 Jack R. Arterburn
Jack R. Arterburn Dirac Twidwell
Dirac Twidwell Carissa L. Wonkka
Carissa L. Wonkka Walter H. Schacht1
Walter H. Schacht1
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 02 October 2019
Sec. Conservation and Restoration Ecology
Volume 7 - 2019 | https://doi.org/10.3389/fevo.2019.00365
Heterogeneity has emerged as a fundamental principle for grassland management and the importance of environmental heterogeneity for biological diversity has raised questions about the appropriateness of grassland practices that seek to promote uniform grassland structure and composition. Principles of uniformity in grassland management reflect a utilitarian target of “managing for the middle” by minimizing both overgrazing and underutilization while avoiding or preventing fire and other disturbances that consume aboveground biomass. We evaluated pioneering efforts to restore fire-grazer interactions via patch burning in an effort to increase spatial heterogeneity at the patch scale in the Nebraska Sandhills, a sandy soil, mixed-grass ecoregion with a long history of “management for the middle.” The application of patch burning did not increase heterogeneity of vegetation structure or composition at the patch-level in the Nebraska Sandhills. Instead, grassland structure exhibited greater temporal variability between growing and dormant seasons and smaller scale variability within fire patches. The low stocking rate and the rapid regrowth of live herbaceous vegetation following fire likely constrained the degree of spatial heterogeneity observed in our study and highlights the challenges of balancing forage supply and demand in semi-arid grasslands.
The rangeland discipline has undergone a recent ideological shift to embrace heterogeneity as a fundamental principle for grassland management (Fuhlendorf et al., 2017). Heterogeneity is the spatial and temporal variability in vegetation structure and composition essential for ecosystem function (Wu and Loucks, 1995; Fuhlendorf and Engle, 2004). Environmental heterogeneity begets greater diversity across multiple trophic levels (MacArthur and MacArthur, 1961; Tews et al., 2004), causing academics and natural resource professionals to question the appropriateness of grassland practices that seek to promote uniform grassland structure and composition (Fuhlendorf et al., 2012). In grasslands, soil and topographic features contribute to inherent diversity of vegetation due to resource availability and plant species competition (Patten and Ellis, 1995; Fuhlendorf and Engle, 2001; Winter et al., 2011). Grassland species evolved within a dynamic, disturbance-driven spatially, and temporally heterogeneous landscape with specialist niche space for flora and fauna (Biondini et al., 1989; Fuhlendorf and Engle, 2004; Tews et al., 2004; McGranahan et al., 2012). Inherent and disturbance-driven heterogeneity have long been minimized in utilitarian grassland management systems that have sought to “manage for the middle” and promote uniform grazing distribution (Fuhlendorf et al., 2012; Twidwell et al., 2013).
Management for uniformity seeks to decrease spatial heterogeneity by distributing moderate grazing pressure more evenly over a grazing unit to minimize the occurrence of overgrazed patches as well as underutilized patches (Fuhlendorf et al., 2009). The rangeland discipline countered long-term overgrazing by adopting utilitarian ideals that promoted uniform grazing at a moderate grazing pressure using fencing and water development to avoid too little or too heavy grazing utilization and disturbance (Fuhlendorf et al., 2006, 2009, 2012; Dunn et al., 2010; Holechek et al., 2011). Therefore, heterogeneity of grassland vegetation managed for spatial uniformity is limited to variability associated with phenological changes over time. Managing for spatial heterogeneity in grasslands has been proposed as an alternative approach to traditional grassland management that is better suited to managing for multiple ecosystem services (Fuhlendorf et al., 2012).
Patch burning has been introduced as a way to transition away from homogeneous grassland management in an effort to increase biodiversity and ecosystem service provisioning while sustaining production value (Fuhlendorf and Engle, 2001, 2004). Patch burning attempts to increase heterogeneity in grasslands managed for uniformity over the last century by restoring fire-grazing interaction (Fuhlendorf and Engle, 2001; Fuhlendorf et al., 2009). By applying spatially and temporally discrete fires and allowing grazers unrestricted access to the patchwork of burned and unburned areas, grazer selection for burned areas creates a heterogeneous landscape of vegetation structure, composition, and biomass in various stages of recovery (Fuhlendorf and Engle, 2001; Anderson et al., 2006; Fuhlendorf et al., 2006). Application of patch burning in tallgrass, mixed-grass, and shortgrass prairies has increased the diversity of small (Fuhlendorf et al., 2010) and large mammal communities (Meek et al., 2008; Augustine and Derner, 2015), increased desirable invertebrate species (Debinski et al., 2011), and decreased insects harmful to livestock (Scasta et al., 2012; Polito et al., 2013) all while improving cattle production (Fuhlendorf and Engle, 2004; Vermeire et al., 2004; Allred et al., 2011b, 2014; Limb et al., 2011), reducing invasive species (Cummings et al., 2007), and contributing to grassland restoration (McGranahan et al., 2014). Although studies have occurred across precipitation, productivity, and topoedaphic gradients (Winter et al., 2011; McGranahan et al., 2012; Augustine and Derner, 2014), few studies have occurred in ecosystems consisting of predominantly sandy soil with lower nutrient, organic matter, and water holding capacities.
The importance of heterogeneity in managing for multiple ecosystem services has led landowner-led organizations and state, federal, and private conservation organizations from across the Nebraska Sandhills to seek management practices to increase plant community variability. Restoration of the fire-grazer interaction in the Nebraska Sandhills has been controversial because the system has a long history of “managing for the middle” (Biondini et al., 1999; Steuter and Hidinger, 1999). Although fire in the Sandhills has become less frequent since EuroAmerican settlement (Steinauer and Bragg, 1987), fire continues to drive grazing site selection irrespective of topoedaphic features including bison selection for recently burned areas (Biondini et al., 1999). However, concerns over disturbances that increase bare ground have limited the application of fire in the Sandhills (Stubbendieck, 1998). Management of the Sandhills reflects the ideology of the rangeland discipline resulting in a structurally homogeneous grassland landscape with the greatest differences in structure and composition occurring as a result of topographic position (i.e., dune vs. interdune; Steuter et al., 1995; Biondini et al., 1999; Schacht et al., 2000), pastoral management history (e.g., differences in timing and intensity of grazing), or the occasional blowout (Stubbendieck et al., 1989).
Our objective was to determine how to restore fire-grazer interactions and increase landscape-level heterogeneity in the Sandhills prairie. The Nebraska Sandhills is an expansive, semiarid ecoregion where land managers have a long history of “managing for the middle” and preventing disturbances (e.g., fire) that expose bare ground. The primary expectation was that a structurally and compositionally heterogeneous landscape would emerge following the implementation of patch burning, consistent with the re-introduction of fire-grazer interactions in other native prairie ecosystems. An alternative expectation, given the long-term legacy of management and the homogeneity of vegetation cover in the Sandhills, is that grassland structure and composition will continue to be spatially homogeneous and more dynamic in time; in this latter case, temporal (interannual) sources of variation (e.g., plant senescence) would exhibit greater control over landscape structure than environmental gradients over space (e.g., topographic position, pastoral management history, and time since focal fire and grazing). We report on the degree of landscape-level heterogeneity created following the initial attempts to restore fire-grazer interactions in the Nebraska Sandhills, and lessons learned that can be implemented in other regions seeking to escape traditions of homogeneity-based management.
