 Nicolas Boulbes
Nicolas Boulbes Eline N. van Asperen
Eline N. van Asperen- 1UMR 7194 Histoire Naturelle de l'Homme Préhistorique, Museum National d'Histoire Naturelle-CNRS-Université de Perpignan Via Domitia, EPCC-CERP, Tautavel, France
- 2Department of Biosciences, Durham University, Durham, United Kingdom
- 3Department of Anthropology, Durham University, Durham, United Kingdom
We present an inventory of the progress of recent research on the biostratigraphy and palaeoecology of the genus Equus sensu lato in Europe. Our discussion starts with the new hypotheses concerning the dispersal and evolution of non-caballine equids of the Early and the beginning of the Middle Pleistocene, focusing on recent discoveries, description of taxa, and revised diagnosis. In particular, we deal with the major debate surrounding the number of lineages, “stenonid horses,” related species and the new subgenus “Sussemionus.” Possible phylogenetic scenarios and relationships with extant species are also taken into consideration. There is consensus that the lineage of true horses emerged in Europe at the beginning of the Middle Pleistocene. Their extensive skeletal plasticity enabled them to survive under a wide range of climatic conditions and environments, from interglacial forests to grass-steppes during glacial episodes. They exhibit homogeneous overall morphology but variability in terms of dental and skeletal proportions. Patterns of general body size, muzzle shape, macro- and microdonty, proportions of the limbs, robustness of the metapodials, and breadth of the third phalanx are influenced by global climate, the characteristics of the vegetation (in particular the degree of openness) and substrate, and are governed by Bergmann's and Allen's rules or other environmental pressures. In addition to palaeoecological information, these ecomorphological adaptations paradoxically provide real biochronological details in a given geographical province. Throughout the Upper Pleistocene, horses underwent a size diminution initiated at the end of the Middle Pleistocene. After the Last Glacial Maximum (LGM), investigation of body size in different regions of Europe demonstrates the existence of latitudinal and longitudinal clines, and the fragmentation of the horse population. The so-called European wild ass, Equus hydruntinus, has a wide geographical distribution but seems to be less ecologically flexible, or, alternatively, more specialized than true horses, with which it is often found in sympatry. The latest palaeogenetic studies place the species into phylogenetic context within the Asiatic wild asses, however, palaeontological evidence points to E. hydruntinus as a separate species. Its tooth morphology varies between biogeographical areas.
Introduction
Over the past 15 years, the discovery of new fossils and the revision of reference collections, added to the increased precision of the dating of certain deposits, have led to a reconsideration of the biochronology of the equids of the Pleistocene (Alberdi et al., 1998; Forsten, 1999a; Eisenmann, 2004a, 2006a, 2010; Alberdi and Palombo, 2013; Palombo and Alberdi, 2017; Van der Made et al., 2017). In addition, the progress of palaeogenetics has made it possible to calibrate evolutionary models proposed by paleontology (Orlando et al., 2013; Jonsson et al., 2014; Bennett et al., 2017).
It is generally considered that the genus Equus, which most likely emerged some 4.0–4.5 Ma (Orlando et al., 2013), includes all monodactyl equids present in Europe, and the stenonid group (horses related to Equus stenonis) was the first to disperse at the Plio-Pleistocene boundary (Equus-Elephant event, Lindsay et al., 1980). Equus livenzovenzis, recorded during the Middle Villafranchian, is for some authors the first species recognized in Eurasia and considered as the phylogenetic root of all the European stenonoid species (Azzaroli, 1992; Alberdi et al., 1998; Palombo and Alberdi, 2017; Bernor et al., 2018; Figure 1). However, its definition remains disputed, in particular because of the heterogeneity of the material of the type locality (Livenzovka, near Rostov-Don, Russia), doubts about the chronology (Forsten, 1998a), and the disagreement about the taxonomy of contemporaneous fossils recorded in Western Europe and attributed to this taxa (i.e., at Montopoli in Italy, and Huélago, El Rincón in Spain; Forsten, 1999a; Eisenmann, 2004a).
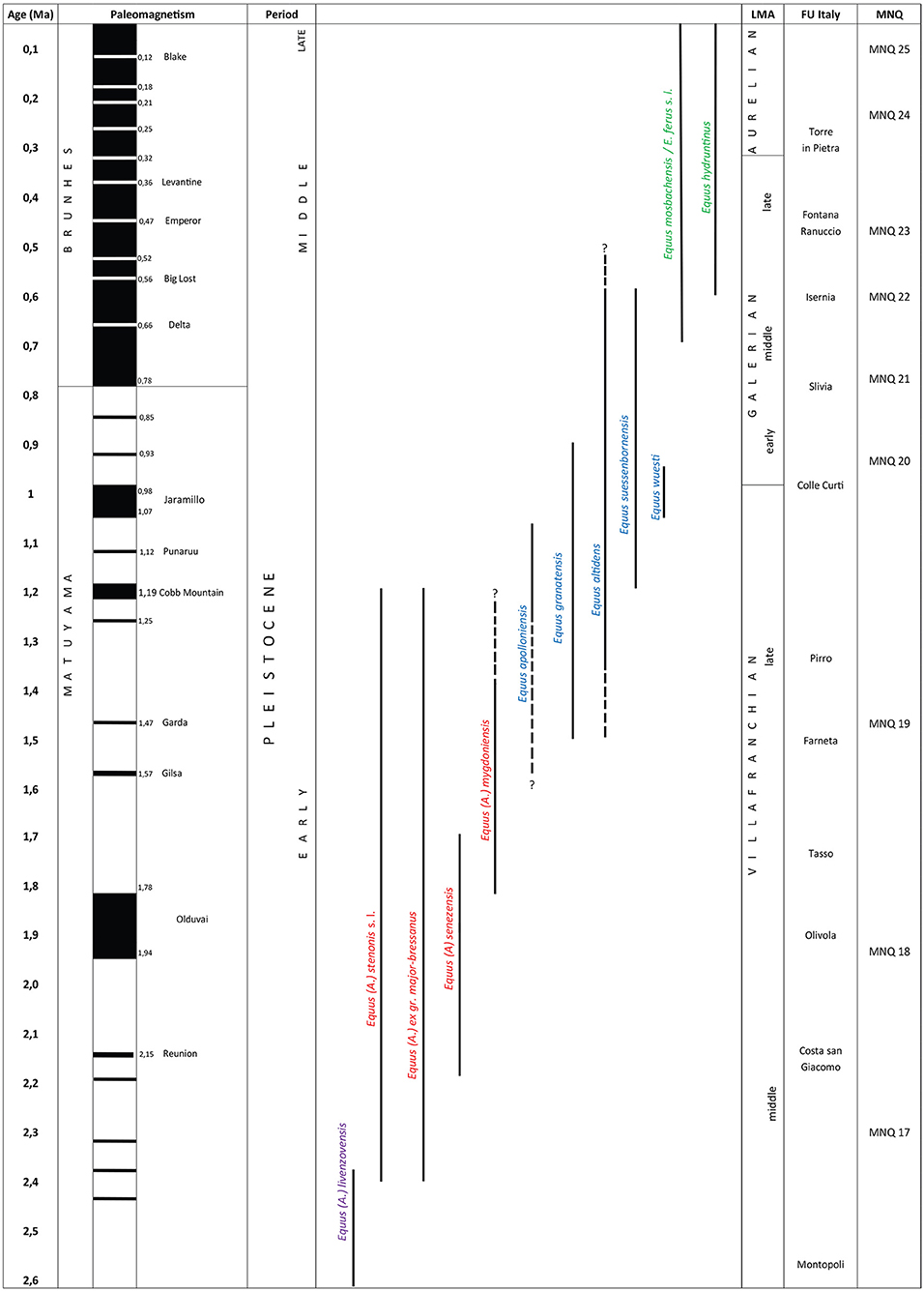
Figure 1. Biochronological overview of the genus Equus s. l. from Europe.
According to Eisenmann and Baylac (2000) and Eisenmann (2006a, 2017), modern equids—genus Equus–can be differentiated from the earlier genera Plesippus and Allohippus (Stenonids) by their basicranial proportions. Following this outline, the first cranium of Equus described in Europe is that of Apollonia in Greek Macedonia (=Equus apolloniensis, Koufos et al., 1997; 1.2–0.9 Ma, Koufos and Kostopoulos, 2016) and the latest cranium of stenonids (Allohippus according to Eisenmann) could be that of Ceyssaguet with an age of 1.2 Ma (Equus stenonis, Aouadi and Bonifay, 2008). Beyond Europe, the crania from Nalaikha (0.9 Ma) in Mongolia [= E. (Hemionus) nalaikhaensis, Kuznetsova and Zhegallo (2009)] and from Kolyma in Siberia at the beginning of the Middle Pleistocene (=E. coliemensis) also belong to Equus (Eisenmann and Kuznetsova, 2004). According to Eisenmann (2006a, 2010, 2017), similarities in the morphology of the teeth suggest, even in the absence of the cranium, a common origin within the branch of ≪ Equus ≫ for certain equids, at least from 1.5 Ma and maybe soon before around 2.5 Ma just above the Gauss-Matuyama limit (Vatera Formation, Greece). These equids (E. granatensis, E. altidens, E. hipparionoides, E. coliemensis, E. suessenbornensis) are grouped together in the subgenus of Equus “Sussemionus” (Eisenmann, 2010). As a consequence, in this model, Allohippus, and Equus could have co-occurred in Europe for more than 1.0 Ma. These characters, however, are not constant, even within the same series. As a consequence, their presence is a good indicator but their absence is not and Allohippus “stenonine” patterns may occur in Sussemiones (Eisenmann and Boulbes, in preparation). During this period, this subgenus had great success from North America to Ethiopia, including both species adapted to relatively dry climatic conditions (Equus granatensis-like), and others adapted to more humid environments (Equus coliemensis-like). Recent molecular and osteological analyses indicate that this subgenus survived until 45,000 BP in Khakassia in southwestern Siberia (Equus odovovi, Eisenmann and Sergej, 2011).
According to Palombo and Alberdi (2017) this splitting of Pleistocene equids from Eurasia in different genus/subgenus names requires a great deal of caution, taking into account the homogeneity but also the large intra- and interspecific variation shown by fossil monodactyl horses. In this paper we will consider the genus Equus sensu lato, taking into account the different points of view of the specialists. The first part is dedicated to the non-caballoid equids from the Early Pleistocene and the start of the Middle Pleistocene. The second part deals with the true or caballoid horses, especially their ecology regarding the high level of plasticity of this group. The final part, about the European wild ass Equus hydruntinus, develops different aspects of its evolution.
Methods
The nomenclature used for the morphological description of the teeth is illustrated in Figure 2. The occlusal measurements were taken from the outer edges of the enamel excluding the cement. Average occlusal size (occlusal surface) can be used to minimize the impact of deformation of the teeth by wear on the analysis. The Protoconal Index (PI) represents the ratio of protoconal length to total length i.e., mesio-distal diameter (IP = Lprot/DMD x 100). Increase in the length of protocone, dissymetry of the double knot and reduction of the vestibular groove (ectoflexid) are generally considered as progressive characters.
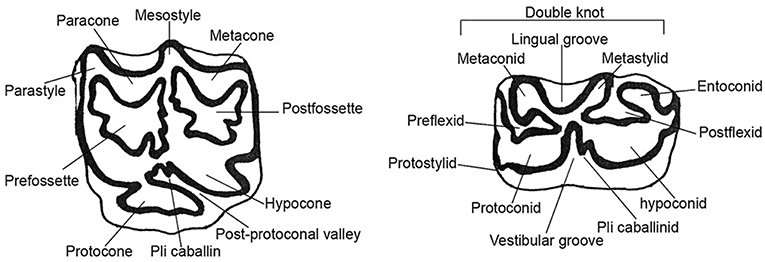
Figure 2. Nomenclature used for upper and lower cheek teeth. Occlusal view.
The measurement system for the postcranial skeleton is based on that established by Eisenmann (2019).
For the metacarpal several indices are calculated: slenderness index 1 (SI 1): breadth at the middle of the diaphysis (3)/maximal length (1) (Alberdi et al., 1998); slenderness index 2 (SI 2): breadth at the distal articular end (11)/maximal length (1) (Koufos, 1992); diaphysis flatness (DF): depth of the diaphysis (4)/breadth of the diaphysis (3) (Eisenmann, 2002); caballine index (CI): distal supra-articular breadth at tuberosities (10)/distal articular breadth (11) (Prat, 1980); proximal flatness (PF): proximal articular breadth (5)/proximal articular depth (6). The development of the sagittal crest is expressed by the Keel Index (KI) (Gromova, 1949) which is the distal maximal antero-posterior diameter of the medial condyle (14) expressed as a percentage of the distal maximal antero-posterior diameter of the keel (12). Gracility, flatness of proximal end, low development of distal tuberosities and protrusion of the keel are usually considered as advanced features (Eisenmann, 1979).
Simpson's ratio diagrams (Simpson, 1941) are used for comparisons of the general conformation i.e., proportion of the limbs (lengths of long bones) and metacarpals. The standard is chosen according to the species to be compared. Equus hemionus onager is most commonly used as a reference for log ratio diagrams; Przewalski's horse for caballoid or true horses; Equus africanus for Equus hydruntinus.
Equids of the Early Pleistocene and the Start of the Middle Pleistocene
Equus stenonis and Relatives
The typical stenonid horses (genus ≪ Allohippus ≫ following Eisenmann) are recorded in Europe from the end of the Pliocene and disappeared at the latest at the end of the Early Pleistocene. Based on an estimate of their body weight, their morphology, and their skeletal proportions, these fossils correspond to multiple species which are unequally documented. According to Eisenmann (2004a) these species formed a monophyletic group characterized by a deep nasal incision (apomorphy) and other characteristics recurring within this group, such as a weakly developed protocone, a rounded double knot, pointed linguaflexid, and a deep ectoflexid on the molars, cups on incisors, and ≪ flattened ≫ metapodials.
Equus stenonis Cocchi 1867
This species was described from material from an unregistered locality in the surroundings of Terranova in the Upper Valdarno Basin (Tuscany, Italy; De Giuli, 1972; Privat Defaus, 1986). According to Forsten (1999a) and Palombo and Alberdi (2017), the many forms of E. stenonis correspond to local variations depending on eco-geographic and maybe chronological factors, and they are little different from the hypodigm of the Upper Valdarno. The revised diagnosis after Alberdi et al. (1998) retains that Equus stenonis corresponds to a medium to large-sized horse with “heavy” proportions characterized by a long stylopod (humerus, femur; Figure 4); a skull with a deep nasal incision, elongated and narrow muzzle and very long diastema; the braincase is small in relation to the length of the face and strongly convex; the upper teeth have a short protocone (Protoconal Index lower than 40, Gromova, 1949; Table 1), the lower teeth have a typical double knot with a rounded metaconid and a slightly angular metastyloid and a deep lingual “V” groove; the vestibular groove is variable, deep or shallow on molars as in the type material (Figure 3A); metapodials are robust and “flattened” (low DF index and high PF index), the distal keel is well-developed with (low KI index; Table 1).
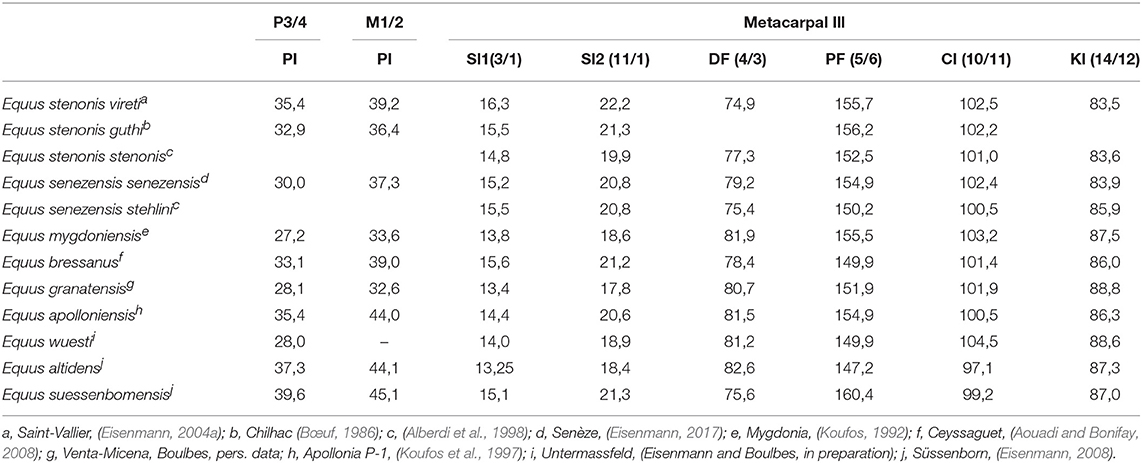
Table 1. Index on teeth and metacarpal for selected Early and Middle Pleistocene equids.
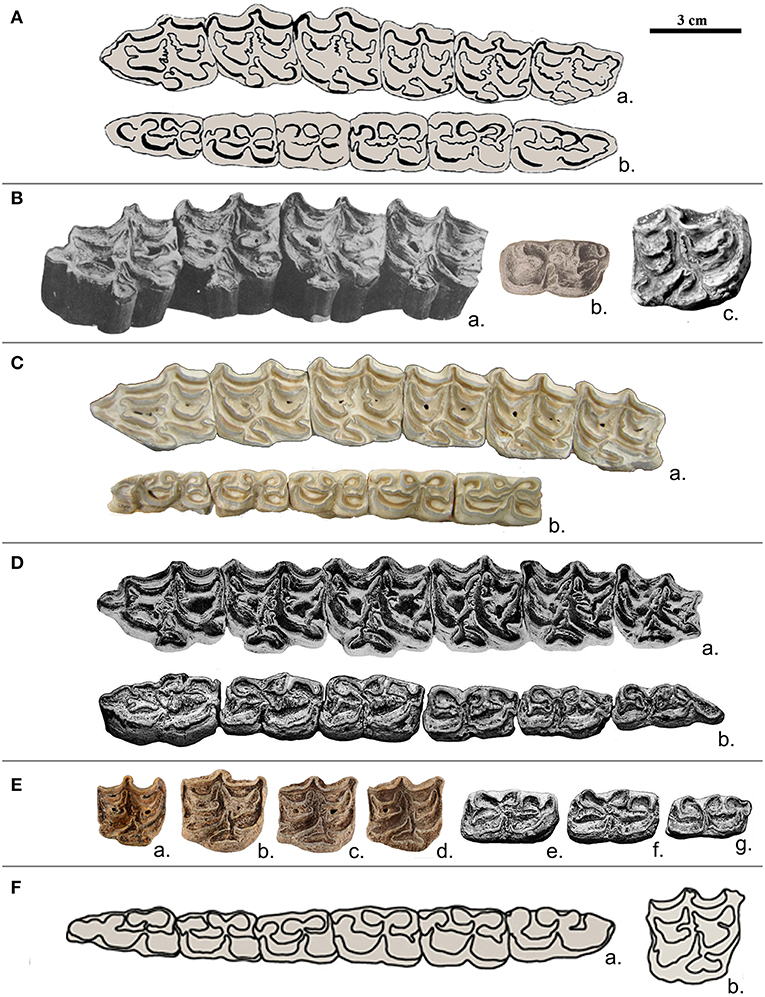
Figure 3. Upper and lower cheek teeth from different Pleistocene equids. Occlusal view. (A) Equus stenonis, Upper Valdarno, a- left P2-M3, Type; b-right P2-M3 (after Azzaroli, 1965 in Caloi, 1997). (B) Equus major, Chagny, a-left P2-M1, b-left M2 (Delafond and Depéret, 1893); c-right P34, se 338, Senèze (Eisenmann, 2017). (C) Equus granatensis, Venta Micena, a-left P2-M3, Type, b-right P3-M3, VM 8297 (Boulbes, pers. photo). (D) Equus suessenbornensis, Süssenborn, a-left upper cheek teeth P2-M3, Type, b-left lower cheek teeth, P2-M3, S 6882 (Eisenmann, 2008). (E) Equus altidens (including E. marxi), Süssenborn, a-right P4, S 1365, M1, S 1366, M2, S 1367, b-left P3, S 1369, P4, S 1368, M2 (S 1370) (Eisenmann, 2019). (F) Equus apolloniensis, Apollonia P-1, a-right P2-M3, APL 147, b-right P4, APL 148 (after Koufos et al., 1997, in Eisenmann and Boulbes, in preparation).
Several subspecies have been recorded (see Palombo and Alberdi, 2017), the samples from the sites in Valdarno and neighboring localities (Matassino, Olivola, about Olduvai event) are usually referred to the nominal subspecies (De Giuli, 1972). The best known are certainly the more robust form from Saint-Vallier (E. stenonis vireti Prat, 1964), descibed in detail with new collection by (Eisenmann, 2004a, “Allohippus” s. vireti), and the stenonid equid from Chilhac (E. stenonis guthi Bœuf, 1986), both dated to 2.5 and around 2.4 Ma (Nomade et al., 2014). The morphology and dimensions of Equus stenonis olivolanus (Caloi, 1997) (Olivola) mainly fall within the range of the nominal subspecies (Palombo and Alberdi, 2017). Equus stenonis pueblensis (Caloi, 1997) (Puebla de Valverde, located over the Reunion event and bellow the Olduvai chron (Sinusía et al., 2004) is considered as a synonym of the form of Chilhac (Alberdi et al., 1998) or a true species “Allohippus” pueblensis (Eisenmann, 2017). In eastern Mediterranean, Equus stenonis is abundant, especially during middle Villafranchian, and belong to one form similar to Saint-Vallier and La Puebla de Valverde (Koufos, 2001); one of the best described sample is that of Sésklo (Thessaly, Greece, MNQ 17; Athanassiou, 2001).
The temporal range of Equus stenonis sensu lato could be 2.5 Ma (Saint-Vallier, Chilhac) to 1.2 Ma (Ceyssaguet; Figure 1).
Small and/or Gracile Stenonids
Originally described as a subspecies of Equus stenonis (Prat, 1964), the small to medium-sized equid from Senèze (2.1–2.2 Ma, Pastre et al., 2015), Equus senezensis, was later elevated to the rank of species (Alberdi et al., 1998; Eisenmann, 2004a). Apart from its small size, Equus senezensis has a morphology (teeth and bones) and general proportions similar to Equus stenonis (Figure 4, Table 1). Alberdi et al. (1998) recognize two geographical subspecies, the nominotypical subspecies, recorded at Senèze, and the subspecies “Equus senezensis stehlini,” including the younger fossils from E. stehlini described by Azzaroli (1965) in the late Villafranchian Upper Valdarno.
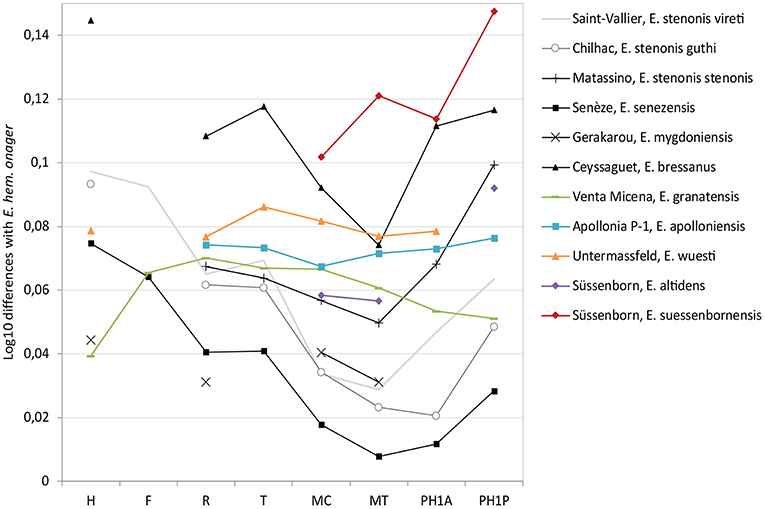
Figure 4. Simpson's ratio diagram of bone lengths of European Early and Middle Pleistocene horses. Reference: Equus hemionus onager (Eisenmann, 1991a). H, Humerus; F, Femur; R, Radius; T, Tibia; MC, Metacarpal; MT, Metatarsal; PH1A, First Anterior Phalanx; PH1P, First Posterior Phalanx. Data: Saint-Vallier (Eisenmann, 2004a); Matassino (Eisenmann, 1999); Senèze (Eisenmann, 2017); Gerakarou (Koufos, 1992); Ceyssaguet (Aouadi, 2001); Apollonia P-1 (Koufos et al., 1997); Venta Micena (Boulbes, pers. data); Untermassfeld (Eisenmann and Boulbes, in preparation); Süssenborn (Eisenmann, 1999, 2008).
