 Ryan Schacht
Ryan Schacht Helen E. Davis3,4
Helen E. Davis3,4- 1Department of Anthropology, East Carolina University, Greenville, NC, United States
- 2Population Sciences, Huntsman Cancer Institute, University of Utah, Salt Lake City, UT, United States
- 3Department of Human Evolutionary Biology, Harvard University, Cambridge, MA, United States
- 4Department of Anthropology, University of Utah, Salt Lake City, UT, United States
Human paternal investment, and that of many other species, is facultatively expressed and dependent on a diverse array of individual, social, and ecological conditions. Well-documented are the various ways in which men invest in offspring and the household. Specifically, local ecology structures pay-offs to male investment and has been shown to be an important predictor of the sexual division of labor. However, while variability in paternal investment has been well-characterized cross-culturally, plasticity within a group in response to changing socioecological conditions remains largely unstudied. To address this, we use recent economic development and market access to explore how changes in socioecology alter behavioral options for men and their resultant investment decisions. Among the monogamous Maya, we find that, associated with the introduction of novel subsistence opportunities and incentives for intensified paternal investment, fathers spend more time in the household, more time in domestic activities and more time interacting with their children. The changes in paternal investment documented here are largely contingent on four conditions: increased efficiency in subsistence brought about by mechanized farming, limited opportunities to engage in wage labor, increased opportunities to invest in offspring quality, and a monogamous mating system. Thus, Maya fathers appear to repurpose found time by furthering investment in their families.
Introduction
Across human societies, men and women typically engage in different yet complementary patterns of parental investment (Murdock and Provost, 1973). This type of role specialization, where males and females engage in sex-specific activities within a pairbond, is observed across many animal taxa (Barta et al., 2014). What is distinctive to humans, however, are the variable ways in which this investment occurs. For example, in some societies males provide nearly all of the calories for subsistence and little childcare (Rasmussen, 1931), in others mothers and fathers contribute relatively equally to household provisioning and men engage considerably in childcare (Griffin and Griffin, 1992; Hewlett, 1993; Kramer, 2009), and still in others men invest little beyond gametes (reviewed in Hewlett and Macfarlan, 2010).
Local ecology, paternity certainty, and opportunities for mating effort have all been shown to be important determinants of paternal involvement because they generally structure pay-offs to male investment across environments (e.g., Marlowe, 2007). However, while variability has been well-described cross-culturally, within-group plasticity in response to changing socioecological conditions remains largely unstudied. Here we target recent economic development and market access in a small-scale Yucatec Maya society to examine the role that changing subsistence options for men play in their parental investment decisions. Below, we briefly review the literature on paternal investment across animal taxa to more broadly situate our research question.
Social Monogamy and Biparental Care
Social monogamy is relatively uncommon across mammals (3–9% of species; Kleiman, 1977; Lukas and Clutton-Brock, 2013), with paternal investment occurring even less frequently (Wright, 1990; Pleck, 1997). This rarity may be driven by sexual conflict, which challenges the emergence of a simple cooperative system of biparental care (Borgerhoff Mulder and Rauch, 2009; Székely, 2014; McNamara and Wolf, 2015). Both parents share the benefits of effort put into raising young, but investing parents may pay an individual opportunity cost or experience lowered survival and/or fertility (Kokko and Jennions, 2008). This raises possible conflict between parents over who pays the costs of care because each is incentivized to minimize their own reproductive costs (e.g., through desertion) at the expense of their partner (Trivers, 1972; Clutton-Brock, 1991; Lessells, 1999; Houston et al., 2005).
Obligate and elevated reproductive costs among females (e.g., gestation, lactation) are commonly referenced to explain the preponderance of maternal care across mammalian taxa (Trivers, 1972). Payoffs to mating multiply are argued to select against paternal care and for males to invest in mating effort (Bateman, 1948). Nonetheless, social monogamy and biparental care are observed across animal taxa and, while rare among mammals (Kleiman, 1977; Wright, 1990), are typical among ~ 90% of bird species (Cockburn, 2006). So why do males invest among some animals and not others? Life history theory offers an evolutionary framework from which to understand variable patterning in paternal investment (Stearns, 1992; Charnov, 1993; Hill, 1993). If resources allocated to reproduction can be directed toward offspring quantity or quality, natural selection is expected to shape investment decisions based on fitness payoffs to time and energy allocated to mating vs. parenting effort.
Under certain circumstances, monogamy can increase male fitness more than deserting a partner and remating (Grafen and Sibly, 1978; Yamamura and Tsuji, 1993; Fromhage et al., 2005; Kokko and Jennions, 2008; Schacht and Bell, 2016). Parental investment theory (Trivers, 1972) outlines several conditions under which male provisioning might evolve: (1) low opportunity costs associated with paternal investment due to, for example, social and ecological factors that reduce male mating opportunities; (2) investment improves offspring survival and/or quality, particularly when payoffs to desertion are low (Dunbar, 1976; Thornhill, 1976; Perrone and Zaret, 1979; Clutton-Brock, 1991; Westneat and Sherman, 1993) and (3) paternity certainty is high, which is necessary for males to avoid squandering investment. Once paternal investment becomes established, specialization of care tasks by males and females may serve to stabilize the pair-bond. Specifically, specialization can lead to synergistic fitness benefits tied to offspring success (Leonetti and Chabot-Hanowell, 2011; Barta et al., 2014). These payoffs both constrain the behavioral options available to a parent and decrease sex-biased asymmetries in the costs of performing a parental investment task. Thus, task specialization strengthens biparental care against invasion by other strategies (Barta et al., 2014).
Role of Fathers
Fathers contribute to their offspring's well-being in a variety of ways. While typically characterized as focused on forms of indirect investment (e.g., resources, calories, protection, monetary investments; Kleiman and Malcolm, 1981), fathers variably engage in the direct care of offspring (e.g., carrying, holding, tending, feeding, and grooming offspring). Among humans, local ecology has been shown to play an important role in mediating payoffs to indirect vs. direct investment (Marlowe, 2007). For example, among the Ache hunter-gatherers of Paraguay, high hunting returns cause food provisioning to earn men higher fitness payoffs (measured as reproductive success) than time spent in other activities, such as childcare (Hill and Kaplan, 1988). Other studies emphasize that variation in male provisioning reflects differences in paternal quality within a population. Aka fathers (Ituri forest hunter-gatherers), for example, hold or are within arm's reach of their infants nearly 50% of the day. But the investments of individual fathers are highly variable and depend on their attractiveness as partners. Poorer, lower status Aka fathers spend more time with their children and provide more childcare than do wealthier, higher status fathers (Hewlett, 1988). Cross-cultural variation in paternal investment can additionally be understood as a response to helper availability (Griffin and Griffin, 1992; Fouts, 2008). For example, among the Agta, foragers native to the Philippines, fathers spend more time in childcare early in a marriage when a mother does not have a daughter old enough to assist (Griffin and Griffin, 1992). Among the Hadza (sub-Saharan hunter-gatherers), husbands spend more time hunting when their wives have diminished foraging efficiency due to a dependent infant (Marlowe, 2003). Maya fathers too respond in this manner and perform all of the agricultural labor when mothers have a nursing infant and also increase their time allocation to food production as family size increases and their children mature (Kramer, 2009).