This study was conducted in the Nebraska Sandhills ecoregion at The Nature Conservancy's Niobrara Valley Preserve located in north-central Nebraska. The Sandhills are a contiguous, 5.6 million ha grassland ecoregion that supports 500,000 cattle and Nebraska's $7.2 billion-a-year cattle industry. The study site is classified as a cold semi-arid climate receiving 591 mm of precipitation annually, with 77% occurring during the April through September growing season (HPRCC, 2015). The mean annual temperature for the site is 10°C, ranging from −3°C in January to 24°C in July, with an average frost free period of 152 days (HPRCC, 2015). Soils at the site are characteristic of Sandhills soils, comprised of Valentine fine sands (mixed, mesic Typic Ustipsamments) featuring a high risk of wind erosion, low water-holding capacity and rapid infiltration of precipitation, with little or no runoff (Official Series Description, 2007). Sands ecological site is the most common, but sandy, choppy sands, and shallow to gravel sites also occur (Pfeiffer and Steuter, 1994; Steuter et al., 1995; WSS, 2015). Vegetation stabilized sand dunes of 0–80% slope are comprised of north and south-facing, gently-rolling dunes (Steinauer and Bragg, 1987; Schacht et al., 2000; Official Series Description, 2007).
Vegetation at the study site is representative of the Sandhills and characterized as Sandhills mixed-grass prairie (Kaul, 1998; Schacht et al., 2000). Differences in topoedaphic features of unburned Sandhills grassland drives plant community composition because of differences in microclimate conditions between dune slopes and interdunes and grazing selection for interdune areas before the reduction in forage necessitates movement to upland dunes (Bragg, 1978; Steuter et al., 1995; Schacht et al., 2000). Warm-season graminoids dominate the Sandhills plant community on both south- and north-facing slopes; however, their deep root systems and low transpiration rates make them better adapted to the drier, south-facing slopes (Steuter, 1987; Schacht et al., 2000). Dominant warm-season graminoids include Andropogon hallii (sand bluestem), Schizachyrium scoparium (little bluestem), and Calamovilfa longifolia (prairie sandreed). Cool-season graminoids are found on north-facing slopes and dominate interdune areas that feature lower light intensities, lower temperatures, and greater water availability because of greater organic matter content (Doliner and Jolliffe, 1979; Schacht et al., 2000). Interdunes are well-suited for cool-season graminoids because the relatively high soil moisture availability at the soil surface early in the growing season is a good match for lower water use efficiency and shallow root systems of the cool-season graminoids (Schacht et al., 2000). Common cool-season graminoids include Carex spp. (sedge), Panicum oligosanths subsp. scribnerianum (Scribner panicum), Hesperostipa spp. (needlegrasses), and Koeleria macrantha (prairie junegrass). Ambrosia psilostachya (western ragweed) and Helianthus pauciflorus (stiff sunflower) are common perennial forbs. Annual forbs and shrubs occur frequently (Schacht et al., 2000).
The Niobrara Valley Preserve includes two independent units that have been grazed year-round by bison for over 20 years (Figure 1). The 5,217 ha west bison unit and the 3,935 ha east bison unit, established in 1988 and 1986, respectively, range from 2.5 to 24 km apart. The original 975 ha east bison unit has expanded several times with the most recent addition of 953 ha in fall 2012. Stocking rate in each unit has remained relatively low at 1 AUM ha−1 for the entire year to follow conservation-based management (Pfeiffer and Steuter, 1994; Biondini et al., 1999). In 2012, during a prolonged drought, herds were reduced to 0.69 AUM ha−1 in the west bison unit and 0.49 AUM ha−1 in the east bison unit (stocking rates based on bison numbers at fall round-ups). By 2015, stocking rates had increased to 1.09 AUM ha−1 in the west bison unit and 1.12 AUM ha−1 in the east bison unit. In an attempt to increase biological diversity, growing and dormant season (about 80% growing season and 20% dormant season) prescribed fire has been applied inconsistently across both units since 1991 at a fire return interval of 7.5 years or greater (Pfeiffer and Steuter, 1994; Biondini et al., 1999). Fine fuel loads range around 1,200 kg/ha and general burning conditions include relative humidity between 25 and 35%, winds 9–25 km h−1, and air temperature between 18 and 26°C (Pfeiffer and Steuter, 1994). Prior to being established, both units were grazed annually by cattle (Steinauer and Bragg, 1987).
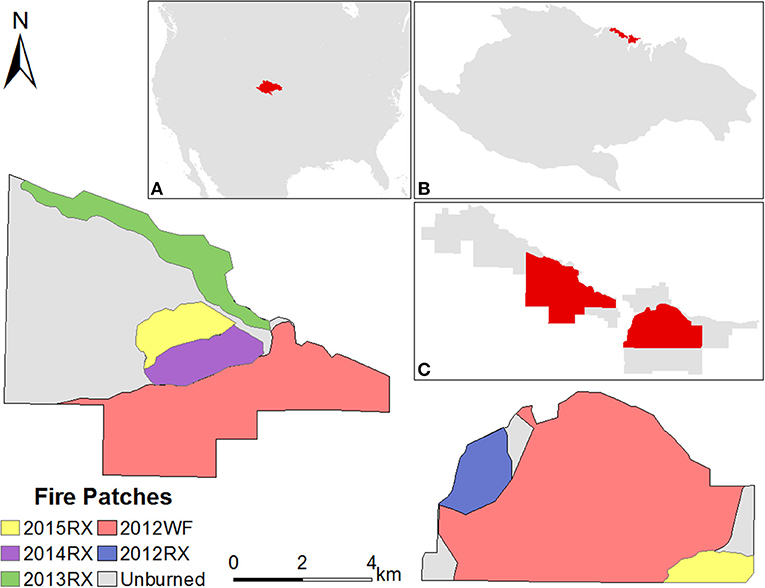
Figure 1. Fire was applied in patches on two continuous Nebraska Sandhills grassland landscapes characterized as homogeneous in vegetation structure and composition after long-term “management for the middle.” RX represents patches burned using prescribed fire in March (2015RX, 2013RX, and 2012RX) and September (2014RX) of each respective year. WF represents the Fairfield Creek Wildfire that burned in July 2012. The unburned patch did not burn in the time period between 2012 and the end of the study. Bison have unrestricted access to all fire patches within each unit. Insets illustrate the location of (A) the Nebraska Sandhills in central North America, (B) the Niobrara Valley Preserve in the Nebraska Sandhills, and (C) the contiguous bison units at the Niobrara Valley Preserve.
Fires occurred during differing seasons and conditions over a 5 year period. In March, the vegetation was primarily dormant and fires were conducted during mild conditions of temperatures below 26°C, wind speed below 32 km h−1, and relative humidity above 20%. In September, live vegetation was transitioning from growing to dormant season. The September fire was conducted during very mild conditions of temperatures below 15°C, wind speed below 16 km h−1, and relative humidity above 60%. The Fairfield Creek Wildfire (part of the Region 24 Complex) burned in late July 2012 when precipitation levels were 75% below average for the 60 days prior to the wildfire (HPRCC, 2015). At the time of the fire, temperatures reached 43°C, relative humidity was as low as 13%, and wind gusts were recorded up to 50 km h−1 (Weather Underground 2012).
We tracked vegetation structure and plant community composition in discrete fire patches that varied in space and time (fire patch size range relative to the landscape unit: 5–87%; Figure 1). Fire patches include patches where prescribed fires were conducted in March 2012, 2013, and 2015, a prescribed fire conducted in September 2014, and a July 2012 wildfire. Bison were allowed immediate access to burned and unburned areas following all fires.
