 Susanne Åkesson
Susanne Åkesson- 1Center for Animal Movement Research, Department of Biology, Lund University, Lund, Sweden
- 2Groningen Institute for Evolutionary Life Sciences, University of Groningen, Groningen, Netherlands
Endogenous programs that regulate annual cycles have been shown for many taxa, including protists, arthropods, fish, mammals and birds. In migration biology, these programs are best known in songbirds. The majority of songbirds rely on a genetic program inherited from their parents that will guide them during their first solo-migration. The phenotypic components of the program are crucial for their individual fitness and survival, and include time components, direction, and distance. This program is constructed to both guide behavior and to regulate flexible responses to the environment at different stages of the annual cycle. The migration program is driven by a circannual rhythm, allowing for, and resetting, carry-over effects. With experience, the migration decisions of individual migrants may be based on information learnt on breeding sites, wintering sites, and en route. At the population level, substantial variation in route choice and timing of migration may be explained by inherited variation of program components, by interactions with environmental and social factors, and by individual learning. In this review we will explore the components of endogenous migration programs and discuss in what ways they can lead to flexibility and variation in migration behavior.
Introduction: Migrations Over Time and Space
Migrations, which we here define as regular periodic movements, have been linked to fluctuations of relevant environmental factors, such as nutrients (Newton, 2008; Häfker et al., 2017), breeding site or mate availability (Barlow et al., 1986; Hodgson and Quinn, 2002; Friesen, 2015), predation risk (McKinnon et al., 2010; Häfker et al., 2017), infection probability (O’Connor et al., 2018), and abiotic survival threats (e.g., severe weather or UV radiation; Hut and Beersma, 2011; Reid et al., 2018). Such environmental fluctuations, and the associated periodic movements of organisms, usually recur on time scales defined by geophysical cycles: annual cycles caused by Earth’s orbit around the sun, diel cycles caused by Earth’s rotation around its axis, lunar cycles caused by the orbit of Moon around Earth, and tidal cycles caused by the combined movements of Earth and Moon (DeCoursey, 2004; Figure 1A). Their common, planetary basis makes these cycles predictable, even if their downstream environmental consequences (e.g., temperature or food availability) may be more variable. The principle predictability of environmental cycles has provided the substrate for organisms to evolve time-keeping mechanisms that are fitted to their life-styles and specific environments.
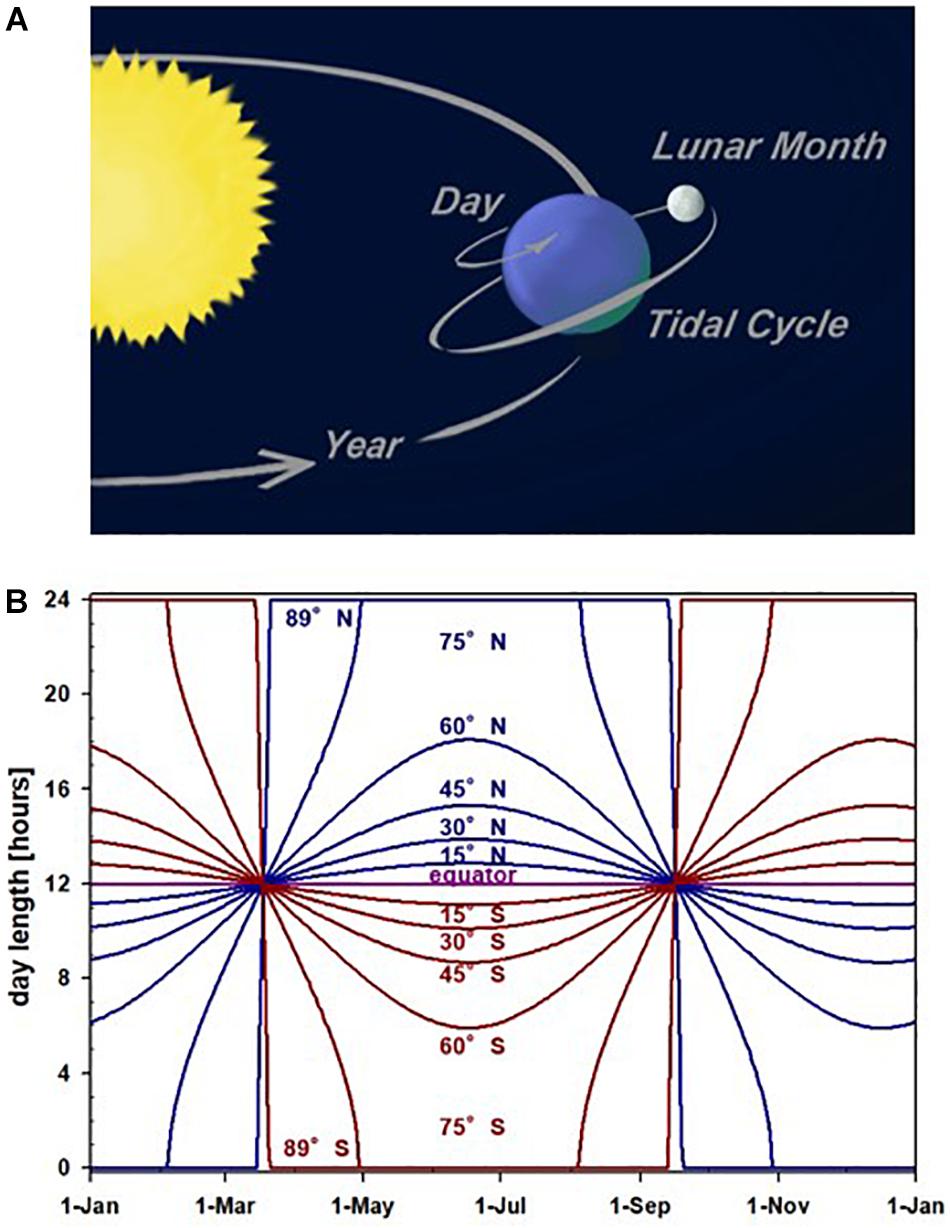
Figure 1. Geophysical basis of periodic migrations. (A) Periodicities arising from planetary movements include annual cycles, diel cycles, and variants of lunar and tidal cycles; image: Edda Starck. (B) Daylength (photoperiod) changes across the year at different latitudes. Daylength changes are locally predictably, but only for organisms that remain at the same latitude.
Organisms use biological time-keeping to predict upcoming changes, to prepare for them, and to spatially relocate in anticipation (Åkesson et al., 2017; Helm et al., 2017). In a nutshell,
these mechanisms integrate internally generated (i.e., endogenous) rhythms and responses to environmental cues into timing programs. In migratory organisms, spatial features, for example direction of travel or magnetic field cues, are often included, so that even naive individuals can perform effective migrations (Kramer, 1957; Åkesson et al., 2017; Reppert and de Roode, 2018).
These intriguing spatio-temporal programs have been extensively studied from a full-cycle perspective in migratory organisms on all four predictable time-scales introduced above (Figure 1A; annual, diel, lunar and tidal; e.g., Barlow et al., 1986; Gwinner, 1996a, b; Åkesson et al., 2017; Häfker et al., 2017). Central questions in this research field are concerned with inheritance of migration programs, their integration of environmental cues, and their physiological and genetic mechanisms (van Noordwijk et al., 2006; Åkesson et al., 2017; Merlin and Liedvogel, 2019). Here, we review key insights, and apply them to address sources of variation in spatiotemporal migratory traits within and among individuals, as well as among populations. Our review is aimed at explaining how endogenous programs can generate, or counteract, variation in migration. Among the countless contributions to the field, we highlight those that in our view are most suitable to achieving this aim, rather than attempting to give a balanced or exhaustive record of this vast research field.
The review actively contributes to a collection of articles that constitute the Frontiers research topic Flexibility in the Migration Strategies of Animals (Senner et al., 2020). Coherently with this Frontiers research topic, we will focus on annual time scales, choosing birds as study subjects, but the principle considerations also hold for other time scales and other periodically migrating organisms. We also streamline the use of terminology in our article as laid out for the entire Frontiers research topic. To facilitate cross-fertilization of ideas, we will first clarify the terminology and conceptual background of migration programs, which are not widely known among field-based researchers. We will then explain in greater detail how migration programs work, how they interact with environmental information, and how they regulate aspects of variation. Thereafter we will discuss additional variation and its possible interpretation. We address these points from theoretical perspectives and illustrate them by selected examples. Due to both our own geographic location and geographic differences in research focus on timing programs, our article is greatly biased toward the northern hemisphere, with emphasis on European data, although we strove to give global examples.
Terminology and Concepts
Flexibility, Plasticity, Variation
The central topic of this Frontiers research topic is flexibility, defined on an individual level as variation in traits that can be reversed in response to an individual’s environment throughout their lives. Such trait reversals can for example be based on experience, can occur in response to inter-annual differences in environmental conditions, or can be a generic part of the annual cycle (i.e., life-cycle staging, sensu Piersma and Drent, 2003). Variation through flexibility is distinguished from polyphenisms (sensu Piersma and Drent, 2003), where individuals differ from each other irreversibly because of genetic differences. It is also distinguished from developmental plasticity, which denotes variation in traits that is irreversibly determined during ontogeny (sensu Piersma and Drent, 2003). We here adopt this terminology for coherence across the Frontiers research topic, diverging from our use elsewhere (Helm et al., 2017).
Additionally, we will introduce a distinction between two types of flexibility. Because migration programs by definition regulate responses to the environment, we consider a substantial proportion of flexibility to be programmed (i.e., resulting from inherited reaction norms; van Noordwijk et al., 2006; Visser et al., 2010; Helm et al., 2017). A remaining part of residual flexibility is not readily explained by our current knowledge of migration programs. On a population level, we will refer to differences in migration as variation because their mechanistic basis is not clear. Differences in individual traits, such as timing or route choice, and in strategies, such as partial or differential migration, could be based on genetic differences, developmental plasticity, or individual flexibility.
Ontogenetic Perspective
Additional emphasis in this Frontiers research topic is on ontogenetic variation in migration. From a perspective of migration programs, the main distinction is between an individual’s first journey and its subsequent migrations. During their first migrations, juveniles are naive and depend on their programs, on social guidance, or on trial and error. In subsequent years, birds will have additional experience (Perdeck, 1958), which may override the initially expressed migration program. A further distinction will be made between open-ended learners and those that adhere to their first experience for their future migrations (Gill et al., 2014).
Carry-Over Effects
A full annual-cycle perspective is at the heart of research on migration programs (Gwinner, 1996b; Briedis et al., 2016), and hence, carry-over effects have long been studied within this field. In adherence to the lay-out for the present Frontiers research topic, we adopt a broad definition of carry-over effects to include all instances when previous history explains current performance, as long as they are functionally important and separated in time (O’Connor et al., 2014). Hence, learning and developmental adjustments mostly fall also under this definition.
From a perspective of timing programs, a distinction is made between carry-over effects within an annual cycle, and those between cycles. In many species, the timing program allows for high variation during some phases of the annual cycle, whereas during others individuals resynchronize to environmental cues and reset their annual timing (e.g., Helm et al., 2005; Conklin et al., 2013; Karagicheva et al., 2016; Briedis et al., 2018; Gow et al., 2019). Thus, we consider carry-over effects within an annual cycle as flexibility that can be regulated by the migration program. In contrast, we view carry-over effects between annual cycles as modifications of the migration program, for example due to experience or to poor state. In either case, we view carry-over effects as an outcome of trade-offs, or, alternatively phrased, of different optimization criteria (Alerstam and Lindström, 1990). Animals may compromise optimal timing for other benefits, such as improved state or additional broods, but may pay costs for suboptimal timing at subsequent annual-cycle stages (see also Senner et al., 2015). Alternatively, they can adhere to timing, at potential costs to state (i.e., departing in poor condition) or to reproduction (e.g., skipping a breeding opportunity to depart in time).
Migratoriness
Species differ greatly in the level of variation of their migrations (Newton, 2008). Generally, spatio-temporal precision and consistency increase with migration distance, with proportion of population members migrating, and with the rigidity of its regulation (Tryjanowski et al., 2005; Newton, 2008). This trend can be captured in the term migratoriness. Migration programs are most useful if birds move between sites that are spatially too distinct to assess environmental conditions of goal areas, and if conditions at the goal areas are sufficiently predictable to facilitate evolution of migration programs. If these conditions are met, birds tend to score high on migratoriness. An example for clear differences in migratory precision of related species with similar ecology are waders on Iceland, where relatively short-distance migrants (e.g., Black-tailed Godwits, Limosa limosa islandica) have substantially advanced arrival time over the last decades (Gill et al., 2014), whereas a long-distance migrant (Whimbrel, Numenius phaeopus islandicus) has not (Carneiro et al., 2019).
Migration Programs for Time and Space
As new data from avian migrations flood in during this golden age of bio-logging (McKinnon and Love, 2018), efforts to distill patterns have invigorated interest in migration programs. Consistent timing is perhaps the most commonly emerging pattern (Altshuler et al., 2013; Briedis et al., 2018; McKinnon and Love, 2018; Carneiro et al., 2019; Gow et al., 2019). Routes are often more variable (Stanley et al., 2012; Vardanis et al., 2016), although in some studies they were more consistent than timing (Vardanis et al., 2011; Sugasawa and Higuchi, 2019). When timing is consistent within individuals, there may be large variation within and between populations, for example in departure date of sympatrically overwintering individuals (e.g., Conklin et al., 2010, 2013; Briedis et al., 2016; cf. Gow et al., 2019).
Much of the new evidence fits well with our current understanding of migration programs. Researchers had long postulated the existence of innate programs to explain why migratory birds do not simply stay at the wintering grounds, and how they return for breeding at the right time of year. Support for innate programs first came from observations by bird fanciers. When wild conspecifics would migrate, caged birds, provided with ample food and shelter, also performed migration-like behaviors (Birkhead, 2008) and directional movements (e.g., Kramer, 1957). This behavior is called migratory restlessness, or Zugunruhe. It is most readily observed in nocturnally migrating species which show bouts of migratory restlessness at night, but similar arguments have also been made for some diurnally migrating species (e.g., Bojarinova and Babushkina, 2015). It is important to note that the restless hopping and flying in cages does not directly represent migration, but rather a captive expression of motivation to migrate (Helm, 2006; Van Doren et al., 2016; Bäckman et al., 2017). However, this frustrated movement state (John Rappole, pers. comm.) often captures important aspects of migration and has been key to our understanding of bird migration. Migratory restlessness has been reported for many northern hemisphere-breeding migrants that breed in Europe, America and Asia. Species included mainly passerines and some other taxa, for example quail (e.g., Helms, 1963; King and Farner, 1963; Gwinner, 1996a, b; Budki et al., 2009; Bertin et al., 2007; Eikenaar et al., 2014; Watts et al., 2016). For tropical and southern hemisphere breeders, only a few Zugunruhe records exist, including intra-tropically migrating yellow–green vireos, Vireo flavoviridis; Styrsky et al., 2004) in the Americas, for stonechats in Africa (Saxicola torquata axillaris, Helm and Gwinner, 2006, which however are locally resident), and for Australian silvereyes (Zosterops lateralis; Chan, 1995).
Although migratory restlessness is not always easy to interpret, its regulation through robustly innate programs was confirmed in migratory birds that were kept under constant conditions of daylength, temperature and food (Gwinner, 1986, 1996b; Holberton and Able, 1992). Over many years Zugunruhe alternated with molt and with reactivation of the reproductive system approximately annually (hence, called circannual). Thereby, it became clear that life-cycle stages including migratory behaviors were driven by an endogenous (i.e., self-generated) circannual rhythm. Furthermore, when tested for their directional preference, birds shifted the orientation of their Zugunruhe activities as appropriate for the corresponding leg of migration (Gwinner and Wiltschko, 1980). A host of additional physiological changes that enhance migration also occurred (e.g., hyperphagia, fuel deposition; King and Farner, 1963; Gwinner, 1996a; Newton, 2008). Migratory activity within this endogenous migration program is encoded in individual birds in relation to migration distance, with short-distance migrants generally expressing shorter periods of migratory restlessness than long-distance migrants (e.g., Berthold, 1973; Berthold and Querner, 1981; Maggini and Bairlein, 2010; Bulte and Bairlein, 2013). Because the endogenous migration program also encodes migration direction, it can lead migratory naïve individuals along routes to population-specific wintering areas (e.g., Helbig, 1991).
Time
Annual Timing
Under constant conditions, without any changes in the captive environment, circannual cycles recur, but their period lengths are ca. 9–15 months, so that life-cycle stages usually drift to occur at earlier or later dates over progressive years (Gwinner, 1996b; Karagicheva et al., 2016). In nature, conversely, annual cycles do not drift, and life-cycle stages recur annually, usually at similar dates. Hence, it became clear that the circannual clock functioned in interaction with environmental cues that synchronize and modify its timing. Therefore, a spate of experimental studies investigated the synchronizing effects of environmental factors on migration programs across the annual cycle (Gwinner, 1996b; Helm et al., 2009).
