 Anne E. Storey
Anne E. Storey Sabina I. Wilhelm
Sabina I. Wilhelm Carolyn J. Walsh
Carolyn J. Walsh- 1Department of Psychology, Memorial University of Newfoundland, St. John’s, NL, Canada
- 2Canadian Wildlife Service, Environment and Climate Change Canada, Mount Pearl, NL, Canada
Mates of biparental species share parental care but they are also predicted to try, within limits, to push for more offspring care from their partners. Here we test (a) whether mates will attempt to exploit their partners less often when resources are scarce so as to not push their partners to their abandonment threshold and (b) whether there are differences in exploitation and compensation strategies of low and high quality partners where the quality assessment is based on within-pair differences in chick-provisioning rates. The same 14 pairs of common murres were observed in a year when capelin fish (the main prey species) were abundant (match year) and in a second year when capelin did not arrive inshore to spawn until the second week after hatching (mismatch year). One murre parent always attends the chick and, in the most common type of interaction, the returning parent feeds the chick, and takes over the brooding role. We consider nest relief interactions to be irregular if they did not follow this sequence for determining which parent will continue or take on the lower-energy brooder role. Two types of irregular nest reliefs were examined: (a) the returning bird does not bring a fish and (b) the brooding bird does not allow a brooding changeover even when the returner brings a fish. Rates of irregular nest reliefs and total visit time increased in the mismatch year after capelin arrived inshore, suggesting that longer co-attendance in good conditions reflects negotiation, rather than the increased resting or “loafing” time as previously proposed. High provisioners initiated fewer irregular nest reliefs than their low provisioning partners during favorable conditions but increased to comparable levels when resources were scarce. Partners' attempts to brood without provisioning were less often refused during unfavorable feeding conditions, suggesting that murres compensated for their mates when they could. The observation that rates of irregular nest reliefs changed with resource availability suggests that negotiation occurs throughout chick rearing and is not a set “sealed bid” at the onset.
Introduction
Although considerable intra-pair cooperation is necessary for successful reproduction in biparental species, individuals should still try to make their mates increase their share of parental work (Jones et al., 2002). However, if pair bonds typically persist across breeding seasons, as in seabirds, individuals should be less likely to exploit their mates, as their partners' long-term survival is critical to their own fitness (Jones et al., 2002). Thus, when resources are limited, breeding seabirds will adjust their parental efforts to prioritize their own survival, and possibly that of their mates, over investment in their chicks (Monaghan et al., 1989; Chastel et al., 1995; Weimerskirch and Lys, 2000; Weimerskirch et al., 2001; but see Kitaysky et al., 1999). On the other hand, individuals in these species should not always compensate fully for any decreased investment by the other parent, as this may lead to evolutionary instability that could be exploited by low investing individuals (Houston et al., 2005). Therefore, seabirds should attempt to negotiate parental duties with their partners but not to decrease their own investment enough that the mate must abandon the breeding attempt (Jones et al., 2002) and/or that could possibly disrupt the pair bond (Moody et al., 2005).
Pair members in seabird species, with their shared care, long lives and long pair bonds are predicted to compensate for their partners until the body condition of one or both partners drops below the threshold at which either partner should abandon the current breeding attempt (Jones et al., 2002). A range of negotiated flexibility should exist between the abandonment threshold and the preferred, less costly level of investment, termed in Jones et al. (2002) the “laziness threshold.” The “laziness threshold” represents the point at which each partner would be doing the least possible to maintain its parental investment while not pushing the partner to the abandonment threshold (Jones et al., 2002). One counterintuitive prediction is that individuals should push their partners to increase care more under good conditions than under poor ones because their mates should be more able to take on extra energetic costs when food is abundant (Jones et al., 2002). Further, signals from the lower quality partner should cause the higher quality partner to assume more parental care when pushed to do more, particularly under good conditions, because costs to itself are less than the risk of the lower quality partner deteriorating in body condition to the abandonment threshold.
An important step to document parental turn taking is determining whether individuals appear to be monitoring their mates' nest provisioning visits and modifying their own visits accordingly (e.g., Bebbington and Hatchwell, 2016; Savage et al., 2017). This step is easier to establish in a seabird species such as the common murre Uria aalge in which, prior to fledging, chicks are continuously attended by at least one parent to minimize predation at their open-cliff, high-density breeding colonies (Ainley et al., 2002). Thus, common murre parents likely have complete information about when their mates visit and whether or not they feed the chick. It is therefore possible to examine the behaviors displayed at nest reliefs, and the times between reliefs to determine what factors influence whether complete turn taking is occurring, that is, whether the nest relief proceeds or not.
Parental care is particularly costly for the common murre for several reasons. In addition to only one parent being able to forage at a time, flight is energetically costly because murres have the highest wing loading of any flying bird, and they can only bring a single fish to the chick each trip (Ainley et al., 2002). Murre parents generally experience mass loss over the chick-rearing period (Harris et al., 2000; Ainley et al., 2002; Wilhelm, 2004) and this mass loss is greater when resources are poor (Wilhelm, 2004).
Parental co-attendance in common murres occurs several times a day when the foraging partner returns to the colony, most often bringing a fish for the chick. An important component of co-attendance is the nest relief, which is initiated when the non-brooding mate returns to the nest site, usually with a fish to provision the chick. While the returning bird controls when the interaction will start by coming to the nest site, the brooding bird controls whether and/or how long it takes for the returning bird to be allowed access to the chick for brooding (the actual nest relief) and then how long it will remain at the nest site after the nest relief. Takahashi et al. (2017) distinguished between normal nest reliefs (returner brings a fish, nest relief occurs, and brooder departs, more than half visits) and irregular nest reliefs. We suggest that three types of irregular nest relief sequences are how murres push their mates to provide more parental care: no nest relief (returner brings a fish but is not allowed to brood), multiple nest reliefs (returner brings fish but brooder does not depart after the nest relief and more nest reliefs follow), and no fish visits (returner fails to bring a fish and a nest relief may or may not follow). Compensation, or allowing the partner to brood in an irregular nest relief, can occur when either the bird returning with a fish departs without a nest relief or when the brooder allows the bird that returns without a fish to initiate a brooding bout. The irregular nest reliefs differ from the normal ones in that the brooder exhibits fewer or delayed bouts of allopreening, and in some cases the interactions are longer (Takahashi et al., 2017).
Given the extensive parental care provided by both parents, it might not be obvious how murres could push their mates to increase care. Of the two major investments in rearing murre chicks, brooding appears to be less energetically expensive than chick feeding (Birt-Friesen et al., 1989; Gabrielsen, 1996). Data from data loggers attached to murres support this contention in that both physiological and behavioral measures are changed by wearing the loggers. Murres have the highest wing loading of any flying bird (Ainley et al., 2002) and thus are affected by the extra load of carrying the loggers. Corticosterone levels were higher in murres with data loggers than controls (Elliott et al., 2012), suggesting that carrying a data logger produces the same effect on corticosterone levels as decreased resources (as in Doody et al., 2008). Murres carrying data loggers decreased their chick provisioning rates, increased the number of no-fish visits and left the colony less often, and in some cases, their mates compensated for their partner's reduced foraging activity (Wanless et al., 1988; Hamel et al., 2004; Paredes et al., 2005). Murres, therefore, may not attempt to reduce their total parental care, but rather to increase the duration of the energetically low-cost brooding component, possibly leaving the other partner to forage more (Takahashi et al., 2017). Continuing the brooding bout even after the mate has provisioned the chick, thus preventing a nest relief, may be how a murre in poor condition (naturally or by having an attached data logger) might push its partner to forage more.
