 E. R. Jasper Wubs
E. R. Jasper Wubs Tom van Heusden
Tom van Heusden Pauline D. Melchers1
Pauline D. Melchers1 T. Martijn Bezemer
T. Martijn Bezemer
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 26 November 2019
Sec. Behavioral and Evolutionary Ecology
Volume 7 - 2019 | https://doi.org/10.3389/fevo.2019.00451
This article is part of the Research Topic The Next Step: Disentangling the Role of Plant-Soil Feedbacks in Plant Performance and Species Coexistence Under Natural Conditions View all 15 articles
Plant-soil feedbacks (PSFs) are important drivers of vegetation composition in nature. Whole-soil inoculations can help to steer plant-community assembly for nature restoration success. However, it is unclear how subsequent colonization by ruderal and late-successional plant species influences PSFs. Here we test the direction and strength of the PSFs for ruderal and target plant species on differently inoculated soils. We hypothesize that inoculation with late-successional field soil promotes positive and negative PSFs for late-successional and ruderal plants, respectively. We conducted a glasshouse experiment with three plant-growth phases. First, we inoculated a common nutrient-rich ex-arable recipient soil with either a heathland, grassland or an arable soil, and grew mixtures of three ruderal, and three late-successional target species in all soils. Subsequently, we divided the experimental units over four new pots and planted half with mixtures of three ruderal and half with mixtures of three late-successional plant species, to simulate local colonization by these species groups. After 9 weeks, we removed the plants and replanted the pots with ruderal and late-successional species mixtures in a full factorial design to quantify the induced PSFs, after a further 9 weeks of growth. We found that ruderal plants developed positive PSF on soils inoculated with arable soil and neutral feedback when soils were inoculated with grassland or heathland inoculum. The positive PSF was most pronounced for Myosotis arvensis, although all ruderal species showed the most positive PSFs on arable-inoculated soils. There was no significant effect of inoculation on the PSF of late-successional species. As a result of changing PSFs, the relative performance of ruderal and late-successional target species became more similar on soils inoculated with grassland or heathland inoculum, leading to higher evenness. We conclude that soil inoculation on nutrient-rich ex-arable soils can affect PSFs. Inoculation with grassland or heathland inoculum prevented ruderal species from developing positive PSF and shifted the competitive balance in favor of late-successional plants. Future studies need to address the longer-term dynamics of soil inoculation-induced shifts in PSFs, particularly because these are expected to develop over longer timescales in the slow-growing late-successional target species, as well as the impact of competitive plant-plant interactions.
Plants and their associated soil biota are continuously interacting and their interplay can lead to net positive and negative effects on plant performance (survival, growth, and reproduction). These phenomena are termed positive and negative plant-soil feedback (PSF), respectively (Bever, 1994; Van der Putten et al., 2013). It is clear that plant-soil feedback effects are important drivers of plant community dynamics, both in the lab (Kulmatiski et al., 2008; Lekberg et al., 2018) and in the field (Bennett et al., 2017; Teste et al., 2017). However, it is becoming increasingly clear that the realized plant-soil feedback is dependent on the local conditions. For instance, plant-soil feedback strengths change with soil type, fertilization and herbivory (Bezemer et al., 2006; Veen et al., 2014; Heinze and Joshi, 2017; Wubs and Bezemer, 2018b). The realized plant-soil feedback is not a property of the plant species per se, but arises through the interplay between plants and their soil biota within the local biotic and abiotic conditions. The latter is illustrated by the observation that PSFs change importantly with the successional stage of the plant as well as the soil (Kardol et al., 2006).
Recently, we applied this contextual knowledge on plant-soil interactions to improve nature restoration success through soil inoculation on sandy soils (Wubs et al., 2016). Previous studies show that late-successional plant species, the typical target species for restoration, develop positive PSF, measured as plant biomass (De Deyn et al., 2003; Carbajo et al., 2011) and this is particularly pronounced in late-successional soils (Kardol et al., 2006). In field experiments it has been shown that introduction of late-successional soil communities can facilitate the establishment and growth of late-successional plant species (Vécrin and Muller, 2003; Pywell et al., 2011; Buisson et al., 2018) and can determine the composition of the developing plant community for decades (Wubs et al., 2016, 2019). While this approach can thus be a successful intervention method, the areas to be restored are still open to potential colonization by plant species from elsewhere and it is unclear how soil inoculation affects the PSFs experienced by subsequently colonizing plants.
Here we test whether inoculation with soils from early- and late-successional systems affect the direction and strength of the PSFs, measured as plant biomass, of ruderal as well as restoration target plant species. We use the sandy glacial deposits in the central parts of the Netherlands as our model system (Kardol et al., 2006; Carbajo et al., 2011; Wubs et al., 2016). In the area, arable farming has been intensive in many places since World War 2, but many farms are no longer economically viable due to the low inherent soil fertility and limits on spatial expansion. Species-rich grasslands and dry heathlands are important national restoration targets for the region, harboring important flora and fauna typical of dry habitats. Both ecosystems are stages along the secondary succession on sandy soils, but under different management regimes (Kardol, 2007). The grasslands arise from the arable fields within a span of 10–30 years under mowing or grazing regimes. The dry heaths in this system are the result of, historically prolonged, sod cutting and grazing. We refer to the arable system as early-succession, and the grassland and heathland both as late-succession.