Equus senezensis stehlini, often considered as a different species, is smaller and characterized by proportionally shorter metapodials, recorded mainly in Italy (late Villafranchian from Casa Frata (1.7 Ma), Upper Valdarno, and Farneta, Val di Chiana; Azzaroli, 1965, 1984; Caloi and Palombo, 1982). During the second half of the Early Pleistocene, E. stehlini replaced E. stenonis in Italy (Azzaroli, 1965). Palombo et al. (2017) tentatively refer the equid from the middle Villafranchian site of Coste San Giacomo (Anagni, central Italy) dated around 2.2 Ma to Equus aff. senezensis stehlini (Figure 1). This recent discovery could roll back the appearance of this form much earlier than expected (Palombo and Alberdi, 2017).
E. mygdoniensis (Koufos, 1992) has been described in the locality of Gerakarou-1 (Mygdonia Basin) in Greek Macedonia (early Late Villafrancian, ~1.8 Ma). It is a small equid with a short, wide muzzle, short dental rows, short protocones (IP index lower than other stenonids, Table 1), and elongated and slender metapodials (Koufos, 1992; low SI indices, Table 1) but with flat proximal end as E. stenonis and unlike E. altidens/granatensis group. According to Eisenmann (2004a), the equid from Gerakarou belongs to the stenonid group (Allohippus) but based on the differences with Equus stenonis (small size, cranial proportions, slender metapodials), the Gerakarou equid, originally described as a subspecies of Equus stenonis (Koufos, 1992), should be given a specific status (Eisenmann, 2004a). This species is likely to be present in other Greek localities of a similar age (e.g., Pyrgos, Krimni 1/2, Riza 1/2; Koufos, 1992) and may be at Tsiotra Vryssi also in the Gerakarou Formation, but with a younger age, intermediate between the localities Gerakarou-1 and Apollonia-1 (Konidaris et al., 2015). E. mygdoniensis is possibly present in Turkey (Denizli basin, Boulbes et al., 2014), Russia (Livenzovka and Psekups), and Valdarno in the wider sense (Eisenmann, 2004a). The stenonoid horses from Libakos and Polylakkos (Western Macedonia, Greece) are very close to E. mygdoniensis (including in Equus altidens, Gkeme et al., 2017). The taxonomic validity of this species is, however, questioned by other authors who consider that most of the fossils reported to it correspond to Equus senezensis/Equus stehlini and/or Equus altidens groups (Alberdi et al., 1998; Forsten, 1999a; Gkeme et al., 2017; Palombo and Alberdi, 2017).
Eisenmann (2017) also supports the close relationship of the Senèze equid with those of Gerakarou (Allohippus mygdoniensis) and Upper Valdarno (Allohippus stehlini), in particular with regard to certain common characteristics such as short-muzzled skulls, but keeps them at a species level.
Very Large Stenonids
There is consensus that during the Villafranchian, sometimes together with medium or small-sized forms, one or several species occurs with a very large size and characteristics that are similar to stenonids (Viret, 1954; Prat, 1980; Azzaroli, 1990; Alberdi et al., 1998; Forsten, 1999a; Eisenmann, 2003, 2004a, 2017; Table 1, Figure 4, “Equus bressanus” from Ceyssaguet”). This material is generally designated under the name Equus major Depéret in (Delafond and Depéret, 1893) ex Boule, poorly defined from the material of Chagny (revised by Viret, 1954; Figure 3B). The stratigraphic position of the fossils is not clearly known. Alternatively, Eisenmann (2017) distinguishes several groups of large (robust or gracile) to very large stenonids but tentatively refers the material from Chagny to the E. suessenbornensis group.
The sporadic presence of large stenonids s. l. is mentioned in several localities in France and in particular at Senèze (Prat, 1980; Eisenmann, 2017); in Germany in Schernfeld (Musil, 1992); in the Netherlands at Tegelen (Viret, 1954); in Great Britain at East Runton (Norfolk) and Italy at Upper Valdarno (Azzaroli, 1990). The equid described at Fonelas P-1 (Guadix-Baza depression, Spain), older than Olduvai episode (around 2 Ma, Arribas et al., 2009), has affinities with Equus cf. major (Garrido, 2008). In eastern Europe, Equus athanasiui described at Tetoiu-Valea Graunceanului (Romania) (Samson, 1975), a species possibly also presents at Oosterschelde, Forest Bed, El Rincón and Ceyssaguet according to Forsten (1999a). A large and gracile stenonian horse, under study, occurs in the lower Pleistocene layers (13 to 11b) of Kozarnika (Bulgaria) (Sirakov et al., 2010).
The small amount of material belonging to this group and the different points of view of specialists make it difficult to establish a biochronological framework and determine the morphometric and temporal range of the species (Alberdi et al., 1991). Musil (1992) considers that the chronological distribution of this Villafranchian species is between 2.2 and 1.7 Ma. However, “E. bressanus” (=Equus major) seems to be present later around 1.2 Ma (Ceyssaguet, Aouadi and Bonifay, 2008). According to Eisenmann (2017), very large “Allohippus” occurs from 2.5 Ma (Oasele, Romania, Samson, 1975) to around the Brunhes-Matuyama limit (Würzburg-Schalksberg, Germany; Mauser, 1987; Figure 1).
E. granatensis/E. altidens Group, Equus wuesti, and Equus suessenbornensis
Equus granatensis Alberdi and Ruiz Bustos (1985) refers to the slender equid of Venta Micena (Orce, Granada), situated above the Olduvai subchron, with an age around 1.5 Ma (Martínez-Navarro et al., 2011). On the upper teeth the protocone is very short (Table 1), often without an anterior part. The shape of the double knot varies from typically rounded stenonine, separated by a pointed and deep lingual groove, to a rather hemionine type with an elongated metaconid and pointed metastylid, separated by a shallow lingual groove (Eisenmann, 1999). The vestibular groove is very deep and often touches the lingual one on molars; the ectoflexid may also be deep on P3 and P4; protostylids are frequent in premolars (Figure 3C). The general proportions of the limbs, different from E. stenonis/E. senezensis, show cursorial adaptations with elongated distal bones (Figure 4). The metapodials are slender (diaphysis and extremities) with a deeper diaphysis and proximal end (high DF index and low PF index), and a distal keel poorly developed in comparison to E. stenonis and E. senezensis (Table 1); as in hemiones the difference in length between the first anterior and posterior phalanges is important (Eisenmann, 1999). In our opinion and following Eisenmann (1999), the morphological differences of the teeth with both E. stenonis and E. altidens (including E. marxi) could justify a specific level. E. granatensis is mainly recorded in different localities in the south of Spain from 1.5 to 0.8 Ma (Figure 1), in particular in the Guadix-Baza basin (Venta Micena, Lachar, Fuensanta, Fuente Nueva 3, Barranco León 5, Marín, 1987; Alberdi and Ruiz Bustos, 1989; Alberdi et al., 1998; Alberdi, 2010) and according to Eisenmann in the site of Cueva Victoria (0.9–0.8 Ma, Gibert et al., 2016), although these fossils were identified as Equus altidens altidens by Alberdi and Piñero (2015). Equus altidens from epivillafrachian levels of Vallparadis (post Jaramillo, around 0.85 Ma, Duval et al., 2015) have short protocones (Aurell-Garrido et al., 2010) as in E. granatensis.
Equus altidens (Von Reichenau, 1915) was described (together with E. suessenbornensis) from the early Middle Pleistocene site of Süssenborn, most likely accumulated during marine oxygen isotope stage (MIS) 16, with an age of 0.65 Ma (Kahlke, 2014). There is disagreement about the number of species present in the site of Süssenborn due to the heterogeneity of the material and the long time sequence of the stratigraphy (Musil, 1969; Forsten, 1986; Eisenmann, 2008). Equus altidens is a medium-sized and slender species. Revised diagnoses (Forsten, 1986; Alberdi and Palombo, 2013) include the third species described at Süssenborn “Equus marxi” (Musil, 1969): the teeth have a small and narrow crown; the morphology of the protocone of the upper teeth is variable, often indented lingually with an elongated distal part, resulting high values of Protoconal Index in comparaison to E. granatensis (Table 1); the mesostyle sometimes divided into two; a deep post-protoconal valley; and a pli caballin moderately developed or even absent on worn molars. Lower teeth often show a pli protostylid fold including P2; the ectostylid is sometimes present but rare; the double knot is asymmetric; the ectoflexid is deep on molars (Figure 3E). The skeleton is gracile and metapodials (similar to E. granatensis) are more slender than in all other stenonids forms; distal keel is less developped and distal tuberosies are not so strong (CI index < 100) than other species (Table 1). Following several authors (Alberdi et al., 1998; Forsten, 1999a; Alberdi and Palombo, 2013) Equus altidens also includes the fossils of the site of Venta Micena renamed Equus altidens granatensis (Alberdi et al., 1998) and those of the small equine of Akhalkalaki in Georgia (1.0 to 0.8 Ma) (=Equus hipparionoides, Vekua, 1986). Eisenmann (1999, 2010) agrees with the proximity of the Süssenborn slender species and the equid from Venta Micena but gives a specific rank to E. granatensis based on the relatively short protocones compared to E. altidens. She considers that Equus altidens is too poorly defined and should be used with caution.
Disagreements make it difficult to trace a reliable biochronology for Equus altidens. The most extensive suggests the concurrent arrival around 1.8 Ma of the species in Western Asia (E. aff. altidens from Dmanisi, Georgia, Lordkipanidze et al., 2007) and in Southeastern Europe (Gkeme et al., 2017). Other possibility is that it dispersed to Europe from China around 1.2 Ma (Van der Made, 2013). According to Alberdi and Palombo (2013) Equus altidens altidens was first recorded in Italy around 1.5 Ma (Selvella and Pirro Nord LFAs). However, the teeth that have the most affinity (deep postprotoconal valley, elongated protocone), with those of the type locality are around and younger than 1 Ma: Bois de Riquet (Bourguignon et al., 2016; Lozano-Fernández et al., 2019), Soleihlac (Prat, 1980; Lacombat, 2005), Pakefield (Lister et al., 2010), Voigstedt (Musil, 1965), Atapuerca Gran Dolina TD6 (Van der Made, 1999), Huescar-1, Cullar de Baza-1 (Alberdi and Ruiz Bustos, 1989). The site of Venosa-Loreto, slightly younger than Notarchirico with an estimate age around 0.6 Ma (Pereira et al., 2015), could represent its last appearance (Alberdi and Palombo, 2013; Figure 1). Some elements (metapodials, teeth) found in the site of Mosbach (0.5 Ma) show affinities with the remains of Süssenborn (Eisenmann, 2008) and merit further investigation.
The equid of the Epivillafranchian site of Untermassfeld (Thuringia, Germany), Equus wuesti (Musil, 2001), represents another species, somewhat larger and less gracile (Table 1), but less distinct in terms of its proportions than Equus altidens (Musil, 2001) (Figure 4). Musil (followed by Lister et al., 2010) considered the Untermassfeld horse as the ancestor of E. altidens, in spite of the presence of the latter species in localities older than the German site (Palombo and Alberdi, 2017). The site is positioned around the Jaramillo polarity subzone with an age of approximately 1.0 Ma (Kahlke, 2006; Figure 1). Unfortunately, no skull material was found and tooth remains are not very numerous whereas the postcranial skeleton is well-represented. Metacarpal has deeper diaphysis and proximal end, the distal tuberosities are very strong (Table 1). Descriptions of new and older material of the species and comparisons confirm that Equus wuesti represents another species inside the Sussemione group close to but different from the equids of Venta Micena (in particular the teeth) and Akhalkalaki (Eisenmann and Boulbes, in preparation).
Equus suessenbornensis (Wüst, 1900) is a large species with a mozaic of features (Musil, 1969; Gromova and Dubrovo, 1975; Forsten, 1986; Azzaroli, 1990; Alberdi et al., 1998; Eisenmann, 2006a). The main characteristic of the teeth is the high level of enamel folding and converging characters with true horses (caballines) regarded as homoplasy. The upper teeth have an elongated and asymmetric protocone (IP index on M12 = 45, Table 1) with a concave lingual border, a wide mesostyle on premolars, a marked groove on styles at least at some distance from the crown, and a wide-based pli caballin. Lower teeth show certain cingulate structures such as protostylid or ectostylid, the vestibular groove is always deep on molars and often in contact with the lingual side (Figure 3D). Metapodials are longer and the extremities are robust; diaphysis and more specially the proximal articulation are “flattened” (high PF index, Table 1); the distal keel is poorly developed (Table 1); the metatarsal is elongated (Figure 4). Some authors have considered Equus suessenbornensis as the first archaic true horse or an evolutionary intermediate between stenonoids and caballoids (Gromova, 1949). However, the double knot appears to be very different from that characteristic in horses, although more derived than in E. stenonis (Forsten, 1986, 1999a; Alberdi et al., 1998; Palombo and Alberdi, 2017). Equus verae (Sher, 1971) of north-eastern Siberia (Chukochya, about Jaramillo event), which is characterized by teeth with a similar morphology and metapodials with identical proportions to Equus suessenbornensis, could present a synonym (Forsten, 1986) or a very similar species (Eisenmann and Kuznetsova, 2004). The cranium from Kolyma (north-east Siberia), referred to E. coliemensis (Lazarev, 1980) and dated to the end of the Early Pleistocene, belongs in the same group (Eisenmann, 2010).
In Europe Equus suessenbornensis is also present in Germany in the Middle Pleistocene site of Voigtstedt (Musil, 1965), in the Czech Republic at Stranska Skala (Musil, 1972), in Italy in several localities between the latest Villafranchian (Farneta and Pirro faunal units) to the middle Galerian (Caloi and Palombo, 1987; Alberdi et al., 1988; Alberdi and Palombo, 2013), in Spain at Fuente Nueva-3, Barranco León-5, Cueva Victoria, Huéscar-1, Cúllar de Baza-1, (Alberdi and Ruiz Bustos, 1989; Alberdi, 2010; Alberdi and Piñero, 2015), in France at the sites Bois de Riquet, Nauterie, and Soleilhac (Prat and Thibault, 1976; Prat, 1980; Lacombat, 2005; Bourguignon et al., 2016), in England in the Cromer Forest Bed Formation, Lister et al., 2010). In Eastern Europe Equus suessenbornensis was likely present in Ukraine (Tiraspol) (Gromova and Dubrovo, 1975), as well as in Romania (Samson, 1975) and in Western Asia in Georgia (Akhalkalaki) which delivered the best-known collection for the species (Vekua, 1986). The presence of this species in older sites as in Italy (Pirro Nord and Selvella, Alberdi and Palombo, 2013) can be debated. The measurements given by the authors of the specimens that belong to this taxon show a relatively small body size, taking into account the variability of the best known sample of Akhalkalakhi (MT III length between 300 and 330 mm). Other localities, for example Dursunlu in Turkey (1.0 and 0.8 Ma) (Yigit, 1998; Güleç et al., 2009), merit further investigation. Indeed, some teeth seem really characteristic of the species.
Taking into account this wide geographical range from Beringia to Western Europe (Eisenmann and Kuznetsova, 2004), the range of the type species based on material similar to the hypodigm (complex enamel, long protocone, and metapodials with robust extremities) (North-East Siberian, Akhalkalaki, Orce, Cueva Victoria, Süssenborn, Venosa) is comprised between the end of Early Plesitocene around 1.2 and 0.6 Ma (Figure 1).
Phylogeny of Equus altidens/granatensis and Equus suessenbornensis
Scholars are divided about the phylogeny of Equus altidens/granatensis and Equus suessenbornensis–a local European evolution or a dispersal event from Asia and or Africa. In the scenario proposed by Eisenmann (2010), cf. Introduction, they belong to the new subgenus of Equus “Sussemionus” and originated probably in North America and are not linked with the stenonid group (Allohippus for instance). Sussemionus is defined in reference to the mixture of morphological characters shared in the fossil equids from Süssenborn and extant hemiones (deep postprotoconal groove, multiple plis caballin or with very large base, sometimes very short protocones on upper teeth, and elongated/bilobated metaconids, presence of stylids and deep ectoflexid on lowers).
However, this point of view is not shared by other authors who consider that both species belong to the stenonid group (Forsten, 1986, 1999a; Azzaroli, 1990, 1992; Alberdi et al., 1998; Alberdi and Palombo, 2013; Palombo and Alberdi, 2017). In this scheme a local evolution in Europe from a stenonid stock is proposed for both species. With an overall trend from robust to gracile, Equus altidens could represent the latest form of the stenonid group (Alberdi et al., 1998; Piñero and Alberdi, 2015). Intermediate chronological forms in relative proportions (Selvella, Libakos, and Pirro Nord) provide support for this model (Van der Made et al., 2017) at least in Italy, but points of view diverge about the taxonomy of the equids from these localities (De Giuli, 1987; Azzaroli, 1990; Forsten, 1999a; Eisenmann, 2004a, 2006a; Alberdi and Palombo, 2013; Gkeme et al., 2017). Alternative scenarios propose that E. altidens evolved from the African species E. tabeti (Aïn Hanech, Algeria), and that it dispersed shortly after 1.8 Ma to Europe (Guerrero-Alba and Palmqvist, 1997).
Concerning Equus suessenbornensis, some authors highlight the phylogenetic link with Equus major (Equus bressanus) (Grossouvre and Stehlin, 1912; Alberdi et al., 1998; Forsten, 1999a; Aouadi and Bonifay, 2008; Palombo and Alberdi, 2017). According to Alberdi et al. (1998), the E. major-E. suessenbornensis monophyletic group may have stemmed from E. livenzovensis. In accordance with this, remains of large equids from the Upper Villafranchian and Lower Galerian are commonly referred to Equus ex. gr. bressanus-suessenbornensis (Rook et al., 2013). Due to the heterogeneity of the large stenonid group (E. major-bressanus) and the hypothesis including E. suessenbornensis in the subgenus Sussemionus, this link remains uncertain.
Ecology of Equus altidens Sensu Lato and Equus suessenbornensis
These two species, in addition to the Süssenborn type locality, are frequently found sympatrically in many other deposits. The association is particularly well-documented in Akhalkalaki, Georgia (1.0 to 0.8 Ma) (Vekua, 1986), in several sites in the Guadix-Baza basin in southern Spain between 1.4 and 0.9 Ma (Alberdi and Ruiz Bustos, 1989; Alberdi et al., 1998; Alberdi, 2010), in the region of Murcia Cueva Victoria and Quibas (Alberdi and Piñero, 2015; Piñero and Alberdi, 2015), in Italy between 1.5 and 0.6 Ma (Alberdi and Palombo, 2013) and in France (Bois de Riquet, Bourguignon et al., 2016; Figure 5). They could both be represented in the Czech Republic at Stranska Skala (Musil, 1995) and in Romania (Samson, 1975). The co-occurrence of the two species seems to be demonstrated in early Middle Pleistocene England at the West Runton and Pakefield sites (Lister et al., 2010). It seems that at lower latitude (Spain, Italy, France) the slender species is more common than Equus suessenbornensis (Alberdi and Piñero, 2015; Bourguignon et al., 2016; Palombo and Alberdi, 2017). It should also be noted that from new discoveries in the site of Untermassfeld Equus wuesti is associated with a large form sharing affinities with E. suessenbornensis group (Eisenmann and Boulbes, in preparation).

Figure 5. Equus altidens (A) and Equus cf. suessenbornensis (B) from Bois de Riquet (US2). (A) Right proximal metacarpal (BDR-X37b-61b); (B) left proximal metacarpal (BDR-Z34d-114).
The large difference in body mass (Figure 5) between Equus altidens and Equus suessenbornensis prevented any interspecific competition between them and led to the use of different ecological niches (Alberdi et al., 1998; Alberdi and Palombo, 2013). According to (Alberdi and Piñero, 2015, p. 325) “the large horses lived on open and dry habitats (steppes), whereas smaller ones lived on more closed and wet habitats (savanna-mosaic, woodlands).” From a locomotor adaptation point of view, the relative length of the Equus altidens/granatensis metapodials in comparison with the proximal bones of the limbs, and their gracility, suggests a more pronounced cursorial capacity than in the other Villafranchian species and the frequenting of more open landscapes and certainly more arid biotopes (Eisenmann, 1984, 2010; Caloi, 1995). In addition, the proportions of the muzzle and the morphology of the row of symphyseal incisors show that these two species occupied a particular ecological niche and exploited different food resources (Caloi, 2002). The relatively short and wide muzzle proportions in Equus altidens are consistent with a highly specialized grazer diet. Equus suessenbornensis has a wide but relatively elongated muzzle that may imply a less strict preference for monocotyledonous short grass. However, during periods of low availability of resources, the diet of these species, in particular Equus suessenbornensis, could have been enriched with a certain amount of dicotyledonous plants (Caloi, 2002). Simple enamel folds are observed in species feeding on hard vegetation, while a more complicated enamel pattern is found in species feeding on softer plants (Gromova, 1949; Eisenmann, 2010). The highly folded teeth of Equus suessenbornensis could be related to more humid conditions (Eisenmann, 2006a). Besides the size and shape of specific skeletal elements, the fossils of Pleistocene equids have also yielded other information on their diets and habitat use. In particular, studies of tooth wear characteristics and isotopic analyses of teeth have given insight into horse behavior. Mesowear describes the tooth wear patterns that result from an individual animal's diet over a large part of its lifespan (Fortelius and Solounias, 2000; Rivals et al., 2007). Tooth microwear reflects the wear characteristics of the foodstuffs the animal consumed in the days before it died (Walker et al., 1978). The microwear of grazers is characterized by high numbers of scratches and low numbers of pits (Solounias and Semprebon, 2002). Feeding habits reconstructions based on mesowear analysis indicate a high level of abrasion in Equus altidens, as demonstrated in the Vallparadis section (level EVT12 and EVT7, respectively, dated to 1.0 Ma and 0.86 Ma) (Strani et al., 2019) and the MIS 17 localities of Pakefield and West Runton in England (Rivals and Lister, 2016). However, microwear scores seem to nuance the strictly “grazer” character of Equus altidens, classified as either grazers or seasonal mixed feeders (Valli et al., 2012; Rivals and Lister, 2016; Strani et al., 2019). Such data are, to our knowledge, not available for E. suessenbornensis.
The Case of Equus apolloniensis
Equus apolloniensis (Koufos et al., 1997), the equid of Apollonia 1 in Greek Macedonia (Platanochori Formation, Mygdonia Basin) dated to about 1.2–0.9 Ma (Koufos and Kostopoulos, 2016), constitutes a different species with many affinities to modern wild asses (Eisenmann and Kuznetsova, 2004; Eisenmann and Boulbes, in preparation). The species is characterized by its medium to large size, a cranium with a short and deep muzzle, short palate, elongated choanae, shallow narial notch, far posteriorly situated orbit, elongated supra-occipital crest; teeth with elongated protocones, deep postprotoconal groove (Table 1; Figure 3F), rounded double knot and shallow ectoflexid on lower molars; and elongated, and not robust metapodials with deeper diaphysis as E. altidens/E. granatensis but “flattened” proximal end as E. stenonis (Table 1); protrusion of the distal is intermediate (Table 1). According to Koufos et al. (1997) the species represents a transitional form from the typical E. stenonis to the Middle Pleistocene horse Equus suessenbomensis. However, the similarities with the current wild donkeys (Equus africanus) are very striking in the mandible, morphology of the teeth (central and bilobed protocone, deep postprotoconal groove, shallow vestibular groove on lower molars), general proportions of the limb bones, and the proportions of the metapodials (Eisenmann and Boulbes, in preparation). The geographical distribution of the species appears to be limited to the eastern Mediterranean, as evidenced by its presence in other localities of the Upper Villafranchian in Greece (Koufos et al., 1997) as in Alikes (1.6 Ma, Kahlke et al., 2011), may be in Tsiotra Vryssi and more certainly in the site of Platanochori-1 (both from Mygdonia basin) similar in age than Apollonia P-1 (Konidaris et al., 2015). Its presence is also suspected in Turkey in the upper travertines of the Denizli region in southwestern Anatolia dated between 1.6 and 1.1 Ma (Boulbes et al., 2014; Lebatard et al., 2014). The species is however evoked at the transition of the Lower to the Middle Pleistocene in Spain in the region of Valencia (Cova del Llentiscle de Vilamarxant, Sarrión Montañana, 2008).
Caballoid or True Horses
There is consensus that the lineage of true or caballoid horses emerged in Europe at the beginning of the Middle Pleistocene (Forsten, 1998b), gradually becoming more numerous and replacing the various stenonid species.
