 Jennifer Morinay
Jennifer Morinay Laure Cauchard
Laure Cauchard Pierre Bize
Pierre Bize Blandine Doligez
Blandine Doligez- 1Laboratoire de Biométrie et Biologie Evolutive UMR 5558, Université de Lyon, Université Lyon 1, CNRS, Villeurbanne, France
- 2Department of Ecology and Genetics/Animal Ecology, Uppsala University, Uppsala, Sweden
- 3School of Biological Sciences, University of Aberdeen, Aberdeen, United Kingdom
In spatio-temporally variable environments, individuals are known to use information for making optimal decisions regarding where and when to breed. Optimal decision making can be complex when relying on multiple information sources with varying levels of reliability and accessibility. To deal with such complexity, different cognitive abilities such as learning and memory might enable individuals to optimally process and use these information sources. Yet, the link between information use and cognitive ability remains unexplored in natural populations. We investigated whether learning performance on a problem-solving task was related to the use of an experimentally manipulated source of social information for nest site selection in wild collared flycatchers (Ficedula albicollis). Collared flycatchers are known to use heterospecific information from their main competitors, the great tits (Parus major). Here, we created a local apparent preference by tits for an artificial nest site feature (a geometric symbol attached to nest boxes occupied by tits) and recorded whether flycatcher pairs chose to settle in nest boxes displaying the same feature as tits (i.e., copied tit apparent preference). Using a problem-solving task requiring opening a door temporarily blocking the nest box entrance, we then measured flycatchers' learning performance during nestling rearing as the number of entrances required to solve the task and enter the nest box twice in a row below a given efficiency threshold. We found that the probability to copy tit preference decreased with decreasing learning performance in females, particularly yearling ones: fast learning females copied tit preference, while slow learning ones rejected it. Male learning performance did not affect copying behavior. Our results showed that learning performance might play an important role in the ability to optimally use information for nest site selection in females: both fast and slow learning females could process this heterospecific information source but used it differently. This could partly explain the link between cognitive abilities and reproductive success reported in previous studies. Whether cognitive abilities may modulate condition-dependent costs of using different information remains to be explored.
Introduction
In most environments, habitat quality varies in time and space, generating environmental uncertainty for individuals, with potentially major fitness consequences (e.g., Franklin et al., 2000; Gunnarsson et al., 2005). Strong selective pressures should therefore favor strategies allowing individuals to reduce this uncertainty by obtaining and using reliable information on habitat quality before making decisions (Danchin et al., 2004; Dall et al., 2005). Many different information sources can be available simultaneously to individuals, such as personal information, acquired from direct observation or sampling of the environment (trial and error tactics), and social information, obtained from observing other individuals' location, behaviors, and success (Danchin et al., 2004; Schmidt et al., 2010). These information sources can vary in reliability (i.e., the extent to which they allow individuals to forecast their own success in the habitat) and accessibility (i.e., the cost at which they can be obtained; Doligez and Boulinier, 2008). They may also provide potentially conflicting information contents (e.g., Grüter et al., 2011). Furthermore, reliability, accessibility, and information contents may themselves depend on individual parameters (e.g., prior knowledge; Kivelä et al., 2014) and/or on the environment (e.g., predation risk; Kendal et al., 2004).
Dealing with multiple information sources for decision making may thus be complex and require different information processing mechanisms. First, individuals may have to prioritize information sources depending on their reliability. For example, individuals may favor the use of personal over social information when the reliability of personal information increases (e.g., when the ecological distance with social information provider increases; Seppänen et al., 2007); conversely, they may favor the use of social over personal information when this reliability decreases (e.g., due to increasing time since information acquisition; van Bergen et al., 2004). Among social information sources, information may be more reliable and favored when provided by conspecifics compared to heterospecifics (Jaakkonen et al., 2015). Second, the information to be prioritized also depends on the context-dependent future rewards of the decision to be made, which implies forecasting the consequences of decisions depending on the context. For example, the use of personal vs. social information can depend on the expected risk of predation: social information may be preferred compared to more reliable personal information for foraging patch choice only when using personal information involves loosing visual contact with conspecifics in a group (Kendal et al., 2004). Adjusting the information used depending on the context was also well-demonstrated in cleaner fish Labroides dimidiatus that alter their cleaning behavior depending on the client fish (Bshary, 2002) or on the audience, in response to potential future clients (Bshary and Grutter, 2006). Third, information use may require long-term memory when the delay between information gathering and use is long (Doligez and Boulinier, 2008). Individuals may use social information cues such as the presence or success of con- or heterospecifics from the previous year to select a breeding site (Doligez et al., 2002; Boulinier et al., 2008; Forsman et al., 2014). In the foraging context, long-term memory has been shown in food-storing birds that hide food reserves during the summer for the next winter (Clayton and Krebs, 1994) but also in non-food-storing birds such as great tits that remembered foraging strategies learned from conspecifics after more than 9 months (Aplin et al., 2014).
The complex use of multiple information sources in a condition- and context-dependent way for optimal decision-making has now been evidenced in various taxa (e.g., in fish, van Bergen et al., 2004; Bshary and Grutter, 2006; birds, Doligez et al., 2008; Kivelä et al., 2014; Jaakkonen et al., 2015; insects, Grüter and Ratnieks, 2011; Grüter et al., 2011; primates, Carter et al., 2013; Kendal et al., 2015). Yet, the mechanisms underlying individuals' ability to cope with such complex information use are still largely unknown. Cognition, broadly defined as all the sensory, neurological, memory, and decision processes used by individuals when interacting with their environment (Shettleworth, 2010), can be expected to play a major role in the ability to optimally process and use information. Cognitive abilities could in particular allow individuals to (i) identify and select the most reliable and/or less costly information cues (e.g., Aplin, 2019), (ii) store and relevantly update information (e.g., Tello-Ramos et al., 2019), (iii) adjust information meaning depending on external factors and individual phenotype (e.g., Kendal et al., 2004), and (iv) use new information by associating cues to fitness-related environmental features (e.g., Loukola et al., 2020). Yet, which cognitive processes enable individuals to optimally use information remains to be investigated in the wild. Among cognitive abilities, learning, defined as the adjustment of behavior to local physical and social conditions through individual experience (Shettleworth, 2010), should in particular allow individuals to efficiently respond to environmental variability by rapidly incorporating new relevant information sources into decision-making processes. Learning can therefore be expected to take a major part in the ability to cope with complex and/or multiple information sources.
Here, we experimentally investigated whether the use of a social information source for nest site selection was related to learning ability measured during nestling rearing in a natural population of a migratory hole-nesting passerine bird, the collared flycatcher. In collared flycatchers and their sister species, pied flycatchers Ficedula hypoleuca, both pair members jointly choose the nest site (Alatalo et al., 1986; Pärt and Qvarnström, 1997). They are well-known to use social information from their main heterospecific resident competitor, the great tit, for making breeding decisions (Seppänen and Forsman, 2007; Forsman et al., 2008) and to gain fitness benefits from this information use (Forsman et al., 2002, 2007). In particular, flycatchers have been experimentally shown to copy great tits' preference for an artificial feature of nesting cavities (e.g., Seppänen and Forsman, 2007; Forsman and Seppänen, 2011; Seppänen et al., 2011). Using the same experimental design, we created an apparent local preference of tits for an artificial nest site feature by attaching geometric symbols on nest boxes occupied by tits. We then recorded whether flycatchers copied this apparent tit preference when choosing their own nest site, i.e., chose a nest box with the same symbol as tits (Seppänen and Forsman, 2007; Seppänen et al., 2011; Morinay et al., 2018). During nestling rearing, we estimated flycatchers' learning performance on a new problem-solving task at the nest by assessing the number of attempts needed to reach a learning threshold when solving the problem over successive trials. If higher cognitive abilities allow individuals to better process and use information, we predict fast learners to be more likely to use social information from tits for nest site selection (i.e., copy the tit apparent preference) than slow or non-learners.
