested these correlations separately for annuals and perennials, consistent significant traits correlations could be observed in perennial species in both fenced and overgrazed meadows (Figure 3). However, for annual species all trait correlations were non-significant in both fenced and overgrazed meadows (Figure 3).
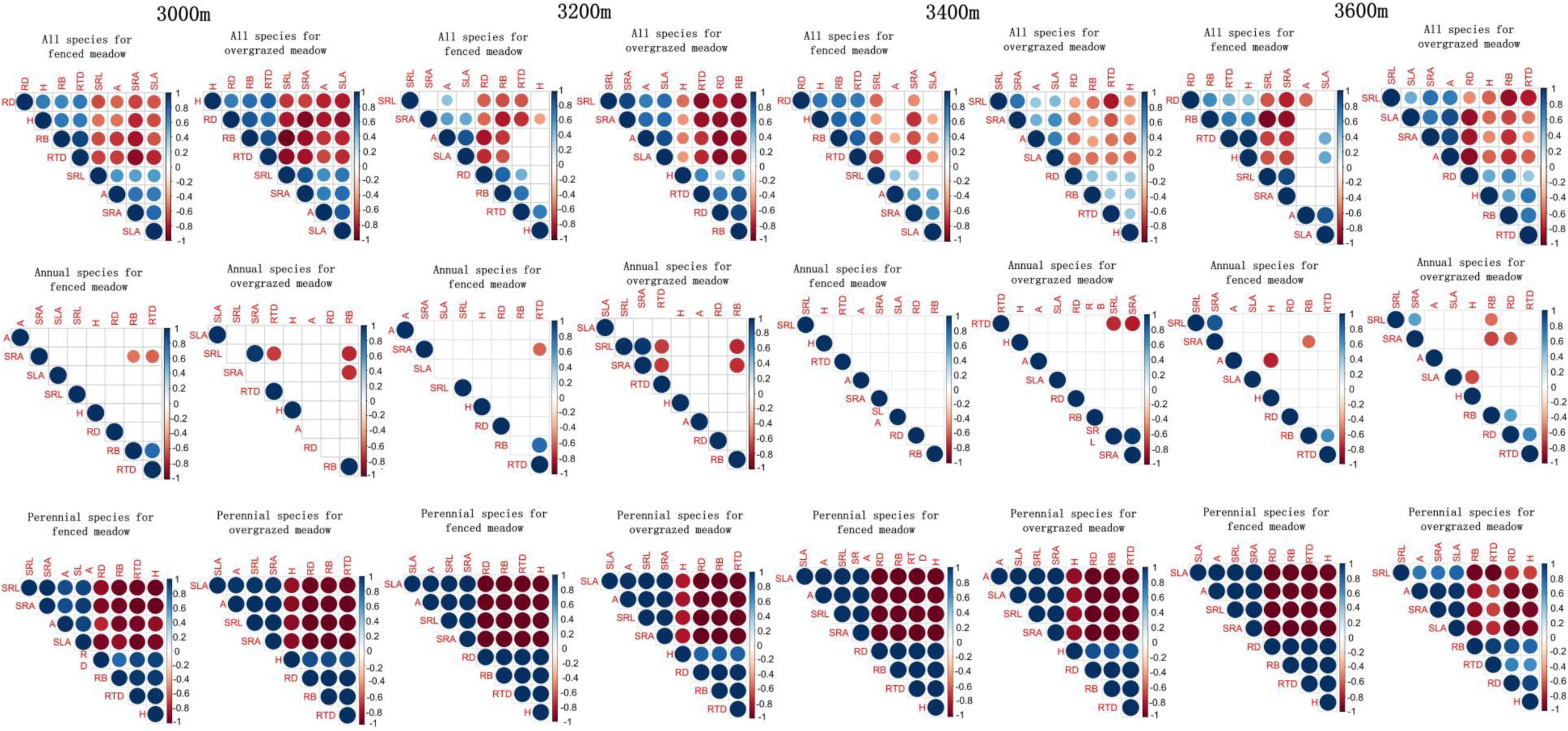
Figure 3. The interrelationships among above-ground traits (H, A, and SLA) and below-ground traits (RD, RB, SRL, SRA, and RTD) in fenced and overgrazed meadows for all species (upper panel), and separately for annuals (middle panel) and perennial species (lower panel) from 3000 to 3600 m elevations.
