 Jennifer N. Phillips
Jennifer N. Phillips W. Justin Cooper3
W. Justin Cooper3 David A. Luther
David A. Luther Elizabeth P. Derryberry
Elizabeth P. Derryberry
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 23 December 2020
Sec. Behavioral and Evolutionary Ecology
Volume 8 - 2020 | https://doi.org/10.3389/fevo.2020.587120
This article is part of the Research Topic Behavioral Adaptations to Life in the City View all 13 articles
Human activity around the globe is a growing source of selection pressure on animal behavior and communication systems. Some animals can modify their vocalizations to avoid masking from anthropogenic noise. However, such modifications can also affect the salience of these vocalizations in functional contexts such as competition and mate choice. Such is the case in the well-studied Nuttall's white-crowned sparrow (Zonotrichia leucophrys nuttalli), which lives year-round in both urban San Francisco and nearby rural Point Reyes. A performance feature of this species' song is salient in territorial defense, such that higher performance songs elicit stronger responses in simulated territorial intrusions; but songs with lower performance values transmit better in anthropogenic noise. A key question then is whether vocal performance signals male quality and ability to obtain high quality territories in urban populations. We predicted white-crowned sparrows with higher vocal performance will be in better condition and will tend to hold territories with lower noise levels and more species-preferred landscape features. Because white-crowned sparrows are adapted to coastal scrub habitats, we expect high quality territories to contain lower and less dense canopies, less drought, more greenness, and more flat open ground for foraging. To test our predictions, we recorded songs and measured vocal performance and body condition (scaled mass index and fat score) for a set of urban and rural birds (N = 93), as well as ambient noise levels on their territories. Remote sensing metrics measured landscape features of territories, such as drought stress (NDWI), greenness (NDVI), mean canopy height, maximum height, leaf area density (understory and canopy), slope, and percent bare ground for a 50 m radius on each male territory. We did not find a correlation between body condition and performance but did find a relationship between noise levels and performance. Further, high performers held territories with lower canopies and less dense vegetation, which are species-preferred landscape features. These findings link together fundamental aspects of sexual selection in that habitat quality and the quality of sexually selected signals appear to be associated: males that have the highest performing songs are defending territories of the highest quality.
Urban habitats are an ever-expanding ecosystem with strong selection pressures for wildlife that inhabit them, and the degree of habitat alteration and pollution often determines whether species are urban adapters or urban avoiders (Blair, 1996; Croci et al., 2008; Aronson et al., 2016). Anthropogenic noise, introduced vegetation, exotic predators, increased risk of disease, and smaller patches of suitable habitat all may affect species composition, the population densities of species, and how individual animals behave in an urban environment (Shochat et al., 2010; Aronson et al., 2014; Marzluff, 2017). For species that do persist in urban ecosystems, these selective pressures may compromise their badges of status and sexually selected signals (Swaddle et al., 2015), hampering individuals' abilities to attain and maintain resources that benefit survival and reproductive success. For example, bird song functions in acquisition and defense of breeding territories and attraction of mates (Catchpole and Slater, 2008), and is known to be under selection pressures from urban noise (Slabbekoorn, 2013). However, links between territory quality, individual quality and signal performance have rarely, if ever, been investigated, and to our knowledge never in the context of anthropogenic disturbance.
Obtaining a good territory is the key to higher fitness for most wildlife. Territories with preferred nesting vegetation result in fewer nest failures (Kaiser et al., 2009) and higher fecundity (Pidgeon et al., 2006). Similarly, territories with higher quality food resources result in higher survival rates (Senar et al., 2002). For birds, males without territories, or “floaters,” scope out territories by assessing habitat quality features like foraging patches and assessing social context as to whether current territory holders are successful nesters (Pärt et al., 2011). How phenotypic traits, such as song, affect the ability to obtain relatively high quality territories is an outstanding question in behavioral ecology (Andersson, 1994; Catchpole and Slater, 2008). Previous research provides some insight – singing longer, increased age, larger body size, and aggressive behavior are often associated with securing territories (e.g., Yasukawa, 1979; Clarkson, 2007). For example, prothonotary warblers (Protonotaria citrea) prefer to nest near wet or flooded areas, and males that sing longer are more likely to attain territories near water; and thus song seems to be used for males to assess the condition of rivals (Clarkson, 2007). Similarly, dominant males in the Paridae family have access to preferred resources (Ekman, 1987; Desrochers, 1989), and male black-capped chickadees (Poecile atricapillus) in high quality mature forest signal their competitive ability with more consistent song structure (van Oort et al., 2006; Grava et al., 2012, 2013) and higher song output (van Oort et al., 2007). Blue-black grassquits (Volatinia jacarina) that sing more often and consistently have territories with higher seed availability (Manica et al., 2014). However, there are still gaps in our knowledge about how song affects the ability of males to attain and defend high quality territories.
Fewer studies have looked at urban habitat quality (Shochat et al., 2006), and especially not fine-grain habitat characteristics of territories, or how these features might vary with performance of sexually selected signals. Instead of fine-grain metrics like types of cover, studies focus on the rural-urban dichotomy. For example, urban house sparrows (Passer domesticus) are smaller and in worse condition than rural birds (Liker et al., 2008), and body size varies with urbanization level (Meillère et al., 2015). Telomere length, which affects life longevity, is also shortened in birds raised in urban conditions (Salmón et al., 2016), but the specific differences between urban and rural conditions (e.g., noise, vegetation) was not examined. Furthermore, none of these aforementioned studies pair song data with habitat data. Only one system to date has investigated whether song traits vary with habitat quality and signaler quality across anthropogenic habitats (Narango and Rodewald, 2016, 2018). In this case, male northern cardinals (Cardinalis cardinalis), body size correlates with song speed and length in rural areas but this pattern does not hold across urban and rural locations (Narango and Rodewald, 2018). Additionally, minimum frequency increased with increased understory density, and urban birds sang faster, longer songs (Narango and Rodewald, 2016, 2018). However, song was not related to provisioning ability or number of offspring, although offspring were in worse condition in cities (Narango and Rodewald, 2018). Therefore, there is much that is not understood about the interaction between habitat characteristics, signaling traits, and body condition of individual birds, especially along urban to rural gradients.
How anthropogenic noise affects sexually selected signals used to attain territories has received much attention in recent years. Many avian species adjust their song pitch in noise with long-term, cultural changes, or real-time plasticity shifts in minimum frequency, and thereby avoid masking by high energy, low frequency background noise (Derryberry et al., 2016; LaZerte et al., 2016; Luther et al., 2016 reviewed in Slabbekoorn, 2013). Oftentimes this shift in minimum frequency is associated with reduced bandwidth (Dowling et al., 2011; Luther et al., 2016; Phillips and Katti, 2020), or changes in note rates (Slabbekoorn and den Boer-Visser, 2006; Cartwright et al., 2014), which can affect performance of a physiological tradeoff between repeated and broadband notes (Podos, 1996), and result in lower quality song performance.
A variety of vocal species have performance limits on the rate at which broadband notes can be repeated because of mechanical constraints (e.g., primates: Clink et al., 2018; mice: Pasch et al., 2011; birds: Podos, 1996, 1997). This tradeoff, known as vocal performance, is used in male-male competition (red-winged blackbirds, Agelaius phoeniceus: Cramer and Price, 2007; banded wrens, Thryophilus pleurostictus: Illes et al., 2006; de Kort et al., 2009; swamp sparrows, Melospiza georgiana: Dubois et al., 2011; Moseley et al., 2013; white-crowned sparrows, Zonotrichia leucophrys: Phillips and Derryberry, 2017a,b, 2018) and female mate choice (swamp sparrows: Ballentine et al., 2004; Lincoln's sparrows, Melospiza lincolnii: Caro et al., 2010; banded wrens: Cramer et al., 2011; canaries, Serinus canaria: Drăgănoiu et al., 2002; singing mice (Scotinomys spp.): Pasch et al., 2011). Limited research has examined the signaler qualities conveyed by vocal performance to receivers, but in nightingales (Luscinia megarhynchos) high performance males are older and sing more consistently (Sprau et al., 2013). In Java sparrows (Lonchura oryzivora) performance increases over time, giving listeners reliable information about the age of the singer (Ota and Soma, 2014). Swamp sparrow males with higher performance also are larger and older (Ballentine, 2009), and aggressive intent is also conveyed through vocal performance (DuBois et al., 2009).
One species that persists across urban and rural areas is Nuttall's white-crowned sparrow (Zonotrichia leucophrys nuttalli). Nutall's white-crowned sparrows are a year-round resident of the narrow strip of coastal scrub in northern and central California, and have been well-documented in the Bay Area of California for the past 50 years (Baptista, 1975; Petrinovich and Patterson, 1982; Dewolfe et al., 1989). This subspecies sings one song type and is known for forming regional dialects (Baptista, 1975). Once a song is crystallized after the first year, songs are highly repeatable (Phillips and Derryberry, 2017b) and used to attain territories (Dewolfe et al., 1989). Before human settlement, San Francisco consisted of mostly dunes and scrub habitat. This native habitat was largely converted to human use in the late 1800's and early 1900's, with remaining natural areas often converted from scrub to forests of blue gum eucalyptus (Eucalyptus globulus), Monterey pine (Pinus radiata), and Monterey cypress (Cupressus macrocarpa) (McClintock, 2001). A long-term study in Point Reyes, California, showed that as Douglas fir (Pseudotsuga menziesii) forest regenerated between 1981 and 2001, densities of breeding Z. l. nuttalli decreased, supporting that this sub-species prefers open scrub habitat rather than dense forests for breeding and foraging (Chase et al., 2005). Despite these changes in habitat, Z. l. nuttalli persists in urban San Francisco, with higher apparent survival, though lower body condition, than rural birds (Phillips et al., 2018b). Further, minimum frequency and vocal performance vary with urban and rural soundscapes (Derryberry et al., 2016; Luther et al., 2016).