Within each patch, two, 300-m transects were sampled using a 0.1-m2 quadrat placed at 3-m intervals. Transect locations were marked with a steel post 10 m directly south and north of each end of the transect to allow repeated sampling. Topographic position was recorded for each quadrat (north-facing slope, south-facing slope, or interdune). Vegetation structure was sampled along each transect over six periods spanning three seasons of vegetation growth. Sampling periods included early growing season in June 2014 and 2015, late growing season in August 2014 and 2015, and dormant season in December 2014 and January 2016. Estimates of percentage foliar cover, litter cover, and bare ground were recorded using cover classes modified from Daubenmire (1959). Measurements of vegetation height and litter depth occurred at three points within each quadrat. Cover and height were recorded for live herbaceous vegetation (live graminoids and live forbs), live graminoids, live forbs, herbaceous standing dead vegetation (characterized as previous years' growth attached at the plant base), and shrub vegetation. Visual obstruction reading (VOR) was measured at a 4-m distance from the center of the quadrat, at a height of 1 m, in the four cardinal directions by recording complete coverage of 5-cm segments (Robel et al., 1970). Plant community composition was sampled along one transect during August sampling periods by estimating percentage foliar cover of each species using classes modified from (Daubenmire, 1959).
We used an unconstrained, distance-based ordination technique known as nonmetric multidimensional scaling (NMDS) to visually describe heterogeneity based on differences in vegetation structure and plant community. NMDS plots samples in multidimensional ordination space based on either structural metrics or plant community composition present in the sample with the distance between samples corresponding to the similarity in structure or composition among samples. This is an iterative approach to representing multidimensional dissimilarity in low dimensional space to assess relative differences (Legendre and Legendre, 1998): the closer the samples are in ordination space, the more similar they are in composition. Given the limitations of natural experiments like ours, which necessarily lack independent application of treatments, for making broad inference, we conducted ordination rather than rigorous statistical analysis because of the potential for spatial autocorrelation to confound the analysis. Samples were averaged for each transect resulting in two transects per burned and not burned sites (n = 2). Vegetation structural metrics and plant community composition were analyzed separately and species that occurred in <5% of samples for both years were omitted from analysis to focus on relationships between dominant species (Biondini et al., 1989; McCune et al., 2002; Laughlin and Fule, 2008; Taylor et al., 2011). Data were plotted in ordination space over time according to sampling period. Convex hulls represented by colored envelopes were added to connect the vertices of sampling units occurring on distinct spatial landscape features (e.g., bison units, fire patches, and topographic positions). This allows us to explore how structural metrics and species composition cluster in space depending on landscape features (e.g., pastoral management history, time since focal fire and grazing, and topographic position). Vegetation structure in each fire patch was ordinated and the ordination scores averaged by sampling period (standard error around the mean for each NMDS axis was calculated for each sampling period and fire patch) to describe temporal variability. We used Permutational Multivariate Analysis of Variance (PERMANOVA) to test for differences in the means of multivariate data among landscape features, sampling periods, and fire patch. We then tested for differences in the variability or dispersion of the data in ordination space among landscape features, sampling periods, and fire patch using analysis of multivariate homogeneity of group dispersions. Multivariate homogeneity of group dispersions compares the average distance to group centroids among the different groups.
We used a linear regression (p < 0.05) and boxplots of structural metrics depicting the mean value and standard error of each metric averaged over transects to visualize changes over time since fire related to phenology. To visualize phenological changes of herbaceous vegetation structural metrics over time, we plotted respective means of live herbaceous, herbaceous standing dead, and live and dead vegetation. All analyses were performed using the R statistical computing software (R Core Team, 2014). NMDS was performed using Bray-Curtis distances in the vegan package (Oksanen et al., 2016). We used the adonis and betadisper functions from the vegan package (Oksanen et al., 2018) to conduct PERMANOVA and multivariate tests for homogeneity of group dispersion.
Patch burning did not create a landscape that was structurally and compositionally more dynamic in space. No landscape features assessed in this study (bison unit, fire patch, or topographic position) contributed to spatial contrasts in structure among burned patches (Figure 2). PERMANOVA results show vegetation structural metrics did not differ on average among bison units, fire patches, or topographic positions; however within-group variability did differ among fire patches (betadisper p < 0.05; Supplementary Table 1). While overall structure did not differ between burned and unburned patches (Supplementary Table 1), variability within unburned areas was lower than variability within all other fire patches (Supplementary Table 2). This difference, while statistically significant, was minimal (Figure 2). Grassland community composition was not temporally dynamic over the 2 years of this study or associated with landscape features (Figure 3). Vegetation structure and composition were each similar across patches of different histories of time since fire (Figures 2, 3, respectively). PERMANOVA results show that plant communities did not differ among units or topographical positions (Supplementary Table 3). Plant community composition did differ significantly, although not substantially among fire patches (Supplementary Table 3, Figure 3), with burned patches being slightly different from unburned. However, this compositional shift did not lead to differences in structure. The lack of spatial patch contrasts is attributed to rapid growth of vegetation following fire. Most vegetation structural metrics rebounded quickly to levels similar to unburned areas (Figure 4). Following fire, structural metrics related to live herbaceous and live graminoids were similar to unburned levels after 300 days. Live forbs and live shrubs remained similar to unburned levels irrespective of days since fire indicating grazing pressure did not impact forb or shrub growth. Herbaceous standing dead and litter cover significantly increased following fire, returning to levels similar to unburned areas after 450 days. Bare ground was greatest immediately following fire before significantly decreasing with time, returning to levels similar to those in unburned areas after 300 days. Litter depth and visual obstruction readings were variable, but did not significantly change after fire.
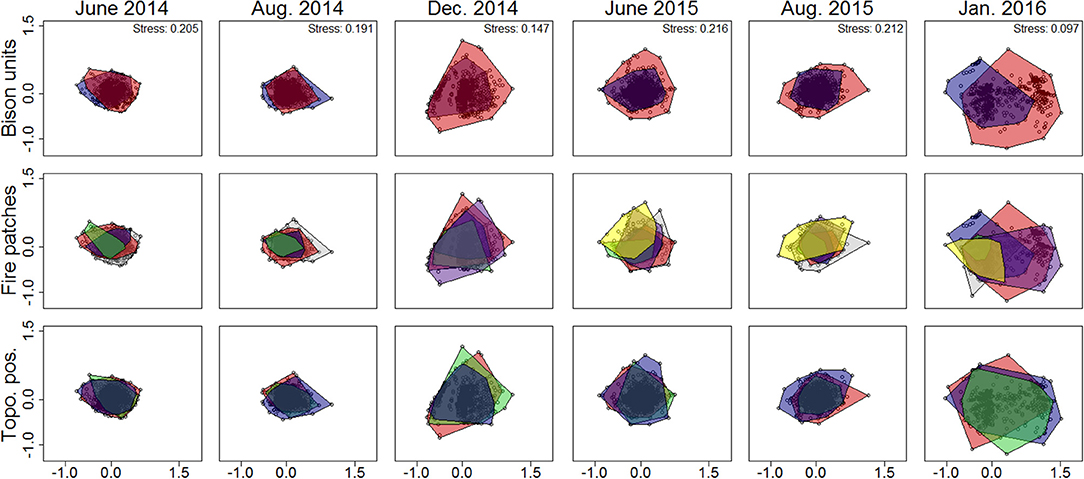
Figure 2. Nonmetric multidimensional scaling (NMDS) ordination of vegetation structural metrics collected over six sampling periods (2014–2016) in a Nebraska Sandhills grassland managed with patch burning and bison grazing. Ordinations were duplicated for each sampling period with color envelopes corresponding to functional patch types. Bison unit color envelopes are red for the west and blue for the east. Fire patch color envelopes are red for 2012WF, blue for 2012RX, green for 2013RX, purple for 2014RX, yellow for 2015RX, and gray for the area unburned between 2012 and the end of the study. Topographic position color envelopes are red for north dunes, blue for south dunes, and green for interdunes. Stress values are included to show how well the ordination summarizes the observed distances among samples (stress >0.3 is a poor representation, 0.2–0.29 is acceptable, 0.1–0.19 is great, and <0.1 is excellent). The distance between points corresponds to the difference between samples with overlapping samples indicating no difference between samples.