Results of PCA revealed that “species of fenced and overgrazed meadows” were significantly separated by above- and below-ground traits measured at all elevations (Figure 4, and Supplementary Table 2). When performing PCA separately for annual and perennial species, we found that perennial species in fenced and overgrazed meadows were significantly separated by all the above- and below-ground traits measured at all elevations (Figure 4 and Supplementary Table 2). In contrast, annual species in fenced and overgrazed meadows could not be significantly separated by any of measured above- and below-ground traits at all elevations (Supplementary Figure 3 and Supplementary Table 2).
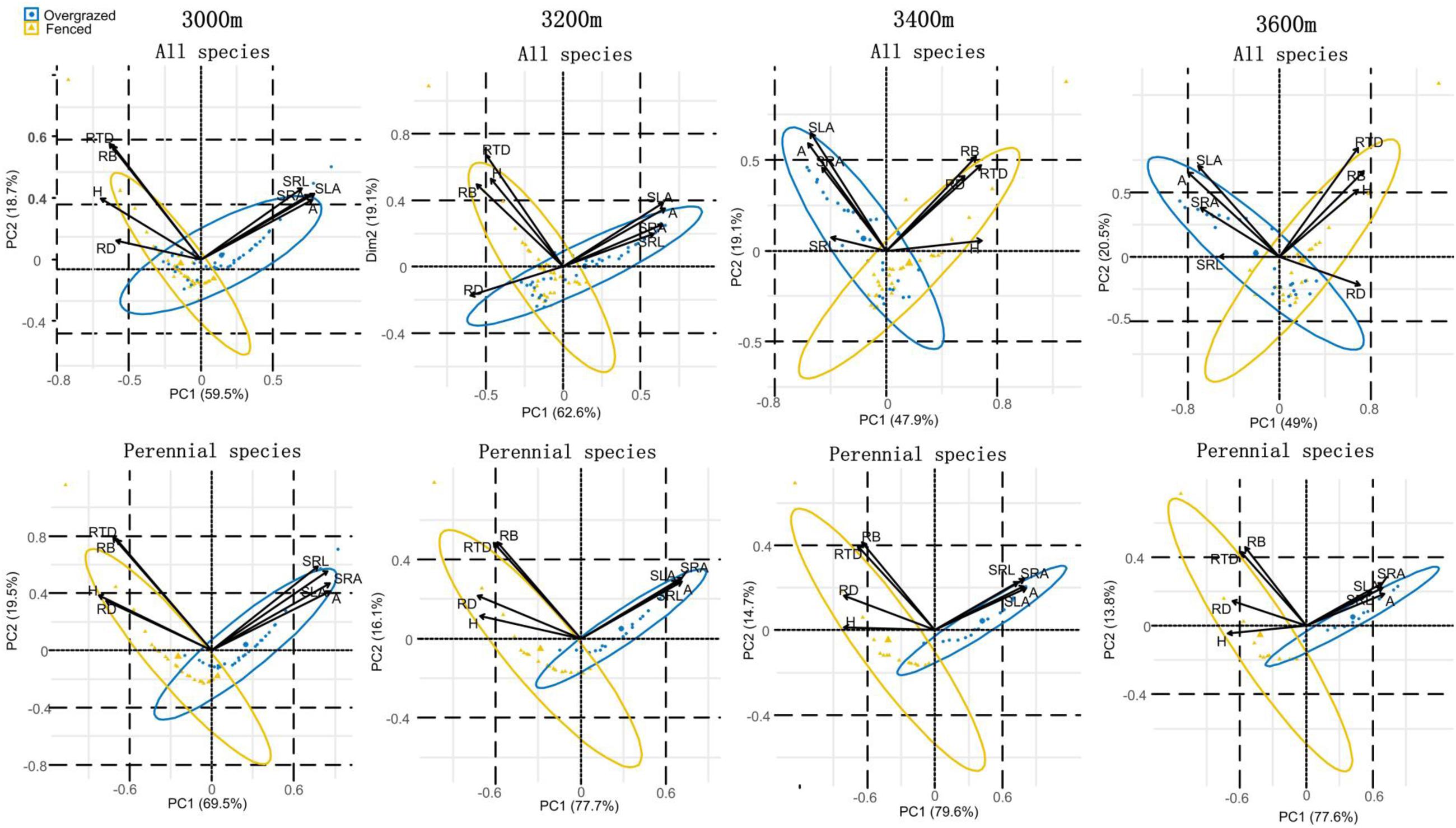
Figure 4. Principal component analysis of below-ground (RD, RB, SRL, SRA, and RTD) and above-ground traits (H, A, and SLA) in fenced and overgrazed meadows for all species, and separately for perennial species from 3000 to 3600 m elevations.