One aspect of turn-taking that has received relatively little attention is the impact of variation in resources on how often birds try to push their mates and whether these attempts are accepted (compensation). Chick-feeding rates and the time both parents spent together in the colony (co-attendance time) were lower in a predator-prey mismatch year of the current study (2000), when capelin (Mallotus villosus) arrived inshore after hatching, compared to two match years (1998 and 1999) when capelin arrival coincided with, or preceded, hatching (Wilhelm et al., 2008). Chick-feeding rates and co-attendance durations increased to the levels of previous years once capelin arrived inshore in the second week after hatching in 2000. This increase in co-attendance time when prey becomes plentiful has been documented in previous murre studies and it has been interpreted as birds having more “loafing” time when prey is plentiful (Cairns et al., 1987; Uttley et al., 1994; Zador and Piatt, 1999). Alternatively, since it is inefficient for both parents to co-attend when only one parent is required, variation in co-attendance time may signal information about the brooder's body condition and hence be a way to request the mate to increase care (Jones et al., 2002).
Here, we examined differences in rates that birds initiated irregular nest reliefs under variable foraging conditions and tested the predictions that under poor foraging conditions murres will initiate fewer irregular nest reliefs and they will resist changeovers for shorter periods, thus resulting in less time overall spent co-attending. We attempted to distinguish between the “loafing” and “negotiation” functions of nest relief behavior by testing whether (a) high quality birds (with higher chick-provisioning rates) both initiated fewer irregular nest reliefs and compensated more than their mates and (b) co-attendance times varied across nest relief types, such that particular irregular nest reliefs (e.g., returner does not bring a fish), would be longer than normal nest reliefs (returner brings a fish and a nest relief follows). The loafing function would be more strongly supported if co-attendance times were evenly distributed across all nest visits and both pair members.
Methods
Study Area
We collected data across three breeding seasons (1998–2000) for common murres breeding on Great Island in Witless Bay, Newfoundland and Labrador, Canada (47° 11′N, 52° 49′W; Wilhelm, 2004). Data are from 28 birds from 14 focal pairs that had surviving chicks in each year. Individuals in these 14 pairs could be individually distinguished as at least one member of the pair was color banded and/or had the distinctive bridled eye band. Twenty-four of 28 (86%) birds were color banded by 1998 and 26/28 by 1999 (93%, the remaining birds were bridled, a marking only seen in about 20% of birds). Given the low divorce rate and behavior particular to mate changing (Moody et al., 2005) as well as the high survival rate of known birds at this site (Robertson et al., 2006), it seems likely that all these pairs consisted of the same individuals in all 3 years. We observed the birds through one-way glass in a wooden blind placed adjacent to the nesting cliff. The birds' behavior appeared to be unaffected by the presence of the blind (i.e., no head tossing or “murring” vocalization, Ainley et al., 2002), with some breeding within 0.5 m of the base of the blind. We minimized disturbance by accessing the blind through a canvas tunnel, entering before dawn and departing after sunset. We sexed the birds behaviorally: copulations were observed during the pre-lay period of six consecutive breeding seasons (no reverse mountings have been recorded for this species). Male murres take their chicks to sea at fledging (Ainley et al., 2002) and no fathers of fledged chicks were observed in the colony after their chicks departed. Finally, we verified sex genetically for adults from the same colony with all birds matching the sex assigned behaviorally (N = 28, Cameron-MacMillan et al., 2007). The proportion of birds at this site that raised a chick successfully to colony departure is generally high (0.75–0.79 fledged chicks/eggs laid; Wilhelm and Storey, 2002). Hence, this site was viewed as being well-established and the current study most likely included many experienced, older birds.
We compared chick-feeding rates to the inshore arrival dates, spawning dates, and approximate densities of capelin, the main species fed to chicks, which were recorded by the same fisherman in capelin diaries spanning across several years, including the years of this study. These diaries were sponsored by the Fisheries and Oceans Canada. The fisherman was instructed to make daily notes using the following checklist: spawning: (1) none observed, (2) spawning activity (light, medium, or intense), (3) presence of dead capelin on the beach or in the water (few, moderate, or heavy concentrations), (4) presence of live fish in the water (three levels: few fish, few small schools, or many or dense schools), and (5) indirect evidence that spawning had occurred within 24 h. These observations were consistent with the timing of peak capelin spawning densities from Bellevue Beach in an adjacent bay (Nakashima, Fisheries, and Oceans Canada, personal communication) and with observations of other biologists in the area. Hatching extended from June 29 to July 12 in 1999 and from June 23 to June 30 in 2000, the two main years in this study. Capelin spawning onset was June 25 and the peak was June 28 locally and July 10 in Bellevue Bay in 1999 whereas, in 2000, capelin were first observed 9 days after the onset of hatching (July 2) and peak spawning did not occur until July 17 (Wilhelm et al., 2008, based on information from Fisheries and Oceans Canada). The year 2000 was the only year (of 8 years) that young chicks were observed to die of starvation early in the season; even the focal pairs in this study (all successful chick-rearers) showed deceased feeding rates early in 2000 compared to 1998 and 1999 (Wilhelm et al., 2008).
Thus, 1998 and 1999 were designated as prey match years and 2000 was designated as a prey mismatch year (Doody et al., 2008; Wilhelm et al., 2008). The focus of this paper is on the contrasting resource years of 1999 and 2000 (henceforth called the match and mismatch years, respectively). Information from 1998 was used with 1999 data to confirm the assignment of high or low provisioning status within pairs. Chick feeding rates were significantly correlated for 1998 and 1999 (r = 0.43, P = 0.014). Based on the average chick provisioning rates for 1998 and 1999, each of the 14 pairs was assigned one high and one low provisioner (categorical variable “provisioner status”), allowing us to make within-pair comparisons of strategies for initiating irregular nest reliefs. High provisioners were equally likely to be male or female.
Colony Observations
We made continuous observations, beginning at dawn (first light at 04:30 h, NDT) and finishing at dusk (last light before 21:30 h, NDT), on 11 days in July and August 1998 (175.8 h total), 15 days in July and August 1999 (238 h total), and 12 days in July 2000 (196 h total). The total number of hours is slightly reduced from the possible total because as the season advanced there were brief periods in morning and evening that we could not accurately see the color bands of departing birds. Two observers were present on each observation day and they switched off every 3 h.