Their extensive skeletal plasticity enabled them to survive under a wide range of climatic conditions and environments, from interglacial forests to grass-steppes during glacial episodes (van Asperen, 2010; Saarinen et al., 2016). Their ubiquitous and often abundant presence in iconic Middle Pleistocene palaeontological and archaeological sites have led to an intensive research focus, with researchers such as Von Reichenau (1915), Gromova (1949), Prat (1968), Nobis (1971), (e.g., Eisenmann et al., 1985; Eisenmann, 1988, 1991a,b; Eisenmann and David, 2002) (e.g., Forsten, 1973, 1999b; Forsten and Moigne, 1998) (e.g., Musil, 1975, 1977, 1978, 1984, 1990, 1991) (Guadelli, 1987, 1991), and Kuzmina (1997) publishing extensively on these fossils in the twentieth century, developing methods that are still in use (especially V. Eisenmann's series of measurements; Eisenmann, 1979, 1980, 1981; Eisenmann et al., 1988; Dive and Eisenmann, 1991). Initially, this research focused primarily on describing the fossils. Later work developed a focus on biostratigraphy (e.g., Eisenmann, 1988, 1991a,b; Forsten, 1998b), which has continued into the twenty-first century (e.g., Cramer, 2002; Boulbes, 2010; van Asperen, 2011, 2012, 2013a; Uzunidis, 2017). Such studies identified patterns of variation in the Middle Pleistocene Equus lineage which were subsequently interpreted as ecomorphological adaptations (van Asperen, 2010; Saarinen et al., 2016). Since they were an important component of the large vertebrate fauna, Middle Pleistocene equids have also been included in broader studies of diet and habitat use (e.g., Bocherens et al., 1997; Rivals et al., 2008, 2009; García García et al., 2009; Feranec et al., 2010; Pushkina et al., 2014; Rivals and Lister, 2016; Uzunidis et al., 2017), resource partitioning and niche separation (Britton et al., 2012), and taphonomy (Rivals et al., 2015).
One Species or Many Species?
To a novice accessing the literature, one thing stands out immediately: the proliferation of equid species names during the Middle Pleistocene (Table 2). Middle Pleistocene caballoid equids exhibit homogeneous overall morphology compared to equids from earlier geological epochs.
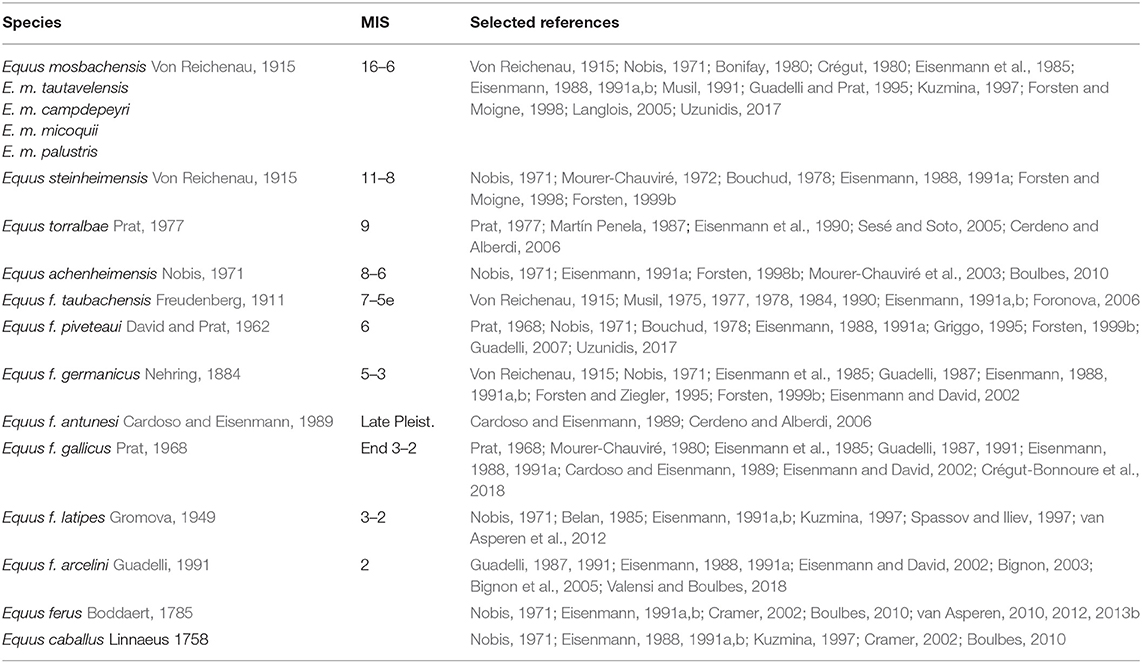
Table 2. Selected species names used in the literature for Middle and Late Pleistocene European caballoid equids (NB many variants on these names also occur but are not included here).
The first true horse in Europe, Equus mosbachensis (Von Reichenau, 1915) (or E. ferus mosbachensis), is perhaps the only one often regarded as a true species (Eisenmann, 1979, 1980; Bonifay, 1980; Crégut, 1980; Eisenmann et al., 1985; Guadelli and Prat, 1995; Langlois, 2005; Boulbes, 2010; Palombo, 2014; Palombo and Alberdi, 2017; Uzunidis et al., 2017). Equus mosbachensis presents some “archaic” morphological characters: presence of the tendon insertion of the anterior brachialis muscle on the inner edge of the diaphysis of the radius, strong supra-articular tuberosities on metapodials, a large size and slender general build. Other characters are specific, such as variation the of protoconal index (low and sub-equal between P4 and M1). Equus mosbachensis is clearly identified in Mauer (Nobis, 1971), dated by combined ESR/U-series and IR analysis at 609 ± 40 ka and correlated to MIS 17/15 (Wagner et al., 2011), then in the eponymous site of Mosbach 2 which yielded a very rich assemblage. Also in Germany, some typically caballine and teeth discovered at Süssenborn (MIS 16, 0.64–0.62 Ma, Kahlke, 2014) could represent some of the oldest evidence (Forsten, 1986; Eisenmann, 2008). West Runton is the oldest site in England that contained material of the species (Lister et al., 2010), of pre-Elsterian age, and is correlated with an MIS 17 fauna (Preece and Parfitt, 2008). Other regions of southern and western Europe also show the presence of this horse at the beginning of the Middle Pleistocene: in Spain in the Middle Pleistocene levels of the Vallparadís section (EVT3) of a slightly more recent age than 0.6 Ma based on the small mammal fauna (Aurell-Garrido et al., 2010; Madurell-Malapeira et al., 2010; Martínez et al., 2014) or in Sierra de Atapuerca (Galeria TG8, Gran Dolina TD10, Van der Made, 2013). In Italy the first true horses are recognized in the Isernia faunal unit (Palombo, 2014) at Notarchirico (about 0.6 Ma; Pereira et al., 2015), and perhaps earlier with the “caballine equid” from Cesi (Brunhes lower to 0.7 Ma, Ficcarelli et al., 1997; Palombo, 2014). There is no chronological consensus on the disappearance of the E. mosbachensis group or even on the taxonomy of the different forms or subspecies ascribed to it (E. m. tautavelensis Crégut, 1980, Caune de l'Arago; E. m. campdepeyrii Guadelli and Prat, 1995, Camp-de-Peyre; E. m. palustris Bonifay, 1980, Lunel-Viel; E. m. micoquii Langlois, 2005, Micoque) because of the numerous regional and chrono/ecotype variations. Equus mosbachensis could extend to the end of Middle Pleistocene during MIS 6 (Guadelli, 2007; Uzunidis, 2017), but populations similar to the type-series are generally Cromerian in age, or up to MIS 9 as in Schöningen (Channel II) for example (Van Kolfschoten et al., 2015; Figure 1).
This is not unexpected, since interspecific variation between extant species of equids is relatively low in comparison with other large mammal lineages, making it difficult to distinguish between modern equid species based on skeletal morphology (Eisenmann, 1979, 1986; Eisenmann and Beckouche, 1986; Dive and Eisenmann, 1991; Eisenmann and David, 2002). However, clear patterns of variability in terms of dental and skeletal proportions can be identified in Middle Pleistocene caballoid equid fossils. This variability has been interpreted as reflecting an evolutionary lineage developing through various (chrono)species (Eisenmann and Kuznetsova, 2004). Whether these can be regarded as species or subspecies, chronospecies or ecomorphs depends largely on an assessment of the extent of this variation between fossil assemblages as compared to intra- and interspecific variation in modern equids (van Asperen, 2013b).
Sexual dimorphism, both in size and in shape, and age-related size and shape variation in adult skeletal elements is insignificant in modern equids (Winans, 1989; van Asperen, 2013b). Whilst male individuals are on average slightly larger than females, body size considerably overlaps between the sexes (Kuzmina, 1997; van Asperen, 2013b). This implies that any size and shape differences that can be observed between fossil assemblages reflect real population differences. However, whilst such size and shape differences can certainly be observed, taken as a whole, the Middle Pleistocene European horse lineage, covering a time span of about 300,000 years, is not more variable than modern ponies or such highly homogeneous groups as Arabian horses or Equus przewalskii (van Asperen, 2013b). Furthermore, there is no unidirectional, evolutionary trend in size and shape. Instead, morphology fluctuates over time (Forsten, 1993; Eisenmann and David, 2002; Eisenmann, 2004b; van Asperen, 2010). In addition to observations on skeletal morphology, studies of equid genetic variation also indicate that Middle Pleistocene equid taxonomy shows a degree of oversplitting (Weinstock et al., 2005; Orlando et al., 2009).
Taking these considerations into account, here we follow the guidelines of the International Commission on Zoological Nomenclature (ICZN, 2003; see also Cramer, 2002; van Asperen, 2013b) and refer to all European late Middle Pleistocene and Upper Pleistocene caballoid equids as Equus ferus Boddaert, 1785, although this taxon, commonly chosen for Pleistocene wild horses, remains questionable (Guadelli and Delpech, 2000; Eisenmann, 2006b). The variation observed between assemblages follows certain patterns that can be interpreted as ecomorphological variation, and the resulting groups are best referred to as ecomorphs, or, at most, subspecies of E. ferus.
Ecomorphological Patterns
The European Middle Pleistocene was characterized by high-amplitude glacial–interglacial cycles, producing an alternation of a wide range of environments ranging from glacial deserts to continental steppe-like environments and temperate forests. Remains of caballoid horses have been found in sediments deposited in virtually all these environments, often in abundance and with good preservation. Although, as we have argued above, all these remains can be assigned to a single species, the variation in temperature, humidity, vegetation openness, and the character of the substrate exerted fluctuating selective pressures on certain skeletal characteristics. Adaptive variation produced several ecomorphs whose morphology reflects the environmental conditions (van Asperen, 2010). The cyclical nature of the climatic changes thus led to cyclical fluctuations in caballoid morphology (Forsten, 1993; Eisenmann and David, 2002; Eisenmann, 2004b; van Asperen, 2010). The influence of these environmental conditions on horse morphology was mediated through diet, thermoregulation and locomotion.
Muzzle
The main differences in cranial proportions of caballine Pleistocene Equids concern the variation in the proportions of the muzzle (Eisenmann, 2014). Despite attempts to detect an evolutionary pattern, the proportions of the muzzle always find an explanation in local climatic conditions. However, whether these differences in proportions result from a micro-adaptive phenomenon or phenotypic plasticity as acclimatization remains unclear. A short and broad muzzle could represent an adaption to a cold climate (following Allen's rule) and grazer specialization. Examples of cold adapted caballoid equids with a short and broad muzzle are found in Middle Pleistocene glacial deposits (Bellai, 1998; Boulbes, 2014; Caune de l'Arago, MIS 14–12, Eisenmann et al., 1985; Lumley et al., 2015) or from the Last Glacial Maximum at the end of the Upper Pleistocene (MIS 2) (Coulet des Roches, E. ferus gallicus, Crégut-Bonnoure et al., 2018). On the other hand, more elongated muzzles often associated with narrower proportions come from temperate climatic conditions as in the case of the interglacial equid from Lunel-Viel (Bonifay, 1980; Eisenmann et al., 1985) or the Late Pleistocene horse from Portugal E. ferus antunesi (Cardoso and Eisenmann, 1989). Intermediate morphs could also reflect interstadial conditions. Reconstruction based on the proportions of the symphysis leads to the same results (Caloi, 2002; Bignon, 2003).
Teeth
Size and Shape
Tooth size and shape is influenced primarily by diet. Equids are generalized grazers (Salter and Hudson, 1979; Guthrie, 1984; Feranec, 2007a; see below). Several features of the dentition of Middle Pleistocene caballoids have been connected with specific dietary pressures. A larger occlusal surface enables the animal to process more food (MacFadden, 1988). Although horses can survive on a low-quality diet, they need a high and relatively diverse food intake (Salter and Hudson, 1979; Duncan et al., 1990). Hindgut fermenters such as horses show an increase in length of the premolar row and a widening of the teeth with an increase in the amount of browse in the diet (Janis, 1990).
In general, tooth size shows relatively little variation over the Middle Pleistocene. And whilst qualitative characteristics of the tooth occlusal surfaces can vary greatly within assemblages, variation between assemblages is insignificant. This may be due to the fact that the dentition is generally less phenotypically plastic and thus reacts more slowly to environmental factors than e.g., limb bones (Hillson, 2005). However, differences in size are sometimes recorded: glacial horses, such as for example those of Arago cave (MIS 14 to MIS 12), exhibit very large teeth (“macrodont”), relative to the size of the postcranial elements (Boulbes, 2014) (Figure 6). An explanation could be that this horse lived under glacial climatic conditions in a steppic environment with low-quality foodstuffs, necessitating a high food intake. A wider occlusal surface is also an advantage when consuming more abrasive grasses (Gromova, 1949; Guadelli, 1987; Eisenmann, 1991b; Delpech et al., 2000). In the case of the Arago horse, a high level of abrasive food is indeed inferred from mesowear and microwear analysis (Kaiser and Franz-Odendaal, 2004; Rivals et al., 2008). Teeth from British sites tend to be somewhat larger than those from continental sites, being relatively wide with relatively long premolar rows (van Asperen, 2010). This suggests a higher food intake and/or a higher proportion of browse in the diet, which may be due to the more oceanic climate, and thus denser and to horses less hospitable interglacial forests, of the British Isles compared to most of continental Europe.

Figure 6. Equus mosbachensis tautavelensis from lower level Q (CMI) of Arago cave. (A) Right maxillary with P4-M3 (G13-GEQ1-1290); (B) Right mandibular with P2-M3 (I8-IZPQ3-2160). Occlusal length of upper P3 (A) is about 36,8 mm.
Protocone length
Eisenmann (1991b) grouped caballoid horses into three types based on the length of protocone of their upper teeth. Longer protocones are more resistant to wear and thus better adapted to abrasive food sources (MacFadden, 1988). Although the size differences are not statistically significant, in general the pattern holds for Middle Pleistocene caballoids, but there are many exceptions (van Asperen, 2009a,b). Horses of Type I have shorter protocones on the upper third and fourth premolars (P3/4) than on the upper first and second molars (M1/2), and are generally found in temperate climates. Kuzmina (1997) correlates long protocones on the upper molars with forest-steppe or forest conditions. Type I is typical for such temperate assemblages as British MIS 9 and 7 material and horse remains from Achenheim, Biache-Saint-Vaast, Weimar-Ehringsdorf and Taubach (Nobis, 1971; Auguste, 1995; van Asperen, 2009a,b), but also characterizes the horse from Abri Suard (layers 5-6, MIS 6) (Prat, 1968; Eisenmann, 1991b). The majority of Upper Pleistocene horses is included in this group (Guadelli, 1987; Eisenmann, 1991b; Fourvel et al., 2014).
Horses of Type II show the opposite pattern, with relatively long protocones on the P3/4 and shorter protocones on the M1/2, and are often correlated with cold climatic conditions. The cold MIS 6 assemblage from Wannen belongs to Type II, but the type seems to be rare. LGM horses from MIS 2 (Magdalenian) are also included in this group (Eisenmann, 1991b).
Finally, horses of Type III, associated with cool to cold environments, have short protocones on both P3/4 and M1/2. The group of Type III horses includes assemblages from temperate sites such as Steinheim, Orgnac 3, Biache-Saint-Vaast, and British MIS 11 sites, as well as sites dating from colder periods, such as the MIS 6 sites of Schweinskopf and Romain-la-Roche (van Asperen, 2009a; Boulbes, 2010).
Some samples do not fit any Type, with long protocones on both the premolars and the molars (e.g., British MIS 6 and Schöningen 13II-4 samples; van Asperen, 2009a). Uzunidis et al. (2017) carried out a direct test of the relationship between protocone length and the abrasiveness of the diet (as reflected in tooth microwear, Solounias and Semprebon, 2002), and found no correlation. Admittedly, their study did not differentiate between premolars and molars, but together these results indicate the length of protocone should be used with caution to infer environmental conditions.
Body Size, Limb Element Proportions, and Robustness of the Limbs
Body size is a complex and highly plastic trait, which is influenced by a large number of factors. Thermoregulatory pressures lead to larger body size in colder (Bergmann's rule) or drier climates, whereas closely related animals in warmer or more humid climates tend to be smaller (Mayr, 1956; James, 1970; Lindstedt and Boyce, 1985; Blackburn et al., 1999). In colder climates, limbs tend to be shorter than in warmer climates (Allen's rule). Together, these body shape changes produce a reduced ratio of surface area to body mass and thus reduced heat loss in colder and drier climates. Furthermore, if a similar bodyweight is supported by shorter legs and muscles, the leg bones will be more robust, which provides further thermoregulatory advantage (Gregory, 1912; Thomason, 1986; Tilkens et al., 2007).
Animals of larger body size have enhanced starvation resistance (Lindstedt and Boyce, 1985; Blackburn et al., 1999). Such animals are more likely to survive seasons of scarcity and profit from increased resource availability after periods of high mortality, although short growing seasons may limit growth potential. In contrast, small body size may facilitate predator avoidance in densely vegetated environments (Bro-Jørgensen, 2008).
Middle Pleistocene
These factors combine to produce a complex pattern of body size and shape variation during the glacial-interglacial cycles of the Middle Pleistocene (Figure 7). Glacial horses are generally smaller and more robust than interglacial horses (Collinge, 2001; Cramer, 2002; Guthrie, 2003; van Asperen, 2010). This is particularly pronounced in British samples from low-productivity periglacial desert environments dating to MIS 6 (Green et al., 1984; Parfitt et al., 1998; Murton et al., 2001). On the other hand, horses from continental sites from MIS 6 (E. achenheimensis, Romain-la-Roche; E. f. piveteaui, Abri Suard 5-6) have a larger body size. The MIS 6 glaciation was one of the most extreme of the Middle Pleistocene (Shackleton, 1987; Reille et al., 2000; Ehlers et al., 2004; Laban and Van der Meer, 2004). In contrast, during the less extreme glacials of MIS 8 and 10, a relatively diverse steppe vegetation (De Beaulieu and Reille, 1995) allowed the horses to grow to larger body size (van Asperen, 2010) (Figure 7).
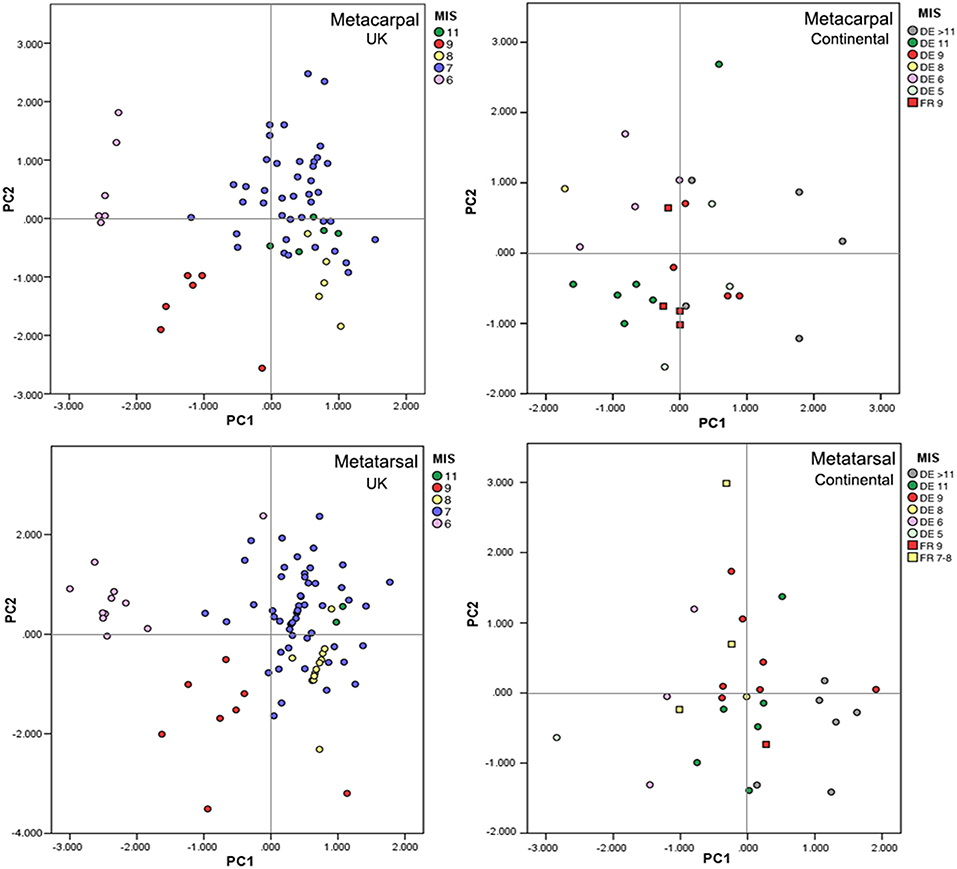
Figure 7. Principal component analysis of complete caballoid equid metacarpals and metatarsals from late Middle Pleistocene British sites and continental Europe; PC2 plotted against PC1 (modified from van Asperen, 2010, Copyright © 2010 Elsevier BV, with permission from Elsevier); PC1 generally relates to overall size, while PC2 tends to reflect robusticity. Supplementary data available in van Asperen (2010).
Within the interglacial assemblages, essentially two morphologies can be distinguished. The mosaic steppe vegetation of phases with a long growing season and a more continental climate presented the horses with an environment that they are extremely well-adapted to. As a consequence, they were able to grow to a large size, with relatively robust limbs, though less robust than their glacial counterparts (van Asperen, 2010). Such large-sized, robust horse remains are found in British sites dating from the later, relatively continental phases of MIS 7 (Schreve, 2001) and on continental MIS 9 and MIS 7 sites. The higher seasonality experienced by populations from continental regions led to somewhat larger body size than in the more oceanic climates of Britain (van Asperen, 2010) (Figure 7).
In contrast, the second group of interglacial horses has relatively small body size and slender limb bones. Assemblages with such characteristics are found in contexts with indications of densely vegetated habitats and oceanic conditions, representing a far less favorable environment for grazing-adapted species. These morphologies are found in British MIS 9 sites and some continental sites of uncertain date, such as “E. steinheimensis” from Steinheim (MIS 11? van Asperen, 2010, 2013a), and the horses from Orgnac (MIS 9–8, Forsten and Moigne, 1998; Moncel et al., 2005) and Lunel-Viel (MIS 11–10, Brugal and Boudadi-Maligne, 2011). The two laters are also characterized by robust diaphysis in relation to substrat and humidity gradient (see below). Small sized horses also present in Acheulean sites in the Iberian Peninsula (E. torralbae, Torralba, Prat, 1977; the horse from the site of cueva del Ángel, MIS 9–7, Falguères et al., 2019) could correspond to the same pattern and/or a latitudinal gradient.
British horse remains dating from MIS 11 and continental horse remains dating from MIS 5 have an intermediate morphology, being larger than the horses in the second group but somewhat more slender than the horses of the first group (van Asperen, 2010). It may be significant that horse remains are scarce in faunal assemblages dating from these phases, indicating that population sizes were small, so that these animals were able to grow to a relatively large size regardless of the probably densely forested habitats they inhabited.
Late Pleistocene
From the end of the Middle Pleistocene and during the Late Pleistocene, the caballoid horses underwent a general decrease in size, which also resulted in the creation of several chronological subspecies (E. f. taubachensis, E. f. germanicus, E. f. gallicus, E. f. arcelini, Prat, 1968; Eisenmann, 1991b; Guadelli, 1991). This trend is for example well-illustrated in the length of the metacarpal (Figure 8) and based on a method using the Variability Size Index V.S.I. (Cramer, 2002; Magniez et al., 2017). The decrease in body size during the Late Pleistocene constitutes a gradual process of adaptation to climatic change, but without speciation (Forsten, 1996). This could be attributed to adaptive pedomorphism or dwarfism (Gould, 1976; Guthrie, 1984; Forsten, 1988, 1996) to maintain population densities in an increasingly fragmented or constraining environment by increasing the rate of population growth (e.g., through the shortening of the gestation period) for the survival of the species Forsten, 1993). This could be an adaptive response to rapid changes in climate during this period (Magniez and Boulbes, 2014). Size reduction among late Pleistocene horses also provides a useful biochronological marker (Magniez and Boulbes, 2014). Even if there are exceptions: the horse of Portel-Ouest cave (levels F) is larger than those of the contemporary sites of MIS 3, perhaps resulting from a regional particularism.