Materials and Methods
Study Species and Population Monitoring
The study was conducted in a natural population of a small hole-nesting passerine bird, the collared flycatcher, breeding on the island of Gotland (Baltic Sea, Sweden, 57°03′ N, 18°18′ E), between April and June from 2012 to 2015. In the study forest patches, collared flycatchers, and titmice (great tits and blue tits Cyanistes caeruleus) breed in sympatry and readily settle in nest boxes provided in excess (Doligez et al., 1999). This allowed us to monitor flycatcher's timing and success of breeding throughout the season, by recording settlement date, laying, and hatching dates, clutch size and fledgling success. Collared flycatchers are migratory and have been shown to use resident titmice as information sources to adjust their own breeding decisions upon arrival on the breeding grounds (Seppänen and Forsman, 2007; Forsman et al., 2008). In particular, flycatchers have been found to use great tits' presence (Kivelä et al., 2014), density (Forsman et al., 2008), phenology (Samplonius and Both, 2017), clutch size (Loukola et al., 2013), and song features (Morinay et al., 2020a) for breeding site selection (along with conspecific social information). The reproductive benefits obtained by using social information from tits may result from settling in higher quality habitat/site as assessed from tit cues or from enhanced access to food resources during nestling rearing and/or social protection from nest predators when settling close to tits (Forsman et al., 2002, 2007). As part of the long-term monitoring of the population, flycatchers breeding in nest boxes were captured (females during incubation and males during nestling provisioning, since both parents feed young), identified (ringed with individually numbered rings if previously unringed) and aged (yearling vs. older based on plumage criteria; Svensson, 1992). Collared flycatchers show plumage sexual dimorphism (males are black and white while females are brown), allowing us to easily sex parents from a distance.
Social Information Use Experiment
The experiment was conducted in 12–17 forest patches (depending on year), containing 29–106 nest boxes each. At the beginning of the breeding season, when tits started settling in nest boxes (i.e., from early April onwards), we chose for each forest patch one (white) geometric symbol between two symbols, either a triangle or a circle, and we attached this symbol around the nest box hole of all boxes occupied by tits in this forest patch. This created a local apparent preference of tits for this artificial nest box feature in the patch. We then randomly attached either a triangle or circle on each unoccupied nest box in the experimental patches so that new-coming birds were provided nest boxes with each of the two symbols in equal proportion in each patch.
When flycatchers arrived on the breeding grounds, on average 2 weeks after tit settlement (i.e., late-April to mid-May), we monitored their nest box choice in terms of the symbol present on the box chosen. We thus recorded whether flycatchers settled in a box with the same symbol than tits in the patch (i.e., they copied local tit apparent preference) or with the other symbol (i.e., they rejected tit apparent preference). To do so, we visited all previously unoccupied nest boxes every second day to detect new settlement of flycatcher or tit pairs (based on nest material used: dry grass for flycatchers and moss with fur or feathers for tits). If a new tit pair settled and the symbol on the box was opposite to the symbol associated with tit nests in the patch, we changed the symbol to match the local apparent tit preference. If a new flycatcher pair settled, we recorded the symbol on the chosen box and removed it to avoid possible information transfer to conspecifics about the pair's choice. We then adjusted the number of each symbol on the remaining unoccupied boxes to keep providing later arriving flycatchers available boxes with each of the two symbols in equal proportion. Keeping an equal proportion was however not always feasible (e.g., for an odd number of empty nest boxes). We thus controlled for the possible resulting deviation by including in our models the ratio of the number of empty boxes with the apparently preferred symbol over the total number of empty boxes −0.5 (i.e., equal proportion) as a covariate (for more details, see Morinay et al., 2018). This experimental design allowed us to disconnect copying behavior (resulting from natural selection for an advantageous information use strategy) from direct fitness benefits of information use because symbols were here associated at random on available nest boxes, i.e., independently from actual box or territory quality. Thus, we investigated here the mere ability of individuals to use heterospecific social information independently from other fitness-related factors.
The symbol associated with tit nests in a given patch was alternated in space, i.e., between patches, and in time, i.e., between years. This allowed us to control for (i) the possible innate preference of hole-nesting birds for a natural circular shape and (ii) the local information gathered in the preceding year by philopatric individuals (i.e., birds that returned to breed in the same forest patch as the preceding year). Furthermore, great tit early reproductive investment (as measured by clutch/brood size on the day of flycatcher settlement) has previously been shown to affect the probability for flycatchers to copy tit apparent preference (Forsman and Seppänen, 2011; Forsman et al., 2012; Morinay et al., 2020b). Flycatchers indeed gather information on tit reproductive investment by directly prospecting tit nests upon their arrival from migration (Forsman and Thomson, 2008; Forsman et al., 2018). This information supposedly reflects tit quality (early breeders for which clutches are complete and/or individuals with overall large clutches being higher quality individuals) and thus the rewards flycatchers may expect when copying their decisions. Accordingly, flycatchers were more likely to copy (respectively, reject) tit apparent preference when local average early reproductive investment of tits in the patch was high (respectively, low) on the day of choice (Forsman and Seppänen, 2011; Seppänen et al., 2011; Loukola et al., 2013; Morinay et al., 2020b). Therefore, we recorded the average great tit clutch/brood size in the forest patch on the settlement day of each flycatcher pair and controlled for this variable in the models testing for the use of social information (here symbol choice) by flycatchers.
Problem-Solving Task and Learning Performance
We measured the individual learning performance of the flycatchers settled in the experimental patches using a problem-solving task (Figure 1) presented at the nest for 1 h during the peak of nestling provisioning, i.e., when nestlings were 6 days old, between 06:00 AM and 04:00 PM. The task featured a door blocking the entrance of the nest box. The participation to the test was therefore motivated here by the access to the young. To access the nestlings, parents had to open the door by perching on one of the three levers attached below the door (Figure 1): only one lever opened the door, either the right or the left one, and the position of the opening lever was randomized among pairs to control for a possible innate preference for one side. Once a parent had entered, the door then closed automatically behind it, allowing us to measure separately each parent's performance in opening the door over successive trials. The parent could afterwards easily get out by pushing the door open from the inside. To limit flycatchers' neophobic response toward the task, we attached on the nest box a fake task with no door and unmoving levers on the evening before the day of the test to let the birds habituate to the device. The 1-h tests were video recorded using a camouflaged camera placed 6–8 m away from the nest box. They were conducted only if nestlings were sufficiently satiated, as measured by begging intensity at the beginning of the test, to avoid starvation if parents were not able to solve the task. If nestlings were strongly begging, the task was not set up and the experimenter came back later during the day or on the next day to check nestling satiation and try again if conditions were more favorable; if this was not the case, no test was conducted and the nest was left out of the study.
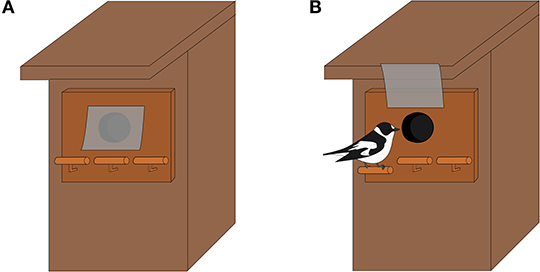
Figure 1. Problem-solving task at the nest used to measure flycatchers' learning ability. (A) The entrance hole is closed by a plastic door but (B) can be opened by the parent (here a male) to enter and feed the nestlings by perching on one of the levers, here the left one. The position of the opening lever, either left or right, was randomized between pairs. See text for the measure of learning performance extracted from the problem-solving test.