We recorded all visits (total N = 1,837; 1999, 961; 2000, 876 visits) and we coded them in terms of whether the returning bird brought a fish (1999; 807, 83% of visits; 2000, 710, 80% of visits). We recorded the length of all interactions from the time of the returner's arrival until one of the birds departed. Spot checks of all nests were taken every half hour so that when the occasional departure was missed (fewer than 5% of interactions), it was assumed to have occurred in the middle of the two spot checks. An interaction or visit was called a normal nest relief if the returning bird brought a fish to the chick and a nest relief followed after which the brooding bird departed from the colony. All other visits were called irregular nest interactions. We recorded whether or not the returner brought a fish so that we could tabulate daily chick-feeding rates for each week for all focal murres.
We compared the frequency of irregular nest reliefs between predator-prey match and mismatch years, based on the assumption that the brooder controls whether a nest relief occurs or not. We calculated a daily frequency for each individual as the combined daily frequency of (a) the brooder not letting the returning partner take over brooding after it provisioned the chick, and (b) the returning bird attempting (successfully or not) to brood without first provisioning the chick. Thus, a brooding bird with a high rate of pushing its mate to do more would have often prevented their partner from brooding after the partner brought a fish and then the same bird as returner would have often tried to initiate a brooding bout without bringing a fish. Attempts to continue or start brooding without first provisioning were classified as successful if the bird's returning partner left the colony to forage without having a brooding bout or if the bird returning without a fish was allowed to brood the chick. A third category of irregular nest relief interactions was the multiple nest relief in brooder did not leave the colony after the nest relief and one or more additional nest exchanges occurred. These multiple nest reliefs comprised only a small proportion of the visits in this study (~4%) and were not part of the frequency analysis but were included in the timing analyses. We calculated weekly rates of initiation of irregular nest reliefs for weeks 1, 2, and 3 after hatching. Weeks were based on hatch dates for each pair so that all birds in week 1 had chicks hatching in the past 7 days and so on for weeks 2 and 3. These data were used in the repeated measures ANOVA analyses.
We obtained body masses in two ways (N = 23): opportunistically when birds stepped on one of the three electronic balances (Ohaus CS-2000 accurate to ±1 gram, modified so that measurements were recorded from within the blind) we had cemented to rocks on the cliff, and also during capture for banding and blood sampling (Wilhelm, 2004). We considered body mass to be a good measure of condition as most fluctuation in mass in murres reflects variation in stored fat (Jacobs et al., 2011) and since the current comparisons are changes within individual adults, no structural differences need to be considered. There was no significant difference in the measurements produced by the scales or at capture, based on 12 murres caught by both methods [capture, mean 932.92 ± 14.1 g; scale, mean 935.25 ± 12.8 g, t(11) = 0.16, p = 0.87]. Body mass was compared for the same birds matched for chick age (range: 7–24 days post-hatch) in 1999 and 2000. If measurements were not available for exactly the same chick age day, masses were adjusted by the average daily mass loss for birds in this colony, that is 5 g per day (up to a maximum of 4 days difference in chick age, Cameron-MacMillan et al., 2007).
Statistical Analyses
We examined rates of initiation of irregular nest reliefs for 14 pairs with early hatching chicks in the mismatch year over weeks (first week, before capelin arrival; second week, moderate capelin density for that year; third week highest capelin density; Wilhelm et al., 2008). We compared these rates to those of the same pairs in the match year over the same weeks relative to the hatching of their chicks. The repeated factors in the repeated measures ANOVA were provisioner status (high and low provisioner in each pair), weeks (one, two, and three) and years (match and mismatch years) with sex as the between factor. We also used repeated measures ANOVA to examine variation in compensation and the differences in mass for high and low provisioners in the 2 years. All interactions were examined but only significant ones are reported. Interactions were examined using t-tests.
Linear mixed-effects models (LMMs) for the entire data set were constructed to examine whether behaviors exhibited during nest relief sequences were significantly influenced by the type of turn-taking sequence (normal and various irregular sequences), the year (match or mismatch), timing of capelin arrival (before or after week 1 of the mismatch year), and provisioner status (high vs. low provisioner within pair). Pair mates were nested within nest site by sex and used as a random factor. Specifically, the total time for the nest relief (or no nest relief) interactions and the times since the last visit by the focal bird and the mate were analyzed. Scaled identity was used as the model covariance structure as suggested for situations in which levels of main variables were not correlated and variances were relatively constant (Beaumont, 2012, https://www.ibm.com/support/knowledgecenter/SSLVMB_24.0.0/spss/advanced/covariance_structures.html).
Linear regression was used to assess which continuous variables best predicted the total time of nest interactions [times since last event (e.g., time since last visit or chick feed by each of the partners), time of day, time in chick rearing]. For all analyses, statistical significance was set at α = 0.05 and we report values as means ± 1 SE. All analyses were performed in SPSS version 25.
Results
The distribution of regular and irregular nest relief interactions can be seen in Table 1. Note that No fish visits are rare early in both years, despite the lower provisioning rate in the mismatch year (Figure 1A). Low provisioners had more irregular nest reliefs (made more no-fish visits and/or refused more nest reliefs) than high provisioners in the match year (Table 2). Murres increased their rates of initiating irregular nest reliefs in the second week of the mismatch year after feeding conditions improved (Figure 1C). In general, attempts to start or continue brooding without first provisioning were more likely to be accepted under poor conditions or by high provisioners. High provisioning murres had less of a decrease in body mass between the match and mismatch years, compared to low provisioners.
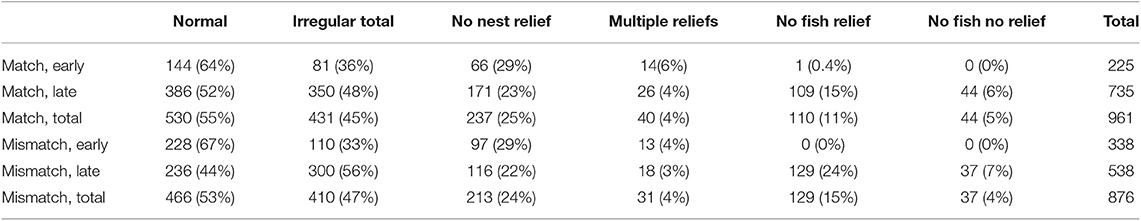
Table 1. Number of nest reliefs (percentage of total) early (week 1, before capelin arrival in the mismatch year) and late in chick rearing for Normal Nest Reliefs (returner brings fish, brooding exchange), Irregular total (four other categories in columns to right), No nest relief (returner brings fish), Multiple nest relief (at same visit by returner), No fish relief (returner does not bring fish, brooding exchange), No fish, no relief (returner does not bring fish, no brooding exchange).