While anthropogenic noise and habitats select for higher pitched, narrow bandwidth songs that transmit better in noise (Slabbekoorn et al., 2007; Grabarczyk and Gill, 2019; Phillips et al., 2020), urban white-crowned sparrows still seem to use vocal performance to assess competitors in urban habitats (Phillips and Derryberry, 2018). Thus, given that performance functions in male-male competition even in noisy urban areas, the question remains whether vocal performance signals a male's quality and ability to obtain higher quality territories under varying levels of noise and urban habitats. Here, we test three hypotheses within the urban-rural matrix in the Bay Area of California using the Nuttall's white-crowned sparrow (Z. l. nuttalli). First, to explore the hypothesis that vocal performance is indicative of male quality, we examine whether physical characteristics associated with male condition are related to vocal performance, namely scaled mass index and fat score. We predict that higher performers are in better body condition and have more fat reserves compared to lower performers. Next, to assess whether noise affects vocal performance across the urban-rural matrix, we examine whether vocal performance varies with noise at three levels: dialect region, location, and territory. We expect that vocal performance decreases as noise levels increase across all dialect regions, locations, and at the territory level (Luther et al., 2016). We additionally examine noise, ecological characteristics and vocal performance between urban and rural locations, and expect that urban areas are noisier and urban birds have lower performance. Lastly, we test if territory quality varies with vocal performance scores using remotely sensed ecological variables to assess habitat quality. We expect that high performers will hold higher quality territories, with less human influenced characteristics like introduced trees, more native scrub for nesting, flat and open ground for foraging, higher greenness, and less drought stress.
In 2016, we identified 93 male white-crowned sparrow territories across urban San Francisco region (N = 60) and rural Point Reyes (N = 33). Six regions were sampled, where each region has a different song dialect. In urbanized areas, the Presidio region sings the San Francisco dialect (N = 28), the Fort Funston region sings a San Francisco and San Francisco-Lake Merced hybrid dialect (N = 19) (Luther and Baptista, 2010), and the Richmond region sings the Berkeley dialect (N = 13) (Baptista, 1975). In Point Reyes, the Limantour region sings the Limantour dialect (N = 14), the Abbott's Lagoon region sings both McClure and Drake dialects (N = 12), and the Commonweal sings the Clear dialect (N = 7) (Baker and Thompson, 1985). Ten locations were sampled and assessed as an individual category to account for spatial correlation, where San Francisco had five locations (Battery East, Battery West, Fort Scott, Lobos Dunes, and Baker Beach) and all other regions only had one location (Fort Funston, Richmond, Commonweal, Abbott's Lagoon, and Limantour). Males were caught with mist-nets and color-banded for individual identification then released at the same locations where they were caught. During capture, JNP measured tarsus using a SPI Polymid Dial Caliper and measured body mass using a Pesola Micro Spring Scale. Fat was scored based on the furcular hollow on a 0–5 scale, where 0 = no fat, 1 = trace amounts, 2 = 1/3 full, 3 = 2/3 full, 4 = even with breast muscle, and 5 = bursting (Kaiser, 1993; Pyle, 1997). One male was recorded but not captured, thus N = 92 for body condition analyses. To account for structural body size, we calculated a scaled mass index following Peig and Green (2009). Briefly, we calculated the standardized major axis (SMA) regression slope of mass vs. tarsus length from a larger historic sample (N = 239) of Nuttall's white crowned sparrows, and used the average tarsus length from the full population (Phillips et al., 2018b). Examination of SMA residual vs. fitted values and QQ plots indicated a linear relationship between tarsus and mass. All work was approved by Tulane University Institutional Animal Care and Use Committee (protocol 0427-R), USGS Bird banding Laboratory Permit (23900), California State Collecting Permit (6799), Golden Gate National Recreation Area Scientific Research and Collecting Permit (GOGA-00079), San Francisco Parks and Recreation Permit (032014), and Point Reyes National Park Scientific Research and Collecting Permit (PORE-0014).
Each male was recorded either immediately prior to capture or on a subsequent day as identified by individual color bands. To record songs, we used a Sony PBR 400 parabolic reflector with a Sennheiser ME62 omnidirectional microphone (Wedemark, Germany), foam windscreen, and Marantz PMD 661 recorder (Kanagawa, Japan). We analyzed approximately 1–10 songs per male (mean ± SD: 4.85 ± 3.90). One song should be sufficient as white-crowned sparrow songs in these populations are highly repeatable (Phillips and Derryberry, 2017b). Songs were recorded at a sampling rate of 44.1 kHz,.wav files, and resampled to 25 kHz for analysis. Using SIGNAL 5 (Beeman, 1998), songs were high-pass filtered at 1,500 Hz to remove low frequency background noise. We measured trill minimum and maximum frequencies at −36 dB from the peak amplitude frequency (time resolution: 10.2 ms, frequency resolution: 97.7 Hz, 256 pt transform). Bandwidth was calculated as the difference between maximum and minimum frequency, and trill rate was measured as the average number of notes per second (Hz). Wilson et al. (2014) suggested the use of the quantile regression method instead of the upper bound regression method when evaluating whether there was a significant trade-off between trill rate and bandwidth in a population of songs. In that review paper, they assessed the family-wide performance limit for Emberizidae and found that both methods (upper bound and quantile regression) recovered a significant trade-off between trill rate and bandwidth. To make sure our results are comparable to previous studies, we used the measurement of vocal deviation to approximate vocal performance, which was measured as the orthogonal distance from the upper-bound regression of 1,572 Passerellidae songs (formerly Emberizidae), y = 0.124x + 7.55 (Podos, 1997). We also conduct all analyses following the recommendations of Wilson et al. (2014) and include the details and results in Supplement 1.
We measured ambient noise levels male territories between 7:00 AM to 12:00 PM with a class 1 Larson-Davis 831 sound level meter, sampling 5 times per second, fast weighting (PCB Piezetronics). Following Brumm (2004), we recorded A-weighted equivalent continuous noise level (LAeq) for 1 min in each cardinal direction from the center of the territory for a total of 4 min, totaling 12,000 samples that are averaged to a single number for that territory. Units of LAeq (DB re 20 μPa) were used because A-weighting covers approximately the range of sound that birds can hear (Dooling et al., 2000) and accounts for fluctuations of noise during the recording. All noise recordings were taken in the absence of wind (3 > Beaufort Wind scale) and actively singing birds.
Remote sensors have proven beneficial in performing repeated standardized measurements over large areas testing for relationships between habitat selection and ecological gradients with high resolution and accuracy (Roughgarden et al., 1991; Smith et al., 2013). Sensors such as Landsat from satellites have been used for decades to estimate environmental variables on the ground and relate to species diversity (Pettorelli et al., 2014; Rocchini et al., 2016). Other sensors such as Light Detection and Ranging (LiDAR) have greatly improved the resolution of these data compared to satellites and are able to capture environmental information at resolutions less than one square meter (Farrell et al., 2013; Davies and Asner, 2014; He et al., 2015). These measures have also been used to designate habitats related to the occurrence of individual species (Clawges et al., 2008) and to understand how habitat heterogeneity relates to richness as a whole (Goetz et al., 2007). The information from these sensors provides great potential to investigate relationships between animals, plants, and environmental variables at both fine and large scales across landscapes.
We used two LiDAR datasets from projects around the San Francisco Bay Area to get accurate vegetation measures across all study sites. One dataset was from the Golden Gate LiDAR Project (GGLP), which was collected for the United States Geological Survey (USGS) in 2010 (OCM Partners, 2010). The second LiDAR dataset was collected by NOAA's Office for Coastal Management in the northern San Francisco Bay area in 2010 (Office for Coastal Management, 2010). LiDAR height returns were normalized to eliminate any elevational difference in points before calculating standard ground metrics using the “lidR” package in R version 3.3.2 (Roussel and David, 2018). In addition to LiDAR, we used imagery collected by Landsat 8 Operational Land Imager (OLI) during August of 2015 to calculate productivity indices. The datasets and specific images were selected to be as close as possible to the time of white-crowned sparrow observations while covering all sites.
A set of nine metrics was selected to determine habitat quality over a 50 m radius from the center of each White-crowned Sparrow territory, as the average territory size in white-crowned sparrows is approximately 1 hectare (Morton, 1992). The leaf area density (LAD) was selected to accurately measure productivity within 50 m of each activity center for two vertical strata above the ground: undergrowth (LAD1: 0.5–2 m) and canopy vegetation (LAD2: >2 m) (Kamoske et al., 2019). Other standard LiDAR and Landsat metrics were calculated for each area as listed in Table 1. The Normalized Difference Water Index (NDWI) was derived from the Landsat 8 OLI imagery, and was used as a surrogate for drought stress, where a high value indicates high water content within vegetation and soil (Gao, 1996). We calculated the mean NDWI values for each territory using the “raster” package in R version 3.3.2 (Robert, 2020). In addition to these habitat measurements, the average slope, aspect, and elevation were derived from the raw LiDAR dataset.