Plant-soil feedbacks are typically studied using two-phase experiments (Kulmatiski et al., 2008; Brinkman et al., 2010). In the first phase, a plant species or group of species is allowed to condition the soil by growing in it, which alters the local soil biotic and abiotic conditions (Ehrenfeld et al., 2005; Van der Putten et al., 2013): the conditioning phase. In the second phase, the test phase, the same (or a different) group of plant species is allowed to grow on that soil and metrics of their performance on conditioned and control soil are recorded (e.g., survival, biomass, reproduction). Based on the difference in plant performance on the conditioned and a control soil the net PSF is calculated (Brinkman et al., 2010), with positive PSF indicating better performance in the conditioned soil than in the control and negative PSF the converse. In this study, we couple a soil inoculation experiment (Phase 1) with a classical PSF experiment, with a conditioning phase (Phase 2) and a test phase (Phase 3). In the first phase we inoculated a common ex-arable soil with either arable, grassland or heathland soil and grew a common plant community of six species to simulate what would happen in a soil inoculation based restoration project (Wubs et al., 2018). After harvest, we then separately grew three ruderal and three target plant species on all the inoculated soils to allow soil conditioning by the ruderal and target species groups, respectively (Phase 2). Finally, after harvest, we again grew the same ruderal and target species groups on each of the conditioned soils in a full factorial design and quantified their performance as shoot biomass production (Phase 3). This design allows us to quantify net PSFs, by comparing performance of plants grown on soils conditioned by the same group of plants to performance on soils conditioned by the other group of plants, across the three different inoculation treatments. We hypothesize that late successional plants experience positive PSF, while ruderal species develop negative PSF (Kardol et al., 2006). Furthermore, when inoculated with late-successional field soil the positive and negative PSFs for late-successional and ruderal plants, respectively, are expected to be exacerbated. This would result in even higher and lower plant biomass for these groups.
We conducted a glasshouse experiment with soils setup in a prior study (Wubs et al., 2018). More details on this part of the methods can be found there. Briefly, in the experiment inocula of three ecosystem types, arable, grassland, and heathland, were sourced from the field (January 2015), with three replicate fields in each type. Within each field an area of 5 × 5 m was selected at least 20 m from the edge of the field. At each corner of the selected area, 5 kg of soil was collected from the upper 10–15 cm. The soil was sieved over a 1 cm mesh to remove stones and large roots. Upon return to the lab, the four samples per field were pooled based on equal amounts of dry weight resulting in homogenized inoculum material of 20 kg per field. These inocula were introduced (1:9 w:w inoculum:soil ratio) into a common ex-arable field soil (total 4 kg/container). The common ex-arable soil was from a field that had been in intensive agricultural use at least since World War 2 until 2004. Then, it was used for extensive wheat cultivation for 2 years prior to the implementation of large-scale nature restoration measures in 2006 (Wubs et al., 2016). The soil was collected from the central part of the field, where the only management consisted of cattle grazing (25–30 cows throughout the year, roaming freely in the entire 160 ha field) and removal of tree seedlings (particularly Betula spp. and Prunus serotina). We collected soil from the organic layer within 10–50 cm depth (~1,300 kg), which was subsequently sieved over a 1 cm mesh to remove major roots and stones and homogenized. The common background soil was sterilized (>25 KGray gamma radiation, Isotron, Ede, the Netherlands) to eliminate the resident soil community. The inocula were not sterilized. The abiotic conditions in the inocula and the common ex-arable soil have been reported in Table S1 of Wubs et al. (2018). Likewise, soil biotic composition data reported previously by Wubs et al. (2016) are available on Figshare (doi: 10.6084/m9.figshare.3435404). The major difference were that bacterial and fungal biomass were lower on the arable soil compared to grassland and heathland (their Table S4) and the microbial and nematode community composition was strongly different (their Figures 1f,g).
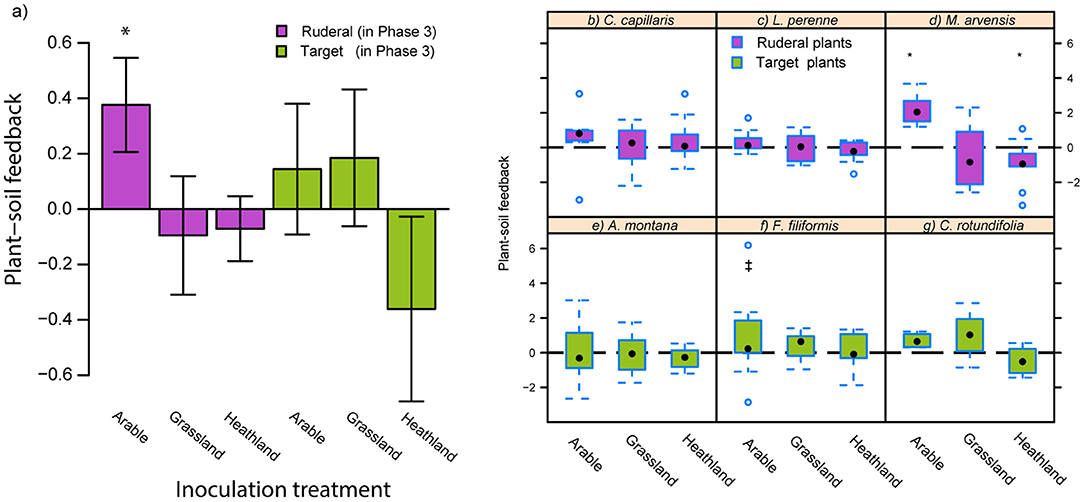
Figure 1. Mean (± SE) plant species group PSFs in Phase 3 based on shoot biomass (a), boxplots showing PSF per species (b–g). The box extends from the lower to the upper quartile, while the thick dot represents the median. The whiskers represent the 5th and 95th percentile. Any data points beyond these cut-offs are individually presented. Plants grew in mixtures of ruderal and late-successional plant species, respectively, in pots in Phase 3. Significance of PSF effects are based on planned contrasts (*p < 0.05, ‡p = 0.0669; Tables S2, S3).