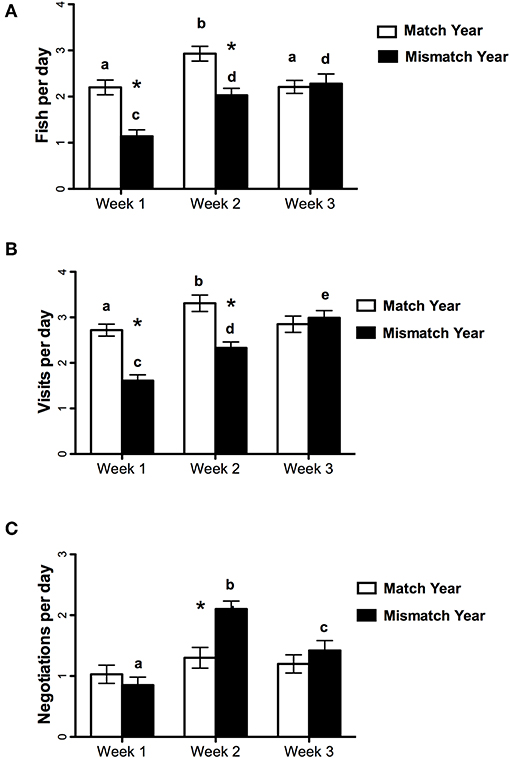
Figure 1. Mean (± SE) frequency of (A) chick provisioning, (B) total visits, and (C) negotiation for weeks 1–3 after chick hatching in the match year (1999) and mismatch year (2000). Different letters within the same year indicate significant differences over weeks whereas * indicates a significant difference between the same week in the 2 years.
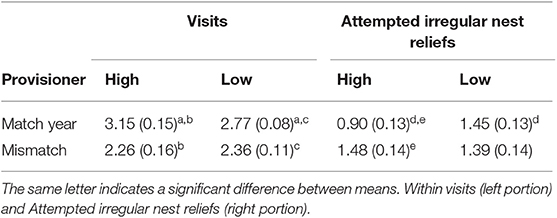
Table 2. Mean (SE) for visits (number per day) and attempted irregular nest reliefs (per day) for the match and mismatch year and for high and low provisioners.
Frequency of Irregular Nest Reliefs in Relation to Resource Abundance and Provisioner Status
Higher provisioning individuals within pairs were equally likely to be male or female. Female provisioning rate was 2.34 (0.11) fish per day in match year and 2.07 (0.11) fish per day in the mismatch year. Male provisioning rate was 2.53 (0.11) fish per day in the match year and 1.93 (0.19) fish per day in the mismatch year. Sex was not a factor in any significant main effects and two-way interactions, and results are reported with sex excluded. The two-way ANOVA with year (match, mismatch), weeks (1, 2, and 3), and provisioner status (high vs. low) as repeated factors showed that provisioning frequency was higher in match year than in the mismatch year [F(1, 13) = 63.84, p < 0.001] and there was a significant difference across weeks [F(2, 26) = 11.73, p = 0.005]. Although the provisioning rate was lower in the first week of both years, the rate was higher in both the first and second weeks of the match year compared to the same weeks in mismatch year [significant week by year interaction, F(2, 26) = 4.79 p = 0.017, first weeks (13) = 5.4, p < 0.001; second weeks, t(13) = 5.4, p < 0.001]. There was no difference in the third weeks (Figure 1A). The same pattern of main effects and interactions was observed for the total number of visits in the 2 years (Figure 1B) except that there was also a year by provisioner status interaction [F(1, 13) = 5.96, p = 0.03, Table 2]. Both high and low provisioners visited more often in the match year than the mismatch year [high provisioners, t(13) = 5.10, p < 0.001, low provisioners, t(13) = 3.58, p = 0.003]. High provisioners visited more than low provisioners in the match year [t(13) = 2.32, p = 0.02] but there was no difference in the mismatch year (p = 0.49).
Mean daily rates of irregular nest reliefs were overall lower in the match year than in the mismatch year [F(1, 13) = 5.92, p = 0.03, primarily due to increases by high provisioners, see the next paragraph] and rates changed across weeks [F(2, 26) = 10.59, p < 0.001]. Analysis of the significant year by week interaction [F(2, 26) = 4.39, p = 0.023] indicated that the rate of irregular nest reliefs did not change over weeks in the match year [F(2, 52) = 0.81, P = 0.5]. In contrast, the rate of irregular nest reliefs in the mismatch year differed across weeks [F(2, 52) = 16.0, P < 0.0001] with a lower rate in week 1, before capelin arrival, than in both weeks after capelin arrival, weeks 2 (p < 0.001) and 3 (p = 0.025). The rate in week 2 of the mismatch year was also higher than in week 3 of the same year (p = 0.008) and than in week 2 of the match year (p < 0.001, Figure 1C).
There was a significant year by provisioner status interaction in rates of initiating irregular nest reliefs [F(1, 13) = 5.74, p = 0.032, Table 2]. High provisioners initiated fewer irregular nest reliefs than low provisioners in the match year (p = 0.005) and their rate increased in the mismatch year (p = 0.003) to be no different from low provisioners in that year (p = 0.67). Rates of initiation of irregular nest reliefs by low provisioners did not differ between years (p = 0.75).
Compensation
We considered that an attempt to get the partner to work harder was successful (i.e., the partner compensated) if the returning bird was allowed to incubate without bringing a fish and if the brooder remained brooding when the mate brought a fish, such that the returning bird departed the colony without having a brooding bout. First, in contrast to the number of fish visits, there was no year difference in the number of no-fish visits [F(1, 26) = 0.58, p = 0.50], between high and low provisioners [F(1, 26) = 4.0, p = 0.06] and the interaction was also not significant [F(1, 26) = 0.32, p = 0.58]. Low provisioners returned without a fish in a higher proportion of visits than high provisioners [F(1, 26) = 5.37, p = 0.03]. The proportion of no-fish visits did not differ by year [F(1, 26) = 1.60, p = 0.22] and the interaction was not significant [F(1, 26) = 0.32, p = 0.53].
There was a higher proportion of no-fish visits with nest reliefs (successful attempts to brood without provisioning) in the mismatch year than match year [F(1, 26) = 5.37, p = 0.03]. Mates returning with a fish were also less often prevented from brooding in the mismatch year compared to the match year [F(1, 26) = 7.34, p = 0.012]. There were fewer brooding preventions in week 1 than in weeks 2 and 3 [F(1, 26) = 6.30, p = 0.019]. The significant year by provisioner status interaction [F(1, 26) = 4.39, p = 0.046] indicated that in the match year, high provisioning returners that brought a fish allowed their mates to continue brooding more often than did low provisioners, whereas there was no difference due to provisioner status the mismatch year.
Timing Components
Total time of each interaction was used in these analyses rather than time to the nest relief as the latter measure would have excluded all the interactions without nest reliefs. The two measures are significantly correlated [r(1, 316) = 0.73, p < 0.0001]. Overall, the total time of nest interactions differed with the type of regular or irregular nest relief [F(4, 1, 832) = 53.95, p < 0.0001]. Normal nest reliefs were shorter than every other nest relief type (multiple nest reliefs, no fish nest interactions with and without nest reliefs, p < 0.001) except for interactions where the returner brings a fish but the brooder does not allow a nest relief (p = 0.28).