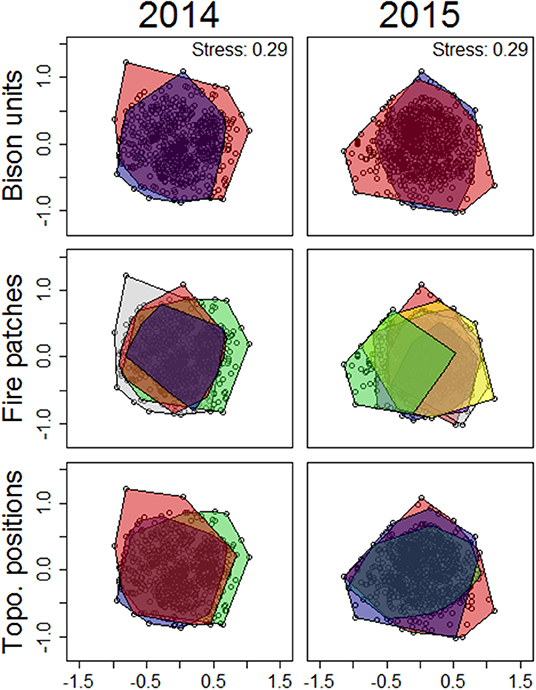
Figure 3. Nonmetric multidimensional scaling (NMDS) ordination of Nebraska Sandhills plant community composition collected in discrete fire patches in August 2014 and 2015. Ordinations were duplicated for each sampling period with color envelopes corresponding to functional patch types. Color envelopes for bison units are red for the west bison unit and blue for the east bison unit. Color envelopes for fire patches are red for the 2012 wildfire, blue for the 2012 prescribed fire, green for the 2013 prescribed fire, purple for the 2014 prescribed fire, yellow for the 2015 prescribed fire, and gray for the area unburned between 2012 and the end of the study. Color envelopes for topographic positions are red for north dunes, blue for south dunes, and green for interdunes. Stress values are included to show how well the ordination summarizes the observed distances among samples (stress >0.3 is a poor representation, 0.2–0.29 is acceptable, 0.1–0.19 is great, and <0.1 is excellent). The distance between points corresponds to the difference between samples with overlapping samples indicating no difference between samples.
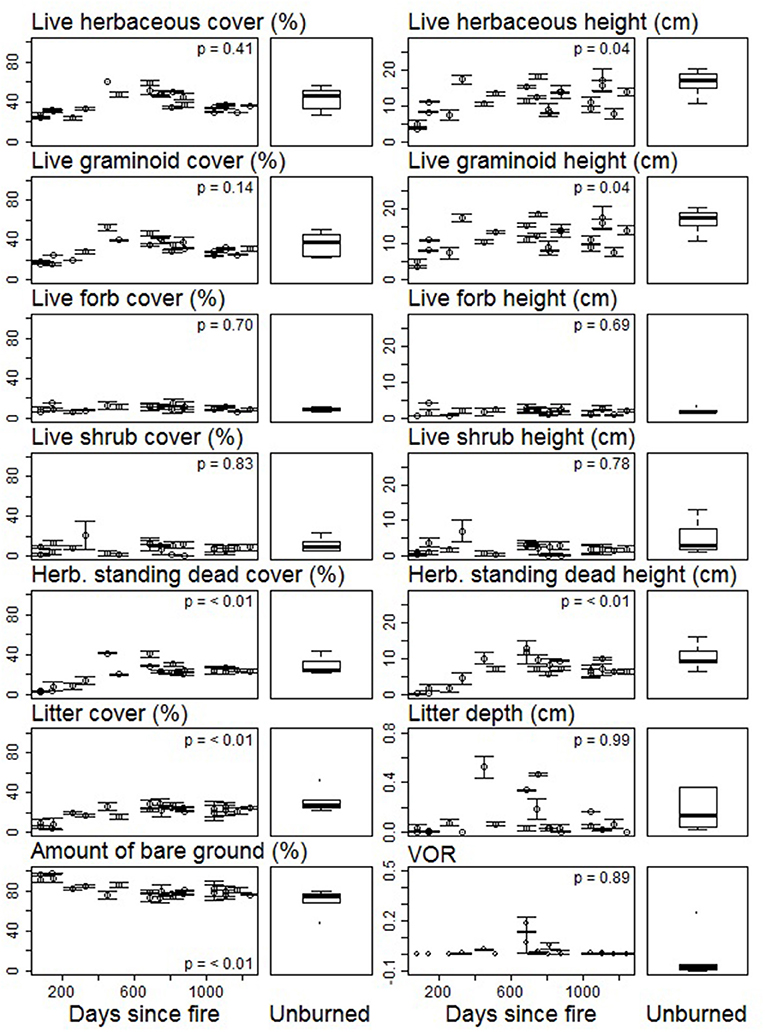
Figure 4. Mean (n = 2) and standard error of structural metrics [estimates of percentage foliar cover, litter cover, and bare ground; vegetation height and litter depth; and visual obstruction reading (VOR)] related to growing season vegetation following fire and grazing in the Nebraska Sandhills grassland.
The structure and composition of this Sandhills grassland exhibited greater temporal variability than spatial variability, which is indicative of a homogeneous grassland at the landscape-scale. Distribution of vegetation structural metrics differed between growing and dormant season sampling periods. Metrics of live vegetation (live herbaceous vegetation, live grasses, live forbs, and live shrubs) were greatest during growing season sampling periods and lowest during dormant sampling periods (Figure 5). Conversely, structural metrics related to herbaceous standing dead vegetation during dormant sampling periods were similar to or greater than those observed during growing season sampling periods (Figure 6). NMDS ordinations all converged with acceptable stress values of <0.3 (which is a measure of how well the low-dimensional ordination reflects the observed multidimensional distances between samples; Legendre and Legendre, 1998). Ordination showed structural metrics during growing season sampling periods were similar within each fire patch, but differences in structural metrics between growing and dormant season occurred during both years of the study, including some separation between years for the dormant season sampling periods (Figure 7). This pattern of homogeneity occurred even after accounting for nested hierarchies among landscape features (e.g., fire patches located within individual bison units; Supplementary Figures 1, 2).
