
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 17 May 2019
Sec. Paleontology
Volume 7 - 2019 | https://doi.org/10.3389/fevo.2019.00176
This article is part of the Research Topic Examining Evolutionary Trends in Equus and its Close Relatives from Five Continents View all 14 articles
 Flavia Strani1,2,3*
Flavia Strani1,2,3* Diana Pushkina4
Diana Pushkina4 Hervé Bocherens5,6
Hervé Bocherens5,6 Luca Bellucci2,3,7
Luca Bellucci2,3,7 Raffaele Sardella1,2,3
Raffaele Sardella1,2,3 Daniel DeMiguel8,9
Daniel DeMiguel8,9The intermontane Anagni Basin (Frosinone, central Italy) is an important region for Italian biochronology and paleoecology due to the presence of two rich fossil assemblages dated to the Early (Coste San Giacomo) and Middle Pleistocene (Fontana Ranuccio). These sites have yielded a vast collection of large fossil mammals with a well-documented presence of fossil equids in both localities (represented mostly by isolated teeth). Coste San Giacomo is dated to around 2.1 Ma, thereby having recorded the effects of the onset of the Quaternary glacial cycles, which led to a gradual trend toward colder and more arid conditions in the Northern Hemisphere. The fossil equids of this site belong to the first group of grazing stenonid equids of the genus Equus that spread to the Italian Peninsula during the so called “Elephant- Equus” event, which marked the appearance of new large mammals living in herds in open and arid environments. The site of Fontana Ranuccio is dated to around 400 ka, close to the MIS 12–11 succession (the “Mid-Brunhes event”), which marked the end of the Middle Pleistocene Transition. The fossil horses from Fontana Ranuccio represent one of the oldest caballoid (or “true horses”) populations of the Italian Peninsula. The Anagni Basin, thus, provides important data to investigate paleoecological adaptations of these groups of equids in response to two critical environmental and climatic shifts of the Pleistocene. We explore their niche occupation by examining long-term dental wear patterns and tooth enamel carbon and oxygen stable isotopic composition. Both taxa appear to have exhibited a narrow dietary niche, displaying a clear abrasive (highly specialized) grass-rich diet. In particular, caballoid equids from Fontana Ranuccio show a more abrasion-dominated mesowear signature. Stenonid equids from Coste San Giacomo exploited broader and more diverse landscapes during the Early Pleistocene, whereas caballoid horses from Fontana Ranuccio appeared to have limited their dietary adaptations to a stricter grazing behavior in more closed environments.
The intermontane Anagni Basin, situated in the central part of the Italian Peninsula, developed between the Late Pliocene and the early Middle Pleistocene (Carrara et al., 1995; Galadini and Messina, 2004). Two rich vertebrate fossil sites, the Early Pleistocene locality of Coste San Giacomo (CSG) and the Middle Pleistocene site of Fontana Ranuccio (FR), were discovered in the 1970s, following different surveys carried out by the Italian Institute of Human Paleontology (IsIPU) (Figure 1). CSG has been dated to around 2.1 Ma based on magnetostratigraphy, pollen, and its record of micro mammals recovered from alluvial deposits (Bellucci et al., 2014). It is one of the few Italian localities that record the terrestrial habitat conditions after the onset of the Pleistocene glaciations that marked the Plio-Pleistocene transition at 2.6 Ma (Lisiecki and Raymo, 2005, 2007). FR fossiliferous layer located within a succession of pyroclastic deposits has been dated to around 0.4 Ma (Pereira et al., 2018), close to the climatic transition of the “Mid-Brunhes Event” (MIS 12–11) with the definitive association of the glacial cycles ruled by a 100 kyr periodicity and the consolidation of the Early-Middle Pleistocene Transition (Head and Gibbard, 2015).
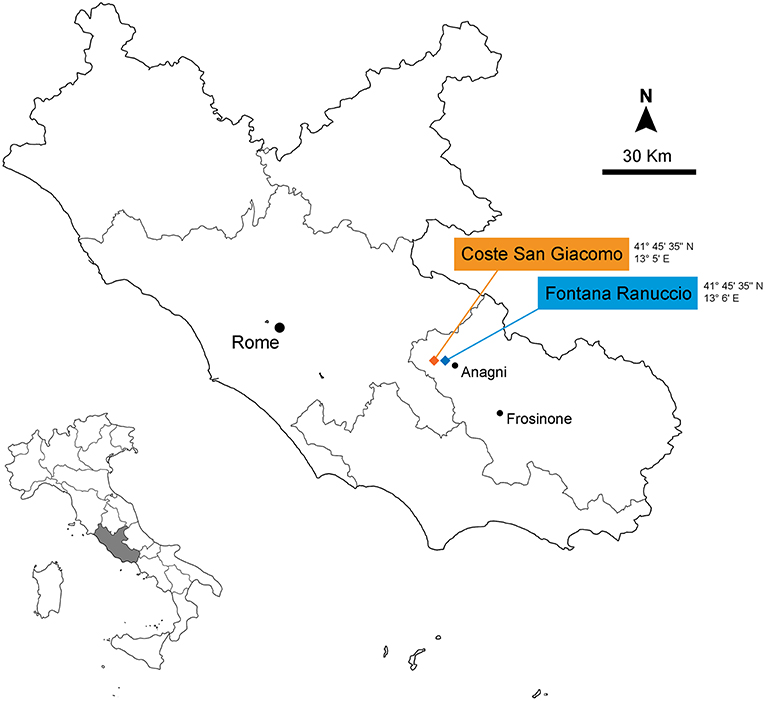
Figure 1. Geographical location of the Coste San Giacomo and Fontana Ranuccio localities.
Importantly, both localities have yielded abundant fossil mammal material, among which equid remains are well-represented (Segre and Ascenzi, 1984; Bellucci et al., 2012, 2014; Strani et al., 2018a). Fossil human teeth were also unearthed in FR (Rubini et al., 2014). The equid remains from CSG were attributed to Equus stenonis by Biddittu et al. (1979), but a recent study (Palombo et al., 2017) points to stronger affinities with the middle-size Equus senezensis and proposes a new provisional identification of the CSG material as E. senezensis aff. E. sen. stehlini. This taxon is among the first groups of stenonid equids (monodactyl horses with a “V” shaped linguaflexid between the metaconid and metasylid of the lower cheek teeth; Forsten, 1988) to reach the Italian Peninsula during the so-called “Elephant- Equus” event with the appearance of new large mammals living in herds in drier habitats (Azzaroli, 1977; Lindsay et al., 1980). Equids' remains from FR have been tentatively classified as Equus cf. E. mosbachensis (Segre and Ascenzi, 1984), belonging to one of the first caballoid (or “true horses,” monodactyl equids with a “U” shaped linguaflexid; Forsten, 1988) populations recorded on the Italian Peninsula after the disappearance of the early Middle Pleistocene stenonid forms (Caloi, 1997; Alberdi and Palombo, 2013).
Both modern and fossil Equus taxa are usually considered highly specialized grazers that consistently occupied similar habitats through the Pleistocene and Holocene (i.e., open grasslands), despite it has been observed that extant species can also include a noticeable amount of browse in their diet (Roeder, 1999) and that some fossil Equus populations displayed less abrasive feeding behaviors (Kaiser and Franz-Odendaal, 2004). The material from the Anagni basin represents an exceptional case to investigate the habitat occupation and resource use of two groups of fossil equids and test the hypothesis that both stenonid and caballoid horses exploited exclusively open habitats and fed mostly on grasses in Central Italian Peninsula. In this study we compare dental remains of E. senezensis aff. E. sen. stehlini from CSG and Equus cf. E. mosbachensis from FR in occlusal surface mesowear, a proxy for dietary abrasiveness in herbivorous mammals (Fortelius and Solounias, 2000), and in stable carbon and oxygen isotope abundances, proxies for habitat openness (Van der Merwe and Medina, 1991) and temperature (Dansgaard, 1964), to shed light on the niche occupation and resource exploitation mechanisms of stenonid and caballoid equids on central Italian Peninsula.
Our data will allow us to obtain comprehensive information on the paleoecological adaptations of the Equus genus in the region following two major climatic events, and to understand if this group could be more flexible in terms of feeding behavior and habitat occupation during the Early and Middle Pleistocene.
Mesowear reflects the cumulative effects at a macroscopical level of the items ingested by herbivores (foods and exogenous particles such as grit or dust) on tooth morphology that are produced in a long period of time compared to the lifespan of the animal (Fortelius and Solounias, 2000). Dental mesowear is a direct signal of a species' diet that varies between browsers, which feed mostly on soft plant resources (e.g., leaves, twigs, green stems), and grazers that clip abrasive vegetation (e.g., grasses) at or near ground level.
Attrition (tooth-to-tooth contact) and abrasion (tooth-to-food contact) are the main factors that influence the occlusal morphology of the teeth, determining the cusps sharpness and the height of the occlusal relief. In browsing animals attrition mainly determines the tooth wear leading to sharper cusps and a higher occlusal relief. In species that feed on more abrasive plant resources the higher level of tooth-to-food contact produces blunter cusps and a lower occlusal relief (Fortelius and Solounias, 2000). Traditional mesowear is limited to upper second molars (M2), but it can be extended to other tooth positions (Kaiser and Solounias, 2003; DeMiguel et al., 2010, 2012; Strani et al., 2015, 2018a,b).