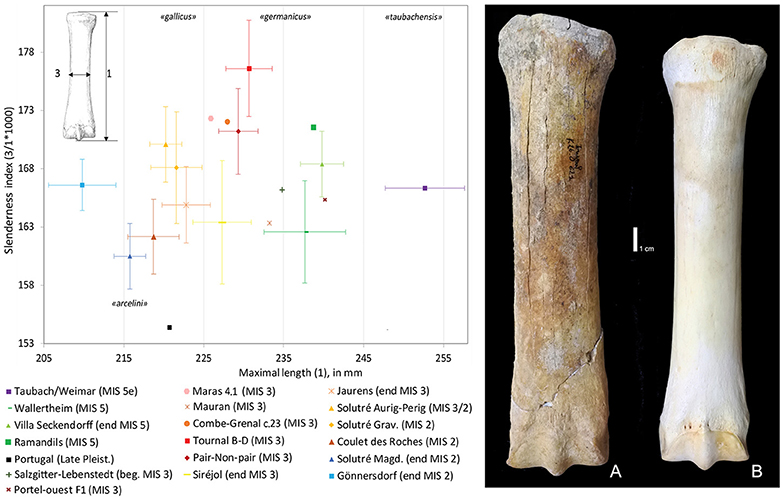
Figure 8. Bivariate diagram of slenderness index (SI 1) in relation to the maximal length of the metacarpal. Slenderness index (SI 1): breadth at the middle of the diaphysis (3)/maximal length (1)*1,000. Confidence interval scale bars (95%) for series with more than 5 specimens. Data: Taubach/Weimar (Eisenmann, pers. comm.); Wallertheim (Nobis, 1971); Villa Seckendorff, Salzgitter-Lebenstedt, Solutré Gravettian, Gönnersdorf (Cramer, 2002); Combe-Grenal, Solutré Aurignacian-Perigordian (Guadelli, 1991); Mauran (Eisenmann and David, 1994); Pair-non-Pair (Prat, 1968); Siréjol, Jaurens (Eisenmann, 2019); Ramandils, Portel-Ouest, Abri du Maras, Tournal B/D, Coulet des Roches (Boulbes, pers. data); Portugal (Cardoso and Eisenmann, 1989). Metacarpals of Late Pleistocene horses. (A) Equus ferus germanicus from Tournal (K26-0-621, level B, MIS 3); (B) Equus ferus gallicus from Coulet des Roches (M4-814, MIS 2).
In addition to this decrease in body size, it is also possible to detect variations in certain skeletal proportions. As during the Middle Pleistocene, the robustness of the diaphysis of the metapodials shows the most important pattern (Figure 8). Following the distribution of the currently recognized species, the robustness of the diaphysis of the metapodials correlates well with moisture levels (Eisenmann, 1984). For example, the metacarpal from the Mousterian layers of Tournal B-D and Abri du Maras (level 4.1) (E. f. germanicus, South of France, MIS 3) have a relatively robust diaphysis, which reflects the high humidity recorded in these levels (Echchat, 2008; Magniez and Boulbes, 2014; Daujeard et al., 2019) (Figure 8A). On the other hand, the metacarpal of the horse from Coulet des Roches (South France, MIS 2) is slender in relation to the hard substrate frequented as demonstrated by other proxies (Crégut-Bonnoure et al., 2018) (Figure 8B). The Late Pleistocene horse from Portugal, E. ferus antunesi, even more gracile, was probably adapted to rather dry climate and to a hard ground (Cardoso and Eisenmann, 1989) (Figure 8). These rapid and reversible variations in the proportions of the diaphysis seem, in this case, to be related to phenotypic plasticity.
Another pattern concerns the variation in the general proportions of the limb and in particular the relationship between the lengths of metacarpal and metarsal (Figure 9). In the lineage of the Upper Pleistocene horses, the length of the metacarpal decreased faster than that of the metatarsal. These differences could reflect an evolution pattern and/or an adaptation to the openess of the environment and the density of the vegetation. Indeed, Eemian (MIS 5e) horses have caricatural proportions with a very long metacarpal compared to the metatarsal; horses from MIS 3 to a lesser degree have a long metacarpal and are often associated with mozaic landscapes; forms from MIS 2, correlated with open landscapes, have a longer metatarsal (Figure 10). Such differences have also been noticed in extant species, the metatarsals are especially short in two species of zebras, Equus burchelli and Equus zebra (Eisenmann, 1979; Groves and Willoughby, 1981), which occasionally frequent wooded environments (Eisenmann, 1984).
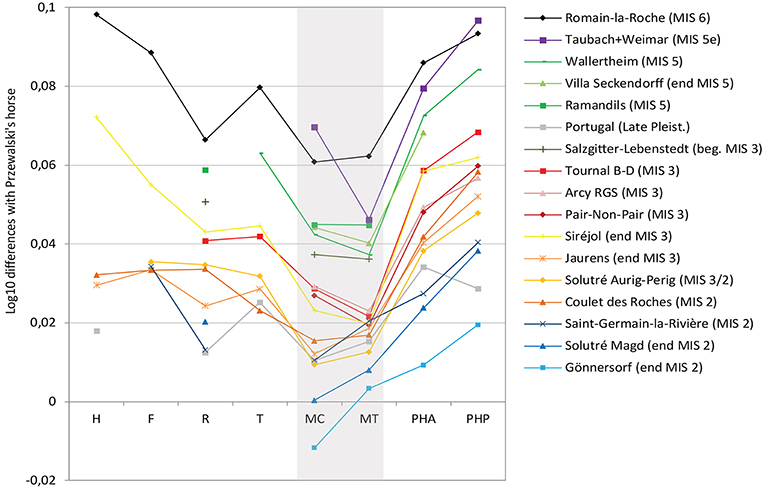
Figure 9. Simpson's ratio diagram of bone lengths of European Late Pleistocene true horses. Reference: Przewalski's horse (Eisenmann, 1991a). H, Humerus; F, Femur; R, Radius; T, Tibia; MC, Metacarpal; MT, Metatarsal; PH1A, First Anterior Phalanx; PH1P, First Posterior Phalanx. Data: Romain-la-Roche (Boulbes, 2010); Taubach-Weimar (Eisenmann, pers. com.); Wallertheim, Villa Seckendorff, Gönnersdorf (Cramer, 2002); Ramandils, Tournal B-D (Boulbes, pers. data); Siréjol (Eisenmann, 2019); Jaurens (Eisenmann, 1991a); Solutré (Guadelli, 1991); Coulet-des-Roches (Crégut-Bonnoure et al., 2018); Saint-Germain-la-Rivière (Prat, 1968).
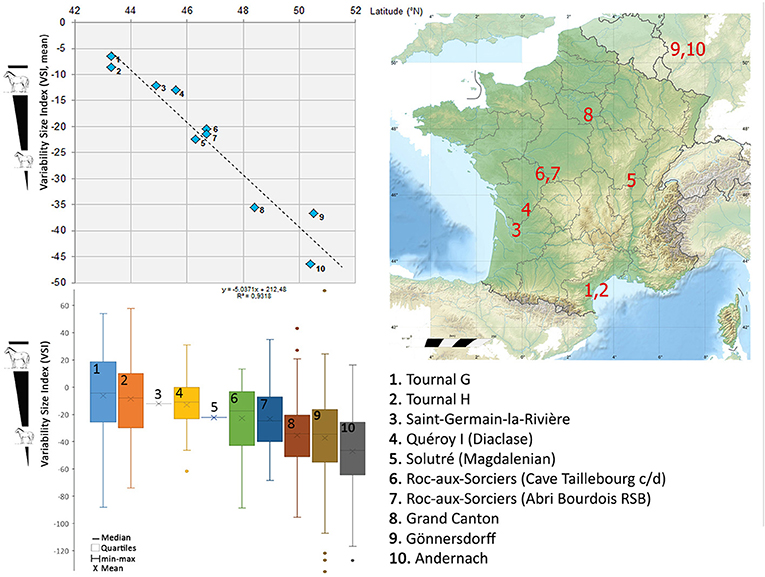
Figure 10. Variability Size Index (VSI, cf. Meadow, 1999) of horse populations from the end of MIS 2 (Magdalenian). Standard: Jaurens (Mourer-Chauviré, 1980; Eisenmann, 2019). Gönnersdorf, Andernach (Cramer, 2002); Tournal (Boulbes, pers. data); Roc-aux-Sorciers (Boulbes and Valensi, unpublished data) Saint-Germain-la-Rivière (Prat, 1968); Quéroy I, Grand Canton (Véra Eisenmann, pers. com.); Solutré Magdalenian (Guadelli, 1991).
Magdalenian cline
The end of the Pleistocene offers a fine chronological resolution to detect size variation with numerous well-dated sites. The size index (V.S.I., see Meadow, 1999) in relation to latitude (Figure 10, Table S1) shows a progressive and statistically significant decrease in the body size of Magdalenian horses from southern France to northern Europe. The pattern of this geographical cline is clearly visible: series coming from the same latitude have an equivalent size. Mediterranean horses are a little larger than those from southwestern France, which are themselves more robust than those from central France; horses from the Paris Basin and the Rhineland in Germany are smaller.
These regional differences can be compared with the 15N nitrogen gradient and could be explained by an earlier onset of climatic improvement in the south compared to the northern and/or continental regions after the Last Glacial Maximum (Drucker et al., 2003, 2012; Stevens and Hedges, 2004; Stevens et al., 2009). Access to more resources in more southerly regions would have favored maintaining an equal size in the south of France. These results reflect the regional mosaic of the Late Glacial horse populations (Kaagan, 2000; Cramer, 2002; Bignon et al., 2005; Bignon and Eisenmann, 2006; Magniez et al., 2017).
Breadth of the Third Phalanx
The third phalanx is the skeletal element that interacts most closely with the substrate in locomotion. In comparison with other equids, caballoid horses have relatively broad third phalanges. Broad third phalanges are thought to be an adaptation to soft substrates, providing a larger contact area and broader distribution of weight (Eisenmann, 1984; Forsten, 1988; Kuzmina, 1997; Foronova, 2006). Although this characteristic has been analyzed for Middle Pleistocene equids (van Asperen, 2010), this interspecific pattern does not seem to be reflected in variation at an intraspecific level.
Study of Late Glacial (LG) horses (Bignon and Eisenmann, 2006) demonstrates the homogeneity in proportions of the third phalanges at a local scale between different areas, according to the authors p. 169 “related to the global climatic context and the local ecology, namely the food exploitation by horses of river banks and marshes vegetation.” New data coming from LGM and LG levels of the site of Coulet des Roches identify slightly narrower proportions than contemporaneous populations, showing that horses at this site frequented less heavy or harder ground habitats (Crégut-Bonnoure et al., 2018) as in the case of the horse from Portugal, E. f. antunesi (Cardoso and Eisenmann, 1989). In contrast, the Late Pleistocene horse E. f. latipes (Gromova, 1949) from the Paleolithic sites of Kostenki, Russian Valley, Don River (Kuzmina, 1997) and Mezin, Desna basin, Ukraine (Belan, 1985) show extremely wide third phalanges.
Diet and Habitat
Regardless of climatic conditions, Middle Pleistocene horse mesowear has the characteristics of a grazer (e.g., Rivals et al., 2008; Rivals and Lister, 2016; Uzunidis et al., 2017). In grazers, the abrasive nature of the foodstuffs consumed, as well as the relatively high amount of grit found on plants growing at ground level, produce an abrasion-dominated wear pattern with round or blunt cusps with low relief (Williams and Kay, 2001). Middle Pleistocene horses often have some of the highest mesowear scores, and therefore the most abrasive diets, among the species represented in the faunal assemblages (e.g., Rivals et al., 2008). However, occasionally horse populations show variation in mesowear, ranging from grazer to mixed feeder with a significant amount of browse in the diet (e.g., Rivals et al., 2009, 2015). Interglacial horse assemblages are often characterized by a higher amount of variation in diet than glacial assemblages (Rivals et al., 2009).
In contrast to their generally grazer-like mesowear signature, Middle Pleistocene horse microwear is more variable, ranging over the entire spectrum from grazer to browser (e.g., Rivals et al., 2009, 2015; Rivals and Lister, 2016). The same assemblage can have a grazer-like mesowear pattern but a browser-like microwear pattern, possibly due to seasonal variation in diet (Uzunidis et al., 2017). One problem with a browsing microwear signature is that it is not possible to distinguish between browsers eating low herbaceous browse (forbs) or trees and shrubs (Rivals and Lister, 2016). Przewalski horses, for example, consume large amounts of dicotyledons (Schulz and Kaiser, 2013).
Animals that feed on different kinds of foodstuffs tend to have different ratios of carbon and nitrogen isotopic values in their tooth and bone collagen. Carbon isotope values relate to the different photosynthetic pathways used by plants, known as the C3, C4, and CAM pathways. The European Middle Pleistocene is strongly dominated by C3 plants. However, within C3 plants, there is further fractionation between woody and herbaceous plants depending on the openness of the landscape, known as the canopy effect (Heaton, 1999; Drucker et al., 2008). As a result, differences in feeding behavior can lead to different δ13C values in herbivores (Feranec, 2007b). Nitrogen isotopic values vary primarily with the trophic level at which the species feeds (Bocherens and Drucker, 2003).
Mean δ13C values for Middle Pleistocene horses tend to reflect grazing in open forest or grassland (e.g., García García et al., 2009; Feranec et al., 2010; Pushkina et al., 2014). However, δ13C values reflecting foraging in more closed habitats also occur, mainly in temperate assemblages such as those from Steinheim (MIS 11?) and Neumark-Nord (MIS 5e; e.g., Britton et al., 2012; Pushkina et al., 2014). Pleistocene equids tend to have slightly lower δ13C values than bovids, which may be due to differences in digestive physiology (Britton et al., 2012). There is also variation in horse δ15N values, to the degree that at some sites, horses appear at a higher trophic level than bovids, their closest competitors for food resources (ibid.). This could be due to the horses consistently selecting lower-quality forage and the way in which their non-ruminant digestive system processes this forage. However, it is more likely that equids and bovids flexibly adapted their feeding strategies to resource availability and the density of herbivores feeding on the same plants (Menard et al., 2002; Britton et al., 2012).
European Wild Ass Equus hydruntinus
Equus hydruntinus, also known as the European wild ass, was first described by Regalia (1907) at the start of the twentieth century from the Upper Paleolithic cave site of Romanelli (Italy), to be precise, in the Upper Complex (≪ Terre brune ≫) attributed to the Epigravettian (Sardella et al., 2018). An exhaustive study of its anatomy and its geographical distribution was made by Stehlin and Graziosi (1935). Many authors have designated Equus altidens (or Equus granatensis Eisenmann, 1999) as the probable ancestor of Equus hydruntinus (Musil, 1969; Forsten, 1986, 1990, 1999a; Azzaroli, 1992; Eisenmann, 1992; Alberdi et al., 1998; Alberdi and Palombo, 2013). This scenario is based in particular on assemblages of intermediate size and chronology (Petralona, Cullar de Baza-1, Venosa) (Van der Made et al., 2017). Affinities with E. tabeti (Aïn Hanech) and E. cf. tabeti (Oubeidiyeh) (Guerrero-Alba and Palmqvist, 1997; Eisenmann, 1999) have also been identified. In contrast, current thinking favors a new arrival from Asia over a local evolution in Europe (Eisenmann, 1992; Palombo and Alberdi, 2017). According to this model, Equus hydruntinus would have entered to occupy the ecological niche left vacant by Equus altidens (Palombo and Alberdi, 2017). At the maximum extent of its geographical expansion in the Late Pleistocene, the distribution of Equus hydruntinus extended from Europe to the south-west of Asia (Middle East, Caucasus) and probably into North Africa (Uerpmann, 1987; Wilms, 1989; Orlando et al., 2006). During the Holocene, the contraction of the steppe, the significant fragmentation of its geographical distribution and distinct subpopulations, and human exploitation progressively caused its disappearance between the Neolithic and the Iron Age (Spassov and Iliev, 2002; Crees and Turvey, 2014), more precisely, according to palaeogenetic information, during the Bronze Age (Bennett et al., 2017). As this hypothesis is subject to certain limitations (cf. Nores et al., 2015), it cannot be excluded that the enigmatic ≪ zebro ≫ mentioned in numerous Medieval manuscripts from the Iberian Peninsula corresponds to Equus hydruntinus (Antunes, 2006).
Systematics and Phylogeny
It was not until the start of the twenty-first century that the enigma of the phylogeny of Equus hydruntinus was significantly resolved through the contribution of studies of ancient DNA. Hitherto, the systematic position of this equid had long been disputed. The species was variously included in the subgenus ≪ Asinus ≫ (Stehlin and Graziosi, 1935; Gromova, 1949), or ≪ Hemionus ≫ (Azzaroli, 1992), placed close to the zebras (Davis, 1980; Bonifay, 1991), seen as the last representative of the Plio-Pleistocene group of the ≪ stenonids ≫ (Forsten, 1986, 1990, 1999a; Forsten and Ziegler, 1995; Alberdi and Palombo, 2013), or even considered as belonging to a separate ≪ Hydruntinus ≫ genus (Radulesco and Samson, 1965). These multiple hypotheses regarding its affiliation are related to the mosaic of characteristics found in the skeleton and dentition of this species: gracility and cursorial proportions of the limbs as in the hemiones, very short protocones as in the ≪ stenonids≫, a deep ectoflexid as in the zebras (Eisenmann and Mashkour, 1999). It was not until the start of the twenty-first century, with the significant study of the morphology of two crania from the site of Kabazi II (Crimea), that the proximity of Equus hydruntinus to the hemiones became clearer (Burke et al., 2003). The rise of palaeogenetics allowed this affiliation to be confirmed by many mitochondrial sequences (Orlando et al., 2006, 2009). The last palaeogenetic studies to date do not definitively support the classification of E. hydruntinus as a distinct species and places it among the Asiatic wild asses (Bennett et al., 2017).
From a morphological and metrical point of view, many characteristics indeed distinguish between Equus hydruntinus and Equus hemionus. The principal differences can be found in the cranium and teeth. The crania from the site of Kabazi II, and above all the complete cranium from the Emine-Bair-Khosar cave, reveal the main differences with E. hemionus: the length of the palate is greater, the muzzle is proportionally very short and relatively wide, the naso-incisival incision is clearly much shorter (Burke et al., 2003; Orlando et al., 2006; Eisenmann et al., 2008; van Asperen et al., 2012) (Figure 11). The species is often referred to as microdont, meaning that the teeth are relatively small in proportion to the postcranial skeleton, but it is also characterized by a high degree of hypsodonty (Stehlin and Graziosi, 1935; Eisenmann and Baryshnikov, 1995; Eisenmann et al., 2008). The mesio-distal length of the M3 is always shorter than that of the M2 (Bonifay, 1991; Boulbes, 2009). The variation in the protoconal index within the tooth row is also different from that of hemiones, with notably an IP which increases from the P4 to the M1 (Eisenmann and Patou, 1980; Cardoso, 1995; Boulbes, 2009). The relative proportions of the limb bones are close to those of hemiones, however, the MC III is proportionally shorter than the MT III (Eisenmann et al., 2008). The frequency of the presence of the small facet for the trapezoid varies between different species of hemiones, but it is more often present than in Equus hydruntinus. The metacarpals and metatarsals of E. hydruntinus have somewhat more robust diaphyses (which are also thicker) (Eisenmann et al., 2008; van Asperen et al., 2012). On the other hand, relatively to the length of the bone (variable 1), the epiphyses (5 to 14) are less developed (Figure 12). Finally, the distal keel (12) shows proportionally less protrusion. Other morphological criteria for the different skeletal elements show the ≪ primitive ≫ character state (Bonifay, 1966, 1991; Prat, 1968; Boulbes and Rillardon, 2010).
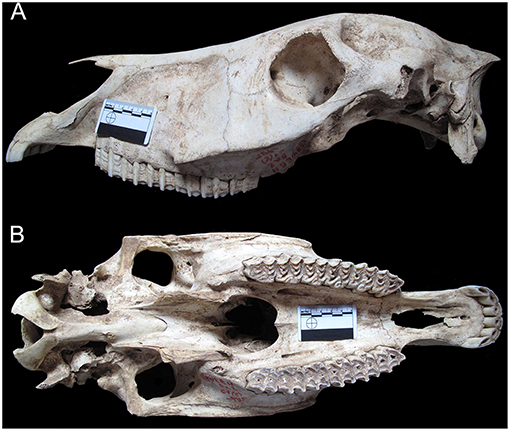
Figure 11. Skull of Equus hydruntinus from the site Emine-Bair-Khosar, Crimea; catalog no. Ba2 384 (photo E. van Asperen; scale in cm; reprinted from van Asperen et al., 2012 © Palaeontological Association February 2012). (A) Lateral view; (B) Occlusal view.
Figure 12. Simpson's ratio diagram of metacarpals from subspecies of extant Hemiones and Equus hydruntinus. Reference: Equus africanus (Eisenmann, 2019). Measurement system (Eisenmann, 2019): 1-Maximum length, 3-Breadth at the middle of the diaphysis, 4-Depth of the diaphysis at the same level, 5-Proximal articular breadth, 6-Proximal articular depth, 7-Diameter of the facet for capitatum, 8-Diameter of the facet for hamatum, 10-Distal supra-articular breadth, 11-Distal articular breadth, 12-Depth of the sagittal crest, 13-Minimum depth of the medial condyle, 14-Maximum depth of the medial condyle. Data: Hemiones, E. hydruntinus from Roterberg and Romanelli (Eisenmann, 2019); E. hydruntinus minor from Lunel-Viel, E. hydruntinus davidi from Arquet (Boulbes, pers. data); Equus hydruntinus from Emine-Bair-Khosar (van Asperen et al., 2012).
Biochronology
The appearance of Equus hydruntinus in the European fossil record is not synchronous across all geographical provinces, which can be partly explained by its rareness.
It seems that the oldest remains of this species in Europe can be found in Spain, in the Middle Pleistocene levels of the section of Vallparadís (E. cf. hydruntinus, level 11, EVT3), which is dated to a little later than 0.6 Ma on the basis of the small mammal fauna (Aurell-Garrido et al., 2010; Madurell-Malapeira et al., 2010; Martínez et al., 2014). In the complex of the Sierra de Atapuerca, Equus cf. hydruntinus was identified at Gran Dolina from level TD 10, correlated with MIS 11-9 (Saladié et al., 2018), in particular from the top of this level (TD 10.2 et TD 10.1), and at Galeria in levels GIIb and GIIIa (Rodríguez et al., 2011; Van der Made, 2013).
In Italy, the first appearance of Equus hydruntinus forms one of the chronological markers of the Middle Aurelian faunal assemblages (Vitinia FU, MIS 8.5 and 7) (Marra et al., 2014, 2018). In France, de caves of Lunel-Viel have long been considered to harbor the oldest remains of the species (Bonifay, 1991). Other fossils from this country could be even older, in particular those from the loess anciens inférieurs at Achenheim (Forsten, 1996, ex E. wernerti ≫) and an isolated tooth from the site of Orgnac 3 (Aouraghe, 1992). Equus hydruntinus is absent from certain very rich fossiliferous sequences, such as those from the Middle Complex of La Caune de l'Arago (MIS 14 to 12) (Moigne et al., 2006). The species is mentioned with caution for the Somme basin in the upper unit (Marne Blanche) of the quarry Carpentier d'Abbeville, recently dated and correlated with MIS 15 (Antoine et al., 2016).
In Germany, Equus hydruntinus is present at Schöningen 13II-4, correlated with the Reinsdorf Interglacial (MIS 9) (Van Kolfschoten et al., 2015) and Steinheim (Forsten, 1999b), the chronological position of which is subject to debate (MIS 11?). In the British Isles, the first appearance of the species is reported during the course of MIS 11 (Pettitt and White, 2012), especially from a number of isolated teeth from the site of Swanscombe (Hoxnian = Holsteinian) (Schreve, 1997; Bridgland and Schreve, 2004).