Linear regression was used to determine what factors might contribute to variation in the total nest interaction time (dependent variable) with predictor variables as time of last visit by self and mate, time of day and time in chick rearing (F = 16.99, P < 0.001; R2 = 0.06). All predictor variables were significant indicating that nest relief sequences were shorter when the mate had been away longer, when the brooder had been sitting longer, when the nest relief took place earlier in the day and when they occurred later in chick rearing (Table 3).
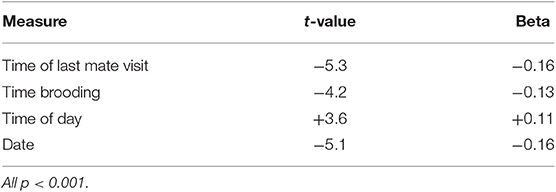
Table 3. Results of linear regression analysis of factors affecting total interaction time.
LMM was used to examine the total time in a nest relief interaction with year, before or after week 1, and nest relief type (all reliefs with fish; normal, no nest relief and multiple nest relief) as the repeated factors and provisioner status as the between subject factor. Of the main effects, only nest relief type was significant [F(2, 999) = 116.7, p < 0.001] and here and in all significant interactions involving this variable, one difference was due to the multiple nest reliefs taking longer than both the normal and no nest relief interactions (all ps < 0.001). There was a significant nest relief type by year interaction [F(2, 999) = 20.7, p = 0.003] indicating that normal nest reliefs were the same duration in both years (p = 0.48) whereas multiple nest reliefs took longer in the mismatch than in the match year [t(39) = 5.50, p < 0.001] and interactions without a nest relief were shorter in the mismatch year than in the match year [t(270) = 3.25, p = 0.001, Figure 2A]. Finally, there was also a significant nest relief by provisioner status interaction wherein multiple nest relief interactions were longer when low provisioners brooded than when high provisioners brooded [t(39) = 3.04, p = 0.004, Figure 2B] but there were no provisioner differences for the other nest relief types (ps > 0.10).
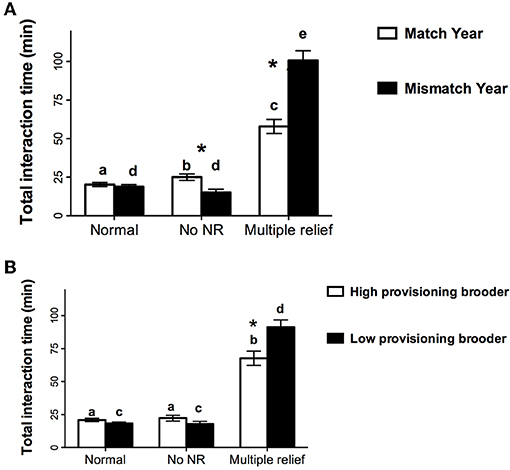
Figure 2. Mean (±SE) time for the three types of nest interactions with fish for (A) the match (1999) and mismatch (2000) years and (B) when the brooder was the high or low provisioning member of the pair. Different letters for bars of the same color indicate significant differences among nest relief types whereas * indicates a significant difference for the same nest relief type between years (A) or between high and low provisioners (B).
There were significant interactions between year and before vs. after the first week [F(1, 999) = 51.2, p < 0.001] and between year and provisioner status [F(1, 999) = 18.64, p < 0.001]. Nest relief interactions were shorter when resources were scarce: early compared to late in mismatch year [t(432) = 4.57, p < 0.001] and early in the mismatch year compared to early in the match year [t(407) = 2.08, p = 0.038]. Nest interactions were also longer later in the mismatch year than they were later in the match year [t(612) = 8.00, p = 0.001, Figure 3A]. Low provisioning brooders had longer nest relief interactions in the mismatch year than they had in the match year [t(543) = 6.02, p < 0.001] and longer nest relief interactions than high provisioners in the mismatch year [t(432) = 3.97, p < 0.001, Figure 3B]. Nest relief durations for high provisioning brooders did not change between years (p = 0.71).
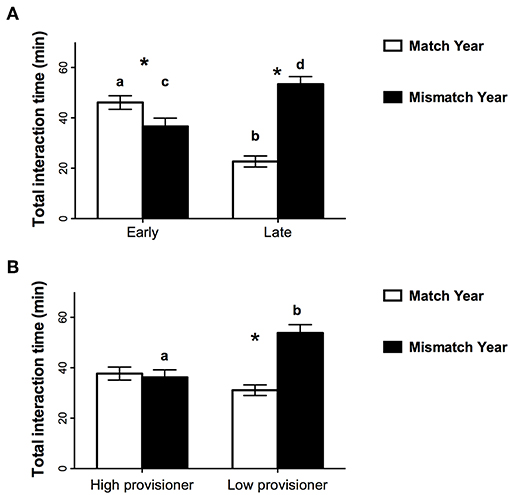
Figure 3. Mean (±SE) time for nest interactions with fish in the match (1999) and mismatch (2000) years (A) before (early) and after (late) capelin arrived inshore and (B) for low and high provisioning brooding murres. Different letters for bars of the same color indicate significant differences between early and late in the season (A) and high and low provisioners (B) whereas * indicates a significant difference between years.
Using LMM, the time between visits was compared for each of the two directly comparable situations: first, when the returner came back with a fish (with and without a nest relief) and second, when it did not (again with and without a nest relief). The analyses only included inter-visit times where the current and previous visits occurred on the same day. Analyses of four inter-visit times were conducted: Time since (a) the last visit by self for the returner and (b) the last visit by the returner's mate, the current brooder, as well as time since (c) the returner brought a fish and (d) since the mate brought a fish. In seven of eight analyses (fish or no fish × four inter-visit times), the times since previous visits were significantly shorter when the brooder prevented a nest relief than when the brooder allowed one (Table 4). We found that the times since the last mate's visit and feed were significantly longer in the mismatch year than in match year, but in cases where no nest relief occurred, there were no year differences in times to last mate visit [significant interaction, F(1, 26) = 4.39, p = 0.046]. In both cases the times since the last visit or feed were greater for the nest relief than no nest relief occurrences (as in the main analyses).
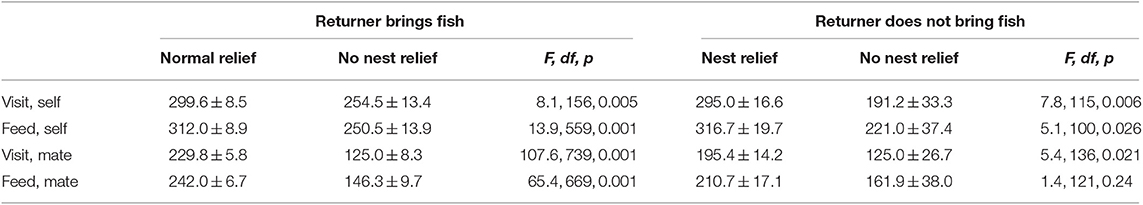
Table 4. Mean ± se for time (min) between visits and feeds for focal birds (self) and their mates.
Variation in Provisioning and Body Condition
Murres categorized as high and low provisioners within each pair during the match year changed their chick-provisioning rates differently during the following predator-prey mismatch year [year × provisioner: F(1, 28) = 4.69, p = 0.04]. Previously high provisioners showed a larger mean decrease in provisioning rate (0.63 ± 0.13 fewer fish per day) than low provisioners (0.23 ± 0.13 fewer fish per day, t = 2.17, P = 0.039, Table 5).
