 Emilie Dion
Emilie Dion Antónia Monteiro
Antónia Monteiro Caroline M. Nieberding
Caroline M. Nieberding- 1Department of Biological Sciences, National University of Singapore, Singapore, Singapore
- 2Yale-NUS College, Singapore, Singapore
- 3Evolutionary Ecology and Genetics Group, Biodiversity Research Center, Earth and Life Institute, UCLouvain, Louvain-la-Neuve, Belgium
We review experimental and theoretical evidence that learning in insects and spiders affects the expression of mate preferences and of sexual signals, the evolution of both traits, and ultimately patterns of assortative mating, and speciation. Both males and females can modify their sexual preferences and signaling based on previous social interactions or the experience of visual, olfactory, gustatory, or auditory signals. Learning takes place during an early life exposure, previous personal sexual experiences or by observing the choices of others, and it can occur sometimes via very short (a few seconds) exposures to individuals or signals. We briefly review some of the molecular mechanisms that mediate learning in insects, as well as theoretical work that assesses how learning impacts the evolution of insect sexual traits and speciation. We suggest that future research should attempt to provide evidence of the adaptive nature of learning, which remains scarce in insects as well as in vertebrates, and explore further the mechanisms of learning in order to probe into their possible transgenerational inheritance. Future studies should also model how this process might further affect the evolution of sexual traits, and provide a unifying terminology for the underlying mechanisms of learning across diverse life-history contexts.
Introduction
Sexual behaviors such as the expression of a mate preference or the expression of a sexual signal are often not fixed but can be modified through social experience in both vertebrates and invertebrates. This leads to animals learning a mate preference or learning to display a sexual signal such as a courtship dance or the release of a pheromone blend. Learning sexual behaviors has been accepted for quite some time in mammals and birds, where most research has been conducted (Hebets and Sullivan-Beckers, 2010; Verzijden et al., 2012; Morand-Ferron and Quinn, 2015; Servedio, 2015; Head et al., 2016), including early work by Konrad Lorenz on learned sexual preferences via imprinting. However, learning in sexual selection remained controversial for insects and other arthropods until recently (Dukas, 2006, 2008a). This stems from insects being thought of as having fixed sexual behaviors due to their short lives and few mating opportunities, limiting their possibilities for learning or its likely adaptive value. However, a large number of more recent studies have illustrated that both insects and spiders modify their behavioral sexual interactions upon previous experience. Furthermore, it is now abundantly clear that many species of insects mate multiply, and have complex brain structures allowing short and long term memory of previous experiences that impact their lifetime mating behavior (Dukas, 2006, 2008a; Chittka and Niven, 2009).
The effect of social experiences, or simply the exposure to a sexual signal such as a pheromone, on the expression of mate preferences and sexual signals has been described in different terms by different authors (Tables 1, 2), from “learning” and “courtship conditioning,” to “mate copying,” to “exposure,” to “premating interaction,” “social learning,” “experience,” “eavesdropping,” “mate preference learning,” and “learned mate recognition,” among a few. This diverse terminology reflects a burgeoning field for insects and spiders over the last decade, as well as the diversity of learning mechanisms that may be at work in these animals. Regardless of terminology (Table 3), what all these cases have in common is a significant change in the expression of mating behaviors that results from a prior social experience or previous exposure to a sexual signal.
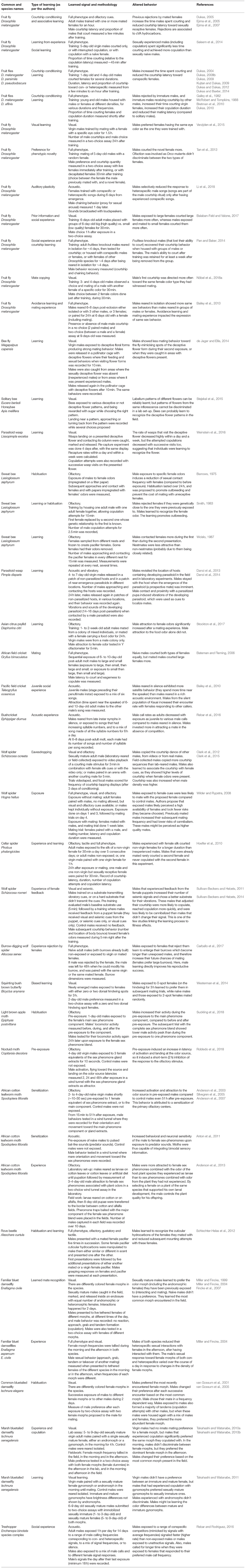
Table 1. A non-exhaustive list of publications about how male insects and spiders learn sexual behaviors.
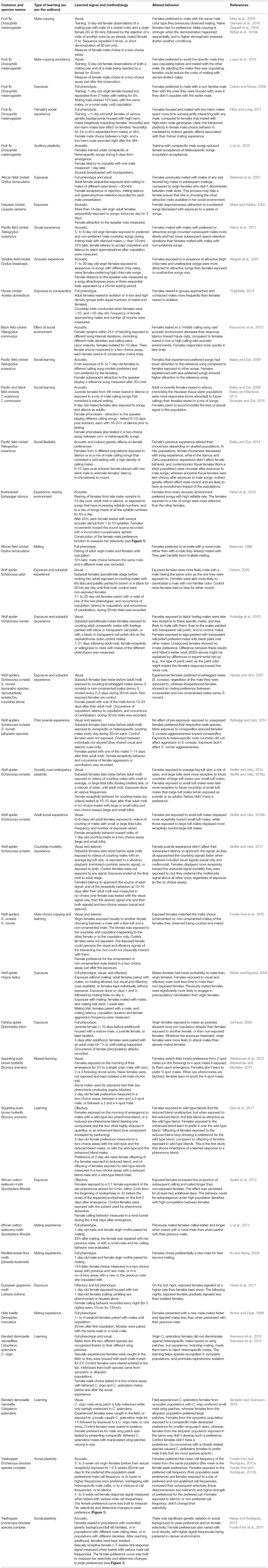
Table 2. A non-exhaustive list of publications about how female insects and spiders learn sexual behaviors.
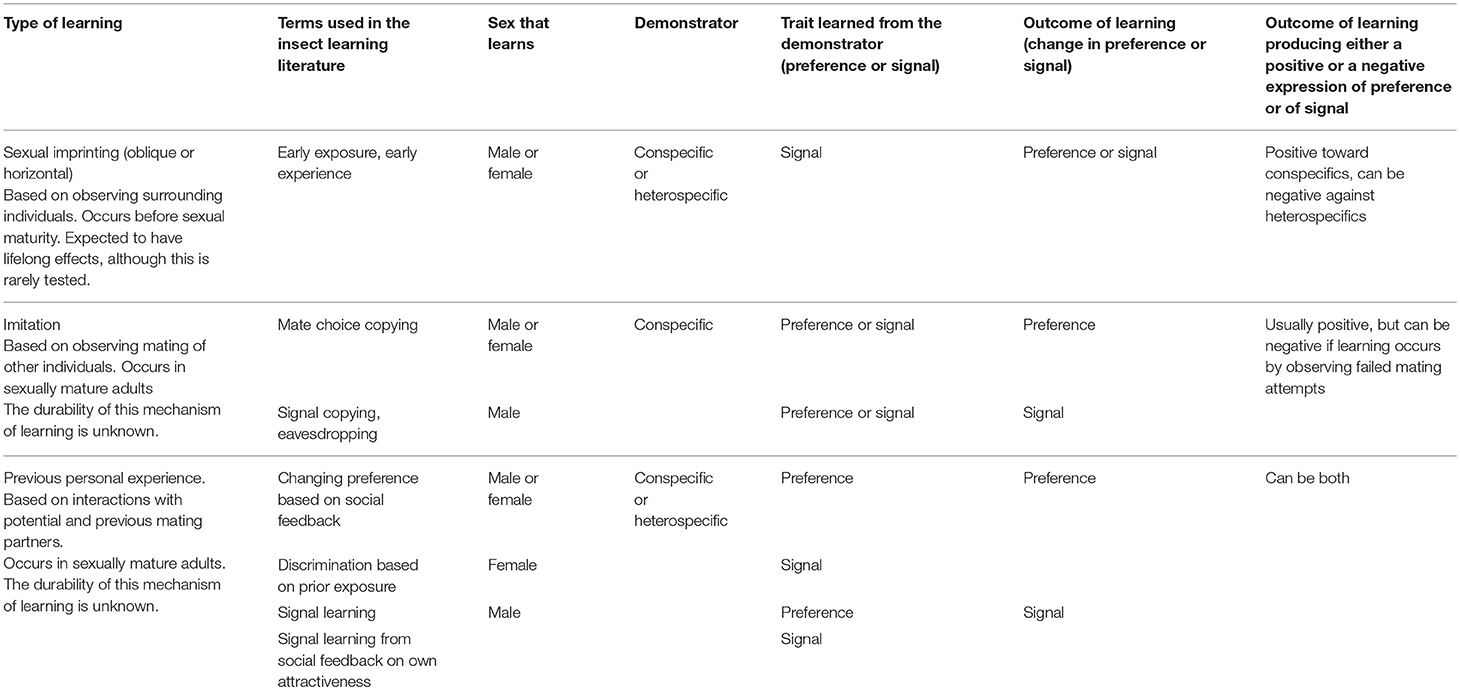
Table 3. Summary of the different types of learning in sexual selection in insects and spiders. Modified from (Yeh and Servedio, 2015) and (Varela et al., 2018).
Our goals in this review are to: (1) highlight the multiple types of information that insects learn that later result in changes in their mating preferences or in the expression of their sexual signals; (2) propose a systematic categorization of the underlying learning mechanisms, by building on a framework developed in vertebrates (Table 3); (3) review some of the molecular mechanisms underlying the learning process; (4) review some of the relevant mathematical models, originally applied to vertebrates, that suggest that learning mechanisms have consequences to the evolution of sexual traits and reproductive isolation in insects and spiders. We end by suggesting opportunities for future research in this field.