Video recordings were analyzed after the field season, blindly with respect to the nest site choice of the flycatcher pair (i.e., whether it copied or rejected tit apparent preference). For each parent, we extracted from the video recording (i) time (in s.) elapsed between the beginning of the test and the first time the individual contacted the task, to control for possible neophobia toward the task despite habituation to the fake task. We also recorded (ii) total time spent on the task (in s.) between the first contact with the task and the first entrance, or until the end of the test for those individuals that never succeeded in entering the nest box during the 1-h test, to control for the level of motivation to solve the task. Finally, for individuals that succeeded in entering, we extracted (iii) the number of times the birds perched on each lever between the first contact with the task and the first entrance, and between subsequent entrances when applicable. Individuals that did not participate to the test (i.e., contacted the task <10 s or <5 times) and those that did not solve the task (i.e., did not enter) were discarded from the analyses.
We scored learning performance using the number of entrances needed to reach a learning threshold. This threshold was based on the number of times the bird perched on any of the levers before entering the nest box. The number of perches used to define the learning threshold was determined from the learning curve of our sample: the mean number of perches before entering showed a strong decrease from the first to the second entrance and then reached a plateau, with no further main change, from the third entrance (Figure 2, Supplementary Table 1). This decrease was accompanied by an increase in the proportion of perches on the opening lever, i.e., correct choices (results not shown here). We thus based our learning threshold on 8 perches, that is, the mean number of perches after which we observed the plateau showing that individuals had on average learnt how to enter (Figure 2). For each entrance, we assessed whether the individual perched <8 times before entering (but we also tested the robustness of our results to this chosen value by repeating all analyses with a threshold based on 6 or 10 perches instead of 8). Because birds may from time to time enter after perching <8 times by chance, we considered that an individual had actually learnt how to enter only if it entered twice in a row with <8 perches per entrance, defining our learning threshold. Individual learning score was then calculated from the number of entrances needed to reach this learning threshold: individuals that entered with <8 perches per entrance both at their first and second entrances (i.e., fastest learning individuals) were assigned a score of 1; those that entered with <8 perches at their second and third entrances were assigned a score of 2; those that entered with <8 perches at their third and fourth entrances were assigned a score of 3; and those that entered with <8 perches at their fourth and fifth entrances were assigned a score of 4. A low score thus corresponded to a low number of entrances needed to reach the learning threshold, thus to fast learning, i.e., high learning performance. Importantly, once an individual reached the learning threshold, the number of perches before subsequent entrances remained low (e.g., 90–95% of individuals with score 1 perched <8 times before the third to fifth entrances), thus validating our choice of two successive entrances with <8 perches to define the threshold.
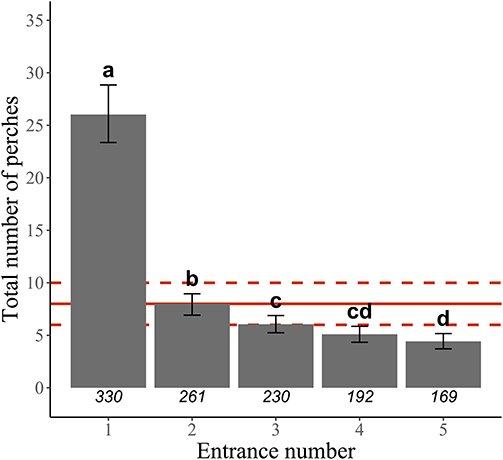
Figure 2. Learning curve showing the decrease in the mean number of perches needed to solve the problem-solving task presented at the nest, i.e., to enter the nest box, over the first five entrances (data restricted to individuals that succeeded in entering the nest box at least once, i.e., solvers, and to the first learning score for individual with repeated scores across years; N = 330 individuals). Predicted means and 95% CI are represented. Bars with different letters have non-overlapping 95% CI. The red solid line corresponds to the value used for defining the learning threshold, i.e., 8 perches or less over two successive entrances (see main text), and the red dashed lines correspond to the two other values used to test the robustness of the results, i.e., 6 and 10 perches, respectively. Sample sizes (number of individuals) are given for each entrance number below each bar. See Supplementary Table 1 for statistics.
To ensure that individuals had the opportunity to learn how to solve the task, we initially included only individuals that entered at least three times (because our learning threshold is based on two entrances in a row). Furthermore, because the total number of entrances over the duration of the test strongly varied among individuals (from 0 to 26 entrances), and because the mean number of perches needed tended to stabilize after the first three entrances (Figure 2), we considered only the first five entrances when individuals entered more than five times to standardize our learning score. Individuals that did not enter with <8 perches twice in a row over up to five entrances were assigned a score of 5. Our initial learning score thus ranged from 1 to 5, with N = 285 individuals scored out of 395 that succeeded to solve the task: 54 with score 1, 119 with score 2, 39 with score 3, 20 with score 4 and 53 with score 5. We exceptionally attributed a score of 1 for three individuals that entered only twice during the test but with <8 perches on both occasions; the results however did not qualitatively change when these three individuals were excluded (results not detailed here). In a second step, we assumed that individuals that entered only once or twice but without reaching the learning threshold could potentially be very slow learners that did not have time to learn within the test duration and we assigned a score of 6 to them (N = 110 individuals). However, because the variability in learning performance for these individuals may be higher than for other score values, we also tested the robustness of our results by repeating all analyses with and without them.
We observed no brood desertion following our learning tests; fledging success (i.e., whether pairs succeeded to fledge at least one young or not), as well as fledgling number and condition (measured as the ratio of body mass over tarsus length) for successful pairs did not differ between pairs that were subjected to our learning test and pairs that were not (Supplementary Material). Thus, our learning tests led to no short- or middle-term adverse effects.
Statistical Analyses
We first explored whether learning performance could be affected by variables often found to influence cognitive performance in previous studies (Griffin and Guez, 2014). Using a Generalized Linear Mixed effects Model (GLMM), we tested whether learning score (i.e., ordinal variable, ranging from 1 [fast] to 5 [slow] or 6 [very slow]—see above) depended on individual sex (male vs. female), age (yearling vs. older individuals, a dichotomy capturing most of the relevant variation in experience in this short-lived species) and their interaction, as well as the latency to return to the box and contact the task for the first time and the total time spent on the task before the first entrance, both mean-centered and scaled (fixed effects). We included the pair as a random effect to account for the potential non-independence of the learning scores of both parents within a pair. Very few individuals were scored for learning performance over several years (31 over 330 ringed individuals in total), and it was not possible to include an individual random effect (quantifying repeatability of learning score) in the model due to convergence issues. Therefore, we retained only the first learning score for each individual with repeated scores.