Among the synchronizing cues, photoperiod, the annually changing light fraction of the 24 h day (Figure 1B), has the strongest effects, and can both, advance or delay the annual cycle. For example, in multi-brooded species, chicks hatch at widely different times of year. The correspondingly different daylengths experienced in early life can then synchronize chicks from consecutive broods. The contribution of the timing program to synchronizing these birds is illustrated by data from captive stonechats (Saxicola spec.) from three regions (Europe, East Africa, Kazakhstan) that bred under naturally changing daylengths (Helm et al., 2005; Figure 2). Chicks that grew up under shorter photoperiods, simulating late hatching at the end of summer, compensated by accelerated postjuvenile development in population-specific ways. Late-hatched of all populations largely caught up with earlier-born conspecifics by advancing autumn Zugunruhe by 0.9 days per day of later hatching. By this genetically programmed compensation mechanism, which was confirmed by field observations from wild conspecifics, stonechats achieved a high level of within-population synchrony (Helm et al., 2005) that counteracted carry-over effects of late hatching. Similar advancement of Zugunruhe for later-hatched chicks was also observed in birds breeding at low latitudes, for example in yellow–green vireos (Styrsky et al., 2004).
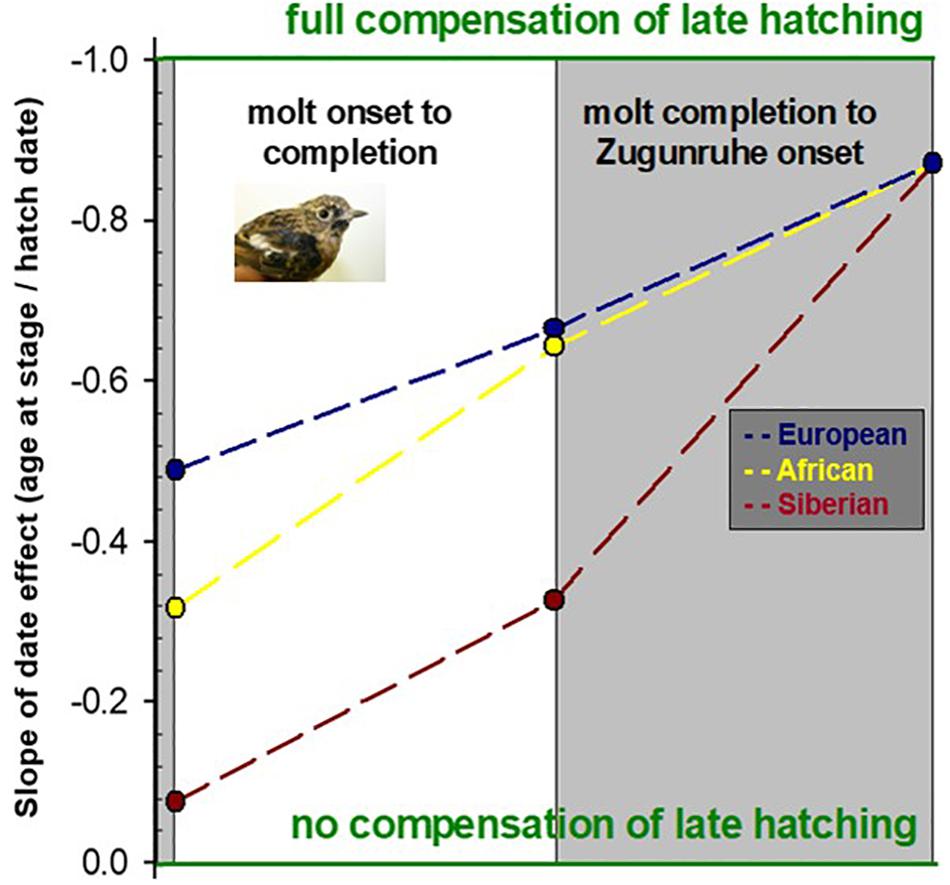
Figure 2. Synchronization of autumn activities. The graph shows data from captive young stonechats from different populations that hatched at different times of year under naturally changing photoperiods. Stonechats from all populations compensated for late hatching by earlier start of molt, faster molt, and younger ages at initiating Zugunruhe. Shown are on the y-axis the slopes of age at the onset and end of molt, and at the start of Zugunruhe, against hatch date on the x-axis (from left to right). Based on Helm et al. (2005). Inlay: stonechat during postjuvenile molt.
Synchronization within populations that counteracts carry-over effects also occurs at the end of overwintering when migrants initiate spring migration (Conklin et al., 2013; Senner et al., 2014; Briedis et al., 2018). For example, recent work on socially migrating tree swallows (Tachycineta bicolor) suggests that differential timing of migration may continue as a domino-effect set by breeding latitude until resynchronization in the winter quarters (Gow et al., 2019). Such resetting effects can be so strong that the non-breeding period has been described as buffering the build-up of carry-over effects (Senner et al., 2014; Briedis et al., 2018). However, for low-latitude wintering grounds, where photoperiod undergoes little change, it is still unclear how such synchronization is achieved, although effective cues have been described for breeding in equatorial birds (e.g., Goymann et al., 2012; Shaw, 2017). A key property of timing programs is that the responses to environmental factors, which they encode, are specific to stage (phase) of the annual cycle (Figure 3). It is intuitively sensible that a migratory bird would respond to a long, warm day differently on the breeding grounds than at its winter quarters. The same environmental cues can thereby cause either advances, delays, or no changes to the annual cycle, depending on time of year. This time-dependence can be shown systematically by plotting timing responses over the annual cycle. Such phase-response curves (Figure 3A) or sensitivity profiles (Figure 3B) have been described by studies of biological rhythms (DeCoursey, 1960; Gwinner, 1996b; Helm et al., 2009) and phenology time series (Thackeray et al., 2016), respectively.
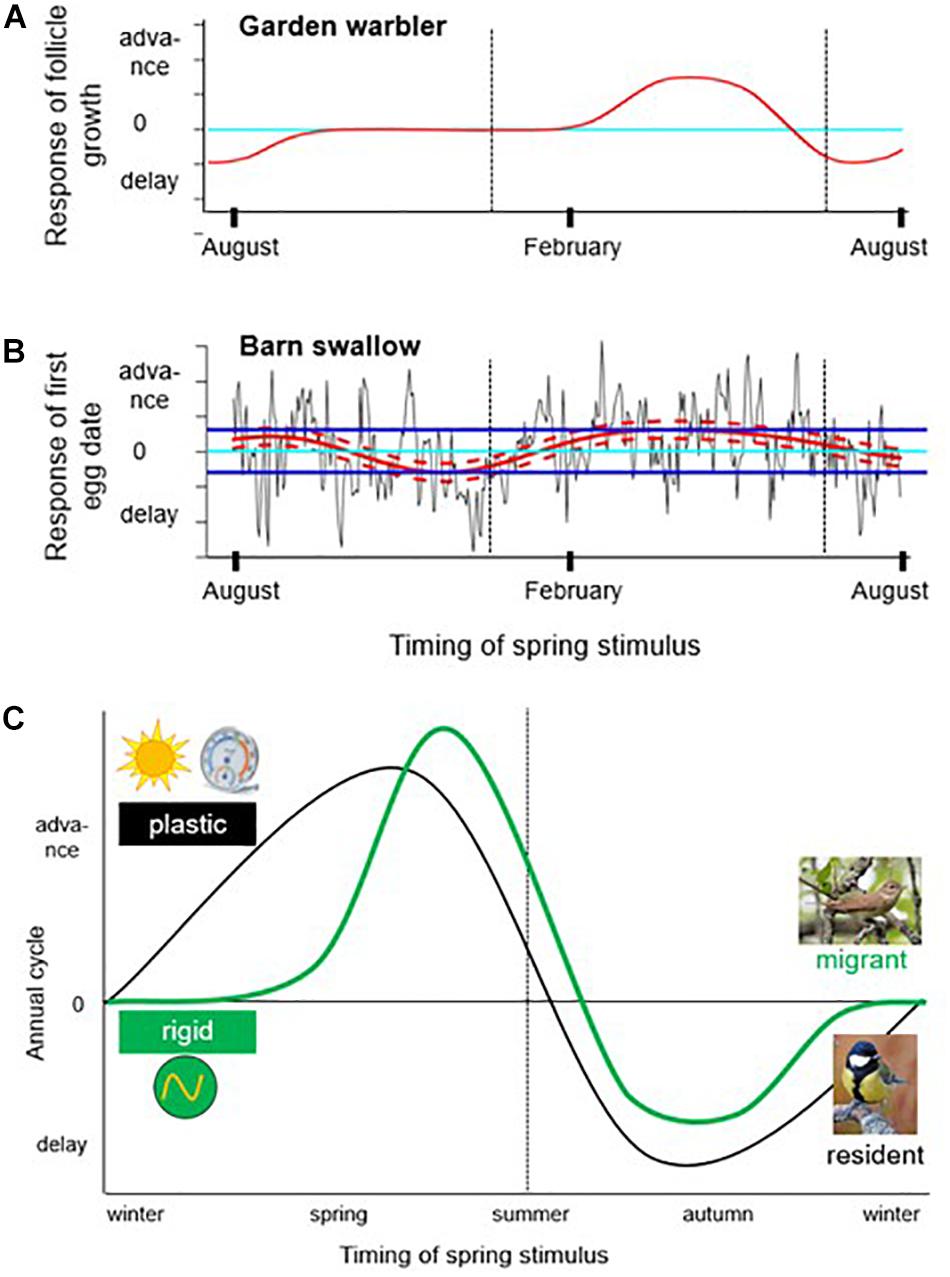
Figure 3. Responses to environmental factors depend on the phase of the annual cycle. Graphs show advance and delay responses of seasonal events (y-axis) to cues experienced at different times of year (x-axis). (A) Alternating responses to temperature and precipitation, depending on the phase of the annual cycle, has been described as the dominant pattern of wild organisms based on 10,000 time series from the UK (Thackeray et al., 2016). Shown is the phase-dependent temperature response of egglaying in wild Barn Swallows (image kindly generated by Dario Massimino). (B) Phase-dependent response of follicle growth to photoperiod in captive Garden Warblers (based on Gwinner, 1996a). (C) Schematic difference between migrants and residents in the phase-specific response to spring-like cues. In late winter and spring, long days and high temperatures advance subsequent life cycle stages, such as spring migration and reproductive activation. Conversely, later in the year the response reverses, and long days and high temperatures delay life-cycle stages. Long-distance migrants (inlay: Garden Warbler) differ from residents (inlay: Great Tit) by a rigid phase during wintering, when long days and high temperatures have little or no effect on the annual cycle; image credit to commons.wikimedia.org: Garden Warbler by Kristjan Osbek, Great Tit by Biillyboy.
Figure 3A shows the response of female reproductive timing (follicle growth) to long days in captive Garden Warblers (Sylvia borin) (Gwinner, 1996b). This usually single-brooded species responds to long days in summer with a shut-down of the reproductive system. Subsequently, reproductive responsiveness is low over winter, but in spring garden warblers respond to long days with reproductive activation and advance their annual cycle. A downregulated response is important for migratory species which in their winter quarters experience conditions that may induce breeding (Hamner and Stocking, 1970; Gwinner, 1996b; Helm et al., 2009). To be sure to return to breeding sites in spring, rather than breed in the winter quarters, it is important to ignore potentially misleading cues at some times of year, while paying close attention to these cues at other times.
A recent, grand-scale study of phenology of wild species in the United Kingdom showed wide-spread, time-dependent sensitivity to ambient temperature and precipitation (Thackeray et al., 2016). The most common pattern was that prior to a phenological event (e.g., breeding), high temperature or precipitation advanced its timing. At earlier dates, response profiles were typically flat, and even earlier, high temperature or precipitation delayed events. Figure 3B shows the phase-dependent response of egglaying to ambient temperature in Barn Swallows (Hirundo rustica; Dario Massimino, pers. comm.; Thackeray et al., 2016). Barn Swallows show advance responses to high temperatures in spring and summer, followed by delay responses during autumn and winter. The broadly similar findings from captive and wild birds emphasize the relevance of phase-specific responses for seasonally appropriate behavior. Differences between the species, in turn, may be due to the more flexible annual behaviors of Barn Swallows. It might be no coincidence that in Barn Swallows, recent cross-hemispheric colonization was observed, associated with complete inversion of the annual cycle (Winkler et al., 2017). Such an inversion is easy to envision if some individuals become sensitive to long and warm days while still on the winter grounds.
Differences between species in response-profiles have important implications for the ability of birds to respond to climate change. Strongly migratory species typically differ from residents by lower flexibility in response to spring cues (Phillimore et al., 2016). Figure 3C shows schematically how flexibility of migrants is specifically reduced in winter compared to resident species. Because of this programming difference, which has likely been adaptive, migrants may now be constrained in their ability to flexibly adjust annual cycles, and instead require evolutionary change (Phillimore et al., 2016). That such change may be possible has recently been shown for Pied Flycatchers (Ficedula hypoleuca; Helm et al., 2019). In this study, a full-annual cycle experiment on captive birds was replicated after 21 years, over which period a wild population had been continuously monitored. Spring activities of both, captive and wild birds, advanced at similar rates (9 and 11 days in 21 years, respectively). In the captive birds, where the full annual cycle was monitored, this advancement occurred selectively during the late winter and early spring phases, suggesting evolutionary acceleration of the circannual clock during winter (Helm et al., 2019).
Diel Timing
Migratory flights in many species occur at night, implying a seasonal change in individuals from almost exclusively daytime activity to nocturnal flight. This shift to additional nocturnality is starkly detectable in the wild, for example on a continental scale using weather radars (Horton et al., 2020), and in captivity as migratory restlessness as described above. Recent studies have indeed indicated that strong Zugunruhe associates with higher probability to migrate in the wild (Eikenaar et al., 2014; Mukhin et al., 2018). However, wild and captive birds also show differences in the extent of night activity during migration seasons. For example, whereas wild birds intersperse migration nights between several nights of rest, captive birds typically show restlessness on most nights of the migration seasons (Åkesson et al., 2017).
Shifts to nocturnality during migration seasons are puzzling, given that in birds and most other organisms, day-night rhythms are stably organized by circadian clocks (Helm et al., 2017; Figure 4). Experimental studies on several species of songbirds have shown that Zugunruhe is, however, organized as part of the avian circadian system, rather than supplanting it (Bartell and Gwinner, 2005; Kumar et al., 2006; Coppack et al., 2008; Coverdill et al., 2008; Mukhin et al., 2018). For example, bouts of Zugunruhe, recognizable by extensive flight behavior, recur rhythmically even under experimental conditions when birds are fully sheltered from environmental information (i.e., receiving continuous dim light, constant temperature and food availability). Several studies have suggested that the birds’ circadian system contains at least two internal drivers of rhythms (i.e., oscillators), of which one produces diurnal and the other nocturnal activity (Bartell and Gwinner, 2005; Mukhin et al., 2018). For much of the year, output from the day-time driver dominates, but during migration seasons, the night-time driver’s activity becomes discernible. The location and functional details of the oscillators driving daytime and nighttime activity are not yet resolved. The avian circadian system consists of several pacemakers (Figure 4) that are interconnected, and are in turn sensitive to multiple environmental sensory inputs as well as to signaling from within the body (Kumar et al., 2006; Cassone, 2014; Helm et al., 2017). The particular responsiveness of Zugunruhe (see Adjusting the Drive to Migrate and Fueling in Response to Geomagnetic Cues) to food availability suggests links of the Zugunruhe oscillator to metabolic signals, and perhaps to brain circuits that are part of the award system (Bartell and Gwinner, 2005; Horton et al., 2019).
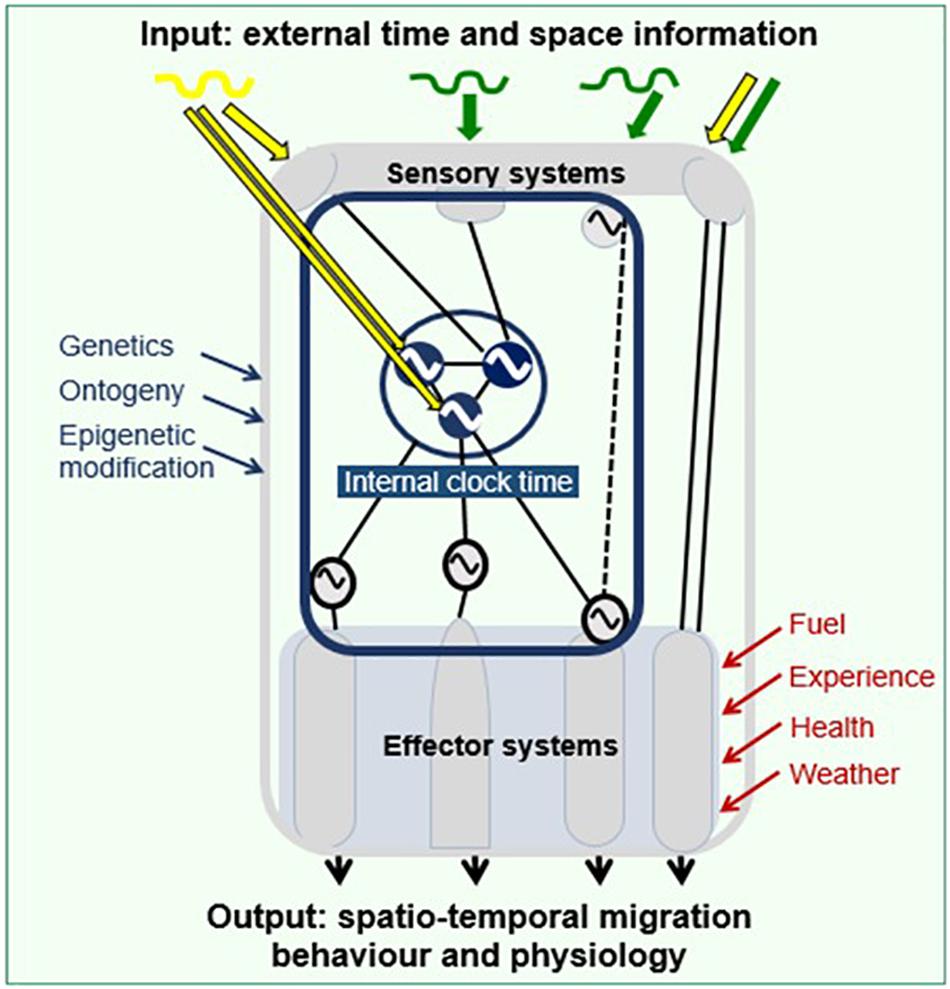
Figure 4. Mechanistic framework of spatio-temporal migration programs. Gray frame delineates the organism. It receives spatio-temporal information from the geophysical (yellow) and the biotic (green) environment, perceived through its sensory systems. This information is integrated with the animal’s biological clock (blue) to generate internal clock time. Clock time modulates effector systems, which integrate additional modifying information (red) from within the body (e.g., fuel reserves) and from the environment (e.g., weather), to set the spatio-temporal migration behavior and physiology. Oscillator symbols indicate biological rhythms (central clock in inner circle, additional clock components peripheral). The clock system is itself modified by different factors (blue); for details see Helm et al. (2017).