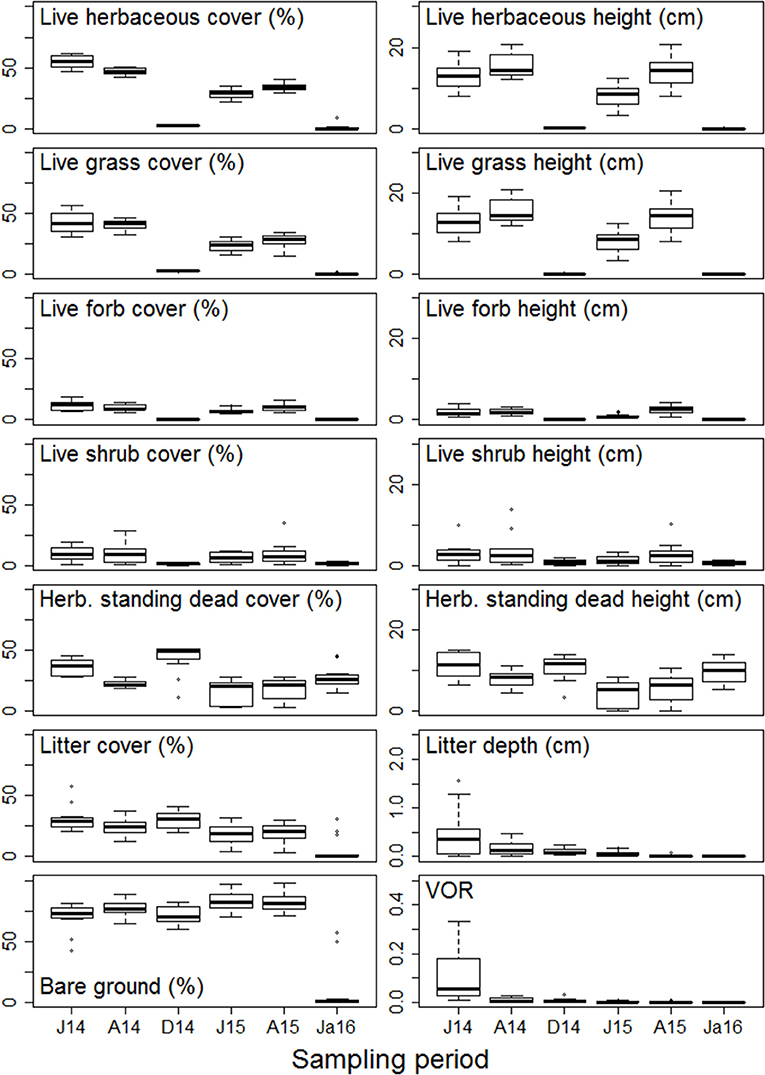
Figure 5. Mean and standard error of vegetation structural metrics [estimates of percentage foliar cover, litter cover, and bare ground; vegetation height and litter depth; and visual obstruction reading (VOR)] collected over six sampling periods in a Nebraska Sandhills grassland managed with patch burning. Sampling periods are: J14, June 2014; A14, August 2014; D14, December 2014; J15, June 2015; A15, August 2015; Ja16, January 2016.
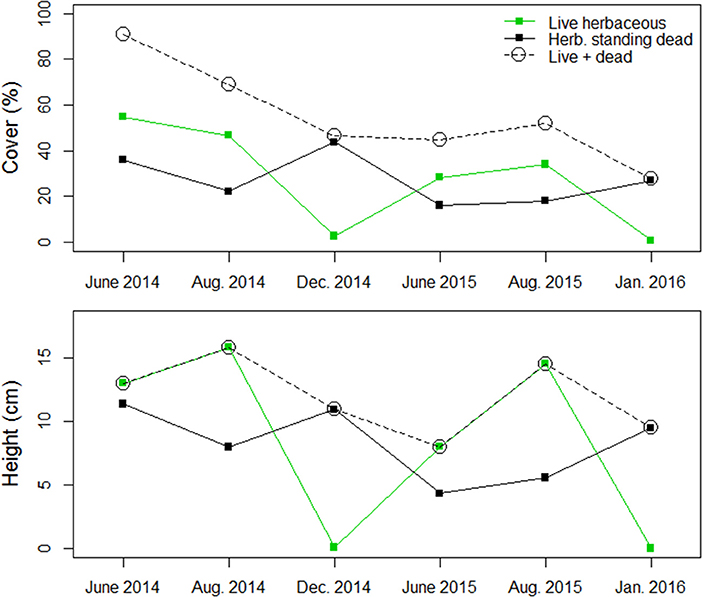
Figure 6. Structural metrics related to live herbaceous and herbaceous standing dead vegetation in the transition from growing to dormant season in a Nebraska Sandhills grassland managed with patch burning.
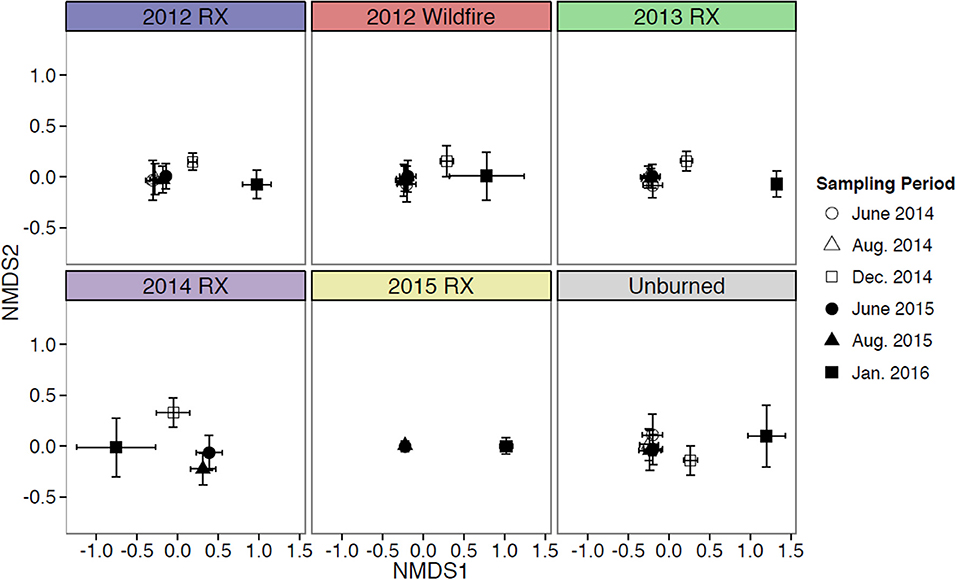
Figure 7. Nonmetric multidimensional scaling (NMDS) ordination of vegetation structural metrics from discrete fire patches in a Nebraska Sandhills grassland managed with patch burning. Shapes within each plot correspond to the six data sampling periods in the early growing, late growing and dormant seasons that were replicated.
Patch burning has been introduced as a method for increasing spatial heterogeneity and biodiversity in grasslands by applying fire in patches on a contiguously grazed landscape with heterogeneity emerging between patches (Fuhlendorf and Engle, 2001). The basis of patch burning is the development of landscape scale heterogeneity with variability in vegetation structure emerging among patches within the landscape, as opposed to finer scales, to provide for multiple ecosystem services (Fuhlendorf and Engle, 2001; Fuhlendorf et al., 2009). We applied patch burning in a sandy soil, mixed-grass landscape that has been managed for uniformity to assess the ability for patch burning to create spatial heterogeneity. The initial applications of patch burning at the Niobrara Valley Preserve resulted in spatially uniform vegetation structure and plant community composition at the patch scale across landscape features. This result is likely owed to the rapid recovery of vegetation in the subsequent growing season following fire and the low stocking rate of bison at the site, constraining the potential to create lasting variability among patches with different fire histories. Instead, heterogeneity emerged temporally according to growing and dormant season conditions where senescence transitioned live herbaceous vegetation to herbaceous standing dead vegetation. Therefore, temporal heterogeneity manifested as an artifact of how vegetation structural metrics were classified in this study.
For patch burning to increase spatial heterogeneity, Allred et al. (2011b) and McGranahan et al. (2012) hypothesize three major conditions are required: (a) discrete fires must be applied in patches on the landscape, (b) fire must be the primary driver of grazing selection, and (c) forage demand must correspond at a moderate grazing pressure to forage supply. Although grazers select for areas recently burned and avoid areas with greater time since fire (Fuhlendorf and Engle, 2004; Allred et al., 2011b), fire can only drive grazing site selection when applied in patches on a contiguous landscape, allowing grazers unrestricted access to select between recently burned areas and unburned areas (Allred et al., 2011b). The ability for fire to reduce grazing selectivity at the plant level is lost when stocking rate does not adequately match the forage production of the burned area. Overstocking results in more uniform grazing as animals graze unburned areas because of depleted forage supply in burned areas. Understocking inhibits patch burning's ability to create heterogeneity by enabling vegetation growth in burned areas to exceed grazing demand allowing grazers to selectively graze within the burned area (Fuhlendorf and Engle, 2004; Cummings et al., 2007; Fuhlendorf et al., 2009).