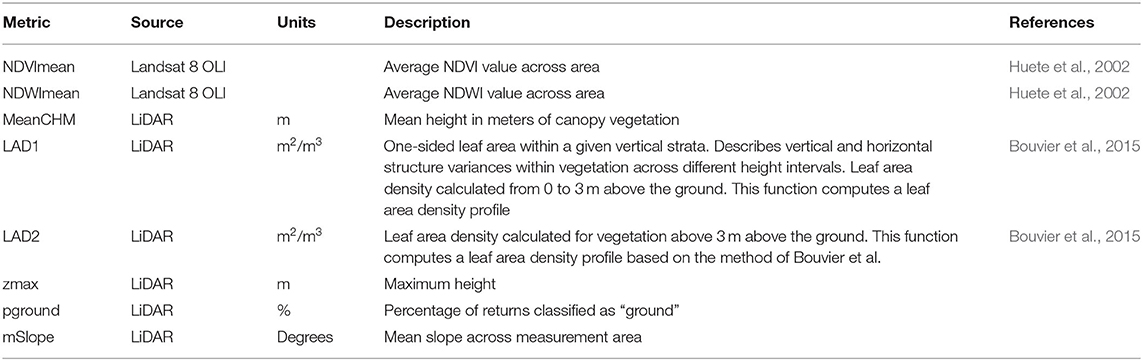
Table 1. Remote sensing variables.
To reduce collinearity of the ecological fixed effects, we examined spatial autocorrelation of 64 ecological variables extracted from remote sensing measures. Because of high correlation of many variables, we first reduced our independent factors to NDVI mean, NDWI mean, percent bare ground cover (pground), mean height of canopy vegetation (MeanCHM), mean slope, mean aspect, elevation, understory leaf area density 0–3 meters (LAD1), canopy leaf area density 3 meters and above (LAD2), overall territory height (Zmax). We further examined collinearity with variance inflation factors (VIF), which showed that all variables had a VIF < 6, so all nine variables were retained in the global model.
All analyses were conducted in R (R Core Team, 2020) using lme4 and AICmodavg to explore candidate models with AICc (Bates et al., 2015; Mazerolle, 2020). First, we examined whether body condition characteristics of individuals correlated to vocal deviation. We used a conservative approach and used our scaled mass index to estimate body condition. We then examined linear mixed models with fixed effects of Fat Score, Scaled Mass Index, and Type (urban/rural), with location as a random effect to account for any spatial autocorrelation (total nine candidate models). Next, we explore linear models to examine whether vocal deviation varied with noise levels (LAeq) at three levels—across dialects (Dialect Noise), across locations (Location Noise), and within territories (Territory Noise). We also explore candidate models for the additive and interactive effect of Type (urban/rural), which totaled eleven candidate models. Additionally, we used t-tests to compare average noise levels and territory characteristics between urban and rural birds. Lastly, to understand how territory quality correlates to vocal performance, we examined linear mixed effect models with deviation as the dependent variable, location as the random effect to take into account spatial variation, and ecological variables measured on territories, including Territory Noise level (LAeq) and Type (urban/rural), as fixed effects. We used the dredge function in MuMIn to explore all possible additive models, and examined candidate models within 2AICc (Arnold, 2010). For all model selection analyses, we used MuMIN to examine the model average of models within 2AICc when necessary (Bartoń, 2020) and report 95% CI intervals. If Type was a significant factor in the model average, we also conducted model selection and model averaging in the urban subset alone and the rural subset alone to clarify directionality of effects. If the random factor Location was equivalent to zero, we instead examined linear models (Bates et al., 2018).
We examined ten candidate models with Location as a random effect, where the top three models were within 2 AICc of each other (Table 2A). The highest ranked model included fat score interacting with Type and Location as a random effect (ER = 2,985, wi = 0.29, Location variance = 7.52, SD = 2.74, Table 2A), followed by a model with Fat + Scaled Mass Index + Type (ER = 2,800, wi = 0.28, Location variance = 15.54, SD = 3.94, Table 2A), and Scaled Mass Index*Type (ER = 2,700, wi = 0.27, Location Variance = 7.48, SD = 2.74). The conditional model average supported strong effects of Type on vocal performance, rather than Fat or Scaled Mass Index (Table 2B). Our analyses with quantile regression performance values also supported the effect of Type over body condition measures (Supplement 1, Table S1). Rural birds had higher performance than urban birds (deviation mean ± SD. Urban: 30.54 ± 4.99, Rural: 24.56 ± 3.16).
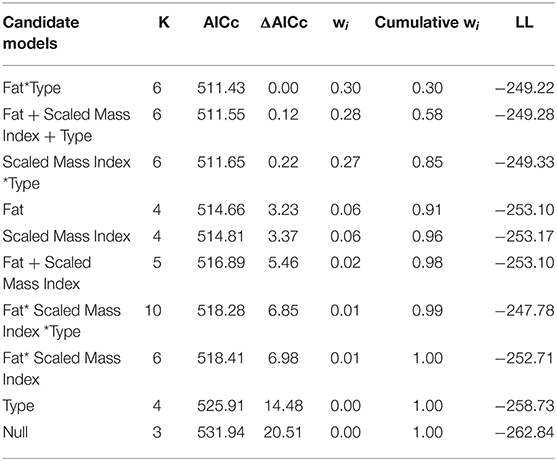
Table 2A. Model selection table for ten candidate models for Vocal Deviation~Body Condition.
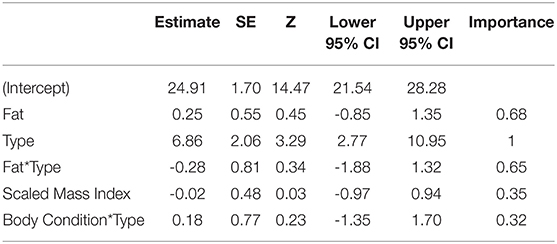
Table 2B. Conditional model average of the top three models within 2AICc for body condition models.
Noise had a negative effect on performance, and was significant at three levels (Dialect Region, Location, and Territory; Figure 1). However, only two candidate models were within 2 AICc, where Dialect Region noise had the most important influence on vocal deviation (β = 0.77, SE = 0.10, F1, 91 = 61.32, 95% CI = 0.58–0.97, Table 3A, Figure 1A), followed by Dialect Region noise + Type (Dialect Region noise: β = 1.06 SE = 0.25, 95% CI = 0.55–1.57; Type: β = −2.82, SE = 2.29, 95% CI = −7.38–1.74; F2, 90 = 31.58, Table 3A, model average Table 3B). Males in quieter locations have higher performance (β = 0.74, SE = 0.09, F1, 91 = 60.97, 95% CI = 0.55–0.93, Figure 1B). Across territories, males that produce higher performance songs defend territories with lower noise levels (β = 0.37, SE = 0.07, F1, 91 = 25.05, 95% CI = 0.22–0.51, Figure 1C). As above, rural birds produced songs with higher vocal performance than did urban birds (β = 6.00, SE = 0.96, t = 6.24, F1, 91 = 38.9, 95% CI = 4.08–7.89). Model selection and model averaging from quantile regression also supported effects of Dialect Region noise, Location noise, and Territory noise on vocal performance (Supplement 1).
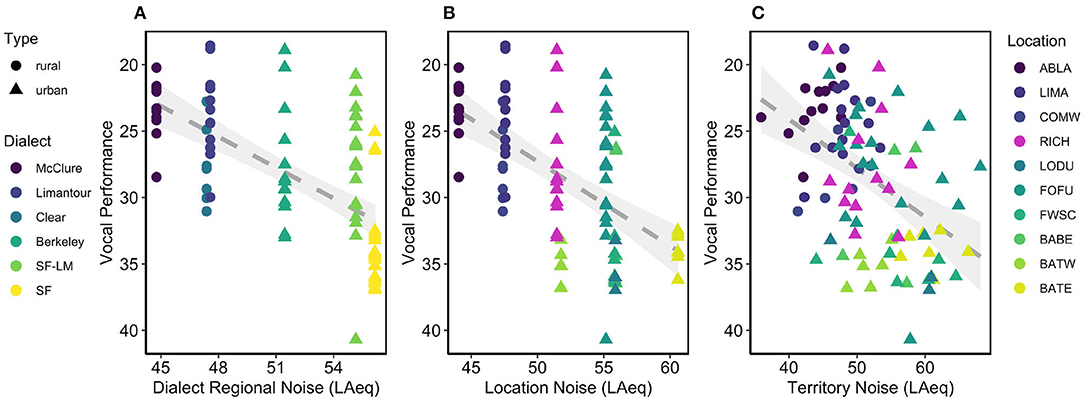
Figure 1. (A) Urban regions have higher noise and lower performance songs. (B) Urban locations are also noisier and have lower performance songs. (C) High performers have quieter territories. Line of best fit and 95% standard error for each model shown in gray.