The method has been developed to examine the diet-related dental wear patterns in ungulates and has been successfully employed to investigate dietary adaptations of different groups of both modern and fossil artiodactyls and perissodactyls (Fortelius and Solounias, 2000; Schulz et al., 2007; DeMiguel et al., 2008, 2011, 2018; Bernor et al., 2014; DeMiguel, 2016; Marín-Leyva et al., 2016; Maniakas and Kostopoulos, 2017). Dental mesowear also proved to be an optimal tool to study equid paleoecological adaptations, allowing to obtain information about the dietary change and evolution of this group from the early forms of the Eocene to the horses of the Pleistocene (Kaiser and Franz-Odendaal, 2004; Mihlbachler et al., 2011; Bernor et al., 2017).
Carbon isotopic (δ13C) abundance in herbivores reflects the presence of C3 and C4 vegetation in lower and mid-latitude habitats as well as a degree of canopy closure vs. openness in wooded high-latitude habitats. C3 plants, or woody vegetation and grasses that have a cool growing season, have δ13C values ranging between −34 and −23‰, with a suggested mean of −27‰ (Calvin and Benson, 1948; Hatch et al., 1967; Smith and Epstein, 1971; Koch, 1998). The canopy effect can result in up to 5‰ differences in δ13C in the same species at one site in pure C3 forest environments between the herbivores feeding in a heavily enclosed wooded environment or on plants from different tree elevation, and the herbivores feeding in more open parkland or grassland environments with higher evapotranspiration (Van der Merwe and Medina, 1991; Bocherens et al., 1999; Feranec and MacFadden, 2006; Drucker et al., 2008; Hofman-Kaminska et al., 2018).
The δ13C values of the carbonate fraction of bioapatite in large mammalian herbivores are higher than or enriched by 14‰ compared to their diet: δ13Cbioapatite = δ13Cdiet + 14‰ (Cerling et al., 1999). In warm temperate and cold boreal environments δ13C values higher than −13‰ in herbivore enamel indicates open landscapes, such as grasslands and steppes, in which the plants have δ13C values higher than −27‰ in diet or ecosystem δ13C values, whereas tooth enamel δ13C values lower than −13‰ indicate woodlands and denser forests (Bocherens, 2014).
Oxygen isotopic (δ18O) abundance of the herbivores' enamel apatite reflects the source and amount of body-water ingested, either free drinking water or obtained from food and plants. The δ18O in the enamel apatite is a proxy for δ18O values of meteoric water, which varies due to the source of precipitation and effects of latitude, altitude, continentality, temperature, and evaporation (Dansgaard, 1964; Longinelli, 1984; Luz et al., 1984; Bryant and Froelich, 1995; Bryant et al., 1996; Kohn, 1996; Sponheimer and Lee-Thorp, 1999; Levin et al., 2006). Crudely, δ18O values of meteoric water are higher in warmer or dryer environments (Dansgaard, 1964). Thus, as a general rule, herbivores feeding in open steppe habitat are expected to have more enriched δ18O values than species from forested cooler and more humid habitat (Sponheimer and Lee-Thorp, 1999). Obligate or water-dependent drinkers usually have δ18O values lower than non-obligate drinkers, being dependent on water availability and seasonality of rainfall. Seasonal variation in rainwater in mid- and high latitude environments and species migration can be significant, producing intra-tooth variations of 3–4‰, or more (e.g., in high-crowned ungulates, horses and bison, Hoppe et al., 2004; Feranec et al., 2009). Oxygen isotopic composition can be affected not only by daily ecology and migration but also by life history and physiology of mammals. Oxygen isotopic compositions are higher in nursing animals during lactation and weaning periods because of milk consumption, which is related to the body water in the mother, and more enriched in 18O due to the preferential loss of 16O through sweat, urine, and expired water vapor (Bryant and Froelich, 1995; Kohn, 1996).
The material studied consists of upper cheek teeth belonging to E. senezensis aff. E. sen. stehlini from CSG and to Equus cf. E. mosbachensis from FR. All the examined fossil material is housed at the Italian Institute of Human Paleontology (IsIPU) (Anagni, Italy). The comparative material was obtained from literature (see in text).
Past studies on the dental wear patterns of E. senezensis aff. E. sen. stehlini and Equus cf. E. mosbachensis from the Anagni Basin examined both upper and lower cheek teeth (Strani et al., 2015, 2018a,b). In this study we examined only upper teeth [P4-M3, following Kaiser and Solounias (2003)] in their occlusal relief (high or low) and cusp shape (sharp, rounded, or blunt) of the apex of the paracone or metacone and scored for a better comparison with available data of both modern and fossil equids because mesowear analysis is often restricted to the upper dentition. Occlusal relief and cusp shape scores were also converted to a single mesowear score (MWS) following the “mesowear ruler” developed for scoring dental mesowear on fossil equids by Mihlbachler et al. (2011). The method is based on seven cusp types (numbered from 0 to 6), ranging in shape from high and sharp (stage 0) to completely blunt with no relief (stage 6). Additionally, a “stage 7” is given to teeth with a convex cusp apex. A total of 30 specimens were scored using this method (E. senezensis aff. E. sen. stehlini N = 20; Equus cf. E. mosbachensis N = 10). Previously unavailable upper teeth of E. senezensis aff. E. sen. stehlini (N = 9) have been added to the original sample analyzed in Strani et al. (2015, 2018b).
Results were compared with mesowear data of four extant equids: plains zebras (Equus quagga) (data from Fortelius and Solounias, 2000 and Rivals and Semprebon, 2010), Grévy's zebras (Equus grevyi) (data from Fortelius and Solounias, 2000), onagers (Equus hemionus), and African free ranging donkeys (Equus asinus) (data from Schulz et al., 2007). To distinguish fossil and modern equid taxa based on mesowear variables (percentages of high relief, rounded, and blunt cusps) a hierarchical cluster analysis was performed using the Ward's method and the Euclidean distance. E. senezensis aff. E. sen. stehlini dental mesowear data were compared to those of E. stenonis from the Early Pleistocene locality of Olivola (~1.8 Ma, north central Italy) (Strani et al., 2018c) and of the Britain sites of Mundesley and East Runton (~1.8 Ma) (Rivals and Lister, 2016). Equus cf. E. mosbachensis dental wear patterns were also compared to those of the fossil Middle Pleistocene caballoid equid population from the central European localities (Steinheim and Heppenloch,MIS 11, Germany) (data from Rivals and Ziegler, 2018), from different Britain sites (Clacton, Hoxne and Swanscombe; MIS 11) (Rivals and Lister, 2016) and from Caune de l'Arago (438 ± 31 ka, France) (data from Rivals et al., 2015a).
Discriminant analysis was performed to examine the resolution of mesowear variables applied to the fossil. The percentages of high relief, rounded and blunt cusps were used as independent variables and two dietary (conservative and radical) classifications of modern ungulate taxa as grouping variables (Fortelius and Solounias, 2000). Equus quagga from Rumuruti (Rivals and Semprebon, 2010), Equus hemionus and Equus asinus (Schulz et al., 2007) were added to the modern taxa with known diets and treated as grazers in both conservative and radical classifications. All analyses were performed using IBM SPSS Statistics 24.
We obtained stable carbon (δ13C) and oxygen isotopic (δ18O) abundances from the upper cheek teeth (upper first or second molars). In horses the isotopic composition in M1 or M2 would reflect nutrients from mother's milk and should be slightly more depleted in 13C and more enriched in 18O compared to isotopic composition of subsequently erupting P2, P3, M3, and P4 that would better reflect solid diet of an adult animal (Hoppe et al., 2004). Here we made comparisons with M1 or M2 from German sites and took into account this difference when comparing samples taken from premolars. We used 3 teeth samples of Equus cf. E. mosbachensis from FR and two E. senezensis aff. E. sen. stehlini from CSG. One FR horse sample was from PM4 and exhibited enrichment by 1‰ in 13C and more enriched in 18O compared to two FR M2. In German sites only E. hydruntinus had permanent teeth compared and the obtained values were within variation ranges of 1 and 2 ‰ in 13C and 18O, respectively.
Enamel samples were either mechanically separated and ground to a finer condition in an agate mortar or drilled with a diamond-impregnated rotary tool. The uppermost surface of the tooth was removed before sampling usually by drilling with a dremel borer. Samples of approximately 20 mg were analyzed. Fossil enamel powders were pre-treated following the method described by Bocherens et al. (1996) but using the Continuous Flow (CF) for collecting carbon dioxide (Jacques et al., 2008; Bocherens et al., 2009). Our experience with fossil material from limestone cave has shown that the pre-treatment used here is strong enough to remove exogenous carbonates but we are aware of possible slight changes of the oxygen isotopic composition of tooth enamel carbonate.