Male Investment in Response to Socioecological Change Among the Maya
While human fathers participate in indirect and direct investment to varying degrees across human societies, how best to characterize male motivations for investment remains a point of contention (e.g., Gurven and Hill, 2009; Hawkes et al., 2010). However, whether male paternal investment is motivated by mating or parenting benefits is not the focus of the research here. Instead we target a largely unaddressed question: how do patterns of paternal investment alter within a population in response to socioecological change and the introduction of novel provisioning and caring opportunities? This is a relevant and timely question given the rapid rate of market entry and integration experienced by many small-scale societies today. Because human paternal investment is facultative and not obligate, as it is for women (Geary, 2000), men have behavioral options in the face of changing conditions. Specifically, among the Maya, the recent introduction of mechanized farming, which allows fathers to refocus their time and energy saved from food production (once reliant on hand-farming techniques) to, for example, childcare, leisure, or pursuing new mating opportunities. Here, we focus on the understudied question of how males allocate time and energy in a changing economic landscape when opportunities for and payoffs to direct and/or indirect investment opportunities are altered.
While males can choose how to allocate their time, payoffs to a particular behavioral strategy depend on marginal returns to offspring investment (e.g., Hurtado and Hill, 1992). As child success becomes more care-dependent and resource intensive, fathers are expected to adjust their investments accordingly (Pennington and Harpending, 1988). Economic development and a market-based economy are associated with intensified male investment in children. In the US, for example, paternal investment, including income and childcare, is associated with better academic performance in childhood and higher socioeconomic status in adulthood (Pleck, 1997; Kaplan et al., 1998; Gray and Anderson, 2010). As economically independent children become more expensive in terms of resources (e.g., school expenses) and childcare, we expect fathers to invest less in other activities tangential to child success (e.g., leisure).
Leveraging data that were collected in a Maya community across two distinct time periods and spanning 20 years (1992 and 2011), we analyze changes in male investment before and after economic development. Traditionally, Maya men spent much of the time outside of the home, away from the village and attending agricultural fields. This limited opportunities for men to engage in childcare and other household activities. An agrarian economy also limits payoffs to concentrated investment in a few children because large families provide agricultural help for food production and foster relational and labor-based wealth (Kramer and Boone, 2002; Kramer, 2005). However, recent infrastructure development (e.g., a paved road connecting the community to a regional highway) has reduced travel time to once distant urban markets and permitted the adoption of mechanized farming. Both of these changes decrease the time fathers need to spend in travel and farming and, consequently, allow them to reallocate this found time in other ways. Thus, here, we center on socioecological changes as important drivers of variation paternal time allocation. Following a life history approach, and potential payoffs to more intensive child investment in response to economic development, we predict that fathers will invest more in the nuclear family unit in the later time period by spending more time with their immediate family and in childcare activities.
Methods
The Population
To examine questions about shifts in paternal investment, we use time allocation data that were collected across two distinct time periods in a remote rural Maya community in the interior of the Yucatan Peninsula, Campeche, Mexico. Economic, demographic, subsistence, and social trends have been studied in this community since 1992. The first sample of behavioral data used in the following analyses was collected during a year-long time allocation study (Kramer, 2005, 2009). At that time, all families made their living as swidden maize farmers. While the household was the unit of production and consumption, labor and food were exchanged across households. The community's isolation and the lack of roads and vehicles limited opportunities to monetize surplus crop production or engage in the market economy through wage labor. Most agricultural field locations were within several kilometers (about a half hour walk) of the village. By the age of eight, children often accompanied their fathers to the family's farmed fields, where they spent up to 20% of their day (Kramer, 2005). All maize processing (shelling, hauling, soaking, grinding, and cooking) occurred within the household.
However, in the mid-2000s, rapid economic development began when a paved road was built linking the community to the regional economy. The road facilitated access to new farming methods, the transportation of crops to market, and people to wage labor jobs. The introduction of tractors and mechanized farming minimized agricultural labor inputs and time spent farming. These changes too expanded the potential for men to pursue different and new subsistence options in and out of the community, which allowed them to generate cash. However, wage labor opportunities were low-paying and temporary and did not allow households to completely transition to a dependency on market jobs. While wage-labor opportunities outside of the village are generally low-skill, parents recognize that education is an important contributing factor for their children to be competitive on this new economic landscape and to secure a well-paying and permanent position in the future. Thus, formal education also has become a priority for most families. To this point, time spent in school among children 6–15 years of age increased by 50% between 1992 and 2011.
While the subsistence economy changed over the 20-year interval, many aspects of reproduction remained the same. During both time periods (1992 and 2011), Maya marriages can be described as life-long and monogamous (Cashdan et al., 2016). Some women (~25% in the 2011 sample) currently use birth control, although mean completed fertility has only slightly declined over the past 20 years (1992 mean = 7.5 ± 1.74, n = 24; 2011 mean = 6.1 ± 3.01, n = 40). Infant survival is estimated to be 96% (IMR = 37/1000) and has not significantly changed between the two time periods (Veile and Kramer, 2018). From ethnographic observations, fathers are generally engaged with their families and enjoy spending time with their children. Moreover, household relationships are largely egalitarian and husbands and wives jointly make decisions about their children's lives and economic futures.
Data Collection
To evaluate how much time Maya fathers invest in their children and how childcare versus provisioning behaviors change before and after economic development, we compare time allocation data from 1992 to 2011. Time budget information is combined with economic and household composition data for the two time periods from databases collected and maintained by KLK. Birth records are used to determine father's and children's ages and are available for both samples. The ages of participants without birth records have been cross checked during multiple annual censuses.
Time allocation data were collected using instantaneous scan sampling techniques. Instantaneous scan sampling is a behavioral observation technique widely used in both non-human animal and human studies to measure the frequency of activities (Washburn and Lancaster, 1968; Altmann, 1974; Borgerhoff Mulder and Caro, 1985; Hames, 1992). Over repeated observations, instantaneous scan sampling is a reliable method to estimate the proportion of time that an individual spends in various activities (Altmann, 1974; Dunbar, 1976; Simpson and Simpson, 1977) and is more accurate than interview or survey methods to estimate time budgets (Reynolds, 1991).