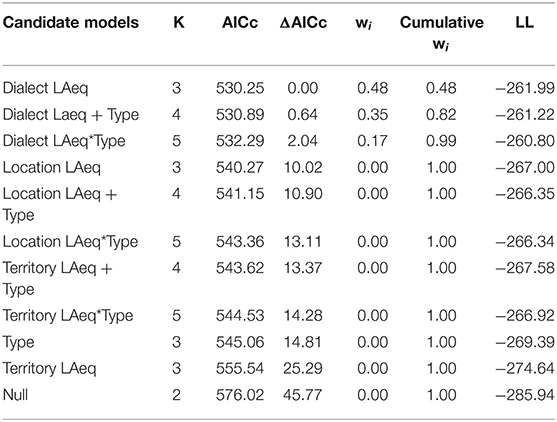
Table 3A. Nine Candidate models for the effects of noise (Dialect Regional Noise, Location Noise, and Territory Noise) and habitat type on vocal deviation.
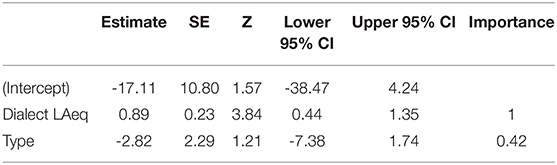
Table 3B. Conditional model average of the top three models within 2AICc.
Urban territories had higher noise levels than rural territories (Table 4). Overall, urban territories had higher leaf area densities in the understory (LAD1) and canopy (LAD2). Canopies of urban territories were taller at maximum height (zmax) and on average (MeanCHM). Rural areas had more open ground (pground) with less sloped terrain (mSlope), more greenness (NDVImean) and more drought stress (NDWImean) (Table 2).
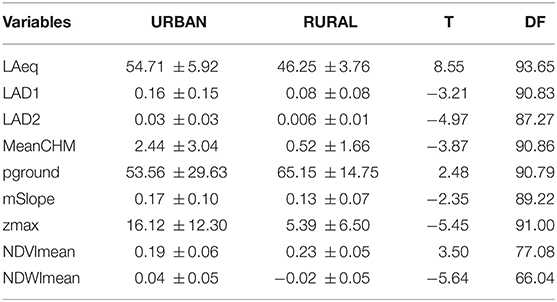
Table 4. Mean and SD for ecological variables for urban and rural samples with results from Welch's t-tests for each variable.
We examined 13 candidate models that were within 2AIC, all of which include Type as a fixed effect, and a null model for a total of 14 models. The highest ranked model included leaf area density for canopy above 3 m (LAD2) and mean slope (mSlope), where both variables had a positive correlation with vocal deviation, meaning lower performers had territories with higher canopy leaf density and steeper slopes (ER = 1,300, Location variance = 3.86, SD = 2.00, Table 5, Figures 2A,B). The next ranked model included leaf area density below 3 m (LAD1, Figure 2C), mean canopy height (MeanCHM, Figure 2D), and maximum height (zmax) (ER = 1,200, Location variance = 4.93, SD = 2.22, Table 5). Model averaged estimates and importance values supported the top model (Table 6, Figure 3), as well as showed significant effects of leaf area density below 3 meters (LAD1) and mean canopy height (Figure 3). Low performers were on territories with steeper slopes, denser and higher average canopy and denser understories (Figures 2, 3). Models estimating performance from quantile regression show similar patterns, where low performers were significantly on territories with steeper slopes and less dense canopies and understories (Supplement 2).
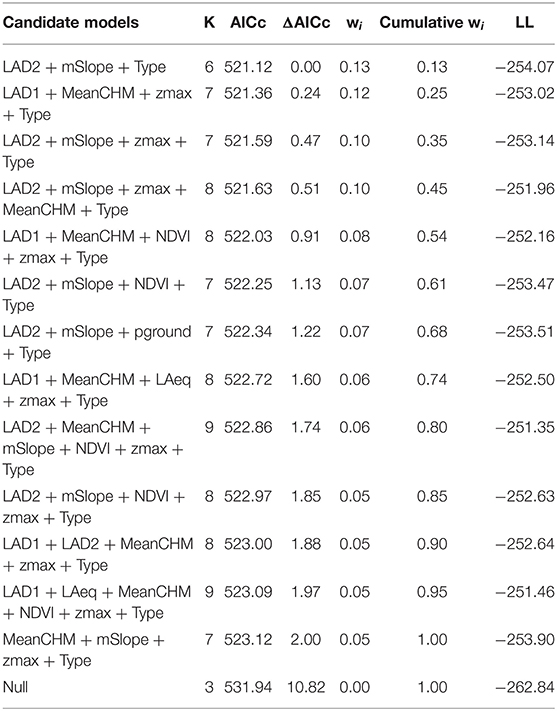
Table 5. Candidate Models for Vocal Deviation ~ Habitat Quality.
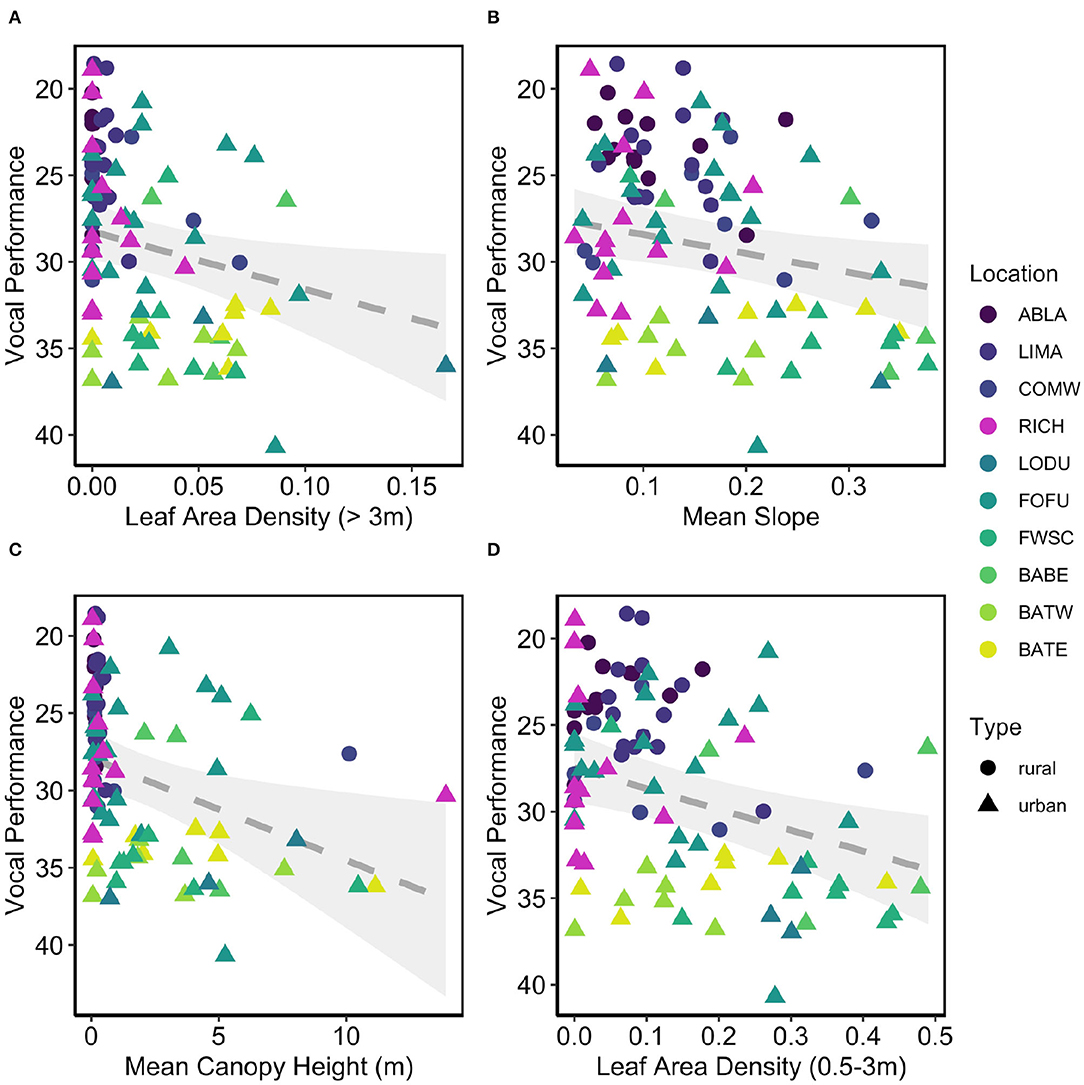
Figure 2. Low performance males (high deviation) have territories with (A) denser canopies, (B) steeper slopes, (C) higher canopies, and (D) denser understories. Gray dotted line and band denotes line of best fit with standard error (A&B candidate model 1, C&D candidate model 6).
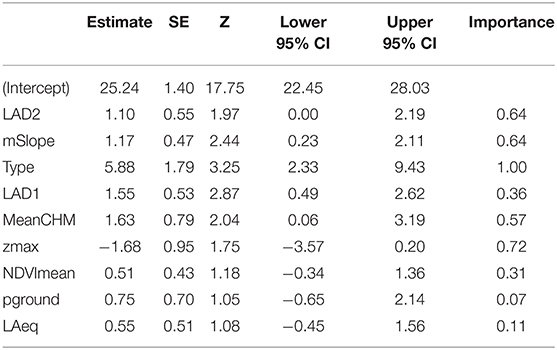
Table 6. Model Averaged Coefficients for Vocal Deviation~Ecological Variables.