In our study, the coupling of a low stocking rate and the subsequent growth of live herbaceous vegetation following fire constrained the ability of patch burning to create spatial heterogeneity by allowing vegetation growth to exceed grazing demand. The average stocking rate in our study (0.85 AUM ha−1) is relatively low compared to a moderate Sandhills stocking rate (1.8 AUM ha−1). Patch burning across the Great Plains has successfully created spatial heterogeneity using moderate to moderate-light stocking rates relative to the region (McGranahan et al., 2012). In our study, fire failed to modify grazing behavior or create spatial variability because of the imbalance between vegetation growth and grazing demand, allowing grazers to selectively graze within burned patches, failing to reduce plant-level grazing selectivity and only increasing within patch variability. When grazing demand corresponds to forage supply, focal grazing in recently burned areas creates a patch of shorter vegetation relative to areas with a greater time since fire, and, thus, spatial heterogeneity.
The balance between forage supply and demand is complicated in semi-arid climates. Annual precipitation variability coincides with variable annual forage production complicating the ability for managers to maintain a constant annual grazing pressure. Lower precipitation fosters slower vegetation growth following fire which decreases grazer attraction to recently burned areas while wet periods increase grazer attraction (McGranahan et al., 2013; Augustine and Derner, 2014). Distance to water in semi-arid grasslands constrains grazing distribution because animals focally graze near water sources to conserve energy (Augustine and Derner, 2014). During our study, precipitation variability, including one of the driest years on record, created a dynamic system of variable forage production and relatively constant annual stocking rate on a patch burned landscape. In other prairie ecosystems, greater landscape-level heterogeneity has occurred in years where forage supply equals forage demand, because grazers have the ability to create grazing lawns of short vegetation structure in recently burned areas compared to areas not recently burned (Fuhlendorf and Smeins, 1999; Fuhlendorf and Engle, 2001). Additionally, drought impacts the emergence of heterogeneity in a patch burning landscape at difference scales (Bielski et al., 2018).
Long-term management (30 years) of the study site bison pastures at a low stocking rate (i.e., low grazing pressure) resulted in spatially homogeneous grassland structure and composition, irrespective of landscape factors (e.g., pastoral management history, time since focal fire and grazing, and topographic position). Landscape management histories, season of fire, and topoedaphic features are known to contribute to Sandhills grassland variability when grazed by cattle. Barnes et al. (1984), Biondini et al. (1989), and Schacht et al. (2000) found that shortgrasses (e.g., Bouteloua gracilis and Poa pratensis) and sedges were more common in the interdunes than on the slopes and dune tops of Sandhills uplands grazed at moderate to heavy stocking rates by cattle. Cattle appear to select the low-lying interdunes and focus grazing on the interdunes resulting in dominance by shortgrasses and sedges (Barnes et al., 1984; Biondini et al., 1989; Schacht et al., 2000). We found no differences on Sandhills grassland grazed by bison in vegetation structure or plant community composition across topoedaphic features. Instead, plant community composition variability emerged among fire patches, but did not translate to among-patch heterogeneity of vegetation structural metrics. Even at low grazing pressure, bison have exhibited more even grazing distribution compared to cattle, which focus grazing near water sources (Allred et al., 2011a) and in favored topoedaphic positions (i.e., interdunes, in the Sandhills; Stubbendieck and Reece, 1992; Schacht et al., 2000). Replacement of heavy cattle grazing with light bison grazing over the previous 30 years at the study site apparently shifted the plant community in the interdune areas from predominantly cool-season, grazing-tolerant species to warm-season, tall grasses similar to those found on the slopes; thereby, decreasing variability of plant community composition associated with topographic position that was reported in previous studies (Barnes et al., 1984; Schacht et al., 2000).
Similar to patch burning in arid climates, time since fire impacted several functional metrics related to previous years' biomass (Augustine and Derner, 2014). Fire reduced herbaceous standing dead metrics and litter cover compared to unburned areas, but both increased as time since fire increased. Inversely, fire increased bare ground compared to unburned areas, but as litter began to accumulate on the ground, bare ground decreased with time since fire increased. These findings are consistent with previous patch burning research in mesic climates, however, the lack of live forb variability following fire is not (Fuhlendorf and Engle, 2004). This is likely owed to the imbalance between vegetation growth and grazing demand allowing graminoids species to rapidly recover and out compete forbs. Conversely, when forage supply corresponds to grazing demand in a burned area, focal grazing in burned areas maintains graminoids at a short, vegetative state, and allows forb species to outcompete and increase in abundance.
Our findings highlight the legacy effect of “management for the middle” to create spatially and temporally uniform landscapes where moderate grazing by cattle attempted to optimize cattle production by reducing variability. Although bison units have been managed independently and fire has been applied to increase biodiversity, the low stocking rates in both bison units likely contributed to the lack of spatial heterogeneity in this study (Fuhlendorf and Engle, 2004). Further, in other grassland systems, conservation-based management using light grazing pressure also resulted in the loss of diversity as a few plant species became dominant by outcompeting others for resources (Towne et al., 2005; Collins and Calabrese, 2012; Scasta et al., 2016). Patch burning has increased heterogeneity and biodiversity in other ecoregions, but alternative patterns of heterogeneity have emerged at multiple scales across patch burning studies (Winter et al., 2011; McGranahan et al., 2012, 2013; Augustine and Derner, 2014). Further research is needed in the Sandhills to identify how fire and grazing interact in a system characterized by high precipitation variability, contrasting topoedaphic features, and sites with different management histories to advance our understanding of how patch burning creates heterogeneity.
All datasets generated for this study are included in the manuscript/Supplementary Files.
JA: study design, data collection, data analysis, and manuscript writing. DT: study design, data analysis, and manuscript editing. CW: data analysis. WS: grant writing and manuscript editing. DW: grant writing.
This work was supported by the Nebraska Environmental Trust (13-176).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We are grateful to Doug Tosoni, Vicki Simonsen, Christine Bielski, Torie Lindsey, Brad Schick, Kyle Schumacher, Cheryl Dunn, and Amanda Hefner for assisting with data collection. We thank the staff of the Niobrara Valley Preserve and Jeremy Hiller for logistical support.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2019.00365/full#supplementary-material
Allred, B. W., Fuhlendorf, S. D., Engle, D. M., and Elmore, R. D. (2011b). Ungulate preference for burned patches reveals strength of fire–grazing interaction. Ecol. Evol. 1, 132–144. doi: 10.1002/ece3.12
Allred, B. W., Fuhlendorf, S. D., and Hamilton, R. G. (2011a). The role of herbivores in Great Plains conservation: comparative ecology of bison and cattle. Ecosphere 2, 1–17. doi: 10.1890/ES10-00152.1
Allred, B. W., Scasta, J. D., Hovick, T. J., Fuhlendorf, S. D., and Hamilton, R. G. (2014). Spatial heterogeneity stabilizes livestock productivity in a changing climate. Agric. Ecosyst. Environ. 193, 37–41. doi: 10.1016/j.agee.2014.04.020
Anderson, R. H., Fuhlendorf, S. D., and Engle, D. M. (2006). Soil nitrogen availability in tallgrass prairie under the fire–grazing interaction. Rangel. Ecol. Manag. 59, 625–631. doi: 10.2111/05-088R2.1
Augustine, D. J., and Derner, J. D. (2014). Controls over the strength and timing of fire–grazer interactions in a semi-arid rangeland. J. Appl. Ecol. 51, 242–250. doi: 10.1111/1365-2664.12186
Augustine, D. J., and Derner, J. D. (2015). Patch burn grazing management in a semiarid grassland: consequences for pronghorn, plains pricklypear, and wind erosion. Rangel. Ecol. Manag. 68, 40–47. doi: 10.1016/j.rama.2014.12.010
Barnes, P., Harrison, A., and Heinisch, S. (1984). Vegetation patterns in relation to topography and edaphic variation Nebraska Sandhills prairie. Fac. Publ. Biol. Sci. 16, 145–158.