Space
Birds tend to follow inherited species- and population-specific migration routes (e.g., Helbig, 1996; Willemoes et al., 2014), which will lead them to suitable stop-over and wintering areas (Fransson et al., 2005; Newton, 2008). The endogenous programs guiding young birds encode compass and space information in relation to the internal clock (Berthold, 1996; Gwinner, 1996a, b; Able, 1980; Åkesson et al., 2014). The three biological compasses used by migratory birds are based on information from the sun and the skylight polarization pattern, stars and the geomagnetic field (e.g., Wiltschko and Wiltschko, 1972; Emlen, 1975; Able, 1980; Schmidt-Koenig, 1990; Åkesson et al., 2014), and their use is tightly connected to the diel and circannual time sense. The sun compass has a time-compensation mechanism enabling compensation for the apparent movement of sun across the sky (Schmidt-Koenig, 1990; Schmidt-Koenig et al., 1991), while the stellar compass encodes direction toward geographic north based on the rotation center of the sky independent of time of day (Emlen, 1967, 1970). The magnetic compass is expressed relative to the angle of inclination providing directions along a north-south axis toward and away from the poles without direct diel time input for its functionality (Wiltschko and Wiltschko, 1972), but changes of courses are expressed at relevant times of year (e.g., Gwinner and Wiltschko, 1978; Wiltschko and Wiltschko, 1992). Perception of the magnetic field seems dependent on specialized photoreceptors activated by a limited range of wavelengths of light involving cryptochrome molecules (Ritz et al., 2009; Muheim et al., 2014). Compass interactions may further lead to recalibrations during migration (e.g., Cochran et al., 2004; Muheim et al., 2006; cf. Åkesson et al., 2015), while during ontogeny a combined experience of geomagnetic information and a rotating star pattern is crucial for birds to express a relevant population-specific migratory direction at the right time of year (Weindler et al., 1996).
Once migrants have started their journey, simple compass mechanisms can sometimes explain the routes they follow (e.g., Kiepenheuer, 1984; Alerstam and Pettersson, 1991; Muheim et al., 2003, 2018; Åkesson and Bianco, 2016; cf. Sokolovskis et al., 2018). For instance, long-distance bird migrants have been proposed to set a course at sunset or sunrise and follow it relative to the position of the sun as they cross longitudes using their time-compensated sun compass without readjusting it for local time during flight (Alerstam and Pettersson, 1991; Alerstam et al., 2001). This mechanism has gained some support from radar observations of long-distance migrating arctic waders (Alerstam et al., 2001). Birds could also use their geomagnetic compass (i.e., inclination compass; Wiltschko and Wiltschko, 1972), and keep track of the apparent inclination angle during long continuous flights, i.e., following magnetoclinic routes (Kiepenheuer, 1984), in many cases leading birds along realistic migration routes as confirmed by tracking data (Åkesson and Bianco, 2016, 2017). Especially challenging situations are met by using any of the alternative compasses in high arctic regions (e.g., Åkesson et al., 2001a; Muheim et al., 2003, 2018; Åkesson and Bianco, 2016). Recently, a comparative study evaluating route simulations demonstrated potential use across the widest latitudinal range for the magnetic compass (i.e., magnetoclinic route; Åkesson and Bianco, 2017).
An increasing number of bird tracking studies have revealed complex course changes throughout the annual cycle (e.g., Sutherland, 1998; Berthold et al., 2004; Åkesson et al., 2012; Willemoes et al., 2014). Such complex routes, involving one or more shifts during migration (Helbig et al., 1989; Willemoes et al., 2014), raise the question of how course shifts are encoded relative to the circannual program in different species and populations of birds. We find experimental support for course shifts expressed at expected times under constant environmental conditions (Gwinner and Wiltschko, 1978). In turn, there is evidence that geomagnetic information also affects the migration program. For example, expression of a relevant course shift required in some species exposure to the geomagnetic information expected at specific latitudes along the migration route (Beck and Wiltschko, 1982, 1988).
Geomagnetic information has also been shown to prompt ecophysiological changes, leading to increased mass increase in response to magnetic parameters associated with sites just in front of a large barrier. These findings, first shown in juvenile Thrush Nightingales (Luscinia luscinia; Fransson et al., 2001), suggest that this response is inherited and encoded in the endogenous migration program. For course shifts and refueling, the endogenous circannual time program seems to be involved in controlling the timing of events and in determining a seasonally correct response to geomagnetic information (Kullberg et al., 2003, 2007; Henshaw et al., 2008). These findings strongly suggest that geomagnetic cues can trigger, advance or delay phases of the migration program, reminiscent of its responsiveness to daylength (Figure 2).
In research on spatial programs, consideration of ontogenetic effects, usually captured by distinguishing naive from experienced migrants, has provided important insights. Whereas the navigational abilities described above hold for all age groups, adherence to the inherited migration program is typically strong in first-time migrants, but may thereafter be supplanted by experience. This has been shown repeatedly in displacement experiments, in which naive migrants followed the blueprint of the inherited program, whereas adults navigated to goal areas they had previously visited. The documented ability to correct for longitudinal displacements of adult birds (Perdeck, 1958; Åkesson et al., 2005; Thorup et al., 2007; Kishkinev et al., 2015), remains to be further explored in juveniles (cf. Åkesson et al., 2005).
Variation in Migration Programs
Variation between individuals and populations in the timing program and its response profile to environmental factors (Figures 2, 3) can take several forms. Individuals can differ from each other in the timing of some phases of the annual cycle, but then resynchronize during a specific phase (see section “Time”). Alternatively, individuals or populations may show shifted timing of the entire annual cycle, as documented for two populations of Collared Flycatchers (Ficedula albicollis) (Briedis et al., 2016). The perhaps most extreme example is the full inversion of the annual cycle in Barn Swallows that colonized the southern hemisphere (Winkler et al., 2017).
Several mechanisms underlie changes in the timing program. If timing is consistent within individuals (i.e., individuals displaying different chronotypes), differences may have a genetic basis. Genetic differences can selectively affect specific phases of the annual cycle. For example, heightened light sensitivity of the reproductive axis (Ramenofsky, 2011; Watts et al., 2018) may advance the timing of spring but not autumn migration, and advanced circannual timing under selection for early breeding occurred specifically in late winter and spring (Helm et al., 2019). Another possibility are epigenetic changes, e.g., methylation of genes involved in biological rhythms (Merlin and Liedvogel, 2019). Ontogenetic effects, such as daylength at birth, affect timing mechanisms in mammals (e.g., Ciarleglio et al., 2011), and may do so also in birds. Additional variation within and between populations may arise from flexible responses to environmental factors that do not change the timing program, but modify its output (see below).
Variation in spatial programs can arise from different sources. Classical studies, notably experiments with European Blackcaps (Sylvia atricapilla) have emphasized genetic determination of directional preference and duration of migratory restlessness (Berthold and Querner, 1981; Berthold et al., 1992; Helbig, 1996). Such polyphenisms are now used in comparative genomic studies aimed at revealing the genetic underpinnings of migration (e.g., Liedvogel et al., 2011; Lundberg et al., 2017). The studies will be important to identify genes involved in encoding variations in space, time and fueling, but also how these genes expressed during migration are regulated.
For individual variation in orientation capacity and compass route-following, the underlying reasons may be related to the perception of the celestial and geomagnetic cues themselves (Muheim et al., 2014), as well as how these cues are encoded in the endogenous migration program. However, we still need to understand exactly how the endogenous migration program interacts with external information, and how birds keep track of space during long migrations throughout the annual cycle. A successful research agenda may be to combine an experimental approach (e.g., Kishkinev et al., 2015; Willemoes et al., 2015; Wikelski et al., 2015; Ilieva et al., 2018) with advanced tracking in the wild (e.g., Willemoes et al., 2014; Bäckman et al., 2017; Sokolovskis et al., 2018; Norevik et al., 2019).
Variation within the population can reveal interesting characteristics, where interactions of the inherited migration phenotype with different environmental factors can lead to the evolution of diverse migration patterns. Phenotypic plasticity may for example lead to advancement of migration timing in response to environmental conditions, in particular in flock-migrating birds (Fraser et al., 2019). Populations may furthermore comprise migratory and resident fractions (i.e., partial migration), or migration may differ between sex and age classes (i.e., differential migration) (Terrill and Able, 1988; Newton, 2008). In partial migration, individual phenotypes may range from completely sedentary to completely migratory (Chapman et al., 2011). Partial and differential migration systems thus enable investigation of effects of selection pressures and fitness consequences of different migration strategies, in particular in long-lived species (Gaillard, 2013; Reid et al., 2018). In different species, this variation can be based on different mechanisms. For example, in European Blackcaps migratory phenotype appears to have a strongly genetic basis (i.e., polyphenism) (Pulido and Berthold, 2010), and in Northern Wheatears (Oenanthe oenanthe; Maggini and Bairlein, 2012) and Dark-eyed Juncos (Junco hyemalis; Holberton, 1993), differences between the sexes are part of the circannual program. In other species, for example Stonechats, differences between migrants and residents are partly environmentally induced (Van Doren et al., 2017). While for most species contributions of genes and environment are unknown, it is likely that both factors are involved.
Mechanistic Integration
Physiological studies, which are beyond the scope of this review, have provided a general picture of the mechanisms of migration programs, although details are still largely unclear. Figure 4 summarizes these findings with an emphasis on timing. It shows schematically how information from the environment affects components of the biological clock that drives the migration program. Spatial cues may also be integrated at this stage. This information is processed in the brain, affects the clock, and prompts a response that is specific to the phase of the annual cycle (Figure 3). Effector systems then fine-tune behavioral and physiological responses by integrating information from within the bird (e.g., its energetic or health state) and from the immediate environment (e.g., weather, food availability) via hormonal pathways (Ramenofsky, 2011; Goymann et al., 2017; Watts et al., 2018). Ultimately, synthetization of this information leads to spatio-temporal migration behavior and physiology. The migration program itself can be modified via genetic change, during ontogeny, and epigenetically.
Programmed Flexibility in Response to Environmental Factors
Within the time window set by the program for migration, decisions about its implementation are sensitive to a range of environmental factors that determine successful migration (Figure 4). Departure time may be adjusted in response to level of fuel reserves, and relative to the expected onward migration route, including distance of barrier crossings (Müller et al., 2018). Responses to these environmental factors are partly inherited, and therefore, we here expand on their effects. We consider some other aspects of flexibility, for example learning, social behavior and responses to weather, to represent residual flexibility.
Adjusting the Drive to Migrate in Response to Food
An important feature of migration is the capacity to prepare for prolonged migratory flights by fueling at stopover sites (Åkesson and Hedenström, 2007). In fact, birds are predicted to spend 1:7 parts of migration time on flight and refueling at stopover, respectively (Hedenström and Alerstam, 1998). Timing and extent of fueling events may be encoded in the endogenous program in relation to expected flight distances (e.g., Fransson et al., 2001; Kullberg et al., 2007), but may also be modified in response to environmental conditions met during flight as well as at stopover sites.
The duration of stop-over, when migrants rest at night, can be predicted by a bird’s body reserves, both in the field (Goymann et al., 2010) and in captivity (Gwinner et al., 1988; Gwinner, 1996a): with increasing reserves, birds are more likely to depart, or to show high levels of Zugunruhe (i.e., migratory drive), respectively. However, this relationship only holds when sufficient local food is available. When food is scarce or inaccessible, birds show the opposite behavior, departing, or showing particularly high Zugunruhe, on low fuel stores (Gwinner et al., 1988). Figure 5 shows how this behavior interacts with the migration program in a series of experiments on garden warblers (Gwinner et al., 1988). In September, during main autumn migration, Zugunruhe was high. When food was temporarily removed, the birds responded by increased night activity (I in Figure 5). On return of ad libitum food, the birds paused Zurunruhe while refeeding (P in Figure 5), and resumed Zugunruhe after regaining body mass (R in Figure 5). In winter, after Zugunruhe had naturally stopped, it was reactivated by food reduction and immediately stopped after food return. These studies show that food availability affects movement decisions of birds during and outside migration seasons, presumably via endocrine pathways (Goymann et al., 2017). The food-induced changes had no effect on the overall migration program of the study birds (Gwinner, 1996a), but likely modify a bird’s actual migration.
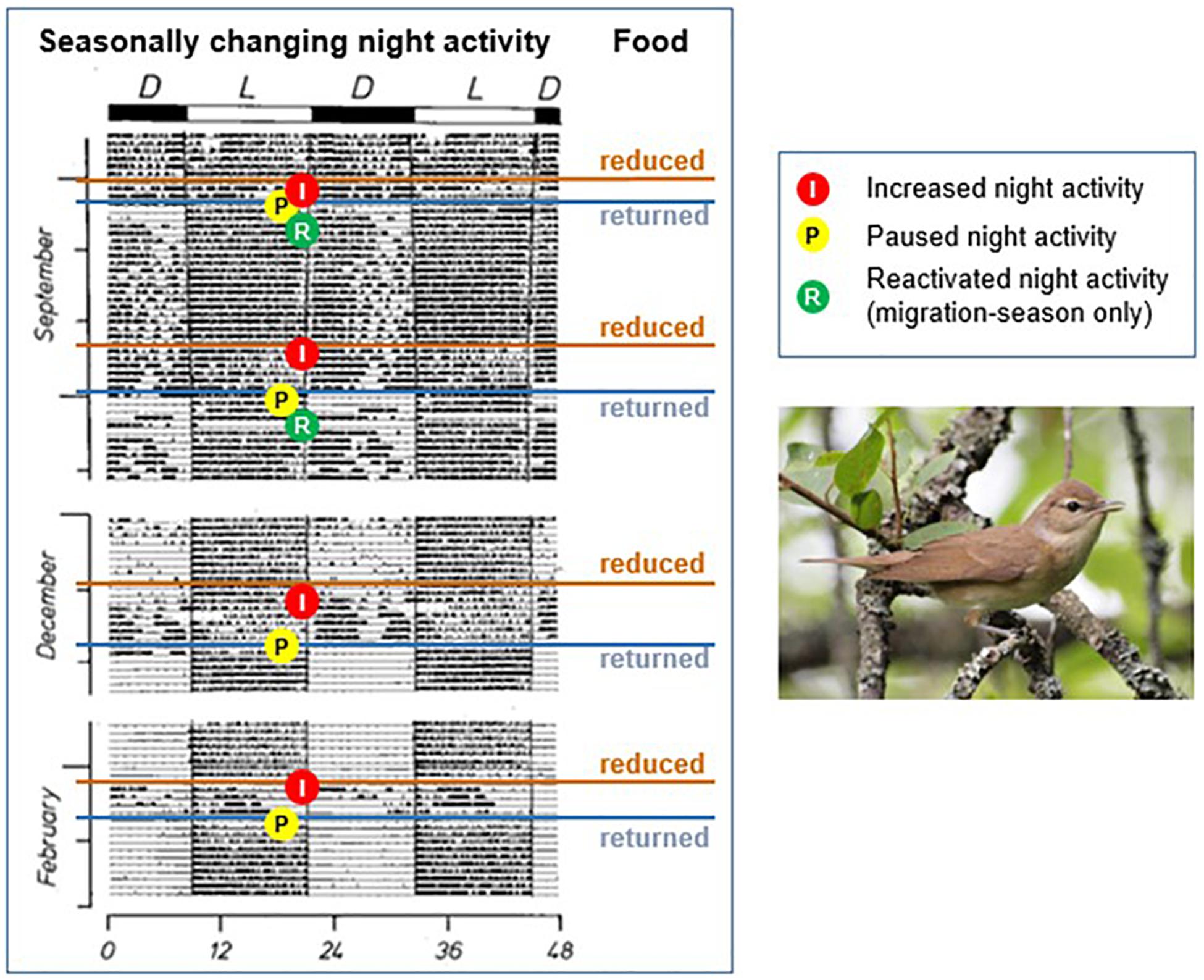
Figure 5. Effects of food on Zugunruhe. Day-night activity shifts of a Garden Warbler in response to experimental changes in food supply. Shown is an actogram which plots activity of the bird (black marks) against time of day (x-axis) for each day of the experiment, represented by a line. Data are shown for two consecutive days: the second day is repeated as the first day on the successive line; actogram reproduced with kind permission of Oecologia. Inlay: image of Garden Warbler from Billyboy, Sweden, wikimedia.org; based on Gwinner et al. (1988).