In the light of these finds and the review of the literature, it seems that the chronological range of E. hydruntinus extends in Europe from the start of the late Middle Pleistocene until the Holocene (Figure 1). Its presence in levels dated toward 0.6 Ma in the North of Spain (and perhaps in the North of France), and in contemporary sites in Transcaucasia (Baryshnikov, 2002) and Acheulean sites from central Syria and the site of C-Spring Azrak in Jordan (Clutton-Brock, 1989; Reynaud Savioz and Morel, 2005) could suggest a first incursion of the species during the MIS 15 interglacial followed later by a second wave of migration from MIS 11-9.
Sub-species or Ecomorphotypes of Equus hydruntinus
Several forms of Equus hydruntinus have been described in the palaeontological literature in addition to the type first described by Stehlin and Graziosi (1935) from Romanelli. The name of the subspecies Equus hydruntinus minor indicates the small body size of the population from Lunel-Viel (Bonifay, 1991). It is possible that this form recurred over the course of the chronological range of E. hydruntinus (Boulbes and Rillardon, 2010). The description of this form was based on abundant material, including a cranium, but its affiliation with Equus hydruntinus has been questioned by certain authors (Azzaroli, 1990; Van der Made et al., 2017). Equus hydruntinus danubiensis, in contrast, represents a large-bodied form described by Samson (1975) from Romania. Eisenmann et al. (2008) also include the fossils of a gracile equid from Petralona (E. petraloniensis, Tsoukala, 1991; Tsoukala and Guérin, 2016), dated to 0.4 Ma, within the variation of the species: “E. hydruntinus petraloniensis.” According to certain authors, the slender equid from Venosa, considered to be one of the last representatives of Equus altidens (Alberdi and Palombo, 2013), could already represent Equus hydruntinus (Forsten, 1999a).
Another form, Equus hydruntinus davidi, defined by Alimen (1946) based on a single tooth from the Mousterian site of Saint-Agneau (Charente), has a longer protocone. This taxon was subsequently used by Prat (1968) to characterize the elongation of this dental character observed in many sites in the south-west of France post-dating the ≪ first Würmian stage ≫. Following Prat's work, Equus hydruntinus davidi has also been identified from other sites in the south of France (Eisenmann and Patou, 1980; Crégut-Bonnoure and Granier, 1982; Guadelli, 1995; Boulbes, 2009). Differences between teeth from Ramandils cave (MIS 5, Rusch et al., 2019) and Crouzade cave (MIS 3, Saos et al., 2019) is for example demonstrative (Figure 13). Another pattern (connected to the variation of protocone) is the reduction of the depth of vestibular groove on lower molars (Boulbes, 2009). These trends (fixed after MIS 3) could reflect a response to environmental pressures (genetic assimilation), the increase in the length of the protocone favored by the consumption of more abrasive food in cold and steppe climatic context (Boulbes, 2009). Guadelli and Delpech (2000) propose that the geographical barrier posed by the Alps could have fostered the independent evolution of western populations compared to those in the Italy-Balkans province, where populations with dental proportions similar to the type from the Romanelli caves persisted. However, isolated observations show that the morphotype with a long protocone could also have been present in Italy (Conti et al., 2010). The upper teeth of the enigmatic Equus graziosii (Azzaroli, 1979) (Maspino, Italy, Late Pleistocene) resemble those of the morph E. hydruntinus davidi (Figure 13F). This species is related to asses (Azzaroli, 1979; Eisenmann, 2006a); proximity with E. hydruntinus has been already pointed by Caloi (1995). This morphotype was also developed on the other side of the Pyrenees in the north-east of Spain, on the site of Teixoneres, dated to MIS 3 (Álvarez-Lao et al., 2017). Equus hydruntinus davidi was also identified from one late Würmian site in Portugal (Cardoso, 1995). We note here the fact that these late populations from the south of France and the north of Spain are more closely similar to hemiones in their morphological characteristics (convergences). The palaeogenetical study (Bennett et al., 2017) supports the hypothesis that hydruntine population could have gone through a bottleneck during the last two glacial periods and that Europe could have been recolonized from a refugial population, as proposed earlier on morphological grounds (Boulbes, 2009).
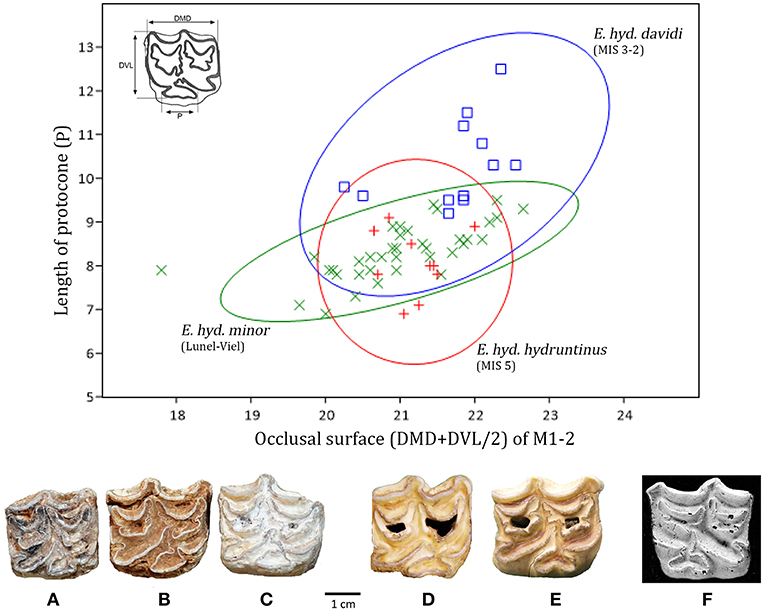
Figure 13. Bivariate plot of protocone length vs. occlusal surface of M1-2. Occlusal surface: average occlusal dimension [(DMD + DVL)/2]. Confidence ellipsoid (95 %). Data: E. hydruntinus minor (Lunel-Viel); E. hydruntinus hydruntinus (Ramandils, MIS 5); E. hydruntinus davidi (Crouzade, Arquet, MIS 3 and Valorgues, MIS 2) (Boulbes, pers. data). Upper teeth of Equus hydruntinus. (A–C) right P3, P4, M1 from Ramandils cave (E. h. hydruntinus); (C, D) right M1 and P4 from Crouzade cave (E. h. davidi); F: right P4 of Equus graziosii (from Eisenmann, 2006a).
Ecology of Equus hydruntinus
Like the hemiones, Equus hydruntinus was adapted to semi-arid, steppic conditions and shows a preference for temperate climates, although it could tolerate limited cold (Prat, 1968; Delpech, 1984; Eisenmann, 1984; Azzaroli, 1990; Bonifay, 1991; Burke et al., 2003; Palombo and Alberdi, 2017). According to Abbazzi et al. (1996), Equus hydruntinus is primarily adapted to dry Mediterranean conditions and does not necessarily depend on the presence of open landscapes. However, a preference for open biotopes is corroborated by the Holocene distribution of the species, reduced especially to open landscapes in southern and eastern Europe (Crees and Turvey, 2014). Its relationship with temperature is more difficult to ascertain. The species shows a preference for more temperate climates (Caloi, 1995; Forsten and Dimitrijevic, 2004), but it is also present in cool or cold climatic contexts (Prat, 1968; Boulbes, 2009). van Asperen et al. (2012) remark that the crania of E. hydruntinus can be distinguished from related equids by their relatively short muzzles, which indicates an adaptation to cold climatic conditions.
At Steinheim, E. hydruntinus is characterized by δ13C values that are clearly higher than those for E. ferus (Pushkina et al., 2014). Variation in the δ18O and δ13C values show that E. hydruntinus “occupied an ecological niche between the true horses, which inhabited shrubland, and the bovids of the grasslands, similar to that of the wooly rhinoceros, although with a slightly higher temperature and more closed vegetation” (p. 175, Pushkina et al., 2014). However, E. hydruntinus seems to have been less tolerant of climatic variations than the true horses, forcing it to adapt or, where appropriate, to migrate. The high chronological resolution of numerous well-dated sites indeed allows us to posit several phases of displacement of its geographical range over the course of the second half of the Late Pleistocene (Delpech, 2003). During the coldest phases of the Last Glacial Maximum (LGM) the species retreated to the peninsulas of southern Europe (Wilms, 1989; Crees and Turvey, 2014), a scenario that would also have characterized preceding glacial episodes (Boulbes, 2009). For example, its presence is confirmed at the start of MIS 4 in a relatively arid climate in Calabria (southern Italy), where a Mediterranean influence persisted (Marra and Bellomo, 2008). During cold episodes, Equus hydruntinus is accompanied by species that are very well-adapted to such a climate, such as for example M. primigenius and C. antiquitatis in the northeast of Spain (Álvarez-Lao et al., 2017), and Saiga tatarica and Rangifer tarandus in the south of France (Crégut-Bonnoure and Paccard, 1997).
Remarks and Conclusion
The main dispersal event concerning Equus in Europe (newcomers, in situ and anagenetic evolutionary processes)—first Equus, turnover during the post-Olduvai late Villafranchian, caballoid horse's emergence—are connected to major climatic change and document turnover of large mammal communities as well-demonstrated by Palombo and Alberdi (2017). The difficulty with regard to equid morphology is to discriminate between evolutionary and ecomorphological traits. The problem is even more complex because recent equids are known for their high level of plasticity and intra-specific variation, while differences between species are relatively low. Progressive characters such as elongation of the protocone, reduction of the ectoflexid, cursorial adaptations, and protrusion of the distal keel of the metapodials are variable at specific level and could have ecological significance. Despite this difficulty to draw up a phylogenetical and biochronological framework, it is possible to calibrate certain general patterns of evolution. Thus, emergence of elongated protocones in Europe for example can be dated between 1.2 and 0.7 Ma in supposed independant lineages (E. suessenbornsis, E. apolloniensis, E. altidens, E. mosbachensis) at the time of fundamental changes in the behavior of glacial cycles during the Quaternary glaciations and the expansion of open and dry environments.
The inventory of the different stenonid forms shows that at the time of the emergence of this group more than 2 million years ago, a wide range of morphologies occur—large, small, robust, slender—as well as combinations of these types. In contrast, the morphology of the teeth (more genetically dependent) seems to be less subject to variation. As a consequence, evolutionary models based on trend of size and/or gracilization are hard to demonstrate. Large and very large stenonids form a heterogeneous group and revision is needed to clarify their taxonomy and temporal distribution.
From the late Early Pleistocene to early Middle Pleistocene many fossils are assigned to Equus altidens. However, the morphology of the teeth of the Süssenborn equid seems too derived to be included in the variation of all the slender equids from this period. E. granatensis is well-defined and the primitive pattern of the teeth (very short protocones and especially very deep ectoflexids on molars and sometimes deep on premolars) casts doubt on an evolution from a stenonid stock. The presence of the more robust E. wuesti at around 1.0 Ma with proportions of limbs and teeth close to E. granatensis complicates the problem. Revision of this group is needed to solve the question of whether E. granatensis was an endemic species from the south of Spain, a primitive representative of E. altidens or an ecomorphotype (Palombo and Alberdi, 2017). The detailed study of the Epivillafranchian key sites of Vallparadis (Aurell-Garrido et al., 2010) and Bois de Riquet (Bourguignon et al., 2016) would certainly provide new information.
The large E. suessenbornensis has a wide geographical range from Beringia to Western Europe which is accompanied with little variation in terms of morphology and proportions. In contrast, slender species split in several geographical provinces. Interesting is the succession and regionalization of co-occurrences of equids in the course of Early and Middle Pleistocene: E. suessenbornensis-E. granatensis/E. altidens or E. wuesti in western and central Europe and E. mygdoniensis-E. apolloniensis around the eastern Mediterranean. The latter seems not to be linked to stenonid s. s. or E. suessenbornensis but could represent a step within the lineage of asses soon after their differentiation (Eisenmann and Boulbes, in preparation) in accordance with DNA and palaeogenetic projections (Jonsson et al., 2014; Bennett et al., 2017). These lineages finally were replaced by the more generalist caballoid horse group at the beginning of Middle Pleistocene.
Despite the overall homogeneous morphology of true horses during the Middle Pleistocene, paradoxically the ecomorphological adaptations described above coupled with some evolving traits provide real biochronological details in a given geographical province. Since the exact combination of the environmental factors that influence horse morphology subtly varies between the different glacials and interglacials, each oxygen isotope stage was characterized by a specific morphology, as outlined above. However, some caution needs to be taken in using horse morphology as a biostratigraphic indicator. First, the patterns sketched above were identified in northwest and central European horses or south of France, and cannot be transposed directly on other regions, such as southern or eastern Europe, where climatic conditions may have been significantly different. Second, due to the relatively homogeneous morphology of Middle Pleistocene horses, assemblages analyzed must be sufficiently large to produce statistically significant results. Although there is clear variation between ecotypes, there is also a significant amount of overlap, so any attempt to determine the affinities of small numbers of specimens must be regarded as preliminary. Finally, due to these overlaps and the small differences between ecotypes, it is advisable to apply a range of analytical methods to new assemblages to identify their morphology. Statistical methods such as principal component analysis and discriminant function analysis can disentangle more subtle variations (van Asperen, 2010, 2012). In this regard, recent developments using geometric morphometric approaches (on enamel patterns in particular) provide good phylogenetic and taxonomic markers (Barrón-Ortiz et al., 2017; Cucchi et al., 2017; Hanot, 2018) that may be complementary to traditional morphometry. Comparisons between the results of different methods can aid in solving the question of the phylogeography and regionalization of horses in Europe. Combining metric/morphological data with studies of tooth wear characteristics (mesowear, microwear) and isotopic analyses of teeth (Saarinen et al., 2016; Uzunidis et al., 2017) is the best way to ensure the robustness of palaeoecological inferences from ecomorphological patterns and their use as a proxy.
Several questions remain to be solved concerning Equus hydruntinus. In the first place, its origin, namely whether it corresponds to a local evolution in Europe or whether it entered Europe in one or more waves of migration from Asia. The detailed study of the oldest fossils (Vallparadis) and comparisons with intermediate forms to E. altidens (Venosa, Petralona) will provide essential information. Secondly, its status as a distinct species or subspecies of Equus hemionus. Paleontology has demonstrated the existence of many distinct morphological characters between Equus hydruntinus and Equus hemionus although most are plesiomorphic. The presence in E. hydruntinus of the mitotype of Equus hemionus (samples from 100 Ka) in association with distinctive morphological characteristics implies that Equus hydruntinus can be seen as an ecomorph or a subspecies of Equus hemionus (Bennett et al., 2017). However, according to the first appearance date of the species (0.6 Ma), it is also not impossible to consider that in the recent history of the taxon (0.1 Ma) gene transfer took place by introgressive hybridization. The sympatric cases noted between the two species in the Eemian, notably in the Binagady site (Azerbaidjan) (Eisenmann and Mashkour, 1999), followed by numerous observations during the Upper Pleistocene of characters convergent with the hemiones could support this hypothesis. The particular form of Equus hydruntinus davidi from southern France and the enigmatic Equus graziosii from Italy require further reflection. Palaeogenetic investigations, not only on the mitochondrial DNA but also the nuclear DNA, will in parallel have to multiply on fossils of various geographical origins. Given the difficulties with their identification, it is also not impossible to rule out the existence of several small, slender equidae in Western Europe including Equus hydruntinus, Equus hemionus and possibly other forms not recognized until now.
Data Availability
All datasets generated for this study are included in the manuscript and/or the Supplementary Files.
Author Contributions
NB and EA wrote sections of the manuscript. Both authors contributed to manuscript revision, read, and approved the submitted version.
Funding
This article was funded by the UMR 7194 HNHP, MNHN-CNRS-UPVD, EPCC-CERP Tautavel.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors are very grateful to the two reviewers who helped to improve the manuscript. Special thanks to the editors G. M. Semprebon, F. Rivals, and R. L. Bernor for inviting us to contribute to this research topic Examining Evolutionary Trends in Equus and its Close Relatives from Five Continents. V. Eisenmann kindly gave us some of her personal data and permission to mention new results about E. apolloniensis and E. wuesti, we are grateful to her. G. D. Koufos gave us some data of equids from Mygdonia Basin, thanks to him. Thanks to H. de Lumley and A. M. Moigne who allowed us (NB) to study the important collections of CERP, at Tautavel. We also thank I. Toro-Moyano and B. Martínez Navarro, for welcoming one of us (NB) a few years ago to study the equid from Venta Micena at the Museo de Prehistoria de Orce. The studies conducted on the sites of Les Ramandils, La Crouzade, and Tournal are undertaken in the framework of a Collective Research Program (PCR) of the French Ministry of Culture Le monde moustérien méditerranéen entre Rhône et Pyrénées, dir. T. Saos. The work on the Denizli Basin in Turkey was done in the context of the First Homo in Turkey research program funded by the CNRS (PICS 2016-2018, dir. A. Vialet and with the collaboration of M. C. Alçiçek). Current excavations carried out in the sites of Caune de l'Arago, dir. C. Perrenoud, Bois de Riquet, dir. L. Bourguignon, l'abri du Maras, dir. M. H. Moncel, Portel-Ouest, dir. G. Becam, La Crouzade, dir. T. Saos, Coulet des Roches, dir. E. Crégut-Bonnoure are supported by the French Ministry of Culture. Research in the site of cueva del Ángel is carried out by the Fundación Instituto de Investigación de Prehistoria y Evolución Humana (FIPEH, dir. C. Barroso Ruiz).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2019.00301/full#supplementary-material
References
Abbazzi, L., Delgado Huertas, A., Iacumin, P., Longinelli, A., Ficcarelli, G., Masini, F., et al. (1996). Mammal changes and isotopic biogeochemistry. An interdisciplinary approach to climatic-environmental reconstructions at the last Pleniglacial/Late glacial transition in the Paglicci cave section (Gargano, Apulia, SE Italy). Il Quaternario 9, 573–580.
Alberdi, M. T. (2010). “Estudio de los caballos de los yacimientos de Fuente Nueva-3 y barranco León-5 (Granada),” in Ocupaciones Humanas en el Pleistoceno Inferior y Medio de Lacuenca de Guadix-Baza Memoria Científica, eds I, Toro, B. Martínez-Navarro, J. Agustí (Junta de Andalucía; Consejería de Cultura; E.P.G. Arqueología Monográfico).
Alberdi, M. T., Caloi, L., and Palombo, M. R. (1988). The Quaternary fauna of Venosa: equids. Bull. Musée d'Anthropol. Préhistorique Monaco 31, 5–39.
Alberdi, M. T., Caloi, L., and Palombo, M. R. (1991). Pleistocene equids from Western Europe: their biostratigraphic and palaeoecological characteristics. Ongulés Ungulates 91, 31–35.
Alberdi, M. T., Ortiz Jauregizar, E., and Prado, J. L. (1998). A quantitative review of european stenonid horses. J. Paleontol. 72, 371–387. doi: 10.1017/S0022336000036350
Alberdi, M. T., and Palombo, M. R. (2013). The late Early to early Middle Pleistocene stenonoid horses from Italy. Q. Int. 288, 25–44. doi: 10.1016/j.quaint.2011.12.005
Alberdi, M. T., and Piñero, P. (2015). “Estudio de los caballos del yacimiento de Cueva Victoria, Pleistoceno Inferior (Murcia),” in Paleontología y Geología de Cueva Victoria. Museo Arqueológico Municipal de Cartagena, eds L. Gibert and E. Ferràndez-Cañadell, 325–358.
Alberdi, M. T., and Ruiz Bustos, A. (1985). Descripción y significado bioestratigráfico y climático de equus e hippopotamus en el yacimiento de Venta Micena (Granada). Estudios Geol. 41, 251–261. doi: 10.3989/egeol.85413-4708
Alberdi, M. T., and Ruiz Bustos, A. (1989). “Taxonomáa y Biostratigrafía de Equidae (Mammalia, Perissodactyla) en la cuenca de Guadix-Baza (Granada),” in Geología y Paleontología de la Cuenca de Guadix-Baza. Trabajos Sobre Neógeno-Cuaternario Vol. 11, eds M. T. Alberdi and F. P. Bonadonna, 239–271.
Álvarez-Lao, D. J., Rivals, F., Sánchez-Hernández, C., Blasco, R., and Rosell, J. (2017). Ungulates from Teixoneres cave (Moià, Barcelona, Spain): presence of cold-adapted elements in NE Iberia during the MIS 3. Palaeogeogr. Palaeoclimatol. Palaeoecol. 466, 287–302. doi: 10.1016/j.palaeo.2016.11.040
Antoine, P., Moncel, M. H., Limondin-Lozouet, N., Locht, J. L., Bahain, J. J., Moreno, D., et al. (2016). Palaeoenvironment and dating of the early acheulean localities from the Somme river basin (Northern France): new discoveries from the high terrace at Abbeville-carriere Carpentier. Q. Sci. Rev. 149, 338–371. doi: 10.1016/j.quascirev.2016.07.035
Antunes, M. T. (2006). “The zebro (equidae) and its extinction in portugal, with an appendix on the noun zebro and the modern zebra,” in Equids in Time and Space, ed M. Mashkour (Oxford: Oxbow Books, 211–236.
Aouadi, N. (2001). Equidés pléistocènes non caballins en Europe du Sud (Thèse de doctorat). Université d'Aix-Marseille, Marseille, France.
Aouadi, N., and Bonifay, M. F. (2008). Les métapodes des chevaux de Ceyssaguet: études morphologique et biométrique. Bull. Musée d'Anthropol. Préhistorique Monaco 48, 17–29.
Aouraghe, H. (1992). Les faunes de grands mammifères du site Pléistocène moyen d'Orgnac 3 (Ardèche, France). Etude paléontologique et palethnographique. Implications paléoécologiques et biostratigraphiques (Thèse de Doctorat). Muséum National d'Histoire Naturelle, Paris, France.
Arribas, A., Garrido, G., Viseras, C., Soria, J. M., Pla, S., et al. (2009). A mammalian lost world in southwest Europe during the Late Pliocene. PLoS ONE 4:e7127. doi: 10.1371/journal.pone.0007127
Athanassiou, A. (2001). New data on the Equus stenonis from the Late Pliocene locality of Sésklo (Thessaly, Greece). Géodiversitas 23, 439–469.
Auguste, P. (1995). Cadres biostratigraphique et paléoécologique du peuplement humain dans la France septentrionale durant le Pléistocène, Apports de l'étude paléontologique des grands mammifères de Biache-Saint-Vaast. Paris: Muséum National d'Histoire Naturelle (Unpublished PhD thesis).
Aurell-Garrido, J., Madurell-Malapeira, J., Alba, D. M., and Moyà-Solà, S. (2010). The stenonian and caballoid equids from the Pleistocene section of Vallparadís (Terrassa, Catalonia, Spain). Cidaris 30, 61–66.
Azzaroli, A. (1965). The two villafranchian horses of the Upper Valdarno. Palaeontogr. Ital. 59, 1–12.
Azzaroli, A. (1979). On a Late Pleistocene ass from Tuscany with notes on the history of ass. Palaeontol. Ital. 71, 27–47.
Azzaroli, A. (1984). On some vertebrate remains of Middle Pleistocene age from the upper valdarno and val di chiana, tuscany. Palaeontogr. Ital. 73, 104–115.
Azzaroli, A. (1990). “The genus Equus in Europe,” in European Neogene Mammal Chronology, eds E. H. Lindsay, V. Fahlbusch, and P. Mein (New York, NY: Plenum Press, 239–256.
Azzaroli, A. (1992). Ascent and decline of monodactyl equids: a case for prehistoric overkill. Ann. Zool. Fennici 28, 151–163.
Barrón-Ortiz, C. I., Rodrigues, A. T., Theodor, J. M., Kooyman, B. P., Yang, D. Y., and Speller, C. F. (2017). Cheek tooth morphology and ancient mitochondrial DNA of late pleistocene horses from the western interior of North America: implications for the taxonomy of North American Late Pleistocene Equus. PLoS ONE 12:e0183045. doi: 10.1371/journal.pone.0183045
Baryshnikov, G. F. (2002). Local biochronology of Middle and Late Pleistocene mammals from the Caucasus. Russ. J. Theriol. 1, 61–67. doi: 10.15298/rusjtheriol.01.1.05
Belan, A. G. (1985). Pozndnepleistotsenovaya shirokopalaya loshad' basseina Desny Mlekopitayuschie Severnoy Evrazii vchetvertichnom periode. Trudy Zoologicheskogo Instituta ANSSSR. 131, 50–63.