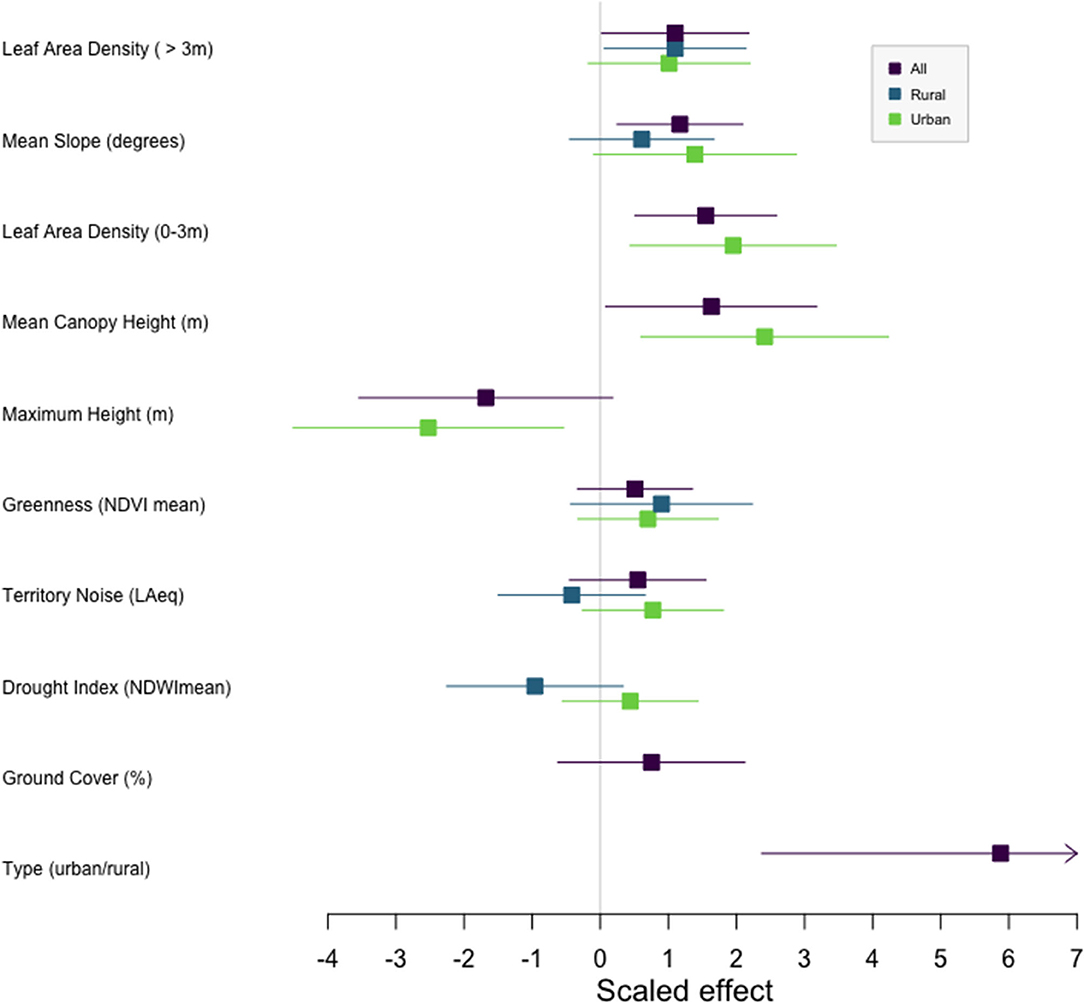
Figure 3. Effects plot for the model average of 13 candidate models across urban and rural locations (All), 10 candidate models within urban areas (Urban), and 7 candidate models within rural areas (Rural). Variables to the left of zero have a negative effect on vocal deviation, and are associated with higher performance songs, while variables to the right have a positive effect on vocal deviation, indicating an association with lower performance songs. Boxes denote mean and lines denote 95% confidence intervals.
Because Type had the strongest effect on deviation, we also ran model selection within habitat types. Within urban habitats, 10 models were within 2AICc (Supplement 2A). Higher performers were on territories with shorter average canopies but taller maximum heights, which includes man-made structures (Supplement 2B, Figure 3). Additionally, urban high performers had less dense understories (Supplement 2B, Figure 3). Within rural habitats, higher performers were on territories with significantly less dense canopies (Supplement 3, Figure 3). Analyses for performance with quantile regression show similar patterns, with urban high performers on territories with less dense understories and lower canopy heights (Supplement 1). Rural birds tended to have less dense canopies (Supplement 1).
Vocal performance of male white-crowned sparrows correlates with background noise levels and landscape features of the breeding territories they defend, but not with body condition. Consistent with our predictions, we found that high performance males hold flatter territories with lower noise levels and short native scrub habitat. Conversely, low performance males hold territories with higher noise levels and dense, taller vegetation, which is a product of human settlement in the coastal regions of the Bay Area (McClintock, 2001). Together, our results suggest that vocal performance may signal the ability to obtain and defend territories that are in favored habitat (i.e., low laying scrub and grassland), which should improve their chance of survival and successful breeding. To our knowledge, only one study shows a similar link between song characteristics and habitat quality (Manica et al., 2014). Since high performance males are on quieter territories, with less dense understories and less canopy cover, selection pressure on song transmission should also be released, reducing constraints on vocal performance. These findings link together fundamental aspects of sexual selection in that habitat quality and the quality of sexually selected signals appear to be associated: males with the highest performing songs are defending territories of the highest quality.
We hypothesized that that high performers may be more competitive at holding territories with better food sources, and therefore be in better body condition. However, we did not find evidence for fat score or scaled mass index correlating with vocal performance. This could be due to sample size, lack of heterogeneity in food availability across our sites, or possibly there are other metrics of body condition that are important that we did not consider in our methods. The sampling year (2016) was a wet year compared to the previous 4 years, which may have erased any large disparities in food or water availability between urban and rural locations. Cities typically present a greater patchwork of available resources, such as bird feeders, green space, and large tracts of cement (Robb et al., 2008), which may create “boom or bust” dynamics, but a wet winter may nullify these dynamics across both urban and rural areas. The 2015-2016 wet season produced 23.21 inches of rain in San Francisco, which is within the average range from 1849 to 2019 (21.86 ± 7.5 inches per season, https://www.ggweather.com/sf/monthly.html). Therefore, our sampling year would have had high productivity for birds, likely due to increased plant and insect food resources related to rainfall (Desante and Geupel, 1987; Dybala et al., 2013), potentially resulting in similar body condition across habitats. Previous studies found body size to vary with urbanization level (Meillère et al., 2015), but whether song performance is related to body condition across urban ecosystems remains to be seen. Future research within long-term research areas will allow for larger sample sizes and the ability to account for yearly variation and would greatly benefit our understanding of the relationship between song and male quality.
Anthropogenic noise interferes with communication (Slabbekoorn, 2013), foraging vigilance (Ware et al., 2015), and reproductive fitness in many species (Injaian et al., 2018; Kleist et al., 2018; Gurule-Small and Tinghitella, 2019). Therefore, there may be a fitness benefit to avoiding noise, such that individuals are likely to be more successful in attracting mates, feeding chicks, and fledging offspring when on territories with lower noise levels. Our study system has consistently shown that urban San Francisco has higher noise levels, particularly at low frequencies as compared to nearby rural Point Reyes (Lee and MacDonald, 2011, 2013; Derryberry et al., 2016; Phillips and Derryberry, 2017b; Phillips et al., 2018b), and that song varies with ambient and background noise levels (Derryberry et al., 2016), including vocal performance characteristics (Luther et al., 2016). Our additional data are in line with previous findings that noise levels are related to vocal performance. Territory noise levels were predictive of vocal performance across dialects, consistent with our previous work within the San Francisco dialect (Luther et al., 2016).
High performers hold flatter territories with more scrub and less trees. These characteristics are beneficial to the scrub adapted Nuttall's white-crowned sparrow (Chase et al., 2005). Flatter ground provides foraging habitat and open space to observe predators, while low shrubs like coyote brush (Baccharus pilularis), lupine spp. (Lupinus spp.), California blackberry (Rubus ursinus), and poison oak (Toxicodendron diversilobum) provide nesting habitat and places to hide from predators. Additionally, many of the areas in San Francisco that retain native scrub are on cliffsides where human development is not possible, so competition for flatter territories is likely high. In this study and in previous work (Phillips et al., 2018a), we have observed that urban parks within the Bay Area are landscaped with taller, non-native shrubs and trees. Our remotely sensed leaf area density measures and mean canopy height measure suggest that low performers are on territories with taller trees and thicker vegetation, which is not preferred habitat for white-crowned sparrows (Chase et al., 2005). This trend is especially notable within the urban habitat. Previously in this system, percent tree cover as measured from Google Earth imagery indicated that more trees are found on urban territories (Phillips et al., 2018a). Together, it seems likely that high performance males are more competitive and thus better able to acquire territories with species-preferred characteristics, such as low laying native scrub with adjacent open space.