Adjusting the Drive to Migrate and Fueling in Response to Geomagnetic Cues
Migratory restlessness can be modified by exposure to geomagnetic field parameters expected to be met en route or at destination areas. This insight, which was previously shown for nocturnally migrating Northern Wheatears (Bulte et al., 2017), was recently also experimentally revealed for diurnally migrating Dunnocks (Prunella modularis) (Ilieva et al., 2018). Migratory restlessness was recorded for individual Dunnocks during a 2-week period, during which one group was kept in the local Swedish geomagnetic field, while two other groups were geomagnetically displaced north (away from wintering area) or south (toward the wintering area in southern France) (Ilieva et al., 2018). The birds showed two peaks of activity throughout the 24 h-cycle, with the longest peak in the morning, associated with migration, and a shorter evening peak associated primarily with feeding (Ilieva et al., 2018). The Dunnocks displaced south reduced the morning migratory restlessness as they were exposed to the geomagnetic parameters, i.e., inclination angle and total field intensity, at the wintering area, while the control birds instead increased the migratory restlessness over the study period (Figure 6; Ilieva et al., 2018). The northern displacement resulted in continued, but somewhat reduced migration activity, suggesting it was not only the magnetic change itself, but also the characteristics of the magnetic parameters (i.e., combination of inclination and total field intensity at expected destination area), that interacted with the endogenous program resulting in reduced migratory restlessness as the winter destination was geomagnetically reached (Ilieva et al., 2018). Like food availability, geomagnetic cues in this example appeared to modulate the output of the migration program, but at least within the study period, not the time course of it.
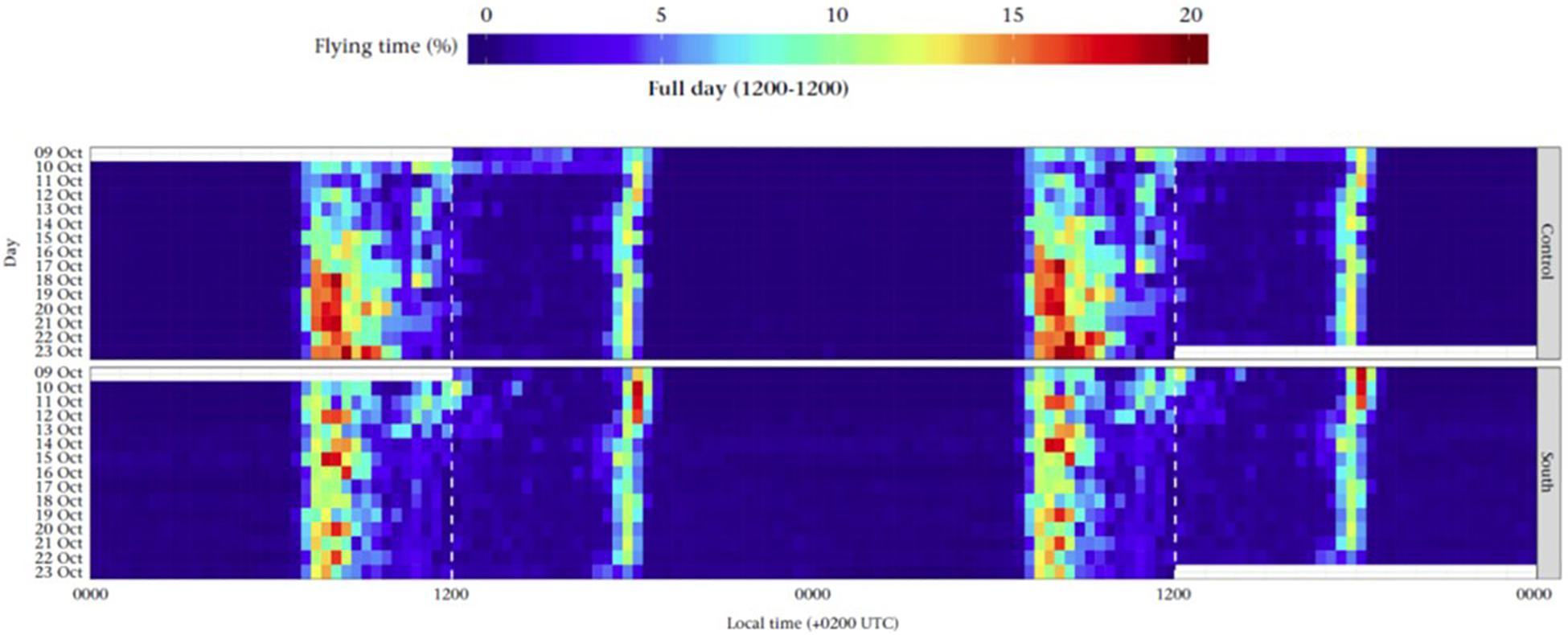
Figure 6. Effects of geomagnetic cues on Zugunruhe. Actograms for first-year migratory Dunnocks monitored from 9 to 24 October 2015 under the local geomagnetic condition (control, top) and under simulated geomagnetic conditions (south displacements, bottom). Each horizontal line shows the mean time spent in flying mode by eight birds per group; days are duplicated for easier viewing as in Figure 5 (except for first and last half-days). Dashed vertical lines indicate 12.00 h local time (14.00 UTC), that is, the time when geomagnetic displacements took place during the first 5 days of the experiment. Figure modified after Ilieva et al. (2018). Actogram reproduced with kind permission from Animal Behaviour.
Adjusting Directions in Response to Geomagnetic Cues
Geomagnetic information can also modify the directional output of the migration program, but its effect differs between species. It is not completely understood why some species express a directional shift encoded with time as the migration season progresses (Gwinner and Wiltschko, 1978), while orientation shifts by other species are only expressed by exposure to changes of geomagnetic information (Beck and Wiltschko, 1988). To understand this we see a need for further studies of different bird species under controlled environmental conditions including magnetic displacements (e.g., Kishkinev et al., 2015).
Feed-back from geomagnetic information helps migratory birds meet a further challenge. Trans-hemispheric long-distance migrants are crossing the geomagnetic equator, which involves a 180°-shift of the angle of inclination, a key-feature of the birds’ magnetic inclination compass (Wiltschko and Wiltschko, 1972). Cage experiments with two long-distance migrating songbirds, the European Garden Warbler and the North American Bobolink (Dolichonyx oryzivorus), have revealed that these birds possess inherited responses where they adaptively change their preferred orientation with respect to the inclination angle, as they are exposed to a horizontal magnetic field simulating a magnetic equator crossing (Beason, 1992; Wiltschko and Wiltschko, 1992). It would be interesting to investigate if this response to magnetic inclination by shifting courses is present in most avian migrants, or if it is characteristic for the long-distance migrants adapted to trans-hemisphere flights.
Once terrestrial birds have initiated migration they may cross landmasses, but also barriers such as seas, mountains and deserts, where they may be unable to land. The inhospitable terrain may challenge their migration performance and stopover use during migration, leading to special adaptations (Åkesson and Hedenström, 2007). At coastal sites, in particular, young birds may hesitate to continue on a sea-crossing and are grounded in large numbers. They may then search for foraging sites and shelter (Alerstam, 1978), and perform reverse migration to inland sites before they continue in the migration direction several days later (e.g., Åkesson et al., 1996b). Temporary reverse migration is typically expressed near coastal barriers leading to more suitable stop-over sites (Åkesson et al., 1996b; Zehnder et al., 2002; Buler and Moore, 2011), whereas at inland locations reverse migration is less common (Åkesson, 1999; Komenda-Zehnder et al., 2002). Temporary reverse migration and movements to nearby stopover sites are predominantly found in birds with low fuel reserves (Åkesson et al., 1996b; Sandberg, 2003; Covino et al., 2015), and its directions are expressed in relation to the geomagnetic field (Sandberg, 1994; Bäckman et al., 1997). Thus, as shown for effects of food shortage and magnetic cues on migratory drive, adaptive responses to barriers seem to be embedded in the migration program.
Flexibility Residual to the Migration Program
Some aspects of flexibility have no clear relationship with migration programs, or appear to contradict, override or supplant them. Major effects are exerted by weather and availability of favorable winds (e.g., Åkesson and Hedenström, 2000; Åkesson et al., 2002; Shamoun-Baranes et al., 2007; Eikenaar and Schmaljohann, 2015; Sjöberg et al., 2015; Van Doren and Horton, 2018). Availability of celestial compass cues (Åkesson et al., 1996a, 2001b) and locations of suitable stopover sites may furthermore have a strong effect on individual route choices when crossing large barriers (Åkesson et al., 2016). A further contributing factor are social effects on migration (Helm et al., 2006). This underrated factor is beginning to be addressed by exciting new data from tracking studies. For example, a recent study on European Bee-Eaters (Merops apiaster) presented migration data from 29 individuals moving in different groups (Dhanjal-Adams et al., 2018). Timing within groups was closely coordinated, and once separated, groups rejoined each other within days, even after a transcontinental journey. These data indicate both strong social effects and a detailed migration program, which could be innate, learned, or resource driven.
Learning and development are forms of flexibility that are still poorly known. For most birds it is unknown how migration performance develops with time. There are examples where adult birds have been shown to migrate faster (Ellegren, 1993), cover longer flight paths per day (Weimerskirch et al., 2006), handle wind drift more efficiently (Thorup et al., 2003), follow shorter routes and have shorter stopover times (Crysler et al., 2016) than juvenile birds. A recent example from a long-term study documents all these parameters for individually tracked migratory Black Kites (Milvus migrans; Figure 7A) across a time span of 1-27 years (Sergio et al., 2014). This study shows that the development of migratory behavior in young birds follows a consistent trajectory, and that the development is more gradual and prolonged than previously assumed (Figures 7B,C). More efficient migration performance was further shown to be promoted by a combination of individual improvement across time and selective mortality occurring most frequently in early phases of life during the pre-breeding migration period (Sergio et al., 2014). Several migration components improved across time, including increased migration speed, shorter stopover time, and increased efficiency to handle cross-winds in adult birds as compared to juveniles (Sergio et al., 2014). The strongest selection occurred on the flanks of the distribution during the early stages of life. Individuals that were able to improve their ability to handle environmental conditions efficiently on migration and to depart progressively earlier than conspecifics, obtained higher breeding and survival rates, leading to a longer life span (Figure 7C).
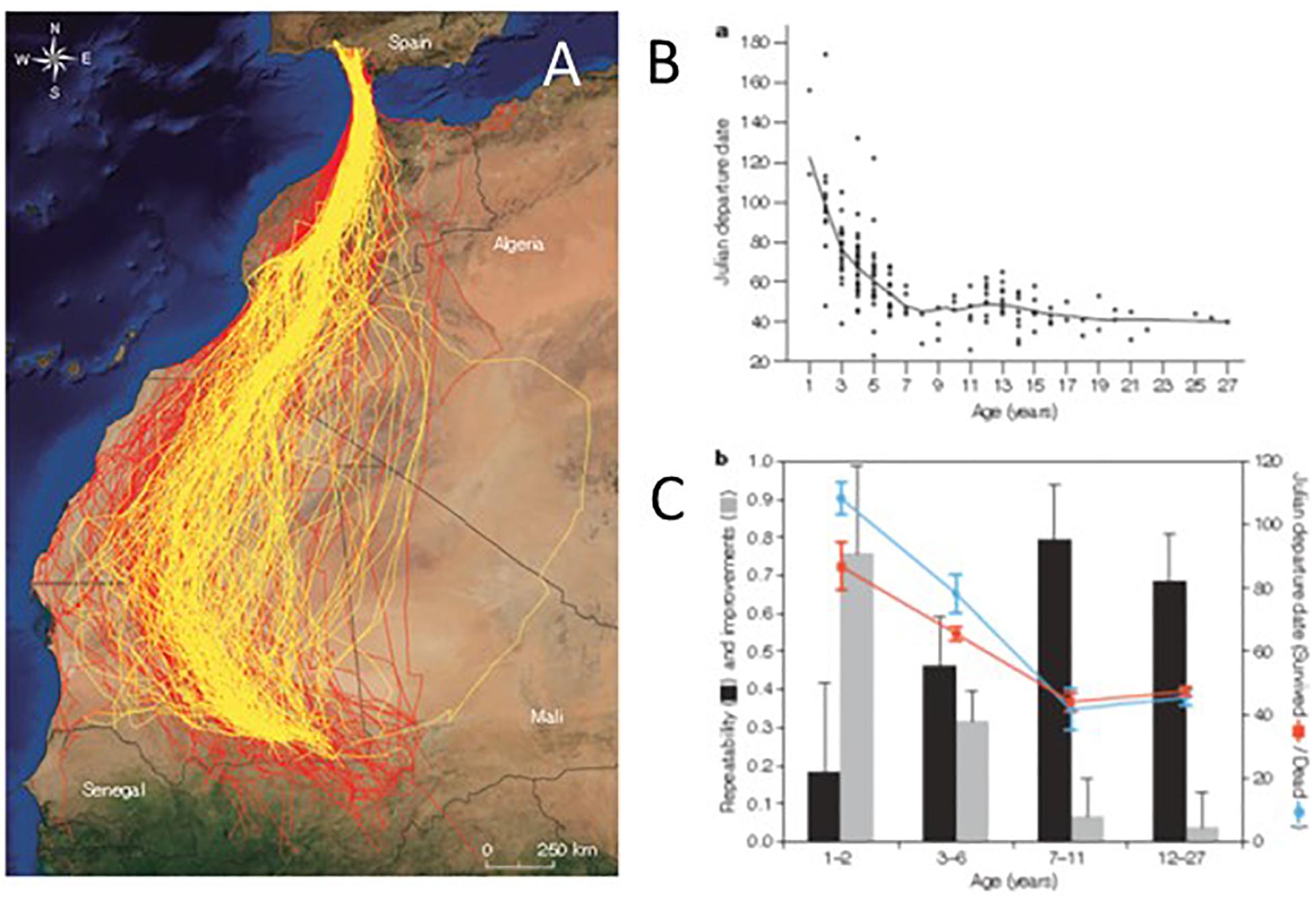
Figure 7. Migration performance and route choices across time for migratory Black Kites tracked for 1–27 years. (A) Migration routes of individually tracked Black Kites from breeding sites in southwestern Spain to wintering areas in West Africa. Pre-breeding tracks are shown in red and post-breeding tracks in yellow. (B) Pre-breeding migration departure date in relation to Julian date advanced with age. (C) Migration performance across and within individuals, illustrating that pre-breeding departure date improved rapidly during the first 7 years of life and then reached a plateau. In the initial years birds that survived within the next year (red) departed earlier (right axis) than those that died (blue). Similarly, within individuals, the repeatability of departure (black bars, left axis) was lowest in the initial years of life, when individual improvements (gray bars, left axis) were highest, and stabilized after birds were 7 years old. The cross-sectional pattern depicted in (B) was concluded by Sergio et al. (2014) to be consistent with both within-individual improvements and selective removal of inferior performers. Figures from Sergio et al. (2014), reprinted by permission from Springer, Nature.
Residual flexibility can either increase, or decrease, variation between individuals in time and space. Tracking data show that in some species delays in timing of breeding and migration departure may be difficult to correct for at later stages of the annual cycle. For instance, migratory Eurasian Nightjars (Caprimulgus caprimulgus) tracked by geolocators reveal that individuals being late in their departure timing in autumn, will continue to be late throughout the annual cycle, while early birds continue to be early (Norevik et al., 2017; Figure 8). It is currently unknown whether these effects persisted beyond the duration of the annual cycle, or whether birds were eventually synchronized, as discussed above (see section “Time”). If differences indeed persisted, they could partly reflect individual differences in chronotype (i.e., polyphenism).