The containers (17 × 17 × 17 cm) were planted with two seedlings each of three ruderal and three late-successional target species (12 plants per pot). Three species were early-successional ruderals: Crepis capillaris (L.) Wallr. (Asteraceae), Lolium perenne L. (Poaceae) and Myosotis arvensis (L.) Hill (Boraginaceae), and three were late-successional, conservation target species: Arnica montana L. (Asteraceae), Festuca filiformis Pourr. (Poaceae) and Campanula rotundifolia L. (Campanulaceae), with one grass and two forbs in each group. Seeds were obtained from commercial suppliers of wild plant seeds (Cruydthoeck, Assen, the Netherlands and B&T World Seeds, Paguignan, France) and germinated (sterilized 1 min. in 5% NaClO solution) on moistened glass beads in a climate chamber (12 h light/dark cycle, 20°C by day and 15°C at night). The mixed plant communities were allowed to grow for 7 weeks (Phase 1) and subsequently shoot biomass was harvested (oven-dried for 48 h at 75°C). In the prior experiment the three inocula types were mixed in different ratio's and the effects of the different ratio's were assessed (Wubs et al., 2018). Here we only used those experimental containers where 100% pure inoculum (i.e., from one field) was introduced. The present experiment used 36 containers (3 soil inoculum types x 3 replicate fields x 4 replicates per field) in Phase 1.
After the Phase 1 harvest, the soil from each container was sieved (4 mm mesh) to remove roots. From each original Phase 1 container, four smaller pots were filled with homogenized soil (900 g, 10 x 10 x 11 cm pots). For each Phase 1 container, two Phase 2 pots were randomly selected and planted with one individual of each of the three ruderal plant species. The other two pots per Phase 1 container were planted with one individual each of the three target species (N = 36 Phase 1 containers × 2 Phase 2 treatments in duplicate = 144). Seeds were germinated as before. Pots were placed in the greenhouse in a random spatial design under the same conditions as described before. Any seedlings that died in the first week were replaced. The pots were hand-weeded every week to remove seedlings emerging from the seedbank, and watered three times per week. The plants were allowed to grow for 9 weeks. Subsequently, shoot biomass of each species was cut and dried (48 h, 75°C), before weighing per species.
For the final phase (Phase 3), the soils of Phase 2 were again sieved (4 mm) to remove plant roots and put back in same size pots. Next, one of the two pots conditioned by ruderals, per Phase 1 container, was randomly selected and planted with ruderals and the other was planted with target plant species. The same was done for the two pots conditioned by target species, again per Phase 1 container. This led to a full factorial design of soil inoculum type with Phase 2 and Phase 3 plant species groups (ruderal or target; N = 36 Phase 1 containers × 2 Phase 2 × 2 Phase 3 treatments = 144) and the soils of independent replicates were kept separate throughout the three phases of the experiment. The same methods were used as above and the plants were allowed to grow for 9 weeks, after which shoot biomass of each species was determined.
Differences in seedling mortality among treatments were analyzed using a generalized linear mixed model with a binomial error distribution. The analysis was conducted at Phase 3 pot level and in case any of the seedlings died this was scored as a case of mortality. Fixed effects included Phase 3 and 2 plant groups, soil inoculation and their interactions, while the random effects consisted of the sampling field of the inoculated soil and the original Phase 1 container.
Plant biomass responses were evaluated at two levels, plant group and individual plant species. The plant group level responses were analyzed using a linear mixed model with the same fixed and random effects as for the mortality model above. In addition, we tested a model where we included Phase 2 shoot biomass as a continuous predictor into the above model and we included all possible interactions. Plant-soil feedbacks were tested directly as planned contrast of biomass production on Phase 2-soils conditioned by ruderal and target species, respectively (Adbi and Williams, 2010). For the individual plant species model, PSF was calculated as the log-ratio of plant biomass in own plant group to the other plant group (Brinkman et al., 2010). The individual plant PSFs were analyzed using a linear mixed effects model, with the same random factors as before. As fixed effects plant species, soil inoculation and their interaction were included.
All analyses were conducted in R v3.5.2 (R Core Team, 2019) and model assumptions were checked graphically. Model heteroscedasticity was modeled explicitly using generalized least squares (Pinheiro and Bates, 2000; Zuur et al., 2009). Linear mixed models were analyzed using the nlme v3.1-137 package (Pinheiro et al., 2017) and the GLMM in package MASS v7.3-51.1 (Venables and Ripley, 2002).
Ruderal plant species developed positive feedback on soil inoculated with arable-soil inoculum, while on grassland and heathland inoculated soil they showed neutral PSFs (Figure 1a; Tables S1, S2). Plant species differed in their PSF responses (Figures 1b–g). The positive PSF in arable-inoculated soil was mostly associated with improved performance in M. arvensis (Figure 1d; Table S3). On heathland soil, M. arvensis developed a weakly negative PSF, which was unique among the tested species. As a group, the late-successional target species did not develop significant feedback in response to soil inoculation, although most negative values occurred on heathland-inoculated soil. In fact, F. filliformis showed a trend for positive PSF on arable-inoculated soil (Figure 1f; Table S3).
Overall, ruderal biomass was higher than that of the late-successional target species (Figure 2; Table S1). However, the difference in performance between ruderal and target species was smaller in heathland-inoculated soil than in arable-inoculated soil (Figure 2A; P3 × Inoc interaction; Table S1). There was no relationship between the pot biomass in Phase 2 and Phase 3 (Figure 3), and Phase 2 biomass did not interact with any experimental treatment (Table S4). The interaction between the two plant groups and inoculation (P3 plant × P2 plant x Inoculation) remained significant, indicating that the way the plant groups conditioned the soil (Phase 2) and affected the test plants (PSF, Phase 3) was dependent on the type of soil inoculated, also when accounting for Phase 2 plant biomass. Seedling mortality was overall low and was not affected by the experimental treatments (Figure S1; Table S1).