Insects and Spiders Learn a Variety of Social Information and Signals That Impact Their Mating Behavior
Learning in the context of mating has been described in a large diversity of spider and insect species (Tables 1, 2). Multiple definitions of learning were used across disciplines, but here, we define learning broadly as being a change in the future sexual behavior of an individual resulting from a previous social experience (Thorpe, 1963; Barron et al., 2015). This definition encompasses all described processes of learning (Table 3), for which the adaptive value of the changed behavior has usually not been demonstrated. Learning often involves changes to mating preferences or to speed of sexual response in females, while it usually involves changes in the level of expression of sexual signals in males. In the majority of investigated cases, and perhaps contrary to what is often assumed, naïve females often do not develop a sexual preference until they are exposed to other members of their species or of closely related species (Figure 1A). The absence of a naïve preference appears to be especially common for visual sexual signals, which have been the topic of most research. However, when an innate sexual preference is observed, females can also modify it (Figure 1B) or become more selective (narrow their preference regarding potential mates) (Figure 1C). In males, learning can lead to changes in the courtship intensity, latency to court, target of courtship, or sometimes the expression of the sexual signals.
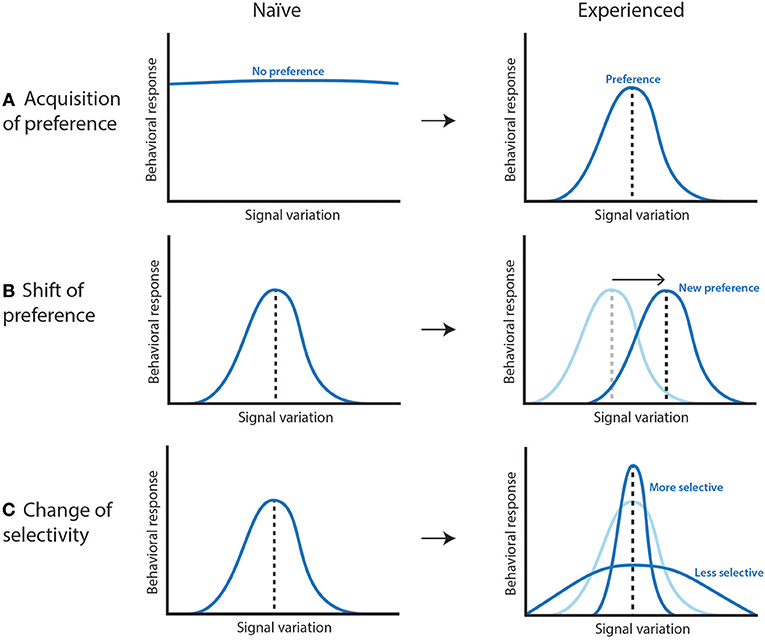
Figure 1. Illustration of how learning can affect a mate preference function. All graphs describe the mate preference of an individual to increasing amount of a sexual signal. (A) Individuals “acquire a preference” when they do not have an innate preference, but acquire one through experience. (B) Individuals “shift their preference” if they have an innate preference for one signal which is changed to another one after experiencing it. (C) A “change of selectivity” involves other types of modifications of the mate preference function, which depends on the individual responsiveness, tolerance and strength of response to a continuously distributed trait. Details on the measure of selectivity can be found in Fowler-Finn and Rodríguez (2012a,b) [Figure adapted from Fowler-Finn and Rodríguez (2012b)].
Most experimental evidence of learning in altering sexual behavior in insects comes from early exposure of sexually immature adults to the phenotypes of surrounding individuals, what has been called “sexual imprinting” in vertebrates. However, sexual behaviors can also vary after the observation of the interaction between other mating individuals, such as in cases of mate-choice copying (e.g., Mery et al., 2009) or imitation of sexual signaling (e.g., Clark et al., 2012, 2015), or from previous sexual interactions with or without mating (Tables 1–3). Most studies do not demonstrate that male or female behavioral shifts have an adaptive value, but such value is often assumed. Below, we illustrate this growing experimental evidence of learning in arthropods organized by the type of learned information: social information with unidentified individual signals (when the full phenotype of the interacting individuals is provided to the “learner”), or of specific signals such as olfactory, tactile, visual, and acoustic including vibratory signals (see Tables 1, 2 for a more complete overview, Figure 2).
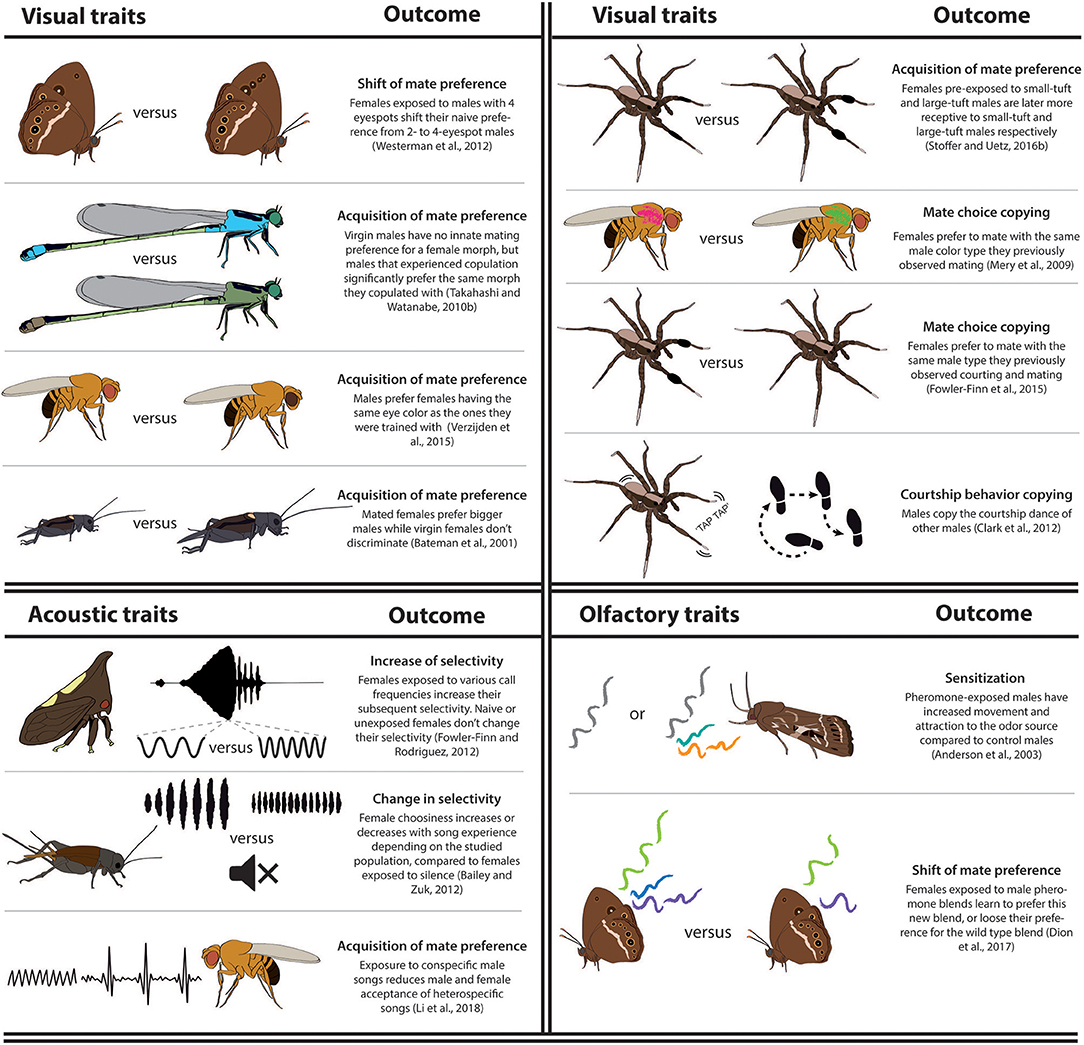
Figure 2. Illustrations of learning sexual traits or preferences in insects and spiders. Learning can produce an acquisition or a shift of preference, or a change in the insect selectivity for the sexual signal. These processes can be due to mate choice or sexual signaling copying or to sensitization, which is an increase of the individual attraction to the stimuli upon exposure. All examples illustrate changes in the mate preference function, except for the courtship behavior copying, which illustrates a change in sexual signaling.
Social Exposure Without Clearly Identified Learned Signals
Authors report that a social experience, with or without mating, affects subsequent sexual interactions of focal individuals. In most of these studies, adults are exposed to the full phenotype of other individuals where the specific assessed and learned signal(s) are not clearly identified. For example, virgin female crickets reared in isolation approach and contact males less frequently than virgin females reared in a group (Tinghitella, 2014). Being housed with groups of males or females induces fruitless knockout Drosophila males, which have lost their ability to court, to recover their courtship behavior, and wild type males to reduce their same-sex sexual behaviors (Bailey et al., 2013; Pan and Baker, 2014). Burrow-digging spider males enlarged their burrows upon rejections by females, which increases their chances of mating, as females prefer larger burrows (Carballo et al., 2017).
The mating status or novelty of the interacting individuals appears to be particularly important in modifying a focal individual's subsequent behavior. For instance, naive female crickets and spiders are more likely to mate, copulate more quickly or cannibalize fewer males than their mated counterparts (Johnson, 2005; Wilder and Rypstra, 2008; Judge et al., 2010). Male fruit flies learn to focus their courtship toward receptive conspecific females, and out-compete sexually inexperienced males, based on previous copulations (Saleem et al., 2014), or based on rejections by mated or virgin females (Dukas, 2005; Ejima et al., 2005; Griffith and Ejima, 2009). These males also learn from rejections from heterospecific females (Dukas, 2004, 2006, 2008b, 2009; Kujtan and Dukas, 2009; Dukas and Dukas, 2012; Dukas and Baxter, 2014) or immature males (Gailey et al., 1982; McRobert and Tompkins, 1988; Bretman et al., 2010). Learning to quickly discriminate receptive from unreceptive individuals is likely adaptive as males can reduce the costs of unsuccessful courting and mating with an unreceptive female or a male. Females in many species also prefer a new male over their previous mate. For instance, in crickets (Bateman, 1998), in moths (Xu and Wang, 2009; Li et al., 2014), in hide beetles (Archer and Elgar, 1999), or in female Drosophila melanogaster, a simple exposure without mating is enough to trigger a similar preference for a new male (Odeen and Moray, 2008; Loyau et al., 2012). This type of learning might also be adaptive as by rejecting the males that they saw copulating, females could reduce the costs of mating with semen-limited males (Loyau et al., 2012).