We then tested whether the probability to copy or reject the apparent tit preference (binary variable: choice of the same vs. opposite symbol than present on boxes occupied by tits in the patch) was related to learning score (considered here as a continuous variable, from 1 to 5 or from 1 to 6) using a GLM. Besides individual's learning score, we included in the model (i) individual's age, to control for possible age-related differences in the probability to copy tit preference as found in previous studies (see also Seppänen and Forsman, 2007 for pied flycatchers; Morinay et al., 2018), (ii) average tit clutch/brood size in the forest patch on the day of flycatcher's nest box choice, and (iii) the deviance to the equal proportion of triangles and circles in the patch (see above). We also included the two-way interaction between individual's learning score and age, to control for possible age-specific relation between learning performance and information use, and the two-way interaction between learning score and tit clutch/brood size, to test whether individuals with higher learning ability may be more prone to use several sources of information simultaneously (here, apparent tit preference and tit early reproductive investment). Because individuals may have been tested several years in the symbol choice experiment, we retained only individuals that were naïve to the symbol experiment, i.e., the first nest box choice with respect to symbols. Finally, the copying behavior being a joint decision by both the male and the female within a pair, we initially intended to include male and female learning scores simultaneously in the model. However, there were only a limited number of pairs for which both partners were naïve to the symbol experiment, scored with the learning test for the first time and were aged (N = 36 pairs). Therefore, we decided to fit sex-specific models including either male or female age and learning score. Yet, we also explored a model with both sexes together on the restricted subset of pairs, and we observed the same trend as for sex-specific models (Supplementary Material). In total, we obtained 232 individuals of known age with both a nest site choice with respect to symbols and a measure of learning performance (130 females and 102 males).
All statistical analyses were performed in R (version 4.0.0, R Core Team, 2020) and we show here posterior means and 95% CI for all estimates. In all models, we initially included year and forest patch as random factors to control for potential spatio-temporal variation in learning performance or copying behavior, but we removed them because (i) the variance associated with year and patch did not differ from zero (lower bound of 95% CI <10−6) or (ii) including these random effects led to convergence issues (both year and forest patch passed stationary tests, but year failed to pass half-width tests despite increased iteration number; heidel.diag diagnosis function, CODA R package; Plummer et al., 2006). Interactions for which 95% CI overlapped zero were also removed (Engqvist, 2005). All models were fitted with the function MCMCglmm (R package MCMCglmm; Hadfield, 2010). Learning score, considered as an ordered factor, and copying probability were fitted with “threshold” families. We used the default wide Normally distributed priors for fixed terms (μ = 0, V = 1010), and expanded priors (V = 1, υ = 1, α.μ = 0, α.V = 1,000) for random terms. We fixed the residual variance to 1 in all models (Hadfield, 2010). We parameterized all models to obtain an effective sample size >2,000 and autocorrelation levels <0.1. We used 3 × 106 iterations, a burn-in of 3 × 105 and a thinning interval of 800 for models fitting learning score, and 105 iterations, a burn-in of 104 and a thinning-interval of 20 for models fitting copying probability. Convergence was assessed visually and by running Gelman and Rubin's convergence diagnosis tests on three MCMC chains with different starting points (gelman.diag function, package CODA, Plummer et al., 2006).
Results
Factors Affecting Learning Performance
Learning score depended neither on individual's sex or age, nor on the latency to contact the task (Table 1). Learning score, i.e., the number of entrances needed to reach down the learning threshold, however increased (and thus learning performance decreased) with increasing time spent on the task before the first entrance: slow learners (with higher score) took more time to enter for the first time than fast learners (estimate [95% CI]: 0.82 [0.46; 1.20] for individuals with learning scores from 1 to 5, and 0.39 [0.21; 0.58] when also including individuals with a learning score of 6; Figure 3, Supplementary Figure 1, Table 1, Supplementary Table 2).
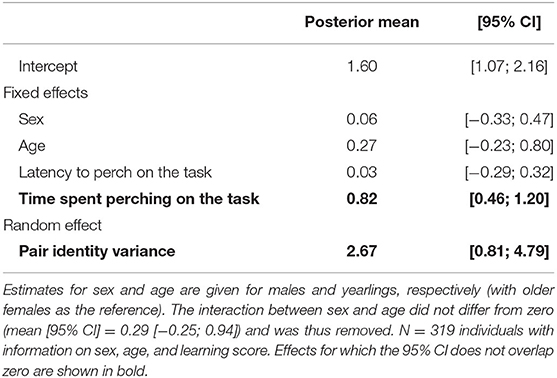
Table 1. Influence of age, sex, latency to perch on the task and time spent perching on the task before the first entrance on the learning score [from 1 (fast learners) to 5 (slow or non-learners)].
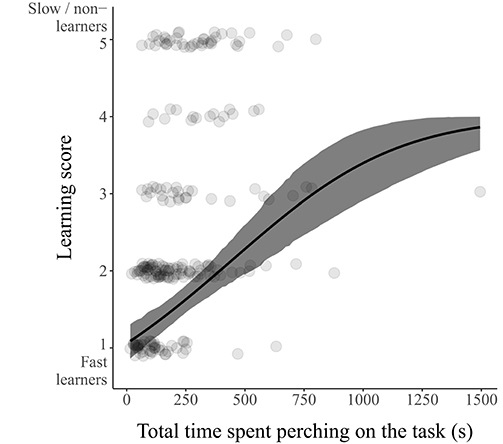
Figure 3. Relation between the total time spent perching on the task before the first entrance and learning score. Low score values indicate fast learning individuals, and high score values slow or non-learning individuals (i.e., learning score increases with decreasing learning performance). Predicted distributions (means and 95% CI) are estimated for the sake of illustration from a model including only time spent perching on the task as a fixed effect. Data points are shown in gray.
Learning Performance and Social Information Use
Among females, learning score affected the probability to copy apparent tit preference. When we considered a learning threshold based on <8 perches per entrance and included females with score from 1 to 5, the effect of learning score was found to differ between yearling and older females (interaction learning score × age; Table 2). Yearling females with lower score, i.e., higher learning performance, were more likely to copy tit apparent preference, while yearling females with higher score, i.e., lower learning performance, were more likely to reject it (Figure 4). This relation between learning performance and probability to copy was however not detected in older females alone, despite a similar decreasing trend (mean [95% CI] = −0.19 [−0.42; 0.04]; Figure 4). Importantly, when considering a learning threshold based on either 6 or 10 perches, or when including females with a score from 1 to 6 (even for the threshold based on 8 perches), the probability to copy tit preference decreased with increasing learning score, i.e., with decreasing learning performance, with no difference between the two age categories (no interaction between learning score and female age; Table 3, Supplementary Table 3, Supplementary Figure 2, 3). The decrease with learning score alone was also retained in the model for the learning threshold based on <8 perches per entrance and females with score from 1 to 5 when the interaction with age was removed (Table 3). Females with lower score were more likely to copy tit apparent preference (mean [95% CI] copying probability for females with score 1 = 0.65 [0.51; 0.79], i.e., >0.5 = random choice), while females with higher score were more likely to reject it (mean [95% CI] copying probability for females with score 6 = 0.34 [0.22; 0.46], i.e., <0.5; Supplementary Figure 3). In addition, for all female models, the probability to copy tit preference also increased with tit clutch/brood size within the patch on the day of female choice (Table 2, Supplementary Table 3; no interaction with age).

Table 2. Influence of individual's age and learning score on the probability to copy tit preference.