For both the 1992 and 2011 observation periods, each participant was drawn from the community census and then observed on at least four separate occasions. During those periods, scan samples were recorded, with each observation period lasting ~4 h. An individual's activity and location were recorded at 15 min intervals. The Maya scan data were collected on a subset of the community (112 individuals or 30% of total population in 1992 and 91 individuals or 18% of total population in 2011), which included 15 fathers in each time period whose average ages were not significantly different (mean age is 36.7 years in 1992 and 34 years in 2011). During a scan sample, individuals were located at specific time intervals, with the researcher recording where they were and what they were doing. Over 400 types of activities were coded, which in addition to childcare, included subsistence (foraging, fishing, hunting, fieldwork), domestic (collecting firewood, water, processing food, cooking, sewing, washing), children's play, social, leisure, and hygiene activities. Scan observations primarily occurred in the household. If a person could not be located during a sampling interval, family members were asked about where the individual was and what he or she was doing. Since several hours were spent with a family during each bout, in many cases a person would be observed leaving and then returning with, for example, a load of firewood or a basket of maize, and location and activity information could be confirmed with the participant. The same methods and suite of hierarchical codes were used to record behaviors in 1992 and 2011. Across the two time periods over 25,000 scan samples were recorded, which were culled to paternal observations (n = 2194 in 1992, n = 1021 in 2011). Observations were aggregated by the location in which they occurred, which was recorded at the time of each scan, to determine the proportion of time fathers spent in each of the four geographic spheres (Figure 1): at home, in the village at large, in their agricultural fields (1–15 km distant from the village), or out of town (in distant, larger communities where men travel > 20 km for wage labor, to buy and sell goods, as well as manage children's school registration and education needs).
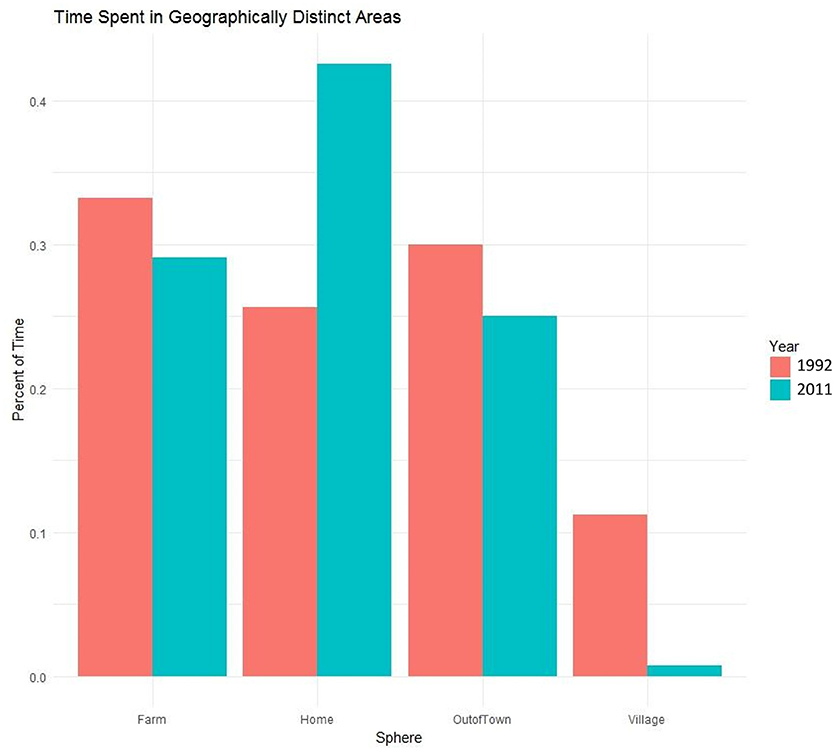
Figure 1. Percentage of time Maya fathers spent in each of four geographically distinct areas in 1992 and 2011.
Within the home a father's time was further categorized into one of three activities: interacting with a child (i.e., direct and indirect childcare), domestic work, or leisure. Direct care (e.g., holding, grooming, carrying, feeding), and indirect care (e.g., helping with homework, disciplining, talking to) were merged into a single childcare category. Domestic work includes food processing and preparation, cleaning, washing, repairing, building, or gardening within the household compound. Leisure was defined as time spent in self-maintenance, bathing, napping, or engaging in social activities with anyone other than a father's children. These suites of activities were then used to calculate the percentage of time fathers engaged in a particular set of behaviors at both time points.
Research protocols and consent procedures were approved by the University of New Mexico's Institutional Review Board (Maya 1992 data) and Harvard University's Institutional Review Board (Maya 2011 data).
Statistical Approach
To evaluate the role of economic development on paternal involvement in the household and investment in children, we first conduct logistic regression to assess whether fathers time allocation budget were different across the two time periods. The year the data were collected is the predictor variable and paternal presence in a particular geographic sphere is the outcome variable. We then use generalized linear mixed models (GLMM) to explore individual predictors of fathers for their presence/absence in a geographic sphere. Fixed effects include: (1) infant in household (<1 year old), (2) number of children in the household (between 1 and 16 years old), (3) the amount of land under cultivation (in hectares) as a measure of wealth, and 4) father's age. Because each father has multiple observations, random effects for “father” are included to account for the nested structure of the data and associated clustering. Random effects allow for heterogeneity in the outcome by individual and for unbalanced observations per person.
To then evaluate how fathers spend their time at home and whether changes occur in childcare activities, we use logistic regression to assess whether activities within the home differ across the two time periods. The year data were collected is the predictor variable and whether a father was observed interacting with his children, engaged in domestic work, or in leisure are the outcome variables. To explain variation among fathers in how they spend time at home, we apply GLMM, with individual predictors of fathers as fixed effects, individuals as random effects, and paternal presence in a particular activity while at home as the outcome.
Analyses were performed in R [35] using lme4 [36]. Multilevel models, where employed, were selected as the best analytic approach because (1) they are appropriate for nested data; (2) intercepts are allowed to vary by the group-level variable (random effect; father); and (3) fixed effects are shared across all groups. All multilevel models include fixed effects for infant in household, number of children in the household, the amount of land under cultivation, and father's age and random effects for individual.
Results
Following economic development, fathers appear to spend more time at home at the expense of time spent in other areas (Figure 1). We find that year is a significant predictor for the probability of spending time in each geographic sphere. For ease of interpretability, we transform the model parameter estimates into odds ratios (OR; Table 1). An OR greater than 1 indicates increased odds that an outcome will occur as the predictor increases. An odds ratio <1 indicates decreased odds of an outcome as the predictor increases. We find that fathers in 2011 are at increased odds to be observed at home (OR = 2.32) and at decreased odds to be spending time in the village (OR = 0.07), at their farm fields (OR = 0.84), or out of town (OR = 0.79; Table 1).

Table 1. Logistic regression with Year as the predictor and geographic areas as outcomes.
Within each period, we then fit generalized linear mixed models to evaluate which paternal characteristics are associated with the probability of a father being observed in a particular geographic sphere. In 1992, and focusing on our significant results, we find that older fathers and those who have an infant in the home are at increased odds to be observed at home (OR = 1.06 and 4.27 respectively; Table 2). Moreover, fathers with more land under cultivation are at increased odds to be found in their agricultural fields (OR = 1.86) and at decreased odds to be out of town (OR = 0.53). Twenty years later, following economic development, these associations are no longer significant. Instead, older fathers and those with an infant are at decreased odds to be found socializing in the village (OR = 0.80 and 0.01 respectively; Table 3). Additionally, fathers with infants are at decreased odds to be observed in their farm fields (OR = 0.26) and those with more children at increased odds to be observed out of town (OR = 1.74; Table 3).