Visual Signals
Multiple studies have shown that naïve individuals have no innate genetic mate preferences for particular visual signals but develop these through learning. In many cases, naïve males direct their courtship toward a wide range of females, while inexperienced females display no preference for a specific male visual trait. For instance, male fruit flies, who initially court both large and small females with equal vigor, will preferentially court one of these female types if previously exposed or mated to them (Balaban-Feld and Valone, 2017). In wolf spiders, juvenile or adult exposure to male tibia types is necessary to limit the female preference to a specific leg tuft size or color (Hebets, 2003, 2007; Rutledge et al., 2010; Stoffer and Uetz, 2015, 2016a,b). Male damselflies learn to prefer the female color morphs they previously interacted with (Miller and Fincke, 1999, 2004; van Gossum et al., 2001; Fincke et al., 2007; Takahashi and Watanabe, 2010b, 2011). Experienced female damselflies learn to reject heterospecific males by recognizing their wing patches (Svensson et al., 2010, 2014; Verzijden and Svensson, 2016). The acquisition of a preference for visual traits has also been reported in fruit flies [eye color, (Verzijden et al., 2015)], butterflies [hindwing ornamentation number, (Westerman et al., 2014)], or crickets [size, (Bateman et al., 2001)] (Figure 2). Recent studies on mate-choice copying showed that virgin individuals tend to prefer male phenotypes with similar color type and ornamentation as the mate choice of another conspecific, e.g., in spiders, (Fowler-Finn et al., 2015) and fruit flies, (Mery et al., 2009; Nöbel et al., 2018a). These cases illustrate that insects can generalize socially learned public information for choosing a mate. Finally, male spiders can also copy the leg-tapping dance of other courting males to increase their likelihood of seducing a female (Clark et al., 2012, 2015; Figure 2). These males reach copulation more quickly and are less likely to be cannibalized (Sullivan-Beckers and Hebets, 2011, 2014).
In the few documented cases of insects displaying innate (naïve) visual signal preferences, learning can be context dependent, and sometimes may override, or even reverse these preferences. For instance, female Bicyclus anynana butterflies can switch their naive preference from males with two UV-reflective spots on their forewings to four spots if they are exposed to these males shortly during sexual maturation Westerman et al. (2012). Importantly, learning is context dependent, as female butterflies learn to avoid, rather than prefer, the novel wing patterns when the male sex pheromone is absent in the training male (Westerman and Monteiro, 2013). This suggests that olfactory communication may trump visual communication in assessing mates, at least in B. anynana butterflies. In addition, learning can be biased in that some sexual signals (supernumerous eyespots) can induce an increased preference while exposure to others (fewer eyespots) does not modify innate mate preferences (Westerman et al., 2012).
Olfactory Signals
In multiple studies, the learned sexual signal that triggers a behavioral change is an olfactory signal. Innate preferences appear to be generally present for olfactory signals in insects, and they can trigger a wide range of behaviors in receivers of many species from the same or the opposite gender. Odor learning can lead to either habituation or to sensitization, which led to opposite behavioral responses, but it is still unclear how the two processes work. Already ~35 years ago, studies reported how male sweat bees avoided mating with previous mates, or with females genetically close to their first mate, by learning to recognize their particular odor (Barrows, 1975; Smith, 1983; Wcislo, 1987). Similar processes were recently reported in rove beetles and fruit flies (Schlechter-Helas et al., 2012; Tan et al., 2013). Such “habituation” to female odor also occurs in male moths, which reduce their level of response to female sex pheromones shortly after exposure to components of the blend (e.g., Robledo et al., 2018; Suckling et al., 2018). On the contrary, increased behavioral sensitivity to sex pheromones, termed “sensitization,” was also described in Spodoptera littoralis moths whereby sexually mature adult males increase their responsiveness and attraction to the odor source 24 h after exposure (Anderson et al., 2003, 2007; Silvegren et al., 2005). Male S. littoralis also learned to prefer a mating site based on a learned plant odor, and reproduced preferentially with females found on the same plant where they grew up as larvae, or on plants where they previously mated (Anderson et al., 2013; Thöming et al., 2013; Proffit et al., 2015). Female B. anynana butterflies not only become sensitized to wild-type male sex pheromone composition, but can also learn to prefer unattractive blends if exposed to these blends during sexual maturation, right after adult emergence (Dion et al., 2017; Figure 2). Female moths also perceive the sex pheromone of other females, which induce them to emit their own sex pheromone earlier and at higher amount than inexperienced individuals (Stelinski et al., 2006; Sadek et al., 2012). Finally, the presence of antiaphrodisiacs, transferred by males onto the female cuticle, and reproductive tract led other males to learn to avoid mated Drosophila females (Ejima et al., 2005, 2007).
Acoustic Signals
Multiple studies have shown a change in sexual behavior upon exposure to acoustic signals, which are often used in species recognition and mate quality assessment in insects (Hedwig, 2016). For example, the rate and number of male calls that female crickets hear as juveniles or during mating significantly affects their preference and their response speed to future mate calls (Wagner et al., 2001; Rebar et al., 2011; Kasumovic et al., 2012). Contrary to individuals exposed to a mixture of call frequencies, females reared in silence respond faster toward a model song mimicking the populations' average calling rate (Bailey and Zuk, 2008, 2009; Bailey and Macleod, 2014; Swanger and Zuk, 2015). Males reared in silence intercept more females attracted to other males' calls and increase their own call rates (Bailey et al., 2010). Changes in a females' response to acoustic experience are variable and population-specific (Bailey and Zuk, 2012). The acoustic environment also impacts female treehopper's preference selectivity (Figure 1C) for male signal frequency and speed (Fowler-Finn and Rodríguez, 2012a,b; Rebar and Rodriguez, 2016; Fowler-Finn et al., 2017). In addition, naïve female Drosophila initially show no preference to the courtship songs (wing vibrations) of conspecific or heterospecific males, but a pre-exposure to conspecific songs makes them prefer this song type (Li et al., 2018). In some parasitoid wasp species, males identify host pupae parasitized by a conspecific using acoustic and vibratory signals, learn their location, and visit them regularly, as a strategy to attain prospective emerging female mates (Danci et al., 2013, 2014). This is one of the few cases where the adaptive value of learning is highlighted.
Molecular Mechanisms of Learning
As detailed above, insects can change their sexual preferences and signaling upon social experiences and exposure to a variety of visual, odor, gustatory, or auditory signals, indicating that sexual behaviors are not fixed but plastic. The underlying molecular mechanisms that control this plasticity, however, are still largely unclear. Below we review a few mechanisms mediating such neural system plasticity.
A social learning experience, such as courtship conditioning, where males experience female rejection in response to courtship, can lead to long-term changes in the behavior of males. This process of long-term memory consolidation in male Drosophila appears to depend on a peak of the ecdysteroid hormone, 20E, that appears immediately after the conditioning (Ishimoto et al., 2009).
Insects can learn to prefer (or avoid) a novel visual signal in a mate via early exposure to that signal but mechanisms of plasticity for preference development have only been explored in a non-sexual context. For instance, mRNA of three opsin genes in worker casts of the ant Camponotus rufipes increased upon exposure of these ants to daylight, as did volume of the three subneuropils of the optic lobe (including lamina, medulla, and lobula) (Yilmaz et al., 2016). A specific increase in UV and green opsin mRNA was also observed in the moth Helicoverpa armigera in response to 6 h exposure to UV light (Yan et al., 2014). These examples suggest that exposure of insects to particular visual signals displayed by the opposite sex could lead to changes in specific opsin expression levels as well as structural changes in the optic lobe, increasing sensitivity to those signals, and perhaps leading to later changes in sexual behaviors and preferences. This remains however to be investigated. Changes in protein expression levels and in cell size, cell number and cell connectivity of higher brain compartments in response to details of color patterns or courtship steps, rather than mere exposure to light of different colors, are also likely taking place but mediating mechanisms are still not known.
Mechanisms of pheromone odor sensitization have been explored to some extent in Spodoptera moths. In these experiments males are being briefly pre-exposed to a scent plume containing one or more components of the pheromone blend that increased their sensitization to the odor relative to naïve males (Anderson et al., 2003). The mechanisms that mediate this sensitization involve increases of the specific olfactory receptor expression and odor binding proteins in the antennae a few hours after the exposure (Wan et al., 2015), increased firing responses of the antennae (López et al., 2017) and of the odor receptor neurons (Guerrieri et al., 2012), as well as changes in the size of the neural compartment processing the pheromone components in the olfactory lobes (Guerrieri et al., 2012). These physiological and structural changes have been hypothesized to lead to long-term memory of the early odor experience and stable changes in behavior (Anderson et al., 2007; Guerrieri et al., 2012).
Males learn to recognize mated or heterospecific Drosophila females thanks to the presence of cuticular hydrocarbons and antiaphrodisiacs transferred by the previous male onto the female's cuticle and reproductive tract (Ejima et al., 2005; Billeter et al., 2009). Recent work has identified neuronal differences across Drosophila species that are responsible for species-specific mate preferences regarding a female cuticular pheromone sensed by the legs of males (Seeholzer et al., 2018). It is possible that this conserved neuronal circuit, which is activated differently across species, is plastic and will be later implicated in learning of novel cuticular pheromones within a species, but this remains to be tested.
Drosophila males produce courtship songs by vibrating their wings. The song frequencies are perceived by the tip of the antennae, which detects air particle oscillations, and are processed by the Johnston's organ, at the base of the antennae (Ishikawa and Kamikouchi, 2016). The mechanisms that mediate the female's development of a song preference are still largely unknown but they involve signaling via the main inhibitory neurotransmitter, gamma-Aminobutyric acid (GABA), acting on the GABAA receptor Rdl in specific neurons (Li et al., 2018).
Currently there is limited evidence that learned mate preferences can be transmitted to the next generation in an insect and even more limited understanding regarding mechanisms. Daughters of B. anynana females exposed to novel sex pheromone blends show naïve preferences similar to those of their exposed mothers and different from non-exposed naïve individuals (Dion et al., 2017), but the mechanisms mediating the inheritance of this learned preference are unknown. Prolonged (5 day) olfactory conditioning in Drosophila was also inherited across two generations, but this work did not test the role of these learned odors on sexual behaviors (Williams, 2016).