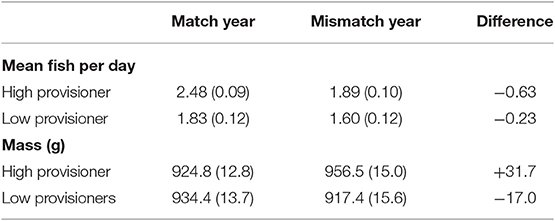
Table 5. Low-provisioning common murres decreased their mean (±SE) daily provisioning rate less and lost more body mass than their high provisioning mates in the predator-prey mismatch year (2000) compared to the match year (1999).
Cost of provisioning and turn-taking strategies was evaluated by comparing the body mass of the same high and low provisioning murres matched for chick age in both years (N = 23). The body mass of low provisioners did not differ by year [F(1, 21) = 0.68, p = 0.42] or provisioning status [F(1, 21) = 0.67, p = 0.42] but the interaction was significant [F(1, 21) = 7.29, p = 0.013]. Mass did not differ by provisioning status in the match year but was significantly higher for high provisioners than for low provioners in the mismatch year (Table 5). Birds with larger drops in feeding rate from the match to the mismatch year had smaller mass decreases (r = −0.51, p = 0.04).
Discussion
The rates of no-fish visits and refused changeovers varied with prey availability in the mismatch year with low rates observed during the first post-hatch week before capelin arrived inshore. Rates increased after the first week in the mismatch year once capelin became more available. In contrast, in the match year, where prey levels were more stable and abundant, there were no weekly changes in rates of irregular nest reliefs. High provisioners initiated fewer irregular nest reliefs than low provisioners in the match year but increased their rate in the mismatch year. High provisioners retained their body mass in the mismatch year whereas low provisioners did not. Overall, more attempts to start or continue brooding without first provisioning were accepted in the mismatch than in the match year.
Although provisioning rates were still lower in week 2 in the mismatch year than in the match year, it is interesting to note that the rate of irregular nest reliefs was higher. This higher rate in week 2 of the mismatch year suggests a rebound effect of increased attempts to push the mate to do more once prey suddenly became more available in the mismatch year. Consistent with these data, nest interactions were shorter early in the mismatch year compared to later that year and early in the match year. Our prediction of fewer irregular nest reliefs early in the mismatch year compared to the match year was not supported, possibly because feeding rates early in the match year, while higher than for early in the mismatch year, were still lower than later in the season. These results generally support the prediction that mates in biparental species should push their partner more when resources are plentiful, as they attempt to perform closer to their “laziness threshold,” and cooperate more with their mates when resources are scarce (Jones et al., 2002). Total visits and fish visits generally mirrored the pattern of irregular nest reliefs in the mismatch year: high when rates of irregular nest reliefs increased and low when resources were limited.
In general, mates accepted their partners' attempts to brood more often in the mismatch year than in the match year: brooders more often allowed nest reliefs when their mates did not bring a fish and returners more often allowed mates to remain brooding even when the returner had provisioned the chick. Since the time between mate visits was also longer in the mismatch year (when prey was harder to find) than in the match year, there are two (not mutually exclusive) possible explanations. First, faster and more frequent nest reliefs may have been allowed in the mismatch year because the brooder was compensating for its returning mate under poor conditions. Alternatively, since mates were away longer on average, nest reliefs may have been more likely to occur because birds with longer brooding bouts in the mismatch year may have been hungrier than in the match year. The exception, that high provisioners accepted more attempts to brood than low provisioners in the match year, fits the rest of these findings: murres compensate when they can (high provisioners in the match year) or must to save their partners (all murres in the mismatch year).
High provisioners initiated fewer irregular nest reliefs than their low-provisioning mates during the predator-prey match year. This difference disappeared the following breeding season, when feeding conditions were unfavorable due to capelin arriving well after chick-hatching; previous high provisioners initiated more irregular nest reliefs than they had during favorable breeding conditions at levels similar to those of the low provisioners. Although all birds brought in fewer fish in the mismatch year than in the match year, high provisioners decreased their provisioning rates more than low provisioners and they experienced less mass loss. Similarly, low provisioners returned to the nest without a fish on a higher proportion of visits than did high provisioners. Taken together these results suggest that high provisioners respond to their partners' attempts to brood by compensating when they can in favorable feeding years. That nest relief patterns change within and between seasons in the same pairs, apparently in response to changes in required foraging effort, suggests on-going negotiation (Lessels and McNamara, 2012; Johnstone et al., 2014) rather than the one-timed “sealed bid” (Houston and Davies, 1985; Schwagmeyer et al., 2002).
Failure to coordinate parental duties can have consequences that are more far reaching than the current breeding season. Divorce in common murres occurs when high-provisioning birds leave mates that have a history of low provisioning and re-pair with high-provisioning recently-widowed neighbors (Moody et al., 2005). Divorce victims have long re-pairing latencies (usually not until the next season, Moody et al., 2005), suggesting that there are serious costs of pushing the mate too much or not responding to the partner's attempts to obtain more brooding time. We propose that co-attendance behavior is the mechanism for negotiating provisioning patterns within long-term pair bonds. Consistent with this proposal, both victim and choosers in divorcing pairs had significantly longer latencies to nest reliefs than stable pairs (Moody et al., 2005), suggesting that both pair members in unstable pairs were unsuccessfully attempting to brood more themselves and push their partners to do more provisioning. It may be useful, therefore, to take a longer view than a single breeding season in order to understand pair dynamics in species with long-term pair bonds.
There was no difference in the mean duration of normal nest reliefs and the sequences in which the brooder did not allow a nest relief after a fish delivery, as was also found for common murres in Takahashi et al. (2017). In the previous study, the main difference between the two types of nest interactions was in the frequency and distribution of allopreeening. In the normal nest relief sequences both partners started allopreening each other early in the interaction at approximately the same time. In contrast, in the no nest relief and no fish sequences, the brooder delayed the onset of allopreening until well after the returner initiated the behavior. Takahashi et al. (2017) concluded that allopreening by the returning mate may facilitate a nest relief whereas delayed onset and low allopreening frequency by the brooder may signal the brooder's attempts to delay or prevent a nest relief. Similarly, Boucaud et al. (2017) showed that duet characteristics in zebra finches (Taeniopygia guttata) predicted whether or not a relief would occur.
Nest reliefs were more likely to occur when a long time had elapsed between the previous visit and the current one, whether or not the returner brought a fish. Mean brooding time was longer (i.e., more time elapsed since the mate had returned to the colony) for successful nest reliefs in the mismatch compared to the match year, whereas time since last visit did not differ by year for unsuccessful attempts to brood. These results suggest that the nest relief proceeds when the brooder has spent a long enough time in the colony to be hungry.