Pretreated (Sample /Enamel) carbonate was reacted with 100% H3PO4 for 4 h at 70°C using a MultiFlow-Geo interfaced with the Elementar IsoPrime 100 IRMS. Final isotopic ratios are reported per mil (‰) calibrated with international standards (IAEA-603 δ13C: 2.46/δ18O: −2.37 and NBS-18 δ13C: −5.014/δ18O: −23.2), as well as three in-house standards. IonOS software (Version 3.2) by Elementar was used to carry out multi-point standard isotope calibration by generating a trend line (y = mx+c) that maps measured vs. expected isotopic results, which is then used to calibrate sample results. The measurement uncertainty was monitored using three in-house standards. The overall analytical precision is higher than 0.1‰ for carbon and better than 0.2‰ for oxygen isotopic values.
Stable isotopic results are expressed as the following standard δ-notation: X = [(Rsample/Rstandard)-1]*1,000, where X is referred to δ13C and δ18O values and R is equivalent to 13C/12C or 18O/16O, respectively. The recorded delta values follow the international reference standards, a “Vienna PeeDee Belemnite” (VPDB) for the carbon and oxygen. Additionally, δ18O values relative to “Vienna Standard Mean Ocean Water” (VSMOW) are given. In addition, the carbonate content (CaCO3%) was determined using the ratio between amount of CO2 released by the reaction, as detected from the peak intensity for mass 44 and the weight of pure carbonate used as a standard, with an analytical error of 0.3%, based on multiple analysis of reference enamel samples.
To calculate values for temperature and drinking water from oxygen isotope values, and convert oxygen carbonate and phosphate fractions, we used and equation from Lécuyer et al. (2010)
The estimates of the δ18O values of drinking water consumed by species were calculated, first, by converting the oxygen isotope composition of structural carbonate to its equivalent in phosphate, using equations that are robust for many different species: δ18OP = 0.71 × δ18OW+22.60 (Delgado Huertas et al., 1995).
Then the oxygen isotope composition of phosphate was converted to that of drinking water of different groups of species, according to formulas below and reformulated based on original data from Skrzypek et al. (2011) based on current relationship between air and water. We calculated temperature (°Celcius) using the formula for continental Europe:
We used both parametric and non-parametric tests with α = 0.05. To identify the sources of significant differences we used ANOVA and among unequal variances we used post hoc Tamhane tests.
The results were compared with the late middle and late Pleistocene horse data from Steinheim, Mauer, Bockstein and Vogelherd (Pushkina et al., 2014).
E. senezensis aff. E. sen. stehlini upper teeth from CSG showed mostly low occlusal relief (75%) and a higher percentage of rounded cusps (45%) over sharp (35%) and blunt cusps (20%) (Table 1). Low occlusal relief and high occurrence of rounded cusps E. senezensis aff. E. sen. stehlini from CSG point to a generally abrasive diet. Equus cf. E. mosbachensis teeth also displayed low occlusal relief and mostly rounded cusps (50%) (Table 1), a pattern pointing to a dominance of abrasion over attrition.
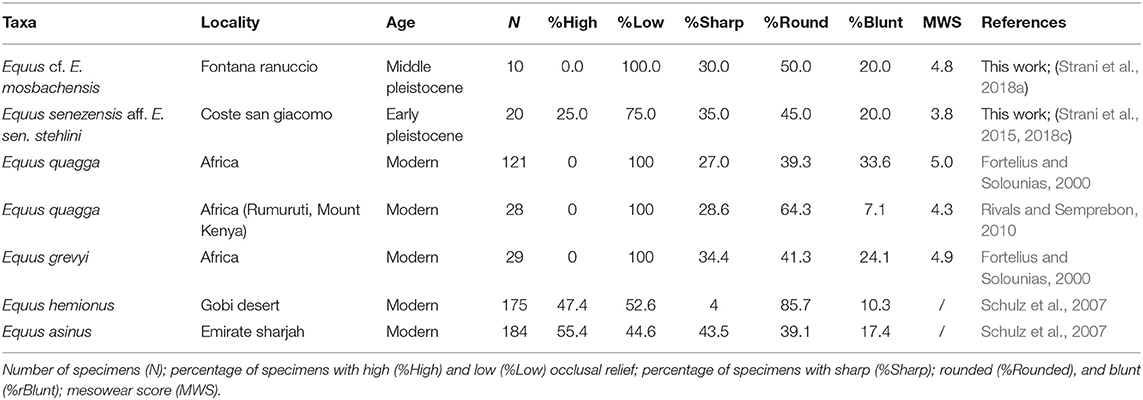
Table 1. Summary of dental mesowear analysis.
E. senezensis aff. E. sen. stehlini was clustered with the onager and the free ranging donkeys (cluster B) both displaying a grazing signature but with E. asinus showing a less abrasion-dominated diet (Schulz et al., 2007) (Figure 2). Equus cf. E. mosbachensis fell in cluster A with modern zebras (Figure 2), which are all characterized by strong abrasion-dominated mesowear patterns (Table 1) (Fortelius and Solounias, 2000; Rivals and Semprebon, 2010). Discriminant analysis performed with the mesowear variables provides a satisfactory dietary discrimination with 71.9% of extant taxa correctly classified according to the conservative classification and 75.4% according to the radical one (70.2 and 75.4%, respectively, in cross-validation). Both E. senezensis aff. E. sen. stehlini and Equus cf. E. mosbachensis are classified as grazers by both the conservative and radical classification (Figures 3A,B).
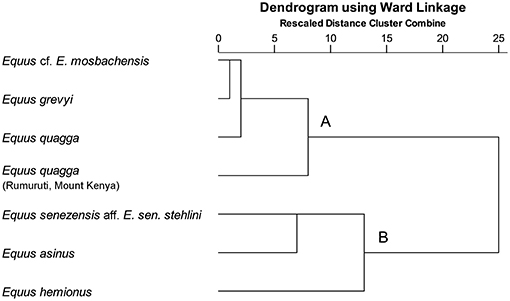
Figure 2. Mesowear hierarchical cluster diagram based on the percentage of high occlusal relief, round and blunt cusps of fossil populations with extant equids. (Data for extant equids from Fortelius and Solounias, 2000; Schulz et al., 2007; Rivals and Semprebon, 2010).
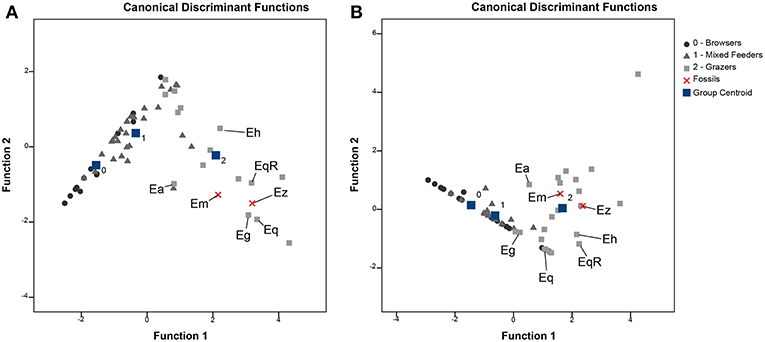
Figure 3. Bivariate diagrams based on discriminant analysis: conservative classification (A) and radical classification (B). Minute abraded brachydont species have been excluded. Fossil taxa Ez, Equus senezensis aff. E. sen. stehlini; Em, Equus cf. E. mosbachensis; Eq, Equus quagga; EqR, Equus quagga (Rumuruti population); Eg, Equus grevyi; Eh, Equus hemionus; Ea, Equus asinus. Group centroids: browsers (0); mixed feeders (1); grazers (2).
E. senezensis aff. E. sen. stehlini displayed an average MWS of 3.8, lower than the one recorded in the upper cheek teeth of E. stenonis from Olivola (5.3), from stenonid horses from East Runton (E. bressanus, 4.5; E. stenonis, 4.0) and from E. stenonis from Mundesley (Figure 4), pointing to a less strictly abrasion-dominated feeding behavior in the stenonid equid from Coste San Giacomo. Equus cf. E. mosbachensis from Fontana Ranuccio, showed a mean MWS of 4.8 higher than the one recorded in all the other caballoid equids from Germany (Steinheim, 3.8; Heppenloch, 4.3), France (Caune de l'Arago, 3.9) and Britain (Swanscome, 3.7; Hoxne, 4.0; Clacton, 4.4) (Figure 5), suggesting an overall more abrasive diet for the Middle Pleistocene fossil caballoid equid of the Anagni Basin.