Mathematical Models Show That Learning Has an Evolutionary Impact
In this section, we first briefly introduce models that assess whether learning itself can evolve and be selected as an evolutionary stable strategy (ESS), i.e., a strategy that if adopted by a population in a given environment cannot be invaded by any alternative strategy that is initially rare. If learning sexual traits is an ESS, then this implies that learning sexual traits is adaptive. Whether learning sexual traits affects their evolution and impacts speciation has also mostly been addressed with theoretical models that we also reviewed in this section.
The first set of models reveal that learning can be selected as an ESS, which is a prerequisite for learning to affect the evolution of sexual signals, mate preferences, and reproductive isolation (reviewed in Galef and Laland, 2005; Vakirtzis, 2011; Verzijden et al., 2012; Dukas, 2013; Servedio and Dukas, 2013; Witte et al., 2015; Head et al., 2016; Kopp et al., 2018; Varela et al., 2018). Overall, for learning to evolve under selection there needs to be genetic variation for learning ability within a species (Mery and Kawecki, 2005). This appears to be the case. An example involves the “rover” and “sitter” alleles at the foraging locus of D. melanogaster that confer different learning abilities to fly larvae when foraging for food (Mery et al., 2007; Papaj and Snell-Rood, 2007; Mery, 2013).
The second set of models assess whether learning affects the evolution of sexual traits and impacts speciation. These models were originally designed for sexual interactions in vertebrates, but here we focused on those models that can be applied to insects (Supplementary Table 1). Most of these models were built on the premise that sexual signals and mate preferences have a genetic basis (e.g., Ritchie, 2000; Shaw, 2000; Noor et al., 2001) that can be modified and be overridden by learning (Supplementary Table 1). The models usually focus on one of the following three underlying mechanisms of learning: “learning by sexual imprinting,” “learning by copying,” and “learning from previous experience” (Table 3 and Supplementary Table 1).
The first group of models focused on imprinting, which occurs when juveniles up to a certain age can learn a sexual preference by observing the phenotypes of surrounding adults (Immelmann, 1975; Head et al., 2016). In insects, the terms “early experience,” or “early exposure” to other individuals of the same generation are used instead of “imprinting” (Table 3). This is primarily because there is still no data on whether or not insects have a fixed period in development or early adulthood when they can learn a preference from a social experience, as in the case of birds, where the term imprinting was first used (Lorenz, 1935). In insects, sexual imprinting of mate preferences can occur between genetically unrelated individuals of the previous or of the same generation, and is termed oblique or horizontal imprinting, respectively (Table 3). Oblique or horizontal imprinting have limited effect on the evolution of sexual preferences and of reproductive isolation, except when spatial structure is taken into account. Spatial structure in models assumes that social learning is only possible between individuals that can perceive each other (Yeh and Servedio, 2015), i.e., that are close in space. Differentiation in social interactions between populations due to spatial isolation is expected to accelerate divergence of sexual preferences, signals, and of reproductive isolation between populations, through coupling of the divergent sexual signals and mate preferences across space (Bailey and Moore, 2012). If populations are exchanging migrants, oblique imprinting of mate preferences cannot produce sympatric speciation (Verzijden et al., 2007), and the populations cannot maintain genetic differentiation in their sexual traits (Yeh and Servedio, 2015). Interestingly, aversive learning of mate preference through oblique imprinting, when individuals learn to avoid a phenotype, was shown to accelerate reproductive isolation and to produce adaptive radiations (Gilman and Kozak, 2015). Finally, imprinting may contribute to reproductive isolation in insects through self-imprinting (i.e., self-referent phenotype matching) that facilitates reinforcement between incipient divergent lineages (Servedio et al., 2009).
Imprinting can also affect the expression of sexual signals, and these learned signals can also contribute to reproductive isolation (Williams and Slater, 1990; Ellers and Slabbekoorn, 2003; Lachlan and Servedio, 2004; Olofsson and Servedio, 2008; Olofsson et al., 2011; Planqué et al., 2014). These models often incorporate a spatial structure, but it is unclear if they can be applicable to insects because they assume that males learn to produce their sexual signals by imitating adults surrounding them during their development, which has so far has been documented only in the wolf spider Schizocosa ocreata (Clark et al., 2012, 2015). These models also assume that assortative mating takes place between females and males that have learned to prefer, or express, a similar sexual signal by experiencing it locally, while assortative mating based on similarly preferred and expressed sexual traits does not occur in insects, as far as we know.
A second mechanism of learning sexual behaviors in insects is to copy another individual's mating decision. Insects that mate in groups, such as promiscuous or lekking species [e.g., some species of ants, bees, paper wasps, and butterflies (Litte, 1979; Wickman and Jansson, 1997; Velthuis et al., 2005; Izzo and Tibbetts, 2012; Prato and Soares, 2013)] can modify their mating preference or the production of their own sexual signals by observing the success of other individuals mating. Most work has focused on “mate choice copying” (“MCC” hereafter), which is usually modeled as “positive” such that individuals (usually females) learn to prefer the phenotype of males that they have observed mating earlier (Servedio and Kirkpatrick, 1996; Santos et al., 2017). The copying behavior itself can spread in a population both through direct (Dugatkin and Höglund, 1995; Stöhr, 1998) or indirect (Servedio and Kirkpatrick, 1996; Santos et al., 2017) selection. MCC has direct selective benefits, if it reduces the sampling costs and/or the error rate of mate choice (Dugatkin and Höglund, 1995; Stöhr, 1998; Agrawal, 2001), but also has indirect selective benefits (Kirkpatrick and Dugatkin, 1994; Servedio and Kirkpatrick, 1996; Santos et al., 2017). These indirect benefits arise because females that copy others are more likely to mate with males that are attractive to other females, spreading in the process genes for attractive sons and genes for daughters with the ability to copy others. MCC can also both increase the variance in male sexual signals (Wade and Pruett-Jones, 1990), and erode genetic variance by eliminating novel or rare male signals, even if these males are fitter than the common males in the population (Kirkpatrick and Dugatkin, 1994). However, when biases in learning are present in mate choice copying, such that females are more strongly affected by experiences involving unusual stimuli (e.g., rare male phenotypes) than those involving standard stimuli (e.g., common male phenotypes), MCC can cause novel male signals to sweep through the population even if there is no inherent preference for the novel trait (Agrawal, 2001). Invasion of a novel sexual signal can also occur when “negative” MCC (aversive learning) is modeled, where females learn to avoid males avoided by other females (Santos et al., 2014). While Kirkpatrick and Dugatkin (1994) suggest that MCC may promote or accelerate population divergence, the role of MCC in reproductive isolation and speciation remains an open question (Varela et al., 2018).
A third learning mechanism occurs through previous (so-called “private” or “personal”) experiences of either courtship or of actual mating events, during which mate preferences and the expression of sexual signals can be learned (Servedio and Dukas, 2013; Morier-Genoud and Kawecki, 2015). Females learning to prefer local or familiar (previously encountered) males increases the rate of divergence between spatially structured populations (Bailey and Moore, 2012), and also in case of a secondary contact (Servedio and Dukas, 2013). In contrast, when males learn to prefer local or familiar females, population divergence can be reduced because competition for accessing these females increases. Heterospecific males, which seldom meet heterospecific females locally, don't learn to prefer them as much as conspecific males, and keep courting and mating with both types of females (Servedio and Dukas, 2013). When males learn to improve the expression of their sexual signals through repeated courtship events, this accelerates the evolution of the sexual signal, even when the signal is costly, and it favors the emergence and spread of a novel male sexual signal, even in the presence of gene flow (Morier-Genoud and Kawecki, 2015).
Finally, some models compared the fitness advantage, or the likelihood of various mechanisms of mate preference learning to spread as evolutionary stable strategies (ESS). Learned mate preferences from previous encounters with potential mates increase fitness compared to other mate selection mechanisms including threshold-based mate preference (Dubois et al., 2012). Furthermore, the advantage provided by learning increases when variance in the quality among males increases locally, and across space or time (Collins et al., 2006). Depending on associated costs, learning mate preferences either through MCC or through previous personal experience can both coexist as an ESS in mixed populations of females displaying either one or the other learning mechanisms (Dubois et al., 2012).
Conclusions and Perspectives
Experimental work on the role of learning in sexual interactions in insects and spiders is a burgeoning field revealing that the traditional view that insects are small robots with mostly innate, genetically fixed sexual behaviors, is now obsolete. Learning in sexual interactions is the rule rather than the exception in every organism tested so far. Innate, genetically fixed sexual preferences and signals are present only in some species, and are more commonly observed in specific modes of communication such as olfactory signals, whereas learning a sexual preference or the expression of a sexual signal is widespread. Learning also affects the expression of innate sexual traits. Learning is usually assumed to be positive, but it can also be negative (i.e., aversive), as well as biased and context-dependent.
Our review revealed the diversity of terminology used by authors to describe experiments involving learning [Table 1, 2, column “Type of learning (as per the authors)”], which can be confusing and prevent the identification of the underlying mechanisms. Hence, we would like to encourage researchers to provide explicit details of their methods as described in Tables 1–3 (e.g., the developmental stage at which the learning happens, the sex of the demonstrator and of the insects that learns). This information will specify the processes of learning used by insects, e.g., sensitization, simple exposure, or conditioning, which will help identify the underlying neurophysiological mechanisms involved.
Finally, most scientists assume that learning sexual traits has evolved under selection, as appears to be the case in vertebrates (Morand-Ferron, 2017) and regarding other behaviors in insects (Nieberding et al., 2018), but there is little to no evidence that learning sexual traits affects insect fitness, particularly in the wild. This is perhaps because one of the first definition of learning included adaptation [learning is an “adaptive change in individual behavior as the result of experience;” Thorpe (1963)]. We encourage field work to complement laboratory experiments with ecologically-relevant setups to quantify the adaptive value of learned sexual interactions across insects. Showing the adaptive value of such learning would explain its prevailing presence in such miniature brained, short lived, organisms.