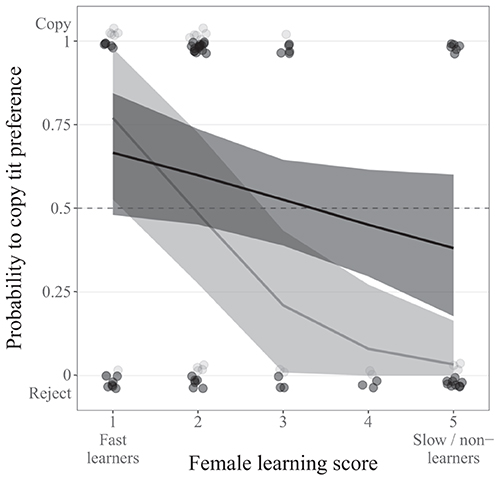
Figure 4. Relation between learning score (from 1 to 5) and the probability to copy apparent tit preference for nest site choice in flycatcher females, depending on female age (light gray: yearlings: N = 25, dark gray: older females: N = 59). Predicted distributions (mean and 95% CI) are estimated from the model including the interaction between learning score and age, with a mean value set for other continuous predictors (see Supplementary Table 2). Data points show actual choices.
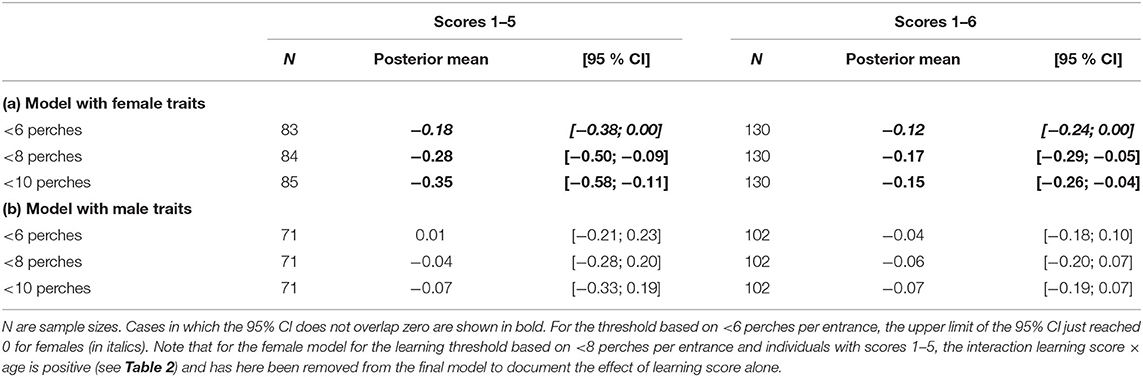
Table 3. Summary of the estimates for the learning score effect on the probability to copy tit preference, when alone (i.e., no interaction with age included in the model) depending on the value considered for defining the learning threshold (<6, 8, or 10 perches per entrance, for two successive entrances) and the individuals considered (individuals with scores 1–5 only or individuals with scores 1–6), for (a) females and (b) males.
Among males, the probability to copy tit preference did not depend on learning score, alone or in interaction with age, whatever the model considered, i.e., for a learning threshold based on either 6, 8, or 10 perches, and when including or not males with a score of 6 (Tables 2, 3, Supplementary Table 4).
Discussion
To make optimal decisions, individuals often have to process multiple, complex, and potentially conflicting information from the environment. Cognitive abilities can therefore be expected to play a crucial role in such information processing. In this study, we used an experimental approach on wild collared flycatchers to test whether variation in learning ability was linked to variation in the use of a source of heterospecific social information (here, apparent tit preference for a nest site feature) for nest site selection. Overall, our results show that females that were faster to learn how to solve a new task at the nest were more likely to copy tit apparent preference while females that were slower to learn were more likely to reject it. This effect seemed more pronounced in yearling females, but because the interaction between learning score and female age was found only when the learning threshold was based on 8 perches before entrance and when females with score 6 were excluded, and in no other case, it did not seem to reflect a strong biological difference. Despite between-pair variance in learning performance, we found no relation between learning score and the probability to copy tit preference in males, whether yearling or older. These results suggest that cognitive abilities (in this study learning ability) can at least partly shape information use for decision-making, here in females, providing a new mechanistic explanation for the positive link between cognitive abilities and fitness components (Cauchard et al., 2017; see also Ducatez et al., 2020) that was previously observed in the wild (e.g., Keagy et al., 2011; Cauchard et al., 2013; Smith et al., 2015).
Linking Cognition and Fitness: The Role of Information Processing
To explain the links between cognitive abilities and fitness traits recently reported in wild populations (e.g., reproductive success: Cauchard et al., 2013; Smith et al., 2015; Ashton et al., 2018; survival: Sonnenberg et al., 2019; Wild et al., 2019), it has been hypothesized that individuals with higher cognitive abilities may achieve higher fitness because of a higher ability to optimally exploit their habitat (Cole et al., 2012; Cauchard et al., 2017). This may rely on a better processing and use of information sources about the environment (e.g., prioritize and memorize information but also learn from either personal or social experience), allowing individuals to optimally choose between alternatives (Mery, 2013). This is in line with previous results showing that higher problem-solving ability was associated with increased nestling provisioning rate (Cauchard et al., 2017), which could involve different habitat use strategies (e.g., range of territory use: Cole et al., 2012). Yet, empirical evidence that cognitive abilities influence the use of complex information for decision making in the wild is still extremely scarce. Our results experimentally show that, in the collared flycatcher, learning performance is related to the use of a manipulated source of information for nest site selection, a crucial decision with major impact on reproductive success. The complex use of social information of heterospecific origin (i.e., from potential competitors) for nest site selection has been demonstrated in previous studies on flycatchers based on the same experimental set up: individuals modulate the use of apparent tit preference for nest site choice depending on tit early reproductive investment (Forsman and Seppänen, 2011; Seppänen et al., 2011; Loukola et al., 2013; Morinay et al., 2020b), but also their own previous experience (Morinay et al., 2018, 2020b) and behavioral traits (Morinay et al., 2020b). In our study, we make one step forward by showing that between-individual variation in the context- and condition-dependent use of this social information source may also depend on variation in learning ability.
One important but still open question is the nature of the cognitive processes underlying our measure of learning ability and how these processes relate to those underlying social information use for decision-making. Identifying these cognitive processes would allow us to better understand on which cognitive trait(s) selective pressures are acting. Although we did not explore here the detailed cognitive processes involved in learning how to solve our task, the test was based on operant conditioning, i.e., a type of associative learning process in which the strength of a behavioral response is modified by reinforcement (leading to response increase) or punishment (leading to response decrease; Skinner, 1938). Here, reinforcement corresponded to door opening, granting access to nestlings, and the rewarded behavior was perching on the opening lever. Such associative learning process is widespread in the animal kingdom (Dickinson, 2012; Morand-Ferron et al., 2015) and should underlie many behavioral decisions during the breeding season, especially those involving repeated individual actions, such as here the visit of many nest boxes needed to associate the symbol to box occupancy by tits and tit early reproductive investment. Much remains to be done to unravel the key underlying cognitive processes (e.g., associative learning, memory, spatial cognition) that are driving between-individual variation in information use processes in the wild and their fitness consequences.