Bielski, C. H., Twidwell, D., Fuhlendorf, S. D., Wonkka, C. L., Allred, B. W., Ochsner, T. E., et al. (2018). Pyric herbivory, scales of heterogeneity and drought. Funct. Ecol. 32, 1599–1608. doi: 10.1111/1365-2435.13083
Biondini, M. E., Steuter, A. A., and Grygiel, C. E. (1989). Seasonal fire effects on the diversity patterns, spatial distribution and community structure of forbs in the Northern Mixed Prairie, USA. Vegetatio 85, 21–31. doi: 10.1007/BF00042252
Biondini, M. E., Steuter, A. A., and Hamilton, R. G. (1999). Bison use of fire-managed remnant prairies. J. Range Manag. 52, 454–461. doi: 10.2307/4003772
Bragg, T. B. (1978). “Effects of burning, cattle grazing, and topography on vegetation of the choppy sands range site in the Nebraska Sandhills prairie,” in Proceedings, 1st International Rangeland Congress (Denver, CO), 248–253.
Collins, S. L., and Calabrese, L. B. (2012). Effects of fire, grazing and topographic variation on vegetation structure in tallgrass prairie. J. Veg. Sci. 23, 563–575. doi: 10.1111/j.1654-1103.2011.01369.x
Cummings, D. C., Fuhlendorf, S. D., and Engle, D. M. (2007). Is altering grazing selectivity of invasive forage species with patch burning more effective than herbicide treatments? Rangel. Ecol. Manag. 60, 253–260. doi: 10.2111/1551-5028(2007)60[253:IAGSOI]2.0.CO;2
Debinski, D. M., Moranz, R. A., Delaney, J. T., Miller, J. R., Engle, D. M., Winkler, L. B., et al. (2011). A cross-taxonomic comparison of insect responses to grassland management and land-use legacies. Ecosphere 2, 1–16. doi: 10.1890/ES11-00226.1
Doliner, L. H., and Jolliffe, P. A. (1979). Ecological evidence concerning the adaptive significance of the C4 dicarboxylic acid pathway of photosynthesis. Oecologia 38, 23–34. doi: 10.1007/BF00347821
Dunn, B. H., Smart, A. J., Gates, R. N., Johnson, P. S., Beutler, M. K., Diersen, M. A., et al. (2010). Long-term production and profitability from grazing cattle in the northern mixed grass prairie. Rangel. Ecol. Manag. 63, 233–242. doi: 10.2111/REM-D-09-00042.1
Fuhlendorf, S. D., and Engle, D. M. (2001). Restoring heterogeneity on rangelands: ecosystem management based on evolutionary grazing patterns we propose a paradigm that enhances heterogeneity instead of homogeneity to promote biological diversity and wildlife habitat on rangelands grazed by livestock. Bio Sci. 51, 625–632. doi: 10.1641/0006-3568(2001)051[0625:RHOREM]2.0.CO;2
Fuhlendorf, S. D., and Engle, D. M. (2004). Application of the fire–grazing interaction to restore a shifting mosaic on tallgrass prairie. J. Appl. Ecol. 41, 604–614. doi: 10.1111/j.0021-8901.2004.00937.x
Fuhlendorf, S. D., Engle, D. M., Elmore, R. D., Limb, R. F., and Bidwell, T. G. (2012). Conservation of pattern and process: developing an alternative paradigm of rangeland management. Rangel. Ecol. Manag. 65, 579–589. doi: 10.2111/REM-D-11-00109.1
Fuhlendorf, S. D., Engle, D. M., Kerby, J., and Hamilton, R. (2009). Pyric herbivory: rewilding landscapes through the recoupling of fire and grazing. Conserv. Biol. 23, 588–598. doi: 10.1111/j.1523-1739.2008.01139.x
Fuhlendorf, S. D., Fynn, R. W. S., McGranahan, D. A., and Twidwell, D. (2017). “Heterogeneity as the basis for rangeland management,” in Rangeland Systems Springer Series on Environmental Management, ed D. D. Briske (Cham: Springer Series on Environmental Management), 169–196. doi: 10.1007/978-3-319-46709-2_5
Fuhlendorf, S. D., Harrell, W. C., Engle, D. M., Hamilton, R. G., Davis, C. A., and Leslie, D. M. (2006). Should heterogeneity be the basis for conservation? grassland bird response to fire and grazing. Ecol. Appl. 16, 1706–1716. doi: 10.1890/1051-0761(2006)016[1706:SHBTBF]2.0.CO;2
Fuhlendorf, S. D., and Smeins, F. E. (1999). Scaling effects of grazing in a semi-arid grassland. J. Veg. Sci. 10, 731–738. doi: 10.2307/3237088
Fuhlendorf, S. D., Townsend, D. E., Elmore, R. D., and Engle, D. M. (2010). Pyric-herbivory to promote rangeland heterogeneity: evidence from small mammal communities. Rangel. Ecol. Manag. 63, 670–678. doi: 10.2111/REM-D-10-00044.1
Holechek, J. L., Pieper, R. D., and Herbel, C. H. (2011). Range Management: Principles and Practices. Englewood Cliffs, NJ: Prentice-Hall, 16–526.
HPRCC (2015). High Plains Regional Climate Center. Available online at: http://www.hprcc.unl.edu/ (accessed March 25, 2016).
Kaul, R. (1998). “Plants,” in An Atlas of the Sand Hills, eds A. Bleed and C. Flowerday (Omaha, NE: Resource Atlas, 127–142.
Laughlin, D. C., and Fule, P. Z. (2008). Wildland fire effects on understory plant communities in two fire-prone forests. Can. J. For. Res. 38, 133–142. doi: 10.1139/X07-118
Legendre, P., and Legendre, L. F. J. (1998). Numerical Ecology, 2nd Edn. Kidlington, UK: Elsevier Science.
Limb, R. F., Fuhlendorf, S. D., Engle, D. M., Weir, J. R., Elmore, R. D., and Bidwell, T. G. (2011). Pyric–herbivory and cattle performance in grassland ecosystems. Rangel. Ecol. Manag. 64, 659–663. doi: 10.2111/REM-D-10-00192.1
MacArthur, R. H., and MacArthur, J. W. (1961). On bird species diversity. Ecology 42, 594–598. doi: 10.2307/1932254
McCune, B., Grace, J. B., and Urban, D. L. (2002). Analysis of Ecological Communities. MJM software design Gleneden Beach, OR. Available online at: https://www.researchgate.net/profile/James_Grace/publication/216769990_Analysis_of_ecological_communities/links/0a85e5318e69b2ae7f000000.pdf (accessed March 31, 2016).