Author Contributions
ED, AM, and CN conceived the scope of the review, collected information from the literature, and contributed to the writing and editing of the manuscript. All authors contributed to revisions.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Maria Servedio for her helpful comments on the mathematical model part of this review, and Astrid Groot and Varvara Vedenina for their invitation to contribute to the special issue. This work was supported by the Ministry of Education, Singapore grants MOE2014-T2-1-146 and MOE2015-T2-2-159 to AM and ED, and by the University of Louvain-la-Neuve (UCL) and the Fédération Wallonie-Bruxelles (Grant ARC 17/22-086) to CN. This is publication BRC 340 of the Biodiversity Research Center at UCL.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2018.00225/full#supplementary-material
References
Agrawal, A. F. (2001). The evolutionary consequences of mate copying on male traits. Behav. Ecol. Sociobiol. 51, 33–40. doi: 10.1007/s002650100401
Anderson, P., Hansson, B. S., Nilsson, U., Han, Q., Sjoholm, M., Skals, N., et al. (2007). Increased behavioral and neuronal sensitivity to sex pheromone after brief odor experience in a moth. Chem. Senses 32, 483–491. doi: 10.1093/chemse/bjm017
Anderson, P., Sadek, M. M., and Hansson, B. S. (2003). Pre-exposure modulates attraction to sex pheromone in a moth. Chem. Senses 28, 285–291. doi: 10.1093/chemse/28.4.285
Anderson, P., Sadek, M. M., Larsson, M., Hansson, B. S., and Thöming, G. (2013). Larval host plant experience modulates both mate finding and oviposition choice in a moth. Anim. Behav. 85, 1169–1175. doi: 10.1016/j.anbehav.2013.03.002
Anton, S., Evengaard, K., Barrozo, R. B., Anderson, P., and Skals, N. (2011). Brief predator sound exposure elicits behavioral and neuronal long-term sensitization in the olfactory system of an insect. PNAS 108, 3401–3405. doi: 10.1073/pnas.1008840108
Archer, M. S., and Elgar, M. A. (1999). Female preference for multiple partners: sperm competition in the hide beetle, Dermestes maculatus (DeGeer). Anim. Behav. 58, 669–675. doi: 10.1006/anbe.1999.1172
Bailey, N. W., Gray, B., and Zuk, M. (2010). Acoustic experience shapes alternative mating tactics and reproductive investment in male field crickets. Curr. Biol. 20, 845–849. doi: 10.1016/j.cub.2010.02.063
Bailey, N. W., Hoskins, J. L., Green, J., and Ritchie, M. G. (2013). Measuring same-sex sexual behaviour: the influence of the male social environment. Anim. Behav. 86, 91–100. doi: 10.1016/j.anbehav.2013.04.016
Bailey, N. W., and Macleod, E. (2014). Socially flexible female choice and premating isolation in field crickets (Teleogryllus spp.). J. Evol. Biol. 27, 170–180. doi: 10.1111/jeb.12285
Bailey, N. W., and Moore, A. J. (2012). Runaway sexual selection without genetic correlations: social environments and flexible mate choice initiate and enhance the Fisher process. Evolution 66, 2674–2684. doi: 10.1111/j.1558-5646.2012.01647.x
Bailey, N. W., and Zuk, M. (2008). Acoustic experience shapes female mate choice in field crickets. Proc. Biol. Sci. 275, 2645–2650. doi: 10.1098/rspb.2008.0859
Bailey, N. W., and Zuk, M. (2009). Field crickets change mating preferences using remembered social information. Biol. Lett. 5, 449–451. doi: 10.1098/rsbl.2009.0112
Bailey, N. W., and Zuk, M. (2012). Socially flexible female choice differs among populations of the Pacific field cricket: geographical variation in the interaction coefficient psi (Psi). Proc. Biol. Sci. 279, 3589–3596. doi: 10.1098/rspb.2012.0631
Balaban-Feld, J., and Valone, T. J. (2017). Prior information and social experience influence male reproductive decisions. Behav. Ecol. 28, 1376–1383. doi: 10.1093/beheco/arx105
Barron, A. B., Hebets, E. A., Cleland, T. A., Fitzpatrick, C. L., Hauber, M. E., and Stevens, J. R. (2015). Embracing multiple definitions of learning. Trends Neurosci. 38, 405–407. doi: 10.1016/j.tins.2015.04.008
Barrows, E. M. (1975). Individually distinctive odors in an invertebrate. Behav. Biol. 15, 57–64. doi: 10.1016/S0091-6773(75)92079-9
Bateman, P. W. (1998). Mate preference for novel partners in the cricket Gryllus bimaculatus. Ecol. Entomol. 23, 473–475. doi: 10.1046/j.1365-2311.1998.00156.x
Bateman, P. W., and Fleming, P. A. (2006). Males are selective too: mating, but not courtship, with sequential females influences choosiness in male field crickets (Gryllus bimaculatus). Behav. Ecol. Sociobiol. 59, 577–581. doi: 10.1007/s00265-005-0083-y
Bateman, P. W., Gilson, L. N., and Ferguson, J. W. H. (2001). Male size and sequential mate preference in the cricket Gryllus bimaculatus. Anim. Behav. 61, 631–637. doi: 10.1006/anbe.2000.1617
Billeter, J.-C., Atallah, J., Krupp, J. J., Millar, J. G., and Levine, J. D. (2009). Specialized cells tag sexual and species identity in Drosophila melanogaster. Nature 461:987. doi: 10.1038/nature08495
Bretman, A., Fricke, C., Hetherington, P., Stone, R., and Chapman, T. (2010). Exposure to rivals and plastic responses to sperm competition in Drosophila melanogaster. Behav. Ecol. 21, 317–321. doi: 10.1093/beheco/arp189
Carballo, M., Baldenegro, F., Bollatti, F., Peretti, A. V., and Aisenberg, A. (2017). No pain, no gain: male plasticity in burrow digging according to female rejection in a sand-dwelling wolf spider. Behav. Processes 140, 174–180. doi: 10.1016/j.beproc.2017.05.007
Chittka, L., and Niven, J. (2009). Are bigger brains better? Curr. Biol. 19, R995–R1008. doi: 10.1016/j.cub.2009.08.023
Clark, D. L., Kizer Zeeff, C., Sabovodny, G., Hollenberg, A., Roberts, J. A., and Uetz, G. W. (2015). The role of social experience in eavesdropping by male wolf spiders (Lycosidae). Anim. Behav. 106, 89–97. doi: 10.1016/j.anbehav.2015.05.001
Clark, D. L., Roberts, J. A., and Uetz, G. W. (2012). Eavesdropping and signal matching in visual courtship displays of spiders. Biol. Lett. 8, 375–378. doi: 10.1098/rsbl.2011.1096
Collins, E. J., McNamara, J. M., and Ramsey, D. M. (2006). Learning rules for optimal selection in a varying environment: mate choice revisited. Behav. Ecol. 17, 799–809. doi: 10.1093/beheco/arl008
Dagaeff, A.-C., Pocheville, A., Nöbel, S., Loyau, A., Isabel, G., and Danchin, E. (2016). Drosophila mate copying correlates with atmospheric pressure in a speed learning situation. Anim. Behav. 121, 163–174. doi: 10.1016/j.anbehav.2016.08.022
Danci, A., Hrabar, M., Ikoma, S., Schaefer, P. W., and Gries, G. (2013). Learning provides mating opportunities for males of a parasitoid wasp. Entomol. Exp. Appl. 149, 229–240. doi: 10.1111/eea.12129
Danci, A., Inducil, C., Takács, S., Schaefer, P. W., and Gries, G. (2014). Mechanism of mate detection in parasitoid wasps: sound and vibratory cues change with the developmental progress of future mates inside host pupal cases. Physiol. Entomol. 39, 292–303. doi: 10.1111/phen.12075
de Jager, M. L., and Ellis, A. G. (2014). Costs of deception and learned resistance in deceptive interactions. Proc. Biol. Sci. 281:20132861. doi: 10.1098/rspb.2013.2861
Dion, E., Pui, L. X., and Monteiro, A. (2017). Early-exposure to new sex pheromone blend alters mate preference in female butterflies and in their offspring. Biorxiv [Preprint]. doi: 10.1101/214635
Dubois, F., Drullion, D., and Witte, K. (2012). Social information use may lead to maladaptive decisions: a game theoretic model. Behav. Ecol. 23, 225–231. doi: 10.1093/beheco/arr179
Dugatkin, L. A., and Höglund, J. (1995). Delayed breeding and the evolution of mate copying in lekking species. J. Theor. Biol. 174, 261–267. doi: 10.1006/jtbi.1995.0097
Dukas, R. (2004). Male fruit flies learn to avoid interspecific courtship. Behav. Ecol. 15, 695–698. doi: 10.1093/beheco/arh068
Dukas, R. (2005). Experience improves courtship in male fruit flies. Anim. Behav. 69, 1203–1209. doi: 10.1016/j.anbehav.2004.08.012
Dukas, R. (2006). Learning in the context of sexual behaviour in insects. Anim. Biol. 56, 125–141. doi: 10.1163/157075606777304258
Dukas, R. (2008a). Evolutionary biology of insect learning. Annu. Rev. Entomol. 53, 145–160. doi: 10.1146/annurev.ento.53.103106.093343
Dukas, R. (2008b). Learning decreases heterospecific courtship and mating in fruit flies. Biol. Lett. 4, 645–647. doi: 10.1098/rsbl.2008.0437
Dukas, R. (2009). Dynamics of learning in the context of courtship in Drosophila persimilis and D. pseudoobscura. Anim. Behav. 77, 253–259. doi: 10.1016/j.anbehav.2008.10.010
Dukas, R. (2010). Causes and consequences of male-male courtship in fruit flies. Anim. Behav. 80, 913–919. doi: 10.1016/j.anbehav.2010.08.017
Dukas, R. (2013). Effects of learning on evolution: robustness, innovation and speciation. Anim. Behav. 85, 1023–1030. doi: 10.1016/j.anbehav.2012.12.030
Dukas, R., and Baxter, C. M. (2014). Mate choosiness in young male fruit flies. Behav. Ecol. 25, 549–552. doi: 10.1093/beheco/aru020
Dukas, R., and Dukas, L. (2012). Learning about prospective mates in male fruit flies: effects of acceptance and rejection. Anim. Behav. 84, 1427–1434. doi: 10.1016/j.anbehav.2012.09.010
Ejima, A., Smith, B. P., Lucas, C., Levine, J. D., and Griffith, L. C. (2005). Sequential learning of pheromonal cues modulates memory consolidation in trainer-specific associative courtship conditioning. Curr. Biol. 15, 194–206. doi: 10.1016/j.cub.2005.01.035
Ejima, A., Smith, B. P., Lucas, C., Van der Goes van Naters, W., Miller, C. J., Carlson, J. R., et al. (2007). Generalization of courtship learning in Drosophila is mediated by cis-vaccenyl acetate. Curr. Biol. 17, 599–605. doi: 10.1016/j.cub.2007.01.053
Ellers, J., and Slabbekoorn, H. (2003). Song divergence and male dispersal among bird populations: a spatially explicit model testing the role of vocal learning. Anim. Behav. 65, 671–681. doi: 10.1006/anbe.2003.2081
Filice, D. C. S., and Long, T. A. F. (2017). Phenotypic plasticity in female mate choice behavior is mediated by an interaction of direct and indirect genetic effects in Drosophila melanogaster. Ecol. Evol. 7, 3542–3551. doi: 10.1002/ece3.2954
Fincke, O. M., Fargevieille, A., and Schultz, T. D. (2007). Lack of innate preference for morph and species identity in mate-searching Enallagma damselflies. Behav. Ecol. Sociobiol. 61, 1121–1131. doi: 10.1007/s00265-006-0345-3
Fowler-Finn, K. D., Cruz, D. C., and Rodríguez, R. L. (2017). Local population density and group composition influence the signal-preference relationship in Enchenopa treehoppers (Hemiptera: Membracidae). J. Evol. Biol. 30, 13–25. doi: 10.1111/jeb.12994
Fowler-Finn, K. D., and Rodríguez, R. L. (2012a). The evolution of experience-mediated plasticity in mate preferences. J. Evol. Biol. 25, 1855–1863. doi: 10.1111/j.1420-9101.2012.02573.x
Fowler-Finn, K. D., and Rodríguez, R. L. (2012b). Experience-mediated plasticity in mate preference: mating assurance in a variable environment. Evolution 66, 459–468. doi: 10.1111/j.1558-5646.2011.01446.x
Fowler-Finn, K. D., Sullivan-Beckers, L., Runck, A. M., and Hebets, E. A. (2015). The complexities of female mate choice and male polymorphisms: elucidating the role of genetics, age, and mate-choice copying. Curr. Zool. 61, 1015–1035. doi: 10.1093/czoolo/61.6.1015
Gailey, D. A., Jackson, F. R., and Siegel, R. W. (1982). Male courtship in Drosophila: the conditioned response to immature males and its genetic control. Genetics 102, 771–782.