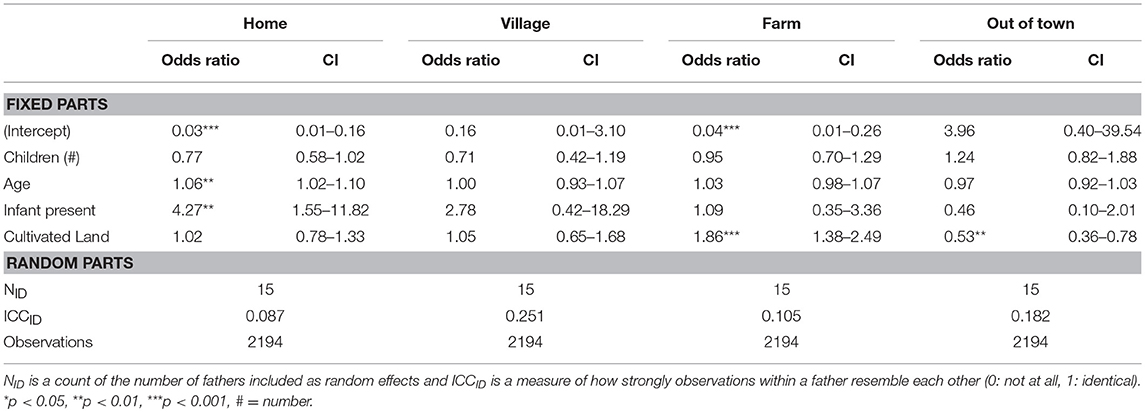
Table 2. Generalized linear multilevel logistic regression with traits of fathers as predictors and geographic areas as outcomes for 1992.
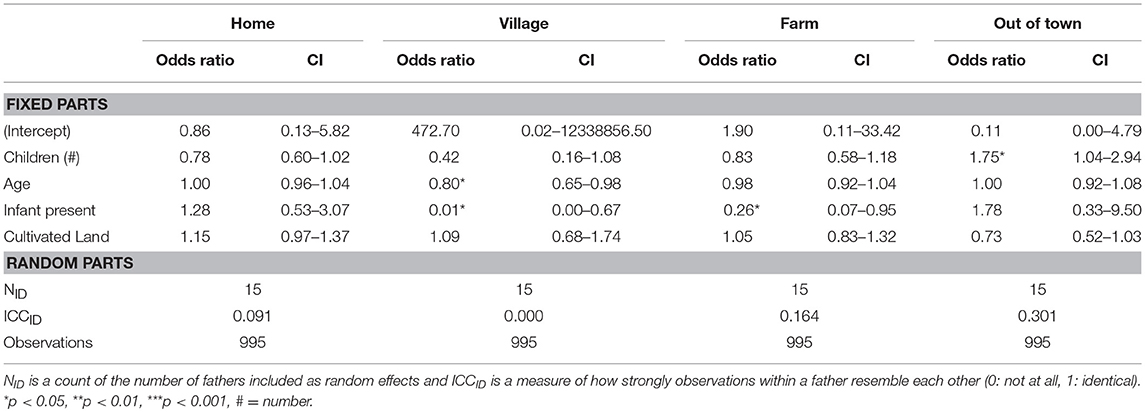
Table 3. Generalized linear multilevel logistic regression with traits of fathers as predictors and geographic areas as outcomes for 2011.
We then ask, when fathers are at home, what are they doing (Figure 2)? A significant shift occurs over the 20-years in whether a father was observed in domestic work, leisure, or child care. In 2011, fathers were at increased odds to be observed in domestic work (OR = 1.45) and interacting with their children (OR = 1.35) and at decreased odds to be observed at leisure (OR = 0.59; Table 4).
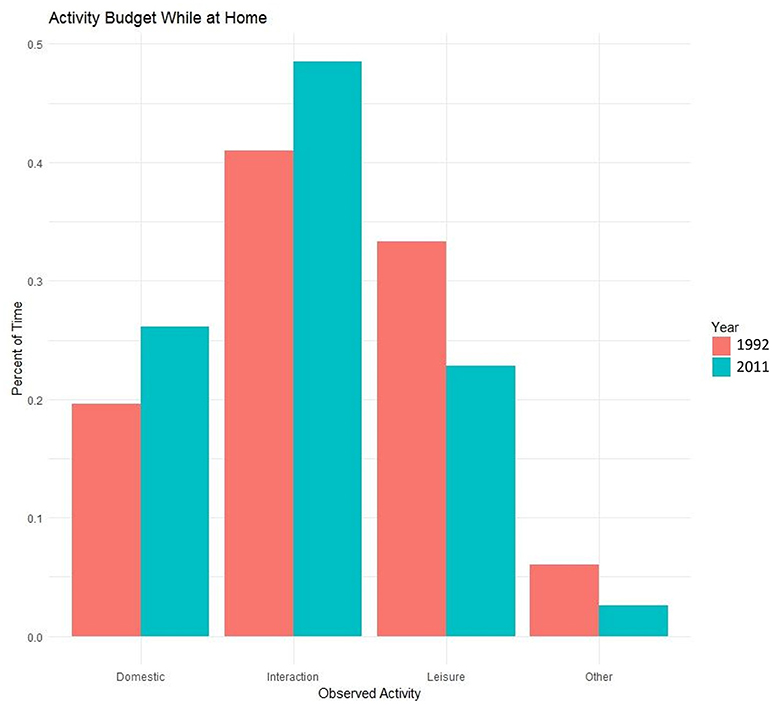
Figure 2. Percentage of time Maya fathers spend in different activities while in the home.

Table 4. Logistic regression with Year as the predictor and activities within home sphere as outcomes.
We then assess the role of fathers' individual traits in predicting how a father spends his time while at home. In 1992, fathers with more children under the age of 16 are at increased odds to be observed in domestic work (OR = 1.35) and at decreased odds to be observed in leisure (OR = 0.64). Odds to be observed at leisure, however, increases with age (OR = 1.04) and if the father has an infant in the home (OR = 4.50; Table 5). In 2011, however, fathers with more land under cultivation are at increased odds to be observed in domestic work (OR = 1.99) and decreased odds to be observed in leisure activities (OR = 0.71), or in childcare (OR = 0.79). Fathers with infants are over five times as likely as those without to be observed in domestic work (OR = 5.57; Table 6).

Table 5. Generalized linear multilevel logistic regression with traits of fathers as predictors and activities at home as outcomes for 1992.
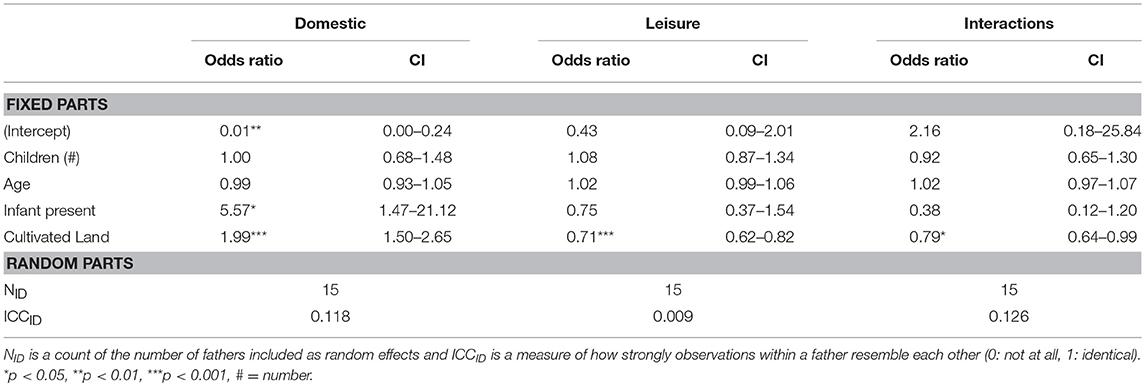
Table 6. Generalized linear multilevel logistic regression with traits of fathers as predictors and activities at home as outcomes for 2011.