High performance males may be on territories with species-preferred scrub habitat because they are more competitive or because their signal transmits better on these more open territories, or both. Taller, denser vegetation results in more reverberation and degradation of trilled signals which over time selects for reduced trill rates and narrow bandwidth (Morton, 1975; Derryberry, 2009; Phillips et al., 2020), whereas trilled vocalizations are less constrained on more open habitats and can evolve to faster rates and wider bandwidth (Wiley, 1991), at least until a physical performance constraint is met (Podos, 1997). Therefore, males with high performance songs may select territories based on transmission properties, or just be more successful during conflict with low performance competitors because their song transmits better and/or indicates higher competitive abilities. Multiple recaptures and recordings of males throughout the breeding season, starting at territory establishment, can inform whether high performance males are more competitive and this is the driver of territory holding ability, or if song transmission alone ultimately leads to higher success at obtaining and defending territories with lower noise levels in species-preferred habitat.
Our results are the first to show a relationship between vocal performance, a physically limited song trait, and territory quality. This study provides a first step toward understanding how urban landscapes and soundscapes affect song evolution and thus reproduction and survival. Future research linking reproductive outcomes across a variety of performers and landscapes will provide important insights into how anthropogenic pollution may affect not only animal behavior but also fitness and the longer-term stability of populations.
All datasets generated for this study are included in the article/supplementary material.
The animal study was reviewed and approved by Tulane University Institutional Animal Care and Use Committee (protocol 0427-R), USGS Bird banding Laboratory Permit (23900), California State Collecting Permit (6799), Golden Gate National Recreation Area Scientific Research and Collecting Permit (GOGA-00079), San Francisco Parks and Recreation Permit (032014), and Point Reyes National Park Scientific Research and Collecting Permit (PORE-0014).
JP, ED, and DL conceived and designed the study. JP collected field data and WC extracted LiDAR and Landsat variables. JP ran the statistics, wrote the manuscript with input, and final approval from WC, ED, and DL. All authors contributed to the article and approved the submitted version.
This work was funded by NSF IOS 1354763, 1354756, and 1827290 and NSF Emerging Frontiers 1823498. JP was supported by a Tulane Gunning Award, Tulane One-Term Dissertation Award, Wilson Ornithological Society Grant, American Ornithologists Union Van Tyne Award, and NSF Postdoctoral Research Fellowship in Biology #1812280.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We are grateful to Mae Berlow and Leanne Norden who assisted with fieldwork, Julia Paschal, Adrianne Seiden, Ted Hagens, Nicole Moody, and Kyu-Min Huh who assisted with song analyses, Point Blue Palomarin Field Station staff and interns, Bill Merkle and Michael Chasse at Golden Gate National Recreation Area, Ben Becker at Point Reyes National Seashore, Doug Bell at East Bay Regional Parks, and Lisa Wayne at San Francisco Parks and Recreation.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.587120/full#supplementary-material
Andersson, M. B. (1994). Sexual Selection. Princeton, NJ: Princeton University Press. doi: 10.1515/9780691207278
Arnold, T. W. (2010). Uninformative parameters and model selection using Akaike's information criterion. J. Wildl. Manage. 74, 1175–1178. doi: 10.1111/j.1937-2817.2010.tb01236.x
Aronson, M. F. J., Nilon, C. H., Lepczyk, C. A., Parker, T. S., Warren, P. S., Cilliers, S. S., et al. (2016). Hierarchical filters determine community assembly of urban species pools. Ecology 97, 2952–2963. doi: 10.1002/ecy.1535
Aronson, M. F. J., Sorte, F. A., La Nilon, C. H., Katti, M., Goddard, M. A., Lepczyk, C. A., et al. (2014). A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. B 281:20133330. doi: 10.1098/rspb.2013.3330
Baker, M. C., and Thompson, D. B. (1985). Song dialects of white-crowned sparrows: historical processes inferred from patterns of geographic variation. Condor 87, 127–141. doi: 10.2307/1367144
Ballentine, B. (2009). The ability to perform physically challenging songs predicts age and size in male swamp sparrows, Melospiza georgiana. Anim. Behav. 77, 973–978. doi: 10.1016/j.anbehav.2008.12.027
Ballentine, B., Hyman, J., and Nowicki, S. (2004). Vocal performance influences female response to male bird song: an experimental test. Behav. Ecol. 15, 163–168. doi: 10.1093/beheco/arg090
Baptista, L. F. (1975). Song dialects and demes in sedentary populations of the white-crowned sparrow (Zonotrichia leucophrys nuttalli). Univ. Calif. Publ. Zool. 105, 1–52.
Bartoń, K. (2020). MuMIn: Multi-Model Inference. R package version 1.43.17. Available online at: https://CRAN.R-project.org/package=MuMIn
Bates, D., Maechler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Bates, D., Kliegl, R., Vasishth, S., and Baayen, H. (2018). Parsimonious Mixed Models. arXiv 1506.04967. Available online at: http://arxiv.org/abs/1506.04967 (accessed November 30, 2020).
Beeman, K. (1998). “Digital signal analysis, editing, and synthesis,” in Animal Acoustic Communication: Sound Analysis and Research Methods, eds S. L. Hopp, M. J. Owren, and C. S. Evans (Berlin: Springer-Verlag).
Blair, R. B. (1996). Land use and avian species diversity along an urban gradient. Ecol. Soc. 6, 506–519. doi: 10.2307/2269387
Bouvier, M., Durrieu, S., Fournier, R., and Renaud, J. (2015). Generalizing predictive models of forest inventory attributes using an area-based approach with airborne las data. Remote Sens. Environ. 156, 322–334. doi: 10.1016/j.rse.2014.10.004
Brumm, H. (2004). The impact of environmental noise on song amplitude in a territorial bird. J. Anim. Ecol. 73, 434–440. doi: 10.1111/j.0021-8790.2004.00814.x
Caro, S. P., Sewall, K. B., Salvante, K. G., and Sockman, K. W. (2010). Female Lincoln's sparrows modulate their behavior in response to variation in male song quality. Behav. Ecol. 21, 562–569. doi: 10.1093/beheco/arq022
Cartwright, L. A., Taylor, D. R., Wilson, D. R., and Chow-Fraser, P. (2014). Urban noise affects song structure and daily patterns of song production in Red-winged Blackbirds (Agelaius phoeniceus). Urban Ecosyst. 17, 561–572. doi: 10.1007/s11252-013-0318-z
Catchpole, C. K., and Slater, P. J. B. (2008). Bird Song: Biological Themes and Variations. 2nd Edn. New York, NY: Cambridge University Press. doi: 10.1017/CBO9780511754791
Chase, M. K., Holmes, A. L., Gardali, T., Ballard, G., Geupel, G. R., and Nur, N. (2005). Two decades of change in a coastal scrub community : songbird responses to plant succession 1. USDA For. Serv. Gen. Tech. Rep. 613–616. Available online at: https://www.fs.fed.us/psw/publications/documents/psw_gtr191/psw_gtr191_0613-0616_chase.pdf
Clarkson, C. E. (2007). Food supplementation, territory establishment, and song in the prothonotary warbler. Wilson J. Ornithol. 119, 342–349. doi: 10.1676/04-073.1
Clawges, R., Vierling, K., Vierling, L., and Rowell, E. (2008). The use of airborne lidar to assess avian species diversity, density, and occurrence in a pine/aspen forest. Remote Sens. Environ. 112, 2064–2073. doi: 10.1016/j.rse.2007.08.023
Clink, D. J., Charif, R. A., Crofoot, M. C., and Marshall, A. J. (2018). Evidence for vocal performance constraints in a female nonhuman primate. Anim. Behav. 141, 85–94. doi: 10.1016/j.anbehav.2018.05.002
Cramer, E. R. A., Hall, M. L., de Kort, S. R., Lovette, I. J., and Vehrencamp, S. L. (2011). Infrequent extra-pair paternity in the banded wren, a synchronously breeding tropical passerine. Condor 113, 637–645. doi: 10.1525/cond.2011.100233
Cramer, E. R. A., and Price, J. J. (2007). Red-winged blackbirds Ageliaus phoeniceus respond differently to song types with different performance levels. J. Avian Biol. 38, 122–127. doi: 10.1111/j.2006.0908-8857.03839.x