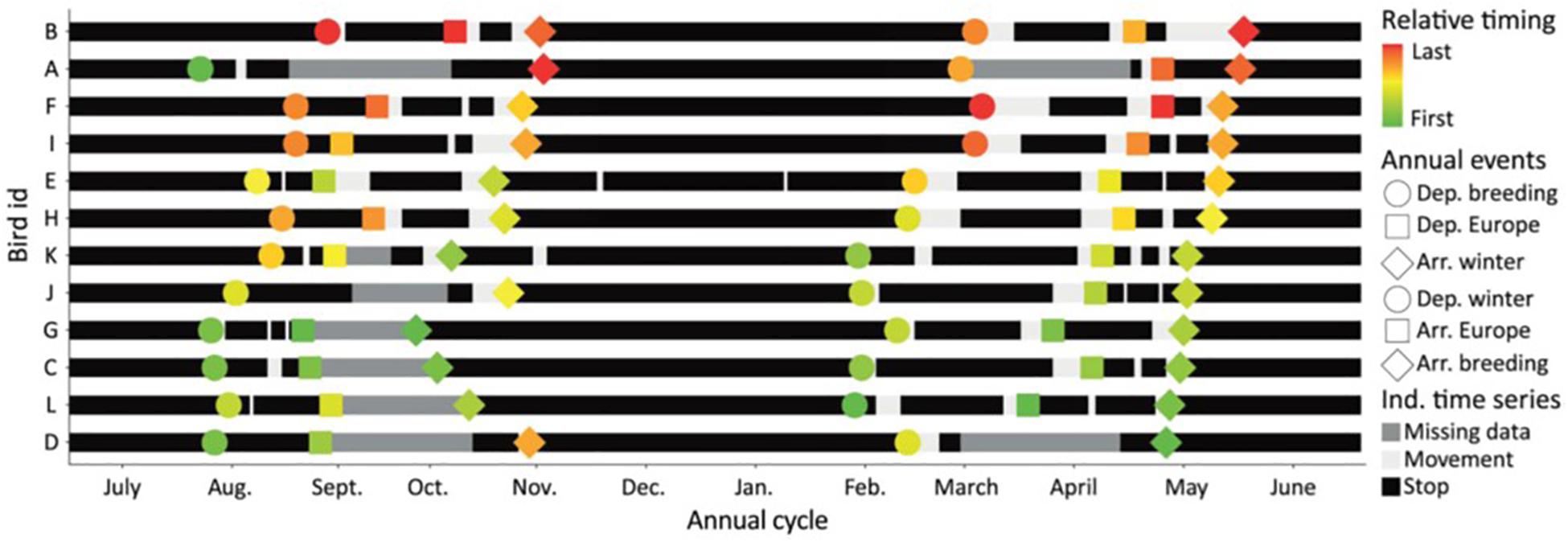
Figure 8. Carry-over effects across the annual cycle for migratory adult European Nightjars. A graphical presentation of distributions of stops and movements in the annual cycle for European Nightjars tracked by microdataloggers, plotted with respect to starting date 1 July. Black bars represent stationary periods and the pale gaps show time of movement. Gray sections refer to periods for which occurrence of stops could not be resolved. The dots show the individual’s timing of the six distinct annual events as shown in inset, and the color gradient shows the order of the birds in each event. Green illustrate the first bird and red the last bird, sorted by the date of arrival to the breeding area. From Norevik et al. (2017).
Illustrating Extremes of Variation in Inherited Spatiotemporal Behavior
The wide range of mechanisms reviewed above can produce highly divergent outcomes. Below, we use extremes of variation within populations, ranging from great consistency to massive differentiation, to illustrate the many axes of variation in migration programs.
Low Within-Population Variation in Cuckoos and Willow Warblers
An example of limited inter-individual variation of a rather complex migration and stopover program comes from satellite tracking of the Common Cuckoo (Cuculus canorus). Cuckoos use a largely fixed sequence of stopover sites and route directions throughout the non-breeding period (Willemoes et al., 2014; Hewson et al., 2016; Figure 9A). Initially the adult birds tracked from northwestern Europe depart toward the southeast in autumn, and stopover in central northern and thereafter southeastern Europe, before they initiate the Sahara crossing on southerly courses (Figure 9A). After the barrier crossing they make a prolonged stopover in eastern Sahel (Willemoes et al., 2014; Hewson et al., 2016; Figure 9A). Later in the season they will proceed to wintering areas further to the south, from where they will initiate northerly movements, following a loop migration in spring via West Africa (Willemoes et al., 2014; Hewson et al., 2016; Figure 9A). The Cuckoo’s somewhat stereotypic sequence of flight steps and stopovers during the non-breeding period is expressed timely with the availability of food resources in the areas visited, and encoded in the endogenous program (Willemoes et al., 2014). The phenotypic expression of this complex spatio-temporal program, and apparent use of goal areas throughout the non-breeding period, open up questions on what specific information is used to identify those areas throughout the annual cycle, and how potentially external information may interact with the migration decisions and route choices. In a displacement experiment with adult cuckoos during autumn migration Willemoes et al. (2015) investigated the capacity to return to the normal migration route, stopover and wintering areas. The displaced adult cuckoos showed some individual variation in the strategy they used to return, but also a capacity to navigate across areas potentially new to them (Willemoes et al., 2015). Juvenile cuckoos have further been shown to migrate later, show larger scatter in route choice and perform migration at slower pace than adults, but still they have been confirmed to reach the expected wintering areas by following their endogenous migration program (Vega et al., 2016).
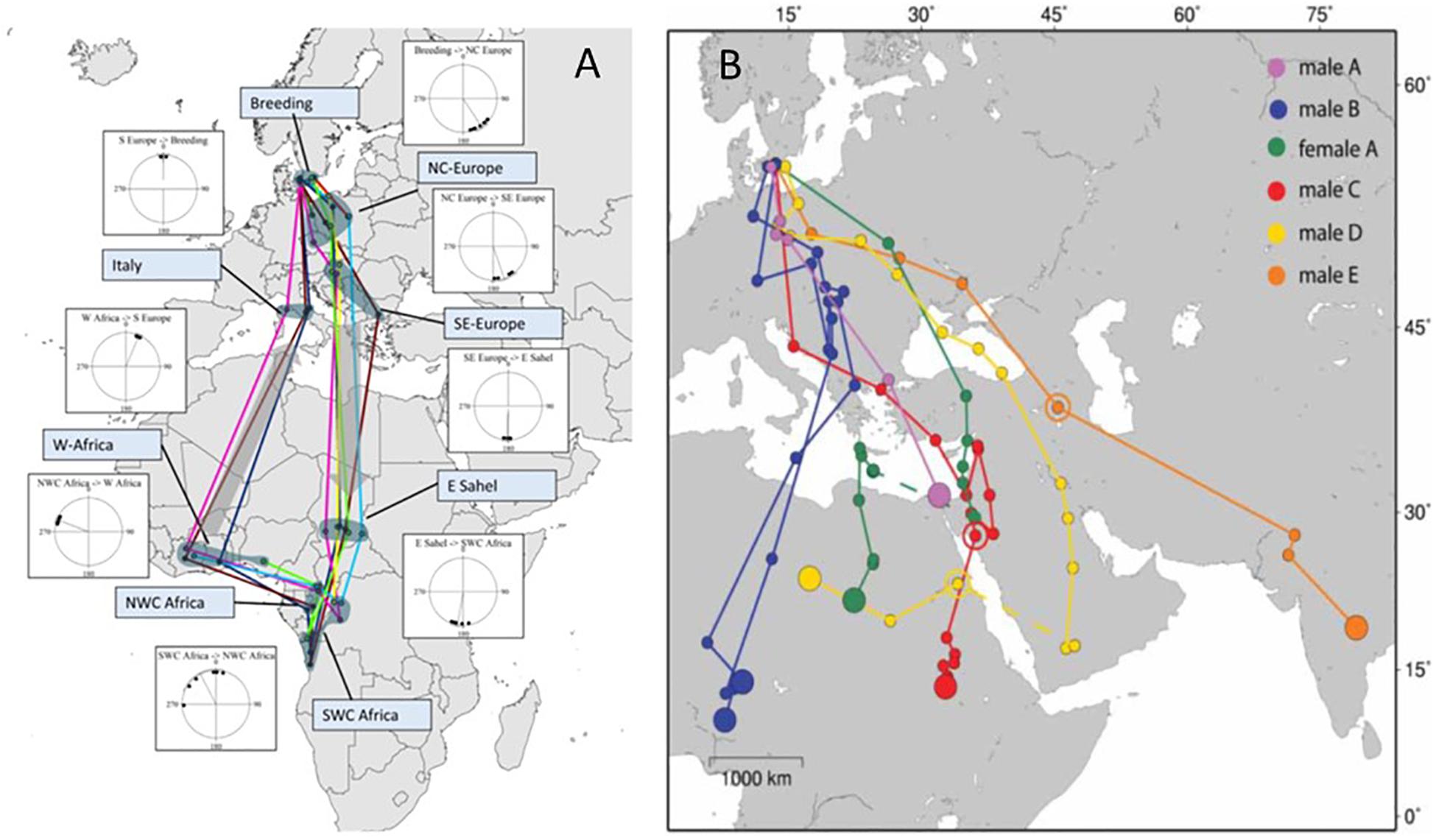
Figure 9. Examples of contrasting variation in migration routes. (A) Common Cuckoo, (B) Little Ringed Plover. (A) Staging areas of eight satellite-tracked adult Common Cuckoos with vector directions between stopovers indicated by inserted orientation diagrams. Lines are connecting staging sites and do not necessarily represent the paths followed. From Willemoes et al. (2014). (B) Autumn migration tracks of Little Ringed Plovers as revealed by geolocators. Filled small circles show three-day means of positional data and filled large circles are mean location for winter positions. Open circles indicate the location of a stopover period. Broken lines indicate unknown movement around the autumn equinox. Note that one individual was tracked for two consecutive migrations using different geolocators (male B). From Hedenström et al. (2013). Maps in Mercator projection.
Another species with highly conserved migration routes are Willow Warblers (Phylloscopus trochilus) (Figure 10). However, these routes are specific to subspecies (P. t. trochilus, acredula, yakutensis) within the species’ vast breeding range in Northern Europe and Asia. Willow warblers, thus, represent an interesting example where the evolution of migration routes, speciation and range expansion may be investigated (Bensch et al., 2009; Lundberg et al., 2017). Thanks to miniaturization of tracking technology we are now able to document migration behavior in these smallest of songbirds from different parts of their range (Lerche-Jørgensen et al., 2017; Sokolovskis et al., 2018; Figure 10), which opens up the possibility to record route constancy and migration timing for birds with known, locally differing, genotypes (Lundberg et al., 2017). Special attention is given to hybrid zones where individuals with different endogenous migration programs breed side by side and may cross-breed, possibly resulting in intermediate directions in hybrids (Helbig, 1991; Delmore and Irwin, 2014). The consistent, but locally differentiated routes of Willow Warblers, thus offer great opportunities for detailed studies of the genetic program encoding migration behavior, and the phenotypic expression of it (Bensch et al., 2009; Ruegg et al., 2014).
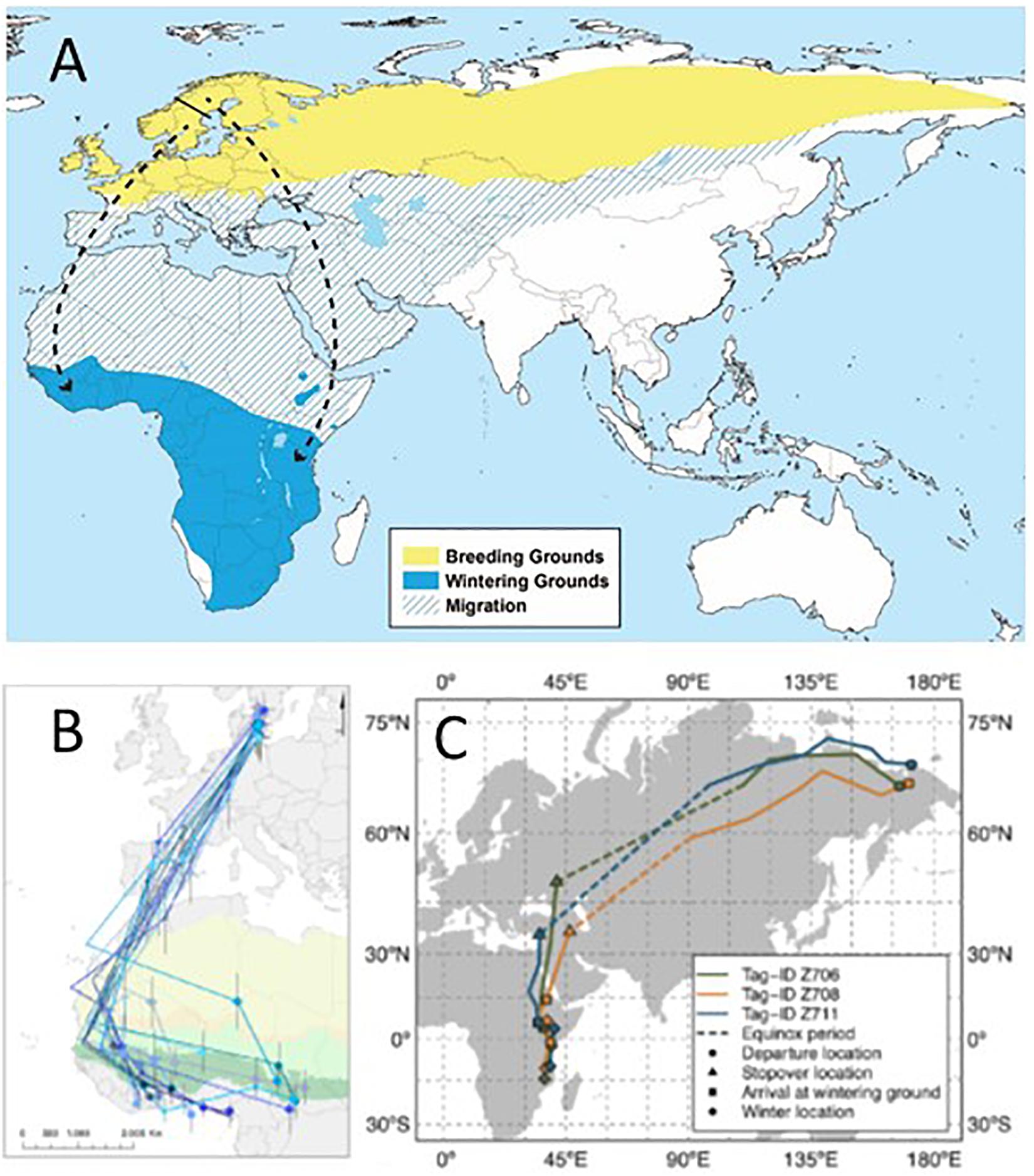
Figure 10. Consistent, population-specific migration routes of Willow Warblers. (A) Worldwide distribution of Willow Warblers. Breeding (yellow) and wintering (blue) ranges and occurrence during migration (hatched). Arrows (broken black) indicate the expected migratory routes of Scandinavian southwest-migrating trochilus and southeast-migrating acredula Willow Warblers, and their approximate initial wintering areas in sub-Saharan Africa. Black bar indicate location of hybrid zone between acredula (north) and trochilus (south) in Scandinavia (Bensch et al., 2009; map courtesy Keith W. Larson, Sweden; Wikimedia.org). (B) Migration of southwest-migrating trochilus Willow Warblers from breeding to wintering grounds (individuals represented by different colors; from Lerche-Jørgensen et al., 2017). (C) Migration of P. t. yakutensis tracked by geolocation from breeding sites in Far East Russia to initial wintering areas in East Africa. From Sokolovskis et al. (2018).
High Within-Population Variation in Plovers and Albatrosses
Conversely, exceptional variation in migration directions, route choice and wintering areas is exemplified within one single study population of the Little Ringed Plover (Charadrius dubius) (Hedenström et al., 2013). In this population, birds breeding side by side within the same gravel pit may migrate to wintering areas from Nigeria in the west to India in the east (Hedenström et al., 2013; Figure 9B). The extreme variation in inherited direction and longitudinal range of wintering area selection call for further understanding of the genetics behind the phenotypic expression of the migration program in birds like the Little Ringed Plover.
Another example for exceptionally high variation comes from partially migratory Wandering Albatross (Diomedea exulans), whose behavior differs from conventional migration systems (Figure 11). The Wandering Albatross is known for its long lifespan, oceanic lifestyle and for breeding on isolated sub-Antarctic Islands. Between reproductive events it spends a sabbatical year at sea before returning to its previous breeding island (Tickell, 1968; Weimerskirch and Wilson, 2000). In this species, differential non-breeding movement strategies have evolved in populations with limited genetic differentiation (Milot et al., 2008). Birds from different breeding colonies may predominantly move to different ocean areas (Weimerskirch et al., 2015; Figure 11). Young wandering albatrosses have further been shown to follow similar routes as adults during their first migration and to spend their first year in sex-specific ocean areas overlapping with the adults, with males moving over twice the distance of females, demonstrating endogenous control of area use (Åkesson and Weimerskirch, 2014).