Bellai, D. (1998). Le cheval du gisement Pléistocène moyen de la Caune de l'Arago (Tautavel, P.-O.). Quaternaire 9, 325–335. doi: 10.3406/quate.1998.1614
Bennett, E. A., Champlot, S., Peters, J., Arbuckle, B. S., Guimaraes, S., Pruvost, M., et al. (2017). Taming the late Quaternary phylogeography of the Eurasiatic wild ass through ancient and modern DNA. PLoS ONE 12:e0174216. doi: 10.1371/journal.pone.0174216
Bernor, R. L., Cirilli, O., Wang, S.-Q., and Rook, L. (2018). Equus cf. livenzovensis from montopoli, Italy (early Pleistocene; MN16b; ca. 2.6 Ma). Bollettino Soc. Paleontol. Ital. 57, 203–216.
Bignon, O. (2003). Diversité et exploitation des équidés au Tardiglaciaire en Europe occidentale. Implications pour les stratégies de subsistance et les modes de vie au Magdalénien et à l'Azilien ancien du Bassin de Paris. (Thèse de Doctorat), Université Paris X-Nanterre, Nanterre, France.
Bignon, O., Baylac, M., Vigne, J.-D., and Eisenmann, V. (2005). Geometric morphometrics and the population diversity of Late Glacial horses in Western Europe (Equus gallicus arcelini): phylogeographic and anthropological implications. J. Archaeol. Sci. 32, 375–391. doi: 10.1016/j.jas.2004.02.016
Bignon, O., and Eisenmann, V. (2006). “Western european Late Glacial horses diversity and its ecological implications,” in Equids in Time and Space, 9th ICAZ Conference, Durham 2002, ed M. Mashkour (Oxford: Oxbow Books, 161–171.
Blackburn, T. M., Gaston, K. J., and Loder, N. (1999). Geographic gradients in body size: a clarification of Bergmann's rule. Divers. Distribut. 5, 165–174. doi: 10.1046/j.1472-4642.1999.00046.x
Bocherens, H., Billiou, D., Patou-Mathis, M., Bonjean, D., Otte, M., and Mariotti, A. (1997). Paleobiological implications of the isotopic signatures (13C, 15N) of fossil mammal collagen in Scladina cave (Sclayn, Belgium). Q. Res. 48, 370–380. doi: 10.1006/qres.1997.1927
Bocherens, H., and Drucker, D. (2003). Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems. Int. J. Osteoarchaeol. 13, 46–53. doi: 10.1002/oa.662
Boddaert, P. (1785). Elenchus Animalium, Vol. I: Sistens Quadrupedia huc Usque Nota, Eorumque Varietates: Ad Ductum Naturae, Quantum Fieri Potuit Disposita. Roterodami: Apud C.R. Hake, 174
Bœuf, O. (1986). L'équidé du site Villafranchien de Chilhac (Haute-Loire, France): Equus stenonis guthi nov. Subsp. Ann. Paléontol. 72, 29–67.
Bonifay, M. F. (1966). Etude paléontologique de la grotte de la Balauzière (Gard). Bull. Mus. Anthrop. Préh. Monaco 13, 91–139.
Bonifay, M. F. (1980). Le cheval du Pléistocène moyen des grottes de Lunel-Viel (hérault) Equus mosbachensis palustris n. ssp. Gallia Préhistoire 23, 233–281. doi: 10.3406/galip.1980.1652
Bonifay, M. F. (1991). “Equus hydruntinus Regalia minor n.ssp. from the caves of Lunel-Viel (Hérault, France),” in Equids in the Ancient World, Vol. II, Beihefte Zum Tübinger Atlas des Vorderen Orients, Reihe A, eds R. H. Meadow and H. P. Uerpmann (Wiesbaden: Naturwissenschaften, 178–216.
Bouchud, J. (1978). Les grands herbivores rissiens de l'Aven II des Abîmes de La Fage, à Noailles (Corrèze). Nouv. Arch. Mus. Hist. Nat. Lyon 16, 9–39.
Boulbes, N. (2009). Étude compare de la denture d'Equus hydruntinus (Mammalia, Perissodactyla) dans le Sud-Est de la France. Implications biogéographiques et biostratigraphiques. Quaternaire 20, 449–465. doi: 10.4000/quaternaire.5319
Boulbes, N. (2010). Le cheval de Romain-la-Roche, Equus achenheimensis (Mammalia, Perissodactyla), contribution à la biochronologie des équidés caballins au Pléistocène moyen. Rev. Paléobiol. 29, 747–770.
Boulbes, N. (2014). “Etude préliminaire du cheval de l'unité archéostratigraphique Q,” dans Caune de l'Arago (Tautavel, Pyrénées-Orientales), Rapport de Fouille 2014, ed C. Perrenoud (SRA-DRAC Languedoc-Roussillon), 48–55.
Boulbes, N., Mayda, S., Titov, V. V., and Alçiçek, M. C. (2014). The Late Villafranchian large mammals from the Denizli Basin travertines (Southwest Anatolia, Turkey). L'Anthropologie 118, 44–73. doi: 10.1016/j.anthro.2014.01.001
Boulbes, N., and Rillardon, M. (2010). “Révision paléontologique et analyse archéozoologique d'Equus hydruntinus (Mammalia, Perissodactyla) de la Baume de Valorgues (St-Quentin-La-Poterie, Gard) et remarques sur l'évolution de la taille de cette espèce du Pléistocène moyen à l'Holocène,” in Histoire d'Equidés, des textes, des images et des os Monographies d'Archéologie Méditerranéenne, Hors-sérien°4, eds A. Gardeisen, E. Furet, and N. Boulbes (Lattes), 9–27.
Bourguignon, L., Crochet, J.-Y., Capdevila, R., Ivorra, J., Antoine, P.-O., Agustí, J., et al. (2016). Bois-de-Riquet (Lézignan-la-Cèbe, Hérault): a late Early Pleistocene archeological occurrence in southern France. Q. Int. 393, 24–40. doi: 10.1016/j.quaint.2015.06.037
Bridgland, D., and Schreve, D. (2004). Quaternary lithostratigraphy and mammalian biostratigraphy of the lower thames terrace system, South-East England. Quaternaire 15, 29–40. doi: 10.3406/quate.2004.1752
Britton, K., Gaudzinski-Windheuser, S., Roebroeks, W., Kindler, L., and Richards, M. P. (2012). Stable isotope analysis of well-preserved 120,000-year-old herbivore bone collagen from the middle palaeolithic site of Neumark-Nord 2, Germany reveals niche separation between bovids and equids. Palaeogeol. Palaeoclimatol. Palaeoecol. 333–334, 168–177. doi: 10.1016/j.palaeo.2012.03.028
Bro-Jørgensen, J. (2008). Dense habitats selecting for small body size: a comparative study on bovids. Oikos 117, 729–737. doi: 10.1111/j.0030-1299.2008.16069.x
Brugal, J. P., and Boudadi-Maligne, M. (2011). Quaternary small to large canids in Europe: taxonomic status and biochronological contribution. Q. Int. 243, 171–182 doi: 10.1016/j.quaint.2011.01.046
Burke, A., Eisenmann, V., and Ambler, G. (2003). The systematic position of Equus hydruntinus, an extinct species of Pleistocene equid. Q. Res. 59, 459–469. doi: 10.1016/S0033-5894(03)00059-0
Caloi, L. (1995). Il genere Equus nell'Italia centrale. Studi Geologici Camerti, Vol. Spec. Biostratigrafia dell'Italia centrale parte b, 469–486.
Caloi, L. (1997). New forms of equids in Western Europe and palaeoenvironmental changes. Geobios 30, 267–284. doi: 10.1016/S0016-6995(97)80233-1
Caloi, L. (2002). The two equids of the Middle Pleistocene of the site Venosa-Loreto (Southern Italy): functional morphology of the cranial remains. Geol. Romana 36, 275–287.
Caloi, L., and Palombo, M. R. (1982). Alcune considerazioni su Equus stenonis Cocchi e Equus stehlini Azzaroli. Paleontol. Stratigr. Evol. 2, 57–61.
Caloi, L., and Palombo, M. R. (1987). Osservazioni sugli Equidi italiani del Pleistocene medio inferior. Geol. Romana 26, 187–221.
Cardoso, J. L. (1995). Presença de Equus hydruntinus Regalia, 1905 no würm recente de Portugal. Comun. Inst. Geol. Mineiro 81, 97–108.
Cardoso, J. L., and Eisenmann, V. (1989). Equus caballus antunesi, nouvelle sous-espèce quaternaire du Portugal. Palaeovertebrata 19, 47–72.
Cerdeno, E., and Alberdi, M. T. (2006). Historia del conocimiento de los perisodáctilos del Neógeno español. Estud. Geol. 62, 481–494. doi: 10.3989/egeol.0662141
Clutton-Brock, J. (1989). “A re-consideration of the fossil fauna from C-Spring, Azraq,” in The Hammer on the Rock: Studies on the Early Paleolithic of Azraq, eds L. Copeland and F. Hours (Oxford: BAR International Series, 391–397.
Collinge, S. E. (2001). Body Size and Community Structure in British Pleistocene Mammals. London: University of London. (Unpublished PhD thesis).
Conti, N., Petronio, C., and Salari, L. (2010). The Late Pleistocene equids from “tana delle iene” (Ceglie Messapica, Brindisi, Southern Italy). Bollettino Soc. Paleontol. Ital. 49, 227–236.
Cramer, B. (2002). Morphometrische untersuchungen and quartären pferden in Mitteleuropa Tübingen: Eberhard-Karls-Universität Tübingen (Unpublished PhD thesis).
Crees, J. J., and Turvey, S. T. (2014). Holocene extinction dynamics of Equus hydruntinus, a late-surviving European megafaunal mammal. Q. Sci. Rev. 91, 16–29. doi: 10.1016/j.quascirev.2014.03.003
Crégut, E. (1980). Equus mosbachensis tautavelensis n. ssp. Un nouvel equidae (Mammalia, Périssodactyla) du gisement Pléistocène moyen anté-rissien de la Caune de l'Arago (Tautavel, Pyrennées-Orientales, France). Géobios 13, 121–127 doi: 10.1016/S0016-6995(80)80019-2
Crégut-Bonnoure, E., Boulbes, N., Desclaux, E., and Marciszak, A. (2018). New insights into the LGM and LG in Southern France (Vaucluse): the mustelids, micromammals and horses from Coulet des Roches. Quaternary 1, 1–47. doi: 10.3390/quat1030019
Crégut-Bonnoure, E., and Granier, J. (1982). À propos d'une dent d'Equus hydruntinus (Mammalia, Perissodactyla) du lœss récent de Collias (Gard, France). Bull. Soc. Linnéenne Lyon 7, 234–240. doi: 10.3406/linly.1982.10550
Crégut-Bonnoure, E., and Paccard, M. (1997). La faune á Saïga tatarica de Chinchon I (Saumanes, Vaucluse). Quaternaire 8, 391–407. doi: 10.3406/quate.1997.1588
Cucchi, T., Mohaseb, A., Peigné, S., Debue, K., Orlando, L., and Mashkour, M. (2017). Detecting taxonomic and phylogenetic signals in equid cheek teeth: towards new palaeontological and archaeological proxies. R. Soc. Open Sci. 4:160997. doi: 10.1098/rsos.160997
Daujeard, C., Vettese, D. B, ritton, K., Béarez, P., Boulbes, N., Crégut-Bonnoure, E., et al. (2019). Neanderthal selective hunting of reindeer? The case study of Abri du Maras (south-eastern France). Archaeol. Anthropol. Sci. 11, 985–1011. doi: 10.1007/s12520-017-0580-8
David, P., and Prat, F. (1962). Sur la présence d'un cheval de grande taille dans certains horizons moustériens de la station de La Chaise de Vouthon (Charente). Bull. Mém. Soc. Archéol. Historique Charente.
Davis, S. J. (1980). Late Pleistocene and Holocene equid remains from Israel. Zool. J. Linnean Soc. 70, 289–312. doi: 10.1111/j.1096-3642.1980.tb00854.x
De Beaulieu, J.-L., and Reille, M. (1995). Pollen records from the velay craters: a review and correlation of the Holsteinian interglacial with isotopic stage 11. Mededelingen Rijks Geologische Dienst 52, 59–70.
De Giuli, C. (1987). Late Villafranchian faunas of ltaly: the Selvella Local Fauna in the southern Chiana valley, Umbria. Palaeontogr. ltal. 74, 11–50.
Delafond, F., and Depéret, C. (1893). Les Terrains Tertiaires de la Bresse et Leurs Gites de Lignites et de Minerais de fer. Etudes des Gites Mineraux de France. Paris: Imprimerie Nationale, 332.
Delpech, F. (1984). Les ongulés en Périgord et Nord-Ouest du Quercy durant le Würm III. Geobios 17, 531–548. doi: 10.1016/S0016-6995(84)80027-3
Delpech, F. (2003). “L'environnement animal des européens au Paléolithique supérieur,” in L'Europe préhistorique. Editions du CTHS, eds R. Desbrosse and A. Thévenin (Paris), 271–289.
Delpech, F., Grayson, D. K., and Rigaud, J. P. (2000). Biostratigraphie et paléoenvironnements du début du Würm récent d'après les grands mammifères de l'abri du Flageolet I (Dordogne, France). Paléo 12, 97–126. doi: 10.3406/pal.2000.1598
Dive, J., and Eisenmann, V. (1991). “Identification and discrimination of first phalanges from Pleistocene and modern Equus, wild and domestic,” in Equids in the Ancient World, Volume II, eds R. H. Meadow and H.-P. Uerpmann (Wiesbaden: Dr Ludwig Reichert Verlag, 278–315.
Drucker, D. G., Bridault, A., and Cupillard, C. (2012). Environmental context of the magdalenian settlement in the Jura mountains using stable isotope tracking (13C, 15N, 34S) of bone collagen from reindeer (Rangifer tarandus). Q. Int. 272–273, 322–332. doi: 10.1016/j.quaint.2012.05.040
Drucker, D. G., Bridault, A., Hobson, K. A., Szuma, E., and Bocherens, H. (2008). Can carbon-13 abundances in large herbivores track canopy effect in temperate and boreal ecosystems? evidence from modern and ancient ungulates. Palaeogeol. Palaeoclimatol. Palaeoecol. 266, 69–82. doi: 10.1016/j.palaeo.2008.03.020
Drucker, D. K., Bocherens, H., and Biliou, D. (2003). Evidence for shifting environmental conditions in Southwestern France from 33 000 to 15 000 years ago derived from carbon-13 and nitrogen-15 abundances in collagen of large herbivores. Earth Planet. Sci. Lett. 216, 163–173. doi: 10.1016/S0012-821X(03)00514-4
Duncan, P., Foose, T. J., Gordon, I. J., Gakahu, C. G., and Lloyd, M. (1990). Comparative nutrient extraction from forages by grazing bovids and equids: a test of the nutritional model of equid/bovid competition and coexistence. Oecologia 84, 411–418. doi: 10.1007/BF00329768
Duval, M., Bahain, J. J., Falguères, C., Garcia, J., Guilarte, V., Grün, R., et al. (2015). Revisiting the ESR chronology of the Early Pleistocene hominid occupation at Vallparadís (Barcelona, Spain). Q. Int. 389, 213–223. doi: 10.1016/j.quaint.2014.08.054
Echchat, R. (2008). Etude stratigraphique, sédimentologique et micromorphologique du remplissage Pléistocène supérieur de la grotte Tournal (Bize-Minervois, Aude) (Thèse de Doctorat). Université de Perpignan, Perpignan, France.
Ehlers, J., Eissmann, L., Lippstreu, L., Stephan, H.-J., and Wansa, S. (2004). “Pleistocene glaciations of North Germany,” in Quaternary Glaciations, Extent and Chronology, Part I Europe, Developments in Quaternary Science, eds J. Ehlers and P. L. Gibbard (Amsterdam: Elsevier, 135–146.
Eisenmann, V. (1979). Les métapodes d'Equus sensu lato (Mammalia, Perissodactyla). Geobios 12, 863–886. doi: 10.1016/S0016-6995(79)80004-2
Eisenmann, V. (1980). “Les chevaux (Equus sensu lato) fossiles et actuels : crânes et dents jugales supérieures,” in Crânes et Dents Jugales Supérieures (Paris: CNRS Editions), 186.
Eisenmann, V. (1981). Etude des dents jugales inférieures des Equus (Mammalia, Perissodactyla) actuels et fossiles. Palaeovertebrata 10, 127–226.
Eisenmann, V. (1984). Sur quelques caractères adaptatifs du squelette d'Equus et leurs implications paléoécologiques. Bull. Mus. Nat. Hist. Nat. 2, 185–195.
Eisenmann, V. (1986). “Comparative osteology of modern and fossil horses, half-asses, and asses,” in Equids in the Ancient World, Vol. II, eds R. H. Meadow, and H -P. Uerpmann (Wiesbaden: Reichert, 67–116.
Eisenmann, V. (1988). Tentative typology and biostratigraphy of some Middle and Late Pleistocene Western European horses, Il Quaternario 1, 103–109.
Eisenmann, V. (1991a). Proportions squelettiques de chevaux quaternaires et actuels. Géobios 13, 25–32. doi: 10.1016/S0016-6995(66)80006-2
Eisenmann, V. (1991b). Les chevaux quaternaires européens (Mammalia, Perissodactyla). Taille Typologie Biostratigraphie Taxonomie Géobios 24, 747–759. doi: 10.1016/S0016-6995(06)80303-7
Eisenmann, V. (1992). Systematic and biostratigraphical interpretation of the equids from Qafzeh, Tabun, Shkul and Kebara (Archeuloyabrudian to Upper Paleolithic of Israel). Archaeozoologia 5, 43–62.
Eisenmann, V. (1999). “Equus granatensis of Venta Micena and evidence for primitive non stenonid horses in the Lower Pleistocene,” in The Hominids and Their Environment During the Lower and Middle Pleistocene of Eurasia. Proceedings of the International Conference of Human Paleontology, eds J. Gibert, F. Sanchez, L. Gibert, and F. Ribot (Orce), 175–190.
Eisenmann, V. (2002). “The primitive horses of the Vatera Formation (Lesbos, Greece),” in Proceedings of the 1st International Workshop “On Late Plio/Pleistocene Extinction and Evolution in the Palearctic. The Vatera site”. Annales Géologiques des Pays Helléniques, 1ère série, Vol. XXXIX (A), 131–153.
Eisenmann, V. (2003). “Gigantic horses. advances in paleontology ‘hen to panta',” Papers in Honour of C. Radulescu P. M. Samson (Bucharest), 31–40.
Eisenmann, V. (2004a). Les équidés (Mammalia, Perissodactyla) de Saint-Vallier (Drôme, France) et les équidés plio-pléistocènes d'Europe. Géobios 37, 279–305. doi: 10.1016/S0016-6995(04)80019-6
Eisenmann, V. (2004b). “Equus: an evolution without lineages?,” in 18th International Senckenberg Conference, VI International Palaeontological Colloquium in Weimar, Late Neogene and Quaternary Biodiversity and Evolution: Regional Developments and Interregional Correlations (Weimar: Terra Nostra, 101–104.
Eisenmann, V. (2006a). Pliocene and Pleistocene equids: palaeontology versus molecular biology. Courier Forschungsinstitut Senckenberg 256, 71–89.
Eisenmann, V. (2006b). “Discriminating equus skulls: the Franck's index and the new palatal index,” in Equids in Time and Space: Papers in Honour of Vera Eisenmann : Proceedings of the 9th Conference of the International Council of Archaeozoology, ed M. Mashkour (Durham), 13, 172–182.
Eisenmann, V. (2008). Süssenborn, Introduction. Available online at: https://vera-eisenmann.com//sussenborn-introduction
Eisenmann, V. (2010). Sussemionus, a new subgenus of Equus (Perissodactyla, Mammalia). Comp Rendus Biol. 333, 235–240. doi: 10.1016/j.crvi.2009.12.013
Eisenmann, V. (2014). Main Caballine Crania Morphs. Available online at: https://vera-eisenmann.com/main-caballine-crania-morphs
Eisenmann, V. (2017). The Senèze Equids. Available online at: https://vera-eisenmann.com/the-seneze-equids-text
Eisenmann, V. (2019). Available online at: http://www.vera-eisenmann.com
Eisenmann, V., Adover, R., Moissenet, E., and Mourer-Chauvire, C. (1990). Le cheval de Monteagudo del Castillo (Province de Teruel, Espagne). Paleontol. Evol. 23, 239–252.
Eisenmann, V., Alberdi, M. T., de Giuli, C., and Staesche, U. (1988). “Methodology,” in Studying Fossil Horses, Collected Papers After the “New York International Hipparion Conference, 1981” eds M. Woodburne and P. Sondaar (Brill: Leiden, 1–71.
Eisenmann, V., and Baryshnikov, G. F. (1995). Equus cf. taubachensis et E. hydruntinus de la grotte de Prolom 2 (Crimée, Ukraine). Bull. Mus. Natl. Hist. Nat. 16, 329–347.
Eisenmann, V., and Baylac, M. (2000). Extant and fossil Equus (Mammalia, Perissodactyla) skulls: a morphometric definition of the subgenus Equus. Zool. Scripta 29, 89–100. doi: 10.1046/j.1463-6409.2000.00034.x
Eisenmann, V., and Beckouche, S. (1986). “Identification and discrimination of metapodials from Pleistocene and modern Equus, wild and domestic,” in Equids in the Ancient World, Vol. II, eds R. H. Meadow and H- P. Uerpmann (Wiesbaden: Reichert, 117–163.
Eisenmann, V., Crégut-Bonnoure, E., and Moigne, A.-M. (1985). Equus mosbachensis et les grands chevaux de la Caune de l'Arago et de Lunel-Viel: crâniologie comparée. Bull. Muséum Natl D'histoire Naturelle 2, 157–173.
Eisenmann, V., and David, F. (1994). Le cheval de Mauran (Haute-Garonne) : stade de transition évolutive entre Equus taubachensis et E. germanicus? in Hommes et bisons du Paléolithique moyen à Mauran (Haute-Garonne), eds C. Farizy, F, David, and J. Jaubert (Paris: CNRS éditions, 41–46.
Eisenmann, V., and David, F. (2002). “Évolution de la taille des chevaux d'Arcy-sur-Cure et de quelques autres chevaux quaternaires,” in L'Aurignacien de la Grotte du Renne, ed B. Schmider (Gallia Préhistoire) Suppl. 34, 97–102.
Eisenmann, V., Howe, J., and Pichardo, M. (2008). Old world hemiones and new world slender species (Mammalia, Equidae). Palaeovertebrata 36, 159–233. doi: 10.18563/pv.36.1-4.159-233
Eisenmann, V., and Kuznetsova, T. V. (2004). Early Pleistocene equids (Mammalia, Perissodactyla) of Nalaikha (Mongolia) and the emergence of modern Equus Linnaeus, 1758. Geodiversitas 26, 535–561.
Eisenmann, V., and Mashkour, M. (1999). The small equids of Binagady (Azerbaidjan) and Qazvin (Iran): E. hemionus binagadensis nov. subsp. and E. hydruntinus. Geobios 32, 105–122. doi: 10.1016/S0016-6995(99)80085-0
Eisenmann, V., and Patou, M. (1980). La faune de la grotte de Félines-Termenès (Aude). Résultats préliminaires et étude détaillée des restes d'Equus hydruntinus (Mammalia, Perissodactyla). L'Anthropologie 84, 633–649.
Eisenmann, V., and Sergej, V. (2011). Unexpected finding of a new Equus species (Mammalia, Perissodactyla) belonging to a supposedly extinct subgenus in Late Pleistocene deposits of Khakassia (southwestern Siberia). Geodiversitas 33, 519–530. doi: 10.5252/g2011n3a5
Falguères, C., Ghaleb, B., Tombret, O., Ben Arous, E., Richard, M., Moigne, A. M., et al. (2019). ESR/U-series dates on Equus teeth from the Middle Pleistocene Acheulean site of Cueva del Angel, Spain. Q. Geochronol. 49, 297–302. doi: 10.1016/j.quageo.2018.02.003
Feranec, R., García, N., Díez, J. C., and Arsuaga, J. L. (2010). Understanding the ecology of mammalian carnivorans and herbivores from Valdegoba cave (Burgos, northern Spain) through stable isotope analysis. Palaeogeol. Palaeoclimatol. Palaeoecol. 297, 263–272. doi: 10.1016/j.palaeo.2010.08.006
Feranec, R. S. (2007a). Ecological generalization during adaptive radiation: evidence from Neogene mammals. Evol. Ecol. Res. 9, 555–577.