Galef, B. G., and Laland, K. N. (2005). Social Learning in Animals: empirical studies and theoretical models. Bioscience 55, 489–499. doi: 10.1641/0006-3568(2005)055[0489:SLIAES]2.0.CO;2
Germain, M., Blanchet, S., Loyau, A., and Danchin, É. (2016). Mate-choice copying in Drosophila melanogaster: impact of demonstration conditions and male–male competition. Behav. Processes 125, 76–84. doi: 10.1016/j.beproc.2016.02.002
Gilman, R. T., and Kozak, G. M. (2015). Learning to speciate: the biased learning of mate preferences promotes adaptive radiation. Evolution 69, 3004–3012. doi: 10.1111/evo.12797
Griffith, L. C., and Ejima, A. (2009). Courtship learning in Drosophila melanogaster: diverse plasticity of a reproductive behavior. Learn. Mem. 16, 743–750. doi: 10.1101/lm.956309
Guerrieri, F., Gemeno, C., Monsempes, C., Anton, S., Jacquin-Joly, E., Lucas, P., et al. (2012). Experience-dependent modulation of antennal sensitivity and input to antennal lobes in male moths (Spodoptera littoralis) pre-exposed to sex pheromone. J. Exp. Biol. 215, 2334–2341. doi: 10.1242/jeb.060988
Harari, A. R., Zahavi, T., and Thiery, D. (2011). Fitness cost of pheromone production in signaling female moths. Evolution 65, 1572–1582. doi: 10.1111/j.1558-5646.2011.01252.x
Head, M. L., Jennions, M. D., and Zajitschek, S. R. K. (2016). Sexual selection: incorporating non-genetic inheritance. Curr. Opin. Behav. Sci. 12, 129–137. doi: 10.1016/j.cobeha.2016.10.005
Hebets, E. (2003). Subadult experience influences adult mate choice in an arthropod: exposed female wolf spiders prefer males of a familiar phenotype. PNAS 100, 13390–13395. doi: 10.1073/pnas.2333262100
Hebets, E. (2007). Subadult female experience does not influence species recognition in the wolf spider schizocosa uetzi stratton 1997. J. Arachnol. 35, 1–10. doi: 10.1636/S05-76.1
Hebets, E., and Sullivan-Beckers, L. (2010). “Mate choice and learning,” in Encyclopedia of Animal Behavior, ed. M.D.B.a.J. Moore. (Amsterdam: Elsevier), 389–393. doi: 10.1016/B978-0-08-045337-8.00364-8
Hebets, E. A., and Vink, C. J. (2007). Experience leads to preference: experienced females prefer brush-legged males in a population of syntopic wolf spiders. Behav. Ecol. 18, 1010–1020. doi: 10.1093/beheco/arm070
Hoefler, C. D., Moore, J. A., Reynolds, K. T., and Rypstra, A. L. (2010). The effect of experience on male courtship and mating behaviors in a cellar spider. Am. Midl. Nat. 163, 255–268. doi: 10.1674/0003-0031-163.2.255
Immelmann, K. (1975). Ecological significance of imprinting and early learning. Annu. Rev. Ecol. Syst. 6, 15–37. doi: 10.1146/annurev.es.06.110175.000311
Ishikawa, Y., and Kamikouchi, A. (2016). Auditory system of fruit flies. Hearing Res. 338, 1–8. doi: 10.1016/j.heares.2015.10.017
Ishimoto, H., Sakai, T., and Kitamoto, T. (2009). Ecdysone signaling regulates the formation of long-term courtship memory in adult Drosophila melanogaster. PNAS 106, 6381–6386. doi: 10.1073/pnas.0810213106
Izzo, A. S., and Tibbetts, E. A. (2012). Spotting the top male: sexually selected signals in male Polistes dominulus wasps. Anim. Behav. 83, 839–845. doi: 10.1016/j.anbehav.2012.01.005
Johnson, J. C. (2005). Cohabitation of juvenile females with mature males promotes sexual cannibalism in fishing spiders. Behav. Ecol. 16, 269–273. doi: 10.1093/beheco/arh152
Judge, K. A., Tran, K.-C., and Gwynne, D. T. (2010). The relative effects of mating status and age on the mating behaviour of female field crickets. Can. J. Zool. 88, 219–223. doi: 10.1139/Z09-139
Kasumovic, M. M., Hall, M. D., and Brooks, R. C. (2012). The juvenile social environment introduces variation in the choice and expression of sexually selected traits. Ecol. Evol. 2, 1036–1047. doi: 10.1002/ece3.230
Kirkpatrick, M., and Dugatkin, L. A. (1994). Sexual selection and the evolutionary effects of copying mate choice. Behav. Ecol. Sociobiol. 34, 443–449. doi: 10.1007/BF00167336
Kopp, M., Servedio, M. R., Mendelson, T. C., Safran, R. J., Rodriguez, R. L., Hauber, M. E., et al. (2018). Mechanisms of assortative mating in speciation with gene flow: connecting theory and empirical research. Am. Nat. 191, 1–20. doi: 10.1086/694889
Kujtan, L., and Dukas, R. (2009). Learning magnifies individual variation in heterospecific mating propensity. Anim. Behav. 78, 549–554. doi: 10.1016/j.anbehav.2009.05.026
Lachlan, R. F., and Servedio, M. R. (2004). Song learning accelerates allopatric speciation. Evolution 58, 2049–2063. doi: 10.1111/j.0014-3820.2004.tb00489.x
Li, X., Ishimoto, H., and Kamikouchi, A. (2018). Auditory experience controls the maturation of song discrimination and sexual response in Drosophila. Elife 7:e34348. doi: 10.7554/eLife.34348
Li, Y.-Y., Yu, J.-F., Lu, Q., Xu, J., and Ye, H. (2014). Female and male moths display different reproductive behavior when facing new versus previous mates. PLoS ONE 9:e109564. doi: 10.1371/journal.pone.0109564
Litte, M. (1979). Mischocyttarus flavitarsis in Arizona: social and nesting biology of a polistine wasp. Zeitschrift für Tierpsychologie 50, 282–312. doi: 10.1111/j.1439-0310.1979.tb01033.x
López, S., Guerrero, A., Bleda, M. J., and Quero, C. (2017). Short-term peripheral sensitization by brief exposure to pheromone components in Spodoptera littoralis. J. Comp. Physiol. A 203, 973–982. doi: 10.1007/s00359-017-1205-5
Lorenz, K. (1935). Der Kumpan in der Umwelt des Vogels. J. für Ornithologie 83, 289–413. doi: 10.1007/BF01905572
Loyau, A., Blanchet, S., Van Laere, P., Clobert, J., and Danchin, E. (2012). When not to copy: female fruit flies use sophisticated public information to avoid mated males. Sci. Rep. 2:768. doi: 10.1038/srep00768
McRobert, S. P., and Tompkins, L. (1988). Two consequences of homosexual courtship performed by Drosophila melanogaster and Drosophila affinis males. Evolution 42, 1093–1097. doi: 10.1111/j.1558-5646.1988.tb02528.x
Mery, F. (2013). Natural variation in learning and memory. Curr. Opin. Neurobiol. 23, 52–56. doi: 10.1016/j.conb.2012.09.001
Mery, F., Belay, A. T., So, A. K., Sokolowski, M. B., and Kawecki, T. J. (2007). Natural polymorphism affecting learning and memory in Drosophila. PNAS 104, 13051–13055. doi: 10.1073/pnas.0702923104
Mery, F., and Kawecki, T. J. (2005). A cost of long-term memory in Drosophila. Science 308:1148. doi: 10.1126/science.1111331
Mery, F., Varela, S. A., Danchin, É, Blanchet, S., Parejo, D., Coolen, I., et al. (2009). Public versus personal information for mate copying in an invertebrate. Curr. Biol. 19, 730–734. doi: 10.1016/j.cub.2009.02.064
Miller, M. N., and Fincke, O. M. (1999). Cues for mate recognition and the effect of prior experience on mate recognition in Enallagma damselflies. J. Insect Behav. 12, 801–814. doi: 10.1023/A:1020957110842
Miller, M. N., and Fincke, O. M. (2004). Mistakes in sexual recognition among sympatric Zygoptera vary with time of day and color morphism (Odonata: Coenagrionidae). Int. J. Odonatol 7, 471–491. doi: 10.1080/13887890.2004.9748233
Morand-Ferron, J. (2017). Why learn? The adaptive value of associative learning in wild populations. Curr. Opin. Behav. Sci 16, 73–79. doi: 10.1016/j.cobeha.2017.03.008
Morand-Ferron, J., and Quinn, J. L. (2015). The evolution of cognition in natural populations. Trends Cogn. Sci. 19, 235–237. doi: 10.1016/j.tics.2015.03.005
Morier-Genoud, R., and Kawecki, T. J. (2015). The effect of learning on the evolution of new courtship behavior: a simulation model. Curr. Zool. 61, 1062–1072. doi: 10.1093/czoolo/61.6.1062
Nieberding, C. M., Van Dyck, H., and Chittka, L. (2018). Adaptive learning in non-social insects: from theory to field work, and back. Curr. Opin. Insect Sci. 27, 75–81. doi: 10.1016/j.cois.2018.03.008
Nöbel, S., Allain, M., Isabel, G., and Danchin, E. (2018a). Mate copying in Drosophila melanogaster males. Anim. Behav. 141, 9–15. doi: 10.1016/j.anbehav.2018.04.019
Nöbel, S., Danchin, E., and Isabel, G. (2018b). Mate-copying for a costly variant in Drosophila melanogaster females. Behav. Ecol. 29, 1150–1156. doi: 10.1093/beheco/ary095
Noor, M. A., Grams, K. L., Bertucci, L. A., and Reiland, J. (2001). Chromosomal inversions and the reproductive isolation of species. PNAS 98, 12084–12088. doi: 10.1073/pnas.221274498
Odeen, A., and Moray, C. M. (2008). Drosophila melanogaster virgins are more likely to mate with strangers than familiar flies. Naturwissenschaften 95, 253–256. doi: 10.1007/s00114-007-0314-3
Olofsson, H., Frame, A. M., and Servedio, M. R. (2011). Can reinforcement occur with a learned trait? Evolution 65, 1992–2003. doi: 10.1111/j.1558-5646.2011.01286.x