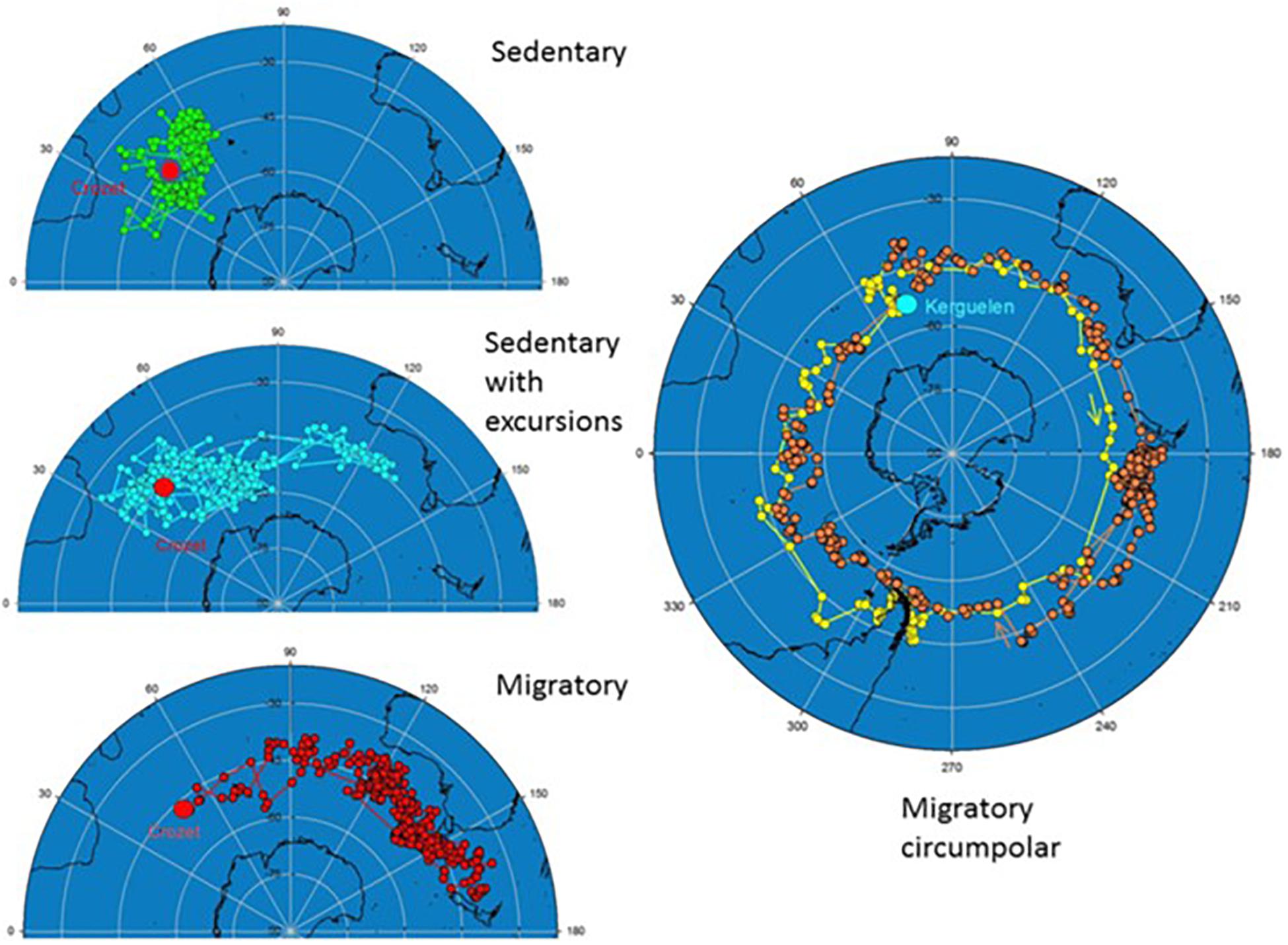
Figure 11. Example tracks for adult Wandering Albatrosses recorded during sabbatical years representing the main movement strategies for birds breeding at Crozet and Kergulean Islands. Movement strategies are sedentary, sedentary with distant excursions of two Crozet birds, and migratory. The full cycle shows extremely long distance migration including two consecutive circumpolar movements of an adult from Kergulean Islands. After an initial rapid flight to Chilean waters where the bird spent 2 months, it moved eastward through the Atlantic and the Indian Ocean (yellow) to reach the Chatham Rise, east of New Zealand, before returning to Kerguelen (orange) through the Pacific and Atlantic. Figures from Weimerskirch et al. (2015), reprinted by permission from Springer Nature//Palgrave.
During the sabbatical year, adult wandering albatrosses follow three alternative movement strategies: sedentary, sedentary with excursions, or migratory (Weimerskirch et al., 2015; Figure 11). The proportion of birds adhering to the different strategies may differ between colonies. Wandering Albatrosses at Kergulean Island are all migratory, while the sedentary strategy is present in Crozet Island birds (Weimerskirch et al., 2015; Figure 11). The migratory strategy is furthermore more commonly used by males than by females (Weimerskirch et al., 2015), and differentially expressed already in young birds (Åkesson and Weimerskirch, 2014). A sedentary strategy, predominantly used by females at Crozet Island, has increased in recent years (Weimerskirch et al., 2015), possibly as a consequence of climate change (Fryxell and Holt, 2013), revealing a potential to adapt to new environmental conditions by phenotypic flexibility or as a consequence of natural selection. A long-term population change in movement strategies including settling down, may in turn lead to bird diversification and speciation as suggested by Rolland et al. (2014).
Conclusion and Outlook
In this review, we have shown how in bird migration, inherited programs and responses to the environment interact. As in other fields of biology, it is time to leave behind old dichotomies between genetics and physiology on the one hand, and ecology on the other. Evolution has brought about an impressive range of solutions to the problem that migration requires both predictive anticipation and flexibility (Figures 9, 11). For some aspects of migration, we begin to understand the birds’ flexibility on the basis of inherited reaction norms that provide solutions which were effective over evolutionary time. These insights refine views of environmental effects on migration, as being dependent on the phase of the annual cycle and a bird’s migration program (Figures 3, 5). They also refine views of carry-over effects as being at least in part permitted, or counter-acted, by migration programs (Figure 2). The combined developments of molecular tools and tracking technology, if applied to rewarding model systems (e.g., Figure 10), are set to greatly foster this understanding. For some other aspects, we currently do not know how decisions are made, and how migrants improve by learning. The study on Black Kites (Figure 7) is one of the most informative about the development of migration performance in individual birds, but calls for follow-up studies on other species (Campioni et al., 2020), in particular those relying strongly on an endogenous program for their first migration. Likewise, the cited study on European Bee-Eaters (Dhanjal-Adams et al., 2018) offers exciting insights on migration of highly social species, which will hopefully be complemented by work on species with different social systems. In addition, recent findings from the European Nightjar call attention to yet another temporal domain that may have been largely overlooked, at least in landbirds. Nocturnally migratory nightjars time fueling and migration events to the different phases of the moon (Norevik et al., 2019), and thereby call for further studies of moon cycle effects on migratory birds, ideally with a circannual perspective (Chapin and Wing, 1959; Cruz et al., 2013).
Ultimately, combined ecological and mechanistic studies may explain why some species strictly adhere to spatio-temporal programs whereas others are flexible, and how selection pressures, from competitive temporal niche segregation (Heim et al., 2018) to climate change, may shape migrations in the future. We hope that our overview will encourage future cross-over between approaches that focus on annual-cycle programs and those that focus on the environment, as jointly these components enable migratory birds to carry out their great journeys.
Author Contributions
Both authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Able, K. P. (1980). “Mechanisms of orientation, navigation and homing,” in Animal Migration, Orientation and Navigation, ed. S. A. Gauthreaux (New York, NY: Academic Press), 283–373. doi: 10.1016/b978-0-08-091833-4.50010-9
Åkesson, S. (1999). Do passerine migrants captured at an inland site perform temporary reverse migration in autumn? Ardea 87, 129–137.
Åkesson, S., Alerstam, T., and Hedenström, A. (1996a). Flight initiation of nocturnal passerine migrants in relation to celestial orientation conditions at twilight. J. Avian Biol. 27, 95–102.
Åkesson, S., and Bianco, G. (2016). Assessing vector navigation in long-distance migrating birds. Behav. Ecol. 27, 865–875. doi: 10.1093/beheco/arv231
Åkesson, S., and Bianco, G. (2017). Route simulations, compass mechanisms and long-distance migration flights in birds. J. Comp. Physiol. A 203, 475–490. doi: 10.1007/s00359-017-1171-y
Åkesson, S., Bianco, G., and Hedenström, A. (2016). Negotiating an ecological barrier: crossing the Sahara in relation to winds by common swifts. Philos. Trans. R. Soc. B 371:20150393. doi: 10.1098/rstb.2015.0393
Åkesson, S., Boström, J., Liedvogel, M., and Muheim, R. (2014). “Animal navigation,” in Animal Movement Across Scales, eds L.-A. Hansson and S. Åkesson (Oxford: Oxford University Press), 151–178.
Åkesson, S., and Hedenström, A. (2000). Wind selectivity of migratory flight departures in birds. Behav. Ecol. Sociobiol. 47, 140–144. doi: 10.1007/s002650050004
Åkesson, S., and Hedenström, A. (2007). How migrants get there: migratory performance and orientation. Bioscience 57, 123–133. doi: 10.1641/b570207
Åkesson, S., Ilieva, M., Karagicheva, J., Rakhimberdiev, E., Tomotani, B., and Helm, B. (2017). Timing avian long-distance migration: from internal clock mechanisms to global flights. Philos. Trans. R. Soc. B 372:20160252. doi: 10.1098/rstb.2016.0252
Åkesson, S., Karlsson, L., Walinder, G., and Alerstam, T. (1996b). Bimodal orientation and the occurrence of temporary reverse bird migration during autumn in south Scandinavia. Behav. Ecol. Sociobiol. 38, 293–302. doi: 10.1007/s002650050245
Åkesson, S., Klaassen, R., Holmgren, J., Fox, J. W., and Hedenström, A. (2012). Migration routes and strategies in a highly aerial migrant, the common swift Apus apus, revealed by light-level geolocators. PLoS One 7:e41195. doi: 10.1371/journal.pone.0041195
Åkesson, S., Morin, J., Muheim, R., and Ottosson, U. (2001a). Avian orientation at steep angles of inclination: experiments with migratory white–crowned sparrows at the magnetic North Pole. Proc. Biol. Sci. 268, 1907–1913. doi: 10.1098/rspb.2001.1736
Åkesson, S., Walinder, G., Karlsson, L., and Ehnbom, S. (2001b). Reed warbler orientation: initiation of nocturnal migratory flights in relation to visibility of celestial cues at dusk. Anim. Behav. 61, 181–189. doi: 10.1006/anbe.2000.1562
Åkesson, S., Morin, J., Muheim, R., and Ottosson, U. (2005). Dramatic orientation shift of white-crowned sparrows displaced across longitudes in the high arctic. Curr. Biol. 15, 1591–1597. doi: 10.1016/j.cub.2005.07.027
Åkesson, S., Odin, C., Hegedüs, R., Ilieva, M., Sjöholm, C., Farkas, A., et al. (2015). Testing avian compass calibration: comparative experiments with diurnal and nocturnal passerine migrants in South Sweden. Biol. Open 4, 35–47. doi: 10.1242/bio.20149837
Åkesson, S., Walinder, G., Karlsson, L., and Ehnbom, S. (2002). Nocturnal migratory flight initiation in reed warblers Acrocephalus scirpaceus: effect of wind on orientation and timing of migration. J. Avian Biol. 33, 349–357. doi: 10.1034/j.1600-048x.2002.02951.x
Åkesson, S., and Weimerskirch, H. (2014). Evidence for sex-segregated ocean distributions of first-winter wandering albatrosses at Crozet Islands. PLoS One 9:e86779. doi: 10.1371/journal.pone.0086779
Alerstam, T. (1978). Reoriented bird migration in coastal areas: dispersal to suitable resting grounds? Oikos 30, 405–408.
Alerstam, T., Gudmundsson, G. A., Green, M., and Hedenström, A. (2001). Migration along orthodromic sun compass routes by arctic birds. Science 291, 300–303. doi: 10.1126/science.291.5502.300
Alerstam, T., and Lindström, Å. (1990). “Optimal bird migration: the relative importance of time, energy, and safety,” in Bird Migration, ed. E. Gwinner (Berlin: Springer), 331–351. doi: 10.1007/978-3-642-74542-3_22
Alerstam, T., and Pettersson, S. G. (1991). Orientation along great circles by migrating birds using a sun compass. J. Theor. Biol. 152, 191–202. doi: 10.1016/s0022-5193(05)80452-7
Altshuler, D. L., Cockle, K. L., and Boyle, W. A. (2013). North American ornithology in transition. Biol. Lett. 9:20120876. doi: 10.1098/rsbl.2012.0876
Bäckman, J., Andersson, A., Alerstam, T., Pedersen, L., Sjöberg, S., Thorup, K., et al. (2017). Activity and migratory flights of individual free-flying songbirds throughout the annual cycle: method and first case study. J. Avian Biol. 48, 309–319. doi: 10.1111/jav.01068
Bäckman, J., Pettersson, J., and Sandberg, R. (1997). The influence of fat stores on magnetic orientation in day-migrating Chaffinch, Fringilla coelebs. Ethology 103, 247–256. doi: 10.1111/j.1439-0310.1997.tb00120.x
Barlow, R. B., Powers, M. K., Howard, H., and Kass, L. (1986). Migration of limulus for mating: relation to lunar phase, tide height, and sunlight. Biol. Bull. 171, 310–329. doi: 10.2307/1541674
Bartell, P. A., and Gwinner, E. (2005). A separate circadian oscillator controls nocturnal migratory restlessness in the songbird Sylvia borin. J. Biol. Rhythms 20, 538–549. doi: 10.1177/0748730405281826
Beason, R. C. (1992). You can get there from here: responses to simulated magnetic equator crossing by the bobolink (Dolichonyx oryzivorus). Ethology 91, 75–80. doi: 10.1111/j.1439-0310.1992.tb00852.x
Beck, W., and Wiltschko, W. (1982). The magnetic field as a reference system for genetically encoded migratory direction in pied flycatchers (Ficedula hypoleuca Pallas). Z. Tierpsychol. 60, 41–46. doi: 10.1111/j.1439-0310.1982.tb01075.x
Beck, W., and Wiltschko, W. (1988). “Magnetic factors control the migratory direction of pied flycatchers (Ficedula hypoleuca, Pallas),” in Acta XX Congressus Internationalis Ornithologici, Vol. 1986, ed. H. Ouellet (Ottawa: University of Ottawa Press), 1955–1962.
Bensch, S., Grahn, M., Müller, N., Gay, L., and Åkesson, S. (2009). Genetic, morphological, and feather isotope variation of migratory willow warblers show gradual divergence in a ring. Mol. Ecol. 18, 3087–3096. doi: 10.1111/j.1365-294X.2009.04210.x
Berthold, P. (1973). Relationships between migratory restlessness and migration distance in six Sylvia species. Ibis 115, 594–599. doi: 10.1111/j.1474-919x.1973.tb01998.x
Berthold, P., Helbig, A. J., Mohr, G., and Querner, U. (1992). Rapid microevolution of migratory behaviour in a wild bird species. Nature 360, 668–670. doi: 10.1038/360668a0
Berthold, P., Kaatz, M., and Querner, U. (2004). Long-term satellite tracking of white stork (Ciconia ciconia) migration: constancy versus variability. J. Ornithol. 145, 356–359. doi: 10.1007/s10336-004-0049-2
Berthold, P., and Querner, U. (1981). Genetic basis of migratory behavior in European warblers. Science 212, 77–79. doi: 10.1126/science.212.4490.77
Bertin, A., Houdelier, C., Richard-Yris, M.-A., Guyomarc’h, C., and Lumineau, S. (2007). Stable individual profiles of daily timing of migratory restlessness in European quail. Chronobiol. Int. 24, 253–267. doi: 10.1080/07420520701283685
Birkhead, T. R. (2008). The Wisdom of Birds. An Illustrated History of Ornithology. London: Bloomsbury.