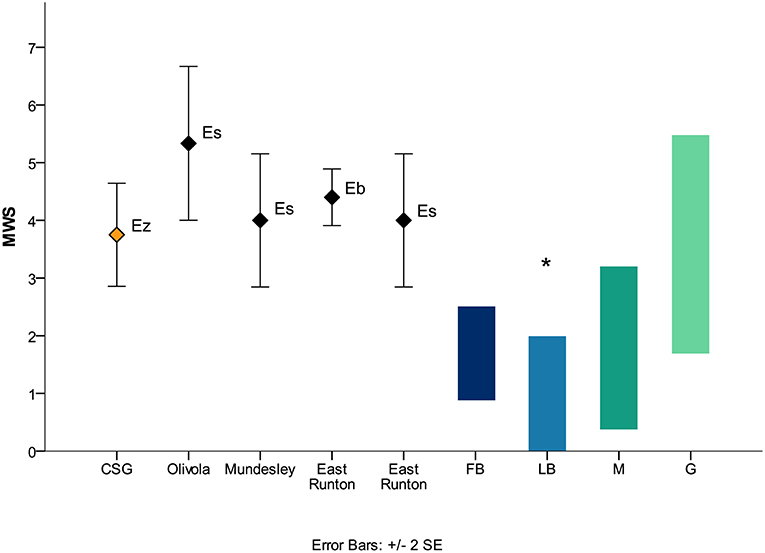
Figure 4. Mesowear results of fossil stenonid equids and extant ungulates. Modern ungulates data from Fortelius and Solounias (2000) (diet information according to the radical classification; minute abraded brachydont species have been included), Rivals and Semprebon (2010). Mundesley and East Runton data from Rivals and Lister (2016). MWS, mesowear score; CSG, Coste San Giacomo; Ez, Equus senezensis aff. E. sen. stehlini; Es, Equus stenonis; Eb, Equus bressanus; FB, fruit browsers; LB, leaf browsers; M, mixed feeders; G, grazers; *, Heterohyrax brucei.
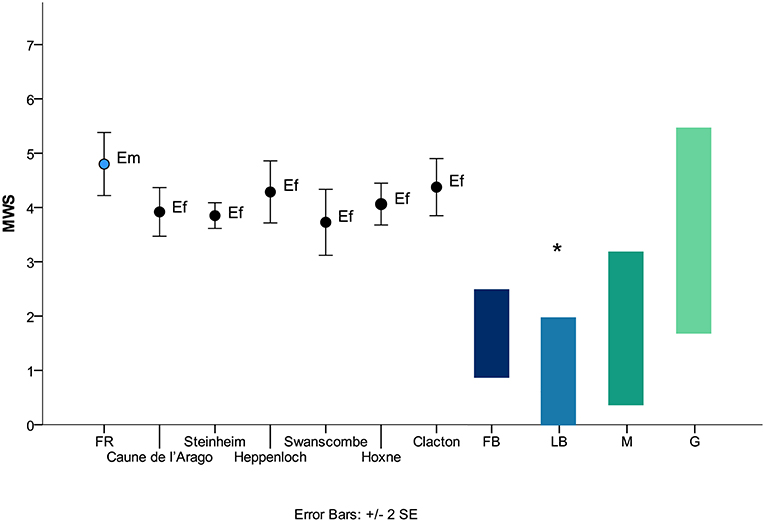
Figure 5. Mesowear results of fossil stenonid equids and extant ungulates. Modern ungulates data from Fortelius and Solounias (2000) (diet information according to the radical classification; minute abraded brachydont species have been included), Rivals and Semprebon (2010). Caune de l'Arago, Steinheim, Heppenloch, Swanscombe, Hoxne and Clacton data from Rivals et al. (2015a), Rivals and Lister (2016) and Rivals and Ziegler (2018). MWS, mesowear score; CSG, Coste San Giacomo; Ez, Equus senezensis aff. E. sen. stehlini; Es, Equus stenonis; Eb, Equus bressanus; FB, fruit browsers; LB, leaf browsers; M, mixed feeders; G, grazers; *, Heterohyrax brucei.
Compared equids E. senezensis aff. E. sen. stehlini from CSG, Equus cf. E. mosbachensis from FR and Mauer, E. ferus/ E. hydruntinus/E. sp. from Steinheim and E. caballus from Bockstein and Vogelherd varied significantly in δ13C and δ18O values (ANOVA [F5, 26 = 8.6, p < 0.001, r2 = 0.55] for δ13C / ecosystem; [F5, 26 = 5.7, p < 0.001, r2 = 0.43] for δ18O VPDB/VSMOW/temperature).
The δ13C values of E. senezensis aff. E. sen. stehlini from the Early Pleistocene CSG were −12.49 and −11.94‰, falling within the values of the late Pleistocene horses from Bockstein and Vogelherd and middle late Pleistocene E. hydruntinus from Steinheim (Table 2) (Figure 6A). The δ18O values of CSG E. senezensis aff. E. sen. stehlini were higher than in the compared German sites −4.02 and −6.19‰ (VPDB), −26.71 and −24.48‰ (CO3 VSMOW) (Table 2) (Figure 6B), 17.76 and 15.6‰ (PO4 VSMOW), respectively, suggesting that the horses used water sources with δ18Owater −6.81 and −9.86‰.
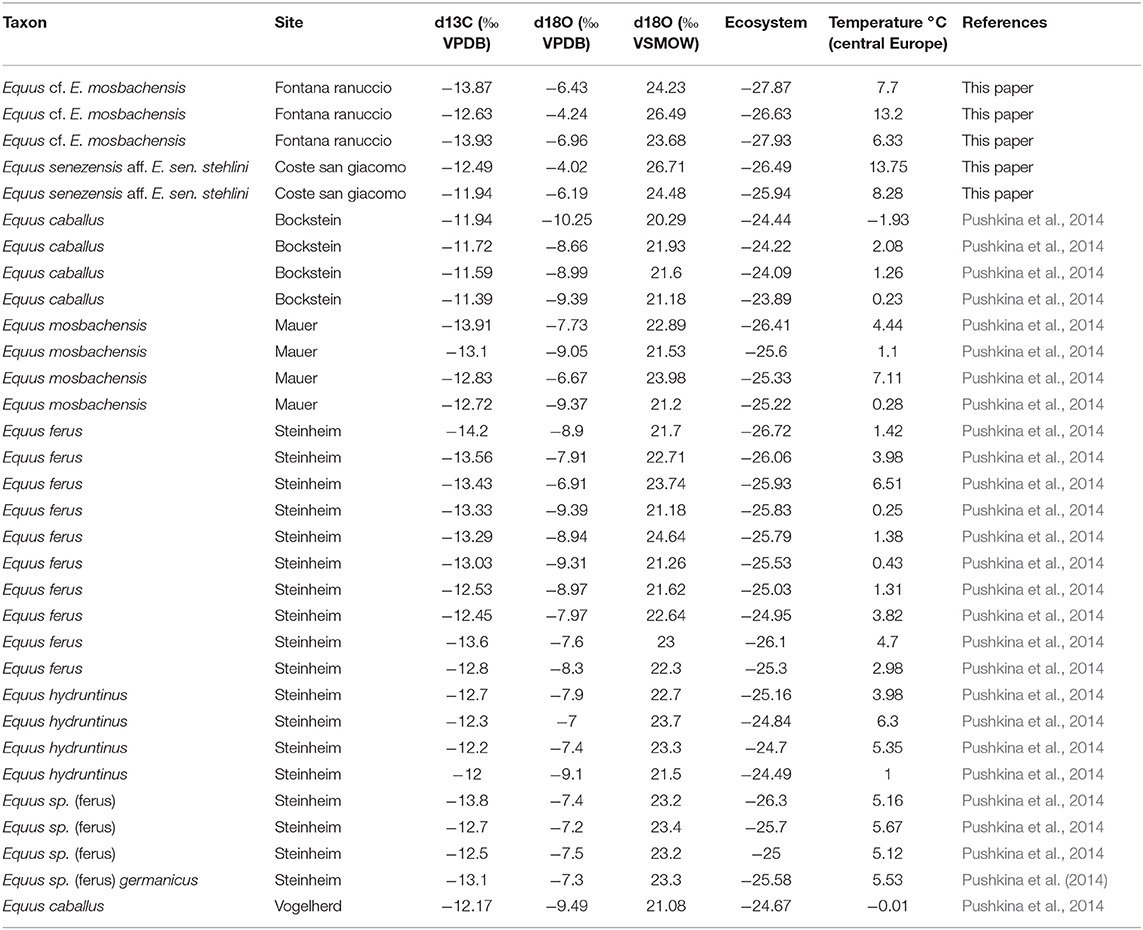
Table 2. Isotopic results for Equus senezensis aff. E. sen. stehlini from CSG, Equus cf. E. mosbachensis from FR, and for Pleistocene equids from German localities [data from Pushkina et al. (2014)].
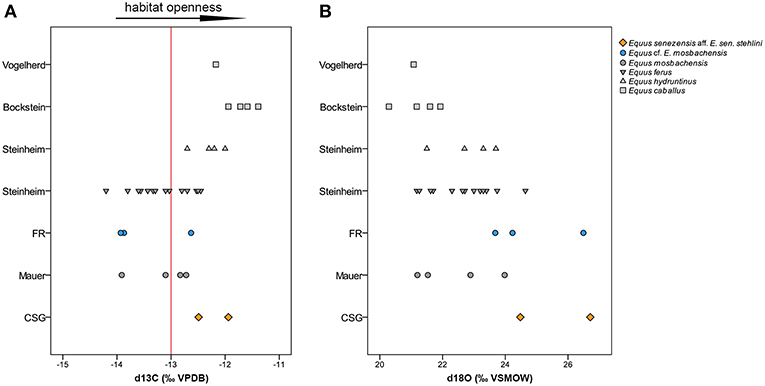
Figure 6. δ13C (A) and δ18O (B) values of the studied taxa compared to fossil equids of central Europe (data from Pushkina et al., 2014). The red line represents the boundary between δ13C values that indicate open (δ13C > −13‰) or closed landscapes (δ13C < −13‰) in warm temperate and cold boreal environments (Bocherens, 2014).