Olofsson, H., and Servedio, M. R. (2008). Sympatry affects the evolution of genetic versus cultural determination of song. Behav. Ecol. 19, 596–604. doi: 10.1093/beheco/arn002
Pan, Y., and Baker, B. S. (2014). Genetic identification and separation of innate and experience-dependent courtship behaviors in Drosophila. Cell 156, 236–248. doi: 10.1016/j.cell.2013.11.041
Papaj, D. R., and Snell-Rood, E. C. (2007). Memory flies sooner from flies that learn faster. PNAS 104, 13539–13540. doi: 10.1073/pnas.0706165104
Planqué, R., Britton, N. F., and Slabbekoorn, H. (2014). On the maintenance of bird song dialects. J. Math. Biol. 68, 505–531. doi: 10.1007/s00285-012-0632-8
Prato, M., and Soares, A. E. E. (2013). Production of sexuals and mating frequency in the stingless bee Tetragonisca angustula (Latreille) (Hymenoptera, Apidae). Neotrop. Entomol. 42, 474–482. doi: 10.1007/s13744-013-0154-0
Proffit, M., Khallaf, M. A., Carrasco, D., Larsson, M. C., and Anderson, P. (2015). 'Do you remember the first time?' Host plant preference in a moth is modulated by experiences during larval feeding and adult mating. Ecol. Lett. 18, 365–374. doi: 10.1111/ele.12419
Rebar, D., Barbosa, F., and Greenfield, M. D. (2016). Acoustic experience influences male and female pre- and postcopulatory behaviors in a bushcricket. Behav. Ecol. 27, 434–443. doi: 10.1093/beheco/arv171
Rebar, D., and Rodríguez, R. L. (2013). Genetic variation in social influence on mate preferences. Proc. Biol. Sci. 280:20130803. doi: 10.1098/rspb.2013.0803
Rebar, D., and Rodriguez, R. L. (2016). Males adjust their signalling behaviour according to experience of male signals and male-female signal duets. J. Evol. Biol. 29:766. doi: 10.1111/jeb.12825
Rebar, D., Zuk, M., and Bailey, N. W. (2011). Mating experience in field crickets modifies pre- and postcopulatory female choice in parallel. Behav. Ecol. 22, 303–309. doi: 10.1093/beheco/arq195
Ritchie, M. G. (2000). The inheritance of female preference functions in a mate recognition system. Proc. Biol. Sci. 267, 327–332. doi: 10.1098/rspb.2000.1004
Robledo, N., Arzuffi, R., and Reyes-Prado, H. (2018). Modification of behavioral response in Copitarsia decolora (Lepidoptera: Noctuidae) due to pre-exposure to sex pheromone and host plant volatiles. Fla. Entomol. 101, 69–73. doi: 10.1653/024.101.0113
Rutledge, J. M., Miller, A., and Uetz, G. W. (2010). Exposure to multiple sensory cues as a juvenile affects adult female mate preferences in wolf spiders. Anim. Behav. 80, 419–426. doi: 10.1016/j.anbehav.2010.05.027
Rutledge, J. M., and Uetz, G. W. (2014). Juvenile experience and adult female mating preferences in two closely related Schizocosa species. J. Arachnol. 42, 170–177. doi: 10.1636/J14-10.1
Sadek, M. M., von Wowern, G., Löfstedt, C., Rosén, W.-Q., and Anderson, P. (2012). Modulation of the temporal pattern of calling behavior of female Spodoptera littoralis by exposure to sex pheromone. J. Insect Physiol. 58, 61–66. doi: 10.1016/j.jinsphys.2011.09.016
Saleem, S., Ruggles, P. H., Abbott, W. K., and Carney, G. E. (2014). Sexual experience enhances Drosophila melanogaster male mating behavior and success. PLoS ONE 9:e96639. doi: 10.1371/journal.pone.0096639
Santos, M., Matos, M., and Varela, S. A. M. (2014). Negative public information in mate choice copying helps the spread of a novel trait. Am. Nat. 184, 658–672. doi: 10.1086/678082
Santos, M., Sapage, M., Matos, M., and Varela, S. A. M. (2017). Mate-choice copying: a fitness-enhancing behavior that evolves by indirect selection. Evolution 71, 1456–1464. doi: 10.1111/evo.13235
Schlechter-Helas, J., Schmitt, T., and Peschke, K. (2012). Learning individual signatures: rove beetle males discriminate unreceptive females by cuticular hydrocarbon patterns. Anim. Behav. 84, 369–376. doi: 10.1016/j.anbehav.2012.05.003
Seeholzer, L. F., Seppo, M., Stern, D. L., and Ruta, V. (2018). Evolution of a central neural circuit underlies Drosophila mate preferences. Nature 559, 564–569. doi: 10.1038/s41586-018-0322-9
Servedio, M. R. (2015). Advances on the interplay of learning and sexual selection. Curr. Zool. 61, 1004–1007. doi: 10.1093/czoolo/61.6.1004
Servedio, M. R., and Dukas, R. (2013). Effects on population divergence of within-generational learning about prospective mates. Evolution 67, 2363–2375. doi: 10.1111/evo.12127
Servedio, M. R., and Kirkpatrick, M. (1996). The evolution of mate choice copying by indirect selection. Am. Nat. 148, 848–867. doi: 10.1086/285959
Servedio, M. R., Sæther, S. A., and Sætre, G.-P. (2009). Reinforcement and learning. Evol. Ecol. 23, 109–123. doi: 10.1007/s10682-007-9188-2
Shaw, K. L. (2000). Interspecific genetics of mate recognition: inheritance of female acoustic preference in Hawaiian crickets. Evolution 54, 1303–1312. doi: 10.1111/j.0014-3820.2000.tb00563.x
Shaw, K. L., and Herlihy, D. P. (2000). Acoustic preference functions and song variability in the Hawaiian cricket Laupala cerasina. Proc. Biol. Sci. 267, 577–584. doi: 10.1098/rspb.2000.1040
Silvegren, G., Lofstedt, C., and Qi Rosen, W. (2005). Circadian mating activity and effect of pheromone pre-exposure on pheromone response rhythms in the moth Spodoptera littoralis. J. Insect Physiol. 51, 277–286. doi: 10.1016/j.jinsphys.2004.11.013
Smith, B. H. (1983). Recognition of female kin by male bees through olfactory signals. PNAS 80, 4551–4553. doi: 10.1073/pnas.80.14.4551
Stejskal, K., Streinzer, M., Dyer, A., Paulus, H. F., and Spaethe, J. (2015). Functional significance of labellum pattern variation in a sexually deceptive orchid (Ophrys heldreichii): evidence of individual signature learning effects. PLoS ONE 10:e0142971. doi: 10.1371/journal.pone.0142971
Stelinski, L., Il'ichev, A., and Gut, L. (2006). Antennal and behavioral responses of virgin and mated oriental fruit moth (Lepidoptera: Tortricidae) females to their sex pheromone. Ann. Entomol. Soc. Am. 99, 898–904. doi: 10.1603/0013-8746(2006)99[898:AABROV]2.0.CO;2
Stockton, D. G., Martini, X., and Stelinski, L. (2017). Male psyllids differentially learn in the context of copulation. Insects 8:16. doi: 10.3390/insects8010016
Stoffer, B., and Uetz, G. W. (2015). The effects of social experience with varying male availability on female mate preferences in a wolf spider. Behav. Ecol. Sociobiol. 69, 927–937. doi: 10.1007/s00265-015-1904-2
Stoffer, B., and Uetz, G. W. (2016a). Social experience affects female mate preferences for a visual trait in a wolf spider. Behav. Ecol. 27, 252–261. doi: 10.1093/beheco/arv143
Stoffer, B., and Uetz, G. W. (2016b). Tuft size matters: the effects of adult visual social experience on female mate preferences in a wolf spider. Behav. Ecol. Sociobiol. 70, 2211–2221. doi: 10.1007/s00265-016-2225-9
Stoffer, B., and Uetz, G. W. (2017). The effects of experience with different courtship modalities on unimodal and multimodal preferences in a wolf spider. Anim. Behav. 123, 187–196. doi: 10.1016/j.anbehav.2016.10.033
Stöhr, S. (1998). Evolution of mate-choice copying: a dynamic model. Anim. Behav. 55, 893–903. doi: 10.1006/anbe.1997.0674
Suckling, D. M., Stringer, L. D., Jimenez-Perez, A., Walter, G. H., Sullivan, N., and El-Sayed, A. M. (2018). With or without pheromone habituation: possible differences between insect orders? Pest Manag. Sci. 74, 1259–1264. doi: 10.1002/ps.4828
Sullivan-Beckers, L., and Hebets, E. A. (2011). Modality-specific experience with female feedback increases the efficacy of courtship signalling in male wolf spiders. Anim. Behav. 82, 1051–1057. doi: 10.1016/j.anbehav.2011.07.040
Sullivan-Beckers, L., and Hebets, E. A. (2014). Tactical adjustment of signalling leads to increased mating success and survival. Anim. Behav. 93, 111–117. doi: 10.1016/j.anbehav.2014.04.021
Svensson, E. I., Eroukhmanoff, F., Karlsson, K., Runemark, A., and Brodin, A. (2010). A role for learning in population divergence of mate preferences. Evolution 64, 3101–3113. doi: 10.1111/j.1558-5646.2010.01085.x
Svensson, E. I., Runemark, A., Verzijden, M. N., and Wellenreuther, M. (2014). Sex differences in developmental plasticity and canalization shape population divergence in mate preferences. Proc. Biol. Sci. 281:20141636. doi: 10.1098/rspb.2014.1636
Swanger, E., and Zuk, M. (2015). Cricket responses to sexual signals are influenced more by adult than juvenile experiences. J. Insect Behav. 28, 328–337. doi: 10.1007/s10905-015-9504-6
Takahashi, Y., and Watanabe, M. (2010a). Diurnal changes in male mate preference to female dimorphism in Ischnura senegalensis (Rambur) (Zygoptera: Coenagrionidae). Odonatologica 39, 159–162.