Croci, S., Butet, A., and Clergeau, P. (2008). Does urbanization filter birds on the basis of their biological traits? Condor 110, 223–240. doi: 10.1525/cond.2008.8409
Davies, A. B., and Asner, G. P. (2014). Advances in animal ecology from 3D-LiDAR ecosystem mapping. Trends Ecol. Evol. 29, 681–691. doi: 10.1016/j.tree.2014.10.005
de Kort, S. R., Eldermire, E. R. B., Cramer, E. R., and Vehrencamp, S. L. (2009). The deterrent effect of bird song in territory defense. Behav. Ecol. 20, 200–206. doi: 10.1093/beheco/arn135
Derryberry, E. P. (2009). Ecology shapes birdsong evolution: variation in morphology and habitat explains variation in white-crowned sparrow song. Am. Nat. 174, 24–33. doi: 10.1086/599298
Derryberry, E. P., Danner, R. M., Danner, J. E., Derryberry, G. E., Phillips, J. N., Lipshutz, S. E., et al. (2016). Patterns of song across natural and anthropogenic soundscapes suggest that white-crowned sparrows minimize acoustic masking and maximize signal content. PLoS ONE 11, e0154456. doi: 10.1371/journal.pone.0154456
Desante, D. F., and Geupel, G. R. (1987). Landbird productivity in central coastal California: the relationship to annual rainfall, and a reproductive failure in 1986. Condor 89, 636–653. doi: 10.2307/1368653
Desrochers, A. (1989). Sex, dominance, and microhabitat use in wintering black-capped chickadees: a field experiment. Ecology 70, 636–645. doi: 10.2307/1940215
Dewolfe, B. B., Baptista, L., and Petrinovich, L. (1989). Song development and territory establishment in Nuttall's white-crowned sparrows. Condor 91, 397–407. doi: 10.2307/1368318
Dooling, R. J., Lohr, B., and Dent, M. L. (2000). “Hearing in birds and reptiles,” in Comparative Hearing: Birds and Reptiles, eds R. J. Dooling, A. N. Popper, and R. R. Fay (New York, NY: Springer-Verlag), 308–359. doi: 10.1007/978-1-4612-1182-2_7
Dowling, J. L., Luther, D. A., and Marra, P. P. (2011). Comparative effects of urban development and anthropogenic noise on bird songs. Behav. Ecol. 23, 201–209. doi: 10.1093/beheco/arr176
Drăgănoiu, T. I., Nagle, L., and Kreutzer, M. (2002). Directional female preference for an exaggerated male trait in canary (Serinus canaria) song. Proc. R. Soc. B 269, 2525–2531. doi: 10.1098/rspb.2002.2192
DuBois, A. L., Nowicki, S., and Searcy, W. A. (2009). Swamp sparrows modulate vocal performance in an aggressive context. Biol. Lett. 5, 163–165. doi: 10.1098/rsbl.2008.0626
Dubois, A. L., Nowicki, S., and Searcy, W. A. (2011). Discrimination of vocal performance by male swamp sparrows. Behav. Ecol. Sociobiol. 65, 717–726. doi: 10.1007/s00265-010-1073-2
Dybala, K. E., Eadie, J. M., Gardali, T., Seavy, N. E., and Herzog, M. P. (2013). Projecting demographic responses to climate change: adult and juvenile survival respond differently to direct and indirect effects of weather in a passerine population. Glob. Chang. Biol. 19, 2688–2697. doi: 10.1111/gcb.12228
Ekman, J. (1987). Exposure and time use in willow tit flocks: the cost of subordination. Anim. Behav. 35, 445–452. doi: 10.1016/S0003-3472(87)80269-5
Farrell, S. L., Collier, B. A., Skow, K. L., Long, A. M., Campomizzi, A. J., Morrison, M. L., et al. (2013). Using LiDAR-derived vegetation metrics for high-resolution, species distribution models for conservation planning. Ecosphere 4, 1–18. doi: 10.1890/ES12-000352.1
Gao, B. (1996). NDWI—A normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens. Environ. 58, 257–266. doi: 10.1016/S0034-4257(96)00067-3
Goetz, S., Steinberg, D., Dubayah, R., and Blair, B. (2007). Laser remote sensing of canopy habitat heterogeneity as a predictor of bird species richness in an eastern temperate forest, USA. Remote Sens. Environ. 108, 254–263. doi: 10.1016/j.rse.2006.11.016
Grabarczyk, E. E., and Gill, S. A. (2019). Anthropogenic noise masking diminishes house wren (Troglodytes aedon) song transmission in urban natural areas. Bioacoustics 29, 518–532. doi: 10.1080/09524622.2019.1621209
Grava, T., Grava, A., and Otter, K. A. (2012). Vocal performance varies with habitat quality in black-capped chickadees (Poecile atricapillus). Behaviour 149, 35–50. doi: 10.1163/156853912X625854
Grava, T., Grava, A., and Otter, K. A. (2013). Habitat-induced changes in song consistency affect perception of social status in male chickadees. Behav. Ecol. Sociobiol. 67, 1699–1707. doi: 10.1007/s00265-013-1580-z
Gurule-Small, G. A., and Tinghitella, R. M. (2019). Life history consequences of developing in anthropogenic noise. Glob. Chang. Biol. 25, 1957–1966. doi: 10.1111/gcb.14610
He, K. S., Bradley, B. A., Cord, A. F., Rocchini, D., Tuanmu, M.-N., Schmidtlein, S., et al. (2015). Will remote sensing shape the next generation of species distribution models? Remote Sens. Ecol. Conserv. 1, 4–18. doi: 10.1002/rse2.7
Huete, A., Didan, K., Miura, T., Rodriguez, E. P., Gao, X., and Ferreira, L. G. (2002). Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 83, 195–213. doi: 10.1016/S0034-4257(02)00096-2
Illes, A. E., Hall, M. L., and Vehrencamp, S. L. (2006). Vocal performance influences male receiver response in the banded wren. Proc. R. Soc. B 273, 1907–1912. doi: 10.1098/rspb.2006.3535
Injaian, A. S., Poon, L. Y., and Patricelli, G. L. (2018). Effects of experimental anthropogenic noise on avian settlement patterns and reproductive success. Behav. Ecol. 29, 1181–1189. doi: 10.1093/beheco/ary097
Kaiser, A. (1993). A New multi-category classification of subcutaneous fat deposits of songbirds (Una Nueva Clasificación, con Multi-categorías, para los Depósitos de Grasa en Aves Canoras). J. F. Ornithol. 64, 246–255.
Kaiser, S. A., Kershner, E. L., and Garcelon, D. K. (2009). “The influence of nest substrate and nest site characteristics on the risk of San Clemente sage sparrow nest failure,” in Proceedings of301the 7th California Islands Symposium (Arcata, CA), 301–313.
Kamoske, A. G., Dahlin, K. M., Stark, S. C., and Serbin, S. P. (2019). Leaf area density from airborne LiDAR: comparing sensors and resolutions in a temperate broadleaf forest ecosystem. For. Ecol. Manage. 433, 364–375. doi: 10.1016/j.foreco.2018.11.017
Kleist, N. J., Guralnick, R., Cruz, A., Lowry, C., and Francis, C. (2018). Chronic anthropogenic noise disrupts glucocorticoid signaling and has multiple effects on fitness in an avian community. Proc. Natl. Acad. Sci. 115, E648–E657. doi: 10.1073/pnas.1709200115
LaZerte, S. E., Slabbekoorn, H., Otter, K. A., Francis, C., Ortega, C., Cruz, A., et al. (2016). Learning to cope: vocal adjustment to urban noise is correlated with prior experience in black-capped chickadees. Proc. R. Soc. B 283, e27052–e27052. doi: 10.1098/rspb.2016.1058
Lee, C., and MacDonald, J. (2011). Baseline ambient sound levels in Point Reyes National Seashore. (Los Angeles, CA: Department of Transportation, Western-Pacific Region).
Lee, C., and MacDonald, J. (2013). Golden Gate National Recreation Area: Acoustical Monitoring 2007/2008. (Fort Collins, CO: National Park service).
Liker, A., Papp, Z., Bókony, V., and Lendvai, Á. Z. (2008). Lean birds in the city: body size and condition of house sparrows along the urbanization gradient. J. Anim. Ecol. 77, 789–795. doi: 10.1111/j.1365-2656.2008.01402.x
Luther, D., and Baptista, L. (2010). Urban noise and the cultural evolution of bird songs. Proc. R. Soc. B 277, 469–473. doi: 10.1098/rspb.2009.1571
Luther, D. A., Phillips, J., and Derryberry, E. P. (2016). Not so sexy in the city: urban birds adjust songs to noise but compromise vocal performance. Behav. Ecol. 27, 332–340. doi: 10.1093/beheco/arv162
Manica, L. T., Maia, R., Dias, A., Podos, J., and Macedo, R. H. (2014). Vocal output predicts territory quality in a neotropical songbird. Behav. Processes 109, 21–26. doi: 10.1016/j.beproc.2014.07.004
Marzluff, J. M. (2017). A decadal review of urban ornithology and a prospectus for the future. Ibis 159, 1–13. doi: 10.1111/ibi.12430
Mazerolle, M. J. (2020). AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c). R package version 2.3-0. Available online at: https://cran.r-project.org/package=AICcmodavg
McClintock, E. (2001). The Trees of Golden Gate Park and San Francisco. ed. R. G. J. Turner Berkeley (California, CA: Heydey Books).