In this study, we found both above-ground and below-ground traits exhibited a similar resource acquisition vs. resource conservation trade-off for responding to overgrazing. Moreover, the response of above- and below-ground traits to overgrazing differed between annual and perennial species, with perennial species determining the response of above- and below-ground traits to overgrazing.
A shift from resource conservation to resource acquisition at above-ground level have resulted higher SLA, A, but lower H after overgrazing as compared to those in the fenced condition (Díaz et al., 2001; Evju et al., 2009; Spasojevic et al., 2010; Laliberté and Tylianakis, 2012; Li et al., 2017). This shift from resource conservation to resource acquisition has also led to significant relationships among these three traits at both fenced and grazed meadows (Wright et al., 2004; Osnas et al., 2013). Here, we observed all fenced meadows, had higher H, but lower SLA and A, compared to overgrazed meadows. We also found significant correlations among these three traits in both fenced and overgrazed meadows at all four elevations. This indicated that a trade-off between resource acquisition vs. resource conservation at above-ground level likely determined plant overgrazing response in Qinghai Tibetan plateau.
A number of studies have found that the trade-off between resource acquisition vs. resource conservation can also be found in below-ground traits (Withington et al., 2006; Mommer and Weemstra, 2012). For example, resource acquisition may warrant high SRL, SRA, but low RTD, RB, and RD to allow fast-growth, whereas, resource conservation may result in high RTD, RB, and RD, but low SRL and SRA (Reich, 2014; Roumet et al., 2016). Likewise, this shift from resource acquisition to resource conservation at below-ground level can also lead to strong correlations among these below-ground traits (Withington et al., 2006; Prieto et al., 2015; de la Riva et al., 2018). Here we found all fenced meadows, had higher SRL, SRA, but lower RTD, RB, and RD, compared to overgrazed meadows. We also found significant correlations among these five below-ground traits. Thus, plants also alter their below-ground traits so that they can shift from resource conservation to resource acquisition to avoid overgrazing, in a manner like shifts in response of above-ground traits.
Principal component analysis revealed that fenced and overgrazed species were significantly separated, which in turn was significantly positively associated with SRL, SRA, A, and SLA, but negatively related RD, RB, RTD, and H. High SRL, SRA, A, and SLA represents high resource acquisition, whereas, high RD, RB, RTD, and H indicates high resource conservation (Wright et al., 2004; Reich, 2014; de la Riva et al., 2018). Thus, the shift from resource conservation to resource acquisition determines the classification of species between fenced and overgrazed meadows. These results together indicate that above- and below- ground traits studied here can capture the key plant response (a shift from resource conservation to resource acquisition) and thus help plants to avoid overgrazing. As a result, these traits can be used to select potential species to re-design overgrazed meadows for restoration in Qinghai-Tibetan Plateau.
In the fenced meadows, resource conservation can also result in low photosynthetic rates and SLA. Moreover, high overall conservation of resources requires conservative root structures such as roots with high root tissue density, root diameter and biomass, and/or low specific root length and area (Eissenstat, 2002; Fort et al., 2013). In contrast, in the overgrazed meadows, resource acquisition relates to high photosynthetic rates and SLA, which requires roots with high specific root length and area, and low root tissue density, diameter and biomass to ensure sufficient nutrient supply to achieve fast plant growth (Reich, 2014; de la Riva et al., 2018). Therefore, significant above-ground -below-ground traits relationship may be observed in both fenced and grazed meadows. Here we observed consistently significant relationships between above- and below- ground traits in both fenced and overgrazed meadows at all elevations. We infer that above-ground traits might act as a proxy for below-ground traits in revealing plant-soil feedbacks and the effects of below-ground traits on ecosystem functioning in overgrazed meadows, as evident in Qinghai-Tibetan Plateau. Meanwhile, above-ground traits can also be a good proxy for below-ground traits to select species to re-design communities to perform restoration of overgrazed meadows.
It has been found that grazing promotes annual over perennial species in global grassland ecosystems (Díaz et al., 2007). Our study supports this trend where we find that abundances for shared annual species in the overgrazed meadows are 4 times as much as those in the fenced meadows. In contrast, abundances for shared perennial species in the overgrazed meadows are merely 1/3 of those in the fenced meadows. However, we observed the responses of above-ground and below-ground traits to overgrazing differ between annual and perennial species. That is because when performing trait variations, trait correlations and PCA analysis separately for annuals and perennials, consistently significant trait variations and trait correlations can still be observed in perennial species. Both aboveground and belowground traits can still significantly classify perennial species between fenced and overgrazed meadows. However, annual species did not have any trait variations and trait correlations in both fenced and overgrazed meadows. Moreover, both aboveground traits and belowground traits cannot differentiate annual species between fenced and overgrazed meadows. Thus, perennial species alter their above-ground and below-ground traits to develop a shift from resource conservation to resource acquisition to avoid overgrazing. While, annual species do not make any response to overgrazing.