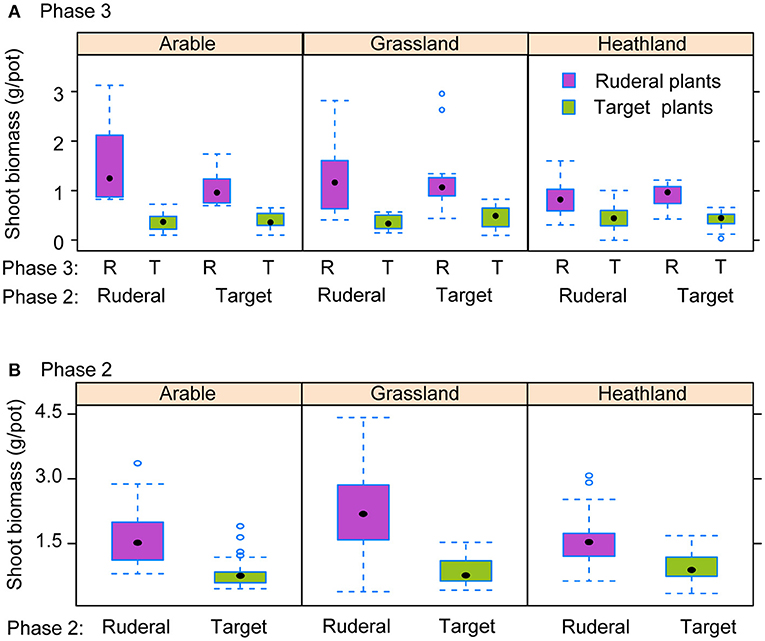
Figure 2. Shoot biomass of ruderal and target plants in Phase 3 (A) and Phase 2 (B), for statistical analyses see Table S1. Phase 2 and Phase 3 indicate the plants that had been grown in the soil, and Inoc indicates the soil type that was inoculated (Arable, Grassland, or Heathland soil). Other conventions follow Figure 1.
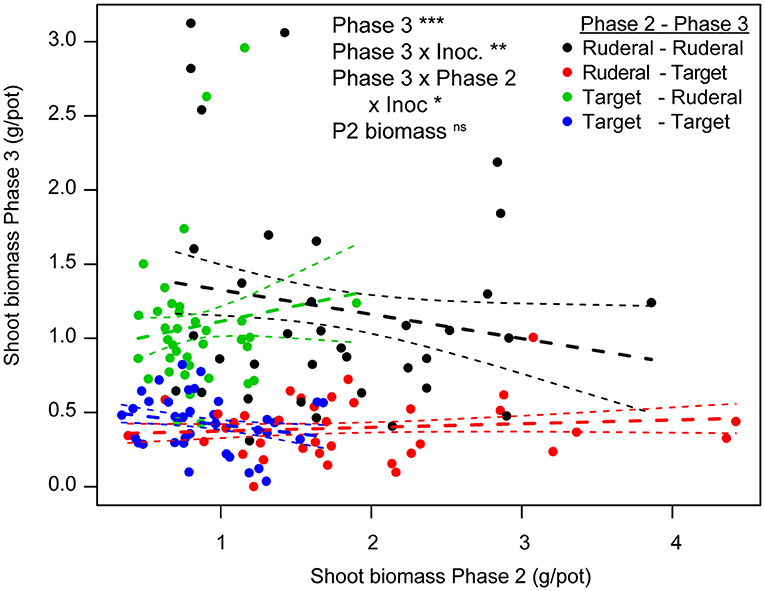
Figure 3. Relationship between shoot biomass per pot in Phase 3 and Phase 2 for each of the four species groups. Significant effects in the mixed model with Phase 2 biomass as a covariate are indicated using stars (*p < 0.05, **p < 0.025, ***p < 0.001), for statistical analysis see Table S4. As a visual aid the fitted relationship of simple linear regressions per feedback group are given as dashed lines (mean ± SE). These analyses show that the effect of Phase 2 biomass was non-significant in all feedback groups.
Our results show that soil inoculation can alter plant-soil feedbacks. However, contrary to our expectation, positive PSFs for target species were not strengthened by inoculation with late-successional soil. Instead, we found that ruderal species had positive PSFs on arable-inoculated soils, in terms of plant biomass. On soil inoculated with late-successional soil, PSFs of ruderal plants became less pronounced and not significantly different from no effect (neutral feedback; Van der Putten et al., 2013). As a group, the late-successional target species developed no significant PSF in this experiment. In the heathland-inoculated soil this led to the most even plant performance between these two groups. Since the positive PSF of ruderal species on arable-inoculated soil was suppressed in pots inoculated with late-successional soil, this suggests that these inoculated soils are robust to colonization by non-target ruderal species.
There was considerable variation in the responses of the individual plant species within the two species groups. For ruderals, the positive PSF on arable-inoculated soil was only clear in the biomass of M. arvensis. The other two ruderal species also had positive mean PSFs on those soils, but these effects were non-significant. Next, we found a trend for a positive PSF in one target species, F. filliformis, on arable-inoculated soils, while we found no other significant responses for late-successional target species. It is well-known that there is large variation in PSF strengths among plant species (Van de Voorde et al., 2011; Cortois et al., 2016) and among plant genotypes (Schweitzer et al., 2008; Evans et al., 2016; Semchenko et al., 2019). Broadly, plant traits associated with resource acquisition, such as specific root length and root diameter, may explain differences in plant-soil feedbacks among ecological groups, directly or via their interaction with soil biota (Lemmermeyer et al., 2015; Cortois et al., 2016; Semchenko et al., 2018). However, the variation in PSFs within plant functional groups and among genotypes of the same species are more likely to be associated with plant traits that regulate the co-evolutionary dynamics of plants and soil-organisms, e.g., via molecular signaling compounds in the roots (e.g., MAMPs and PAMPs; Jones and Dangl, 2006). The differences in these traits may lead to differential susceptibility of plant species and genotypes to the soil biota that are introduced via soil inoculation (Van der Putten et al., 2013; Bardgett et al., 2014; Wubs et al., 2016).