Feranec, R. S. (2007b). Stable carbon isotope values reveal evidence of resource partitioning among ungulates from modern-3-dominated ecosystems in North America. Palaeogeol. Palaeoclimatol. Palaeoecol. 252, 575–585. doi: 10.1016/j.palaeo.2007.05.012
Ficcarelli, G., Abbazzi, L., Albianelli, A., Bertini, A., Coltorti, M., Magnatti, M., et al. (1997). Cesi, an early Middle Pleistocene site in the Colfiorito basin (Umbro-Marchean Apennine), central Italy. J. Q. Sci. 12, 507–518. doi: 10.1002/(SICI)1099-1417(199711/12)12:6<507::AID-JQS329>3.0.CO;2-E
Foronova, I. (2006). “Late quaternary equids (genus Equus) of South-Western and Southcentral Siberia,” in Equids in Time and Space, Papers in Honour of Véra Eisenmann, Proceedings of the 9th Conference of the International Council of Archaeozoology, ed M. Mashkour (Durham), 20–30.
Forsten, A. (1973). The evolution of some morphological characters of the upper cheek teeth of the fossil horses. Evolution 27, 36–43. doi: 10.1111/j.1558-5646.1973.tb05915.x
Forsten, A. (1986). A review of the Süssenborn horses and the origin of Equus hydruntinus Regalia. Quatärpalaläontologie 6, 43–52.
Forsten, A. (1988). Middle Pleistocene replacement of stenonid horses by caballoid horses – ecological implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 65, 23–33. doi: 10.1016/0031-0182(88)90109-5
Forsten, A. (1993). Size decrease in Late Pleistocene–Holocene caballoid horses (genus Equus), intra- or interspecific evolution? A discussion of alternatives. Q. Int. 19, 71–75. doi: 10.1016/1040-6182(93)90025-B
Forsten, A. (1996). The pleistocene horses from Achenheim, near Strasbourg (Alsace, France), and a comparison with contemporaneous finds, neues jahrbuch für Geologie und Paläontologie. Abhandlungen 201, 163–177. doi: 10.1127/njgpa/201/1996/163
Forsten, A. (1998a). The fossil horses (Equidae, Mammalia) from the Plio-Pleistocene of Liventsovka near Rostov-Don, Russia. Geobios 31, 645–657. doi: 10.1016/S0016-6995(98)80052-1
Forsten, A. (1998b). Equus species as stratigraphic markers, reality or wishful thinking? Q. Sci. Rev. 17, 1097–1100.
Forsten, A. (1999a). A review of Equus stenonis Cocchi (Perissodactyla, Equidae) and related forms. Q. Sci. Rev. 18, 1373–1408. doi: 10.1016/S0277-3791(98)00073-0
Forsten, A. (1999b). “The horses (genus Equus) from the Middle Pleistocene of Steinheim, Germany,” in Mammoths and the Mammoth Fauna: Studies of an Extinct Ecosystem, eds G. Haynes, J. Klimowicz, and J. W. F. Reumer (Germany: DEINSEA, 147–154.
Forsten, A., and Dimitrijevic, V. (2004). Pleistocene horses (genus Equus) in the central Balkans. Geoloski Anali Balkanskog Poluostrva 65, 55–75. doi: 10.2298/GABP0301055F
Forsten, A., and Moigne, A. M. (1998). The horse from the Middle Pleistocene of Orgnac-3 (Ardèche, France). Quaternaire 9, 315–323. doi: 10.3406/quate.1998.1613
Forsten, A., and Ziegler, R. (1995). The horses (Mammalia, Equidae) from the early Wuermian of Villa Seckendorff, Stuttgart-Bad Cannstadt, Germany. Stuttgarter Beiträge Naturkunde Ser. B. 224, 1–22.
Fortelius, M., and Solounias, N. (2000). Functional characterization of ungulate molars using the abrasion-attrition wear gradient: a new method for reconstructing paleodiets. Am. Museum Novitates 3301, 1–36. doi: 10.1206/0003-0082(2000)301<0001:FCOUMU>2.0.CO;2
Fourvel, J. B., Fosse, P., Fernandez, P., and Antoine, P. O. (2014). La grotte de Fouvent, dit l'Abri Cuvier (Fouvent-le-Bas, Haute-Saône, France) : analyse taphonomique d'un repaire du Pléistocène supérieur (OIS3). Paléo 25, 79–99
Freudenberg, W. (1911). Beiträge zur Gliederung des Quartärs von Weinheim a. d. Bergstrasse, Maure b. Heidelberg, Jockgrim i. d. Pfalz u. a. und seine Bedeutung für den Bau der oberrheinischen Tiefebene. Notizblatt des Vereins für Erdkunde und der Grossh. Geologischen Landesanstalt IV 32, 76–149.
García García, N., Feranec, R. S., Arsuaga, J. L., Bermúdez de Castro, J. M., and Carbonell, E. (2009). Isotopic analysis of the ecology of herbivores and carnivores from the Middle Pleistocene deposits of the Sierra de Atapuerca, northern Spain. J. Archaeol. Sci. 36, 1142–1151. doi: 10.1016/j.jas.2008.12.018
Garrido, G. (2008). “Los peridacáctilos [Equus cf. major Depéret, 1893 y Stephanorhinus etruscus (Falconer, 1859)] del yacimiento Plioceno Superior terminal de Fonelas P-1 (Cuenca de Guadix, Granada),” in Vertebrados del Plioceno Superior Terminal en el Suroeste de Europa: Fonelas P-1 y el Proyecto Fonelas Cuadernos del Museo Geominero, ed A. Arribas, 10, 553–595.
Gibert, L., Scott, G. R., Scholz, D., Budsky, A., Ferràndez, C., Ribot, F., et al. (2016). Chronology for the Cueva Victoria fossil site (SE Spain): evidence for Early Pleistocene afro-iberian dispersals. J. Hum. Evol. 90, 183–197. doi: 10.1016/j.jhevol.2015.08.002
Gkeme, A., Koufos, G. D., and Kostopoulos, D. S. (2017). “The Early Pleistocene stenonoid horse from Libakos (Western Macedonia, Greece): Biochronological and palaeoecological implications and dispesal events,” in 15th RCMNS Congress (Athens).
Gould, S. J. (1976). “Paleontology plus ecology as palaeobiology,” in Theoretical Ecology: Principles and Applications, ed R. M. May (New Jersey, NY: Blackwell Scientific Publications, 218–236.
Green, C. P., Coope, G. R., Currant, A. P., Holyoak, D. T., Ivanovich, M., Jones, R. L., et al. (1984). Evidence of two temperate episodes in Late Pleistocene deposits at Marsworth, UK. Nature 309, 778–781. doi: 10.1038/309778a0
Gregory, W. K. (1912). Notes on the principles of quadrupedal locomotion and on the mechanism of the limbs in hoofed animals. Ann. N. Y. Acad. Sci. 22, 267–294. doi: 10.1111/j.1749-6632.1912.tb55164.x
Griggo, C. (1995). Significations paléoenvironnementales des communautés animales pléistocènes reconnues dans l'abri Suard (Charente) et la grotte de bois-ragot (Vienne) : essai de quantification de variables climatiques (Thèse de Doctorat), Université de Bordeaux, Bordeaux, France.
Gromova, V. I. (1949). Istorija loshadej (roda Equus) v Starom Svete. Chast' 1. obzor i opisanie form. Trudy Paleontol. Inst. Akademii Nauk SSSR 17.
Gromova, V. I., and Dubrovo, I. A. (1975). Fossile Pferde aus dem faunenkomplex von Tiraspol. Quartärpaläontologie 1, 107–124.
Grossouvre, A., and Stehlin, H. G. (1912). Les sables de Rosieres, près Saint-Florent (Cher). Bull. Soc. Géol. France 4, 194–212.
Groves, C. P., and Willoughby, D. P. (1981). Studies on the taxonomy and phylogeny of the genus Equus. Mammalia 45, 321–354. doi: 10.1515/mamm.1981.45.3.321
Guadelli, J. L. (1987). Contribution à l'étude des zoocénoses préhistoriques en Aquitaine (Würm ancien et interstade würmien) (Thèse de doctorat). Université Bordeaux, Bordeaux, France.
Guadelli, J. L. (1991). Les chevaux de Solutré (Saône et Loire, France). Cahier Quater. 16, 261–336.
Guadelli, J. L. (1995).“Étude des équidés,” in La Grotte de Pégourié, Camiac-du-Causse, Lot. Préhistoire Quercinoise, Supplément, ed M. R. Seronie-Vivien, 99–148.
Guadelli, J. L. (2007). “Etude paléontologique des grands mammifères,” in Les Dossiers de l'Archéo-logis, 3, eds I, Sainte-Anne, Sinzelles, Polignac, Haute-Loire. Le Paléolithique moyen de l'unité (Laussonne: CDERAD, 163–188.
Guadelli, J. L., and Delpech, F. (2000). “Les grands mammiferes du debut du Paleolithique superieur a Temnata,” in Temnata Cave. Excavation in Karlukovo Karst Area, eds B. Ginter, J. K. Kozlowski, J. L. Guadelli, and H. Laville, 53–158.
Guadelli, J. L., and Prat, F. (1995). Le cheval du gisement Pléistocène moyen de Camp-de-Peyre (Sauveterre-la-Lémance, Lot-et-Garonne). Equus mosbachensis campdepeyri nov. ssp. Paléo 7, 85–121. doi: 10.3406/pal.1995.1209
Guerrero-Alba, S., and Palmqvist, P. (1997). Estudio morfométrico del caballo de Venta Micena (Orce, Granada) y su comparación con los équidos modernos y del Plio-Pleistoceno en el viejo y nuevo mundo. Paleontol. Evol. 30–31, 93–148.
Güleç, E., White, T., Kuhn, S. L., Özer, I., Sagr, M., Yilmaz, H., et al. (2009). The Lower Pleistocene lithic assemblage from Dursunlu (Konya), Central Anatolia, Turkey. Antiquity 83, 11–22. doi: 10.1017/S0003598X00098057
Guthrie, R. D. (1984). “Mosaics, allelochemics and nutrients, an ecological theory of Late Pleistocene megafaunal extinctions,” in Quaternary Extinctions, A Prehistoric Revolution, eds P. S. Martin and R. G. Klein (Tucson: University of Arizona Press, 259–298.
Guthrie, R. D. (2003). Rapid body size decline in alaskan Pleistocene horses before extinction. Nature 426, 169–171. doi: 10.1038/nature02098
Hanot, P. (2018). Morphological diversity of modern and past domestic equids: complete skeletons as a marker of function and cultural practices (Unpublished PhD thesis). Muséum National d'Histoire Naturelle, Paris, France.
Heaton, T. H. E. (1999). Spatial, species and temporal variations in the 13C/12C ratios of C3 plants: implications for palaeodiet studies. J. Archaeol. Sci. 26, 637–649. doi: 10.1006/jasc.1998.0381
ICZN (International Commission on Zoological Nomenclature) (2003). Opinion 2027 (Case 3010), usage of 17 specific names based on wild species which are pre-dated by or contemporary with those based on domestic animals (Lepidoptera, Osteichthyes, Mammalia): conserved. Bull. Zool. Nomencl. 60, 81–84.
James, F. C. (1970). Geographic size variation in birds and its relationship to climate. Ecology 51, 365–390. doi: 10.2307/1935374
Janis, C. M. (1990). Correlation of cranial and dental variables with dietary preferences in mammals: a comparison of macropodoids and ungulates. Mem. Queensl. Museum 28, 349–366.
Jonsson, H., Schubert, M, Seguin-Orlando, A., Ginolhac, A., Petersen, L., et al. (2014). Speciation with gene flow in equids despite extensive chromosomal plasticity. Proc. Natl. Acad. Sci. U.S.A. 111, 18655–18660. doi: 10.1073/pnas.1412627111
Kaagan, L. M. (2000). The horse in Late Pleistocene and Holocene Britain (Thesis). University of London, London, United Kingdom.
Kahlke, R. D. (2006). Untermassfeld - a Late Early Pleistocene (Epivillafranchian) Fossil Site Near Meiningen (Thuringia, Germany) and its Position in the Development of the European Mammal Fauna. British Archaeological Reports, International Series 1578. Oxford: Archaeopress.
Kahlke, R. D. (2014). The origin of eurasian mammoth faunas (mammuthus–coelodonta faunal complex). Q. Sci. Rev. 96, 32–49. doi: 10.1016/j.quascirev.2013.01.012
Kahlke, R. D., Garcia, N., Kostopoulos, D., Lacombat, F., Lister, A., Mazza, P., et al. (2011). Western palaearctic palaeoenvironmental conditions during the early and early middle pleistocene inferred from large mammal faunal communities, and implications for hominin dispersal in Europe. Q. Sci. Rev. 30, 1368–1395. doi: 10.1016/j.quascirev.2010.07.020
Kaiser, T. M., and Franz-Odendaal, T. A. (2004). A mixed-feeding Equus species from the Middle Pleistocene of South Africa. Q. Res. 62, 316–323. doi: 10.1016/j.yqres.2004.09.002
Konidaris, G. E., Tourloukis, V., Kostopoulos, D. S., Thompson, N., Giusti, D., Michailidis, D., et al. (2015). Two new vertebrate localities from the Early Pleistocene of Mygdonia basin (Macedonia, Greece): preliminary results. Comp. Rendus Palevol. 14, 353–362. doi: 10.1016/j.crpv.2015.05.004
Koufos, G. D. (1992). Early Pleistocene equids from Mygdonia basin (Macedonia, Greece). Palaeontogr. Ital. 79, 167–199.
Koufos, G. D. (2001). The Villafranchian mammalian faunas and biochronology of Greece. Boll. Soc. Paleontol. Ital. 40, 217–223.
Koufos, G. D., and Kostopoulos, D. S. (2016). “The pliocene large mammal record in Greece: implication for early human dispersal into Europe,” in Palaeoanthropology of the Balkans and Anatolia: Human Evolution and its Context, eds K. Harvati and M. Roksandis (Dortrecht: Springer, 269–280.
Koufos, G. D., Kostopoulos, D. S., and Sylvestrou, I. A. (1997). Equus apolloniensis n. sp. (Mammalia, Equidae) from the latest Villafranchian locality of Apollonia, macedonia, Greece. Paleontologia i Evolucio 30–31, 49–76.
Kuzmina, I. E. (1997). “Horses of North Eurasia from the Pliocene till the present time, Russian academy of Sciences,” in Proceedings of the Zoological Institute 273, ed N. K. Vereschagin (Saint-Petersburg) [in Russian with English summary].
Kuznetsova, T. V., and Zhegallo, V. I. (2009). Equus (Hemionus) nalaikhaensis from the Pleistocene of Mongolia, the Earliest Kulan of Central Asia. Paleontol. J. 43, 574–583. doi: 10.1134/S0031030109050141
Laban, C., and Van der Meer, J. J. M. (2004). “Pleistocene glaciation in The Netherlands,” in Quaternary Glaciations, Extent and Chronology, Part I Europe, Developments in Quaternary Science. eds J, Ehlers and P. L. Gibbard (Amsterdam: Elsevier, 251–260.
Lacombat, F. (2005). Les grands mammifères fossiles du Velay. les collections paléontologiques du Plio-Pléistocène du musée crozatier, Le Puy-en-Velay. Ann. Amis Musée Crozatier. 13–14, 208.
Langlois, A. (2005). Le cheval du gisement Pleistocene Moyen de La Micoque (Les Eyzies-de-Tayac, Dordogne). Equus mosbachensis micoquii nov. ssp. Paléo. 17, 73–110.
Lazarev, P. A. (1980). Antropogenovye Loshadi Iakutii. [= Anthropogene Horses of Iakutia]. Moscow: Nauka.
Lebatard, A. E., Alçiçek, M. C., Rochette, P., Khatib, S., Vialet, A., Boulbes, N., et al. (2014). 1.2-million-year-old Homo cranium from kocabaş (Denizli, Turkey). Earth Planet. Sci. Lett. 398, 8–18. doi: 10.1016/j.epsl.2013.12.031
Lindsay, E. H., Opdyke, N. D., and Johnson, N. M. (1980). Pliocene dispersal of the horse Equus and Late Cenozoic mammalian dispersal events. Nature 287, 135–138. doi: 10.1038/287135a0
Lindstedt, S. L., and Boyce, M. S. (1985). Seasonality, fasting endurance, and body size in mammals. Am. Nat. 125, 873–787. doi: 10.1086/284385
Lister, A. M., Parfitt, S. A., Owen, F. J., Collinge, S. E., and Breda, M. (2010). Metric analysis of ungulate mammals in the early Middle Pleistocene of Britain, in relation to taxonomy and biostratigraphy: II: cervidae, equidae and suidae. Q. Int. 228, 157–179. doi: 10.1016/j.quaint.2010.05.014
Lordkipanidze, D., Jashashvili, T., Vekua, V., Ponce de Leon, M. S., Zollikofer, C. P. E., Rightmire, G. P., et al. (2007). Postcranial evidence from early Homo from Dmanisi, Georgia. Nature 449, 305–310. doi: 10.1038/nature06134
Lozano-Fernández, I., Blain, H. A., Agustí, J., Piñero, P., Barsky, D., Ivorra, J., et al. (2019). New clues about the late Early Pleistocene peopling of western Europe: small vertebrates from the Bois-de-Riquet archeo-paleontological site (Lézignan-La-Cèbe, southern France). Q. Sci. Rev. 219, 187–203. doi: 10.1016/j.quascirev.2019.07.015
Lumley, H., Fontaneil, C., Grégoire, S., Batalla, G., Caumon, G., Celiberti, V., et al. (2015). Caune de l'arago. Tautavel-en-Roussillon, Pyrénées-Orientales, France. Tome VI. Individualisation Des Unités Archéostratigraphiques. Paris: CNRS Éditions, 641.
MacFadden, B. J. (1988). Fossil horses from “Eohippus” (Hyracotherium) to Equus, 2: rates of dental evolution revisited. Biol. J. Linnean Soc. 35, 37–48. doi: 10.1111/j.1095-8312.1988.tb00457.x
Madurell-Malapeira, J., Minwer-Barakat, R., Alba, D. M., Garcés, M., Gómez, M., Aurell-Garrido, J., et al. (2010). The Vallparadís section (Terrassa, Iberian peninsula) and the latest Villafranchian faunas of Europe. Q. Sci. Rev. 29, 2972–2982. doi: 10.1016/j.quascirev.2010.09.020
Magniez, P., and Boulbes, N. (2014). Environmental at the transition Middle and Late Paleolithic in south of France. The archeosequence of the Tournal cave (Bize-Minervois, France). Q. Int. 337, 43–63. doi: 10.1016/j.quaint.2013.08.021
Magniez, P., Boulbes, N., and Brugal, J. P. (2017). “Variations de la taille corporelle chez les ongulés pléistocènes: implications paléoécologiques et taphonomiques,” in TaphonomieS, ed J. P. Brugal (Paris: Éditions des archives contemporaines).
Marín, M. (1987). Equus stenonis granatensis en el Pleistoceno Inferior de Venta Micena (Granada, España). Paleontol. Evol. Mem. Esp. 1, 255–282.
Marra, A. C., and Bellomo, E. (2008). Equus hydruntinus from the Upper Pleistocene (MIS 4) of calabria, Southern Italy. Boll. Soc. Paleontol. Ital. 47, 1–6.
Marra, F., Nomade, S., Pereira, A., Petronio, C., Salari, L., Sottili, G., et al. (2018). A review of the geologic sections and the faunal assemblages of Aurelian mammal age of Latium (Italy) in the light of a new chronostratigraphic framework. Q. Sci. Rev. 181, 173–199. doi: 10.1016/j.quascirev.2017.12.007
Marra, F., Pandolfi, L., Petronio, C., Di Stefano, G., Gaeta, M., and Salari, L. (2014). Reassessing the sedimentary deposits and vertebrate assemblages from Ponte Galeria area (Roma, central Italy): an archive for the Middle Pleistocene faunas of Europe. Earth Sci. Rev. 139, 104–122. doi: 10.1016/j.earscirev.2014.08.014
Martín Penela, A. J. (1987). Los grandes mamíferos del yacimiento Achelense de La Solana de Zamborino, Fonelas (Granada, España). Antrop. Paleoec. Hum. 5, 29–188.
Martínez, K., Garcia, J., Burjachs, F., Yll, R., and Carbonell, E. (2014). Early human occupation of Iberia: the chronological and palaeoclimatic inferences from Vallparadís (Barcelona, Spain). Q. Sci. Rev. 85, 136–146. doi: 10.1016/j.quascirev.2013.12.002
Martínez-Navarro, B., Ros-Montoya, S., Espigares, M. P., and Palmqvist, P. (2011). Presence of Asian Origin Bovini, Hemibos sp. aff. H. gracilis and Bison sp., at the Early Pleistocene site of Venta Micena (Orce, Spain). Q. Int. 243, 54–60. doi: 10.1016/j.quaint.2011.05.016
Mauser, M. (1987). Geologische und Palaeontologische Untersuchungen an der altpleistozänen SäugetierFundstelle Würzburg-Schalksberg. Münchner Geowissenschaftliche Abhandlungen Reihe A 11:77.
Mayr, E. (1956). Geographical character gradients and climatic adaptation, Evolution 10, 105–108. doi: 10.1111/j.1558-5646.1956.tb02836.x
Meadow, R. H. (1999). “The use of size index scaling techniques for research on archaeozoological collections from the middle east,” in Historia Animalium ex Ossibus: Beiträge aur Paläoanatomie, Archäologie, Ägyptologie, Ethnologie und Geschichte der Tiermedizin, eds C. Becker, H. Manhart, J. Peters, and J. Schibler (Rahden: Leidorf, 285–300.
Menard, C., Duncan, P., Fleurance, G., Georges, J. Y., and Lila, M. (2002). Comparative foraging and nutrition of horses and cattle in European wetlands. J. Appl. Ecol. 39, 120–133. doi: 10.1046/j.1365-2664.2002.00693.x
Moigne, A. M., Palombo, M. R., Belda, V., Heriech-Briki, D., Kacimi, S., Lacombat, F., et al. (2006). Les faunes de grands mammifères de la Caune de l'Arago (Tautavel) dans le cadre biochronologique des faunes du Pléistocène Moyen Italien. L'Anthropologie 110, 788–831. doi: 10.1016/j.anthro.2006.10.011
Moncel, M.-H., Moigne, A.-M., and Combier, J. (2005). Pre-neandertal behaviour during isotopic stage 9 and the beginning of stage 8, new data concerning fauna and lithics in the different occupation levels of Orgnac 3 (Ardèche, South-East France): occupation types. J. Archaeol. Sci. 32, 1283–1301. doi: 10.1016/j.jas.2005.03.014
Mourer-Chauviré, C. (1972). Etude de nouveaux restes de vertébrés provenant de la carrière Fournier à Châtillon-Saint-Jean. III. Artiodactyles, chevaux, oiseaux. Bull. l'Assoc. Franç. L'étude Quater. 9, 271–305. doi: 10.3406/quate.1972.1209
Mourer-Chauviré, C. (1980). Le gisement pléistocène supérieur de la grotte de Jaurens, à Nespouls, Corrèze, France : les Equidés (Mammalia, Perissodactyla). Nouvel. Arch. Muséum d'Hist. Nat. Lyon 18, 17–60.
Mourer-Chauviré, C., Philippe, M., Quinif, Y., Chaline, J., Debard, E., Guérin, C., et al. (2003). Position of the palaeontological site Aven I des Abîmes de La Fage, at Noailles (Corrèze, France), in the European Pleistocene chronology. Boreas 32, 521–531. doi: 10.1080/03009480301811
Murton, J. B., Baker, A., Bowen, D. Q., Caseldine, C. J., Coope, G. R., Currant, A. P., et al. (2001). A late Middle Pleistocene temperate-periglacial-temperate sequence (Oxygen Isotope Stages 7-5e) near Marsworth, Buckinghamshire, UK. Q. Sci. Rev. 20, 1787–1825. doi: 10.1016/S0277-3791(01)00004-X
Musil, R. (1965). Die Equidenreste aus dem Pleistozän von Voigstedt. Paläontologische abhandlungen. Abt. A Paläozool. 3, 435–449.
Musil, R. (1969). Die Equidenreste aus dem Pleistozän von Süssenborn bei Weimar. Paläontologische abhandlungen. Abt. A Paläozool. 1969, 617–666.
Musil, R. (1975). “Die equiden aus dem travertin von Ehringsdorf,” in Das Pleistozän von Weimar-Ehringsdorf, Teil II, Abhandlungen des Zentralen Geologischen Instituts, Paläontologische Abhandlungen, ed H -D. Kahlke (Berlin), 265–335.
Musil, R. (1977). Die Equidenreste aus den travertinen von Taubach. Quartärpaläontologie 2, 237–264.
Musil, R. (1978). Die fossilen equiden-reste aus den travertinen von Burgtonna in Thüringen. Quartärpaläontologie 3, 137–138.