Takahashi, Y., and Watanabe, M. (2010b). Mating experience affecting male discrimination between sexes and female morphs in Ischnura senegalensis (Rambur) (Zygoptera: Coenagrionidae). Odonatologica 39, 47–56.
Takahashi, Y., and Watanabe, M. (2011). Male mate choice based on ontogenetic colour changes of females in the damselfly Ischnura senegalensis. J. Ethol. 29, 293–299. doi: 10.1007/s10164-010-0257-6
Tan, C. K., Lovlie, H., Greenway, E., Goodwin, S. F., Pizzari, T., and Wigby, S. (2013). Sex-specific responses to sexual familiarity, and the role of olfaction in Drosophila. Proc. Biol. Sci. 280:20131691. doi: 10.1098/rspb.2013.1691
Thöming, G., Larsson Mattias, C., Hansson Bill, S., and Anderson, P. (2013). Comparison of plant preference hierarchies of male and female moths and the impact of larval rearing hosts. Ecology 94, 1744–1752. doi: 10.1890/12-0907.1
Tinghitella, R. M. (2014). Male and female crickets modulate their courtship behaviour depending on female experience with mate availability. Anim. Behav. 91, 9–15. doi: 10.1016/j.anbehav.2014.02.022
Vakirtzis, A. (2011). Mate choice copying and nonindependent mate choice: a critical review. Ann. Zool. Fenn. 48, 91–107. doi: 10.5735/086.048.0202
van Gossum, H., De Bruyn, L., and Stoks, R. (2005). Reversible switches between male–male and male–female mating behaviour by male damselflies. Biol. Lett. 1, 268–270. doi: 10.1098/rsbl.2005.0315
van Gossum, H., Stoks, R., and Bruyn, L. D. (2001). Reversible frequency–dependent switches in male mate choice. Proc. Biol. Sci. 268, 83–85. doi: 10.1098/rspb.2000.1333
Varela, S. A. M., Matos, M., and Schlupp, I. (2018). The role of mate-choice copying in speciation and hybridization. Biol. Rev. Camb. Philos. Soc. 93, 1304–1322. doi: 10.1111/brv.12397
Velthuis, H. H. W., Koedam, D., and Imperatriz-Fonseca, V. L. (2005). The males of Melipona and other stingless bees, and their mothers. Apidologie 36, 169–185. doi: 10.1051/apido:2005014
Verzijden, M. N, Lachlan Robert, F., and Servedio Maria, R. (2007). Female mate-choice behavior and sympatric speciation. Evolution 59, 2097–2108. doi: 10.1111/j.0014-3820.2005.tb00920.x
Verzijden, M. N., Abbott, J. K., von Philipsborn, A. C., and Loeschcke, V. (2015). Male Drosophila melanogaster learn to prefer an arbitrary trait associated with female mating status. Curr. Zool. 61, 1036–1042. doi: 10.1093/czoolo/61.6.1036
Verzijden, M. N., and Svensson, E. I. (2016). Interspecific interactions and learning variability jointly drive geographic differences in mate preferences. Evolution 70, 1896–1903. doi: 10.1111/evo.12982
Verzijden, M. N., ten Cate, C., Servedio, M. R., Kozak, G. M., Boughman, J. W., and Svensson, E. I. (2012). The impact of learning on sexual selection and speciation. Trends Ecol. Evol. 27, 511–519. doi: 10.1016/j.tree.2012.05.007
Wade, M. J., and Pruett-Jones, S. G. (1990). Female copying increases the variance in male mating success. PNAS 87, 5749–5753. doi: 10.1073/pnas.87.15.5749
Wagner, W. E., Smeds, M. R., and Wiegmann, D. D. (2001). Experience affects female responses to male song in the variable field cricket Gryllus lineaticeps (Orthoptera, Gryllidae). Ethology 107, 769–776. doi: 10.1046/j.1439-0310.2001.00700.x
Wan, X., Qian, K., and Du, Y. (2015). Synthetic pheromones and plant volatiles alter the expression of chemosensory genes in Spodoptera exigua. Sci. Rep. 5:17320. doi: 10.1038/srep17320
Wcislo, W. T. (1987). The role of learning in the mating biology of a sweat bee Lasioglossum zephyrum (Hymenoptera: Halictidae). Behav. Ecol. Sociobiol. 20, 179–185. doi: 10.1007/BF00299731
Weinstein, A. M., Davis, B. J., Menz, M. H. M., Dixon, K. W., and Phillips, R. D. (2016). Behaviour of sexually deceived ichneumonid wasps and its implications for pollination in Cryptostylis (Orchidaceae). Biol. J. Linn. Soc. 119, 283–298. doi: 10.1111/bij.12841
Westerman, E. L., Chirathivat, N., Schyling, E., and Monteiro, A. (2014). Mate preference for a phenotypically plastic trait is learned, and may facilitate preference-phenotype matching. Evolution 68, 1661–1670. doi: 10.1111/evo.12381
Westerman, E. L., Hodgins-Davis, A., Dinwiddie, A., and Monteiro, A. (2012). Biased learning affects mate choice in a butterfly. PNAS 109, 10948–10953. doi: 10.1073/pnas.1118378109
Westerman, E. L., and Monteiro, A. (2013). Odour influences whether females learn to prefer or to avoid wing patterns of male butterflies. Anim. Behav. 86, 1139–1145. doi: 10.1016/j.anbehav.2013.09.002
Wickman, P.-O., and Jansson, P. (1997). An estimate of female mate searching costs in the lekking butterfly Coenonympha pamphilus. Behav. Ecol. Sociobiol. 40, 321–328. doi: 10.1007/s002650050348
Wilder, S. M., and Rypstra, A. L. (2008). Prior encounters with the opposite sex affect male and female mating behavior in a wolf spider (Araneae, Lycosidae). Behav. Ecol. Sociobiol. 62, 1813–1820. doi: 10.1007/s00265-008-0610-8
Williams, J. M., and Slater, P. J. B. (1990). Modelling bird song dialects: the influence of repertoire size and numbers of neighbours. J. Theor. Biol. 145, 487–496. doi: 10.1016/S0022-5193(05)80483-7
Williams, Z. M. (2016). Transgenerational influence of sensorimotor training on offspring behavior and its neural basis in Drosophila. Neurobiol. Learn. Mem. 131, 166–175. doi: 10.1016/j.nlm.2016.03.023
Witte, K., Kniel, N., and Kureck, I. M. (2015). Mate-choice copying: Status quo and where to go. Curr. Zool. 61, 1073–1081. doi: 10.1093/czoolo/61.6.1073
Xu, J., and Wang, Q. (2009). A polyandrous female moth discriminates against previous mates to gain genetic diversity. Anim. Behav. 78, 1309–1315. doi: 10.1016/j.anbehav.2009.09.028
Yan, S., Zhu, J., Zhu, W., Zhang, X., Li, Z., Liu, X., et al. (2014). The expression of three opsin genes from the compound eye of Helicoverpa armigera (Lepidoptera: Noctuidae) is regulated by a circadian clock, light conditions and nutritional status. PLoS ONE 9:e111683. doi: 10.1371/journal.pone.0111683
Yeh, D. J., and Servedio, M. R. (2015). Reproductive isolation with a learned trait in a structured population. Evolution 69, 1938–1947. doi: 10.1111/evo.12688
Yilmaz, A., Lindenberg, A., Albert, S., Grubel, K., Spaethe, J., Rossler, W., et al. (2016). Age-related and light-induced plasticity in opsin gene expression and in primary and secondary visual centers of the nectar-feeding ant Camponotus rufipes. Dev. Neurobiol. 76, 1041–1057. doi: 10.1002/dneu.22374
Keywords: mate choice, preference, selectivity, signal, social experience, adaptive value, sexual selection
Citation: Dion E, Monteiro A and Nieberding CM (2019) The Role of Learning on Insect and Spider Sexual Behaviors, Sexual Trait Evolution, and Speciation. Front. Ecol. Evol. 6:225. doi: 10.3389/fevo.2018.00225
Received: 01 August 2018; Accepted: 05 December 2018;
Published: 08 January 2019.
Edited by:
Astrid T. Groot, University of Amsterdam, NetherlandsReviewed by:
Valentina Zizzari, Universität Koblenz Landau, GermanyMatthew R. E. Symonds, Deakin University, Australia
Copyright © 2019 Dion, Monteiro and Nieberding. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Emilie Dion, ZGlvbi5lbWlsaWVAeW1haWwuY29t
Antónia Monteiro, YW50b25pYS5tb250ZWlyb0BudXMuZWR1LnNn
Caroline M. Nieberding, Y2Fyb2xpbmUubmllYmVyZGluZ0B1Y2xvdXZhaW4uYmU=