Earlier studies showed that late-successional species elicit positive PSFs on late-successional soils and ruderal species negative PSFs (De Deyn et al., 2003; Kardol et al., 2006; Kulmatiski et al., 2017). In contrast, our results show that for some species the PSF effects may act differently, i.e., via reduced positive PSFs for ruderal species. Interestingly, the selected target species are largely the same in our experiment and for De Deyn et al. (2003) and Kardol et al. (2006). As we used seeds from ex-situ cultivated populations, it could be that the plants established plant-soil interactions that are weaker than those produced by co-evolved plant and soil populations (Felker-Quinn et al., 2011; Evans et al., 2016) leading to different net PSFs. Alternatively, plant-soil feedbacks are known to vary across environmental gradients (De Deyn et al., 2004; Bezemer et al., 2006; Manning et al., 2008) and our common ex-arable soil had substantially higher P-Olsen (78.3 ± 6.71 mg P kg−1 soil) and soil organic matter (5.9 ± 0.2%) content than the soil used in these previous studies. At this point we can only speculate about the causes underlying the differences between these studies. Nevertheless, our results do highlight that even though changes in species abundance in response to soil inoculation (Carbajo et al., 2011; Wubs et al., 2016) may follow the expectations derived from greenhouse PSF experiments (De Deyn et al., 2003; Kardol et al., 2006; Kulmatiski et al., 2017), there may be a different combination of feedbacks in operation (Heinze et al., 2016).
The observed plant-soil feedbacks can in principle have been mediated by both abiotic and biotic changes induced in the soil through plant conditioning (Ehrenfeld et al., 2005; Van der Putten et al., 2013). Nevertheless, we suggest that the observed effects were biotically mediated for two reasons. First, we inoculated a common nutrient rich background soil with a limited amount of inoculum, so that differences in abiotic factors were diluted, thus limiting the scope for nutrient limitation. Secondly, Phase 2 plant biomass showed no correlation with Phase 3 biomass, directly nor in interaction with the experimental treatments, suggesting that abiotic factors such as nutrient limitation did not have an overriding role in our study (e.g., Kardol et al., 2006). The three ecosystem types used to inoculate the soil differ in their soil biotic community composition (Kardol et al., 2005; Van der Wal et al., 2006; Wubs et al., 2016): the grassland and heathland have higher microbial biomass than the arable soil, the grassland has more arbuscular mycorrhizae than the heathland, and the species composition of microbes and nematodes is distinct. Furthermore, data from the same study system showed that soil fungi better explain plant responses than soil abiotic factors (Wubs and Bezemer, 2018a). This is in line with results from other systems showing that the soil biota, and particularly fungi, play a prominent role in determining plant-soil feedback strengths (e.g., Kulmatiski et al., 2017; Mommer et al., 2018; Semchenko et al., 2018). Based on these arguments, we suggest that the effects observed in this experiment are due to plant-induced differences in the different inoculated soil communities, although we cannot rule out plant-induced abiotic effects.
Our study was intended as a proof of principle of how soil inoculations may alter plant-soil feedbacks for use in restoration. There are however two important limitations for direct translation of our results to restoration projects in the field. Our experiment was conducted on a sterilized common soil and it is well-known that establishment success of inoculated soil biota depends on the abundance and diversity of the resident soil community (Van Elsas et al., 2012; Mallon et al., 2015). The effects may therefore be smaller when the resident soil community is left intact by not sterilizing the common soil, as would be the case in restoration projects. However, in previous soil inoculation trials on undisturbed field soils, we did observe that novel soil biota could be successfully introduced and alter local soil and plant community composition (Wubs et al., 2016, 2019) and therefore the local resident community may not be a strong barrier to establishment of late-successional soil biota. Secondly, as is the case with most plant-soil feedback studies (Kulmatiski et al., 2008; Van der Putten et al., 2013), we only documented plant biomass responses over a period of 9 weeks. It is unclear how these short-term responses translate into longer-term fitness differences in the field (Trinder et al., 2013; Heinze et al., 2016; Kulmatiski et al., 2016) as plant-soil feedback strengths may change with plant ontogeny and may take longer to develop in later-successional species (Hawkes et al., 2013; Bezemer et al., 2018; Dudenhöffer et al., 2018). Field observations suggest that the reciprocal interactions among plants and soil biota strengthen over time (Meyer et al., 2016; Wubs et al., 2019) and extrapolation may thus be possible. Nevertheless, field based quantifications of plant-soil feedback strength, e.g., using phytometer plants, are needed to test our conclusions under conditions relevant for restoration.
An important aspect of our study is that both in the conditioning and in the response phases we used plant communities rather than individual plants or monocultures of plants. Most PSF studies have been carried out with soil conditioned by individual plants or monocultures (Kulmatiski et al., 2008). Plant competition can alter plant-soil feedbacks (Casper and Castelli, 2007; Kardol et al., 2007; Jing et al., 2015; Xue et al., 2018), for instance via altered root exudation (Bais et al., 2006), the interaction between resource depletion and defense (Lind et al., 2013), and because of altered food web interactions driven by different species mixtures (Bezemer et al., 2010; Bakker et al., 2013; Kulmatiski et al., 2014). In nature, plants typically do not grow in isolation or in monocultures, and we urge that an important step forward in PSF research is understanding these mixed community feedbacks.
Our results show that soil inoculation alters the nature of individual species' plant-soil feedback (PSF) when these species are grown in competition with other species. The effects were species specific and differed from effects observed in other experiments. This suggests that net inoculation effects on natural vegetation depend importantly on the individual PSFs induced by the soil inoculation, which in turn depend on the soil biotic community and abiotic conditions the plants are growing in. Inoculation with late-successional soil led to suppression of the positive PSF of ruderal species found on arable-inoculated soils. Thus, we conclude that inoculation with late successional soil can reduce the colonization by non-target plant species from elsewhere if our results can be extrapolated to non-sterilized soils and long-term differences in plant fitness.