Influence of Individual Factors on the Link Between Learning and Information Use
The relation observed here between learning performance and the use of heterospecific social cues for nest site choice depended on sex: the probability to copy tit apparent preference decreased with decreasing learning performance (i.e., with increasing number of entrances to reach down the learning threshold) only in females. Male learning performance had no effect on copying behavior here, even though nest site choice is a joint behavior by both pair members (Pärt, 1994; Pärt and Qvarnström, 1997). Previous studies in collared and pied flycatchers suggest that females often use social information collected at large spatial scales for breeding site selection (e.g., local tit phenology at the forest patch scale: Samplonius and Both, 2017; tit song quality at the scale of a group of nest sites: Morinay et al., 2020a). In contrast, males are more prone to use social information obtained at small scales (e.g., nest reproductive success at the territory scale: Pärt and Doligez, 2003). The link between learning performance and probability to copy tit preference in females is in line with this difference, because the social information provided here relies on associating tit occupancy to the symbol attached to the box at a large (patch) spatial scale. Conversely, males secure a territory encompassing several adjacent nest boxes that most often display both symbols, because our experimental protocol avoided large spatial aggregations of same-symbol boxes. Therefore, the strong male philopatry to breeding patches (e.g., Pärt, 1994, 1995; Germain et al., 2017) and the previously observed effects of male age, experience, and aggressiveness on the probability to copy tit preference for nest-site choice (Morinay et al., 2018, 2020b) may be a response to strong male-male competition. Males familiar with their environment may have an easy access to local heterospecific social information, thus requiring no strong learning ability to process this information for choosing a site. Conversely, females are expected to gain advantage in choosing a nest site over larger scales (Greenwood, 1980), possibly favoring the use of more complex quality-related social cues, requiring higher cognitive abilities for processing these cues. Hence, the sex-specific relation observed in the present study between learning performance and use of social information may result from the different selective pressures acting on both sexes for settlement decisions. Alternatively, females with higher learning performance may be more efficient in assessing, and mating with, more competitive (aggressive) males (Qvarnström et al., 2000), which themselves choose nest boxes with the symbol matching tit preference (Morinay et al., 2020b); in this case, the relation between learning performance and use of social information in females would be indirect. Further work is needed to explore the consequences of learning performances of paired individuals on complex social information use for joint nest site selection.
Even though age-dependent use of social information has been repeatedly reported in previous studies on this population (e.g., Doligez et al., 2004; Kivelä et al., 2014; Morinay et al., 2020a), including studies using the same experimental symbol design (Morinay et al., 2018, 2020b), the age difference in the effect of female learning score on the probability to copy tit preference was not strongly supported. Thus, here, differences between yearling and older females in either past experience, current competitive level, or time constraints linked to the timing of arrival from migration did not seem to strongly affect the link between learning performance and information use.
Only fast-learning females could have been expected to succeed in optimally and/or quickly processing available social information, and thus be more prone to copy tit preference. Yet, slow-learning females were not simply choosing a nest site at random with respect to tit apparent preference: they were actually more likely to reject it compared to a random choice. This suggests that, although all females could access this source of information, they may have processed and/or used it differently for subsequent choices depending on their learning ability. One hypothesis is that slow-learning females may be less competitive than fast-learning ones because of a lower ability to exploit the habitat, e.g., because of a lower ability to use other information sources. By rejecting tit preference, slow-learning females would then avoid expected increased competition with dominant great tits. In this case, the decision-making process of both fast and slow learners may lead to an optimal nest site choice with respect to competitive ability (see e.g., Doligez et al., 1999 for another example of age-related opposite use of social information with respect to competitive ability). Tit apparent preference as a source of social information may be accessible to all individuals regardless of their learning ability; but the relation observed between the probability to copy tit preference and learning performance could indirectly result from the impairment of acquiring other information sources in slow-learning individuals.
This study evidences a link between learning performance and the use of (here heterospecific) social information for nest site choice in the wild. Yet, this relationship appears more complex than initially thought, with sex- (and possibly age-) dependent effects potentially reflecting different consequences for subsequent choices. By reporting a clear relationship between learning performance and social information use, our study moves one step forward toward understanding the mechanisms linking cognition and fitness in the wild. This provides support for the role of cognitive abilities in using complex information for decision-making, with major ecological and evolutionary implications.
Data Availability Statement
The data supporting the findings of this study are openly available in FigShare at http://doi.org/10.6084/m9.figshare.13229081.
Ethics Statement
Birds were caught, handled, and ringed under a license from the Ringing Centre of the Museum in Stockholm (471:M015 to BD). Behavioral experiments were conducted in accordance with international standards on animal welfare and complied with local and national regulations.
Author Contributions
LC and BD designed the study. JM, LC, and BD carried out the field work. LC and PB supervised the data extraction and JM carried out the statistical analyses. JM and BD drafted the manuscript and all authors revised it.
Funding
This work was funded by the ANR (Evol-Cog project, ANR-19-CE02-0007), the Centre National pour la Recherche Scientifique (PICS) and the Region Rhône-Alpes (CIBLE programme) to BD; the NSERC (postdoctoral fellowship), the ABS (student research award), the BOU and the BES (research grants), and the SCO (Fred Cook award) to LC; the Ministère de l'Enseignement Supérieur et de la Recherche, the Department of Ecology and Genetics from Uppsala University and Stiftelsen för Zoologisk Forskning, the Région Auvergne Rhone-Alpes (Explora'Doc mobility grants) and by the University of Lyon (ATER fellowship and IDEX mobility grant) to JM; the Pearcy Sladen Memorial Trust and Carnegie Trust (travel grants), and the BOU and BES (research grants) to PB.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Jukka Forsman for helping with the symbol experiment protocol, and Lars Gustafsson for granting us access to the field site. We also thank all land owners for letting us conduct our experiments in their forest patches, and all field workers and students who contributed to data collection and extraction.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.559690/full#supplementary-material