In summary, we report seven key findings of relevance to male paternal investment behaviors. After the introduction of economic development, which facilitated access to urban markets and alternative ways to allocate time, fathers spend: (1) more time at home; (2) less time in their agricultural fields and out of town: (3) less time in social activities with other men in the village, particularly when they have an infant in the home; (4) more time interacting with their children (5) more time in domestic work; (6) less time in leisure; and (7) an increasing amount of time out of town in response to more children in the home.
Discussion
Paternal investment can range from genetic inheritance (Savalli and Fox, 1998; Hunt and Simmons, 2000), to pre- or postnatal provisioning (Clutton-Brock, 1991; Kaplan and Lancaster, 2003), to assisting in social development and direct care (Alberts and Altmann, 1995; Kaplan et al., 2000; Marlowe, 2000; Shenk and Scelza, 2012). Among the Maya recent economic development has introduced new subsistence opportunities that subsequently altered the payoff structure for paternal investment. Traditionally, Maya fathers spent much of their day either farming or outside of the community selling agricultural goods. However, the introduction of a paved road, vehicles, and mechanized farming allow the same amount of resources (either food, or cash produced by the sale of agricultural goods) to be generated in less time. These developments have also ushered in rapid social change and a transition from an agrarian economy with few opportunities for formal education to one that is recognized, community-wide, as increasingly skills-based, with children now spending much more of their day in school as a consequence (Kramer, 2005; Veile and Kramer, 2017; Kramer and Veile, 2018; Urlacher and Kramer, 2018).
For this project, we leveraged insight from life history theory as well as research on paternal investment in industrialized populations. Now that Maya fathers have their time open to other pursuits, what do they do with this found time? We predicted that in response to economic development and a transition to a skills-based and education dependent economy, fathers would devote a greater proportion of their time budget to the needs of their children. Economic development and a market-based economy have been shown to incentivize intense investment in children, both in terms of indirect and direct investment (Pleck, 1997; Kaplan et al., 1998). As child success becomes more care dependent and resource intensive, fathers are expected to adjust their investments accordingly. Consistent with this expectation we found that fathers indeed began investing more in the nuclear family unit by spending more time at home, more time interacting with their children, and less time in leisure. Thus, children today are getting a larger share of their father's time budget than they did in the past.
The complementary nature of the division of labor as well as normatively enforced monogamy among the Maya results in few investment options outside of the pair-bond for men. Marriage to a single partner is the avenue toward adulthood, household formation, and the production of children. Over the 20-year period considered here, divorce and out of wedlock birth have never been reported. Thus, generalizing from this case study, we would predict that, in a monogamous population with few opportunities for males to earn fitness benefits through other means, when opportunities arise to augment offspring quality, fathers will respond by intensifying investment in their children.
Monogamy describes the Maya mating pattern, however, sexual exclusivity within marriage is not a human universal, neither for men nor women (Neel, 1972; Beckerman and Valentine, 2002; Anderson, 2006; Scelza, 2013). In societies where monogamy is either not the norm or not enforced, economic development may lead men to invest less in their children. This is likely in societies where lifelong monogamy is atypical and where payoffs to direct and/or indirect care by men are outweighed by benefits to mating effort. Among Caribbean households, for example, both paternity uncertainty and limited male economic opportunities have been offered to explain why men are typically peripheral to family structure (and why offspring success appears to be more dependent on female kinship networks; reviewed in Gray and Brown, 2015). Moreover, within polygynous pastoral societies, fathers may focus turning new found wealth into additional partners, as opposed to intensified investments in children (Hedges et al., 2016). Here, again, life history theory offers an evolutionary framework from which to understand variable patterning in paternal investment (Stearns, 1992; Charnov, 1993; Hill, 1993). Because male investment is facultative, when fitness payoffs toward time and energy allocated to mating effort outweigh parenting effort, we expect men to invest less in direct and indirect care.
While the nuclear family (i.e., mother, father, and dependent children as a self-contained economic entity) is often thought to be the fundamental human unit, this family structure is typical of industrialized rather than small-scale societies. It is well documented that the human family exhibits remarkable flexibility within and across populations and social organization typical of cooperative breeders (Hrdy, 2009; Russell and Lummaa, 2009; Kramer, 2010; van Schaik and Burkart, 2010; Kramer and Russell, 2015). While some form of pair-bonding is observed cross-culturally, with mothers as the primary infant caregiver, extended kin, and alloparental support are also necessary for offspring success. However, economic development may lead to the economic nuclearization of the household (Keilman, 1987). Among the Maya, as fathers spend less time in other activities unrelated to indirect and direct provisioning (e.g., leisure) and less time out of the household socializing, male relational wealth is likely to decrease. Large networks are indeed important for managing resource needs of large families, particularly under conditions of resources scarcity. However they can also limit the ability of households to accumulate wealth necessary for child success where education is costly, yet necessary in a skills-based economy (Stack, 1974). Thus we may see the emergence of the nuclear family as an independent economic unit among the Maya as fathers intensify their investment in fewer children and spend less time and resources on both extended-kin and non-kin. Possibly of concern in the current Maya context, particularly given that fertility has remained high, is that nuclear households may not yet be self-sufficient, and so child outcomes may suffer in response to smaller sharing networks until fertility declines.
Economic development across a broad swath of small-scale societies is currently occurring very rapidly. However, we do not expect outcomes to be uniform across place. Optimal levels of paternal investment are expected to vary by socioecological factors, including subsistence type, social organization, and mating system. The changes in paternal investment documented here are largely contingent on four conditions: increased efficiency in subsistence brought about by mechanized farming, limited opportunities to engage in wage labor, increased opportunities to invest in offspring quality, and a monogamous mating system. Because males can choose how to allocate their time, payoffs to a particular strategy are expected to be sensitive to marginal returns to offspring investment. Fathers with additional time on their hands as a consequence of mechanized farming are not universally expected to invest more in their children. This time could be reallocated to mating effort, leisure, socializing, or in a number of other ways. However, among the Maya, as a consequence of monogamous marriage and a transition to a skills-based economy, fathers are directing “found time” to their children. In other small-scale societies, economic development in contrast has opened up opportunities for fathers to work outside of the community, which has led to fathers to adopt market jobs as opportunities to provision the family, and thus spend more time away from their children (Mattison et al., 2014). However, in rural Mexico, these positions are low-paying and temporary and cannot alone support a household. As evidence of this, no father in the 2011 sample had given up farming for wage labor. However, from interviews with fathers in the community, child success is increasingly recognized as care-dependent and resource intensive in response to the emergence of a skills-based economy. Thus, we conclude that this change in the patterning of child success, coupled with few mating opportunities outside of marriage, incentivize fathers to funnel investments toward their children.