The δ13C values of Equus cf. E. mosbachensis from the Middle Pleistocene FR ranged from −13.93 to −12.63‰ (Table 2) (Figure 6A), which is similar to E. mosbachensis from Mauer and equids from Steinheim. Equus cf. E. mosbachensis differed from the late Pleistocene Equus ferus (post hocs Tamhane p = 0.004 δ13C/ ecosystem).
The δ18O values of FR Equus cf. E. mosbachensis varied between −6.96 and −4.24‰ (VPDB), −23.68 and −26.49‰ (CO3 VSMOW) (Table 2) (Figure 6B), 14.83 and 17.54‰ (PO4 VSMOW), respectively, indicating that water sources used had the δ18Owater values of −10.94 and −7.12‰.
The only peculiar difference between the Italian and German sites is demonstrated in the temperature calculations, indicating much higher temperatures for both CSG and FR (6.2–12°C) than for the German sites (−0–6.8°C) (Table 2).
Mesowear and stable isotopic analyses suggest that horses in both sites inhabited rather open and dry environments with abrasive C3 vegetation as a dietary source for the majority. As obligate drinkers, however, horses could have used different water sources in more humid and closed, or more open and arid environments, which could have been at slightly different elevation and subjected to seasonal variation in water availability. Both E. senezensis aff. E. sen. stehlini from CSG and Equus cf. E. mosbachensis from FR display dental mesowear patterns highly consistent with a grazing feeding behavior, according to the results reported in Strani et al. (2015, 2018a,b,c), where both upper and lower cheek teeth were analyzed. Differences, however, are observed in terms of niche occupation in the Early Pleistocene stenonid equid, displaying a less abrasion-dominated diet in a more open landscape of steppe, shrubland, or light forest compared to the Middle Pleistocene caballoid species that fed strictly on abrasive sources in a more closed habitat of woodland or shrubland. Temperature-wise these localities did not appear to have differed. These findings allow us to provide information on the paleoecology of the two groups of fossil equids of the Anagni Basin, hereby shedding light on their ecological response to the major climatic events that marked the Early and Middle Pleistocene: the onset of the Pleistocene glaciation, which led to a gradual trend toward cooler climatic conditions and subsequent reduction of sub-tropical vegetation in Eurasia (Fortelius et al., 2006; Combourieu-Nebout et al., 2015), and strengthening of the Early-Middle Pleistocene Transition, which led to an increase of interglacial temperatures (Head and Gibbard, 2015). Isotopic results obtained for E. senezensis aff. E. sen. stehlini indicate that this equid occupied open and dry habitats similarly to the Late Pleistocene horses and middle Late Pleistocene E. hydruntinus from central Europe (Pushkina et al., 2014). Dental mesowear patterns are compatible with a grazing behavior similarly to E. stenonis populations, e.g., from Sésklo (MN17 biozone, Early Pleistocene, Greek Peninsula) (Rivals and Athanassiou, 2008) and from Olivola (~1.8 Ma, Italian Peninsula) (Strani et al., 2018b).
E. stenonis is a common species of both Italian and Greek Peninsulas associated with fossil ungulates showing generally mixed or grazing dietary adaptations in open and dry environments (Rivals and Athanassiou, 2008; Strani et al., 2018b). In contrast, E. senezensis aff. E. sen. stehlini from CSG is associated with artiodactyls showing a wider range of feeding behaviors in a more heterogeneous landscape comprised of wetlands, forests and grasslands (Strani et al., 2015, 2018c), while displaying a less abrasion-dominated dental mesowear pattern than E. stenonis from Olivola (Figure 4). Similar conditions are observed in East Runton, where E. bressanus displayed long-term grazing (according to molar mesowear) and short-term browsing diet (according to tooth microwear) and is grouped with other ungulates with a variety of feeding behaviors (Rivals and Lister, 2016).
The stenonid Equus altidens from the late Early Pleistocene Southern European site of Vallparadís Estació (layer EVT7) also displayed a certain dietary plasticity with a short-term mixed feeding behavior (Strani et al., 2019). E. senezensis aff. E. sen. stehlini is also one of the best represented ungulates from CSG, whereas fewer individuals have been recorded in Olivola (Strani et al., 2018b). This suggests that this equid, while feeding mostly on grasses, may have also accessed more diverse food items leading to a greater intake of softer plant resources compared to the similarly aged E. stenonis from Sésklo and to the later stenonid equids from Olivola. This scenario is consistent with the onset of a gradual trend toward cooler climatic conditions after the initiation at the Plio-Pleistocene boundary (2.6 Ma) of the Quaternary glaciations regulated by a 41 kyr periodicity (Lisiecki and Raymo, 2005), which led to the augmentation of open habitats and the disappearance of sub-tropical vegetation in the European continent (Bertini, 2003; Bertini, 2006, Fortelius et al., 2006; Bertini, 2010, 2013; Kahlke et al., 2011; Combourieu-Nebout et al., 2015). This process probably affected later Northern Europe as the paleoenvironmental conditions of East Runton appear to be more heterogeneous compared to the coeval locality of Olivola.
The more abrasion-dominated mesowear patterns of Equus cf. E. mosbachensis point to a stricter grazing diet, and are comparable to the ones observed in modern zebra populations, which feed almost exclusively on grasses (Bodenstein et al., 2000; Fortelius and Solounias, 2000; Rivals and Semprebon, 2010; Kartzinel et al., 2015). The same trend is observed also in the Middle Pleistocene caballoid equids from other European localities, which also display abrasion-dominated mesowear signatures (Rivals et al., 2008, 2015a; Rivals and Lister, 2016; Rivals and Ziegler, 2018). A similar trend from less abrasion-dominated to strictly grazing diet is observed even in Early and Middle Pleistocene equid populations of Britain (Rivals and Lister, 2016).
Isotope analysis points instead to a preference for slightly more closed environments in Equus cf. E. mosbachensis from FR similarly to E. mosbachensis from Mauer and Middle and Late Pleistocene equids from Steinheim (Pushkina et al., 2014) in comparison to E. senezensis aff. E. sen. stehlini. Long-term grazing behavior and short-term less abrasive diets are observed in E. ferus from the MIS 11 German localities of Hoppenloch and Steinheim (Rivals and Ziegler, 2018) and from the MIS 9 site Schöningen (Rivals et al., 2015b), suggesting that also in central Europe Middle Pleistocene horses appeared to have successfully exploited less open habitats.
FR Equus cf. E. mosbachensis is associated with ungulates displaying a wide range of feeding behavior with a noticeable abundance of browsing and mixed feedings cervids (Strani et al., 2018a,d). A similar condition is observed in Steinheim, where herbivores with diverse diets in humid and more closed habitats are recorded (Pushkina et al., 2014; Rivals and Ziegler, 2018). Isotopic analysis from the ungulates of Mauer also suggest warm and densely wooded landscape for this locality (Pushkina et al., 2014). On the contrary, fossil horses from the layer G (MIS 12) of Caune de l'Arago Cave (France) and from Hoppenloch are associated with a lower range of dietary traits pointing to drier environmental settings (Rivals et al., 2008; Rivals and Ziegler, 2018). This suggests that Equus cf. E. mosbachensis from the Anagni Basin occupied an ecological niche similar to the fossil horses from Steinheim in comparable environmental conditions, as grazers that feed also in more closed habitats. The higher MWS recorded in the FR population (4.8) compared to that of the Steinheim (3.8) (Figure 5), suggests that the fossil horses of central Italian Peninsula had a stricter grazing behavior compared to central European populations.
FR archeopaleontological layer with its fossil assemblage is dated to around “Mid-Brunhes event” (Biddittu et al., 1979; Muttoni et al., 2009; Pereira et al., 2018), which is associated with an increase in the amplitude of the interglacials and glacial cycles (Lang and Wolff, 2011; McClymont et al., 2013), and with one of the warmest and longest interglacial periods of the last 400 ka, registered during MIS 11 (Weirauch et al., 2008). According to palynological data from the MIS 11 Boiano Section (central Italian Peninsula) these warmer conditions led to the expansion of closed canopy forests with high summer precipitations (Combourieu-Nebout et al., 2015). The paleoecological information for the fossil horse of FR, suggests that in this scenario the Middle Pleistocene caballoid equids from central Italian Peninsula adapted to the reduction of open grasslands by exploiting also more closed habitats in heterogeneous landscapes. A possible competition with other large gregarious ungulates, such as Bos primigenius and Stephanorhinus sp., could have also played a key role in the niche occupation of this equid. The higher temperature gradient recorded by the isotopic analysis of the equid dental material compared to those observed in Steinheim, also can imply warmer conditions that characterized the Anagni Basin during this interval. This is concordant with the hypothesis of the Italian Peninsula acting as a southern refugium for mammal taxa and human populations during colder periods and extreme glacials of the Pleistocene (Manzi et al., 2011; Manzi, 2016).