References
Alatalo, R. V., Lundberg, A., and Glynn, C. (1986). Female pied flycatchers choose territory quality and not male characteristics. Nature 323, 152–153. doi: 10.1038/323152a0
Aplin, L. M. (2019). Culture and cultural evolution in birds: a review of the evidence. Anim. Behav. 147, 179–187. doi: 10.1016/j.anbehav.2018.05.001
Aplin, L. M., Farine, D. R., Morand-Ferron, J., Cockburn, A., Thornton, A., and Sheldon, B. C. (2014). Experimentally induced innovations lead to persistent culture via conformity in wild birds. Nature 518, 538–541. doi: 10.1038/nature13998
Ashton, B. J., Ridley, A. R., Edwards, E. K., and Thornton, A. (2018). Cognitive performance is linked to group size and affects fitness in Australian magpies. Nature 554, 364–367. doi: 10.1038/nature25503
Boulinier, T., McCoy, K. D., Yoccoz, N. G., Gasparini, J., and Tveraa, T. (2008). Public information affects breeding dispersal in a colonial bird: kittiwakes cue on neighbours. Biol. Lett. 4, 538–540. doi: 10.1098/rsbl.2008.0291
Bshary, R. (2002). Biting cleaner fish use altruism to deceive image-scoring client reef fish. Proc. R. Soc. B Biol. Sci. 269, 2087–2093. doi: 10.1098/rspb.2002.2084
Bshary, R., and Grutter, A. S. (2006). Image scoring and cooperation in a cleaner fish mutualism. Nature 441, 975–978. doi: 10.1038/nature04755
Carter, A. J., Marshall, H. H., Heinsohn, R., and Cowlishaw, G. (2013). Personality predicts decision making only when information is unreliable. Anim. Behav. 86, 633–639. doi: 10.1016/j.anbehav.2013.07.009
Cauchard, L., Angers, B., Boogert, N. J., Lenarth, M., Bize, P., and Doligez, B. (2017). An experimental test of a causal link between problem-solving performance and reproductive success in wild great tits. Front. Ecol. Evol. 5:107. doi: 10.3389/fevo.2017.00107
Cauchard, L., Boogert, N. J., Lefebvre, L., Dubois, F., and Doligez, B. (2013). Problem-solving performance is correlated with reproductive success in a wild bird population. Anim. Behav. 85, 19–26. doi: 10.1016/j.anbehav.2012.10.005
Clayton, N. S., and Krebs, J. R. (1994). Memory for spatial and object-specific cues in food-storing and non-storing birds. J. Comp. Physiol. A 174, 371–379. doi: 10.1007/BF00240218
Cole, E. F., Morand-Ferron, J., Hinks, A. E., and Quinn, J. L. (2012). Cognitive ability influences reproductive life history variation in the wild. Curr. Biol. 22, 1808–1812. doi: 10.1016/j.cub.2012.07.051
Dall, S. R. X., Giraldeau, L.-A., Olsson, O., McNamara, J. M., and Stephens, D. W. (2005). Information and its use by animals in evolutionary ecology. Trends Ecol. Evol. 20, 187–193. doi: 10.1016/j.tree.2005.01.010
Danchin, E., Giraldeau, L.-A., Valone, T. J., and Wagner, R. H. (2004). Public information: from nosy neighbors to cultural evolution. Science 305, 487–491. doi: 10.1126/science.1098254
Dickinson, A. (2012). Associative learning and animal cognition. Philos. T. R. Soc B 367, 2733–2742. doi: 10.1098/rstb.2012.0220
Doligez, B., Berthouly, A., Doligez, D., Tanner, M., Saladin, V., Bonfils, D., et al. (2008). Spatial scale of local breeding habitat quality and adjustment of breeding decisions. Ecology 89, 1436–1444. doi: 10.1890/07-0113.1
Doligez, B., and Boulinier, T. (2008). “Habitat selection and habitat suitability preferences,” in Behavioral Ecology. Vol. [3] of Encyclopedia of Ecology, Vol. 5, eds S. E. Jørgensen and B. D. Fath (Oxford: Elsevier), 1810–1830. doi: 10.1016/B978-008045405-4.00015-X
Doligez, B., Danchin, E., and Clobert, J. (2002). Public information and breeding habitat selection in a wild bird population. Science 297, 1168–1170. doi: 10.1126/science.1072838
Doligez, B., Danchin, E., Clobert, J., and Gustafsson, L. (1999). The use of conspecific reproductive success for breeding habitat selection in a non-colonial, hole-nesting species, the collared flycatcher. J. Anim. Ecol. 68, 1193–1206. doi: 10.1046/j.1365-2656.1999.00362.x
Doligez, B., Pärt, T., Danchin, E., Clobert, J., and Gustafsson, L. (2004). Availability and use of public information and conspecific density for settlement decisions in the collared flycatcher. J. Anim. Ecol. 73, 75–87. doi: 10.1111/j.1365-2656.2004.00782.x
Ducatez, S., Lefebvre, L., Sayol, F., Audet, J., Clayton, D., and Adamo, S. (2020). Host cognition and parasitism in birds : a review of the main mechanisms. Front. Ecol. Evol. 8:102. doi: 10.3389/fevo.2020.00102
Engqvist, L. (2005). The mistreatment of covariate interaction terms in linear model analyses of behavioural and evolutionary ecology studies. Anim. Behav. 70, 967–971. doi: 10.1016/j.anbehav.2005.01.016
Forsman, J. T., Hjernquist, M. B., Taipale, J., and Gustafsson, L. (2008). Competitor density cues for habitat quality facilitating habitat selection and investment decisions. Behav. Ecol. 19, 539–545. doi: 10.1093/beheco/arn005
Forsman, J. T., Kivelä, S. M., Jaakkonen, T., Seppänen, J.-T., Gustafsson, L., and Doligez, B. (2014). Avoiding perceived past resource use of potential competitors affects niche dynamics in a bird community. BMC Evol. Biol. 14, 175. doi: 10.1186/s12862-014-0175-2
Forsman, J. T., and Seppänen, J.-T. (2011). Learning what (not) to do: testing rejection and copying of simulated heterospecific behavioural traits. Anim. Behav. 81, 879–883. doi: 10.1016/j.anbehav.2011.01.029
Forsman, J. T., Seppänen, J.-T., and Mönkkönen, M. (2002). Positive fitness consequences of interspecific interaction with a potential competitor. Proc. R. Soc. B 269, 1619–1623. doi: 10.1098/rspb.2002.2065
Forsman, J. T., Seppänen, J.-T., Mónkkónen, M., Thomson, R. L., Kivelä, S. M., Krams, I., et al. (2018). Is it interspecific information use or aggression between putative competitors that steers the selection of nest-site characteristics? A reply to Slagsvold and Wiebe. J. Avian Biol. 49:01558. doi: 10.1111/jav.01558
Forsman, J. T., Seppänen, J.-T., and Nykänen, I. L. (2012). Observed heterospecific clutch size can affect offspring investment decisions. Biol. Lett. 8, 341–343. doi: 10.1098/rsbl.2011.0970
Forsman, J. T., and Thomson, R. L. (2008). Evidence of information collection from heterospecifics in cavity-nesting birds. Ibis 150, 409–412. doi: 10.1111/j.1474-919X.2007.00773.x
Forsman, J. T., Thomson, R. L., and Seppänen, J.-T. (2007). Mechanisms and fitness effects of interspecific information use between migrant and resident birds. Behav. Ecol. 18, 888–894. doi: 10.1093/beheco/arm048
Franklin, A. B., Anderson, D. R., Gutiérrez, R. J., and Burnham, K. P. (2000). Climate, habitat quality, and fitness in northern spotted owl populations in Northwestern California. Ecol. Monogr. 70, 539–590. doi: 10.1890/0012-9615(2000)070[0539:CHQAFI]2.0.CO;2
Germain, M., Pärt, T., and Doligez, B. (2017). Lower settlement following a forced displacement experiment: nonbreeding as a dispersal cost in a wild bird? Anim. Behav. 133, 109–121. doi: 10.1016/j.anbehav.2017.09.001
Greenwood, P. J. (1980). Mating systems, philopatry and dispersal in birds and mammals. Anim. Behav. 28, 1140–1162. doi: 10.1016/S0003-3472(80)80103-5
Griffin, A. S., and Guez, D. (2014). Innovation and problem solving: a review of common mechanisms. Behav. Proces. 109, 121–134. doi: 10.1016/j.beproc.2014.08.027
Grüter, C., Czaczkes, T. J., and Ratnieks, F. L. W. (2011). Decision making in ant foragers (Lasius niger) facing conflicting private and social information. Behav. Ecol. Sociobiol. 65, 141–148. doi: 10.1007/s00265-010-1020-2