Limitations
While behavioral observation and time allocation data are a “gold-standard” in the study of behavior, this approach also has limitations. Because of the intensive time investment required for data collection, the sample of fathers for both time periods is small, potentially giving only a partial view of paternal behavior. To minimize bias, fathers included in the study were drawn from a community-wide census. One source of bias that we were not able to account for was migration-bias. Our data come from men who have chosen to remain in the community, which may select for particular kinds of men. And while not a limitation per-se, but because of our focus on fathers, we here leave out many other family members. For example, mother's time budgets are changing as well. In 2011, mothers spent over 70% of their time in the home, an increase from 1992, with comparatively little time in other areas (Figure 3). This increase is in part due to older children spending more time in school and so are less available as helpers. While fathers spent more time at home in 2011 than in 1992, they still engaged in calorie and income generating activities outside of the home. Mothers have taken over more of the care of young children since 1992, and in particular spend more time in the direct care of infants (Kramer and Veile, 2018). This is consistent with other studies that find an association between market integration and young children being cared for more often by their mothers (Valeggia, 2009). What this means for future fertility patterns as well as for gender relations, as women potentially transition into a fully domestic role, is the target of future research.
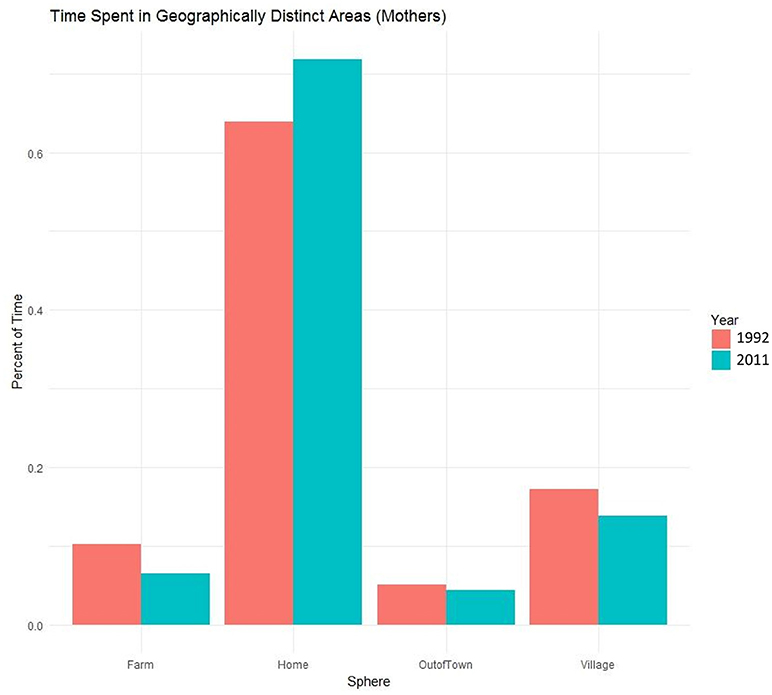
Figure 3. Percentage of time Maya mothers spent in each of four geographically distinct areas in 1992 and 2011.
Conclusion
Maternal investment is necessary for infants and young children to survive in all but the most modern of human societies (Kramer, 2010). In contrast, human paternal investment, and that of many other species, is facultatively expressed and dependent on a diverse array of individual, social, and ecological conditions (Westneat and Sherman, 1993), making it highly variable (Cashdan, 1993; Kaplan and Lancaster, 2003; Geary, 2005; Marlowe, 2007; Gray and Anderson, 2010). To this point, cross-culturally, fathers provide between 16% and <1% of the direct care received by a child (Kramer, 2010). Among the monogamous Maya, we find that incentives for intensified paternal investment, driven by the introduction of novel subsistence opportunities, are associated with fathers spending more time in the household, more time in domestic activities and more time interacting with their children. Thus, Maya fathers appear to repurpose found time by furthering investment in their families. These changes appear contingent on 1) increased efficiency in feeding offspring (indirect investment) such that more time is available for alternative activities; 2) limited opportunities to reinvest found time away from home (e.g., wage labor); 3) marginal benefits to invest in offspring (e.g., payoffs to education); 4) monogamy, which constrains mating effort. In sum, here, in response to changing requirements for offspring independence, we find fathers to be responsive and accordingly spending less time in activities not directly relevant to parenting effort.
Author Contributions
The coauthors collectively conceived the idea and edited the manuscript. RS and KK wrote the manuscript. KK collected the data, HD created the database, and RS performed the analyses.
Funding
We are grateful for financial support from the National Science Foundation (1632338) and the Wenner-Gren Foundation.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Alberts, S. C., and Altmann, J. (1995). Preparation and activation: determinants of age at reproductive maturity in male baboons. Behav. Ecol. Sociobiol. 36, 397–406. doi: 10.1007/BF00177335
Altmann, J. (1974). Observational study of behavior: sampling methods. Behaviour 49, 227–266. doi: 10.1163/156853974X00534
Anderson, K. (2006). How well does paternity confidence match actual paternity? Evidence from worldwide nonpaternity rates. Curr. Anthropol. 47, 513–520. doi: 10.1086/504167
Barta, Z., Székely, T., Liker, A., and Harrison, F. (2014). Social role specialization promotes cooperation between parents. Am. Nat. 183, 747–761. doi: 10.1086/676014
Beckerman, S., and Valentine, P. (2002). Cultures of Multiple Fathers: The Theory and Practice of Partible Paternity in Lowland South America. Gainesville, FL: University Press of Florida.
Borgerhoff Mulder, M., and Caro, T. M. (1985). The use of quantitative observational techniques in anthropology. Curr. Anthropol. 26, 323–335. doi: 10.1086/203277
Borgerhoff Mulder, M., and Rauch, K. L. (2009). Sexual conflict in humans: variations and solutions. Evol. Anthropol. 18, 201–214. doi: 10.1002/evan.20226
Cashdan, E. (1993). Attracting mates: effects of paternal investment on mate attraction strategies. Evol. and Hum. Behav. 14.1, 1–23. doi: 10.1016/0162-3095(93)90014-9
Cashdan, E., Kramer, K. L., Davis, H. E., Padilla, L., and Greaves, R. D. (2016). Mobility and navigation among the Yucatec Maya. Hum. Nat. 27, 35–50. doi: 10.1007/s12110-015-9250-7
Charnov, E. L. (1993). Life History Invariants: Some Explorations of Symmetry in Evolutionary Ecology. Oxford University Press.
Clutton-Brock, T. H. (1991). The Evolution of Parental Care. Princeton, NJ: Princeton University Press.