The datasets generated for this study are available on request from the corresponding author.
EW and TB designed the experiment. TH and PM collected the data. EW analyzed the data and wrote the first draft. All authors contributed to the final version of the paper.
EW was funded by a NWO-Rubicon grant (Netherlands Organization for Scientific Research, contract no. 019.181EN.01). TB was funded by a NWO-Vici grant (865.14.006). This is NIOO-KNAW publication 6848.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2019.00451/full#supplementary-material
Adbi, H., and Williams, L. J. (2010). “Contrast analysis,” in Encyclopedia of Research Design, ed N. Salkind (Thousand Oaks, CA: Sage), 243–251.
Bais, H. P., Weir, T. L., Perry, L. G., Gilroy, S., and Vivanco, J. M. (2006). The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 57, 233–266. doi: 10.1146/annurev.arplant.57.032905.105159
Bakker, M. G., Otto-Hanson, L., Lange, A. J., Bradeen, J. M., and Kinkel, L. L. (2013). Plant monocultures produce more antagonistic soil Streptomyces communities than high-diversity plant communities. Soil Biol. Biochem. 65, 304–312. doi: 10.1016/j.soilbio.2013.06.007
Bardgett, R. D., Mommer, L., and De Vries, F. T. (2014). Going underground: root traits as drivers of ecosystem processes. Trends Ecol. Evol. 29, 692–699. doi: 10.1016/j.tree.2014.10.006
Bennett, J. A., Maherali, H., Reinhart, K. O., Lekberg, Y., Hart, M. M., and Klironomos, J. (2017). Plant-soil feedbacks and mycorrhizal type influence temperate forest population dynamics. Science 355, 181–184. doi: 10.1126/science.aai8212
Bever, J. D. (1994). Feedback between plants and their soil communities in an old field community. Ecology 75, 1965–1977. doi: 10.2307/1941601
Bezemer, T. M., Fountain, M. T., Barea, J. M., Christensen, S., Dekker, S. C., Duyts, H., et al. (2010). Divergent composition but similar function of soil food webs of individual plants: plant species and community effects. Ecology 91, 3027–3036. doi: 10.1890/09-2198.1
Bezemer, T. M., Jing, J., Bakx-Schotman, J. M. T., and Bijleveld, E.-J. (2018). Plant competition alters the temporal dynamics of plant-soil feedbacks. J. Ecol. 106, 2287–2300. doi: 10.1111/1365-2745.12999
Bezemer, T. M., Lawson, C. S., Hedlund, K., Edwards, A. R., Brook, A. J., Igual, J. M., et al. (2006). Plant species and functional group effects on abiotic and microbial soil properties and plant-soil feedback responses in two grasslands. J. Ecol. 94, 893–904. doi: 10.1111/j.1365-2745.2006.01158.x
Brinkman, E. P., Van der Putten, W. H., Bakker, E.-J., and Verhoeven, K. J. F. (2010). Plant–soil feedback: experimental approaches, statistical analyses and ecological interpretations. J. Ecol. 98, 1063–1073. doi: 10.1111/j.1365-2745.2010.01695.x
Buisson, E., Jaunatre, R., Römermann, C., Bulot, A., and Dutoit, T. (2018). Species transfer via topsoil translocation: lessons from two large Mediterranean restoration projects. Restor. Ecol. 26, S179–S188. doi: 10.1111/rec.12682
Carbajo, V., den Braber, B., van der Putten, W. H., and De Deyn, G. B. (2011). Enhancement of late successional plants on ex-arable land by soil inoculations. PloS ONE 6:e21943. doi: 10.1371/journal.pone.0021943
Casper, B. B., and Castelli, J. P. (2007). Evaluating plant–soil feedback together with competition in a serpentine grassland. Ecol. Lett. 10, 394–400. doi: 10.1111/j.1461-0248.2007.01030.x
Cortois, R., Schröder-Georgi, T., Weigelt, A., van der Putten, W. H., and De Deyn, G. B. (2016). Plant–soil feedbacks: role of plant functional group and plant traits. J. Ecol. 104, 1608–1617. doi: 10.1111/1365-2745.12643
De Deyn, G. B., Raaijmakers, C. E., and Van der Putten, W. H. (2004). Plant community development is affected by nutrients and soil biota. J. Ecol. 92, 824–834. doi: 10.1111/j.0022-0477.2004.00924.x
De Deyn, G. B., Raaijmakers, C. E., Zoomer, H. R., Berg, M. P., De Ruiter, P. C., Verhoef, H. A., et al. (2003). Soil invertebrate fauna enhances grassland succession and diversity. Nature 422, 711–713. doi: 10.1038/nature01548
Dudenhöffer, J.-H., Ebeling, A., Klein, A.-M., and Wagg, C. (2018). Beyond biomass: soil feedbacks are transient over plant life stages and alter fitness. J. Ecol. 106, 230–241. doi: 10.1111/1365-2745.12870
Ehrenfeld, J. G., Ravit, B., and Elgersma, K. (2005). Feedback in the plant-soil system. Annu. Rev. Environ. Resour. 30, 75–115. doi: 10.1146/annurev.energy.30.050504.144212
Evans, J. A., Lankau, R. A., Davis, A. S., Raghu, S., and Landis, D. A. (2016). Soil-mediated eco-evolutionary feedbacks in the invasive plant Alliaria petiolata. Funct. Ecol. 30, 1053–1061. doi: 10.1111/1365-2435.12685
Felker-Quinn, E., Bailey, J. K., and Schweitzer, J. A. (2011). Soil biota drive expression of genetic variation and development of population-specific feedbacks in an invasive plant. Ecology 92, 1208–1214. doi: 10.1890/10-1370.1
Hawkes, C. V., Kivlin, S. N., Du, J., and Eviner, V. T. (2013). The temporal development and additivity of plant-soil feedback in perennial grasses. Plant Soil 369, 141–150. doi: 10.1007/s11104-012-1557-0
Heinze, J., and Joshi, J. (2017). Plant–soil feedback effects can be masked by aboveground herbivory under natural field conditions. Oecologia 186, 235–246. doi: 10.1007/s00442-017-3997-y
Heinze, J., Sitte, M., Schindhelm, A., Wright, J., and Joshi, J. (2016). Plant-soil feedbacks: a comparative study on the relative importance of soil feedbacks in the greenhouse vs. the field. Oecologia 181, 559–569. doi: 10.1007/s00442-016-3591-8
Jing, J., Bezemer, T. M., and van der Putten, W. H. (2015). Interspecific competition of early successional plant species in ex-arable fields as influenced by plant–soil feedback. Basic Appl. Ecol. 16, 112–119. doi: 10.1016/j.baae.2015.01.001
Jones, J. D. G., and Dangl, J. L. (2006). The plant immune system. Nature 444, 323–329. doi: 10.1038/nature05286
Kardol, P. (2007). Plant and Soil Community Assembly in Secondary Succession on Ex-Arable Land. Fundamental and Applied Approaches. Ph.D. thesis, Wageningen University, Wageningen.