Overgrazing is common from low elevation (3000 m) to high elevation (4000 m) (Cui and Graf, 2009). Here our results clearly observed that at all four elevations (3000, 3200, 3400, and 3600 m) annual and perennial species have consistent responses to overgrazing. Thus, we conclude that overgrazing can result in largely removal of perennial species, which force perennial specie to develop above- and below-ground traits that are highly associated with high resource acquisition to avoid overgrazing. In contrast, overgrazing promotes annual species, thus annual species do not necessarily make any response to overgrazing. These results filled the gap of the effects of life-forms (annual vs. perennial) on species-specific response at both above-, and below-ground levels during overgrazing. More important these findings have important implications for the sustainable ecosystem management and recovery of overgrazed ecosystems in Qinghai-Tibetan Plateau and beyond. First, since above-ground traits can be a good proxy for below-ground traits in revealing key overgrazing responses of plants, we can use above-ground traits and trait-based species selection model (Laughlin, 2014) to select many potential perennial species that have high resource acquisition ability from natural meadow ecosystems for increasing diversity of overgrazed meadows. Above-ground traits and trait-based species selection model can also be used to select potential perennial species for specific sustainable ecosystem management. It has been found that decreased species diversity and biomass resulting from overgrazing is the key driver of soil N loss and greenhouse gas emission (e.g., N2O) in Qinghai-Tibetan Plateau (Liu et al., 2018). As a result, active seeding of these selected perennial species may have high potential to diminish soil N loss and N2O emission. Moreover, using above-ground traits to select potential perennial species that have high resource acquisition and high biomass can provide feed for cultivating as many yaks or sheep as possible to meet ever-increasing food demand, while simultaneously increasing ecosystem productivity and soil C storage. Similarly, selecting potential perennial legume species that have high resource acquisition can help in increasing soil nitrogen retention. In addition, selecting desirable species with high resource acquisition to undesirable weeds may help control weeds in overgrazed meadows.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
HZ, SP, and CW designed the research. HZ, QH, and KJ performed the research. HZ and SP analyzed the data and wrote the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
This work was funded by Scientific Research Project of Ecological Restoration of Baopoling Mountain in Sanya, the National Natural Science Foundation of China (41905094 and 41991285) and a start-up fund from Hainan University [KYQD (ZR) 1876].
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This manuscript has been released as a pre-print at https://www.biorxiv.org/content/10.1101/771444v1 (Zhang and Long, 2019).
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.573948/full#supplementary-material
Asner, G. P., Elmore, A. J., Olander, L. P., Martin, R. E., and Harris, A. T. (2004). Grazing systems, ecosystem responses, and global change. Annu. Rev. Environ. Resour. 29, 261–299. doi: 10.1146/annurev.energy.29.062403.102142
Bergmann, J., Ryo, M., Prati, D., Hempel, S., and Rillig, M. C. (2017). Root traits are more than analogues of leaf traits: the case for diaspore mass. New Phytol. 216, 1130–1139. doi: 10.1111/nph.14748
Briske, D. D. (1999). “Plant traits determining grazing resistance: why have they proved so elusive?,” in Proceedings of the VIth International Rangeland Congress, eds D. Eldridge and D. Freudenberger (Charters Towers, QLD: Australian Rangeland Society), 901–905.
Chu, C. J., Wang, Y. S., Du, G. Z., Maestre, E. T., Luo, Y. J., and Wang, G. (2007). On the balance between niche and neutral processes as drivers of community structure along a successional gradient: insights from alpine and sub-alpine meadow communities. Ann. Bot. 100, 807–812.