Cockburn, A. (2006). Prevalence of different modes of parental care in birds. Proc. Biol. Sci. 273, 1375–1383. doi: 10.1098/rspb.2005.3458
Dunbar, R. I. M. (1976). Some aspects of research design and their implications in the observational study of behaviour. Behaviour 58, 58–78.
Fouts, H. N. (2008). Father involvement with young children among the Aka and Bofi foragers. Cross Cult. Res. 42, 290–312. doi: 10.1177/1069397108317484
Fromhage, L., Elgar, M. A., and Schneider, J. M. (2005). Faithful without Care: the evolution of monogyny. Evolution 59, 1400–1405. doi: 10.1111/j.0014-3820.2005.tb01790.x
Geary, D. C. (2000). Evolution and proximate expression of human paternal investment. Psychol. Bull. 126, 55–77. doi: 10.1037/0033-2909.126.1.55
Geary, D. C. (2005). The Origin of Mind: Evolution of Brain, Cognition, and General Intelligence. Washington, DC: American Psychological Association.
Grafen, A., and Sibly, R. (1978). A model of mate desertion. Anim. Behav. 26, 645–652. doi: 10.1016/0003-3472(78)90131-8
Gray, P. B., and Anderson, K. G. (2010). Fatherhood: Evolution and Human Paternal Behavior. Cambridge: Harvard University Press.
Gray, P. B., and Brown, E. (2015). Fatherhood in St. Kitts: patterns and predictors of partnership and paternal dynamics in a Caribbean island. Fathering 13:18. doi: 10.3149/fth.1301.18
Griffin, P. B., and Griffin, M. B. (1992). “Fathers and childcare among the Cagayan Agta,” in Foundations of Human Behavior. Father-Child Relations: Cultural and Biosocial Contexts, ed B. S. Hewlett (Hawthorne, NY: Aldine de Gruyter), 297–320.
Gurven, M., and Hill, K. (2009). Why do men hunt? A reevaluation of “Man the Hunter” and the sexual division of labor. Curr. Anthropol. 50, 51–74. doi: 10.1086/595620
Hames, R. (1992). “Time allocation,” in Evolutionary Ecology and Human Behavior, eds E. A. Smith and B. Winterhalder (New York, NY: Aldine De Gruyter), 203–235.
Hawkes, K., O'Connell, J. F., and Coxworth, J. E. (2010). Family provisioning is not the only reason men hunt: a comment on gurven and hill. Curr. Anthropol. 51, 259–264. doi: 10.1086/651074
Hedges, S., Mulder, M. B., James, S., and Lawson, D. W. (2016). Sending children to school: rural livelihoods and parental investment in education in northern Tanzania. Evol. Hum. Behav. 37, 142–151. doi: 10.1016/j.evolhumbehav.2015.10.001
Hewlett, B. (1988). “Sexual selection and paternal investment among Aka pygmies,” in Human Reproductive Behavior: A Darwinian Perspective, edS L. Betzig, M. Borgerhoff Mulder and P. Turke (Cambridge: Cambridge University Press), 263–276
Hewlett, B., and Macfarlan, S. (2010). “Fathers' roles in hunter-gatherer and other small scale cultures,” in The Role of the Father in Child Development, 5th Edn., ed M. E. Lamb (Hoboken, NJ: John Wiley & Sons), 413–434.
Hewlett, B. S. (1993). Intimate Fathers: The Nature and Context of Aka Pygmy Paternal Infant Care. Ann Arbor, MI: University of Michigan Press.
Hill, K. (1993). Life history theory and evolutionary anthropology. Evol. Anthropol. 2, 78–88. doi: 10.1002/evan.1360020303
Hill, K., and Kaplan, H. (1988). Tradeoffs in male and female reproductive strategies among the Ache: Part 1.“ Hum. Reprod. Behav. 277–290.
Houston, A. I., Székely, T., and McNamara, J. M. (2005). Conflict between parents over care. Trends Ecol. Evol. 20, 33–38. doi: 10.1016/j.tree.2004.10.008
Hrdy, S. B. (2009). Mothers and Others: The Evolutionary Origins of Mutual Understanding. Cambridge, MA: Belknap Press of Harvard University Press.
Hunt, J., and Simmons, L. (2000). Maternal and paternal effects on offspring phenotype in the dung beetle Onthophagus taurus. Evolution 54, 936–941. doi: 10.1111/j.0014-3820.2000.tb00093.x
Hurtado, A. M., and Hill, K. R. (1992). “Paternal effect on offspring survivorship among Ache and Hiwi hunter-gatherers: Implications for modeling pair-bond stability,” in Foundations of Human Behavior. FatherChild Relations: Cultural and Biosocial Contexts, ed B. S. Hewlett (Hawthorne, NY: Aldine de Gruyter), 31–55.
Kaplan, H., Hill, K., Lancaster, J., and Hurtado, A. M. (2000). A theory of human life history evolution: diet, intelligence, and longevity. Evol. Anthrop. 9, 156–185. doi: 10.1002/1520-6505(2000)9:4<156::AID-EVAN5>3.0.CO;2-7
Kaplan, H. S., and Lancaster, J. B. (2003). “An evolutionary and ecological analysis of human fertility, mating patterns, and parental investment,” in National Research Council (US) Panel for the Workshop on the Biodemography of Fertility and Family Behavior, Offspring: Human Fertility Behavior in Biodemographic Perspective, eds K. W. Wachter and R. A. Bulatao (Washington, DC: National Academies Press).
Kaplan, H. S., Lancaster, J. B., and Anderson, K. G. (1998). “Human parental investment and fertility: the life histories of men in Albuquerque,” in Men in Families: When Do They Get Involved? What Difference Does It Make? eds A. Booth and A. C. Crouter (Mahwah, NJ: Lawrence Erlbaum Associates Publishers), 55–109.
Keilmans, N. (1987). Recent trends in family and household composition in Europe. Eur. J. Popul. 3, 297–325. doi: 10.1007/BF01796903
Kleiman, D. G., and Malcolm, J. R. (1981). “The evolution of male parental investment in mammals,” in Parental Care in Mammals, eds D. J. Gubernick and P. H. Klopfer (Boston, MA: Springer).
Kokko, H., and Jennions, M. D. (2008). Parental investment, sexual selection and sex ratios. J. Evol. Biol. 21, 919–948. doi: 10.1111/j.1420-9101.2008.01540.x
Kramer, K. (2009). “Does it take a family to raise a child? Cooperative breeding and the contributions of Maya siblings, parents and older adults in raising children,” in Substitute Parents: Biological and Social Perspectives on Alloparenting in Human Societies, 1st Edn., eds G. Bentley and R. Mace (Berghahn Books), 77–99. Available online at: www.jstor.org/stable/j.ctt9qch9m.9
Kramer, K., and Boone, J. (2002). Why intensive agriculturalists have higher fertility: a household energy budget approach. Curr. Anthropol. 43, 511–517. doi: 10.1086/340239
Kramer, K. L. (2010). Cooperative breeding and its significance to the demographic success of humans. Annu. Rev. Anthropol. 39, 417–436. doi: 10.1146/annurev.anthro.012809.105054
Kramer, K. L., and Russell, A. F. (2015). Was monogamy a key step on the Hominin road? Reevaluating the monogamy hypothesis in the evolution of cooperative breeding. Evol. Anthropol. 24, 73–83. doi: 10.1002/evan.21445
Kramer, K. L., and Veile, A. (2018). Infant allocare in traditional societies. Physiol. Behav. 93(Pt A), 117–126. doi: 10.1016/j.physbeh.2018.02.054
Leonetti, D. L., and Chabot-Hanowell, B. (2011). The foundation of kinship: Households. Hum. Nat. 22, 16–40. doi: 10.1007/s12110-011-9111-y
Lessells, C. (1999). “Sexual Conflict,” in Levels of Selection in Evolution, ed L. Keller (Princeton, NJ: Princeton University Press), 75–99.