Bojarinova, J., and Babushkina, O. (2015). Photoperiodic conditions affect the level of locomotory activity during autumn migration in the Long-tailed Tit (Aegithalos c. caudatus). Auk 132, 370–379. doi: 10.1642/auk-14-155.1
Briedis, M., Hahn, S., Gustafsson, L., Henshaw, I., Träff, J., Král, M., et al. (2016). Breeding latitude leads to different temporal but not spatial organization of the annual cycle in a long-distance migrant. J. Avian Biol. 47, 743–748. doi: 10.1111/jav.01002
Briedis, M., Krist, M., Král, M., Voigt, C. C., and Adamík, P. (2018). Linking events throughout the annual cycle in a migratory bird—non-breeding period buffers accumulation of carry-over effects. Behav. Ecol. Sociobiol. 72:93. doi: 10.1093/oso/9780198824268.003.0008
Budki, P., Rani, S., and Kumar, V. (2009). Food deprivation during photosensitive and photorefractory life-history stages affects the reproductive cycle in the migratory Red-headed Bunting (Emberiza bruniceps). J. Exp. Biol. 212, 225–230. doi: 10.1242/jeb.024190
Buler, J. J., and Moore, F. R. (2011). Migrant–habitat relationships during stopover along an ecological barrier: extrinsic constraints and conservation implications. J. Ornithol. 152, 101–112. doi: 10.1007/s10336-010-0640-7
Bulte, M., and Bairlein, F. (2013). Endogenous control of migratory behavior in Alaskan northern wheatears Oenanthe oenanthe. J. Ornithol. 154, 567–570. doi: 10.1007/s10336-012-0920-5
Bulte, M., Heyers, D., Mouritsen, H., and Bairlein, F. (2017). Geomagnetic information modulates nocturnal migratory restlessness but not fueling in a long distance migratory songbird. J. Avian Biol. 48, 75–82. doi: 10.1111/jav.01285
Campioni, L., Dias, M. P., Granadeiro, J. P., and Catry, P. (2020). An ontogenetic perspective on migratory strategy of a long-lived pelagic seabird: timings and destinations change progressively during maturation. J. Anim. Ecol. 89, 29–43. doi: 10.1111/1365-2656.13044
Carneiro, C., Gunnarsson, T. G., and Alves, J. A. (2019). Why are whimbrels not advancing their arrival dates into Iceland? Exploring seasonal and sex-specific variation in consistency of individual timing during the annual cycle. Front. Ecol. Evol. 7:248. doi: 10.3389/fevo.2019.00248
Cassone, V. M. (2014). Avian circadian organization: a chorus of clocks. Front. Neuroendocrinol. 35, 76–88. doi: 10.1016/j.yfrne.2013.10.002
Chan, K. (1995). Diurnal and nocturnal pattern of activity in resident and migrant Silvereyes Zosterops lateralis. Emu 95, 41–46. doi: 10.1071/mu9950041
Chapin, J. P., and Wing, L. W. (1959). The Wideawake calendar, 1953 to 1958. Auk 76, 153–158. doi: 10.2307/4081773
Chapman, B. B., Brönmark, C., Nilsson, J. -Å., and Hansson, L.-A. (2011). The ecology and evolution of partial migration. Oikos 120, 1764–1775. doi: 10.1111/j.1600-0706.2011.20131.x
Ciarleglio, C. M., Axley, J. C., Strauss, B. R., Gamble, K. L., and McMahon, D. G. (2011). Perinatal photoperiod imprints the circadian clock. Nat. Neurosci. 14, 25–27. doi: 10.1038/nn.2699
Cochran, W. W., Mouritsen, H., and Wikelski, M. (2004). Migrating songbirds recalibrate their magnetic compass daily from twilight cues. Science 304, 405–408. doi: 10.1126/science.1095844
Conklin, J. R., Battley, P. F., and Potter, M. A. (2013). Absolute consistency: individual versus population variation in annual-cycle schedules of a long-distance migrant bird. PLoS One 8:e54535. doi: 10.1371/journal.pone.0054535
Conklin, J. R., Battley, P. F., Potter, M. A., and Fox, J. W. (2010). Breeding latitude drives individual schedules in a trans-hemispheric migrant bird. Nat. Commun. 1:67. doi: 10.1038/ncomms1072
Coppack, T., Becker, S. F., and Becker, P. J. J. (2008). Circadian flight schedules in night-migrating birds caught on migration. Biol. Lett. 4, 619–622. doi: 10.1098/rsbl.2008.0388
Coverdill, A. J., Bentley, G. E., and Ramenofsky, M. (2008). Circadian and masking control of migratory restlessness in gambel’s white-crowned sparrow (Zonotrichia leucophrys gambelii). J. Biol. Rhythms 23, 59–68. doi: 10.1177/0748730407311456
Covino, K. M., Holberton, R. L., and Morris, S. R. (2015). Factors influencing migratory decisions made by songbirds on spring stopover. J. Avian Biol. 46, 73–80. doi: 10.1111/jav.00463
Cruz, S. M., Hooten, M., Huyvaert, K. P., Proaño, C. B., Anderson, D. J., Afanasyev, V., et al. (2013). At–sea behavior varies with lunar phase in a nocturnal pelagic seabird, the swallow-tailed gull. PLoS One 8:e56889. doi: 10.1371/journal.pone.0056889
Crysler, Z. J., Ronconi, R. A., and Taylor, P. D. (2016). Differential fall migratory routes of adult and juvenile Ipswich Sparrows (Passerculus sandwichensis princeps). Mov. Ecol. 4:3. doi: 10.1186/s40462-016-0067-8
DeCoursey, P. (1960). Daily light sensitivity rhythm in a rodent. Science 131, 33–35. doi: 10.1126/science.131.3392.33
DeCoursey, P. (2004). “The behavioral ecology and evolution of biological timing systems,” in Chronobiology: Biological Timekeeping, eds J. C. Dunlap, J. J. Loros, and P. Decoursey (Sunderland, MA: Sinauer), 27–65.
Delmore, K. E., and Irwin, D. E. (2014). Hybrid songbirds employ intermediate routes in a migratory divide. Ecol. Lett. 17, 1211–1218. doi: 10.1111/ele.12326
Dhanjal-Adams, K. L., Bauer, S., Emmenegger, T., Hahn, S., Lisovski, S., and Liechti, F. (2018). Spatiotemporal group dynamics in a long-distance migratory bird. Curr. Biol. 28, 2824–2830. doi: 10.1016/j.cub.2018.06.054
Eikenaar, C., Klinner, T., Szostek, K. L., and Bairlein, F. (2014). Migratory restlessness in captive individuals predicts actual departure in the wild. Biol. Lett. 10:20140154. doi: 10.1098/rsbl.2014.0154
Eikenaar, C., and Schmaljohann, H. (2015). Wind conditions experienced during the day predict nocturnal restlessness in a migratory songbird. Ibis 157, 125–132. doi: 10.1111/ibi.12210
Ellegren, H. (1993). Speed of migration and migratory flight lengths of passerine birds ringed during autumn migration in Sweden. Ornis Scand. 24, 220–228.
Emlen, S. T. (1967). Migratory orientation in the indigo bunting, Passerina cyanea. Part II: mechanism of celestial orientation. Auk 84, 463–489. doi: 10.2307/4083330
Emlen, S. T. (1970). Celestial rotation: its importance in the development of migratory orientation. Science 170, 1198–1202.
Emlen, S. T. (1975). “Migration, orientation, and navigation,” in Avian Biology, eds D. S. Farner and J. R. King (Cambridge, MA: Academic Press), 129–219.
Fransson, T., Jakobsson, S., Johansson, P., Kullberg, C., Lind, J., and Vallin, A. (2001). Bird migration: magnetic cues trigger extensive refuelling. Nature 414, 35–36. doi: 10.1038/35102115
Fransson, T., Jakobsson, S., and Kullberg, C. (2005). Non-random distribution of ring recoveries from trans-Saharan migrants indicates species-specific stopover sites. J. Avian Biol. 36, 6–11. doi: 10.1111/j.0908-8857.2005.03471.x
Fraser, K. C., Shave, A. C., de Greef, E., Siegrist, J., and Garroway, C. J. (2019). Individual variability in migration timing can explain long-term, population-level advances in a songbird. Front. Ecol. Evol. 7:324. doi: 10.3389/fevo.2019.00324
Friesen, V. L. (2015). Speciation in seabirds: why are there so many species…and why aren’t there more? J. Ornithol. 156, 27–39. doi: 10.1007/s10336-015-1235-0
Fryxell, J., and Holt, R. (2013). Environmental change and the evolution of migration. Ecology 94, 1274–1279. doi: 10.1890/12-0668.1
Gaillard, J.-M. (2013). Assessing fitness consequences of migratory tactics requires longterm individually based monitoring. Ecology 94, 1261–1264. doi: 10.1890/12-0710.1
Gill, J. A., Alves, J. A., Sutherland, W. J., Appleton, G. F., Potts, P. M., and Gunnarsson, T. G. (2014). Why is timing of bird migration advancing when individuals are not? Proc. R. Soc. Lond. B 281:20132161. doi: 10.1098/rspb.2013.2161
Gow, E. A., Burke, L., Winkler, D. W., Knight, S. M., Bradley, D. W., Clark, R. G., et al. (2019). A range-wide domino effect and resetting of the annual cycle in a migratory songbird. Proc. Biol. Sci. 286:20181916. doi: 10.1098/rspb.2018.1916
Goymann, W., Helm, B., Jensen, W., Schwabl, I., and Moore, I. T. (2012). A tropical bird can use the equatorial change in sunrise and sunset times to synchronize its circannual clock. Proc. Biol. Sci. 279, 3527–3534. doi: 10.1098/rspb.2012.0743
Goymann, W., Lupi, S., Kaiya, H., Cardinale, M., and Fusani, L. (2017). Ghrelin affects stopover decisions and food intake in a long-distance migrant. Proc. Natl. Acad. Sci. U.S.A. 114, 1946–1951. doi: 10.1073/pnas.1619565114
Goymann, W., Spina, F., Ferri, A., and Fusani, L. (2010). Body fat influences departure from stopover sites in migratory birds: evidence from whole-island telemetry. Biol. Lett. 6, 478–481. doi: 10.1098/rsbl.2009.1028
Gwinner, E. (1986). Circannual Rhythms. Endogenous Annual Clocks in the Organization of Seasonal Processes. Berlin: Springer.
Gwinner, E. (1996a). Circadian and circannual programmes in avian migration. J. Exp. Biol. 199, 39–48.
Gwinner, E. (1996b). Circannual clocks in avian reproduction and migration. Ibis 138, 47–63. doi: 10.1111/j.1474-919x.1996.tb04312.x
Gwinner, E., Schwabl, H., and Schwabl-Benzinger, I. (1988). Effects of food-deprivation on migratory restlessness and diurnal activity in the garden warbler Sylvia borin. Oecologia 77, 321–326. doi: 10.1007/BF00378037
Gwinner, E., and Wiltschko, W. (1978). Endogenously controlled changes in migratory direction of the Garden Warbler Sylvia borin. J. Comp. Physiol. A 125, 267–273. doi: 10.1007/bf00656605
Gwinner, E., and Wiltschko, W. (1980). Circannual changes in migratory orientation of the garden warbler, Sylvia borin. Behav. Ecol. Sociobiol. 7, 73–78. doi: 10.1007/bf00302521
Häfker, N. S., Meyer, B., Last, K. S., Pond, D. W., Hüppe, L., and Teschke, M. (2017). Circadian clock involvement in zooplankton diel vertical migration. Curr. Biol. 27, 2194–2201. doi: 10.1016/j.cub.2017.06.025
Hedenström, A., Klaassen, R. H., and Åkesson, S. (2013). Migration of the little ringed plover Charadrius dubius breeding in South Sweden tracked by geolocators. Bird Study 60, 466–474. doi: 10.1080/00063657.2013.843635
Heim, W., Eccard, J. A., and Bairlein, F. (2018). Migration phenology determines niche use of East Asian buntings (Emberizidae) during stopover. Curr. Zool. 64, 681–692. doi: 10.1093/cz/zoy016
Helbig, A. J. (1991). Inheritance of migratory direction in a bird species: a cross-breeding experiment with SE-and SW-migrating blackcaps (Sylvia atricapilla). Behav. Ecol. Sociobiol. 28, 9–12.
Helbig, A. J. (1996). Genetic basis, mode of inheritance and evolutionary changes of migratory directions in Palearctic warblers (Aves: Sylviidae). J. Exp. Biol. 199, 49–55.
Helbig, A. J., Berthold, P., and Wiltschko, W. (1989). Migratory orientation in blackcaps (Sylvia atricapilla): population-specific shifts of direction during the autumn. Ethology 82, 307–315. doi: 10.1111/j.1439-0310.1989.tb00510.x
Helm, B. (2006). Zugunruhe of migratory and non-migratory birds in a circannual context. J. Avian Biol. 37, 533–540. doi: 10.1111/j.2006.0908-8857.03947.x
Helm, B., and Gwinner, E. (2006). Migratory restlessness in an equatorial nonmigratory bird. PLoS Biol. 4:e110. doi: 10.1371/journal.pbio.0040110
Helm, B., Gwinner, E., and Trost, L. (2005). Flexible seasonal timing and migratory behavior: results from Stonechat breeding programs. Ann. N. Y. Acad. Sci. 1046, 216–227. doi: 10.1196/annals.1343.019
Helm, B., Piersma, T., and Van der Jeugd, H. (2006). Sociable schedules: interplay between avian seasonal and social behaviour. Anim. Behav. 72, 245–262. doi: 10.1016/j.anbehav.2005.12.007
Helm, B., Schwabl, I., and Gwinner, E. (2009). Circannual basis of geographically distinct bird schedules. J. Exp. Biol. 212, 1259–1269. doi: 10.1242/jeb.025411
Helm, B., Van Doren, B. M., Hoffmann, D., and Hoffmann, U. (2019). Evolutionary response to climate change in migratory pied flycatchers. Curr. Biol. 29, 3714–3719. doi: 10.1016/j.cub.2019.08.072
Helm, B., Visser, M. E., Schwartz, W., Kronfeld-Schor, N., Gerkema, M., Piersma, T., et al. (2017). Two sides of a coin: ecological and chronobiological perspectives of timing in the wild. Philos. Trans. R. Soc. B 372:20160246. doi: 10.1098/rstb.2016.0246
Helms, C. W. (1963). “The annual cycle and Zugunruhe in birds,” in Proceedings of the XIIIth International Ornithological Congress Ithaca, (Ithaca, NY: American Ornithologists’ Union), 925–939.
Henshaw, I., Fransson, T., Jakobsson, S., Lind, J., Vallin, A., and Kullberg, C. (2008). Food intake and fuel deposition in a migratory bird is affected by multiple as well as single-step changes in the magnetic field. J. Exp. Biol. 211, 649–653. doi: 10.1242/jeb.014183
Hewson, C. M., Thorup, K., Pearce-Higgins, J. W., and Atkinson, P. W. (2016). Population decline is linked to migration route in the Common Cuckoo. Nat. Commun. 7:12296. doi: 10.1038/ncomms12296
Hodgson, S., and Quinn, T. P. (2002). The timing of adult sockeye salmon migration into fresh water: adaptations by populations to prevailing thermal regimes. Can. J. Zool. 80, 542–555. doi: 10.1139/z02-030
Holberton, R. L. (1993). An endogenous basis for differential migration in the dark-eyed junco. Condor 95, 580–587. doi: 10.2307/1369602
Holberton, R. L., and Able, K. P. (1992). Persistence of circannual cycles in a migratory bird held in constant dim light. J. Comp. Phys. A 171, 477–481.
Horton, K. G., La Sorte, F. A., Sheldon, D., Lin, T.-Y., Winner, K., Bernstein, G., et al. (2020). Phenology of nocturnal avian migration has shifted at the continental scale. Nat. Clim. Change 10, 63–68. doi: 10.1038/s41558-019-0648-9
Horton, W. J., Jensen, M., Sebastian, A., Praul, C. A., Albert, I., and Bartell, P. A. (2019). Transcriptome analyses of heart and liver reveal novel pathways for regulating songbird migration. Sci. Rep. 9:6058. doi: 10.1038/s41598-019-41252-8
Hut, R. A., and Beersma, D. G. M. (2011). Evolution of time-keeping mechanisms: early emergence and adaptation to photoperiod. Philos. Trans. R. Soc. Lond. B Biol. Sci. 366, 2141–2154. doi: 10.1098/rstb.2010.0409
Ilieva, M., Bianco, G., and Åkesson, S. (2018). Effect of geomagnetic field on migratory activity in a diurnal passerine migrant, the dunnock, Prunella modularis. Anim. Behav. 146, 79–85. doi: 10.1242/bio.014779
Karagicheva, J., Rakhimberdiev, E., Dekinga, A., Brugge, M., Koolhaas, A., Ten Horn, J., et al. (2016). Seasonal time keeping in a long-distance migrating shorebird. J. Biol. Rhythms 31, 509–521. doi: 10.1177/0748730416655929
Kiepenheuer, J. (1984). The magnetic compass mechanism of birds and its possible associations with the shifting course direction of migrants. Behav. Ecol. Sociobiol. 14, 81–99. doi: 10.1007/bf00291900
King, J. R., and Farner, D. S. (1963). The relationship of fat deposition to zugunruhe and migration. Condor 65, 200–223. doi: 10.2307/1365665
Kishkinev, D., Chernetsov, N., Pakhomov, A., Heyers, D., and Mouritsen, H. (2015). Eurasian reed warblers compensate for virtual magnetic displacement. Curr. Biol. 25, R822–R824.
Komenda-Zehnder, S., Liechti, F., and Bruderer, B. (2002). Is reverse migration a common feature of nocturnal bird migration?-an analysis of radar data from Israel. Ardea 90, 325–334.