McGranahan, D. A., Engle, D. M., Fuhlendorf, S. D., Winter, S. J., Miller, J. R., and Debinski, D. M. (2012). Spatial heterogeneity across five rangelands managed with pyric-herbivory. J. Appl. Ecol. 49, 903–910. doi: 10.1111/j.1365-2664.2012.02168.x
McGranahan, D. A., Engle, D. M., Fuhlendorf, S. D., Winter, S. L., Miller, J. R., and Debinski, D. M. (2013). Inconsistent outcomes of heterogeneity-based management underscore importance of matching evaluation to conservation objectives. Environ. Sci. Policy 31, 53–60. doi: 10.1016/j.envsci.2013.03.005
McGranahan, D. A., Henderson, C. B., Hill, J. S., Raicovich, G. M., Wilson, W. N., and Smith, C. K. (2014). Patch burning improves forage quality and creates grass-bank in old-field pasture: results of a demonstration trial. Southeast. Nat. 13, 200–207. doi: 10.1656/058.013.0203
Meek, M. G., Cooper, S. M., Owens, M. K., Cooper, R. M., and Wappel, A. L. (2008). White-tailed deer distribution in response to patch burning on rangeland. J. Arid Environ. 72, 2026–2033. doi: 10.1016/j.jaridenv.2008.06.002
Official Series Description (2007). VALENTINE Series. Official Series Description. Available online at: https://soilseries.sc.egov.usda.gov/OSD_Docs/V/VALENTINE.html (accessed August 4, 2016).
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., et al. (2018). vegan: Community Ecology Package. R package version. 2.4-6. Available online at: https://CRAN.R-project.org/package=vegan
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O'Hara, R. B., et al. (2016). vegan: Community Ecology Package. Available online at: https://cran.r-project.org/web/packages/vegan/index.html (accessed March 11, 2016).
Patten, R. S., and Ellis, J. E. (1995). Patterns of species and community distributions related to environmental gradients in an arid tropical ecosystem. Vegetatio 117, 69–79. doi: 10.1007/BF00033260
Pfeiffer, K. E., and Steuter, A. A. (1994). Preliminary response of Sandhills prairie to fire and bison grazing. J. Range Manag. 47, 395–397. doi: 10.2307/4002337
Polito, V. J., Baum, K. A., Payton, M. E., Little, S. E., Fuhlendorf, S. D., and Reichard, M. V. (2013). Tick abundance and levels of infestation on cattle in response to patch burning. Rangel. Ecol. Manag. 66, 545–552. doi: 10.2111/REM-D-12-00172.1
R Core Team (2014). R: The R Project for Statistical Computing. Available online at: https://www.r-project.org/ (accessed March 11, 2016).
Robel, R. J., Briggs, J. N., Dayton, A. D., and Hulbert, L. C. (1970). Relationships between visual obstruction measurements and weight of grassland vegetation. J. Range Manag. 23, 295–297. doi: 10.2307/3896225
Scasta, J. D., Duchardt, C., Engle, D. M., Miller, J. R., Debinski, D. M., and Harr, R. N. (2016). Constraints to restoring fire and grazing ecological processes to optimize grassland vegetation structural diversity. Ecol. Eng. 95, 865–875. doi: 10.1016/j.ecoleng.2016.06.096
Scasta, J. D., Engle, D. M., Talley, J. L., Weir, J. R., Stansberry, J. C., Fuhlendorf, S. D., et al. (2012). Pyric-herbivory to manage horn flies (Diptera: Muscidae) on cattle. Southwest. Entomol. 37, 325–334. doi: 10.3958/059.037.0308
Schacht, W. H., Volesky, J. D., Bauer, D., Smart, A., and Mousel, E. (2000). Plant community patterns on upland prairie in the eastern Nebraska sandhills. Agron. Publ. 339, 43–58.
Steinauer, E. M., and Bragg, T. B. (1987). Ponderosa pine (Pinus ponderosa) invasion of Nebraska Sandhills prairie. Am. Midl. Nat. 118, 358–365. doi: 10.2307/2425792
Steuter, A., and Hidinger, L. (1999). Comparative ecology of bison and cattle on mixed-grass prairie. Gt. Plains Res. J. Nat. Soc. Sci. 9, 329–342.
Steuter, A. A. (1987). C3/C4 Production shift on seasonal burns: northern mixed prairie. J. Range Manag. 40, 27–31. doi: 10.2307/3899356
Steuter, A. A., Steinauer, E. M., Hill, G. L., Bowers, P. A., and Tieszen, L. L. (1995). Distribution and diet of bison and pocket gophers in a Sandhills prairie. Ecol. Appl. 5, 756–766. doi: 10.2307/1941983
Stubbendieck, J. (1998). “Range Management,” in An Atlas of the Sand Hills, eds A. Bleed and C. Flowerday (Omaha, NE: Resource Atlas), 239–245.
Stubbendieck, J., Flessner, T. R., and Weedon, R. (1989). “Blowouts in the Nebraska Sandhills: the habitat of Penstemon haydenii,” in Proceedings of the North Americn Prairie Conference (Lincoln, NE).
Stubbendieck, J. L., and Reece, P. E. (1992). EC92-124 Nebraska handbook of range management. Hist. Mater. Univ. Neb. Linc. Ext. 1624.
Taylor, C. A Jr., Twidwell, D., Garza, N. E., Rosser, C., Hoffman, J. K., et al. (2011). Long-term effects of fire, livestock herbivory removal, and weather variability in Texas semiarid savanna. Rangel. Ecol. Manag. 65, 21–30. doi: 10.2111/REM-D-10-00124.1
Tews, J., Brose, U., Grimm, V., Tielbörger, K., Wichmann, M. C., Schwager, M., et al. (2004). Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. J. Biogeogr. 31, 79–92. doi: 10.1046/j.0305-0270.2003.00994.x
Towne, E. G., Hartnett, D. C., and Cochran, R. C. (2005). Vegetation trends in tallgrass prairie from bison and cattle grazing. Ecol. Appl. 15, 1550–1559. doi: 10.1890/04-1958
Twidwell, D., Allred, B. W., and Fuhlendorf, S. D. (2013). National-scale assessment of ecological content in the world's largestland management framework. Ecosphere 4, 1–27. doi: 10.1890/ES13-00124.1
Vermeire, L. T., Mitchell, R. B., Fuhlendorf, S. D., and Gillen, R. L. (2004). Patch burning effects on grazing distribution. Rangel. Ecol. Manag. 57, 248–252. doi: 10.2111/1551-5028(2004)057[0248:PBEOGD]2.0.CO;2
Winter, S. L., Fuhlendorf, S. D., Goad, C. L., Davis, C. A., and Hickman, K. R. (2011). Topoedaphic variability and patch burning in sand sagebrush shrubland. Rangel. Ecol. Manag. 64, 633–640. doi: 10.2111/REM-D-11-00068.1
WSS (2015). Web Soil Survey (WSS) Provides Soil Data and Information Produced by the National Cooperative Soil Survey. Available online at: http://websoilsurvey.nrcs.usda.gov/ (accessed May 24, 2016).
Keywords: grazing pressure, heterogeneity, patch burning, Sandhills grassland, sandy soil ecosystems, traditional grassland management
Citation: Arterburn JR, Twidwell D, Wonkka CL, Schacht WH and Wedin DA (2019) Restoring Fire-Grazer Interactions to Pursue Heterogeneity in Sandhills Prairie. Front. Ecol. Evol. 7:365. doi: 10.3389/fevo.2019.00365
Received: 14 May 2019; Accepted: 17 September 2019;
Published: 02 October 2019.
Edited by:
David Jack Coates, Department of Biodiversity, Conservation and Attractions (DBCA), AustraliaReviewed by:
Carl R. Gosper, Department of Biodiversity, Conservation and Attractions (DBCA), AustraliaCopyright © 2019 Arterburn, Twidwell, Wonkka, Schacht and Wedin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jack R. Arterburn, amFjay5hcnRlcmJ1cm5AdW5sLmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.