Lukas, D., and Clutton-Brock, T. H. (2013). The evolution of social monogamy in mammals. Science 341, 526–530. doi: 10.1126/science.1238677
Marlowe, F. (2000). Paternal investment and the human mating system. Behav. Proc. 51, 45–61. doi: 10.1016/S0376-6357(00)00118-2
Marlowe, F. W. (2003). A critical period for provisioning by Hadza men: Implications for pair bonding. Evol. Hum. Behav/. 24, 217–229. doi: 10.1016/S1090-5138(03)00014-X
Marlowe, F. W. (2007). Hunting and gathering: the human sexual division of foraging labor. Cross Cult. Res. 41, 170–195. doi: 10.1177/1069397106297529
Mattison, S. M., Scelza, B., and Blumenfield, T. (2014). Paternal investment and the positive effects of fathers among the matrilineal Mosuo of Southwest China. Am. Anthropol. 116, 591–610. doi: 10.1111/aman.12125
McNamara, J. M., and Wolf, M. (2015). Sexual conflict over parental care promotes the evolution of sex differences in care and the ability to care. Proc. Biol. Sci. 282:20142752. doi: 10.1098/rspb.2014.2752
Murdock, G. P., and Provost, C. (1973). Factors in the division of labor by sex: a cross-cultural analysis. Ethnology 12, 203–225. doi: 10.2307/3773347
Neel, J. V. (1972). The genetic structure of a tribal population, the Yanomama Indians. Ann. Hum. Genet. 35, 255–259. doi: 10.1111/j.1469-1809.1957.tb01399.x
Pennington, R., and Harpending, H. (1988). Fitness and fertility among Kalahari! Kung. Am. J. Phys. Anthropol. 77, 303–319. doi: 10.1002/ajpa.1330770304
Perrone, M., and Zaret, T. M. (1979). Parental care patterns of fishes. Am. Nat. 113, 351–361. doi: 10.1086/283394
Rasmussen, K. (1931). The Netsilik Eskimos: Social life and Spiritual Culture. Norwalk, CT: Ams Press.
Reynolds, P. (1991). Dance Civet Cat. Child Labour in the Zambezi Valley. Athens: Ohio University Press.
Russell, A. F., and Lummaa, V. (2009). Maternal effects in cooperative breeders: from hymenopterans to humans. Philos. Trans. R. Soc. B Biol. Sci. 364, 1143–1167. doi: 10.1098/rstb.2008.0298
Savalli, U. M., and Fox, C. W. (1998). Genetic variation in paternal investment in a seed beetle. Anim. Behav. 56, 953–961. doi: 10.1006/anbe.1998.0853
Scelza, B. A. (2013). Choosy but not chaste: multiple mating in human females. Evol. Anthropol. 22, 259–269. doi: 10.1002/evan.21373
Schacht, R., and Bell, A. (2016). The evolution of monogamy in response to partner scarcity. Nat. Sci. Commun.6:32472. doi: 10.1038/srep32472
Shenk, M. K., and Scelza, B. A. (2012). Paternal investment and status-related child outcomes: timing of father's death affects offspring success. J. Biosoc. Sci. 44, 549–569. doi: 10.1017/S0021932012000053
Simpson, M., and Simpson, A. (1977). One-zero and scan methods for sampling behaviour. Anim. Behav. 25, 726–731. doi: 10.1016/0003-3472(77)90122-1
Székely, T. (2014). Sexual conflict between parents: offspring desertion and asymmetrical parental care. Cold Spring Harb. Perspect. Biol. 6:a017665. doi: 10.1101/cshperspect.a017665
Thornhill, R. (1976). Sexual selection and paternal investment in insects. Am. Nat. 110, 153–163. doi: 10.1086/283055
Trivers, R. L. (1972). Parental investment and sexual selection. in Sexual Selection and the Descent of Man, 1871-1971, eds B. Campbell (Chicago: Aldine), 136–179.
Urlacher, S. S., and Kramer, K. L. (2018). Evidence for energetic tradeoffs between physical activity and childhood growth across the nutritional transition. Sci. Rep. 8:369. doi: 10.1038/s41598-017-18738-4
Valeggia, C. R. (2009). “Flexible caretakers. responses of toba families in transition,” in substitute Parents. Biological and Social Perspectives on Alloparenting in Human Societies, eds G. Bentley and R. Mace (New York, NY: Berghahn Press), 100–114.
van Schaik, C. P., and Burkart, J. M. (2010). “Mind the gap: cooperative breeding and the evolution of our unique features,” in Mind the Gap, eds P. Kappeler and J. Silk (Berlin; Heidelberg: Springer).
Veile, A., and Kramer, K. L. (2017). Childhood body mass is positively associated with cesarean birth in Yucatec Maya subsistence farmers. Am. J. Hum. Biol. 29 :e22920. doi: 10.1002/ajhb.22920
Veile, A., and Kramer, K. L. (2018). “Shifting weanling's optimum: breastfeeding ecology and infant health in Yucatan,” in Breastfeeding. New Anthropological Approaches, eds C. Tomori, A. Palmquist and E. Quinn (London: Routledge Press), 170–184.
Washburn, S., and Lancaster, C. (1968). The evolution of human hunting. in Man the Hunter, eds R. Lee and L. DeVore (Chicago: Aldine de Gruyter), 293–320.
Westneat, D. F., and Sherman, P. W. (1993). Parentage and the evolution of parental behavior. Behav. Ecol. 4, 66–77. doi: 10.1093/beheco/4.1.66
Wright, P. C. (1990). Patterns of paternal care in primates. Int. J. Primatol. 11, 89–102. doi: 10.1007/BF02192783
Keywords: paternal investment, mating effort, division of labor, Maya, life history theory
Citation: Schacht R, Davis HE and Kramer KL (2018) Patterning of Paternal Investment in Response to Socioecological Change. Front. Ecol. Evol. 6:142. doi: 10.3389/fevo.2018.00142
Received: 11 May 2018; Accepted: 28 August 2018;
Published: 17 October 2018.
Edited by:
Alexander G. Ophir, Cornell University, United StatesReviewed by:
Peter B. Gray, University of Nevada, Las Vegas, United StatesSiobhan Mary Mattison, University of New Mexico, United States
Copyright © 2018 Schacht, Davis and Kramer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ryan Schacht, c2NoYWNodHIxOEBlY3UuZWR1