Kramer, G. (1957). Experiments on bird orientation and their interpretation. Ibis 99, 196–227. doi: 10.1111/j.1474-919x.1957.tb01947.x
Kullberg, C., Henshaw, I., Jakobsson, S., Johansson, P., and Fransson, T. (2007). Fuelling decisions in migratory birds: geomagnetic cues override the seasonal effect. Proc. R. Soc. B Biol. Sci. 274, 2145–2151. doi: 10.1098/rspb.2007.0554
Kullberg, C., Lind, J., Fransson, T., Jakobsson, S., and Vallin, A. (2003). Magnetic cues and time of season affect fuel deposition in migratory thrush nightingales (Luscinia luscinia). Proc. R. Soc. B Biol. Sci. 270, 373–378. doi: 10.1098/rspb.2002.2273
Kumar, V., Rani, S., and Singh, B. (2006). Biological clocks help reduce the physiological conflicts in avian migrants. J. Ornithol. 147, 281–286. doi: 10.1007/s10336-006-0055-7
Lerche-Jørgensen, M., Willemoes, M., Tøttrup, A. P., Snell, K. R. S., and Thorup, K. (2017). No apparent gain from continuing migration for more than 3000 kilometres: willow warblers breeding in Denmark winter across the entire northern Savannah as revealed by geolocators. Mov. Ecol. 5:17. doi: 10.1186/s40462-017-0109-x
Liedvogel, M., Åkesson, S., and Bensch, S. (2011). The genetics of migration on the move. Trends Ecol. Evol. 26, 561–569. doi: 10.1016/j.tree.2011.07.009
Lundberg, M., Liedvogel, M., Larson, K., Sigeman, H., Grahn, M., Wright, A., et al. (2017). Genetic differences between willow warbler migratory phenotypes are few and cluster in large haplotype blocks. Evol. Lett. 1, 155–168. doi: 10.1002/evl3.15
Maggini, I., and Bairlein, F. (2010). Endogenous rhythms of seasonal migratory body mass changes and nocturnal restlessness in different populations of northern wheatear Oenanthe oenanthe. J. Biol. Rhythms 25, 268–276. doi: 10.1177/0748730410373442
Maggini, I., and Bairlein, F. (2012). Innate sex differences in the timing of spring migration in a songbird. PLoS One 7:e31271. doi: 10.1371/journal.pone.0031271
McKinnon, E. A., and Love, O. P. (2018). Ten years tracking the migrations of small landbirds: lessons learned in the golden age of bio-logging. Auk 135, 834–856. doi: 10.1642/auk-17-202.1
McKinnon, L., Smith, P. A., Nol, E., Martin, J. L., Doyle, F. I., Abraham, K. F., et al. (2010). Lower predation risk for migratory birds at high latitudes. Science 327, 326–327. doi: 10.1126/science.1183010
Merlin, C., and Liedvogel, M. (2019). The genetics and epigenetics of animal migration and orientation: birds, butterflies and beyond. J. Exp. Biol. 222:jeb191890. doi: 10.1242/jeb.191890
Milot, E., Weimerskirch, H., and Bernatchez, L. (2008). The seabird paradox: dispersal, genetic structure and population dynamics in a highly mobile, but philopatric albatross species. Mol. Ecol. 17, 1658–1673. doi: 10.1111/j.1365-294X.2008.03700.x
Muheim, R., Åkesson, S., and Alerstam, T. (2003). Compass orientation and possible migration routes of passerine birds at high arctic latitudes. Oikos 103, 341–349. doi: 10.1034/j.1600-0706.2003.12122.x
Muheim, R., Boström, J., Åkesson, S., and Liedvogel, M. (2014). “Sensory mechanisms of animal orientation and navigation,” in Animal Movement Across Scales, eds L. A. Hansson and S. Åkesson (Oxford: Oxford University Press), 179–194. doi: 10.1093/acprof:oso/9780199677184.003.0010
Muheim, R., Phillips, J. B., and Åkesson, S. (2006). Polarized light cues underlie compass calibration in migratory songbirds. Science 313, 837–839. doi: 10.1126/science.1129709
Muheim, R., Schmaljohann, H., and Alerstam, T. (2018). Feasibility of sun and magnetic compass mechanisms in avian long-distance migration. Mov. Ecol. 6:8. doi: 10.1186/s40462-018-0126-4
Mukhin, A., Kobylkov, D., Kishkinev, D., and Grinkevich, V. (2018). Interrupted breeding in a songbird migrant triggers development of nocturnal locomotor activity. Sci. Rep. 8:5520. doi: 10.1038/s41598-018-23834-0
Müller, F., Eikenaar, C., Crysler, Z. J., Taylor, P. D., and Schmaljohann, H. (2018). Nocturnal departure timing in songbirds facing distinct migratory challenges. J. Anim. Ecol. 87, 1102–1115. doi: 10.1111/1365-2656.12821
Norevik, G., Åkesson, S., Andersson, A., Bäckman, J., and Hedenström, A. (2019). The lunar cycle drives migration of a nocturnal bird. PLoS Biol. 17:e3000456. doi: 10.1371/journal.pbio.3000456
Norevik, G., Åkesson, S., and Hedenström, A. (2017). Migration strategies and annual space-use in an Afro-Palaearctic aerial insectivore – the European nightjar Caprimulgus europaeus. J. Avian Biol. 48, 738–747. doi: 10.1111/jav.01071
O’Connor, C. M., Norris, D. R., Crossin, G. T., and Cooke, S. J. (2014). Biological carryover effects: linking common concepts and mechanisms in ecology and evolution. Ecosphere 5:art28. doi: 10.1890/es13-00388.1
O’Connor, E. A., Cornwallis, C. K., Hasselquist, D., Nilsson, J. Å., and Westerdahl, H. (2018). The evolution of immunity in relation to colonization and migration. Nat. Ecol. Evol. 2, 841–849. doi: 10.1038/s41559-018-0509-3
Perdeck, A. C. (1958). Two types of orientation in migrating starlings, Sturnus vulgaris L., and chaffinches, Fringilla coelebs L., as revealed by displacement experiments. Ardea 55, 194–202.
Phillimore, A. B., Leech, D. I., Pearce-Higgins, J. W., and Hadfield, J. D. (2016). Passerines may be sufficiently plastic to track temperature-mediated shifts in optimum lay date. Glob. Change Biol. 22, 3259–3272. doi: 10.1111/gcb.13302
Piersma, T., and Drent, J. (2003). Phenotypic flexibility and the evolution of organismal design. Trends Ecol. Evol. 18, 228–233. doi: 10.1093/icb/42.1.68
Pulido, F., and Berthold, P. (2010). Current selection for lower migratory activity will drive the evolution of residency in a migratory bird population. Proc. Natl. Acad. Sci. U.S.A. 107, 7341–7346. doi: 10.1073/pnas.0910361107
Ramenofsky, M. (2011). “Hormones in migration and reproductive cycles of birds,” in Hormones and Reproduction of Vertebrates, eds D. O. Norris and K. H. Lopez (London: Academic Press), 205–237. doi: 10.1016/b978-0-12-374932-1.00046-9
Reid, J. M., Travis, J. M. J., Daunt, F., Burthe, S. J., Wanless, S., and Dytham, C. (2018). Population and evolutionary dynamics in spatially structured seasonally varying environments. Biol. Rev. 93, 1578–1603. doi: 10.1111/brv.12409
Reppert, S. M., and de Roode, J. C. (2018). Demystifying monarch butterfly migration. Curr. Biol. 28, R1009–R1022. doi: 10.1016/j.cub.2018.02.067
Ritz, T., Wiltschko, R., Hore, P. J., Rodgers, C. T., Stapput, K., Thalau, P., et al. (2009). Magnetic compass of birds is based on a molecule with optimal directional sensitivity. Biophys. J. 96, 3451–3457. doi: 10.1016/j.bpj.2008.11.072
Rolland, J., Jiguet, F., Joansson, K. A., Condamine, F. L., and Morlon, H. (2014). Settling down of seasonal migrants promotes bird diversification. Proc. Biol. Sci. 281:20140473. doi: 10.1098/rspb.2014.0473
Ruegg, K., Anderson, E. C., Boone, J., Pouls, J., and Smith, T. B. (2014). A role for migration-linked genes and genomic islands in divergence of a songbird. Mol. Ecol. 23, 4757–4769. doi: 10.1111/mec.12842
Sandberg, R. (1994). Interaction of body condition and magnetic orientation in autumn migrating robins, Erithacus rubecula. Anim. Behav. 47, 679–686. doi: 10.1006/anbe.1994.1092
Sandberg, R. (2003). “Stored fat and the migratory orientation of birds,” in Avian Migration, eds P. Berthold, E. Gwinner, and E. Sonnenschein (Berlin: Springer), 515–525. doi: 10.1007/978-3-662-05957-9_35
Schmidt-Koenig, K., Ganzhorn, J. U., and Ranvaud, R. (1991). “The sun compass,” in Orientation in Birds, ed. P. Berthold (Basel: Birkhäuser), 1–15.
Senner, N. R., Conklin, J. R., and Piersma, T. (2015). An ontogenetic perspective on individual differences. Proc. Biol. Sci. 282:20151050. doi: 10.1098/rspb.2015.1050
Senner, N. R., Hochachka, W. M., Fox, J. W., and Afanasyev, V. (2014). An exception to the rule: carry-over effects do not accumulate in a long-distance migratory bird. PLoS One 9:e86588. doi: 10.1371/journal.pone.0086588
Senner, N. R., Morbey, Y. E., and Sandercock, B. K. (2020). Editorial: the degree of flexibility in the migration strategies of animals. Front. Ecol. Evol, 8.
Sergio, F., Tanferna, A., De Stephanis, R., Jiménez, L. L., Blas, J., Tavecchia, G., et al. (2014). Individual improvements and selective mortality shape lifelong migratory performance. Nature 515, 410–413. doi: 10.1038/nature13696
Shamoun-Baranes, J., van Loon, E., Liechti, F., and Bouten, W. (2007). Analyzing the effect of wind on flight: pitfalls and solutions. J. Exp. Biol. 210, 82–90. doi: 10.1242/jeb.02612
Shaw, P. (2017). Rainfall, leafing phenology and sunrise time as potential Zeitgeber for the bimodal, dry season laying pattern of an African rain forest tit (Parus fasciiventer). J. Ornithol. 158, 263–275. doi: 10.1007/s10336-016-1395-6
Sjöberg, S., Alerstam, T., Åkesson, S., Schulz, A., Weidauer, A., Coppack, T., et al. (2015). Weather and fuel reserves determine departure and flight decisions in passerines migrating across the Baltic Sea. Anim. Behav. 104, 59–68. doi: 10.1016/j.anbehav.2015.02.015
Sokolovskis, K., Bianco, G., Willemoes, M., Solovyeva, D., Bensch, S., and Åkesson, S. (2018). Ten grams and 13,000 km on the wing–route choice in willow warblers Phylloscopus trochilus yakutensis migrating from Far East Russia to East Africa. Mov. Ecol. 6:20. doi: 10.1186/s40462-018-0138-0
Stanley, C. Q., MacPherson, M., Fraser, K. C., McKinnon, E. A., and Stutchbury, B. J. M. (2012). Repeat tracking of individual songbirds reveals consistent migration timing but flexibility in route. PLoS One 7:e40688. doi: 10.1371/journal.pone.0040688
Styrsky, J. D., Berthold, P., and Robinson, W. D. (2004). Endogenous control of migration and calendar effects in an intratropical migrant, the yellow–green vireo. Anim. Behav. 67, 1141–1149. doi: 10.1016/j.anbehav.2003.07.012
Sugasawa, S., and Higuchi, H. (2019). Seasonal contrasts in individual consistency of oriental honey buzzards’ migration. Biol. Lett. 15:20190131. doi: 10.1098/rsbl.2019.0131
Sutherland, W. J. (1998). Evidence for flexibility and constraint in migration systems. J. Avian Biol. 29, 441–446.
Terrill, S. B., and Able, K. P. (1988). Bird migration terminology. Auk 105, 205–206. doi: 10.1093/auk/105.1.205
Thackeray, S. J., Henrys, P. A., Hemming, D., Bell, J. R., Botham, M. S., Burthe, S., et al. (2016). Phenological sensitivity to climate across taxa and trophic levels. Nature 535, 241–245. doi: 10.1038/nature18608
Thorup, K., Alerstam, T., Hake, M., and Kjellén, N. (2003). Bird orientation: compensation for wind drift in migrating raptors is age dependent. Proc. Biol. Sci. 270(Suppl._1), S8–S11.
Thorup, K., Bisson, I. A., Bowlin, M. S., Holland, R. A., Wingfield, J. C., Ramenofsky, M., et al. (2007). Evidence for a navigational map stretching across the continental US in a migratory songbird. Proc. Natl. Acad. Sci. U.S.A. 104, 18115–18119. doi: 10.1073/pnas.0704734104
Tickell, W. L. N. (1968). The biology of the great albatrosses, Diomedea exulans and Diomedea epomophora. Antarctic. Res. Ser. 12, 1–55.
Tryjanowski, P., Kuźniak, S., and Sparks, T. H. (2005). What affects the magnitude of change in first arrival dates of migrant birds? J. Ornithol. 146, 200–205. doi: 10.1007/s10336-005-0079-4
Van Doren, B., and Horton, K. G. (2018). A continental system for forecasting bird migration. Science 361, 1115–1118. doi: 10.1126/science.aat7526
Van Doren, B., Liedvogel, M., and Helm, B. (2016). Programmed and flexible: long-term Zugunruhe data highlight the many axes of variation in avian migratory behaviour. J. Avian Biol. 48, 155–172. doi: 10.1111/jav.01348
Van Doren, B. M., Horton, K. G., Dokter, A. M., Klinck, H., Elbin, S. B., and Farnsworth, A. (2017). High-intensity urban light installation dramatically alters nocturnal bird migration. Proc. Natl. Acad. Sci. U.S.A. 114, 11175–11180. doi: 10.1073/pnas.1708574114
van Noordwijk, A. J., Pulido, F., Helm, B., Coppack, T., Delingat, J., Dingle, H., et al. (2006). A framework for the study of genetic variation in migratory behaviour. J. Ornithol. 147, 221–233. doi: 10.1007/s10336-005-0047-z
Vardanis, Y., Klaassen, R. H., Strandberg, R., and Alerstam, T. (2011). Individuality in bird migration: routes and timing. Biol. Lett. 7, 502–505. doi: 10.1098/rsbl.2010.1180
Vardanis, Y., Nilsson, J. -Å., Klaassen, R. H., Strandberg, R., and Alerstam, T. (2016). Consistency in long-distance bird migration: contrasting patterns in time and space for two raptors. Anim. Behav. 113, 177–187. doi: 10.1016/j.anbehav.2015.12.014
Vega, M. L., Willemoes, M., Thomson, R. L., Tolvanen, J., Rutila, J., Samaš, P., et al. (2016). First-time migration in juvenile common cuckoos documented by satellite tracking. PLoS One 11:e0168940. doi: 10.1371/journal.pone.0168940
Visser, M. E., Caro, S. P., van Oers, K., Schaper, S. V., and Helm, B. (2010). Phenology, seasonal timing and circannual rhythms: towards a unified framework. Philos. Trans. R. Soc. Lond. B Biol. Sci. 365, 3113–3127. doi: 10.1098/rstb.2010.0111
Watts, H. E., Cornelius, J. M., Fudickar, A. M., Pérez, J., and Ramenofsky, M. (2018). Understanding variation in migratory movements: a mechanistic approach. Gen. Comp. Endocrinol. 256, 112–122. doi: 10.1016/j.ygcen.2017.07.027
Watts, H. E., Robart, A. R., Chopra, J. K., Asinas, C. E., Hahn, T. P., and Ramenofsky, M. (2016). Seasonal expression of migratory behavior in a facultative migrant, the pine siskin. Behav. Ecol. Sociobiol. 71:9.
Weimerskirch, H., Åkesson, S., and Pinaud, D. (2006). Postnatal dispersal of wandering albatrosses Diomedea exulans: implications for the conservation of the species. J. Avian Biol. 37, 23–28. doi: 10.1111/j.0908-8857.2006.03675.x
Weimerskirch, H., Delord, K., Guitteaud, A., Phillips, R. A., and Pinet, P. (2015). Extreme variation in migration strategies between and within wandering albatross populations during their sabbatical year, and their fitness consequences. Sci. Rep. 5:8853. doi: 10.1038/srep08853
Weimerskirch, H., and Wilson, R. P. (2000). Oceanic respite for wandering albatrosses. Nature 406, 955–956. doi: 10.1038/35023068
Weindler, P., Wiltschko, R., and Wiltschko, W. (1996). Magnetic information affects the stellar orientation of young bird migrants. Nature 383, 158–160. doi: 10.1038/383158a0
Wikelski, M., Arriero, E., Gagliardo, A., Holland, R. A., Huttunen, M. J., Juvaste, R., et al. (2015). True navigation in migrating gulls requires intact olfactory nerves. Sci. Rep. 5:17061. doi: 10.1038/srep17061
Willemoes, M., Blas, J., Wikelski, M., and Thorup, K. (2015). Flexible navigation response in common cuckoos Cuculus canorus displaced experimentally during migration. Sci. Rep. 5:16402. doi: 10.1038/srep16402
Willemoes, M., Strandberg, R., Klaassen, R. H., Tøttrup, A. P., Vardanis, Y., Howey, P. W., et al. (2014). Narrow-front loop migration in a population of the common cuckoo Cuculus canorus, as revealed by satellite telemetry. PLoS One 9:e83515. doi: 10.1371/journal.pone.0083515
Wiltschko, W., and Wiltschko, R. (1972). Magnetic compass of European robins. Science 176, 62–64. doi: 10.1126/science.176.4030.62
Wiltschko, W., and Wiltschko, R. (1992). Migratory orientation: magnetic compass orientation of garden warblers (Sylvia borin) after a simulated crossing of the magnetic equator. Ethology 91, 70–74. doi: 10.1111/j.1439-0310.1992.tb00851.x
Winkler, D. W., Gandoy, F. A., Areta, J. I., Iliff, M. J., Rakhimberdiev, E., Kardynal, K. J., et al. (2017). Long-distance range expansion and rapid adjustment of migration in a newly established population of barn swallows breeding in Argentina. Curr. Biol. 27, 1080–1084. doi: 10.1016/j.cub.2017.03.006
Keywords: circannual rhythm, endogenous programs, geomagnetic field, migration, navigation, songbirds
Citation: Åkesson S and Helm B (2020) Endogenous Programs and Flexibility in Bird Migration. Front. Ecol. Evol. 8:78. doi: 10.3389/fevo.2020.00078
Received: 30 July 2019; Accepted: 10 March 2020;
Published: 26 March 2020.
Edited by:
Nathan R. Senner, University of South Carolina, United StatesReviewed by:
Elizabeth A. Gow, University of Guelph, CanadaFranz Bairlein, Institute of Avian Research, Germany
Copyright © 2020 Åkesson and Helm. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Susanne Åkesson, c3VzYW5uZS5ha2Vzc29uQGJpb2wubHUuc2U=; Barbara Helm, Yi5oZWxtQHJ1Zy5ubA==
†These authors have contributed equally to this work