Kardol, P., Bezemer, T. M., and Van der Putten, W. H. (2006). Temporal variation in plant-soil feedback controls succession. Ecol. Lett. 9, 1080–1088. doi: 10.1111/j.1461-0248.2006.00953.x
Kardol, P., Bezemer, T. M., van der Wal, A., and van der Putten, W. H. (2005). Successional trajectories of soil nematode and plant communities in a chronosequence of ex-arable lands. Biol. Conserv. 126, 317–327. doi: 10.1016/j.biocon.2005.06.005
Kardol, P., Cornips, N. J., van Kempen, M. M. L., Bakx-Schotman, J. M. T., and van der Putten, W. H. (2007). Microbe-mediated plant-soil feedback causes historical contingency effects in plant community assembly. Ecol. Monogr. 77, 147–162. doi: 10.1890/06-0502
Kulmatiski, A., Anderson-Smith, A., Beard, K. H., Doucette-Riise, S., Mazzacavallo, M., Nolan, N. E., et al. (2014). Most soil trophic guilds increase plant growth: a meta-analytical review. Oikos 123, 1409–1419. doi: 10.1111/oik.01767
Kulmatiski, A., Beard, K. H., Grenzer, J., Forero, L., and Heavilin, J. (2016). Using plant-soil feedbacks to predict plant biomass in diverse communities. Ecology 97, 2064–2073. doi: 10.1890/15-2037.1
Kulmatiski, A., Beard, K. H., Norton, J. M., Heavilin, J. E., Forero, L. E., and Grenzer, J. (2017). Live long and prosper: plant–soil feedback, lifespan, and landscape abundance covary. Ecology 98, 3063–3073. doi: 10.1002/ecy.2011
Kulmatiski, A., Beard, K. H., Stevens, J. R., and Cobbold, S. M. (2008). Plant-soil feedbacks: a meta-analytical review. Ecol. Lett. 11, 980–992. doi: 10.1111/j.1461-0248.2008.01209.x
Lekberg, Y., Bever, J. D., Bunn, R. A., Callaway, R. M., Hart, M. M., Kivlin, S. N., et al. (2018). Relative importance of competition and plant–soil feedback, their synergy, context dependency and implications for coexistence. Ecol. Lett. 21, 1268–1281. doi: 10.1111/ele.13093
Lemmermeyer, S., Lörcher, L., Van Kleunen, M., and Dawson, W. (2015). Testing the plant growth-defense hypothesis belowground: do faster-growing herbaceous plant species suffer more negative effects from soil biota than slower-growing ones? Am. Nat. 186, 264–271. doi: 10.1086/682005
Lind, E. M., Borer, E., Seabloom, E., Adler, P., Bakker, J. D., Blumenthal, D. M., et al. (2013). Life-history constraints in grassland plant species: a growth-defence trade-off is the norm. Ecol. Lett. 16, 513–521. doi: 10.1111/ele.12078
Mallon, C. A., Elsas, J. D., and van Salles, J. F. (2015). Microbial invasions: the process, patterns, and mechanisms. Trends Microbiol. 23, 719–729. doi: 10.1016/j.tim.2015.07.013
Manning, P., Morrison, S. A., Bonkowski, M., and Bardgett, R. D. (2008). Nitrogen enrichment modifies plant community structure via changes to plant–soil feedback. Oecologia 157, 661–673. doi: 10.1007/s00442-008-1104-0
Meyer, S. T., Ebeling, A., Eisenhauer, N., Hertzog, L., Hillebrand, H., Milcu, A., et al. (2016). Effects of biodiversity strengthen over time as ecosystem functioning declines at low and increases at high biodiversity. Ecosphere 7:e01619. doi: 10.1002/ecs2.1619
Mommer, L., Cotton, T. E. A., Raaijmakers, J. M., Termorshuizen, A. J., Ruijven, J., van Hendriks, M., et al. (2018). Lost in diversity: the interactions between soil-borne fungi, biodiversity and plant productivity. New Phytol. 218, 542–553. doi: 10.1111/nph.15036
Pinheiro, J. C., and Bates, D. M. (2000). Mixed-Effects Models in S and S-PLUS. New York, NY: Springer Verlag. doi: 10.1007/978-1-4419-0318-1
Pinheiro, J. C., Bates, D. M., DebRoy, S., and Sarkar, D. R Development Core Team (2017). nlme: Linear and Nonlinear Mixed Effects Models. Available online at: https://cran.r-project.org/web/packages/nlme/index.html
Pywell, R. F., Meek, W. R., Webb, N. R., Putwain, P. D., and Bullock, J. M. (2011). Long-term heathland restoration on former grassland: the results of a 17-year experiment. Biol. Conserv. 144, 1602–1609. doi: 10.1016/j.biocon,0.2011.02.010
R Core Team (2019). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Schweitzer, J. A., Bailey, J. K., Fischer, D. G., LeRoy, C. J., Lonsdorf, E. V., Whitham, T. G., et al. (2008). Plant–soil–microorganism interactions: heritable relationship between plant genotype and associated soil microorganisms. Ecology 89, 773–781. doi: 10.1890/07-0337.1
Semchenko, M., Leff, J. W., Lozano, Y. M., Saar, S., Davison, J., Wilkinson, A., et al. (2018). Fungal diversity regulates plant-soil feedbacks in temperate grassland. Sci. Adv. 4:eaau4578. doi: 10.1126/sciadv.aau4578