This work offers important updated information on the paleoecological adaptations of Pleistocene equids of the Italian Peninsula. Overall Early Pleistocene stenonid horses grazed in open landscapes but demonstrated less abrasion-dominated diet compared to the later forms, suggesting the existence of more heterogeneous environmental conditions during the early Early Pleistocene before the Gelasian/Calabrian (middle/late Villafranchian) transition. In the same region, Middle Pleistocene caballoid populations displayed more abrasion-dominated long-term feeding behavior, having occupied less open habitats (at the edge between grasslands and woodlands) than stenonid forms of the Early Pleistocene, both in possibly warmer climatic conditions compared to French and German localities of the same age. Temperature calculations, however, should be considered with caution, since the sample size is quite small and based only on horses, and oxygen stable isotope variation apart from higher temperature can be associated with other factors, e.g., water sources from different elevation and subjected to seasonal variation in water availability.
Obtained data on the niche occupation of the stenonid Equus of the Anagni Basin support the spread of open landscapes in the Italian Peninsula following the initiation of the Pleistocene glacial cycles, suggesting, however, that these equids could have successfully adapted to exploit also more heterogeneous resources in the long-term timescale. Data from the caballoid Equus support the expansion of more closed environments in the region after the MIS 12–11 transition, though horses seemed to feed almost exclusively on abrasive vegetation possibly to avoid competition with other large ungulates that exploited softer food resources.
All authors have provided substantial contribution to the article. FS conceived the ideas and designed the approach. FS collected and elaborated the mesowear data. DP collected and performed the isotopic analysis with the help of HB, FS, and DP led the writing of the manuscript with the support of HB and DD. RS and LB provided helpful comments and inputs for the manuscript. All authors have approved the manuscript and agree with its submission to the journal.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This research has been supported by Avvio alla Ricerca 2017 (Ref. AR11715C7CE78D4F to FS) and the Spanish Agencia Estatal de Investigación (Ministerio de Economía, Industria y Competitividad)–European Regional Development Fund of the European Union (CGL2016-76431-P, AEI/FEDER EU). We thank the Italian Institute of Human Paleontology for preserving the collection and allowing the study of the material object of this investigation. We are also very grateful to Gina Semprebon and Mikael Fortelius for providing the raw mesowear data of modern equids (Equus quagga and Equus grevyi) and other extant ungulates We also acknowledge to Raymond L. Bernor for inviting us to contribute to the Examining Evolutionary Trends in Equus and its Close Relatives from Five Continents Research Topic. Finally, we thank the two reviewers for their helpful comments and suggestions.
Alberdi, M., and Palombo, M. (2013). The late Early to early Middle Pleistocene stenonoid horses from Italy. Quat. Int. 288, 25–44. doi: 10.1016/j.quaint.2011.12.005
Azzaroli, A. (1977). The Villafranchian stage in Italy and the Plio-Pleistocene boundary. Giornale di Geologia 41, 61–79.
Bellucci, L., Bona, F., Corrado, P., Magri, D., Mazzini, I., Parenti, F., et al. (2014). Evidence of late gelasian dispersal of African fauna at Coste san Giacomo (Anagni Basin, central Italy): early Pleistocene environments and the background of early human occupation in Europe. Quat. Sci. Rev. 96, 72–85. doi: 10.1016/j.quascirev.2013.10.011
Bellucci, L., Mazzini, I., Scardia, G., Bruni, L., Parenti, F., Segre, A. G., et al. (2012). The site of Coste San Giacomo (Early Pleistocene, central Italy): palaeoenvironmental analysis and biochronological overview. Quat. Int. 267, 30–39. doi: 10.1016/j.quaint.2011.04.006
Bernor, R., Göhlich, U., Harzhauser, M., and Semprebon, G. (2017). The Pannonian C hipparions from the Vienna Basin. Palaeogeogr. Palaeoclimatol. Palaeoecol. 476, 28–41. doi: 10.1016/j.palaeo.2017.03.026
Bernor, R., Semprebon, G., and Damuth, J. (2014). Maragheh ungulate mesowear: interpreting paleodiet and paleoecology from a diverse fauna with restricted sample sizes. Ann. Zool. Fennici. 51, 201–208. doi: 10.5735/086.051.0220
Bertini, A. (2003). Early to Middle Pleistocene changes of the Italian Flora and vegetation in the light of a chronostratigraphic framework. Il Quaternario 16, 19–36.
Bertini, A. (2010). Pliocene to Pleistocene palynoflora and vegetation in Italy: state of the art. Quat. Int. 225, 5–24. doi: 10.1016/j.quaint.2010.04.025
Bertini, A. (2013). Climate and vegetation in the Upper Valdarno Basin (central Italy) as a response to Northern Hemisphere insolation forcing and regional tectonics in the late Pliocene–early Pleistocene. Ital. J. Geosci. 132, 137–148. doi: 10.3301/IJG.2012.18
Biddittu, I., Cassoli, P. F., Radicati di Brozzolo, F., Segre, A. G., Segre-Naldini, E., and Villa, I. (1979). Anagni, a K-Ar dated lower and middle pleistocene site, Central Italy: preliminary report. Quaternaria 21, 53–71.
Bocherens, H. (2014). “Diet and ecology of the Scladina Neanderthal child: insights from stable isotopes,” in The Juvenile Neandertal Facial Remains from Scladina Cave, Vol 134. ed M. Toussaint, (Liège: ERAUL), 345–356.
Bocherens, H., Billiou, D., Patou-Mathis, M., Otte, M., Bonjean, D., Toussaint, M., et al. (1999). Palaeoenvironmental and palaeodietary implications of isotopic biogeochemistry of late interglacial Neandertal and mammal bones in Scladina Cave (Belgium). J. Archaeol. Sci. 26, 599–607. doi: 10.1006/jasc.1998.0377
Bocherens, H., Jacques, L., Ogle, N., Moussa, I., Kalin, R., Vignaud, P., et al. (2009). Reply to the comment by A. Zazzo, W.P Patterson and T.C. Prokopiuk on “Implications of diagenesis for the isotopic analysis of Upper Miocene large mammalian herbivore tooth enamel from Chad” by Jacques et al. (2008). Palaeogeography, Palaeoclimatology, Palaeoecology 266, 200–210. Palaeogeogr. Palaeoclimatol. Palaeoecol. 277, 269–271. doi: 10.1016/j.palaeo.2009.02.003
Bocherens, H., Koch, P. L., Mariotti, A., Geraads, D., and Jaeger, J.-J. (1996). Isotopic biogeochemistry (13C, 18O) of mammal enamel from African Pleistocene hominid sites: implications for the preservation of paleoclimatic isotopic signals. Palaios 11, 306–318. doi: 10.2307/3515241
Bodenstein, V., Meissner, H. H., and van Hoven, W. (2000). Food selection by Burchell's zebra and blue wildebeest in the Timbavati area of the Northern Province Lowveld. S. Afr. J. Wildl. Res. 30, 63–72.
Bryant, J. D., and Froelich, P. N. (1995). A model of oxygen isotope fractionation in body water of large mammals. Geochim. Cosmochim. Acta 59, 4523–4537. doi: 10.1016/0016-7037(95)00250-4
Bryant, J. D., Koch, P. L., Froelich, P. N., Showers, W. J, and Genna, B. J. (1996). Oxygen isotope partitioning between phosphate and carbonate in mammalian apatite. Geochim. Cosmochim. Acta 60, 5145–5148. doi: 10.1016/S0016-7037(96)00308-0
Caloi, L. (1997). New forms of equidsin Western Europe and palaeoenvironmental changes. Geobios 30, 267–284. doi: 10.1016/S0016-6995(97)80233-1
Calvin, M., and Benson, A. A. (1948). The path of carbon in photosynthesis. Science 107, 476–480. doi: 10.1126/science.107.2784.476
Carrara, C., Frezzotti, M., and Giraudi, C. (1995). “Stratigrafia plio-quaternaria,” in Lazio Meridionale, Sintesi Delle Ricerche Geologiche Multidisciplinari. ed C. Carrara (Roma: ENEA), 62–85.