Musil, R. (1990). Pferdefunde (Equus taubachensis und Equus scythicus) aus der Kulna- Höhle in Mähren. Weimar: Museum für Ur- und Frühgeschichte Thüringens.
Musil, R. (1991). Pferde aus Bilzingsleben, in Bilzingsleben IV, Homo erectus, Seine Kultur Und Seine Umwelt, eds K. E. W. Fischer, W-D. Guenther, D. Heinrich, R, Mania, R. Musil, and T. Nötzold (Berlin: Landesamt für Denkmalpflege und Archäologie Sachsen-Anhalt, 103–130.
Musil, R. (1992). Die Pferde aus der Oberpliozänen spaltenfüllung Schernfeld bei eichstätt. MitteilungenBayerisch. Staatssamml. Paläontol. Historische Geol. 32, 115–162.
Musil, R. (1995). Large fauna of Talus cones at the stránská Skála Hill. in Stránská Skála Hill, Excavation of Open-Air Sediments 1964–1972, ed R. Musil (Brno: Moravian Museum; Anthropos 26, 65–83.
Musil, R. (2001). “Die equiden-reste aus dem Unterpleistozän von Untermaßfeld. in Das Pleistozän von Untermassfeld bei Meiningen (Thüringen). Teil 2. Monographien des Römisch-Germanischen Zentralmuseums Mainz 40, ed R. D. Kahlke (Bonn: Dr. Rudolf Habelt, 557–587.
Nehring, A. (1884). Fossile Pferde aus Deutschen Diluvial-Ablagerungen und ihre Beziehungen zu den lebenden Pferden: Ein Beitrag zur Geschichte des Hauspferdes, Vol. 13. Landwirtschaftliches Jahrbuch, 81–160.
Nobis, G. (1971). Vom Wildpferd zum Hauspferd, Studien zur Phylogenie Pleistozäner Equiden Eurasiens und das Domestikationsproblem unserer Hauspferde, Fundamenta Reihe B, Band 6. Köln: Bõhlau Verlag.
Nomade, S., Pastre, J. F., Guillou, H., Faure, M., Guérin, G., Delson, E., et al. (2014). 40Ar/39Ar constraints on some French landmark Late Pliocene to Early Pleistocene large mammalian paleofauna: paleoenvironmental and paleoecological implications. Q. Geochronol. 21, 2–15. doi: 10.1016/j.quageo.2012.12.006
Nores, C., Morales, M. A., Llorente Rodríguez, L., Bennett, E. A., and Geigl, E. M. (2015). The Iberian zebro: what kind of a beast was it? Anthropozoologica 50, 21–32. doi: 10.5252/az2015n1a2
Orlando, L., Ginolhac, A., Zhang, G., Froese, D., Albrechtsen, A., Stiller, M., et al. (2013). Recalibrating Equus evolution using the genome sequence of an early Middle Pleistocene horse. Nature 499, 74–78. doi: 10.1038/nature12323
Orlando, L., Mashkour, M., Burke, A., Douady, J. C., Eisenmann, V., and Hänni, C. (2006). Geographic distribution of an extinct equid (Equus hydruntinus: Mammalia, Equidae) revealed by morphological and genetical analyses of fossils. Mol. Ecol. 15, 2083–2093. doi: 10.1111/j.1365-294X.2006.02922.x
Orlando, L., Metcalf, J. L., Alberdi, M. T., Telles-Antunes, M., Bonjean, D., Otte, M., et al. (2009). Revising the recent evolutionary history of equids using ancient DNA. Proc. Natl. Acad. Sci. U.S.A. 106, 21754–21759. doi: 10.1073/pnas.0903672106
Palombo, M. R. (2014). Deconstructing mammal dispersals and faunal dynamics in SW europe during the quaternary. Q. Sci. Rev. 96, 50–71. doi: 10.1016/j.quascirev.2014.05.013
Palombo, M. R., and Alberdi, M. T. (2017). Light and shadows in the evolution of South European stenonoid horses. Fossil Imprint 73, 115–140. doi: 10.2478/if-2017-0006
Palombo, M. R., Alberdi, M. T., Bellucci, L., and Sardella, R. (2017). An intriguing middle-sized horse from Coste San Giacomo (Anagni Basin, central Italy). Q. Res. 87, 347–362. doi: 10.1017/qua.2017.6
Parfitt, S. A., Owen, F., and Keen, D. H. (1998). “Pleistocene stratigraphy, vertebrates and mollusca, black rock, Brighton,” in The Quaternary of Kent and Sussex, Field Guide. Quaternary Research Association The Quaternary of Kent and Sussex, Field Guide, eds J. B. Murton, C. A. Whiteman, M. R. Bates, D. R. Bridgland, A. J. Long, M. B. Roberts, et al. (London: Quaternary research association, 146–150.
Pastre, J. F., Debard, E., Nomade, S., Guillou, H., Faure, M., Guérin, C., et al. (2015). Nouvelles données géologiques et téphrochronologiques sur le gisement paléontologique du maar de Senèze (Pléistocène inférieur, Massif Central, France). Quaternaire 26, 225–244. doi: 10.4000/quaternaire.7368
Pereira, A., Nomade, S., Voinchet, P., Bahain, J. J., Falguères, C., and Garon, H. (2015): The earliest securely dated hominin fossil in Italy evidence of acheulian occupation during glacial MIS 16 at Notarchirico (Venosa, Basilicata, Italy). J. Q. Sci. 30, 639–650. doi: 10.1002/jqs.2809.
Pettitt, P. B., and White, M. J. (2012). The British Palaeolithic: Human Societies at the Edge of the Pleistocene World. London: Routledge.
Piñero, P., and Alberdi, M. T. (2015). Estudio de los caballos del yacimiento de Quibas, Pleistoceno inferior final (Abanilla, Murcia, España). Estudios Geológicos 71:e034. doi: 10.3989/egeol.41863.348
Prat, F. (1964). Contribution à la classification des Equidés villafranchiens. P.v. Soc. Linn. Bordeaux 101, 14–32.
Prat, F. (1968). Recherche sur les équides pléistocènes en France (Thèse de doctorat). Université de Bordeaux, Bordeaux, France.
Prat, F. (1977). L'Equidé du gisement acheuleen de Torralba (Soria, Espagne). INQUA. Suppl. Bull. AFEQ 50, 33–46.
Prat, F. (1980). Les Equidés Villafranchiens en France. Genre Equus. Centre National de la Recherche Scientifique, Centre régional de publication de Bordeaux. Cahiers du Quaternaire 2:290.
Prat, F., and Thibault, C. (1976). Le gisement de Nauterie à la Romieu (Gers). Fouilles de 1967 à 1973, Nauterie I. Mémoires du Muséum National d'Histoire Naturelle, Ssérie C, t. 35 (Paris: Éditions du Muséum), 96.
Preece, R. C., and Parfitt, S. A. (2008). “The cromer-forest bed formation: some recent developments relating to early human occupation and lowland glaciation,” in The Quaternary of Northern East Anglia. Field Guide, eds I. Candy, J. R. Lee and A. M. Harrison (London: Quaternary Research Association), 60–83.
Privat Defaus, J. (1986). The Villafranchian Equines from the Upper Valdarno. Riv. It. Paleont. Strat. 91, 519–536.
Pushkina, D., Bocherens, H., and Ziegler, R. (2014). Unexpected palaeoecological features of the Middle and Late Pleistocene large herbivores in southwestern Germany revealed by stable isotopic abundances in tooth enamel. Q. Int. 339–340, 164–178. doi: 10.1016/j.quaint.2013.12.033
Radulesco, C., and Samson, P. (1965). Sur la présence de Hydruntinus hydruntinus (Regalia) en roumanie. Quaternaria 7, 219–234.
Regalia, E. (1907). Sull'Equus (Asinus) hydruntinus Regalia della grotta di Romanelli (Castro, Lecce). Arch. Antropol. Etnol. 37, 375–390.
Reille, M., De Beaulieu, J.-L., Svobodova, H., Andrieu-Ponel, V., and Goeury, D. (2000). Pollen analytical biostratigraphy of the last five climatic cycles from a long continental sequence from the Velay region (Massif Central, France). J. Q. Sci. 15, 665–685. doi: 10.1002/1099-1417(200010)15:7<665::AID-JQS560>3.0.CO;2-G
Reynaud Savioz, N., and Morel, P. (2005). La faune de Nadaouiyeh Aïn Askar (Syrie centrale, Pléistocène moyen): aperçu et perspectives. Rev. Paléobiol. 10, 31–35.
Rivals, F., Julien, M.-A., Kuitems, M., Van Kolfschoten, T., Serangeli, J., Drucker, D., et al. (2015). Investigation of equid paleodiet from Schöningen 13II-4 through dental wear and isotopic analyses: archaeological implications. J. Hum. Evol. 89, 129–137. doi: 10.1016/j.jhevol.2014.04.002
Rivals, F., and Lister, A. M. (2016). Dietary flexibility and niche partitioning of large herbivores through the Pleistocene of Britain. Q. Sci. Rev. 146, 116–133. doi: 10.1016/j.quascirev.2016.06.007
Rivals, F., Mihlbachler, M. C., and Solounias, N. (2007). Effect of ontogenetic-age distribution in fossil samples on the interpretation of ungulate paleodiets using the mesowear method. J. Vertebrate Paleontol. 27, 763–767. doi: 10.1671/0272-4634(2007)27[763:EOODIF]2.0.CO;2
Rivals, F., Schulz, E., and Kaiser, T. M. (2008). Climate-related dietary diversity of the ungulate faunas from the Middle Pleistocene succession (OIS 14-12) at the Caune de l'Arago (France). Paleobiology 34, 117–127. doi: 10.1666/07023.1
Rivals, F., Schulz, E., and Kaiser, T. M. (2009). Late and Middle Pleistocene ungulate dietary diversity in western europe indicate variations of Neanderthal paleoenvironments through time and space. Q. Sci. Rev. 28, 3388–3400. doi: 10.1016/j.quascirev.2009.09.004
Rodríguez, J., Burjachs, F., Cuenca-Bescós, G., García, N., Van der Made, J., Pérez González, A., et al. (2011). One million years of cultural evolution in a stable environment at Atapuerca (Burgos, Spain). Q. Sci. Rev. 30, 1396–1412. doi: 10.1016/j.quascirev.2010.02.021
Rook, L., Croitor, R., Delfino, M., Ferretti, M., Gallai, G., and Pavia, M. (2013). The Upper Valdarno Plio-Pleistocene vertebrate record: an historical overview, with notes on palaeobiology and stratigraphic significance of some important taxa. Italian J. Geosci. 132, 104–125. doi: 10.3301/IJG.2012.16
Rusch, L., Boulbes, N., Lartigot-Campin, A. S., Testu, A., Pois, V., Bahain, J. J., et al. (2019). Contexte paléoenvironnemental et chronologique des occupations néandertaliennes de la grotte des Ramandils (Port-la-Nouvelle, Aude, France): apport des restes de grands mammifères. Quaternaire 30, 151–165. doi: 10.4000/quaternaire.11717
Saarinen, J., Eronen, J., Fortelius, M., Seppä, H., and Lister, A. M. (2016). Patterns of diet and body mass of large ungulates from the Pleistocene of Western Europe, and their relation to vegetation. Palaeontol. Electron. 32A, 1–58. doi: 10.26879/443
Saladié, P., Rodríguez-Hidalgo, A., Marín, J., Vallverdú i Poch, J., and Carbonell, E. (2018). The top of the Gran Dolina (Atapuerca, Spain) sequence: a zooarchaeological and occupational perspective. Q. Sci. Rev. 195, 48–71. doi: 10.1016/j.quascirev.2018.07.010
Salter, R. E., and Hudson, R. J. (1979). Feeding ecology of feral horses in western Alberta. J. Range Manage. 32, 221–225. doi: 10.2307/3897127
Samson, P. (1975). Les équidés fossiles de Roumanie (Pliocène moyen - Pléistocène supérieur). Geol. Romana 14, 165–354.
Saos, T., Grégoire, S., Bahain, J. J., Higham, T., Moigne, A. M., Testu, A., et al. (2019). The Middle and Upper Palaeolithic of la Crouzade cave (Gruissan, Aude, France): new excavations and a chronostratigraphic framework. Session Q26 XVI:peopling dynamics in the Mediterranean area between 45 and 39 ky ago: state of art and new data. Q. Int.
Sardella, R., Mazzini, I., Giustini, F., Mecozzi, B., Brilli, M., Iurino, D. A., et al. (2018). Grotta Romanelli (Southern Italy, Apulia): legacies and issues in excavating a key site for the Pleistocene of the Mediterranean. Riv. It. Paleontol. Strat. 124, 247–264. doi: 10.13130/2039-4942/9983
Sarrión Montañana, I. (2008). Presencia de “Homo” en el Pleistoceno inferior y Medio Valenciano. Archivo Prehistoria Levantina 27, 9–24.
Schreve, D. C. (1997). Mammalian biostratigraphy of the later middle pleistocene in Britain (Unpublished PhD thesis). University of London, London, United Kingdom.
Schreve, D. C. (2001). Mammalian evidence from Middle Pleistocene fluvial sequences for complex environmental change at the oxygen isotope substage level. Q. Int. 79, 65–74. doi: 10.1016/S1040-6182(00)00123-3
Schulz, E., and Kaiser, T. M. (2013). Historical distribution, habitat requirements and feeding ecology of the genus Equus (Perissodactyla). Mammal Rev. 43, 111–123. doi: 10.1111/j.1365-2907.2012.00210.x
Sesé, C. Y., and Soto, E. (2005). Mamíferos del yacimiento del Pleistoceno medio de Ambrona (Soria, España): análisis faunístico e interpretación paleoambiental (2005). En Los yacimientos paleoliticos de Torralba y Ambrona (Soria). Un siglo de investigaciones arqueologicas. Zona Arqueol. 5, 258–280.
Shackleton, N. J. (1987). Oxygen isotopes, ice volume and sea level. Q. Sci. Rev. 6, 183–190. doi: 10.1016/0277-3791(87)90003-5
Sher, A. V. (1971). Mlekopitaiushchie i Stratigrafia Pleistotsena Krainego Severo-Vostoka SSSR i Severnoi Ameriki. Nauka, Moskva.
Sinusía, C., Pueya, E. L., Azanza, B., and Pocoví, A. (2004). Datación magnetoestratigráfica del yacimiento paleontológico de la Puebla de Valverde (Teruel). Geotemas 6, 339–342.
Sirakov, N., Guadelli, J. L., Ivanova, S., Sirakova, S., Boudadi-Maligne, M., Dimitrova, I., et al. (2010). An ancient continuous human presence in the Balkans and the beginnings of the settlement of western Eurasia. The lower Pleistocene example of lower Palaeolithic in Kozarnika cave (North-western Bulgaria). Q. Int. 223–224, 94–106. doi: 10.1016/j.quaint.2010.02.023
Solounias, N., and Semprebon, G. (2002). Advances in the reconstruction of ungulate ecomorphology with application to early fossil equids. Am. Museum Novitates. 3366, 1–49. doi: 10.1206/0003-0082(2002)366<0001:AITROU>2.0.CO;2
Spassov, N., and Iliev, N. (1997). “The wild horses of eastern Europe and the polyphyletic origin of the domestic horse,” in Proceedings of the 7th ICAZ Conference (Konstanz, 26/09/1994-01/10/1994), Vol. 25–26, eds M. Kokabi and J. Wahl (Paris: L'Homme et l'Animal, Société de Recherche Interdisciplinaire), 753–760.
Spassov, N., and Iliev, N. (2002). “The animal bones from the prehistoric necropolis near Durankulak (NE Bulgaria) and the latest record of Equus hydruntinus Regalia,” in Durankulak II ed H. Todorova, 313–324.
Stehlin, H.-G., and Graziosi, P. (1935). Ricerche sugli Asinidi fossili d'Europa. Mémoires Soc. Paléontol. Suisse 56, 1–73.
Stevens, R. E., and Hedges, R. E. M. (2004). Carbon and nitrogen stable isotope analysis of northwest European horse bone and tooth collagen, 40,000 BP-present: palaeoclimatic interpretations. Q. Sci. Rev. 23, 977–991. doi: 10.1016/j.quascirev.2003.06.024
Stevens, R. E., O'Connell, T. C., Hedges, R. E. M., and Street, M. (2009). Radiocarbon and stable isotope investigations at the central Rhineland sites of Gönnersdorf and Andernach-martinsberg, Germany. J. Hum. Evol. 57, 131–148. doi: 10.1016/j.jhevol.2009.01.011
Strani, F., DeMiguel, D., Alba, D. M., Moyà-Solà, S., Bellucci, L., Sardella, R., et al. (2019). The effects of the “0.9 Ma event” on the mediterranean ecosystems during the early-middle pleistocene transition as revealed by dental wear patterns of fossil ungulates. Q. Sci. Rev. 210, 80–89. doi: 10.1016/j.quascirev.2019.02.027
Thomason, J. J. (1986). The functional morphology of the manus in the tridactyl equids Merychippus and Mesohippus. J. Verteb. Paleontol. 6, 143–161.
Tilkens, M. J., Wall-Scheffler, C., Weaver, T. D., and Steudel-Numbers, K. (2007). The effects of body proportions on thermoregulation: an experimental assessment of Allen's rule. J. Hum. Evol. 53, 286–291. doi: 10.1016/j.jhevol.2007.04.005
Tsoukala, E. (1991). Contribution to the study of the Pleistocene fauna of large mammals (Carnivora, Perissodactyla, Artiodactyla) from Petralona cave, Chalkidiki (N. Greece). Prelim. Rep. 312, 331–336.
Tsoukala, E., and Guérin, C. (2016). The Rhinocerotidae and Suidae of the Middle Pleistocene from Petralona cave (Macedonia, Greece). Acta Zool. Bulgarica 68, 243–264.
Uerpmann, H. (1987). The ancient distribution of ungulate mammals in the Middle East. Naturwissenschaften 27, 1–173.
Uzunidis, A. (2017). Grands herbivores de la fin du Pléistocène moyen au début du Pléistocène supérieur dans le sud de la France. Implications anthropologiques pour la lignée néandertalienne (Thèse de doctorat). Aix-Marseille Université, Marseille, France.
Uzunidis, A., Rivals, F., and Brugal, J.-P. (2017). Relation between morphology and dietary traits in horse jugal upper teeth during the Middle Pleistocene in southern France. Quaternaire 28, 303–312. doi: 10.4000/quaternaire.8256
Valensi, P., and Boulbes, N. (2018). The magdalenian fauna from Roc-aux-Sorciers, a sculpted rock shelter (Vienne, France). Main archaeozoological results. Quaternaire 29, 75–80. doi: 10.4000/quaternaire.8639
Valli, A. M. F., Palombo, M. R., and Alberdi, M. T. (2012). How homogeneous are microwear patterns on a fossil horse tooth? preliminay test on a premolar of Equus altidens from Barranco Leon 5 (Spain). Alpine Mediterran. Q. 25, 25–33.
van Asperen, E. N. (2009a). An outline of late Middle Pleistocene horse biostratigraphy, a contribution to the chronology and ecology of the European Early and Middle Palaeolithic. University of York, York (Unpublished PhD thesis).
van Asperen, E. N. (2009b). Caballoid horses and late Middle Pleistocene biostratigraphy of the British Isles. Quaternaire 20, 437–448. doi: 10.4000/quaternaire.5309
van Asperen, E. N. (2010). Ecomorphological adaptations to climate and substrate in late middle Pleistocene caballoid horses. Palaeogeogr. Palaeoclimatol. Palaeoecol. 297, 584–596. doi: 10.1016/j.palaeo.2010.09.007
van Asperen, E. N. (2011). Distinguishing between the late Middle Pleistocene interglacials of the British isles: a multivariate approach to horse biostratigraphy. Q. Int. 231, 110–115. doi: 10.1016/j.quaint.2010.06.029
van Asperen, E. N. (2012). Late Middle Pleistocene horse fossils from northwestern Europe as biostratigraphic indicators. J. Archaeol. Sci. 39, 1974–1983. doi: 10.1016/j.jas.2012.02.025
van Asperen, E. N. (2013a). Position of the Steinheim interglacial sequence within the marine oxygen isotope record based on mammal biostratigraphy. Q. Int. 292, 33–42. doi: 10.1016/j.quaint.2012.10.045
van Asperen, E. N. (2013b). Implications of age variation and sexual dimorphism in modern equids for Middle Pleistocene equid taxonomy. Int. J. Osteoarchaeol. 23, 1–12. doi: 10.1002/oa.1231
van Asperen, E. N., Stefaniak, K., Proskurnyak, L., and Ridush, B. (2012). Equids from Emine-Bair-Khosar cave (Crimea, Ukraine): co-occurrence of the stenonid Equus hydruntinus and the caballoid E. ferus latipes based on skull and postcranial remains. Palaeontol. Electron. 15:28. doi: 10.26879/280
Van der Made, J. (1999). Ungulates from Atapuerca TD6. J. Hum. Evol. 37, 389–413. doi: 10.1006/jhev.1998.0264
Van der Made, J. (2013). First description of the large mammals from the locality of Penal, and updated faunal lists for the Atapuerca ungulates - Equus altidens, Bison and human dispersal into Western Europe. Q. Int. 295, 36–47. doi: 10.1016/j.quaint.2012.03.001
Van der Made, J., Rosell, J., and Blasco, R. (2017). Faunas from Atapuerca at the Early-Middle Pleistocene limit: the ungulates from level TD8 in the context of climatic change. Q. Int. 433, 296–346. doi: 10.1016/j.quaint.2015.09.009
Van Kolfschoten, T., Buhrs, E., and Verheijen, I. (2015). The larger mammal fauna from the Lower Paleolithic Schöningen spear site and its contribution to hominin subsistence. J. Hum. Evol. 89, 138–153. doi: 10.1016/j.jhevol.2015.09.014
Vekua, A. K. (1986). The Lower Pleistocene mammalian fauna of Akhalkalaki (Southern Georgia, USSR). Palaeontogr. Ital. 74, 63–96.
Viret, J. (1954). Le lœss à bancs durcis de Saint-Vallier (Drôme) et sa faune de mammiferes villafranchiens. Nouv. Arch. Mus. Hist. Nat. Lyon 4, 1–266.
Von Reichenau, W. (1915). Beiträge zur näheren Kenntnis fossiler Pferde aus Deutschem Pleistozän, inbesondere über die Entwicklung und die Abkaustadien des Gebisses vom Hochterrassenpferd. Equus mosbachensis v. R, Darmstadt, Germany.
Wagner, G. A., Maul, L. C., Lösher, M., and Schreiber, H. D. (2011). Mauer – the type site of Homo heidelbergensis: palaeoenvironment and age. Q. Sci. Rev. 30, 1464–1473. doi: 10.1016/j.quascirev.2010.01.013
Walker, A., Hoeck, H. N., and Perez, L. (1978). Microwear of mammalian teeth as an indicator of diet. Science 201, 908–910. doi: 10.1126/science.684415
Weinstock, J., Willerslev, E., Sher, A., Tong, W., Ho, S. Y. W., Rubenstein, D., et al. (2005). Evolution, systematics, and phylogeography of Pleistocene horses in the new world: a molecular perspective. PLoS Biol. 3:e241. doi: 10.1371/journal.pbio.0030241
Williams, S. H., and Kay, R. F. (2001). A comparative test of adaptive explanations for hypsodonty in ungulates and rodents. J. Mammal. Evol. 8, 207–229. doi: 10.1023/A:1012231829141
Winans, M. C. (1989). “A quantitative study of the North American fossil species of the genus Equus,” in The Evolution of Perissodactyls, eds D. R. Prothero and R. M. Schoch (Oxford: Oxford University Press, 262–297.
Keywords: Equus, Pleistocene, Europe, biochronology, ecomorphology, stenonid horses, caballoid horses, Equus hydruntinus
Citation: Boulbes N and van Asperen EN (2019) Biostratigraphy and Palaeoecology of European Equus. Front. Ecol. Evol. 7:301. doi: 10.3389/fevo.2019.00301
Received: 31 March 2019; Accepted: 26 July 2019;
Published: 10 September 2019.
Edited by:
Florent Rivals, Catalan Institute for Research and Advance Studies (ICREA), SpainReviewed by:
Maria Teresa Alberdi, National Museum of Natural Sciences (MNCN), SpainJean-Luc Guadelli, UMR5199 De la Prehistoire A l'actuel Culture, Environnement et Anthropologie (PACEA), France
Copyright © 2019 Boulbes and van Asperen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nicolas Boulbes, bmljb2xhcy5ib3VsYmVzQGNlcnB0YXV0YXZlbC5jb20=