Semchenko, M., Nettan, S., Sepp, A., Zhang, Q., Abakumova, M., Davison, J., et al. (2019). Soil biota and chemical interactions promote co-existence in co-evolved grassland communities. J. Ecol. 107, 2611–2622. doi: 10.1111/1365-2745.13220
Teste, F. P., Kardol, P., Turner, B. L., Wardle, D. A., Zemunik, G., Renton, M., et al. (2017). Plant-soil feedback and the maintenance of diversity in Mediterranean-climate shrublands. Science 355, 173–176. doi: 10.1126/science.aai8291
Trinder, C. J., Brooker, R. W., and Robinson, D. (2013). Plant ecology's guilty little secret: understanding the dynamics of plant competition. Funct. Ecol. 27, 918–929. doi: 10.1111/1365-2435.12078
Van de Voorde, T. F. J., Van der Putten, W. H., and Bezemer, T. M. (2011). Intra- and interspecific plant–soil interactions, soil legacies and priority effects during old-field succession. J. Ecol. 99, 945–953. doi: 10.1111/j.1365-2745.2011.01815.x
Van der Putten, W. H., Bardgett, R. D., Bever, J. D., Bezemer, T. M., Casper, B. B., Fukami, T., et al. (2013). Plant–soil feedbacks: the past, the present and future challenges. J. Ecol. 101, 265–276. doi: 10.1111/1365-2745.12054
Van der Wal, A., Van Veen, J. A., Smant, W., Boschker, H. T. S., Bloem, J., Kardol, P., et al. (2006). Fungal biomass development in a chronosequence of land abandonment. Soil Biol. Biochem. 38, 51–60. doi: 10.1016/j.soilbio.2005.04.017
Van Elsas, J. D., Chiurazzi, M., Mallon, C. A., Elhottovā, D., Krištufek, V., and Salles, J. F. (2012). Microbial diversity determines the invasion of soil by a bacterial pathogen. Proc. Nat. Acad. Sci. USA 109, 1159–1164. doi: 10.1073/pnas.1109326109
Vécrin, M. P., and Muller, S. (2003). Top-soil translocation as a technique in the re-creation of species-rich meadows. Appl. Veg. Sci. 6, 271–278. doi: 10.1111/j.1654-109X.2003.tb00588.x
Veen, G. F., De Vries, S., Bakker, E. S., Van der Putten, W. H., and Olff, H. (2014). Grazing-induced changes in plant–soil feedback alter plant biomass allocation. Oikos 123, 800–806. doi: 10.1111/j.1600-0706.2013.01077.x
Venables, W. N., and Ripley, B. (2002). Modern Applied Statistics with S. New York, NY: Springer Science+Business Media. doi: 10.1007/978-0-387-21706-2
Wubs, E. R. J., and Bezemer, T. M. (2018a). Plant community evenness responds to spatial plant-soil feedback heterogeneity primarily through the diversity of soil conditioning. Funct. Ecol. 32, 509–521. doi: 10.1111/1365-2435.13017
Wubs, E. R. J., and Bezemer, T. M. (2018b). Temporal carry-over effects in sequential plant–soil feedbacks. Oikos 127, 220–229. doi: 10.1111/oik.04526
Wubs, E. R. J., Melchers, P. D., and Bezemer, T. M. (2018). Potential for synergy in soil inoculation for nature restoration by mixing inocula from different successional stages. Plant Soil 433, 147–156. doi: 10.1007/s11104-018-3825-0
Wubs, E. R. J., Van der Putten, W. H., Bosch, M., and Bezemer, T. M. (2016). Soil inoculation steers restoration of terrestrial ecosystems. Nature Plants 2:16107. doi: 10.1038/nplants.2016.107
Wubs, E. R. J., Van der Putten, W. H., Mortimer, S. R., Korthals, G. W., Duyts, H., Wagenaar, R., et al. (2019). Single introductions of soil biota and plants generate long-term legacies in soil and plant community assembly. Ecol. Lett. 22, 1145–1151. doi: 10.1111/ele.13271
Xue, W., Berendse, F., and Bezemer, T. M. (2018). Spatial heterogeneity in plant–soil feedbacks alters competitive interactions between two grassland plant species. Funct. Ecol. 32, 2085–2094. doi: 10.1111/1365-2435.13124
Keywords: colonization, nature restoration, plant-community assembly, plant-soil feedback, soil inoculation
Citation: Wubs ERJ, van Heusden T, Melchers PD and Bezemer TM (2019) Soil Inoculation Steers Plant-Soil Feedback, Suppressing Ruderal Plant Species. Front. Ecol. Evol. 7:451. doi: 10.3389/fevo.2019.00451
Received: 21 May 2019; Accepted: 11 November 2019;
Published: 26 November 2019.
Edited by:
Mauro Santos, Autonomous University of Barcelona, SpainReviewed by:
Pilar Andrés, Ecological and Forestry Applications Research Center (CREAF), SpainCopyright © 2019 Wubs, van Heusden, Melchers and Bezemer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: E. R. Jasper Wubs, amFzcGVyLnd1YnNAdXN5cy5ldGh6LmNo
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.