Cui, X., and Graf, H. F. (2009). Recent land cover changes on the Tibetan Plateau: a review. Clim. Change 94, 47–61. doi: 10.1007/s10584-009-9556-8
de la Riva, E. G., Marañón, T., Pérez-Ramos, I. M., Navarro-Fernández, C. M., Olmo, M., and Villar, R. (2018). Root traits across environmental gradients in Mediterranean woody communities: are they aligned along the root economics spectrum? Plant Soil 424, 35–48. doi: 10.1007/s11104-017-3433-4
Díaz, S., Lavorel, S., Mcintyre, S., Falczuk, V., Casanoves, F., Milchunas, D. G., et al. (2007). Plant trait responses to grazing – a global synthesis. Glob. Change Biol. 13, 313–341. doi: 10.1111/j.1365-2486.2006.01288.x
Díaz, S., Noy-Meir, I., and Cabido, M. (2001). Can grazing response of herbaceous plants be predicted from simple vegetative traits? J. Appl. Ecol. 38, 497–508. doi: 10.1046/j.1365-2664.2001.00635.x
Dong, Q. M., Zhao, X. Q., Wu, G. L., and Chang, X. F. (2015). Optimization yak grazing stocking rate in an alpine grassland of Qinghai-Tibetan Plateau, China. Environ. Earth Sci. 73, 2497–2503. doi: 10.1007/s12665-014-3597-7
Eissenstat, D. (2002). Root structure and function in an ecological context. New Phytol. 148, 353–354. doi: 10.1046/j.1469-8137.2000.00781.x
Evju, M., Austrheim, G., Halvorsen, R., and Mysterud, A. (2009). Grazing responses in herbs in relation to herbivore selectivity and plant traits in an alpine ecosystem. Oecologia 161, 77–81. doi: 10.2307/40310116
Feng, C., Wang, Z., Zhu, Q., Fu, S., and Chen, H. Y. H. (2018). Rapid increases in fine root biomass and production following cessation of anthropogenic disturbances in degraded forests. Land Degrad. Dev. 29, 461–470. doi: 10.1002/ldr.2878
Fort, F., Jouany, C., and Cruz, P. (2013). Root and leaf functional trait relations in Poaceae species: implications of differing resource-acquisition strategies. J. Plant Ecol. 41, 107–127. doi: 10.1093/jpe/rts034
Klumpp, K., Fontaine, S., Attard, E., Le Roux, X., Gleixner, G., and Soussana, J. F. (2009). Grazing triggers soil carbon loss by altering plant roots and their control on soil microbial community. J. Ecol. 97, 876–885. doi: 10.1111/j.1365-2745.2009.01549.x
Kramer-Walter, K. R., Bellingham, P. J., Millar, T. R., Smissen, R. D., Richardson, S. J., and Laughlin, D. C. (2016). Root traits are multidimensional: specific root length is independent from root tissue density and the plant economic spectrum. J. Ecol. 104, 1299–1310. doi: 10.1111/1365-2745.12562
Laliberté, E., and Tylianakis, J. M. (2012). Cascading effects of long-term land-use changes on plant traits and ecosystem functioning. Ecology 93, 145–155. doi: 10.1890/11-0338.1
Laughlin, D. C. (2014). Applying trait-based models to achieve functional targets for theory-driven ecological restoration. Ecol. Lett. 17, 771–784.
Li, W., Xu, F., Zheng, S., Taube, F., and Bai, Y. (2017). Patterns and thresholds of grazing-induced changes in community structure and ecosystem functioning: species-level responses and the critical role of species traits. J. Applied. Ecol. 54, 963–975. doi: 10.1111/1365-2664.12806
Liu, S., Zamanian, K., Schleuss, P. M., Zarebanadkouki, M., and Kuzyakov, Y. (2018). Degradation of Tibetan grasslands: consequences for carbon and nutrient cycles. Agric. Ecosyst. Environ. 252, 93–104.
McInenly, L. E., Merrill, E. H., Cahill, J. F., and Juma, N. G. (2010). Festuca campestris alters root morphology and growth in response to simulated grazing and nitrogen form. Funct. Ecol. 24, 283–292. doi: 10.1111/j.1365-2435.2009.01642.x
Mommer, L., and Weemstra, M. (2012). The role of roots in the resource economics spectrum. New Phytol. 195, 725–727. doi: 10.1111/j.1469-8137.2012.04247.x
Mouillot, D., Villeger, S., Scherer-Lorenzen, M., and Mason, N. W. H. (2011). Functional structure of biological communities predicts ecosystem multifunctionality. PLoS One 6:e17476. doi: 10.1371/journal.pone.0017476
Orwin, K. H., Buckland, S. M., Johnson, D., Turner, B. L., Smart, S., Oakley, S., et al. (2010). Linkages of plant traits to soil properties and the functioning of temperate grassland. J. Ecol. 98, 1074–1083. doi: 10.1111/j.1365-2745.2010.01679.x
Orwin, K. H., Mason, N. W. H., Jordan, O. M., Lambie, S. M., Stevenson, B. A., and Mudge, P. L. (2018). Season and dominant species effects on plant trait-ecosystem function relationships in intensively grazed grassland. J. Appl. Ecol. 55, 236–245.