Our results suggest a re-interpretation of the function of the increased co-attendance times seen in murres under good feeding conditions. Changes in co-attendance time in response to changes in resources has traditionally been viewed in terms of amount of resting or “loafing time” (Cairns et al., 1987; Monaghan et al., 1994; Uttley et al., 1994; Zador and Piatt, 1999). Consistent with this view, Harding et al. (2007) suggest that extra colony time for murres under good foraging conditions may particularly benefit individuals that are more efficient foragers. Our data suggest an alternative: under good feeding conditions, low provisioners are initiating more irregular nest reliefs and they are benefiting from the compensation from their mates. Nest relief interactions take significantly longer under good conditions than under poor ones, suggesting that, in the former, mates express greater conflict with each other. Thus, a considerable proportion of co-attendance may be ongoing negotiation rather than loafing.
Less mass decrease in high provisioners than in low provisioners suggests that high provisioners have a greater mass cushion in that they can change behavior when resources are less abundant and not lose additional body mass in ways that low provisoners, existing closer to abandonment threshold proposed by Jones et al. (2002), cannot. Results in Takahashi et al. (2017) support this contention: brooding murres that delayed nest reliefs had lower body mass and higher betahydroxybutyrate levels (a measure of mass loss as stored lipids are mobilized). Taken together, the results suggest the provisioner status used here, based on behavioral differences, has a physiological basis.
Conclusions
We present evidence that the rates of initiating irregular nest reliefs change with resources, increasing as the resource base improves. Supporting evidence was strongest within the mismatch year and less in line with our predictions between years. High provisioning birds change more than low provisioners in that they compensate for their partners under good conditions but take care of themselves (increase attempts to brood more, decrease provisioning and maintain body mass) when resources are limited, which suggests that they have more of a buffer than their low provisioning mates. The greater co-attendance time when foraging conditions are good result from visits being longer (early vs. late in the mismatch year, particularly for low provisioners) and more frequent (match vs. mismatch year). Our data support the idea that extensive co-attendance under good conditions may be more about negotiation rather than extra loafing time since behavior varied with nest relief type and individual quality. However, if the “loafing time” idea were to be developed to include quality differences that reflect which individuals need to forage less and/or rest more, loafing and negotiation might not be incompatible explanations for the variation in co-attendance behavior. It is interesting to note that what we call a mismatch year in this paper is not nearly as bad as some of the subsequent years (Storey et al., 2017; Takahashi et al., 2017). As the timing and abundance of capelin spawning become more variable with climate-change induced temperatures fluctuations, more frequent serious mismatch years are expected to occur.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Ethics Statement
The animal study was reviewed and approved by the Institutional Animal Care Committee of Memorial University. Permits were granted by the Government of Newfoundland and Labrador and the Canadian Wildlife Service (CWS) of Environment Canada.
Author Contributions
AS, SW, and CW made substantial contributions to data acquisition and interpretation, as well as manuscript preparation. AS and CW contributed to data analysis.
Funding
Research was funded by NSERC Postgraduate Scholarships and Memorial University Fellowships (SW and CW) and an NSERC Discovery Grant to AS.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Maureen Cameron-McMillan, Allison Moody, Joel Heath, and Kelly Squires for assistance in gathering the behavioral data and Tommy Reddick for boat transport. We are grateful to Gregory Robertson of CWS for logistic support and to Ian Jones for advice on scale design.
References
Ainley, D. G., Nettleship, D. N., Carter, H. R., and Storey, A. E. (2002). “Common murres (Uria aalge),” in The Birds of North America, eds A. F. Poole and F. B. Gill (Ithaca, NY: Cornell Lab of Ornithology). doi: 10.2173/bna.666
Beaumont, R. (2012). Analysing Repeated Measures With Linear Mixed Models (Random Effects Models) (1) Getting Familiar With the Linear Mixed Models (LMM) Options in SPSS. Available online at: http://www.floppybunny.org/robin/web/virtualclassroom/stats/statistics2/repeated_measures_1_spss_lmm_intro.pdf (accessed April 3, 2019).
Bebbington, K., and Hatchwell, B. J. (2016). Coordinated parental provisioning is related to feeding rate and reproductive success in a songbird. Behav. Ecol. 27, 652–659. doi: 10.1093/beheco/arv198
Birt-Friesen, V. L., Montevecchi, W. A., Cairns, D. K., and Macko, S. A. (1989). Activity-specific metabolic rates of free-living northern gannets and other seabirds. Ecology 70, 357–367. doi: 10.2307/1937540
Boucaud, I. C. A., Perez, E. C., Ramos, L. S., Griffith, S. C., and Vignal, C. (2017). Acoustic communication in zebra finches signals when mates will take turns with parental duties. Behav. Ecol. 28, 645–656. doi: 10.1093/beheco/arw189
Cairns, D. K., Bredin, K. A., and Montevecchi, W. A. (1987). Activity budgets and foraging ranges of breeding common murres. Auk 104, 218–224. doi: 10.1093/auk/104.2.218
Cameron-MacMillan, M. L., Walsh, C. J., Wilhelm, S. I., and Storey, A. E. (2007). Male chicks cost more to rear than females in a mongamous seabird, the common murre. Behav. Ecol. 18, 81–85. doi: 10.1093/beheco/arl048
Chastel, O., Weimerskirch, H., and Jouventin, P. (1995). Body condition and seabird reproductive performance: a study of three petrel species. Ecology 76, 2240–2246. doi: 10.2307/1941698
Doody, L. M., Wilhelm, S. I., McKay, D. W., Walsh, C. J., and Storey, A. E. (2008). The effects of variable foraging conditions on common murre (Uria aalge) parental behavior and corticosterone concentrations. Horm. Behav. 53, 140–148. doi: 10.1016/j.yhbeh.2007.09.009
Elliott, K. H., McFarlane-Tranquilla, L., Burke, C. M., Hedd, A., Montevecchi, W. A., and Anderson, W. G. (2012). Year-long deployments of small geolocators increase corticosterone levels in murres. Mar. Ecol. Prog. Ser. 466, 1–7. doi: 10.3354/meps09975
Gabrielsen, G. W. (1996). “Energy expenditure of breeding common murres,” in Studies of High-Latitude Seabirds. 4. Trophic Relationships Andenergetics of Endotherms in Cold Ocean Systems, ed W. A. Montevecchi (Ottawa, ON: Canadian Wildlife Service Occasional Paper Number 91, 49–58.
Hamel, N. J., Parrish, J. K., and Conquest, L. L. (2004). Effects of tagging on behavior, provisioning, and reproduction in the common murre (Uria aalge), a diving seabird. Auk 121, 1161–1171. doi: 10.1093/auk/121.4.1161
Harding, A. M., Piatt, J. F., Schmutz, J. A., Shultz, M. T., Van Pelt, T. I., Kettle, A. B., et al. (2007). Prey density and the behavioral flexibility of a marine predator: the common murre (Uria aalge). Ecology 88, 2024–2033 doi: 10.1890/06-1695.1
Harris, M. P., Wanless, S., and Webb, A. (2000). Changes in body mass of Common Guillemots Uria aalge in southeast Scotland throughout the year: implications for the release of cleaned birds. Ringing Migr. 20, 134–142. doi: 10.1080/03078698.2000.9674235
Houston, A. I., and Davies, N. B. (1985). “The evolution of cooperation and life history in the dunnock, Prunella modularis,” in Behavioural Ecology, eds R. M. Sibly and R. M. Smith (Oxford: Blackwell Scientific), 471–487.