Grüter, C., and Ratnieks, F. L. W. (2011). Honeybee foragers increase the use of waggle dance information when private information becomes unrewarding. Anim. Behav. 81, 949–954. doi: 10.1016/j.anbehav.2011.01.014
Gunnarsson, T. G., Gill, J. A., Newton, J., Potts, P. M., and Sutherland, W. J. (2005). Seasonal matching of habitat quality and fitness in a migratory bird. Proc. R. Soc. B Biol. Sci. 272, 2319–2323. doi: 10.1098/rspb.2005.3214
Hadfield, J. D. (2010). MCMC methods for multi-response generalized linear mixed models: The MCMCglmm R package. J. Stat. Softw. 33, 1–22. doi: 10.18637/jss.v033.i02
Jaakkonen, T., Kivelä, S. M., Meier, C. M., and Forsman, J. T. (2015). The use and relative importance of intraspecific and interspecific social information in a bird community. Behav. Ecol. 26, 55–64. doi: 10.1093/beheco/aru144
Keagy, J., Savard, J. F., and Borgia, G. (2011). Complex relationship between multiple measures of cognitive ability and male mating success in satin bowerbirds, Ptilonorhynchus violaceus. Anim. Behav. 81, 1063–1070. doi: 10.1016/j.anbehav.2011.02.018
Kendal, R. L., Coolen, I., and Laland, K. N. (2004). The role of conformity in foraging when personal and social information conflict. Behav. Ecol. 15, 269–277. doi: 10.1093/beheco/arh008
Kendal, R. L., Hopper, L. M., Whiten, A., Brosnan, S. F., Lambeth, S. P., Schapiro, S. J., et al. (2015). Chimpanzees copy dominant and knowledgeable individuals: implications for cultural diversity. Evol. Hum. Behav. 36, 65–72. doi: 10.1016/j.evolhumbehav.2014.09.002
Kivelä, S. M., Seppänen, J.-T., Ovaskainen, O., Doligez, B., Gustafsson, L., Mönkkönen, M., et al. (2014). The past and the present in decision-making: the use of conspecific and heterospecific cues in nest site selection. Ecology 95, 3428–3439. doi: 10.1890/13-2103.1
Loukola, O. J., Gatto, E., Híjar-Islas, A. C., and Chittka, L. (2020). Selective interspecific information use in the nest choice of solitary bees. Anim. Biol. 70, 1–11. doi: 10.1163/15707563-20191233
Loukola, O. J., Seppänen, J.-T., Krams, I., Torvinen, S. S., and Forsman, J. T. (2013). Observed fitness may affect niche overlap in competing species via selective social information use. Am. Nat. 182, 474–483. doi: 10.1086/671815
Mery, F. (2013). Natural variation in learning and memory. Curr. Opin. Neurobiol. 23, 52–56. doi: 10.1016/j.conb.2012.09.001
Morand-Ferron, J., Hamblin, S., Cole, E. F., Aplin, L. M., and Quinn, J. L. (2015). Taking the operant paradigm into the field: associative learning in wild great tits. PLoS ONE 10:e0133821. doi: 10.1371/journal.pone.0133821
Morinay, J., Forsman, J. T., and Doligez, B. (2020a). Heterospecific song quality as social information for settlement decisions: an experimental approach in a wild bird. Anim. Behav. 161, 103–113. doi: 10.1016/j.anbehav.2020.01.002
Morinay, J., Forsman, J. T., Germain, M., and Doligez, B. (2020b). Behavioural traits modulate the use of heterospecific social information for nest site selection: experimental evidence from a wild bird population. Proc. R. Soc. B Biol. Sci. 287:20200265. doi: 10.1098/rspb.2020.0265
Morinay, J., Forsman, J. T., Kivelä, S. M., Gustafsson, L., and Doligez, B. (2018). Heterospecific nest site copying behavior in a wild bird: assessing the influence of genetics and past experience on a joint breeding phenotype. Front. Ecol. Evol. 5:167. doi: 10.3389/fevo.2017.00167
Pärt, T. (1994). Male philopatry confers mating advantage in migratory collared flycatchers. Anim. Behav. 48, 401–409. doi: 10.1006/anbe.1994.1254
Pärt, T. (1995). The importance of local familiarity and search costs for age- and sex-biased philopatry in the collared flycatcher. Anim. Behav. 49, 1029–1038. doi: 10.1006/anbe.1995.0132
Pärt, T., and Doligez, B. (2003). Gathering public information for habitat selection: prospecting birds cue on parental activity. Proc. Biol. Sci. 270, 1809–1813. doi: 10.1098/rspb.2003.2419
Pärt, T., and Qvarnström, A. (1997). Badge size in collared flycatchers predicts outcome of male competition over territories. Anim. Behav. 54, 893–899. doi: 10.1006/anbe.1997.0514
Plummer, M., Best, N., Cowles, K., and Vines, K. (2006). CODA: convergence diagnosis and output analysis for MCMC. R News 6, 7–11.
Qvarnström, A., Griffith, S. C., and Gustafsson, L. (2000). Male-male competition and parental care in collared flycatchers (Ficedula albicollis): An experiment controlling for differences in territory quality. Proc. R. Soc. B Biol. Sci. 267, 2547–2552. doi: 10.1098/rspb.2000.1319
R Core Team (2020). R: A Language and Environment for Statistical Computing. Available online at: https://www.r-project.org/ (accessed May 25, 2020).
Samplonius, J. M., and Both, C. (2017). Competitor phenology as a social cue in breeding site selection. J. Anim. Ecol. 86, 615–623. doi: 10.1111/1365-2656.12640
Schmidt, K. A., Dall, S. R. X., and van Gils, J. A. (2010). The ecology of information: an overview on the ecological significance of making informed decisions. Oikos 119, 304–316. doi: 10.1111/j.1600-0706.2009.17573.x
Seppänen, J.-T., and Forsman, J. T. (2007). Interspecific social learning: novel preference can be acquired from a competing species. Curr. Biol. 17, 1248–1252. doi: 10.1016/j.cub.2007.06.034
Seppänen, J.-T., Forsman, J. T., Mönkkönen, M., Krams, I., and Salmi, T. (2011). New behavioural trait adopted or rejected by observing heterospecific tutor fitness. Proc. R. Soc. B Biol. Sci. 278, 1736–1741. doi: 10.1098/rspb.2010.1610
Seppänen, J.-T., Forsman, J. T., Mönkkönen, M., and Thomson, R. L. (2007). Social information use is a process across time, space, and ecology, reaching heterospecifics. Ecology 88, 1622–1633. doi: 10.1890/06-1757.1
Smith, C., Philips, A., and Reichard, M. (2015). Cognitive ability is heritable and predicts the success of an alternative mating tactic. Proc. R. Soc. B Biol. Sci. 282:20151046. doi: 10.1098/rspb.2015.1046
Sonnenberg, B. R., Branch, C. L., Pitera, A. M., Bridge, E., and Pravosudov, V. V. (2019). Natural selection and spatial cognition in wild food-caching mountain chickadees. Curr. Biol. 29, 670–676.e3. doi: 10.1016/j.cub.2019.01.006
Tello-Ramos, M. C., Branch, C. L., Kozlovsky, D. Y., Pitera, A. M., and Pravosudov, V. V. (2019). Spatial memory and cognitive flexibility trade-offs: to be or not to be flexible, that is the question. Anim. Behav. 147, 129–136. doi: 10.1016/j.anbehav.2018.02.019
van Bergen, Y., Coolen, I., and Laland, K. N. (2004). Nine-spined sticklebacks exploit the most reliable source when public and private information conflict. Proc. Biol. Sci. 271, 957–962. doi: 10.1098/rspb.2004.2684
Keywords: breeding site choice, collared flycatcher, Ficedula albicollis, heterospecific social information, information processing, problem-solving task, trial and error learning
Citation: Morinay J, Cauchard L, Bize P and Doligez B (2020) The Role of Cognition in Social Information Use for Breeding Site Selection: Experimental Evidence in a Wild Passerine Population. Front. Ecol. Evol. 8:559690. doi: 10.3389/fevo.2020.559690
Received: 06 May 2020; Accepted: 05 November 2020;
Published: 27 November 2020.
Edited by:
Sasha Raoul Xola Dall, University of Exeter, United KingdomReviewed by:
Wiebke Schuett, University of Sussex, United KingdomAmanda R. Ridley, University of Western Australia, Australia
Copyright © 2020 Morinay, Cauchard, Bize and Doligez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Laure Cauchard, bGF1cmUuY2F1Y2hhcmRAYWJkbi5hYy51aw==
†These authors have contributed equally to this work
‡Present address: Jennifer Morinay, Area Avifauna Migratrice, Istituto Superiore per la Protezione e la Ricerca Ambientale (ISPRA), Ozzano dell'Emilia, Italy