Osnas, J. L. D., Lichstein, J. W., Reich, P. B., and Pacala, S. W. (2013). Global leaf trait relationships: mass, area, and the leaf economics spectrum. Science 340, 741–744. doi: 10.1126/science.1231574
Prieto, I., Roumet, C., Cardinael, R., Dupraz, C., Jourdan, C., Kim, J. H., et al. (2015). Root functional parameters along a land-use gradient: evidence of a community-level economics spectrum. J. Ecol. 103, 361–373. doi: 10.1111/1365-2745.12351
Reich, P. B. (2014). The world-wide ‘fast–slow’ plant economics spectrum: a traits manifesto. J. Ecol. 102, 275–301. doi: 10.1111/1365-2745.12211
Roumet, C., Birouste, M., Picon-Cochard, C., Ghestem, M., Osman, N., Vrignon-Brenas, S., et al. (2016). Root structure–function relationships in 74 species: evidence of a root economics spectrum related to carbon economy. New Phytol. 210, 815–826. doi: 10.1111/nph.13828
Roumet, C., Urcelay, C., and Díaz, S. (2006). Suites of root traits differ between annual and perennial species growing in the field. New. Phytol. 170, 357–368. doi: 10.1111/j.1469-8137.2006.01667.x
Spasojevic, M. J., Aicher, R. J., Koch, G. R., Marquardt, E. S., Mirotchnick, N., Troxler, T. G., et al. (2010). Fire and grazing in a mesic tallgrass prairie: impacts on plant species and functional traits. Ecology 91, 1651–1659. doi: 10.2307/25680406
Ward, S. E., Smart, S. M., Quirk, H., Tallowin, J. R. B., Mortimer, S. R., Shiel, R. S., et al. (2016). Legacy effects of grassland management on soil carbon to depth. Glob. Change Biol. 22, 2929–2938. doi: 10.1111/gcb.13246
Westoby, M. (1999). “The LHS strategy in relation to grazing and fifire,” in Proceedings of the VIth International Rangeland Congress, eds D. Eldridge and D. Freudenberger (Charters Towers, QLD: Australian Rangeland Society), 893–896.
Withington, J. M., Reich, P. B., Oleksyn, J., and Eissenstat, D. M. (2006). Comparisons of structure and life span in roots and leaves among temperate trees. Ecol. Monogr. 76, 381–397.
Wright, I. J., Reich, P. B., Westoby, M., Ackerly, D. D., Baruch, Z., Bongers, F., et al. (2004). The worldwide leaf economics spectrum. Nature 428, 821–827. doi: 10.1038/nature02403
Zhang, H., Gilbert, B., Wang, W., Liu, J., and Zhou, S. (2013). Grazer exclusion alters plant spatial organization at multiple scales, increasing diversity. Ecol. Evol. 3, 3604–3612. doi: 10.1002/ece3.743
Zhang, H., John, R., Zhu, S., Liu, H., Xu, Q., Qi, W., et al. (2018). Shifts in functional trait–species abundance relationships over secondary subalpine meadow succession in the Qinghai–Tibetan Plateau. Oecologia 188, 547–557. doi: 10.1007/s00442-018-4230-3
Zhang, H., and Long, W. (2019). Can the response of above- and below-ground traits to overgrazing vary between annual and perennial species? bioRxiv [Preprint] doi: 10.1101/771444
Keywords: above-and below-ground traits, grazing avoidance strategy, land use, life form, Qinghai-Tibetan Plateau, resource conservation vs resource acquisition trade-off
Citation: Zhang H, He Q, Pandey SP, Jiang K and Wang C (2021) Can Overgrazing Responses Be Disentangled by Above- and Below-Ground Traits? Front. Ecol. Evol. 9:573948. doi: 10.3389/fevo.2021.573948
Received: 18 June 2020; Accepted: 04 January 2021;
Published: 28 January 2021.
Edited by:
Orsolya Valkó, Centre for Ecological Research, Hungarian Academy of Sciences, HungaryReviewed by:
Jennifer Lesley Silcock, The University of Queensland, AustraliaCopyright © 2021 Zhang, He, Pandey, Jiang and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chen Wang, Y2hlbi53YW5nQHNjYmcuYWMuY24=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.