Meillère, A., Brischoux, F., Parenteau, C., and Angelier, F. (2015). Influence of urbanization on body size, condition, and physiology in an urban exploiter: a multi-component approach. PLoS ONE 10:13568. doi: 10.1371/journal.pone.0135685
Morton, E. S. (1975). Ecological sources of selection on avian sounds. Am. Nat. 109, 17–34. doi: 10.1086/282971
Morton, M. L. (1992). Effects of sex and birth date on premigration biology, migration schedules, return rates and natal dispersal in the mountain white-crowned sparrow. Condor 94, 117–133. doi: 10.2307/1368802
Moseley, D. L., Lahti, D. C., and Podos, J. (2013). Responses to song playback vary with the vocal performance of both signal senders and receivers. Proc. R. Soc. B 280, 1–9. doi: 10.1098/rspb.2013.1401
Narango, D. L., and Rodewald, A. D. (2016). Urban-associated drivers of song variation along a rural-urban gradient. Behav. Ecol. 27, 608–616. doi: 10.1093/beheco/arv197
Narango, D. L., and Rodewald, A. D. (2018). Signal information of bird song changes in human-dominated landscapes. Urban Ecosyst. 21, 41–50. doi: 10.1007/s11252-017-0698-6
Ota, N., and Soma, M. (2014). Age-dependent song changes in a closed-ended vocal learner: elevation of song performance after song crystallization. J. Avian Biol. 45, 566–573. doi: 10.1111/jav.00383
Pärt, T., Arlt, D., Doligez, B., Low, M., and Qvarnström, A. (2011). Prospectors combine social and environmental information to improve habitat selection and breeding success in the subsequent year. J. Anim. Ecol. 80, 1227–1235. doi: 10.1111/j.1365-2656.2011.01854.x
Pasch, B., George, A. S., Campbell, P., and Phelps, S. M. (2011). Androgen-dependent male vocal performance influences female preference in Neotropical singing mice. Anim. Behav. 82, 177–183. doi: 10.1016/j.anbehav.2011.04.018
Peig, J., and Green, A. J. (2009). New perspectives for estimating body condition from mass/length data: The scaled mass index as an alternative method. Oikos 118, 1883–1891. doi: 10.1111/j.1600-0706.2009.17643.x
Petrinovich, L., and Patterson, T. L. (1982). The white-crowned sparrow: stability, recruitment, and population structure in the Nuttall subspecies (1975-1980). Auk 99, 1–14. doi: 10.2307/4086016
Pettorelli, N., Laurance, W. F., O'brien, T. G., Wegmann, M., Nagendra, H., and Turner, W. (2014). Satellite remote sensing for applied ecologists: opportunities and challenges. J. Appl. Ecol. 51, 839–848. doi: 10.1111/1365-2664.12261
Phillips, J. N., Berlow, M., and Derryberry, E. P. (2018a). The effects of landscape urbanization on the gut microbiome: an exploration into the gut of urban and rural white-crowned sparrows. Front. Ecol. Evol. 6:148. doi: 10.3389/fevo.2018.00148
Phillips, J. N., and Derryberry, E. P. (2017a). Equivalent effects of bandwidth and trill rate: support for a performance constraint as a competitive signal. Anim. Behav. 132, 209–215. doi: 10.1016/j.anbehav.2017.08.012
Phillips, J. N., and Derryberry, E. P. (2017b). Vocal performance is a salient signal for male–male competition in white-crowned sparrows. Auk 134, 564–574. doi: 10.1642/AUK-17-2.1
Phillips, J. N., and Derryberry, E. P. (2018). Urban sparrows respond to a sexually selected trait with increased aggression in noise. Sci. Rep. 8:7505. doi: 10.1038/s41598-018-25834-6
Phillips, J. N., Gentry, K., Luther, D. A., and Derryberry, E. (2018b). Surviving in the City: higher apparent survivial for urban birds but worse condition on noisy territories. Ecosphere 9:e02440. doi: 10.1002/ecs2.2440
Phillips, J. N., and Katti, M. (2020). Anthropogenic noise affects winter song structure of a long distance migrant, Gambel's White-crowned Sparrow. J. Urban Ecol. 6, 1–9. doi: 10.1093/jue/juaa003
Phillips, J. N., Rochefort, C., Lipshutz, S., Derryberry, G. E., Luther, D., and Derryberry, E. P. (2020). Increased attenuation and reverberation are associated with lower maximum frequencies and narrow bandwidth of bird songs in cities. J. Ornithol. 161, 593–608. doi: 10.1007/s10336-020-01751-2
Pidgeon, A. M., Radeloff, V. C., and Mathews, N. E. (2006). Contrasting measures of fitness to classify habitat quality for the black-throated sparrow (Amphispiza bilineata). Biol. Conserv. 132, 199–210. doi: 10.1016/j.biocon.2006.03.024
Podos, J. (1996). Motor constraints on vocal development in a songbird. Anim. Behav. 51, 1061–1070. doi: 10.1006/anbe.1996.0107
Podos, J. (1997). A performance constraint on the evolution of trilled vocalizations in a songbird family (Passeriformes: Emberizidae). Evolution 51, 537–551. doi: 10.1111/j.1558-5646.1997.tb02441.x
R Core Team, (2020). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available online at: https://www.R-project.org/
Robb, G. N., McDonald, R. A., Chamberlain, D. E., and Bearhop, S. (2008). Food for thought: supplementary feeding as a driver of ecological change in avian populations. Front. Ecol. Environ. 6, 476–484. doi: 10.1890/060152
Robert, J. H. (2020). Raster: Geographic Data Analysis and Modeling. R package version 3.3-7. Available online at: https://CRAN.R-project.org/package=raster
Rocchini, D., Boyd, D. S., Féret, J.-B., Foody, G. M., He, K. S., Lausch, A., et al. (2016). Satellite remote sensing to monitor species diversity: potential and pitfalls. Remote Sens. Ecol. Conserv. 2, 25–36. doi: 10.1002/rse2.9
Roughgarden, J., Running, S. W., and Matson, P. A. (1991). What does remote sensing do for ecology? J. Ecol. 72, 1918–1922. doi: 10.2307/1941546
Roussel, J.-R., and David, A. (2018). Airborne LiDAR Data Manipulation and Visualization for Forestry Applications. Version 1.5.0.
Salmón, P., Nilsson, J. F., Nord, A., Bensch, S., and Isaksson, C. (2016). Urban environment shortens telomere length in nestling great tits, Parus major. Biol. Lett. 12, 254–260. doi: 10.1098/rsbl.2016.0155
Senar, J. C., Conroy, M. J., and Borras, A. (2002). Asymmetric exchange between populations differing in habitat quality: a metapopulation study on the citril finch. J. Appl. Stat. 29, 425–441. doi: 10.1080/02664760120108791
Shochat, E., Lerman, S., and Fernández-Juricic, E. (2010). Birds in urban ecosystems: population dynamics, community structure, biodiversity, and conservation. Urban Ecosyst. Ecol. Agron. Monogr. 55, 75–86. doi: 10.2134/agronmonogr55.c4
Shochat, E., Warren, P. S., Faeth, S. H., McIntyre, N. E., and Hope, D. (2006). From patterns to emerging processes in mechanistic urban ecology. Trends Ecol. Evol. 21, 186–191. doi: 10.1016/j.tree.2005.11.019
Slabbekoorn, H. (2013). Songs of the city: noise-dependent spectral plasticity in the acoustic phenotype of urban birds. Anim. Behav. 85, 1089–1099. doi: 10.1016/j.anbehav.2013.01.021
Slabbekoorn, H., and den Boer-Visser, A. (2006). Cities change the songs of birds. Curr. Biol. 16, 2326–2331. doi: 10.1016/j.cub.2006.10.008
Slabbekoorn, H., Yeh, P., and Hunt, K. (2007). Sound transmission and song divergence : a comparison of urban and forest acoustics. Condor 109, 67–78. doi: 10.1093/condor/109.1.67
Smith, T. B., Harrigan, R. J., Kirschel, A. N. G., Buermann, W., Saatchi, S., Blumstein, D. T., et al. (2013). Predicting bird song from space. Evol. Appl. 6, 865–874. doi: 10.1111/eva.12072
Sprau, P., Roth, T., Amrhein, V., and Naguib, M. (2013). The predictive value of trill performance in a large repertoire songbird, the nightingale Luscinia megarhynchos. J. Avian Biol. 44, 567–574. doi: 10.1111/j.1600-048X.2013.00113.x
Swaddle, J. P., Francis, C. D., Barber, J. R., Cooper, C. B., Kyba, C. C. M., Dominoni, D. M., et al. (2015). A framework to assess evolutionary responses to anthropogenic light and sound. Trends Ecol. Evol. 30, 550–560. doi: 10.1016/j.tree.2015.06.009
van Oort, H., Otter, K. A., Fort, K. T., and Holschuh, C. I. (2006). Habitat quality, social dominance and dawn chorus song output in black-capped chickadees. Ethology 112, 772–778. doi: 10.1111/j.1439-0310.2006.01228.x
van Oort, H., Otter, K. A., Fort, K. T., and McDonell, Z. (2007). Habitat, dominance, and the phenotypic quality of male black-capped chickadees. Condor 109, 88–96. doi: 10.1093/condor/109.1.88
Ware, H. E., McClure, C. J. W., Carlisle, J. D., and Barber, J. R. (2015). A phantom road experiment reveals traffic noise is an invisible source of habitat degradation. Proc. Natl. Acad. Sci. USA. 112:201504710. doi: 10.1073/pnas.1504710112
Wiley, R. H. (1991). Associations of song properties with habitats for territorial oscine birds of eastern North America. Am. Nat. 138, 973–993. doi: 10.1086/285263
Wilson, D. R., Bitton, P. P., Podos, J., and Mennill, D. J. (2014). Uneven sampling and the analysis of vocal performance constraints. Am. Nat. 183, 214–228. doi: 10.1086/674379
Keywords: vocal performance, territory quality, male quality, urban ecology, birdsong, noise pollution
Citation: Phillips JN, Cooper WJ, Luther DA and Derryberry EP (2020) Territory Quality Predicts Avian Vocal Performance Across an Urban-Rural Gradient. Front. Ecol. Evol. 8:587120. doi: 10.3389/fevo.2020.587120
Received: 24 July 2020; Accepted: 24 November 2020;
Published: 23 December 2020.
Edited by:
Andrew James Jonathan MacIntosh, Kyoto University, JapanReviewed by:
Luis Sandoval, University of Costa Rica, Costa RicaCopyright © 2020 Phillips, Cooper, Luther and Derryberry. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jennifer N. Phillips, am5waGlsbGkyNkBnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.