Houston, A. I., Székely, T., and McNamara, J. M. (2005). Conflict between parents over care. Trends Ecol. Evol. 20, 33–38. doi: 10.1016/j.tree.2004.10.008
Jacobs, S. R., Edwards, D. B., Ringrose, J., Elliott, K. H., Weber, J.-M., and Gaston, A. J. (2011), Changes in body composition during breeding: reproductive strategies of three species of seabirds under poor environmental conditions. Comp. Biochem. Physiol. B 158, 77–82. doi: 10.1016/j.cbpb.2010.09.011
Johnstone, R. A., Manica, A., Fayet, A. L., Stoddard, M. C., Rodriguez-Gironés, M. A., and Hinde, C. A. (2014). Reciprocity and conditional cooperation between great tit parents. Behav. Ecol. 25, 216–222. doi: 10.1093/beheco/art109
Jones, K. M., Graeme, D., Ruxton, D., and Monaghan, P. (2002). Model parents: is full compensation for reduced partner nest attendance compatible with stable biparental care? Behav. Ecol. 13, 838–843. doi: 10.1093/beheco/13.6.838
Kitaysky, A. S., Wingfield, J. C., and Piatt, J. F. (1999). Dynamics of food availability, body condition and physiological stress response in breeding Black-legged Kittiwakes. Funct. Ecol. 13, 577–584. doi: 10.1046/j.1365-2435.1999.00352.x
Lessels, C. M., and McNamara, J. M. (2012). Sexual conflict over parental investment in repeated bouts: negotiation reduces overall care. Proc. R. Soc. B 279, 1506–1514. doi: 10.1098/rspb.2011.1690
Monaghan, P., Uttley, J. D., Burns, M. D., Thaine, C., and Blackwood, J. (1989). The relationship between food supply, reproductive effort and breeding success in Arctic Terns Sterna paradisaea. J. Anim. Ecol. 58, 261–274. doi: 10.2307/4999
Monaghan, P., Walton, P., Wanless, S., Uttley, J. D., and Burns, M. D. (1994). Effects of prey abundance on the foraging behaviour, diving efficiency and time allocation of breeding guillemots Uria aalge. Ibis 136, 241–222. doi: 10.1111/j.1474-919X.1994.tb01087.x
Moody, A. T., Wilhelm, S. I., Camerson-MacMillan, M. L., Walsh, C. J., and Storey, A. E. (2005). Divorce in common murres (Uria aalge): relationship to parental quality. Behav. Ecol. Sociobiol. 57, 224–230. doi: 10.1007/s00265-004-0856-8
Paredes, R., Jones, I. L., and Bonness, D. (2005). Reduced parental care, compensatory behavior and reproductive costs of thick-billed murres equipped with data loggers. Anim. Behav. 69, 197–208. doi: 10.1016/j.anbehav.2003.12.029
Robertson, G. J., Storey, A. E., and Wilhelm, S. I. (2006). Local survival rates of common murres breeding in witless bay, newfoundland. J. Wildlife Manage. 70, 584–587.
Savage, J. L., Browning, L. E., Manica, A., Russell, A. F., and Johnstone, R. A. (2017). Turn-taking in cooperative offspring care: by-product of individual provisioning behavior or active response rule? Behav. Ecol. Sociobiol. 71:162. doi: 10.1007/s00265-017-2391-4
Schwagmeyer, P. L., Mock, D. G., and Parker, G. A. (2002). Biparental care in house sparrows: negotiation or sealed bid? Behav. Ecol. 13, 713–721. doi: 10.1093/beheco/13.5.713
Storey, A. E., Ryan, M. G., Fitzsimmons, M. G., Kouwenberg, A.-L., Takahashi, L. S., Robertson, G. J., et al. (2017). Balancing personal maintenance with parental investment in a chick-rearing seabird: physiological indicators change with foraging conditions. Conserv. Physiol. 5, 1–12. doi: 10.1093/conphys/cox055
Takahashi, L. S., Storey, A. E., Wilhelm, S. I., and Walsh, C. J. (2017). Turn-taking ceremonies in a colonial seabird: does behavioral variation signal individual condition? Auk 134, 530–541. doi: 10.1642/AUK-17-26.1
Uttley, J. D., Walton, P., Monaghan, P., and Austin, G. (1994). The effects of food abundance on breeding performance and adult time budgets of guillemots Uria aalge. Ibis 136, 205–213. doi: 10.1111/j.1474-919X.1994.tb01086.x
Wanless, S., Harris, M. P., and Morris, J. A. (1988). The effect of radio transmitters on the behavior or common murres and razorbills during chick rearing. Condor 90, 816–823. doi: 10.2307/1368838
Weimerskirch, H., and Lys, P. (2000). Seasonal changes in the provisioning behaviour and mass of male and female wandering albatrosses in relation to the growth of their chick. Polar Biol. 23, 733–744. doi: 10.1007/s003000000144
Weimerskirch, H., Zimmermann, L., and Prince, P. A. (2001). Influence of environmental variability on breeding effort in a long-lived seabird, the Yellow-nosed Albatross. Behav. Ecol. 12, 22–30. doi: 10.1093/oxfordjournals.beheco.a000374
Wilhelm, S. I. (2004). Behavioural and physiological responses of breeding common murres (Uria aalge): exploring inter-annual variability within individuals (doctoral thesis). Memorial University of Newfoundland, St. John's, NL, Canada.
Wilhelm, S. I., and Storey, A. E. (2002). Influence of cyclic pre-lay attendance of synchronous breeding in common murres. Waterbirds 25, 156–163. doi: 10.1675/1524-4695(2002)025[0156:IOCPAO]2.0.CO;2
Wilhelm, S. I., Walsh, C. J., and Storey, A. E. (2008). Time budgets of common murres vary in relation to changes in inshore capelin availability. Condor 110, 316–324. doi: 10.1525/cond.2008.8440
Keywords: negotiation, parental investment, food availability, common murre, Uria aalge, nest relief
Citation: Storey AE, Wilhelm SI and Walsh CJ (2020) Negotiation of Parental Duties in Chick-Rearing Common Murres (Uria aalge) in Different Foraging Conditions. Front. Ecol. Evol. 7:506. doi: 10.3389/fevo.2019.00506
Received: 17 June 2019; Accepted: 13 December 2019;
Published: 14 January 2020.
Edited by:
Camilla Anne Hinde, Wageningen University and Research, NetherlandsReviewed by:
Katarzyna Wojczulanis-Jakubas, University of Gdansk, PolandNina Jane Karnovsky, Pomona College, United States
Copyright © 2020 Storey, Wilhelm and Walsh. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anne E. Storey, YXN0b3JleUBtdW4uY2E=