Cerling, T. E., Harris, J. M., and Leakey, and, M. G. (1999). Browsing and grazing in elephants: the isotope record of modern and fossil proboscideans. Oecologia 120, 364–374. doi: 10.1007/s004420050869
Combourieu-Nebout, N., Bertini, A., Russo-Ermolli, E., Peyron, O., Klotz, S., Montade, V., et al. (2015). Climate changes in the central Mediterranean and Italian vegetation dynamics since the Pliocene. Rev. Palaeobot. Palynol. 218, 127–147. doi: 10.1016/j.revpalbo.2015.03.001
Dansgaard, W. (1964). Stable isotopes in precipitation. Tellus 16, 436–468. doi: 10.3402/tellusa.v16i4.8993
Delgado Huertas, A., Iacumin, P., Stenni, B., Sanchez Chillon, B., and Longinelli, A. (1995). Oxygen isotope variations of phosphate in mammalian bone and tooth enamel. Geochim. Cosmochim Acta 59, 4299–4305. doi: 10.1016/0016-7037(95)00286-9
DeMiguel, D. (2016). Disentangling adaptive evolutionary radiations and the role of diet in promoting diversification on islands. Sci. Rep. 6:29803. doi: 10.1038/srep29803
DeMiguel, D., Azanza, B., and Morales, J. (2010). Trophic flexibility within the oldest Cervidae lineage to persist through the Miocene Climatic Optimum. Palaeogeogr. Palaeoclimatol. Palaeoecol. 289, 81–92. doi: 10.1016/j.palaeo.2010.02.010
DeMiguel, D., Azanza, B., and Morales, J. (2011). Paleoenvironments and paleoclimate of the Middle Miocene of central Spain: a reconstruction from dental wear of ruminants. Palaeogeogr. Palaeoclimatol. Palaeoecol. 302, 452–463. doi: 10.1016/j.palaeo.2011.02.005
DeMiguel, D., Azanza, B., and Morales, J. (2018). Regional impacts of global climate change: a local humid phase in central Iberia in a late Miocene drying world. Palaeontology 62, 77–92. doi: 10.1111/pala.12382
DeMiguel, D., Fortelius, M., Azanza, B., and Morales, J. (2008). Ancestral feeding state of ruminants reconsidered: earliest grazing adaptation claims a mixed condition for Cervidae. BMC Evol. Biol. 8, 1–13. doi: 10.1186/1471-2148-8-13
DeMiguel, D., Quiralte, V., Azanza, B., Montoya, P., and Morales, J. (2012). Dietary behaviour and competition for vegetal resources in two Early Miocene pecoran ruminants from Central Spain. Geodiversitas 34, 425–443. doi: 10.5252/g2012n2a10
Drucker, D. G., Bridault, A., Hobson, K. A., Szuma, E., and Bocherens, H. (2008). Can carbon- 13 abundances in large herbivores track canopy effect in temperate and boreal ecosystems? Evidence from modern and ancient ungulates. Palaeogeogr. Palaeoclimatol. Palaeoecol. 266, 69–82. doi: 10.1016/j.palaeo.2008.03.020
Feranec, R. S., Hadly, E. A., and Paytan, A. (2009). Stable isotopes reveal seasonal competition for resources between late Pleistocene bison (Bison) and horse (Equus) from Rancho La Brea, southern California. Palaeogeogr. Palaeoclimatol. Palaeoecol. 271, 153–160. doi: 10.1016/j.palaeo.2008.10.005
Feranec, R. S., and MacFadden, B. J. (2006). Isotopic discrimination of resource partitioning among ungulates in C3-dominated communities from the Miocene of Florida and California. Paleobiology 32, 191–205. doi: 10.1666/05006.1
Forsten, A. (1988). Middle Pleistocene replacement of stenonid horses by caballoid horses — ecological implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 65, 23–33. doi: 10.1016/0031-0182(88)90109-5
Fortelius, M., Eronen, J., Liu, L., Pushkina, D., Tesakov, A., Vislobokova, I., et al. (2006). Late Miocene and Pliocene large land mammals and climatic changes in Eurasia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 238, 219–227. doi: 10.1016/j.palaeo.2006.03.042
Fortelius, M., and Solounias, N. (2000). Functional characterization of ungulate molars using the abrasion-attrition wear gradient: a new method for reconstructing paleodiets. Am. Mus. Novit. 3301, 1–35. doi: 10.1206/0003-0082(2000)301<0001:FCOUMU>2.0.CO;2
Galadini, F., and Messina, P. (2004). Early-Middle Pleistocene eastward migration of the Abruzzi Apennine (central Italy) extensional domain. J. Geodyn. 37, 57–81. doi: 10.1016/j.jog.2003.10.002
Hatch, M. D., Slack, C. R., and Johnson, H. S. (1967). Further studies on a new pathway of photosynthetic carbon dioxide fixation in sugarcane, and its occurrence in other species. Biochem. J. 102, 417–422. doi: 10.1042/bj1020417
Head, M., and Gibbard, P. (2015). Early–Middle Pleistocene transitions: linking terrestrial and marine realms. Quat. Int. 389, 7–46. doi: 10.1016/j.quaint.2015.09.042
Hofman-Kaminska, E., Bocherens, H., Borowik, T., Drucker, D. G., and Kowalczyk, R. (2018). Stable isotope signatures of large herbivore foraging habits across Europe. PLoS ONE 13:e0190723. doi: 10.1371/journal.pone.0190723
Hoppe, K. A., Stover, S. M., Pascoe, J. R., and Amundson, R. (2004). Tooth enamel biomineralization in extant horses: implications for isotopic microsampling. Palaeogeogr. Palaeoclimatol. Palaeoecol. 206, 355–365. doi: 10.1016/j.palaeo.2004.01.012
Jacques, L., Ogle, N., Moussa, I., Kalin, R., Vignaud, P., Brunet, M., et al. (2008). Implications of diagenesis for the isotopic analysis of upper Miocene large mammalian herbivore tooth enamel from Chad. Palaeogeogr. Palaeoclimatol. Palaeoecol. 266, 200–210. doi: 10.1016/j.palaeo.2008.03.040
Kahlke, R.-D., García, N., Kostopoulos, D. S., Lacombat, F., Lister, A. M., Mazza, P. P. A., et al. (2011). Western Palaearctic palaeoenvironmental conditions during the Early and early Middle Pleistocene inferred from large mammal communities, and implications for hominin dispersal in Europe. Quat. Sci. Rev. 30, 1368–1395. doi: 10.1016/j.quascirev.2010.07.020
Kaiser, T., and Franz-Odendaal, T. (2004). A mixed-feeding Equus species from the middle pleistocene of South Africa. Quat. Res. 62, 316–323. doi: 10.1016/j.yqres.2004.09.002
Kaiser, T. M., and Solounias, N. (2003). Extending the tooth mesowear method to extinct and extant equids. Geodiversitas 25, 321–345.
Kartzinel, T. R., Chen, P. A., Coverdale, T. C., Erickson, D. L., Kress, W. J., Kuzmina, M. L., et al. (2015). DNA metabarcoding illuminates dietary niche partitioning by African large herbivores. Proc. Natl. Acad. Sci. U.S.A. 112, 8019–8024. doi: 10.1073/pnas.1503283112
Koch, P. L. (1998). Isotopic reconstruction of past continental environments. Ann. Rev. Earth Planetary Sci. 26, 573–613. doi: 10.1146/annurev.earth.26.1.573
Kohn, M. J. (1996). Predicting animal d18O: accounting for diet and physiological adaptation. Geochim. Cosmochim. Acta 60, 4811–4829. doi: 10.1016/S0016-7037(96)00240-2
Lang, N., and Wolff, E. W. (2011). Interglacial and glacial variability from the last 800 ka in marine, ice and terrestrial archives. Clim. Past. 7, 361–380. doi: 10.5194/cp-7-361-2011
Lécuyer, C., Balter, V., Martineau, F., Fourel, F., Bernard, A., Amiot, R., et al. (2010). Oxygen isotope fractionation between apatite-bound carbonate and water determined from controlled experiments with synthetic apatites precipitated at 10–37 C. Geochim. Cosmochim. Acta 74, 2072–2081. doi: 10.1016/j.gca.2009.12.024
Levin, N. E., Cerling, T. E., Passey, B. H., Harris, J. M., and Ehleringer, J. R. (2006). A stable isotope aridity index for terrestrial environments. Proc. Natl. Acad. Sci. U.S.A 103, 11201–11205. doi: 10.1073/pnas.0604719103
Lindsay, E. H., Opdyke, N. O., and Johnson, N. M. (1980). Pliocene dispersal of the horse Equus and late Cenozoic mammalian dispersal events. Nature 287, 135–138. doi: 10.1038/287135a0
Lisiecki, L. E., and Raymo, M. E. (2005). A Pliocene-Pleistocene stack of 57 globally distributed benthic δ18O records. Paleoceanography 20, 1–17. doi: 10.1029/2004PA001071
Lisiecki, L. E., and Raymo, M. E. (2007). Plio-Pleistocene climate evolution: trends and transition in glacial cycle dynamics. Quat. Sci. Rev. 26, 56–69. doi: 10.1016/j.quascirev.2006.09.005
Longinelli, A. (1984). Oxygen isotopes in mammal bone phosphate: a new tool for paleohydrological and paleoclimatological research? Geochim. Cosmochim. Acta 48, 385–390. doi: 10.1016/0016-7037(84)90259-X
Luz, B., Kolodny, Y., and Horowitz, M. (1984). Fractionation of oxygen isotopes between mammalian bone-phosphate and environmental drinking water. Geochim. Cosmochim. Acta 48, 1689–1693. doi: 10.1016/0016-7037(84)90338-7
Maniakas, I., and Kostopoulos, D. (2017). Morphometric-palaeoecological discrimination between Bison populations of the western Palaearctic. Geobios 50, 155–171. doi: 10.1016/j.geobios.2017.01.001
Manzi, G. (2016). Humans of the Middle Pleistocene: the controversial calvarium from Ceprano (Italy) and its significance for the origin and variability of Homo heidelbergensis. Quat. Int. 411, 254–261. doi: 10.1016/j.quaint.2015.12.047
Manzi, G., Magri, D., and Palombo, M. (2011). Early–Middle Pleistocene environmental changes and human evolution in the Italian peninsula. Quat. Sci. Rev. 30, 1420–1438. doi: 10.1016/j.quascirev.2010.05.034
Marín-Leyva, A., DeMiguel, D., García-Zepeda, M., Ponce-Saavedra, J., Arroyo-Cabrales, J., Schaaf, P., et al. (2016). Dietary adaptability of Late Pleistocene Equus from West Central Mexico. Palaeogeogr. Palaeoclimatol. Palaeoecol. 441, 748–757. doi: 10.1016/j.palaeo.2015.10.019
McClymont, E. L., Sosdian, S. M., Rosell-Mel,é, A., and Rosenthal, Y. (2013). Pleistocene seasurface temperature evolution: early cooling, delayed glacial intensification, and implications for the mid-Pleistocene climate transition. Earth-Sci. Rev. 123, 173–193. doi: 10.1016/j.earscirev.2013.04.006
Mihlbachler, M., Rivals, F., Solounias, N., and Semprebon, G. (2011). Dietary change and evolution of horses in North America. Science 331, 1178–1181. doi: 10.1126/science.1196166
Muttoni, G., Scardia, G., Kent, D., Swisher, C., and Manzi, G. (2009). Pleistocene magnetochronology of early hominin sites at Ceprano and Fontana Ranuccio, Italy. Earth Planet. Sci. Lett. 286, 255–268. doi: 10.1016/j.epsl.2009.06.032
Palombo, M. R., Alberdi, M. T., Bellucci, L., and Sardella, R. (2017). An intriguing middle-sized horse from Coste San Giacomo (Anagni Basin, central Italy). Quat. Res. 87, 347–362. doi: 10.1017/qua.2017.6
Pereira, A., Nomade, S., Moncel, M., Voinchet, P., Bahain, J., Biddittu, I., et al. (2018). Integrated geochronology of Acheulian sites from the southern Latium (central Italy): Insights on human-environment interaction and the technological innovations during the MIS 11-MIS 10 period. Quat. Sci. Rev. 187, 112–129. doi: 10.1016/j.quascirev.2018.03.021
Pushkina, D., Bocherens, H., and Ziegler, R. (2014). Unexpected palaeoecological features of the Middle and Late Pleistocene large herbivores in southwestern Germany revealed by stable isotopic abundances in tooth enamel. Quat. Int. 339–340, 164–178. doi: 10.1016/j.quaint.2013.12.033
Rivals, F., and Athanassiou, A. (2008). Dietary adaptations in an ungulate community from the late Pliocene of Greece. Palaeogeogr. Palaeoclimatol. Palaeoecol. 265, 134–139. doi: 10.1016/j.palaeo.2008.05.001
Rivals, F., Julien, M., Kuitems, M., Van Kolfschoten, T., Serangeli, J., Drucker, D., et al. (2015b). Investigation of equid paleodiet from Schöningen 13 II-4 through dental wear and isotopic analyses: Archaeological implications. J. Hum. Evol. 89, 129–137. doi: 10.1016/j.jhevol.2014.04.002
Rivals, F., and Lister, A. (2016). Dietary flexibility and niche partitioning of large herbivores through the Pleistocene of Britain. Quat. Sci. Rev. 146, 116–133. doi: 10.1016/j.quascirev.2016.06.007
Rivals, F., Prignano, L., Semprebon, G., and Lozano, S. (2015a). A tool for determining duration of mortality events in archaeological assemblages using extant ungulate microwear. Sci. Rep. 5:17330. doi: 10.1038/srep17330
Rivals, F., Schulz, E., and Kaiser, T. (2008). Climate-related dietary diversity of the ungulate faunas from the middle Pleistocene succession (OIS 14-12) at the Caune de l'Arago (France). Paleobiology 34, 117–127. doi: 10.1666/07023.1
Rivals, F., and Semprebon, G. (2010). What can incisor microwear reveal about the diet of ungulates? Mammalia 74, 401–406. doi: 10.1515/mamm.2010.044
Rivals, F., and Ziegler, R. (2018). High-resolution paleoenvironmental context for human occupations during the Middle Pleistocene in Europe (MIS 11, Germany). Quat. Sci. Rev. 188, 136–142. doi: 10.1016/j.quascirev.2018.03.026
Roeder, J. (1999). Zur Ökologie des Bergzebras im südlichen Namibia. [dissertation/master's thesis], Ernst-Moritz-Arndt-University, Greifswald.
Rubini, M., Cerroni, V., Festa, G., Sardella, R., and Zaio, P. (2014). A revision of hominin fossil teeth from Fontana Ranuccio (Middle Pleistocene, Anagni, Frosinone, Italy). J. Hum. Evol. 77, 204–216. doi: 10.1016/j.jhevol.2014.09.002
Schulz, E., Kaiser, T., Stubbe, A., Stubbe, M., Samjaa, R., Batsaikhan, N., et al. (2007). Comparative demography and dietary resource partitioning of two wild ranging asiatic equid populations. Explor. Biol. Res. Mongol. 10, 77–90.
Segre, A. G., and Ascenzi, A. (1984). Fontana Ranuccio: Italy's earliest Middle Pleistocene hominid site. Curr. Anthropol. 25, 230–233. doi: 10.1086/203117
Skrzypek, G., Wiśniewski, A., and Grierson, P. F. (2011). How cold was it for Neanderthals moving to Central Europe during warm phases of the last glaciation? Quat. Sci. Rev. 30, 481–487. doi: 10.1016/j.quascirev.2010.12.018
Smith, B. N., and Epstein, S. (1971). Two categories of 13C/12C ratios for higher plants. Plant Physiol. 47, 380–384. doi: 10.1104/pp.47.3.380
Sponheimer, M., and Lee-Thorp, J. A. (1999). The ecological significance of oxygen isotopes in enamel carbonate. J. Archaeol. Sci. 26, 723–728. doi: 10.1006/jasc.1998.0388
Strani, F., DeMiguel, D., Alba, D., Moyà-Solà, S., Bellucci, L., Sardella, R., et al. (2019). The effects of the “0.9 Ma event” on the Mediterranean ecosystems during the Early-Middle Pleistocene transition as revealed by dental wear patterns of fossil ungulates. Quat. Sci. Rev. 210, 80–89. doi: 10.1016/j.quascirev.2019.02.027
Strani, F., DeMiguel, D., Bellucci, L., and Sardella, R. (2018b). Dietary response of early Pleistocene ungulate communities to the climate oscillations of the Gelasian/Calabrian transition in Central Italy. Palaeogeogr. Palaeoclimatol. Palaeoecol. 499, 102–111. doi: 10.1016/j.palaeo.2018.03.021
Strani, F., DeMiguel, D., Bona, F., Sardella, R., Biddittu, I., Bruni, L., et al. (2018a). Ungulate dietary adaptations and palaeoecology of the Middle Pleistocene site of Fontana Ranuccio (Anagni, Central Italy). Palaeogeogr. Palaeoclimatol. Palaeoecol. 496, 238–247. doi: 10.1016/j.palaeo.2018.01.041
Strani, F., DeMiguel, D., Sardella, R., and Bellucci, L. (2015). Paleoenvironments and climatic changes in the Italian Peninsula during the Early Pleistocene: evidence from dental wear patterns of the ungulate community of Coste San Giacomo. Quat. Sci. Rev. 121, 28–35. doi: 10.1016/j.quascirev.2015.05.008
Strani, F., DeMiguel, D., Sardella, R., and Bellucci, L. (2018c). Resource and niche differentiation mechanisms by sympatric Early Pleistocene ungulates: the case study of Coste San Giacomo. Quat. Int. 481, 157–163. doi: 10.1016/j.quaint.2017.08.064
Strani, F., Profico, A., Manzi, G., Pushkina, D., Raia, P., Sardella, R., et al. (2018d). MicroWeaR: a new R package for dental microwear analysis. Ecol. Evol. 8, 7022–7030. doi: 10.1002/ece3.4222
Van der Merwe, N. J., and Medina, E. (1991). The canopy effect, carbon isotope ratios and foodwebs in Amazonia. J. Archaeol. Sci. 18, 249–259. doi: 10.1016/0305-4403(91)90064-V
Keywords: mesowear, isotopes, equids, Equus, Pleistocene, paleoecology, paleoenvironment
Citation: Strani F, Pushkina D, Bocherens H, Bellucci L, Sardella R and DeMiguel D (2019) Dietary Adaptations of Early and Middle Pleistocene Equids From the Anagni Basin (Frosinone, Central Italy). Front. Ecol. Evol. 7:176. doi: 10.3389/fevo.2019.00176
Received: 29 January 2019; Accepted: 01 May 2019;
Published: 17 May 2019.
Edited by:
Gina Marie Semprebon, Bay Path University, United StatesReviewed by:
Nikos Solounias, New York Institute of Technology, United StatesCopyright © 2019 Strani, Pushkina, Bocherens, Bellucci, Sardella and DeMiguel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Flavia Strani, ZmxhdmlhLnN0cmFuaUB1bmlyb21hMS5pdA==; ZmxhdmlhLnN0cmFuaUBnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.